
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Plant Sci. , 02 December 2024
Sec. Plant Abiotic Stress
Volume 15 - 2024 | https://doi.org/10.3389/fpls.2024.1481372
This article is part of the Research Topic Managing Metal Toxicity in Plants and Soil: Strategies for Stress Mitigation and Remediation View all 9 articles
 Anas Iqbal1
Anas Iqbal1 Xiaoyuan Chen2Rayyan Khan3Maid Zaman4
Xiaoyuan Chen2Rayyan Khan3Maid Zaman4 Aamir Hamid Khan5Marcin Kiedrzyński5Mohamed Ebaid6
Aamir Hamid Khan5Marcin Kiedrzyński5Mohamed Ebaid6 Abdulwahed Fahad Alrefaei7
Abdulwahed Fahad Alrefaei7 Sobhi F. Lamlom8Xiangru Tang1*
Sobhi F. Lamlom8Xiangru Tang1* Muhammad Zeeshan1,2*
Muhammad Zeeshan1,2*Cadmium (Cd) pollution in arable soils and its accumulation in rice plants have become a global concern because of their harmful effects on crop yield and human health. The in-situ stabilization method which involves the application of organic amendments such as vermicompost (VC), is frequently utilized for the remediation of Cd-contaminated soils. This study investigated the effects of VC on the soil chemical properties and the physio-biochemical functions of fragrant rice, as well as nitrogen (N) metabolism and assimilatory enzyme activities, 2-acetyl-1-pyrroline (2AP) content in rice grains, and the grain yields of fragrant rice cultivars, i.e., Xiangyaxiangzhan (XGZ) and.Meixiangzhan-2 (MXZ-2) under Cd stress condition. Four doses of VC (.VC1 = 0, VC2 = 3 t. ha-1, VC3 = 4 t ha-1, and VC4 = 6 t ha-1) and two levels of Cd (0 and 25 mg Cd kg-1) were used in this study. Our results showed that VC supplementation significantly (p < 0.05) improved soil characteristics, including soil organic carbon, available N, total N, phosphorus (P), and potassium (K). Furthermore, VC enhanced plant physiological and biochemical attributes in fragrant rice, such as net photosynthetic rate (Pn), nitrate reductase (NR), nitrite reductase (NiR), glutamine synthetase (GS), glutamate oxoglutarate aminotransferase (GOGAT) enzyme activities, protein contents, amino acid, and 2-acetyl-1-pyrroline (2AP) contents under Cd stress condition. Specifically, the VC-amended treatment, Cd2 + VC3, led to an 86.75% increase in Pn and 2AP, and a 60.05% and 77.55% increase in grain yield for MXZ-2 and XGZ cultivars, respectively, compared to Cd-only treated plants (Cd2 + VC1). In addition, VC application significantly (p < 0.05) decreased the Cd uptake and accumulation in rice plants. The correlation analysis indicated that leaf physiological activity and biochemical traits are strongly correlated with soil qualitative traits, suggesting that improved soil health leads to enhanced leaf physiological activity, N metabolism, grain 2AP content, and grain yields. Among the treatments, Cd2 + VC3 showed the best performance in terms of soil fertility and rice quality and production. Consequently, our study indicates that using VC in soils may benefit rice growers by improving soil fertility and supporting sustainable rice productivity and quality in soils contaminated with Cd.
Fragrant rice is considered premium quality and is globally recognized for its distinct aroma and flavor (Bryant and McClung, 2011). Fragrant rice cultivars release unique aromatic components that set them apart from non-fragrant rice cultivars (Jezussek et al., 2002). Several volatile compounds have been identified in fragrant rice, such as 2AP, 4,5-epoxy-(E)-2-decennial, 2-amino acetophenone, 4-vinyl phenol, 4-vinyl-guaiacol, octanal, decanal and hexanal (Jezussek et al., 2002). However, 2AP is noted as to primary contributor to the grain aroma (Mo et al., 2019). Soil quality degradation poses a significant challenge to sustainable crop production, with heavy metal toxicity being a major factor in the decline of soil fertility (Keesstra et al., 2016; Iqbal et al., 2024). Heavy metals, particularly Cd, are among the most hazardous metals owing to their high toxicity and significant bioaccumulation in cereal plants (Singh et al., 2020). Whereas, rice, a food source for approximately 3.5 billion people globally (Dabral et al., 2019), is particularly vulnerable to Cd stress. Cd accumulation in arable soil is caused by industrial processes such as waste discharge, fertilization, mining, and smelting (Islam et al., 2017; Tang et al., 2019; Seleiman et al., 2020). Cd is more soluble and mobile than other metals, thus it is easily absorbed by plants, translocated, and deposited in various parts of plants (Chen et al., 2018; Adil et al., 2020). Furthermore, Cd is often not recyclable and difficult to eliminate from the soil, and it can transfer to cereal grains via the soil-plant-food cycle, posing a health risk to humans (Rizwan et al., 2016; Seleiman et al., 2020). In China, about 2.78 × 109 m2 of farming land is contaminated by Cd (Xue et al., 2017; Huang et al., 2019). Cd inputs into the soil also adversely affect soil biodiversity and its associated ecosystem function (Haider et al., 2021). Thus, Cd has gained a lot of attention in arable soil because of its toxicity, accessibility, and long life in living organisms (Rizwan et al., 2016).
High levels of Cd in agricultural soils can detrimentally impact soil health, physio-chemical properties, and plant metabolism, leading to diminished crop growth, productivity, and quality (Mitra et al., 2018; Iqbal et al., 2023). Cd also hampers photosynthesis in plants and reduces the uptake of essential nutrients, thereby decreasing agricultural yields (Tran and Popova, 2013; Chen et al., 2018). Furthermore, Cd stress induces morpho-physiological and biochemical, alterations in plants, such as reduced root growth, stomatal density, and N metabolism enzyme activities (Bari et al., 2019; Huybrechts et al., 2020). The photosynthetic apparatus is particularly susceptible to Cd-induced damage, as chlorophyll production, crucial for photosynthesis, is compromised by Cd toxicity, impairing the photosynthetic process (Parmar et al., 2013; Li et al., 2010). Cd stress disrupts mitochondrial function in plants by disrupting redox balance and promoting the production of reactive oxygen species (ROS), which damage membrane lipids and alter metabolic activities (Chen et al., 2018; Huybrechts et al., 2020). The ROS produced under stress conditions are responsible for cellular oxidative damage and genotoxicity (Khan et al., 2022). Consequently, Cd, one of the most hazardous contaminants, requires particular attention to control its mobility in agricultural soils. Rice, a staple cereal crop for approximately 3.5 billion people globally (Dabral et al., 2019), is particularly vulnerable. A significant portion of Cd in the food chain originates from agricultural products, as Cd accumulates in soil plants via roots and enters the food supply, posing health risks to human immune, neurological, and reproductive systems (Parmar et al., 2013; Adil et al., 2020).
The utilization of organic amendments such as cattle manure, biochar, and compost represent an eco-friendly strategy for preventing Cd contamination (Gu et al., 2019; Hamid et al., 2020). However, these methods are often deemed impractical due to their associated costs and the potential introduction of additional pollutants (Pramanik et al., 2018). Vermicompost (VC), a nutrient-dense fertilizer, has emerged as a prevalent choice for rehabilitating metal-polluted agricultural soils (Wang et al., 2018; Zhang et al., 2020). VC is not only environmentally friendly but also a non-toxic amendment that enriches soil with essential nutrients and growth-promoting substances. In a systematic review by Oyege and Balaji Bhaskar (2023), it was reported that the supplementation of solo VC into agricultural systems enhances soil quality including enhanced permeability, aeration, drainage, and water-holding capacity and improves microbial biodiversity, ultimately boosting crop yield. Additionally, VC has been shown converting unavailable nutrients into available forms and supplies micro and macronutrients to plants, as well as it has elevated sulfur level than mineral fertilizer that can further enhance plant growth (Hoque et al., 2022; Shen et al., 2022). Alam et al. (2020) demonstrated that VC is superior to spent mushroom and organic fertilizers in mitigating the accumulation and uptake of Cd and other metals in plants. The maximum Cd2+ absorption rate of 170.70 mg g-1 by VC suggests its potential as an in-situ sorbent for Cd-treated soils (Zhu et al., 2017). In addition, VC is more effective than plant compost in reducing heavy metal levels in soil and its uptake in plants due to its maximum capacity, high specific area, strong cation exchange capacity, and enrichment in the active structural group (Li et al., 2021). Furthermore, it application can influence soil physical and biochemical properties, altering the chemical speciation of Cd in the soil (Wang et al., 2018) and increased the soil pH, thereby decreasing Cd bioavailability (Rafiq et al., 2014). Cd availability is sensitive to soil pH and exhibits a negative correlation with it. In alkaline conditions, Cd is present as CdHCO3+ or CdCO3 forms that are less bioavailable (Sauvé et al., 2000; Khaokaew et al., 2011; Shahid et al., 2017). Post-application, VC contributes polysaccharides, and mucilage from earthworms and microbes, and enhances soil physical structure, including aeration, porosity, aggregate stability, and drainage, all of which promote crop root development and nutrient uptake (Lim et al., 2015). VC is also a substantial source of both micronutrients and macronutrients for plants, leading to improved soil mineral content and increased plant growth and yield (Maji et al., 2017; Dubey et al., 2020). However, there is a lack of research assessing the impact of VC on paddy soil characteristics, the uptake of Cd by fragrant rice, the physiological and oxidative stress defense mechanisms, and rice yield under Cd stress.
This study explored the potential of VC as a soil conditioner for remediating Cd-polluted soils. The research focused on aromatic rice cultivars MXZ-2 and XGZ, which are popular in southern China for their desirable taste and flavor (Luo et al., 2020; Zhang et al., 2022). As a semi-aquatic tropical species grown in flooded fields, rice is particularly susceptible to Cd uptake and accumulation in its tissues (Wu et al., 2014). The main objectives of this study were (1) to investigate the effect of VC on the chemical properties of paddy soil, plant physiological attributes, and grain yield under Cd stress, (2) to determine the influence of VC on Cd accumulation and biochemical parameters in aromatic rice, including N metabolism related enzyme activities, N assimilation, and grain 2-acetyl-1-pyrroline (2AP) content, and (3) to elucidate the relationship between soil fertility, plant physiological processes, N assimilation, grain yield, and 2AP production in aromatic rice. We hypothesized that VC application could enhance soil health, reduces Cd accumulation in aromatic rice, thereby improve growth, quality, and yield. The findings are anticipated to contribute to establish a conceptual framework for safe and sustainable crop production in Cd-contaminated agricultural lands.
A pot experiment was conducted at South China Agricultural University in Guangzhou, China. The paddy soil from the specified rice field (0-15 cm depth) exhibited mild acidity, with a pH of 5.88. Additionally, the soil contained 0.93 g kg-1 of total phosphorus (TP), 140.45 mg kg-1 of available N (AN), and 1.19 g kg-1 of total N. Further details regarding the soil characteristics are provided in Supplementary Table S1.
In the current study, two aromatic rice varieties i.e., MXZ-2 and XGZ, showed differential responses to Cd stress were used. These cultivars were collected from the College of Agriculture, South China Agricultural University. The experiment was conducted using a complete block design having three replications during the late growing season (July-November) of 2023. Each plastic pot was filled by 15 kg of paddy soil collected from an unpolluted rice field at a depth of 15 cm. Four levels of VC were tested: VC1 = 0, VC2 = 2 t ha-1, VC3 = 4 t ha-1, and VC4 = 6 t ha-1, in conjunction with two Cd doses (Cd1 = 0 and Cd2 = 25 mg Cd kg-1 soil). The experiment comprised eight treatments: (1) Cd1VC1 = 0 Cd + 0 VC, (2) Cd1VC2 = 0 Cd + 2 t ha-1 VC, (3) Cd1VC3 = 0 Cd + 4 t ha-1 VC, (4) Cd1VC4 = 0 Cd + 6 t ha-1 VC, (5) Cd2VC1 = 25 mg Cd kg-1 soil + 0 VC, (6) Cd2VC2 = 25 mg Cd kg-1 soil + 2 t ha-1 VC, (7) Cd2VC3 = 25 mg Cd kg-1 soil + 4 t ha-1 VC, and (8) Cd2VC4 = 25 mg Cd kg-1 soil + 6 t ha-1 VC. Cd and VC were thoroughly mixed before seedling transplantation. The Cd treatment concentration was chosen based on previous studies in our lab (Kanu et al., 2017; Imran et al., 2021). The Cd used were CdCl2.2.5H2O, which was purchased from Sigma Aldrich, China. The seeds of two fragrant rice cultivars were cultivated in a plastic-trays, with each tray containing three hills. After 24 d, the uniform-sized seedlings were then transplanted into pots in mid-July, and the rice crops were harvested in November. The NPK fertilizer application rate was 300:150:300 (kg ha-1), with 1.80 g of N as urea, 0.90 g of P2O5 as superphosphate, and 2.20 g of KCl used. Uniform flooding irrigation was maintained from the time of seedling planting until physiological maturity.
A core sampler was utilized to extract soil samples from a depth of 15 cm, before the planting of seedlings and post-harvest. The collected soil samples were subsequently divided into two distinct parts: one portion was allocated for soil nutrient assessment, while the other was designated for molecular analysis (data are not provided in this study) and preserved at -80°C. Soil organic carbon (SOC) was analyzed using the oxidation method with K2Cr2O7-H2SO4, as described by Wang et al. (2003). For TN analysis, 200 mg soil samples were weighted using salicylic acid-sulfuric acid hydrogen peroxide according to Ohyama (1991), and finally, TN was calculated according to the micro-Kjeldahl technique recommended by Jackson (1956). In addition, soil pH, AN, total P, and total K were measured using the techniques detailed by Lu (2000).
On the seventh day of the heading stage, gas exchange attributes, such as transpiration rate (Tr) and net photosynthesis (Pn) were measured at sunny conditions using a portable photosynthesis system (LI-6800 System; Li-COR) to assess leaf physiological activity. In addition, the activity of NiR and NR enzymes in rice spikelets tissue was quantified employing the NIR-2-G and NR-2-Y assay kits, respectively, supplied by Biotechnology Co. Ltd., China. Additionally, GS and GOGAT enzyme activities in spikelets tissue were determined using the GS-2-Y and GOGAT-2-Y assay kits from the same manufacturer.
The leaves samples were homogenized in distilled water, cooked for 15 minutes, and filtered to determine NO3- and NH4+ levels. The NO3- concentrations in leaves were determined using a previous approach (Cataldo et al., 1975). Whereas, the NH4+ content was measured using the Nessler reagent method (Molins-Legua et al., 2006). In addition, total N concentration was determined by using Kjeldahl method, as described by Barbano et al. (1990).
The soluble protein was measured in leaves by bovine serum albumin according to Bradford technique (Bradford, 1976). In addition, the proline content in leaves tissues was calculated according to the procedure of Bates et al. (1973). Total amino acid content in rice leaves was measured as described in the previous procedure (Barbano et al., 1990).
The shoot and grain samples were dried, ground, and subsequently processed using a 4:1 (v/v) mixture of HNO3- and HClO4. Following dilution to a final volume of 25 mL, Cd levels in these tissues were analyzed using a flame atomic absorption spectrometer, by the methodology previously described by Cao et al. (2014).
The 2AP levels in grains of two fragrant rice cultivars were determined by a synchronized distillation and extraction procedure and a Gas Chromatograph Mass Spectrometer according to the procedure of Mo et al. (2019).
Two fragrant rice cultivars were tested for grain yield and yield features. The rice grains were sun-dried to a moisture level of 12-14%. During the reproductive stage, the number of productive tillers was recorded. To determine the thousand-grain weight, 1000 rice grains were taken and weighed.
All the data were collected in Microsoft Office Excel 2023 and all the data were showed as mean + standard errors. The collected data were subjected to one-way analysis of variance (ANOVA) for completely randomized designs, using Statistix 8.1 software (Analytical Software, Tallahassee, FL, USA). Before statistical analysis, the results were normalized using the arcsine transformation. Significant differences among the groups means for variables significantly affected by experimental factors were determined by Tukey’s HSD test at P < 0.05. Figures were prepared with OriginPro 2021.
The application of VC significantly (p < 0.05) improved various soil attributes such as TN, AN, SOC, pH, TK and TP, in soil treated with Cd- (, as detailed in Table 1. The VC supplementation alleviated the adverse effects of Cd on paddy soil health, with the most pronounced effect observed in all evaluated parameters at high VC amendments. Among the treatments, pots without Cd treatment (Cd1 + VC3) had higher values of soil quality attributes (i.e., pH, TN, AN, and SOC), whereas the soloCd2 pots had the lowest values. In comparison to the Cd2 alone treatment, the VC amendment treatments (Cd2 + VC3) enhanced the soil SOC, AN, TN and pH by 37.45%, 42.98%, 23.95% and pH 23.95% respectively. Likewise, lower VC inputs also increased each examined parameters, albeit to a lesser extent than the corresponding VC amendments under Cd stress.
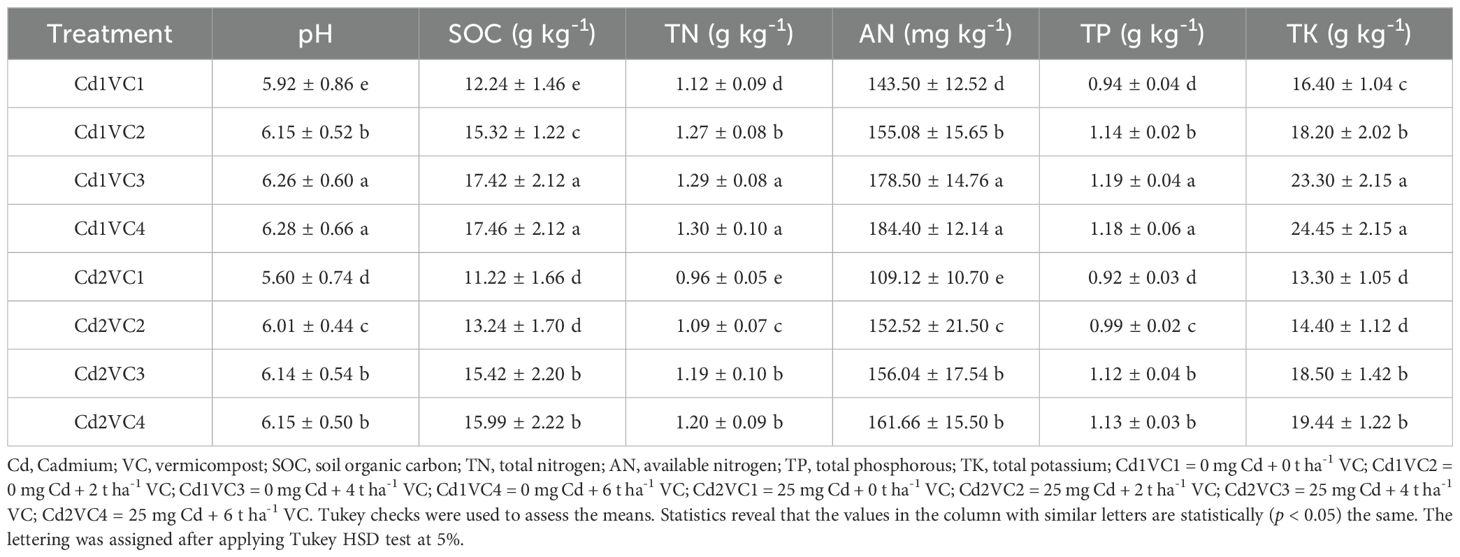
Table 1. Influence of VC additions on soil fertility under Cd stress condition.
The study examined the impact of VC application on the photosynthetic parameters of two fragrant rice varieties, such as XGZ and MXZ-2, under Cd stress condition. Significant variations in photosynthesis were observed, as shown in Figure 1. In plants subjected to Cd stress, VC supplementation improved photosynthetic parameters, including net Pn and Tr, during the reproductive phase. Both cultivars exhibited a comparable response to the treatments. Specifically, the Cd2 + VC3 treatment led to an 86.75% increase in Pn and a 102.20% increase in Tr for MXZ-2, and a 70.05% and 81.22% increase for XGZ, respectively, compared to the Cd2 + VC1 treatment, as illustrated in Figure 1. Additionally, low doses of VC significantly enhanced physiological activity in the leaves of fragrant rice under Cd treatment conditions.
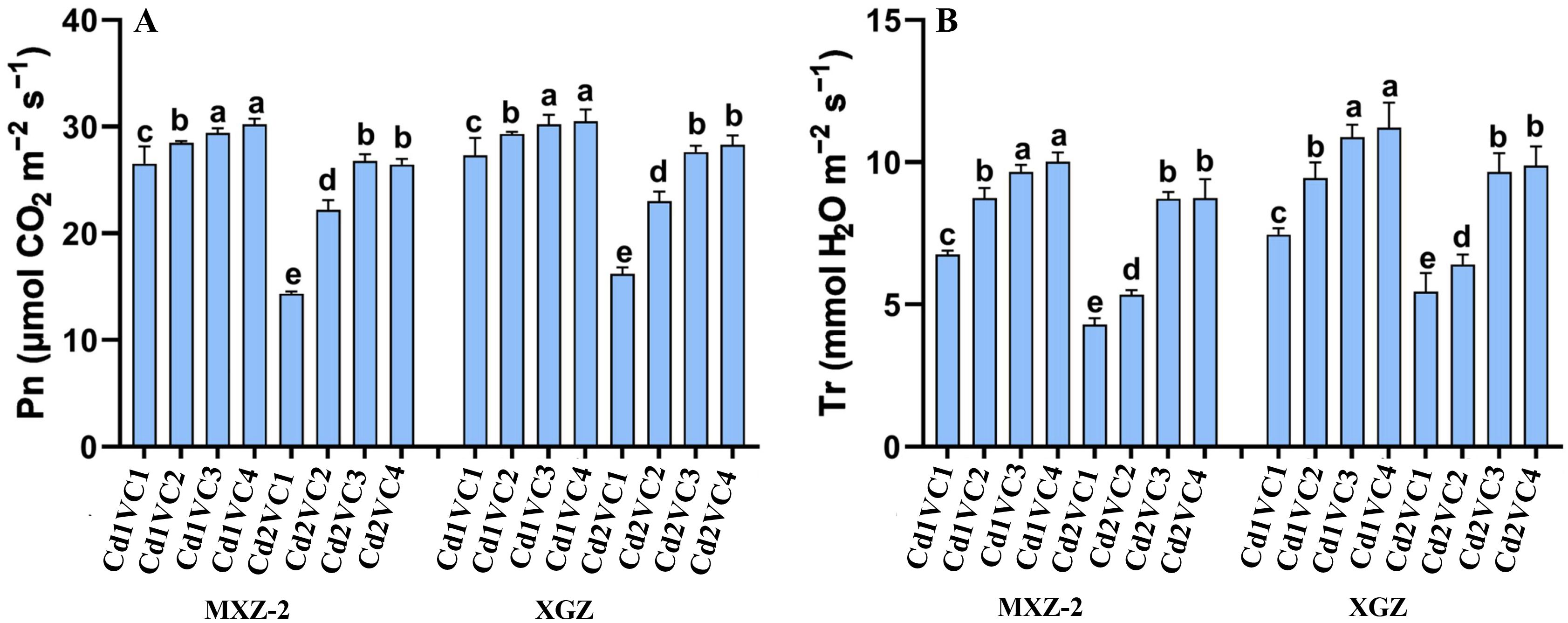
Figure 1. Effect of VC application on Pn (A) and Tr rate (B) of two fragrant rice cultivars (MXZ-2 and XGZ) at heading stage subjected to Cd treatment. Tukey HSD test were applied to relate the means of the treatments. Bars with distinct letter combinations show significant differences at (P < 0.05). See Table 1 for treatment details.
The study examined the impact of VC supplementation on N assimilation and accumulation in plants subjected to Cd toxicity (Figure 2). Enzyme activities associated with N metabolism, including NiR, NR, GOGAT, and GS, were notably diminished under Cd treatment compared to non-Cd exposed plants. Cultivar variations were observed, with XGZ showing a less pronounced decline in enzyme activities, suggesting greater tolerance to Cd stress. Interestingly, VC supplementation mitigated Cd treatment, particularly under high VC treatments, and enhanced the activities of N metabolism enzymes in leaves. Both cultivars exhibited a similar response pattern to the treatments. Specifically, the Cd2 + VC3 treatment led to a substantial increase in NR (95.66% and 90.05%), NiR (62.15% and 50.50%), GS (61.10% and 49.66%), and GOGAT (58.45% and 60.78%) activities in MXZ-2 and XGZ, respectively, compared to plants treated with Cd alone. Furthermore, plants treated with lower concentrations of VC displayed significantly higher N metabolism enzyme activities than those exposed solely to Cd treatment.
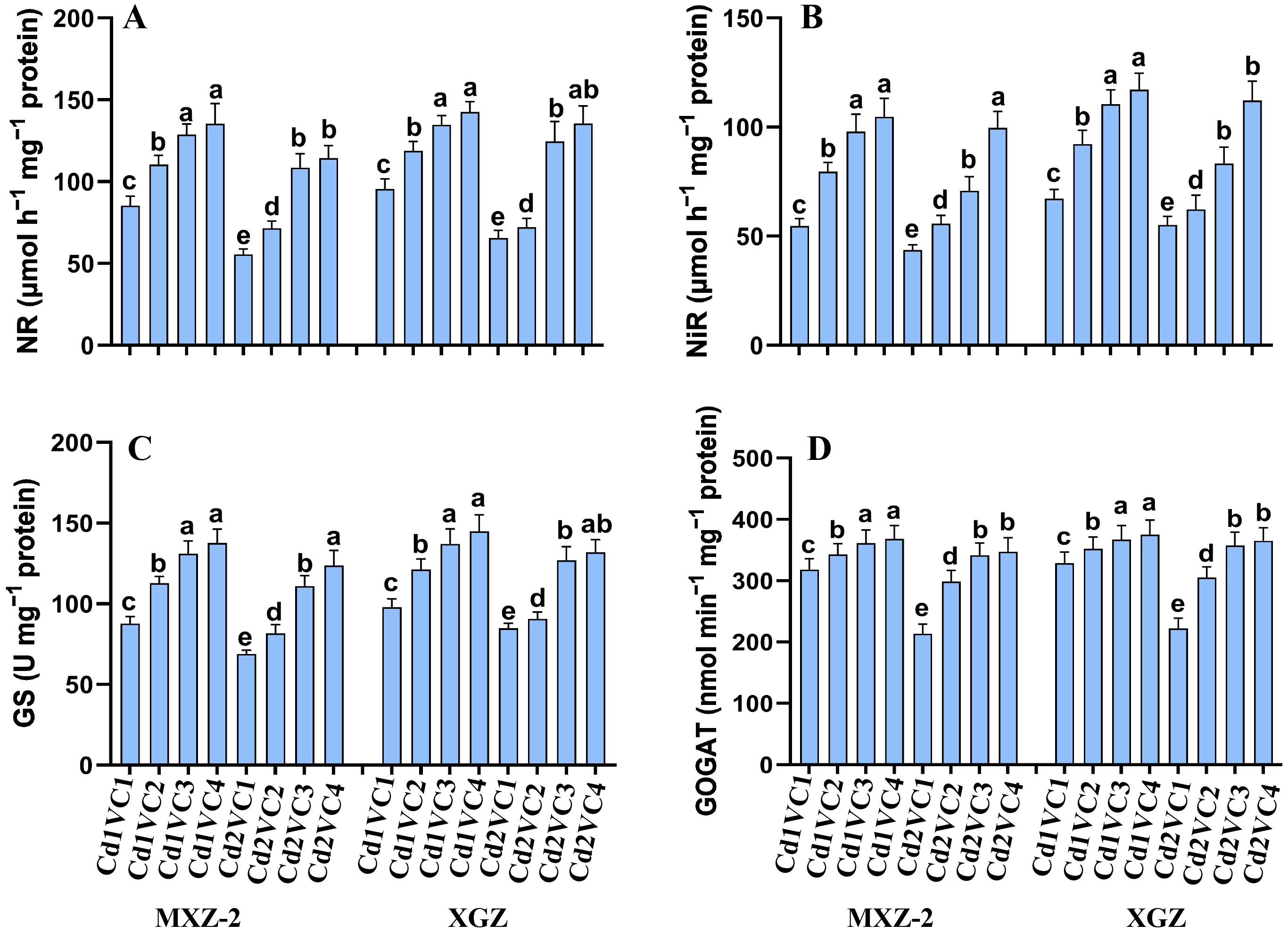
Figure 2. Effect of VC on N metabolism enzyme activities, (A) nitrate reductase, (B) nitrite reductase, (C) glutamine synthetase, and (D) glutamate oxoglutarate aminotransferase in spiklets of two fragrant rice cultivars (MXZ-2 and XGZ) subjected to Cd treatment. Tukey HSD test were applied to relate the means of the treatments. Error bars are standard errors of the mean. At P < 0.05, bars with distinct letter combinations show significant differences. See Table 1 for treatment details.
The study revealed that Cd stress significantly reduced the levels of NO3-, NH4+, and TN content in aromatic rice cultivars, as depicted in Figure 3. Conversely, the supplementation of VC mitigated the detrimental impact of Cd stress on rice plants, enhancing both inorganic N and total N content. Notably, both cultivars exhibited a comparable response to the treatments. The lowest concentrations of NO3-, NH4+, and TN were observed in plants subjected to the Cd2 + VC1 treatment. Among the treatments, the Cd2 + VC3 treatment led to a substantial increase in NO3- (74.44% and 58.05%), NH4+ (65.50% and 88.66%), and plant TN content (147.50% and 123.85%) in MXZ-2 and XGZ, respectively, compared to plants treated with Cd2 + VC1. Furthermore, plants in pots treated with lower concentrations of VC demonstrated significantly higher N metabolism enzyme activity than those subjected to Cd treatment alone.
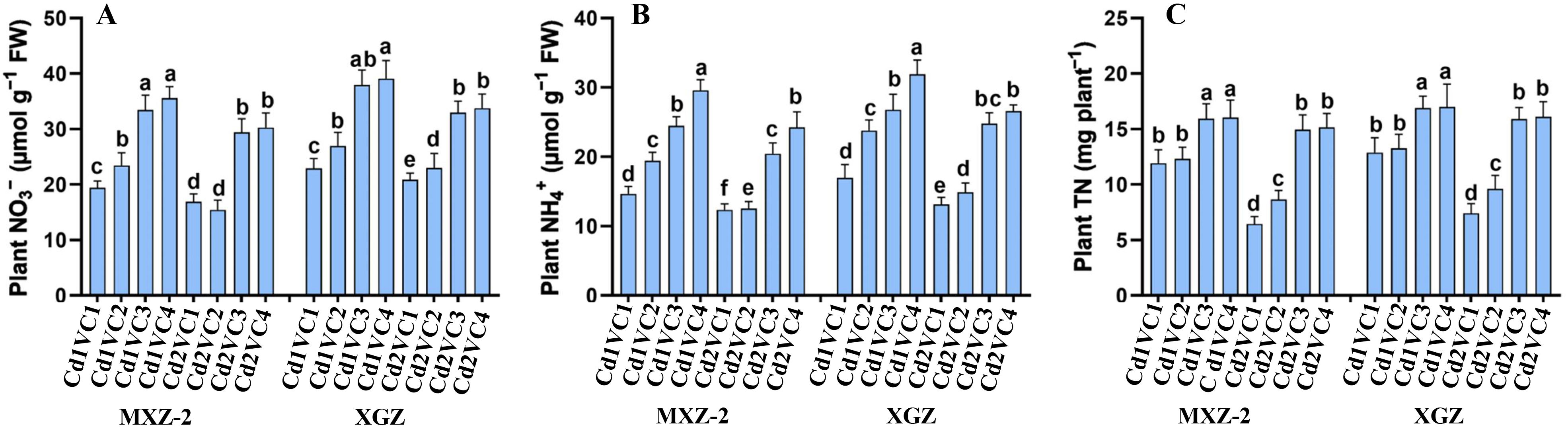
Figure 3. Effect of VC application on plant inorganic N and total N content such as NO3- (A), NH4+ (B) and TN (C) in leaves of two fragrant rice cultivars (MXZ-2 and XGZ) subjected to Cd treatment. Tukey HSD test were applied to relate the means of the treatments. Error bars are standard errors of the mean. At P < 0.05, bars with distinct letter combinations show significant differences. See Table 1 for treatment details.
The production of soluble proteins, proline, and amino acids in leave were varied significantly between cultivars under Cd treatment when treated with VC, as indicated in Table 2. In Cd-treated plants, proline levels were markedly elevated compared to those in non-Cd-treated plants. However, VC application mitigated Cd-induced stress and decreased proline synthesis. Specifically, proline content was decreased by 30.74% and 23.25% in the XGZ and MXZ-2 cultivars, respectively, in the Cd2 + VC3 treatment compared to the Cd2 + VC1 treatment. Similarly, a lower concentration of VC significantly reduced proline content in Cd-treated plants. Moreover, the supplementation of VC alleviated Cd stress in fragrance rice, significantly (p < 0.05). For instance, compared to the Cd2 + VC1 treatment, the Cd2 + VC3 treatment increased leaf total protein and amino acid content by 25.95% and 29.62% in XGZ, and by 68.70% and 49.50% in MXZ-2, respectively. Additionally, the results indicated that MXZ-2 exhibited lower proline and soluble protein content than XGZ, suggesting that MXZ-2 may be more susceptible to stress conditions.
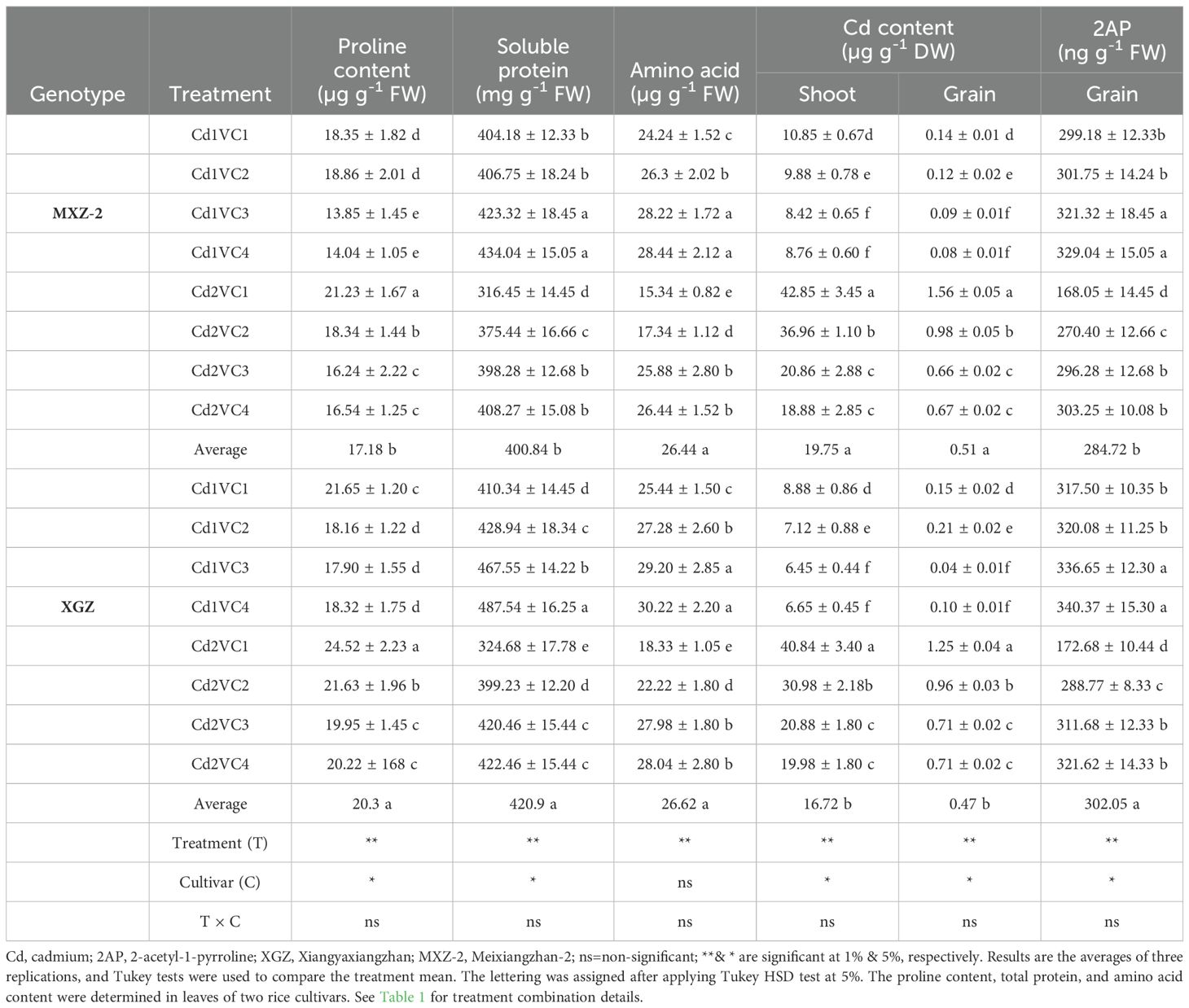
Table 2. Influence of VC on fragrant rice biochemical attributes and Cd accumulations under Cd stress condition.
The uptake and accumulation of Cd in the shoots and grains of two fragrant rice varieties were significantly elevated under Cd-treated soil conditions (Table 2). However, the application of VC declined Cd-related toxicity and substantially diminishes Cd uptake in rice tissue. The Cd content in rice tissue exhibits the following trend: shoot > grains.
High level of VC amendments, particularly Cd1 + VC4, resulted in the lowest Cd accumulation in rice tissue. The application of VC significantly decreased Cd concentrations in both leaves and stems compared to the Cd2 treatment alone. Specifically, Cd2 combined with VC3 decreased Cd accumulation by 105.50% in the shoots and 136.66% in the grains of the MXZ-2 rice cultivar, while in the XGZ cultivar, reduction were 95.55% in shoots and 100.20% in grains. These data indicate that a high dosage of VC significantly diminishes Cd accumulation in rice plants. Furthermore, the results suggest that the XGZ cultivar exhibits greater resistance to Cd than the MXZ-2 cultivar.
The Cd-stressed plants significantly declined the 2AP content in rice grains. However, VC supply mitigated the Cd-induced toxicity and significantly improved the grain 2AP content (Table 2). Of the treatments, higher 2AP content was observed in the Cd1 + VC4 treated plants. All the treatments followed a similar pattern for both cultivars. Whereas, the lowest 2AP content in grains was noted in the Cd2 treatment only. Relative to alone Cd2 treatment, high VC supplementation (VC3) treatment into Cd-stressed plants enhanced the 2AP content by 76.50% and 80.46% in XGZ and MXZ-2 cultivars, respectively. Similarly, low VC treatment also increased the 2AP content in rice grains under Cd stress condition.
The Cd treatment significantly decreased the yield and yield traits of both varieties (Table 3). Nevertheless, VC additions counteracted the Cd stress and significantly improved grain yield and yield attributes. Interestingly, the treatments followed a similar pattern for both cultivars, and higher grain yield was noted in the Cd1 + VC4 treatment. Related to Cd-only plants, VC input treated plants, i.e., Cd2 + VC3 enhanced the productive tillers, thousand-grain weight, and yield by (53.22% and 49.50%) and (60.05%, and 77.55%) in XGZ and MXZ-2 cultivars, respectively. Similarly, other low VC doses also increased grain yield and yield attributes in a Cd-treated condition.
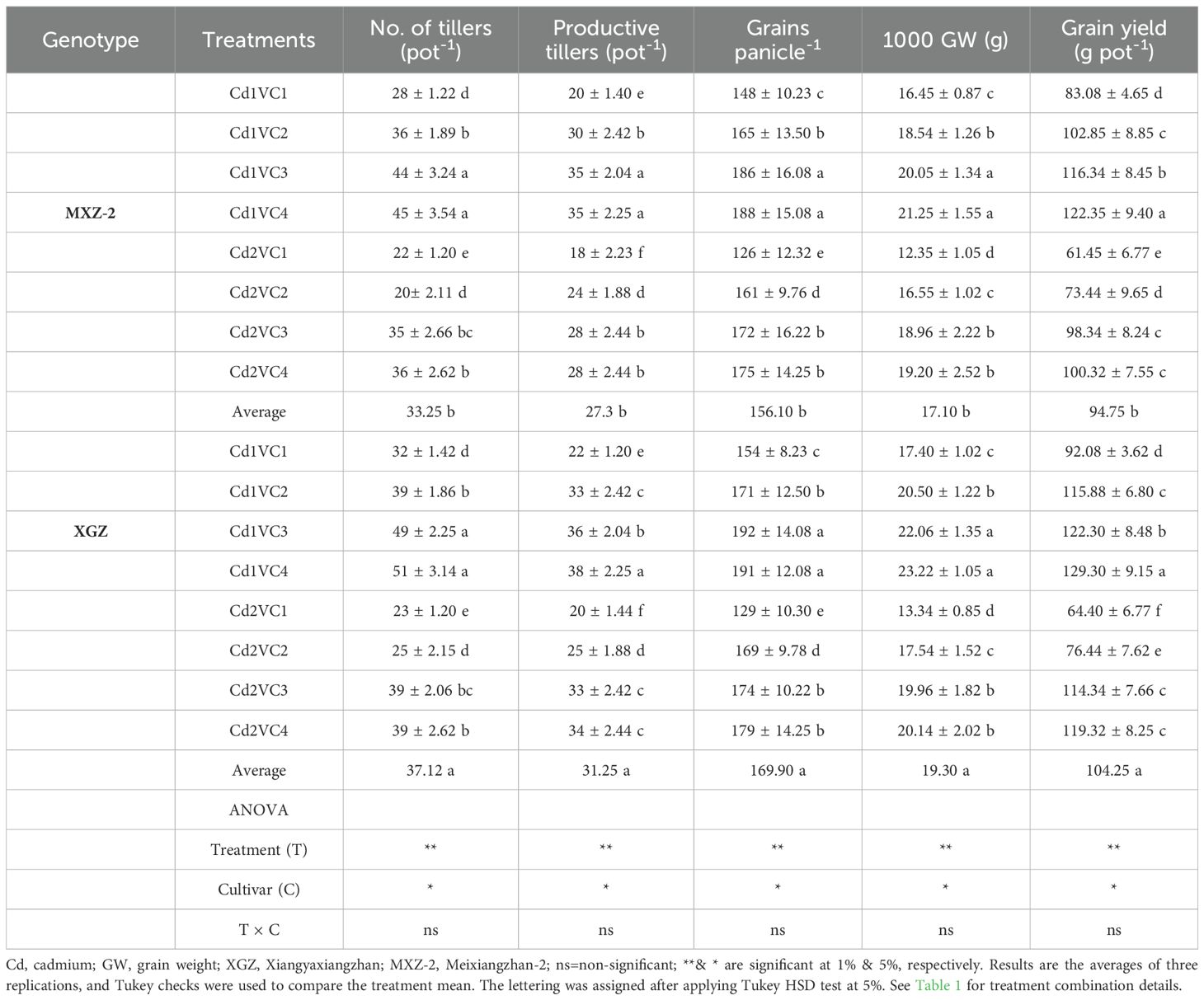
Table 3. Influence of VC supply on rice cultivars on yield and yield traits in a Cd-stressed soil.
Heavy metals, specifically Cd, often disrupt soil fertility, reduces soil pH and affect variety of morphological, and physio-biochemical characteristics of plants including growth, photosynthesis, N assimilation, and accumulation (Hussain et al., 2020). Therefore, mitigating Cd stress in soil and its impact on plant growth, and development is a crucial objective for plant researchers. One effective and environmentally friendly approach is in situ stabilization, which involves immobilizing Cd through the application of organic fertilizers such as cattle dung, VC, and biochar (Hamid et al., 2020; Iqbal et al., 2024). In this study, we investigated the effect of VC application on the chemical properties of paddy soil, as well as the physiological and biochemical characteristics of plants, 2AP content in grains, and the yield of fragrance rice grown under Cd stress condition.
The current investigation, reveals that the application of VC significantly improves the quality of soil affected by Cd stress. Our findings suggest that increasing the biodegradation rate of VC enhances soil quality by gradually releasing essential nutrients for plant uptake. We observed a notable increase in soil pH with the addition of VC compared to untreated and Cd-treated soil. In contrast, Ni et al. (2018) noted that nitrification, resulting from the heavy use of synthetic N fertilizers, produces hydrogen ions (H+), which subsequently reduces soil pH. The acidic nature of synthetic N may contribute to this pH reduction, as posited by Adekiya et al. (2020). Our study indicates that Cd2 alone treatment also enhanced the soil acidity, compared to VC application treatments. These differences in Cd accumulation in rice grains in different soils may be attributed to disparities in Cd phytoavailability, which is influenced by soil pH. Specifically, lower soil pH levels correlate with increased Cd accumulation in rice grains, and conversely, higher pH levels result in reduced Cd uptake (Rafiq et al., 2014). Furthermore, Ok et al. (2011a; b) observed similar trends in rice cultivated in Cd-contaminated soils where amendments that altered soil pH were applied. In line with our findings, previous studies have also noted low pH levels in the presence of Cd toxicity (Jin et al., 2020). Conversely, the addition of organic N has been shown to substantially decrease soil acidity (Iqbal et al., 2019). Correspondingly, our research indicates that VC supplementation markedly increases soil pH. Hydroxyl ions (OH-) are generated from charged functional groups in organic additives and the hydrolysis of CaCO3 interacts with H+ ions, thereby increasing the soil pH. These hydroxyl ions encompass phenolic, hydroxyl, and carboxyl groups (Gul et al., 2015). The majority of plant species thrive in soil with a pH adjusted to near neutrality by the application of VC (Fernandez-Bayo et al., 2009).
The improvements in soil chemical properties observed with high-VC treatments in this study can be attributed to the compost rich organic matter content and its variety of essential plant nutrients (Tejada et al., 2010). Liang et al. (2017) noted that metals generally do not easily dissolve or move through soil with high pH. Consequently, the elevated pH of the soil following VC fertilizer application may have significantly contributed to the retardation of Cd migration in the present experiment, and the enhanced mineral content of the soil promotes robust plant growth. Furthermore, the improved mineral content of the soil ensures vigorous plant growth. Lim et al. (2015) reported that the VC application promotes the secretion of mucus by earthworms, polysaccharides, and microorganisms, which enhance the soil’s physical structure and are crucial for plant root development and nutrient absorption. These physical improvements encompass increased porosity, improved drainage, and aeration, and enhanced aggregate stability. Additionally, the soil chemical properties were positively correlated with plant physiological and biochemical characteristics, as well as the yield of fragrance rice grains (Figure 4). This analysis indicated that the enhancements in soil quality are directly linked to plant growth and development. In summary, the application of VC can enhance soil fertility and crop production while also mitigating the mobility of Cd in paddy soils.
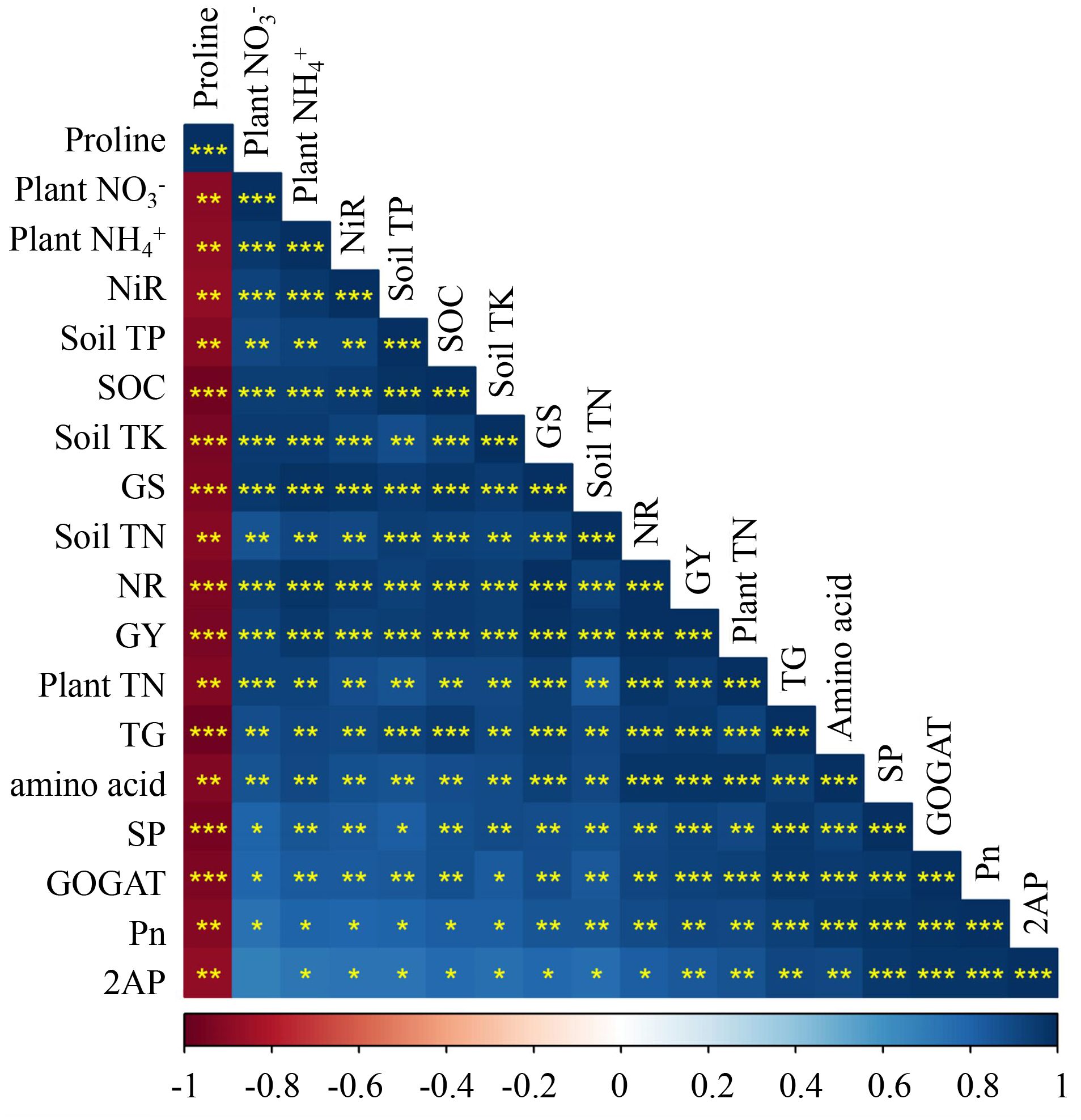
Figure 4. The correlation analysis between soil chemical traits, plant physiological, biochemical, and grain yield of rice under Cd toxicity to the application of different VC amendments. 2AP (2-acetyl-1- pyrroline), Pn (net photosynthesis rate), NR (nitrate reductase), NiR (nitrite reductase), GS (glutamine synthetase), GOGAT (glutamate oxoglutarate aminotransferase), SP (soluble protein), TN (total nitrogen), GY (grain yield), TK (total potassium), TP (total phosphorous). Asterisks (*), (**), and (***), indicate significant differences at P < 0.05, P < 0.01 and P < 0.001 respectively.
Photosynthesis is the main element of plant physiological activity and productivity by enhancing crop growth and biomass accumulation (Khan et al., 2017). In this experiment, the VC enhanced the plant’s photosynthetic efficiency compared to the Cd stress environment (Figures 1, 2). The enhancement in leaf photosynthetic activity induced under VC application could be primarily attributed to the boosted soil fertility in the current work (Table 1), faster release of soil nutrients from VC in the early growth stages and gradual and slow release of crop-related nutrients from VC throughout the crop period (Yang et al., 2015; Iqbal et al., 2021). Photosynthesis experienced a strong reaction to water and soil health (Makoto and Koike, 2007). An adequate supply of water and nutrients reduces the concentration of water-soluble nutrients and mitigates the root-derived stress signal abscisic acid (ABA), leading to the opening of leaf stomata and enhancing their water potential and physiological activity (Daszkowska-Golec and Szarejko, 2013). Additional evidence is provided by the present study linear regression analysis, which indicates a highly significant correlation between soil chemical properties and leaf photosynthetic activity (Figure 4).
NO3- and NH4+ are the primary forms of N that plants use for growth activities, and N uptake and assimilation is a vital metabolic activity that not only controls crop growth and development but also plays a critical role in crop survival in a polluted soil (Imran et al., 2019). In N assimilation, NO3- is converted into NO2- and NH4+ by the sequential activity of N metabolizing enzymes, NR and NiR, whereas NH4+ is directly converted into amino acids and protein by the concerted actions of GS and GOGAT (Balotf et al., 2016). In the current study, solo supplementation of VC increased the NO3-, NH4+ and total N maybe because VC application significantly influence the essential nutrient particularly N and P in the soil (Ramazanoglu, 2024). The application of VC has been shown to significantly influence plant growth through its substantial impact on key soil properties. Specifically, VC has been found to enhance processes such as mineralization and nitrification, thereby increasing the availability of essential mineral nutrients to plants compared to other organic fertilizers (Karlsons et al., 2016). Furthermore, studies indicated that the application of VC results in elevated levels of soil organic matter, microbial activity and N concentration, the latter being a critical nutrient for plant growth (Akhzari and Pessarakli, 2017; Erdal and Ekinci, 2020; Iqbal et al., 2024). These findings collectively underscore the potential of VC as a potent soil amendment for improvement of plant physiological and enzymatic activities. In contrast, Cd stress considerably reduced NR and NiR enzyme activities in plants (Figure 2). A possible explanation for this is that the Cd stress significantly reduced the soil fertility and NO3- content in rice leaf tissues (Table 1, Figure 3). However, the VC addition increased the N metabolism enzyme activities, including NR and NiR. Interestingly, in the current work, VC supply also increased the GOGAT and GS activity and simultaneously enhanced amino acid and SP contents in Cd-stressed plants, leading to Cd tolerance mechanism (Table 2). The main reason for the improvements in N metabolism activities was mainly due to enhanced plant physiological activity in Cd-contaminated soils in the presence of VC.
In this work, Cd stress considerably reduced the total N in both cultivars (Figure 3). The main reason might be due to significantly decreased NO3- level and N assimilation, N enzymes activities in the current study under Cd stress condition. Similarly, earlier research also stated that Cd stress reduced N accumulation (Campbell, 2001; Hussain et al., 2020). However, the VC supply decreased the side effects of Cd stress on soil health and crop growth and development and significantly (p < 0.05) increased the soil nutritive status and plant physio-biochemical activity under Cd-stressed conditions. This improvement in soil fertility and plant growth and physiological activity ultimately increased plant N uptake and assimilation in rice, and these VC-related side effects are nearly to be accomplished by enough NO3- accumulation, upgrading the activity of N assimilation enzymes and accumulating nitrogenous compounds (i.e., amino acid and proline) in this study. Similarly, Sun et al. (2012), also stated that several N metabolism enzymes play a key part in plant N accumulation. In this work, the greater activity of N assimilation enzymes in VC-added plants. The N metabolism enzymes, i.e., GOGAT and GS in the reproductive stage were positively correlated to N assimilation and grain yield and quality. Ceusters et al. (2019), reported that soil N availability has a strong relationship with plant N uptake and enhanced N metabolism enzyme activities. Furthermore, the Cd stress reduced the 2AP level and yield of cultivars, while VC alleviated the Cd stress and enhanced the 2AP level and yield of fragrant varieties (Table 2). A possible explanation is that Cd stress reduced N metabolic activity and thus reduced plant TN content in different organs. However, VC amendments counteracted the Cd-related inhibitory effect on N assimilation and utilization by enough N uptake and strengthened N assimilation enzyme activities. In addition, the plant physiological and biochemical attributes were highly positively correlated with 2AP production in grains of both fragrant rice cultivars (Figure 4). This analysis revealed that the improvement in rice physiological attributes are closely linked to grain quality. In conclusion, the application of VC can improve the soil fertility and plant physicochemical attributes, which in turn positively impacts crop quality. Our findings are also in agreement with the previous findings indicating a strong positive correlation between a plant physiological and biochemical traits and both 2AP and grain yield of rice (Yang et al., 2012; Mo et al., 2019).
The cytoplasm of plants contains proline, a compound that modulates osmotic pressure by adjusting cellular water potential (Muneer et al., 2011). Our findings indicate that under Cd stress, the proline concentration in rice leaves significantly increases (Table 2). Elevated proline levels in plants are commonly associated with heavy metal stress, particularly Cd, as stressed plants exhibit increased resilience (Bauddh and Singh, 2012). The degradation of plant proteins may correlate with heightened proline levels, which could serve as an indicator of plant tissue damage (Elmer and White, 2018). Our results are consistent with Elmer and White (2018), who reported that increased protease activity leads to protein deficiency in response to Cd stress. However, the application of VC significantly enhanced protein content and 2AP in plant grains, counteracting the adverse effects of Cd stress (Table 2). Our study suggests that the addition of organic amendments to soil enhances its fertility, thereby promoting plant physiological and biochemical processes through the improved uptake and accumulation of essential nutrients. Furthermore, VC application reduced leaf proline content and bolstered plant defense mechanisms by enhancing physiological activity, demonstrating a mitigating effect on maintaining osmotic balance in plants exposed to Cd pollution.
The current study demonstrates that the application of VC significantly reduces Cd uptake in plant organs such as shoots and grains, as shown in Table 2. This effect is primarily attributed to VC’s ability to decrease Cd’s accessibility and mobility. VC acts as a soil conditioner by promoting metal precipitation and complexation, thereby providing plants with nutrients and organic matter while concurrently diminishing the mobility and bioavailability of metals in the soil (Deng et al., 2017). Furthermore, due to its elevated cation exchange capacity and surface area, VC is considered a promising agent for stabilizing heavy metals in soil (Wang et al., 2018; Ding et al., 2021). Additionally, Wan et al. (2020) observed that the supplementation of organic fertilizers led to a substantial reduction in Cd levels in rice grains, ranging from 7.8 to 79.3%. Similarly, Tang et al. (2015) found that the addition of organic amendments reduced heavy metal content in the roots and shoots of plants.
Cd stress significantly reduced the yield and yield components in both cultivars examined. Conversely, the application of VC markedly enhanced rice yields and yield components, as shown in Table 3. Enhancements in crop production are closely associated with enhanced soil fertility (Iqbal et al., 2022, 2024). Organic fertilizers improve soil quality and fertility, thereby promoting plant growth, development, quality, and yield (Iqbal et al., 2022). In this study, nutritional values of the soil were increased upon application of VC into the soil (Table 1), which facilitated aromatic physiological activity, growth, and yield by providing essential nutrients during the growth phase. This was corroborated by the correlation analysis conducted, which indicated that soil quality parameters were highly positively correlated with the plant’s physiological and biochemical attributes, ultimately influencing yield and yield characteristics (Figure 4). Iqbal et al. (2022) demonstrated that variations in rice yield are closely associated with soil fertility. In summary, the use of VC may enhance soil fertility and plant physical-biochemical attributes, directly impacting and improving crop productivity.
Our study offer novel insights into how VC supplementation enhances 2AP content and grain yield in rice under Cd stress by improving N assimilation and utilization. We found that Cd stress significantly impeded soil fertility and plant physiological processes, including the activities of N metabolism related enzymes (NR, NiR, GS, and GOGAT), as well as 2AP production and grain yield in rice. However, the application of VC effectively counteracted the Cd-induced decline in soil fertility and plant physio-biochemical characteristics, resulting in improved 2AP levels and grain yield in both rice cultivars studied. The results indicated that VC application immobilized Cd in paddy soil, enhancing soil quality and reducing Cd accumulation in rice tissues (shoots and grains), while also decreasing the uptake of essential nutrients from the soil. Our findings suggest that VC supplementation enhances plant physiological efficiency and biochemical status by mitigating the deleterious effects of Cd on plant health, potentially through decreased Cd uptake and improved leaf photosynthetic efficiency, soluble protein levels, and regulation of nitrogenous compounds such as amino acids and proline. As a result, our study demonstrates that VC amendments can improve N uptake and assimilation, leading to increased 2AP levels and grain yield in rice under Cd stress conditions.
The original contributions presented in the study are included in the article/Supplementary Material, further inquiries can be directed to the corresponding author/s.
AI: Conceptualization, Data curation, Formal analysis, Investigation, Methodology, Visualization, Writing – original draft. XC: Validation, Writing – review & editing. RK: Data curation, Investigation, Writing – review & editing. MaZ: Formal analysis, Methodology, Writing – review & editing. AK: Methodology, Writing – review & editing. MK: Writing – review & editing. ME: Writing – review & editing. AA: Formal analysis, Writing – review & editing. SL: Funding acquisition, Visualization, Writing – review & editing. XT: Funding acquisition, Resources, Supervision, Writing – review & editing. MuZ: Formal analysis, Methodology, Visualization, Writing – review & editing.
The author(s) declare financial support was received for the research, authorship, and/or publication of this article. This work was funded by the Guangzhou Science and Technology Projects (202103000075 and 202102100008), National Natural Science Foundation of China (No. W2433093) and Researchers Supporting Project (No. RSP2025R218), King Saud University, Riyadh, Saudi Arabia.
In addition, we thank the research station staff of SCAU for their cooperation in conducting this experiment.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fpls.2024.1481372/full#supplementary-material
Adekiya, A. O., Agbede, T. M., Ejue, W. S., Aboyeji, C. M., Dunsin, O., Aremu, C. O., et al. (2020). Biochar, poultry manure and NPK fertilizer: sole and combine application effects on soil properties and ginger (Zingiber officinale Roscoe) performance in a tropical Alfisol. Open Agric. 5, 30–39. doi: 10.1515/opag-2020-0004
Adil, M. F., Sehar, S., Chen, G., Chen, Z. H., Jilani, G., Chaudhry, A. N., et al. (2020). Cadmium-zinc crosstalk delineates toxicity tolerance in rice via differential genes expression and physiological/ultrastructural adjustments. Ecotoxicol. Environ. Saf. 190, 110076. doi: 10.1016/j.ecoenv.2019.110076
Akhzari, D., Pessarakli, M. (2017). Effects of vermicompost and urea fertilizers on qualitative and quantitative characteristics of vetiveria zizanioides stapf. grown under drought stress conditions. J. Plant Nutr. 40, 2063–2075. doi: 10.1080/01904167.2017.1346126
Alam, M., Hussain, Z., Khan, A., Khan, M. A., Rab, A., Asif, M., et al. (2020). The effects of organic amendments on heavy metals bioavailability in mine impacted soil and associated human health risk. Scientia Hortic. 262, 109067. doi: 10.1016/j.scienta.2019.109067
Balotf, S., Kavoosi, G., Kholdebarin, B. (2016). Nitrate reductase, nitrite reductase, glutamine synthetase, and glutamate synthase expression and activity in response to different nitrogen sources in nitrogen-starved wheat seedlings. Biotechnol. Appl. Biochem. 63, 220–229. doi: 10.1002/bab.2016.63.issue-2
Barbano, D. M., Clark, J. L., Dunham, C. E., Flemin, R. J. (1990). Kjeldahl method for determination of total nitrogen content of milk: collaborative study. J. Assoc. Off. Anal. Chem. 73, 849–859. doi: 10.1093/jaoac/73.6.849
Bari, M. A., Akther, M. S., Reza, M. A., Kabir, A. H. (2019). Cadmium tolerance is associated with the root-driven coordination of cadmium sequestration, iron regulation, and ROS scavenging in rice. Plant Physiol. Biochem. 136, 22–33. doi: 10.1016/j.plaphy.2019.01.007
Bates, L. S., Waldren, R. P. A., Teare, I. D. (1973). Rapid determination of free proline for water-stress studies. Plant Soil 39, 205–207. doi: 10.1007/BF00018060
Bauddh, K., Singh, R. P. (2012). Growth, tolerance efficiency and phytoremediation potential of Ricinus communis (L.) and Brassica juncea (L.) in salinity and drought affected cadmium contaminated soil. Ecotoxicol. Environ. Saf. 85, 13–22. doi: 10.1016/j.ecoenv.2012.08.019
Bradford, M. M. (1976). A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 72, 248–254. doi: 10.1016/0003-2697(76)90527-3
Bryant, R., McClung, A. (2011). Volatile profiles of aromatic and non-aromatic rice cultivars using SPME/GC–MS. Food Chem. 124, 501–513. doi: 10.1016/j.foodchem.2010.06.061
Campbell, W. (2001). Structure and function of eukaryotic NAD (P) H: nitrate reductase. Cell. Mol. Life Sci. 58, 194–204. doi: 10.1007/PL00000847
Cao, F., Wang, R., Cheng, W., Zeng, F., Ahmed, I. M., Hu, X., et al. (2014). Genotypic and environmental variation in cadmium, chromium, lead and copper in rice and approaches for reducing the accumulation. Sci. Total Environ. 496, 275–281. doi: 10.1016/j.scitotenv.2014.07.064
Cataldo, D. A., Maroon, M., Schrader, L. E., Youngs, V. L. (1975). Rapid colorimetric determination of nitrate in plant tissue by nitration of salicylic acid. Commun. Soil Sci. Plant Anal. 6, 71–80. doi: 10.1080/00103627509366547
Ceusters, N., Valcke, R., Frans, M., Claes, J. E., Van den Ende, W., Ceusters, J. (2019). Performance index and PSII connectivity under drought and contrasting light regimes in the CAM orchid Phalaenopsis. Front. Plant Sci. 10, 461163. doi: 10.3389/fpls.2019.01012
Chen, Q., Lu, X., Guo, X., Pan, Y., Yu, B., Tang, Z., et al. (2018). Differential responses to Cd stress induced by exogenous application of Cu, Zn or Ca in the medicinal plant Catharanthus roseus. Ecotoxicol. Environ. Saf. 157, 266–275. doi: 10.1016/j.ecoenv.2018.03.055
Dabral, S., Varma, A., Choudhary, D. K., Bahuguna, R. N., Nath, M. (2019). Biopriming with Piriformospora indica ameliorates cadmium stress in rice by lowering oxidative stress and cell death in root cells. Ecotoxicol. Environ. Saf. 186, 109741. doi: 10.1016/j.ecoenv.2019.109741
Daszkowska-Golec, A., Szarejko, I. (2013). Open or close the gate–stomata action under the control of phytohormones in drought stress conditions. Front. Plant Sci. 4, 138. doi: 10.3389/fpls.2013.00138
Ding, Z., Kheir, A. M., Ali, O. A., Hafez, E. M., ElShamey, E. A., Zhou, Z., et al. (2021). A vermicompost and deep tillage system to improve saline-sodic soil quality and wheat productivity. J. Environ. Manag. 277, 111388. doi: 10.1016/j.jenvman.2020.111388
Deng, X., Wu, C., Li, Q., Li, W. (2017). “Effect of vermicompost on soil enzyme activity of coastal saline soil in water spinach plantation,” in 2017 6th international conference on energy, environment and sustainable development (ICEESD 2017). 419–422 (Atlantis Press). doi: 10.2991/iceesd-17.2017.79
Dubey, R. K., Dubey, P. K., Chaurasia, R., Singh, H. B., Abhilash, P. C. (2020). Sustainable agronomic practices for enhancing the soil quality and yield of Cicer arietinum L. under diverse agroecosystems. J. Environ. Manage. 262, 110284. doi: 10.1016/j.jenvman.2020.110284
Elmer, W., White, J. C. (2018). The future of nanotechnology in plant pathology. Annu. Rev. Phytopathol. 56, 111–133. doi: 10.1146/annurev-phyto-080417-050108
Erdal, İ., Ekinci, K. (2020). Effects of composts and vermicomposts obtained from forced aerated and mechanically turned composting method on growth, mineral nutrition and nutrient uptake of wheat. J. Plant Nutr. 43, 1343–1355. doi: 10.1080/01904167.2020.1727506
Fernández-Bayo, J. D., Nogales, R., Romero, E. (2009). Assessment of three vermicomposts as organic amendments used to enhance diuron sorption in soils with low organic carbon content. Eur. J. Soil Sci. 60, 935–944. doi: 10.1111/j.1365-2389.2009.01176.x
Gu, J. F., Zhou, H., Tang, H. L., Yang, W. T., Zeng, M., Liu, Z. M., et al. (2019). Cadmium and arsenic accumulation during the rice growth period under in situ remediation. Ecotoxicol. Environ. Saf. 171, 451–459. doi: 10.1016/j.ecoenv.2019.01.003
Gul, S., Whalen, J. K., Thomas, B. W., Sachdeva, V., Deng, H. (2015). Physico-chemical properties and microbial responses in biochar-amended soils: mechanisms and future directions. Agricult. Ecosyst. Environ. 206, 46–59. doi: 10.1016/j.agee.2015.03.015
Haider, F. U., Liqun, C., Coulter, J. A., Cheema, S. A., Wu, J., Zhang, R., et al. (2021). Cadmium toxicity in plants: Impacts and remediation strategies. Ecotoxicol. Environ. Saf. 211, 111887. doi: 10.1016/j.ecoenv.2020.111887
Hamid, Y., Tang, L., Hussain, B., Usman, M., Lin, Q., Rashid, M. S., et al. (2020). Organic soil additives for the remediation of cadmium contaminated soils and their impact on the soil-plant system: A review. Sci. Total Environ. 707, 136121. doi: 10.1016/j.scitotenv.2019.136121
Hoque, T. S., Hasan, A. K., Hasan, M. A., Nahar, N., Dey, D. K., Mia, S., et al. (2022). Nutrient release from vermicompost under anaerobic conditions in two contrasting soils of Bangladesh and its effect on wetland rice crop. Agriculture 12, 376. doi: 10.3390/agriculture12030376
Huang, Y., Wang, L., Wang, W., Li, T., He, Z., Yang, X. (2019). Current status of agricultural soil pollution by heavy metals in China: A meta-analysis. Sci. Total Environ. 651, 3034–3042. doi: 10.1016/j.scitotenv.2018.10.185
Hussain, S., Khaliq, A., Noor, M. A., Tanveer, M., Hussain, H. A., Hussain, S., et al. (2020). "Metal toxicity and nitrogen metabolism in plants: an overview." in Carbon nitrogen cycling Soil. In: Datta, R., Meena, R., Pathan, S., Ceccherini, M. (eds). doi: 10.1007/978-981-13-7264-3_7
Huybrechts, M., Hendrix, S., Bertels, J., Beemster, G. T., Vandamme, D., Cuypers, A. (2020). Spatial analysis of the rice leaf growth zone under controlled and cadmium-exposed conditions. Environ. Exp. Bot. 177, pp.104120. doi: 10.1016/j.envexpbot.2020.104120
Imran, M., Hussain, S., Rana, S. R., Saleem, M. H., Rasul, F., Ali, K. H., et al. (2021). Molybdenum improves 2-acetyl-1-pyrroline, grain quality traits and yield attributes in fragrant rice through efficient nitrogen assimilation under cadmium toxicity. Ecotoxicol. Environ. Saf. 211, 111911. doi: 10.1016/j.ecoenv.2021.111911
Imran, M., Sun, X., Hussain, S., Ali, U., Rana, M. S., Rasul, F., et al. (2019). Molybdenum-induced effects on nitrogen metabolism enzymes and elemental profile of winter wheat (Triticum aestivum L.) under different nitrogen sources. Int. J. Mol. Sci. 20, 3009. doi: 10.3390/ijms20123009
Iqbal, A., Ali, I., Yuan, P., Khan, R., Liang, H., Wei, S., et al. (2022). Combined application of manure and chemical fertilizers alters soil environmental variables and improves soil fungal community composition and rice grain yield. Frontier Microbiol. 13, 856355. doi: 10.3389/fmicb.2022.856355
Iqbal, A., He, L., Khan, A., Wei, S., Akhtar, K., Ali, I., et al. (2019). Organic manure coupled with inorganic fertilizer: An approach for the sustainable production of rice by improving soil properties and nitrogen use efficiency. Agronomy 9, 651. doi: 10.3390/agronomy9100651
Iqbal, A., Hussain, Q., Mo, Z., Hua, T., Mustafa, A. E. Z. M., Tang, X. (2024). Vermicompost supply enhances fragrant-rice yield by improving soil fertility and eukaryotic microbial community composition under environmental stress conditions. Microorganisms 12, 1252. doi: 10.3390/microorganisms12061252
Iqbal, A., Khan, A., Green, S.J., Ali, I., He, L., Zeeshan, M., et al. (2021). Long-term straw mulching in a no-till field improves soil functionality and rice yield by increasing soil enzymatic activity and chemical properties in paddy soils. J. Plant Nutr. Soil Sci. 184, 622–634. doi: 10.1002/jpln.202100089
Iqbal, A., Mo, Z., Pan, S. G., Qi, J. Y., Hua, T., Imran, M., et al (2023). Exogenous tiO2 nanoparticles alleviate cd toxicity by reducing cd uptake and regulating plant physiological activity and antioxidant defense systems in rice (Oryza sativa L.). Metabolites 13, 765. doi: 10.3390/metabo13060765
Islam, M. N., Taki, G., Nguyen, X. P., Jo, Y. T., Kim, J., Park, J. H. (2017). Heavy metal stabilization in contaminated soil by treatment with calcined cockle shell. Environ. Sci. pollut. Res. 24, 7177–7183. doi: 10.1007/s11356-016-8330-5
Jezussek, M., Juliano, B. O., Schieberle, P. (2002). Comparison of key aroma compounds in cooked brown rice varieties based on aroma extract dilution analyses. J. Agric. Food Chem. 50, 1101–1105. doi: 10.1021/jf0108720
Jin, Z., Zhang, M., Li, R., Zhang, X., Wang, G., Liu, X, et al. (2020). Spent mushroom substrate combined with alkaline amendment passivates cadmium and improves soil property. Environ. Sci. Pollut. Res. 27, 16317–16325. doi: 10.1007/s11356-020-08099-3
Kanu, A. S., Ashraf, U., Mo, Z., Fuseini, I., Mansaray, L. R., Duan, M., et al. (2017). Cadmium uptake and distribution in fragrant rice genotypes and related consequences on yield and grain quality traits. J. Chem. 137, 1–9. doi: 10.1155/2017/1405878
Karlsons, A., Osvalde, A., Andersone-Ozola, U., Ievinsh, G. (2016). Vermicompost from municipal sewage sludge affects growth and mineral nutrition of winter rye (secale cereale) plants. J. Plant Nutr. 39, 765–780. doi: 10.1080/01904167.2015.1087566
Keesstra, S. D., Bouma, J., Wallinga, J., Tittonell, P., Smith, P., Cerdà, A., et al. (2016). The significance of soils and soil science towards realization of the United Nations Sustainable Development Goals. Soil 2, 111–128. doi: 10.5194/soil-2-111-2016
Khan, A., Najeeb, U., Wang, L., Tan, D. K. Y., Yang, G., Munsif, F., et al. (2017). Planting density and sowing date strongly influence growth and lint yield of cotton crops. Field Crops Res. 209, 129–135. doi: 10.1016/j.fcr.2017.04.019
Khan, S., Sehar, Z., Fatma, M., Mir, I. R., Iqbal, N., Tarighat, M. A., et al. (2022). Involvement of ethylene in melatonin-modified photosynthetic-N use efficiency and antioxidant activity to improve photosynthesis of salt grown wheat. Physiol. Plant. 174, e13832. doi: 10.1111/ppl.v174.6
Khaokaew, S., Chaney, R. L., Landrot, G., Ginder-Vogel, M., Sparks, D. L. (2011). Speciation and release kinetics of cadmium in an alkaline paddy soil under various flooding periods and draining conditions. Environ. Sci. Technol. 45, 4249–4255. doi: 10.1021/es103971y
Li, J., Chang, Y., AL-Huqail, A. A., Ding, Z., Al-Harbi, M. S., Ali, E. F., et al. (2021). Effect of manure and compost on the phytostabilization potential of heavy metals by the halophytic plant wavy-leaved saltbush. Plants 10, 2176. doi: 10.3390/plants10102176
Li, J., Lu, Y., Shim, H., Deng, X., Lian, J., Jia, J., et al. (2010). Use of the BCR sequential extraction procedure for the study of metal availability to plants. J. Environ. Monit. 12, 466–471. doi: 10.1039/B916389A
Liang, L., Liu, W., Sun, Y., Huo, X., Li, S., Zhou, Q. (2017). Phytoremediation of heavy metal contaminated saline soils using halophytes: current progress and future perspectives. Environ. Rev. 25, 269–281. doi: 10.1139/er-2016-0063
Lim, S. L., Wu, T. Y., Lim, P. N., Shak, K. P. Y. (2015). The use of vermicompost in organic farming: overview, effects on soil and economics. J. Sci. Food Agric. 95, 1143–1156. doi: 10.1002/jsfa.2015.95.issue-6
Lu, R. K. (2000). Analytical Methods of Soil and Agrochemistry (Beijing: China Agricultural Science and Technology Press).
Luo, H., He, L., Du, B., Pan, S., Mo, Z., Duan, M., et al. (2020). Biofortification with chelating selenium in fragrant rice: Effects on photosynthetic rates, aroma, grain quality and yield formation. Field Crops Res. 255, 107909. doi: 10.1016/j.fcr.2020.107909
Maji, D., Misra, P., Singh, S., Kalra, A. (2017). Humic acid rich vermicompost promotes plant growth by improving microbial community structure of soil as well as root nodulation and mycorrhizal colonization in the roots of Pisum sativum. Appl. Soil Ecol. 110, 97–108. doi: 10.1016/j.apsoil.2016.10.008
Makoto, K., Koike, T. (2007). Effects of nitrogen supply on photosynthetic and anatomical changes in current-year needles of Pinus koraiensis seedlings grown under two irradiances. Photosynthetica 45, 99–104. doi: 10.1007/s11099-007-0015-3
Mitra, S., Pramanik, K., Sarkar, A., Ghosh, P. K., Soren, T., Maiti, T. K. (2018). Bioaccumulation of cadmium by Enterobacter sp. and enhancement of rice seedling growth under cadmium stress. Ecotoxicol. Environ. Saf. 156, 183–196. doi: 10.1016/j.ecoenv.2018.03.001
Mo, Z., Li, Y., Nie, J., He, L., Pan, S., Duan, M., et al. (2019). Nitrogen application and different water regimes at booting stage improved yield and 2-acetyl-1-pyrroline (2AP) formation in fragrant rice. Rice 12, pp.1–pp16. doi: 10.1186/s12284-019-0328-4
Molins-Legua, C., Meseguer-Lloret, S., Moliner-Martinez, Y., Campíns-Falcó, P. (2006). A guide for selecting the most appropriate method for ammonium determination in water analysis. TrAC Trends Anal. Chem. 25, 282–290. doi: 10.1016/j.trac.2005.12.002
Muneer, S., Qadri, T. N., Siddiqi, T. O. (2011). Cytogenetic and biochemical investigations to study the response of Vigna radiata to cadmium stress. Afr. J. Plant Sci. 5, 183–192. doi: 10.5897/AJPS.9000057
Ni, K., Shi, Y. Z., Yi, X. Y., Zhang, Q. F., Fang, L., Ma, L. F., et al. (2018). Effects of long-term nitrogen application on soil acidification and solution chemistry of a tea plantation in China. Agric. Ecosyst. Environ. 252, 74–82. doi: 10.1016/j.agee.2017.10.004
Ohyama, T. (1991). Analytical procedures of N, P, K contents in plant and manure materials using H2SO4-H2O2 Kjeldahl digestion method. Bull. Facul. Agric. Niigata Univ 43, 111–120.
Ok, Y. S., Kim, D. K., Skousen, J. G., Lee, J. S., Cheong, Y. W., S.J.and Yang, J. E. (2021b). Ameliorants to immobilize Cd in rice paddy soils contaminated by abandoned metal mines in Korea. Environ. Geochem. Health 33, 23–30. doi: 10.1007/s10653-010-9364-0
Ok, Y. S., Usman, A. R. A., Lee, S. S., Abd El-Azeem, S. A. M., Choi, B., Hashimoto, Y., et al. (2011a). Effects of rapeseed residue on lead and cadmium availability and uptake by rice plants in heavy metal contaminated paddy soil. Chemosphere 85, 677–682. doi: 10.1016/j.chemosphere.2011.06.073
Oyege, I., Balaji Bhaskar, M. S. (2023). Effects of vermicompost on soil and plant health and promoting sustainable agriculture. Soil Sys. 7, 101. doi: 10.3390/soilsystems7040101
Parmar, P., Kumari, N., Sharma, V. (2013). Structural and functional alterations in photosynthetic apparatus of plants under cadmium stress. Bot. Stud. 54, 1–6. doi: 10.1186/1999-3110-54-45
Pramanik, K., Mitra, S., Sarkar, A., Maiti, T. K. (2018). Alleviation of phytotoxic effects of cadmium on rice seedlings by cadmium resistant PGPR strain Enterobacter aerogenes MCC 3092. J. Hazard. Mater. 351, 317–329. doi: 10.1016/j.jhazmat.2018.03.009
Rafiq, M. T., Aziz, R., Yang, X., Xiao, W., Rafiq, M. K., Ali, B., et al. (2014). Cadmium phytoavailability to rice (Oryza sativa L.) grown in representative Chinese soils. A model to improve soil environmental quality guidelines for food safety. Ecotoxicol. Environ. Saf. 103, 101–107. doi: 10.1016/j.ecoenv.2013.10.016
Ramazanoglu, E. (2024). Effects of vermicompost application on plant growth and soil enzyme activity in wheat (Triticum aestivum L.) monitored by thermal imaging. Cogent Food Agric. 10. doi: 10.1080/23311932.2024.2373872
Rizwan, M., Ali, S., Abbas, T., Zia-ur-Rehman, M., Hannan, F., Keller, C., et al. (2016). Cadmium minimization in wheat: a critical review. Ecotoxicol. Environ. Saf. 130, 43–53. doi: 10.1016/j.ecoenv.2016.04.001
Sauvé, S., Hendershot, W., Allen, H. E. (2000). Solid-solution partitioning of metals in contaminated soils: dependence on pH, total metal burden, and organic matter. Environ. Sci. Technol. 34, 1125–1131. doi: 10.1021/es9907764
Seleiman, M. F., Ali, S., Refay, Y., Rizwan, M., Alhammad, B. A., El-Hendawy, S. E. (2020). Chromium resistant microbes and melatonin reduced Cr uptake and toxicity, improved physio-biochemical traits and yield of wheat in contaminated soil. Chemosphere 250, 126239. doi: 10.1016/j.chemosphere.2020.126239
Shahid, M., Dumat, C., Khalid, S., Niazi, N. K., Antunes, P. M. (2017). Cadmium bioavailability, uptake, toxicity and detoxification in soil-plant system. Rev. Environ. contam. Toxicol. 241, 73–137. doi: 10.1007/398_2016_8
Shen, Z., Yu, Z., Xu, L., Zhao, Y., Yi, S., Shen, C., et al. (2022). Effects of vermicompost application on growth and heavy metal uptake of barley grown in mudflat salt-affected soils. Agronomy 12, 1007. doi: 10.3390/agronomy12051007
Singh, P., Singh, I., Shah, K. (2020). Alterations in antioxidative machinery and growth parameters upon application of nitric oxide donor that reduces detrimental effects of cadmium in rice seedlings with increasing days of growth. South Afr. J. Bot. 131, 283–294. doi: 10.1016/j.sajb.2020.02.022
Sun, Y., Ma, J., Sun, Y., Xu, H., Yang, Z., Liu, S., et al. (2012). The effects of different water and nitrogen managements on yield and nitrogen use efficiency in hybrid rice of China. Field Crops Res. 127, 85–98. doi: 10.1016/j.fcr.2011.11.015
Tang, X., Li, X., Liu, X., Hashmi, M. Z., Xu, J., Brookes, P. C. (2015). Effects of inorganic and organic amendments on the uptake of lead and trace elements by Brassica chinensis grown in an acidic red soil. Chemosphere. 119, 177–183. doi: 10.1016/j.chemosphere.2014.05.081
Tang, J., Zhang, J., Ren, L., Zhou, Y., Gao, J., Luo, L., et al. (2019). Diagnosis of soil contamination using microbiological indices: A review on heavy metal pollution. J. Environ. Manage. 242, 121–130. doi: 10.1016/j.jenvman.2019.04.061
Tejada, M., Gómez, I., Hernández, T., García, C. (2010). Utilization of vermicomposts in soil restoration: effects on soil biological properties. Soil Sci. Soc. America J. 74, 525–532. doi: 10.2136/sssaj2009.0260
Tran, T. A., Popova, L. P. (2013). Functions and toxicity of cadmium in plants: recent advances and future prospects. Turkish J. Bot. 37, 1–13. doi: 10.3906/bot-1112-16
Wan, Y., Huang, Q., Wang, Q., Yu, Y., Su, D., Qiao, Y., et al. (2020). Accumulation and bioavailability of heavy metals in an acid soil and their uptake by paddy rice under continuous application of chicken and swine manure.J. Hazard. Mater. 384, 121293. doi: 10.1016/j.jhazmat.2019.121293
Wang, S., Tian, H., Liu, J., Pan, S. (2003). Pattern and change of soil organic carbon storage in China: 1960s–1980s. Tellus B: Chem. Phys. Meteor. 55, 416–427. doi: 10.1034/j.1600-0889.2003.00039.x
Wang, Y., Xu, Y., Li, D., Tang, B., Man, S., Jia, Y., et al. (2018). Vermicompost and biochar as bio-conditioners to immobilize heavy metal and improve soil fertility on cadmium contaminated soil under acid rain stress. Sci. Total Environ. 621, 1057–1065. doi: 10.1016/j.scitotenv.2017.10.121
Wu, M., Wang, P. Y., Sun, L. G., Zhang, J. J., Yu, J., Wang, Y. W., et al. (2014). Alleviation of cadmium toxicity by cerium in rice seedlings is related to improved photosynthesis, elevated antioxidant enzymes and decreased oxidative stress. Plant Growth Regul. 74, 251–260. doi: 10.1007/s10725-014-9916-x
Xue, S., Shi, L., Wu, C., Wu, H., Qin, Y., Pan, W., et al. (2017). Cadmium, lead, and arsenic contamination in paddy soils of a mining area and their exposure effects on human HEPG2 and keratinocyte cell-lines. Environ. Res. 156, 23–30. doi: 10.1016/j.envres.2017.03.014
Yang, B., Xiong, Z., Wang, J., Xu, X., Huang, Q., Shen, Q. (2015). Mitigating net global warming potential and greenhouse gas intensities by substituting chemical nitrogen fertilizers with organic fertilization strategies in rice–wheat annual rotation systems in China: A 3-year field experiment. Ecol. Eng. 81, 289–297. doi: 10.1016/j.ecoleng.2015.04.071
Yang, S., Zou, Y., Liang, Y., Xia, B., Liu, S., Ibrahim, M., et al. (2012). Role of soil total nitrogen in aroma synthesis of traditional regional aromatic rice in China. Field Crops Res. 125, 151–160. doi: 10.1016/j.fcr.2011.09.002
Zhang, J., Sun, X., Xu, M., Zhao, X., Yang, C., Li, K., et al. (2022). A Self-amplifying ROS-sensitive prodrug-based nanodecoy for circumventing immune resistance in chemotherapy-sensitized immunotherapy. Acta Biomater. 149, 307–320. doi: 10.1016/j.actbio.2022.06.035
Zhang, W., Du, W., Wang, F., Xu, H., Zhao, T., Zhang, H., et al. (2020). Comparative study on Pb2+ removal from aqueous solutions using biochars derived from cow manure and its vermicompost. Sci. Total Environ. 716, 137108. doi: 10.1016/j.scitotenv.2020.137108
Keywords: vermicompost, cadmium toxicity, fragrant rice, 2-Acetyl-1-pyrroline, leaf physiological activity, N metabolism, metalloid stress
Citation: Iqbal A, Chen X, Khan R, Zaman M, Khan AH, Kiedrzyński M, Ebaid M, Alrefaei AF, Lamlom SF, Tang X and Zeeshan M (2024) Vermicompost application improves leaf physiological activity, 2-acetyl-1-pyrroline, and grain yield of fragrant rice through efficient nitrogen assimilation under Cd stress. Front. Plant Sci. 15:1481372. doi: 10.3389/fpls.2024.1481372
Received: 15 August 2024; Accepted: 29 October 2024;
Published: 02 December 2024.
Edited by:
Isabel Nogues, National Research Council (CNR), ItalyReviewed by:
Hongmei Cai, Huazhong Agricultural University, ChinaCopyright © 2024 Iqbal, Chen, Khan, Zaman, Khan, Kiedrzyński, Ebaid, Alrefaei, Lamlom, Tang and Zeeshan. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Muhammad Zeeshan, MTE2MTYxMDJAemp1LmVkdS5jbg==; Xiangru Tang, dGFuZ3hyQHNjYXUuZWR1LmNu
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.