 Muluken Demelie Alemu1,2†
Muluken Demelie Alemu1,2† Shiran Ben-Zeev1†
Shiran Ben-Zeev1† Vered Barak1Yusuf Tutus3†
Vered Barak1Yusuf Tutus3† Ismail Cakmak3†
Ismail Cakmak3† Yehoshua Saranga1*†
Yehoshua Saranga1*†- 1R. H. Smith Faculty of Agriculture, Food and Environment, The Hebrew University of Jerusalem, Rehovot, Israel
- 2Crop Research, Ethiopian Institute of Agricultural Research, Addis Ababa, Ethiopia
- 3Faculty of Engineering and Natural Sciences, Sabanci University, Istanbul, Türkiye
Climate change is becoming a global challenge, threating agriculture’s capacity to meet the food and nutritional requirements of the growing population. Underutilized crops present an opportunity to address climate change and nutritional deficiencies. Tef is a stress-resilient cereal crop, producing gluten-free grain of high nutritional quality. However, knowledge is lacking on tef’s diversity of grain nutritional properties, their interaction with environmental conditions (e.g., water availability) and the underlying genomic loci. We assessed the effect of water availability on tef grain nutrient concentrations and identify the associated genomic loci. A collection of 223 tef genotypes, a subset of tef diversity panel 300 (TDP-300), were grown in the field under well-watered and water-limited conditions in 2021, and phenotyped for 11 traits including: grain protein and mineral concentrations and seed color. A genome-wide association study was conducted using 28,837 single-nucleotide polymorphisms (SNPs) and phenotypic data to identify marker–trait associations (MTAs). Tef grain nutrient concentrations exhibited wide genetic diversity with a significant influence of environment. Protein and most micronutrients were more concentrated under water-limited conditions, whereas most macronutrients were higher in the well-watered environment. A total of 59 SNPs were associated with one or more of the studied traits, resulting in 65 MTAs detected under both water treatments, and providing insights into the genetic basis of grain nutrients. Five SNPs reflected multiple associations, with four detecting the same trait under both treatments (multiple-environment effect), and one associated with both Zn and K (pleiotropic effect). In addition, two pairs of closely linked SNPs reflected multiple-environment effects. While multiple-environment associations provide greater support for the integrity of these MTAs, the pleiotropic locus hints at a common mechanism controlling two mineral ions. The identified MTAs shed new light on the genomic architecture of tef’s nutritional properties and provide the basis to enhance tef grain nutritional quality alongside drought resilience.
1 Introduction
With millions of people suffering from malnutrition due to protein and micronutrient deficiencies (FAO, 2020; Stevens et al., 2022), the capacity of agriculture to meet food and nutritional requirements of a growing population in the face of climate change is becoming a global concern (FAO, 2023). Underutilized (orphan) crop species, which are not widely cultivated, present outstanding nutritional value and a high tolerance to abiotic and biotic stresses (Cheng, 2018; Bekkering and Tian, 2019; Hunter et al., 2019). Although they are considered underutilized crops, some are staple crops in their origin and center of diversity. These crops offer the potential to improve food and nutrition security for millions of people (Mabhaudhi et al., 2019; Siddique et al., 2021), but they have not been sufficiently studied or improved (Tadele, 2019; Allaby, 2021; Milla and Osborne, 2021; Alemu et al., 2024b).
Tef (Eragrostis tef (Zucc.) Trotter) is a C4 cereal crop with a genome size of 622 Mb (VanBuren et al., 2020). Ethiopia is tef’s origin and center of diversity (Vavilov, 1951), where it is the most important food and feed crop (D’Andrea, 2008; Assefa et al., 2017; Chanyalew et al., 2019; Tadele, 2019), and the primary ingredient of injera, the traditional Ethiopian flat bread (Abewa et al., 2019; Ligaba-Osena et al., 2021). Tef is a prestigious cereal crop with outstanding nutritional value, producing gluten-free grains, rich in minerals (Fe, Cu, Mn, Zn, B, Mo, Na, Ca, Mg, K, P, S), carbohydrates, fat, protein, essential amino acids, vitamins and dietary fiber (Abewa et al., 2019; Ligaba-Osena et al., 2021; Shumoy et al., 2018; Tietel et al., 2020; Villanueva et al., 2022; Zhu, 2018). The concentrations of minerals, carbohydrates, fat, proteins and fiber in tef are comparable to or higher than those in other cereal crops (Saturni et al., 2010; Hager et al., 2012; Ligaba-Osena et al., 2021). Thus, tef is recognized as a most important food crop to combat malnutrition, including mitigation of iron-deficiency anemia, particularly in developing nations (Daba, 2017; Abewa et al., 2019). Its favorable nutritional composition and health benefits have contributed to a global interest in tef as a healthy superfood, especially for gluten-intolerant and sensitive individuals (Assefa et al., 2011; Zhu, 2018; Tietel et al., 2020; Ligaba-Osena et al., 2021).
Tef seed is broadly grouped into white and brown colors (Jifar et al., 2015), varying from ivory white to pale white and light brown to dark brown, respectively (Asefa et al., 2023). Seed color is associated with nutritional properties, where brown tef grain is superior to white grain with respect to most minerals and antioxidants (Abebe et al., 2007; Dame, 2020; Tietel et al., 2020), whereas white grain is superior with respect to essential amino acids and protein (Gebru et al., 2019).
Drought is a primary abiotic stress, significantly limiting crop development and productivity (El Sabagh et al., 2020; Vadez et al., 2024). Drought also influences grain quality, either positively or negatively (Stagnari et al., 2016), by modifying morphological, physiological and biochemical characteristics of crop species (El Sabagh et al., 2020; Dietz et al., 2021). Assessing crop grain quality under different environments and understanding the effects of gene-by-environment interactions on quality traits is crucial for crop improvement. Despite extensive documentation on the effects of drought stress on growth, development and production of various crops, its effects on grain quality have not been sufficiently investigated (Stagnari et al., 2016; Chadalavada et al., 2022). Recently, attention has been drawn to various effects of drought on grain quality of cereal crops using high-throughput phenotyping (El Sabagh et al., 2020; Chadalavada et al., 2022; Kamal et al., 2023). Tef grain nutrient concentrations are influenced by genotype and environmental factors (climatic and edaphic variations) (Abewa et al., 2019), but we are not aware of any prior study on the effect of contrasting water regimes on tef grain quality.
Genome-wide association studies (GWASs) have been used for a range of plant species to dissect complex traits in diversity panels (Atwell et al., 2010; Huang and Han, 2014). GWASs have been extensively applied for staple cereal crops, but they are only beginning to be used for underutilized crops (Chapman et al., 2022). Recently, GWASs have been successfully applied to several orphan crops, including tef (Woldeyohannes et al., 2022; Alemu et al., 2024b). High-throughput genotyping and phenotyping techniques provide opportunities to examine the genomic loci underlying grain quality (El Sabagh et al., 2020; Chadalavada et al., 2022; Kamal et al., 2023). Although the genomic dissection of grain nutrients in crops remains limited, especially in underutilized crops, grain nutritional quality has been targeted in numerous cereal-breeding programs, including foxtail millet (Jaiswal et al., 2019), finger millet (Puranik et al., 2020), sorghum (Kamal et al., 2023), barley (Nyiraguhirwa et al., 2022), wheat (Liu et al., 2021) and rice (Islam et al., 2022).
Tef genotypes hold a rich gene pool and substantial genetic variation with respect to nutrition, yet this crop remains untapped due to a lack of understanding of its genotypic diversity in nutrient concentrations and bioavailability (Ligaba-Osena et al., 2021), and of the genomic loci underlying grain nutritional properties (Ereful et al., 2022). A better understanding of the diversity in tef grain nutrient concentrations, their interaction with environmental conditions and genomic architecture is crucial for the development of nutritious tef varieties.
We previously reported on the assembly of a tef diversity panel 300 (TDP-300) and its utilization for a GWAS of productivity, phenology, lodging and morphophysiological traits under contrasting water regimes (Alemu et al., 2024b). In the current study we assessed tef grain nutritional properties, including protein, micronutrient and macronutrient concentrations, to identify their responses to water availability and their underlying genomic loci. The combination of our previous (Alemu et al., 2024b) and current GWASs shed new light on tef’s responses to water stress and provide solid grounds for further studies and the development of novel tef varieties combining drought resilience with high grain yield and nutritional quality.
2 Materials and methods
2.1 Plant materials and genotyping
A wide collection of tef genotypes, collected by the USDA in Ethiopia and maintained in the Israel Plant Gene Bank, was used for our current study. Genotype by sequencing and subsequent data processing and filtering resulted in 28,837 single-nucleotide polymorphism (SNP) markers called across 297 accessions, hereafter termed TDP-300. Further details on genotyping, SNP quality and population structure are presented in Alemu et al. (2024b). A subset of 223 genotypes from the TDP-300 grown in the field under well-watered (WW) and water-limited (WL) treatments were phenotyped for grain protein, micronutrients and macronutrients.
2.2 Growth conditions and phenotyping
Tef genotypes were grown in 2021, in the field at the Kvutzat Shiller farm in Israel (31.879° N, 34.777° E), under WW (total seasonal water applied: 438.3 mm) and WL (total seasonal water applied: 211.7 mm) conditions. The experiment consisted of two main plots, one per treatment, each comprising of three replicates per genotype (2 m2 plot size) in a randomized block design. Soil type at the experimental location was clay, with the upper layer (0–30 cm depth) at sowing time containing 8.1, 18.7 and 60.9 mg/kg soil N, P and K, respectively. Liquid fertilizer (Shefer 5-3-8, ICL, Israel) was applied via the irrigation system between seedling establishment and flowering onset at rates of 52, 30 and 85 kg/ha N, P2O5 and K2O, respectively. No rainfall occurred during the experimental seasons, and average temperatures (min/max) were 16.8/31.3°C. Complete details of the experimental layout, environmental conditions and management are described in Alemu et al. (2024b).
2.2.1 Grain protein
Grain protein concentrations were estimated with near-infrared spectrometer (NIRS) DS2500 (FOSS, Hillerød, Denmark). Whole-grain samples were dried at 40°C for 24 h, stored in a sealed plastic bag and cooled to room temperature. Approximately 10 g dried grain of each sample was transferred into a small sample cup (6 cm diameter), scanned, and its near-infrared spectral reflectance across 1100–2498 nm was recorded at 2-nm intervals, using Mosaic Solo software (FOSS Analytical, Hillerod, Denmark).
A preliminary calibration for the NIRS, based on 51 grain samples of various genotypes and treatments from two experiments conducted in two locations in 2019, was already available. In addition, 74 grain samples were prepared by bulking the three replicates of randomly selected genotypes from both treatments of our current experiment; 33 of these were added to the calibration set (for a total of 84 samples) and the remaining 41 samples were used for validation. Nitrogen (N) concentrations of the calibration and validation samples were determined at Bactochem laboratories (Ness Ziona, Israel) by the Kjeldahl method, using block digestion and steam distillation (Kjeldahl, 1883). Total protein content was then calculated by multiplying the N content by a conversion factor of 6.25. The laboratory protein concentrations of the 84 calibration samples were associated with the respective spectral data using the WinISI 4 software (FOSS Analytical, Hillerød, Denmark). Calibration curve coefficient (R2) and cross-validation (1-VR) were 0.99 and 0.98, respectively, whereas the R2 of the validation set was 0.90. The calibration models were then applied to the entire set of tef grain samples (3 replicates per genotype under each treatment) to determine their protein concentration.
2.2.2 Grain minerals
Samples for determining tef grain mineral concentrations were prepared by combining 0.5 g of grain from each of the three replicates of a genotype under a specific treatment (WW or WL). The grain samples were first thoroughly washed with tap water and then with deionized water to remove the adhering soil dusts, chaff and other possible contaminants. The washed samples were dried at about 40°C in a forced-draft oven to a constant weight and then subjected to analyses of the targeted mineral nutrients (Fe, Zn, Cu, Mn, Ca, Mg, K, P, and S). The samples were digested in a closed-vessel microwave system (MarsExpress; CEM Corp; Matthews, NC, USA) in 2 ml of 30% (v/v) premium-grade H2O2 (Merck, EMSURE®, Darmstadt, Germany) and 5 ml of 65% (v/v) premium-grade HNO3 (Merck, EMSURE®, Darmstadt, Germany) as described by (Yazici et al., 2021). All dilutions required were performed using ultra-pure water (electrical resistance 18.2 MΩ). The measurement of the mineral nutrients in the acid digests was conducted by inductively coupled plasma optical emission spectroscopy (ICP-OES; Agilent 5110 Vertical Dual View, Agilent Technologies, Inc, Santa Clara, CA, USA). The measurements were verified by using certified standard reference material (SRM) obtained from the National Institute of Standards and Technology (NIST, Gaithersburg, MD, USA).
2.2.3 Grain color
Grain colors were scored visually into two broad categories: white and brown (Supplementary Figure S1) and used both as an independent variable in the analysis of variance (ANOVA) and a dependent variable in the GWAS (scored 1 and 2 for brown and white seeds, respectively).
2.3 Statistical analyses
JMP Pro 17.0.0 (SAS Institute Inc., 1989–2021) was used for ANOVA. A full factorial nested ANOVA, including environment (E), seed color (SC), genotypes (G, nested in SC) and interactions, was used to analyze the replicated protein results and calculate the least squares mean values. For the non-replicated (bulk) grain minerals, two-way ANOVA was used to analyze the effects of G and E, whereas G * E interactions were visualized as raincloud plots, using “ggrain” package in R (Allen et al., 2021). The distribution of phenotypic traits was presented by density plot using the “ggplot2” package in R (Wickham et al., 2016). The associations among the tested grain nutrients, as well as grain yield (GY) and thousand seed weight (TSW), in WW and WL environments were assessed using Pearson’s correlation coefficient (Peterson et al., 2014) in JMP Pro 17.0.0 and principal component analysis (PCA) “factoextra” package in R (R Core Team, 2020). All traits were tested for normality using the Shapiro–Wilk test and in the few cases of traits that were not normally distributed, Box–Cox transformation procedure was used (JMP Pro 17.0), resulting in normal distribution of all traits.
2.4 Genome-wide associations
Phenotypic (11 traits) and genotypic (28,837 SNPs) data were subjected to GWAS using the Genome Association and Prediction Integrated Tool (GAPIT3) genetics statistical package (Wang and Zhang, 2021) in R (R Core Team, 2020). A multi-locus GWAS model, Bayesian-Information and Linkage-Disequilibrium Iteratively Nested Keyway (BLINK) (Huang et al., 2019), was selected due to the lowest rate of false positives as observed in the QQ plots. The effect of population structure was corrected for by including the kinship matrix (K-model) which was calculated using the VanRaden method (VanRaden, 2008). Principal components (PCs) were not included in the association model due to the low clustering tendency of the population (Alemu et al., 2024b). To minimize the rate of false positives, we used the stringent Bonferroni-corrected threshold (0.05/number of SNPs = 1.7E-06) (Bonferroni, 1936) to determine significant marker–trait associations (MTAs). Once the association between a certain SNP and a trait met this stringent threshold, a putative MTA was considered if the association of the same SNP with other traits or the same trait under a different environment met a less stringent threshold (p < 1E-04).
Neighboring MTAs were considered within the same linkage block (i.e., locus) when the linkage between them (r²) was greater than 0.5 and their physical distance smaller than 97,242 bp, the distance at which linkage disequilibrium (LD) decay decreases to below 0.2 (Remington et al., 2001).
The SNPs that were associated with grain nutrient concentrations across multiple environments or multiple traits (pleiotropic) were subjected to the tef genome browser (https://genomevolution.org/coge/GenomeInfo.pl?gid=50954) to identify candidate genes co-located with or nearest to the selected MTAs. The candidate genes were blasted in the National Center for Biotechnology Information (NCBI) database (http://www.ncbi.nlm.nih.gov/) to identify their homologous functions.
3 Results
3.1 Tef grain nutrient concentrations
The tef genotypes tested in this study exhibited wide variation in grain concentrations of protein, micronutrient (Fe, Zn, Cu and Mn) and macronutrient (Ca, Mg, K, P and S) under both WW and WL treatments (Figure 1), as well as across grain colors (brown and white) (Figure 2).
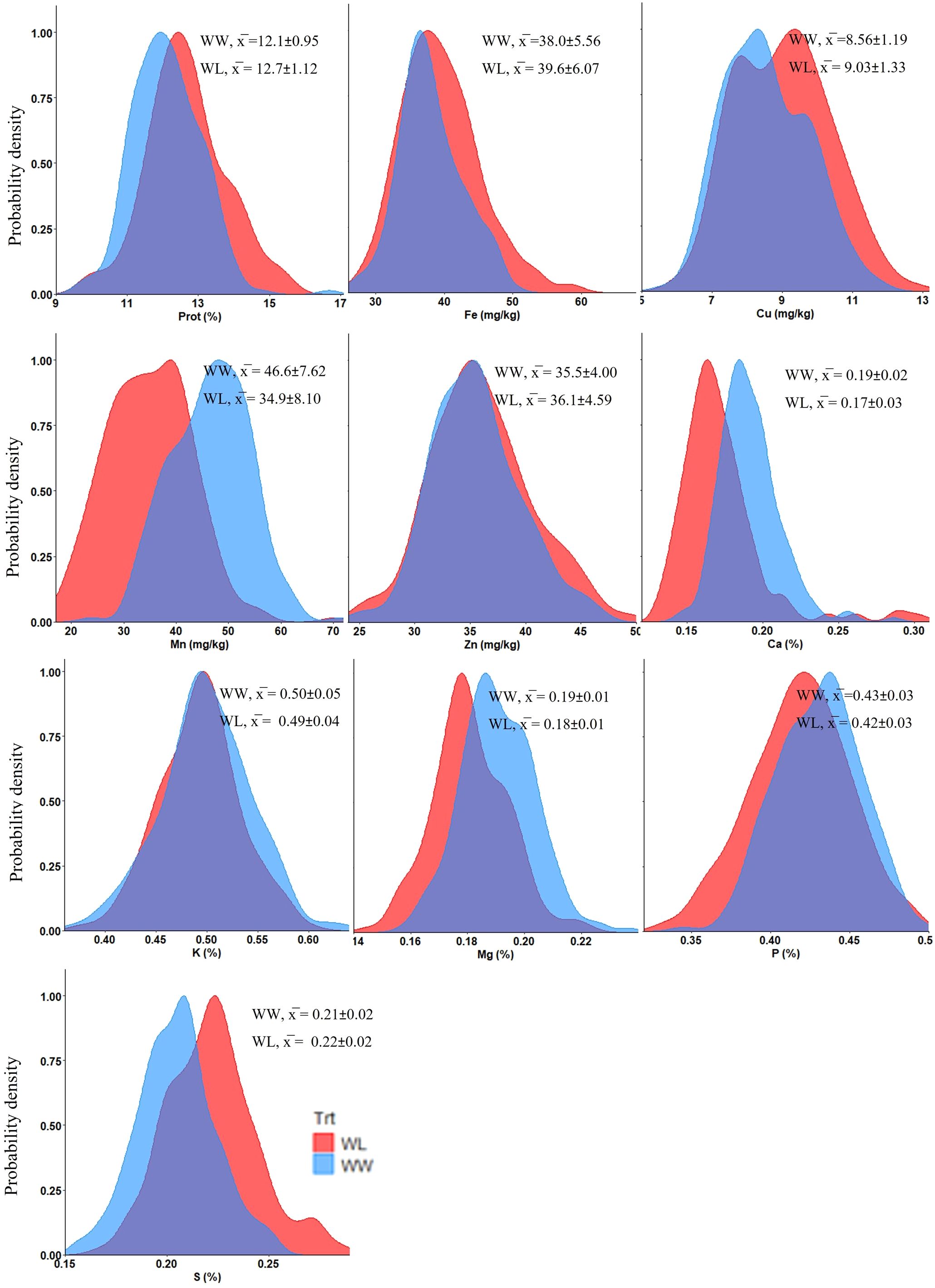
Figure 1. Density plots presenting the distribution of tef grain nutrient concentrations under well-watered (WW) and water-limited (WL) treatments in 2021. Prot, protein. x̅ indicates mean values ± SD.
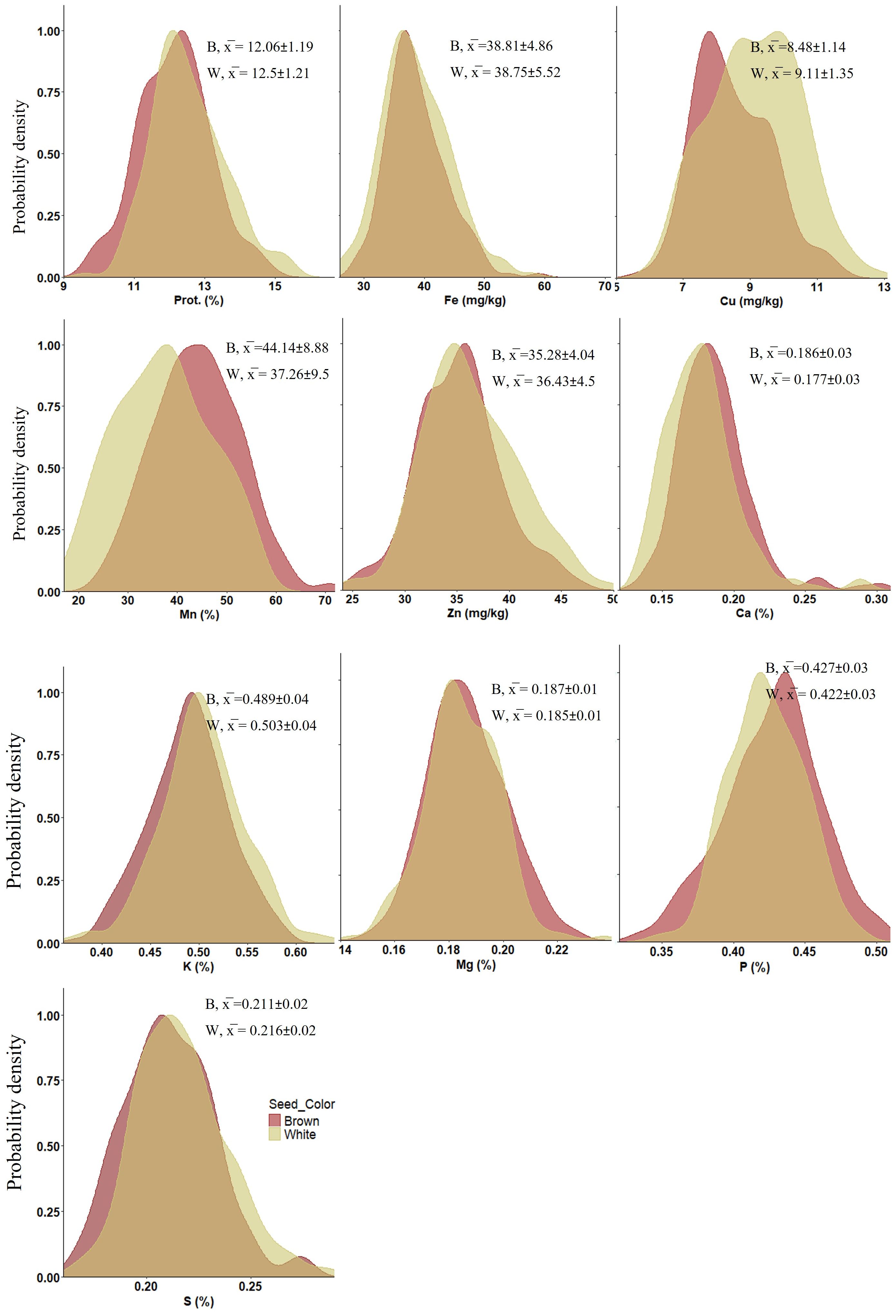
Figure 2. Density plots presenting the distribution of nutrient concentrations (averaged across treatments) in tef seeds of different colors: brown (B) (n = 114) and white (W) (n = 109) in 2021. Prot, protein. x̅ indicates mean values ± SD.
Water availability had a highly significant effect (p < 0.001) on most mineral nutrients, except Zn which exhibited a lower effect (p < 0.05) and K, which did not show any significant effect (Supplementary Table S1). Grain nutrient concentrations were either positively or negatively affected by water availability (Supplementary Table S1; Figure 1). Protein and most micronutrients (Fe, Zn and Cu) exhibited higher concentrations under WL vs. WW conditions, whereas Mn exhibited the opposite trend (Supplementary Table S1; Figure 1). On the other hand, most macronutrients (Ca, Mg and P) had higher concentrations under the WW vs. WL treatment, except for S which exhibited the opposite trend and K which, as already noted, was not affected (Supplementary Table S1; Figure 1). The treatment effects on most nutrient concentrations were in the range of 1–10%, except for Mn which exhibited a 25% lower concentration under the WL vs. WW treatment.
ANOVA revealed significant differences for all grain nutrient concentrations between genotypes (p < 0.001), as well as between grain colors (p < 0.01), except Fe which did not exhibit significant variation between seed colors (Figure 2; Supplementary Table S1). Protein, Cu, Zn, K and S exhibited higher concentrations in white-seeded tef genotypes. Conversely, Mn, Ca, Mg and P were at higher concentrations in the brown seed genotypes. While most differences in nutrient concentrations between seed colors were in the range of 0–7%, the most pronounced difference was noted for Mn, with an about 18% higher concentration in the brown-seeded vs. white-seeded genotypes.
Treatment-by-genotype interaction, assessed for the replicated protein data by ANOVA, revealed a significant effect (p < 0.01), whereas the treatment-by-seed color interaction was not significant (Supplementary Table S1). For the non-replicated micronutrients and macronutrients, which could not be subjected to full factorial ANOVA, raincloud plots (Supplementary Figure S2) suggested considerable treatment-by-genotype and treatment-by-seed color interactions for most minerals.
3.2 Association between phenotypic traits
Correlations among grain nutrients were positive and significant in most cases, non-significant in a few cases, and negative and significant in a single case (Mn vs. Cu under WL) (Supplementary Table S2). In contrast, the correlations between grain yield and grain nutrients were negative in most cases, non-significant in several other cases and positive in three cases (most pronounced for K vs. GY under WW conditions). Finally, the correlations between TSW and grain nutrients were usually non-significant, except three cases of negative correlations (S, Mg and P under WW conditions) and two cases of positive correlation (Fe under both treatments).
PCA, conducted separately for each environment, indicated variations in the principal components (PCs) between treatments (Figure 3). The two main PCs (eigenvalues >1) explained a total of 47.7% and 45.4% of the variation under WW and WL treatments, respectively. PC1 (X-axis) explained 32.5% and 29.8% of the variation and PC2 (Y-axis) explained 15.2% and 15.6% of the variation under WW and WL treatments, respectively. Most of the traits showed positive loadings for PC1 under both the WW and WL treatments, whereas grain yield was negatively loaded in PC1 under both treatments, thus supporting the correlation analyses. Fe and P exhibited the highest contribution to the variation under both treatments. High contributions were also noted for K under WL conditions and for protein under the WW treatment.
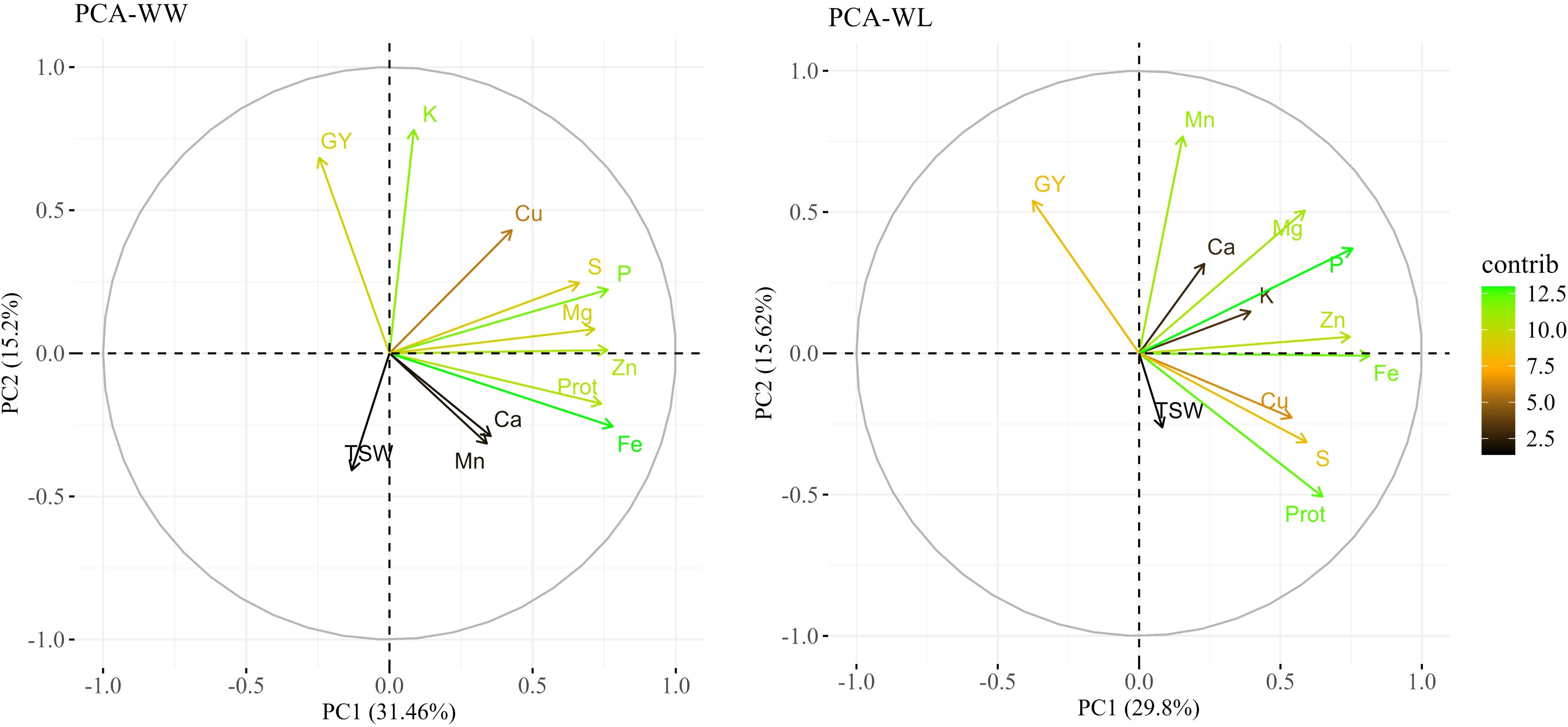
Figure 3. Principal component analysis (PCA) of grain nutrients and yield-related traits in a subset of tef diversity panel 300 (TDP-300) under well-watered (WW, left) and water-limited (WL, right) conditions. PCA included grain mineral nutrients and protein (Prot.) concentrations, grain yield (GY), and thousand seed weight (TSW). Biplot vectors showing trait loadings for PC1 and PC2, with color indicating trait contribution to variation.
3.3 Genome-wide association
A total of 59 significant MTAs (meeting the Bonferroni-corrected threshold, p < 1.7E-06) were identified for seed color and nutrient concentrations in a subset of TDP-300 under WW and WL treatments (Tables 1, 2). An additional four MTAs that met a less stringent threshold (p < 1E-04) but overlapped with significant MTAs were also considered and defined as putative MTAs. A greater number of significant MTAs was detected under the WL vs. WW treatment (Table 1). MTAs were unevenly distributed across 19 of the 20 tef chromosomes, with Chr 9B showing no significant association (Figure 4), and their abundance was rather similar on subgenome A (28 SNPs) and subgenome B (31 SNPs). Most tef grain nutritional traits showed significant MTAs under both WW and WL conditions, except for Ca and Zn, which exhibited MTAs only for the WL treatment (Table 1).
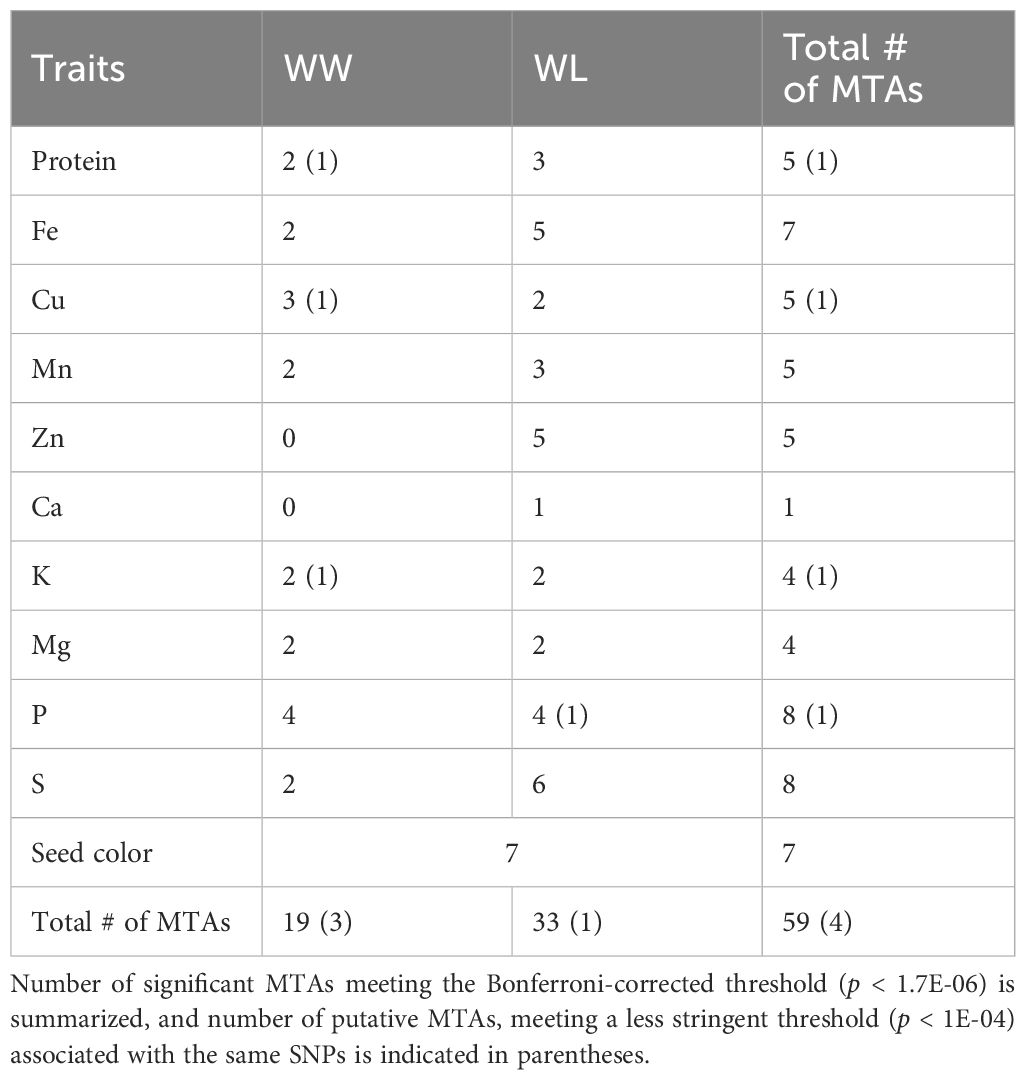
Table 1. Summary of marker–trait associations (MTAs) detected for tef grain nutrient concentrations under well-watered (WW) and water-limited (WL) conditions in 2021.
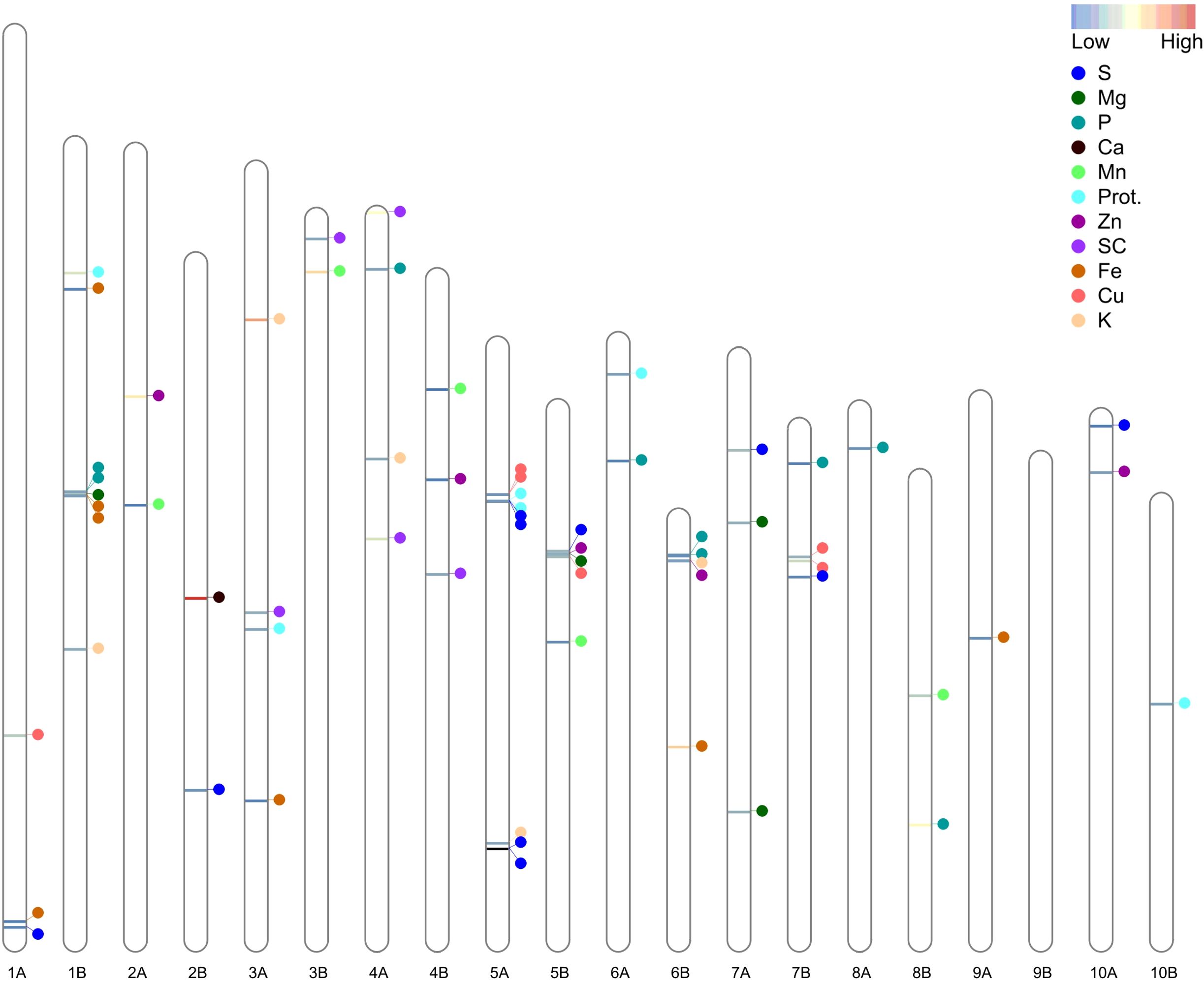
Figure 4. Distribution of 63 (significant and putative) marker–trait associations (MTAs) across the 20 tef chromosomes. Prot., protein; SC, seed color. The color of the horizontal bars on the chromosomes indicates the relative percentage of explained variation (PEV).
The detected (significand and putative) MTAs involved 58 SNPs, of which 53 were associated with a single trait under one environment, 4 were associated with the same trait under two environments, and 1 was associated with Zn and K (pleiotropic effect) (Table 2). LD analysis (Supplementary Figure S4) detected two groups of closely linked of MTAs (underlined in Table 2). Among them, the 1B linkage group (two SNPs) was associated with Fe under both environments and Mg under the WW treatment, whereas the 5A linkage group (three SNPs) was associated with S and protein, both under the two environments.
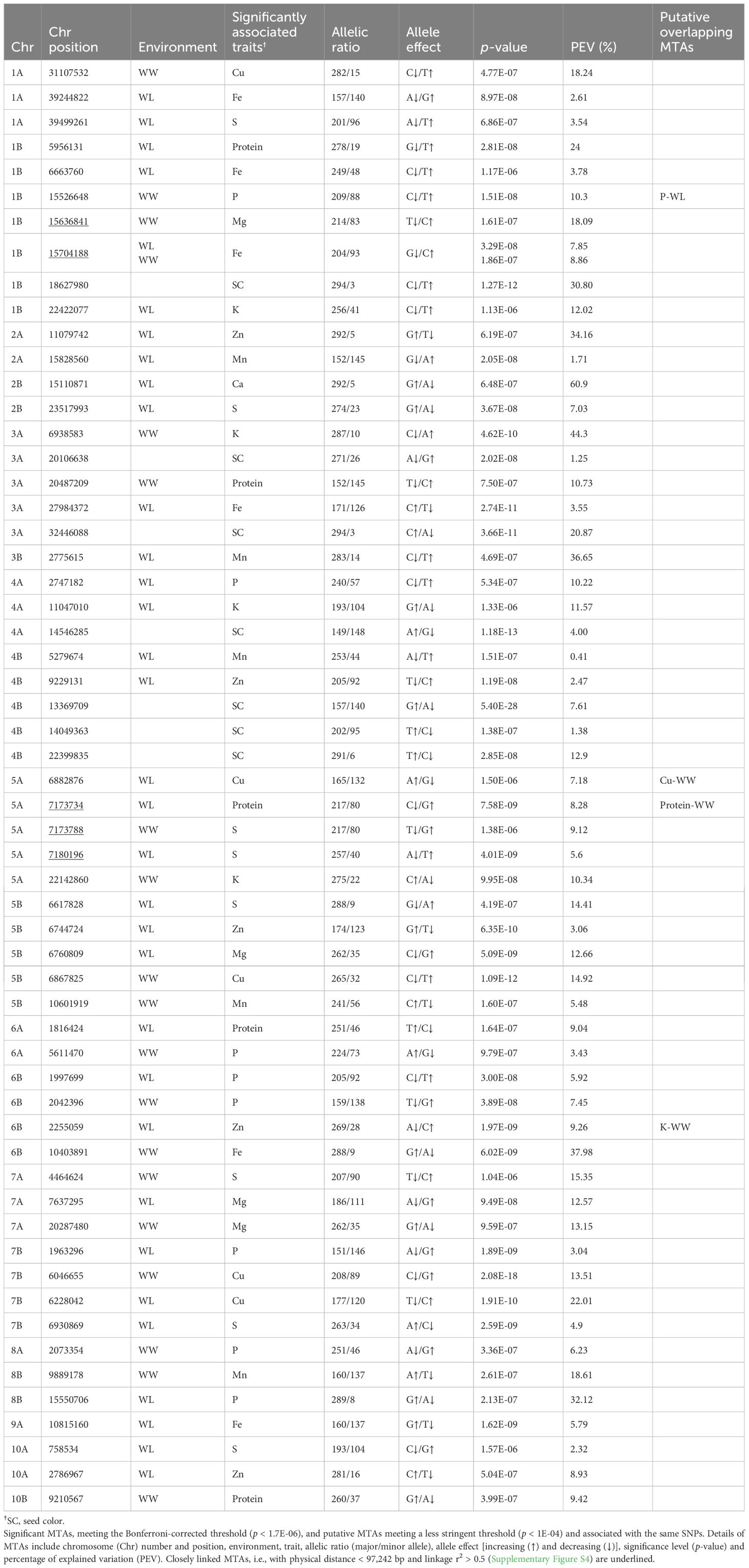
Table 2. List of marker–trait associations (MTAs) detected for tef seed color and nutrient concentrations under well-watered (WW) and water-limited (WL) conditions in 2021.
Five significant (and one putative) MTAs were found for grain protein (Tables 1, 2), with two of them involving the same SNP (5A_7173734) under both treatments (Table 2). The significant MTAs detected for protein explained 8.28–24% of the phenotypic variation, with the minor allele increasing protein concentration in three cases and the major allele increasing protein concentration in two cases.
A total of 22 significant (and one putative) MTAs were detected for grain micronutrients (Tables 1, 2). Two SNPs (1B_15704188 and 5A_6882876) were each associated with the same mineral (Fe and Cu, respectively) under both treatments. The MTAs detected for micronutrients explained between 0.4% and 37.98% of the phenotypic variation, with the lowest and highest variation observed for Mn and Fe, respectively. In 13 cases, the minor alleles increased the concentration of grain micronutrients, whereas in 9 cases, the major alleles increased the concentration of micronutrients.
A total of 25 significant (and two putative) MTAs were detected for macronutrients (Tables 1, 2). One SNP (1B_15526648) was associated with P under both treatments. The MTAs associated with macronutrients explained 2.3% to 60.9% of the phenotypic variation, with the lowest and highest variation observed for S and Ca, respectively. In most cases, the minor alleles increased the concentration of macronutrients (Table 2).
Seven significant MTAs were detected for seed color (Tables 1, 2). The significant MTAs explained 11% to 29.36% of the phenotypic variation for this trait. In most cases, the major alleles were associated with white seeds, whereas the minor alleles were associated with brown seeds (Table 2).
Five SNPs that exhibited either multiple-environment or pleiotropic associations (1B_15526648, 1B_15704188, 5A_6882876, 5A_7173734 and 6B_2255059) were prioritized to identify candidate genes. These SNPs were significantly associated with the following respective homologous genes: PTI1-like tyrosine-protein kinase 1, cell division control protein 48 homolog E-like, polygalacturonase inhibitor 1-like, ADP-ribosylation factor GTPase-activating protein AGD, and cytochrome b-c1 complex subunit 6 (Supplementary Table S3). These genes play roles in disease resistance, immune response against fungal polygalacturonases, controlling membrane-trafficking events, and electron transport, respectively.
4 Discussion
Global human diet is highly reliant on cereal crops, changes in their yield or quality can affect the food and nutrition security of millions of people (Galani et al., 2022, IFPRI, 2024). Drought stress slows grain-filling rate and shortens filling duration, thereby decreasing crop yields and nutritional properties (Sehgal et al., 2018). The effects of drought on yield and quality may vary depending on the crop species, genotype, environment, growth stage and stress severity (Stagnari et al., 2016; El Sabagh et al., 2020). While the effects of drought stress on the productivity of various crops have been extensively studied, the impact on grain quality and its genomic basis have been less investigated (Stagnari et al., 2016; Chadalavada et al., 2022). Recent development of high-throughput tools has created opportunities to examine the effects of drought on grain nutrient concentrations (El Sabagh et al., 2020; Chadalavada et al., 2022; Kamal et al., 2023). Nevertheless, there are only a few published studies on the effects of environmental factors on tef nutritional properties (Abewa et al., 2019) and their underlying genomic loci (Ereful et al., 2022), usually based on a small number of genotypes. Hence, the current study seems to be the first to investigate tef grain quality in a large number of genotypes under contrasting environments (WW and WL), and to implement a GWAS to detect the associated genomic loci.
4.1 Genetic diversity in grain nutrient concentrations
Grain mineral concentrations are the outcome of various physiological processes, including root uptake, translocation, redistribution within the plant tissues, remobilization and accumulation in the grain (Cakmak and Kutman, 2018; Marcos-Barbero et al., 2021). In the current study, grain nutrient concentrations exhibited wide variation among the tested genotypes across the two irrigation treatments (WW and WL) (Figure 1), thus reflecting the genetic diversity in tef germplasm. In previous studies, grain nutrient concentrations varied significantly across tef accessions in the field (Ereful et al., 2022) and greenhouse (Ligaba-Osena et al., 2021). The ranges of protein, micronutrients and macronutrients recorded in our study are mostly in agreement with previous reports on grain nutritional properties of tef grown under common field conditions in Ethiopia (Daba, 2017; Abewa et al., 2019), under irrigated field conditions in Israel (Tietel et al., 2020) and in greenhouse in Washington, USA (Ligaba-Osena et al., 2021). A clear deviation from this general agreement was found for Fe concentration, which in previous field studies (Daba, 2017; Abewa et al., 2019; Tietel et al., 2020), exhibited up to ~20-fold the concentration found in our study, most probably due to soil contamination in the grain samples (Stangoulis and Sison, 2008; Baye et al., 2014), but soil properties might also have some effect. Nevertheless, our results confirm the superiority of tef grain mineral contents, including Fe, in comparison with published results for other cereal crops (Abebe et al., 2007; Baye et al., 2014; Dame, 2020; Ligaba-Osena et al., 2021).
Cereals crop grains have various colors, and contain substantial levels of pigments associated with grain nutrient content (Francavilla and Joye, 2020). Significant variations in nutrient concentrations have been observed between colored grains in maize (Martínez-Martínez et al., 2019) and wheat (Li et al., 2023). In the current study, differently colored tef grains differed significantly in all nutrient concentrations except Fe (Supplementary Figure S1; Supplementary Table S1), suggesting either a physiological or genetic association between grain color and nutrient uptake and/or accumulation. The variation in most nutrients’ concentrations between grain colors ranged from 0–7%, with the exception of Mn which exhibited 18% higher concentration in brown compared to white grains. Our results also indicated significantly higher Ca, Mg and P concentrations in the brown seeds, whereas protein, Cu, Zn, K and S were more plentiful in white-seeded genotypes. In previous studies, brown tef grain was found superior to white grain with respect to most minerals (Abebe et al., 2007; Dame, 2020; Tietel et al., 2020), while protein concentration was higher in white tef grain (Gebru et al., 2019).
Soil moisture represents a key physical factor affecting directly transport of nutrients to root surfaces through mass flow and diffusion in soils, root-nutrient contact and uptake and shoot transport (Brouder and Volenec, 2023; Coskun and White, 2023). In wheat, abiotic stress (drought and heat) negatively affects grain yield while increasing protein and mineral concentrations (Ben Mariem et al., 2021; Galani et al., 2022), however, in a wild emmer wheat collection no effect of drought was evident on protein, Fe or Zn content (Peleg et al., 2008). In the current study, water regimes had a significant effect on tef grain nutrient concentrations, with the exception of K (Figure 2; Supplementary Table S1), which might be related to preferential deposition of K in the stem tissues of plants. Published reports show that up to 70% of K in aboveground plant parts is transported and deposited in the stem tissue of various plants, such as wheat and rice (Zorb et al., 2014; Xu et al., 2018). Seasonal water application in our field experiment was reduced by ~50% under WL compared to WW conditions, resulting in an average reduction of 42% in grain yield (Alemu et al., 2024b). However, the effects of this severe drought treatment on grain quality attributes were relatively modest, usually ranging between 1 and 10% (Figure 2). Protein and most micronutrients exhibited higher concentrations under WL compared to WW conditions, providing partial compensation for the yield reduction, while most macronutrients exhibited the opposite trend. Among grain nutrients, Mn was the most influenced by water availability and exhibited a 25% lower concentration under the WL vs. WW treatment. It has been well-documented that root Mn uptake and shoot Mn accumulation are strongly affected by water regime. Most commonly, plant Mn concentrations show significant reductions with decreases in soil water status due to rapid oxidation of Mn to chemical forms unavailable for root uptake (Tao et al., 2007; Schmidt et al., 2016; Alejandro et al., 2020). It was previously reported that tef grain nutrient concentrations are influenced by the soil’s physicochemical properties (Abewa et al., 2019). However, we are not aware of any prior study on the effect of drought on tef grain quality.
In tef, GY was negatively associated with most grain nutrient concentrations under both treatments (Figure 3; Supplementary Table S2), suggesting that higher yield under the WW treatment was counterbalanced by reduced nutrient concentration, as previously reported for wheat (Marcos-Barbero et al., 2021). In the current study, significant positive associations were observed in most cases under both treatments between grain protein and mineral concentrations, as well as among the various grain minerals. Notably, highly significant positive correlations were found between protein and Zn and protein and Fe (Supplementary Table S2). Previously, it has been shown that Zn, Fe and protein are co-localized in the same grain fractions of wheat, leading to suggestion that grain protein acts as a sink for Zn and Fe (Cakmak et al., 2010). Similar correlations between grain mineral concentrations have been previously reported in tef (Ligaba-Osena et al., 2021) and wheat (Gomez-Becerra et al., 2010; Marcos-Barbero et al., 2021), suggesting synergic physiological mechanisms or common genetic control.
4.2 MTAs identified for grain nutrients
A total of 59 significant and 4 putative MTAs were identified for seed color and grain nutrients under WW and WL environments (Table 2; Supplementary Figure S3). MTAs were distributed across 19 out of the 20 tef chromosomes, the exception being Chr 9B (Figure 4). Similar to our previous tef GWAS on productivity and drought-adaptive traits (Alemu et al., 2024b), a greater number of significant MTAs were detected under the WL vs. WW treatment (Table 2). This suggests that genes induced by drought and possibly involved in drought-adaptive responses (Alemu et al., 2024b) influence nutrient uptake and accumulation in the grains. GWAS has been used in various crops to identify loci underlying grain quality (Kimani et al., 2020; Puranik et al., 2020; Chandra et al., 2024). However, reports on genomic dissection of nutritional quality in orphan crops are rather limited, and we are not aware of any previous genomic dissection of tef grain nutritional quality under contrasting water regimes.
4.3 Genomic loci highlight multiple-environment and pleiotropic effects
Five genomic regions reflected multiple-environment associations, including four cases of the same SNP (5A_7173734, 1B_15704188, 5A_6882876 and 1B_15526648) and one case of two closely linked SNPs (5A_7173788–7180196) associated with the same trait (protein, Fe, Cu, P and S, respectively) across two environments. Similarly, multiple-environment associations have been identified in tef for productivity and drought-adaptive traits across irrigation regimes (Alemu et al., 2024b), as well as for productivity and phenology across various locations in Ethiopia (Woldeyohannes et al., 2022). These multiple-environment associations reflect the effect of highly reliable constitutive genes underlying various grain quality traits, which are very likely to prove valuable for marker-assisted selection (McLeod et al., 2023) to improve tef adaptation to diverse environments.
Three genomic regions reflected pleiotropic effects, including one case on a single locus (6B_2255059) associated with both Zn (under WL) and K (under WW) and two groups of closely linked SNPs, 1B_15636841–15704188 and 5A_7173734–7180196, associated with Mg + Fe and protein + S, respectively. Whereas pleiotropism between Zn and K was supported by significant phenotypic association only under WL conditions (Figure 3; Supplementary Table S2), the other two pleiotropic effects were supported by highly significant correlations across both environments. Similarly, pleiotropic loci involving GY, lodging and heading time were identified in TDP-300 in our previous study (Alemu et al., 2024b). These pleiotropic relationships and significant associations suggest the existence of genetic correlations between traits (Mathew et al., 2019; Chebib and Guillaume, 2021; Alemu et al., 2024b). Pleiotropy can indicate a shared etiology between traits (Lee et al., 2021; Von Berg et al., 2022) due to common genetic factors, such as the same gene(s) controlling both traits or closely linked genes (Fang et al., 2017; Chebib and Guillaume, 2021).
Identifying genes associated with a specific trait is among the most concrete tasks in genomics and pre-breeding. In keeping the scope of this work to identify genomic regions associated with nutritional quality, we chose to address genes that are co-located with or nearest to multiple-environment or pleiotropic loci. These genes were found to play various biological functions related to stress resistance, membrane trafficking and electron transport (Supplementary Table S3). PTI1-like Tyrosine-Protein Kinase 1 is involved disease resistance signaling mechanism (Dang et al., 2019). Cell Division Control Protein 48 Homolog E-like (CDC48) plays a crucial role in protein quality control and regulation of immune responses in plants (Copeland et al., 2016). Polygalacturonase inhibitors (PGIPs) has a role in defending against pathogens, particularly by inhibiting enzymes that degrade plant cell walls (De Lorenzo and Ferrari, 2002). Although these three genes are not directly involved in grain quality, their role stress responses can indirectly influence grain development and quality. ADP-ribosylation factor GTPase-activating protein AGD is regulating nutrient transport, storage, and cellular organization in plants (Stefano et al., 2010). Similarly, cytochrome b-c1 complex subunit 6 is involved in energy metabolism through the mitochondrial electron transport chain (Smith et al., 2012). The last two genes, playing important roles in energy production, nutrient transport and storage, and metabolic processes, seem to be involved in grain development, nutrient accumulation and quality. It is our hope that future studies will use the data presented here to further explore the associated genes and their specific functions and support the development of high quality tef varieties.
5 Concluding remarks
Under the current climate change scenario, drought and heat stress are becoming the most severe constraints to crop production and quality, thus threating worldwide food and nutrition security. Therefore, a global focus on stress-resilient, high-yielding, nutritious crops is critical. Underutilized crops, such as tef, present an outstanding opportunity to increase crop diversity, promote climate resilience and boost nutritional quality. However, genomic and physiological dissection of tef under abiotic stress is still limited. In our previous study on tef, we focused on the genetic diversity and a genomic dissection of productivity and drought-adaptive traits (Alemu et al., 2024b), as well as on the whole plant’s physiological responses to drought (Alemu et al., 2024a). These are supplemented by the current work on nutrient concentrations in tef grains under contrasting water regimes.
Grain nutrient concentration depends on genotype, environment and management, all contributing to the particularly wide variation between genotypes, but also between grain colors and irrigation regimes. Both seed color and irrigation treatment exhibited an inconsistent effect (either increasing or decreasing) on grain nutrients, generally up to 10% in magnitude. As discussed above, it is interesting to note that Mn exhibited the greatest responses to both water regime and seed color.
Genomic loci detected in multiple environments reflect the stability and dependability of the genomic region for nutritional trait, whereas pleiotropic associations provide new insights into the complex genetic basis of grain nutrients, both can potentially facilitate breeding of novel tef cultivars. The pleiotropism and phenotypic associations between grain nutrients may facilitate simultaneous selection and improvement of tef nutritional properties.
Our previous (Alemu et al., 2024b) and current genomic dissection studies, as well as high throughput physiological phenotyping study (Alemu et al., 2024a), shed new light on tef responses to stress and may contribute to the main targets in tef breeding, as well as to further studies and a deeper understanding of tef genomics and physiology. Overall, tef productivity, drought-responsive traits and grain nutrient qualities are highly influenced by the environment and farming system. Although our studies were conducted in an irrigated Mediterranean environment, the genomic results may contribute to the development of drought-resistant, high-yielding and nutritious tef varieties, for rained conditions in Ethiopia and elsewhere.
Data availability statement
The datasets presented in this study can be found in online repositories. The names of the repository/repositories and accession number(s) can be found below: https://www.ncbi.nlm.nih.gov/, PRJNA1063642 and https://zenodo.org/records/14267904, for genomic and phenotypic data, respectively. Seeds of the tef diversity panel (TDP-300) are available upon reasonable request from the corresponding author.
Author contributions
MDA: Conceptualization, Formal analysis, Investigation, Methodology, Writing – review & editing, Data curation, Writing – original draft. SB-Z: Investigation, Writing – review & editing. VB: Investigation, Writing – review & editing, Methodology. YT: Investigation, Methodology, Writing – review & editing. IC: Investigation, Methodology, Writing – review & editing. YS: Conceptualization, Formal analysis, Funding acquisition, Investigation, Methodology, Project administration, Supervision, Validation, Writing – review & editing.
Funding
The author(s) declare financial support was received for the research, authorship, and/or publication of this article. This study was supported by The Israel Innovation Authority, Challenge Program (grant no. 73546).
Acknowledgments
We gratefully acknowledge the Heinrich Bonnenberg Scholarship awarded to MDA, and the Robert H. Smith Foundation for doctoral fellowships awarded to MDA and SB-Z. The authors wish to acknowledge the Israel Plant Gene Bank for providing the tef germplasm collection. We thank the Kvutzat Shiller farm for providing the platform for the field experiment. YS is the incumbent of the Haim Gvati Chair in Agriculture. This article was submitted as a preprint to BioRxiv as Alemu et al. (2024c).
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fpls.2024.1458408/full#supplementary-material
References
Abebe, Y., Bogale, A., Hambidge, K. M., Stoecker, B. J., Bailey, K., Gibson, R. S. (2007). Phytate, zinc, iron and calcium content of selected raw and prepared foods consumed in rural Sidama, Southern Ethiopia, and implications for bioavailability. J. Food Compos. Anal. 20, 161–168. doi: 10.1016/j.jfca.2006.09.003
Abewa, A., Adgo, E., Yitaferu, B., Alemayehu, G., Assefa, K., Solomon, J. K. Q., et al. (2019). Teff grain physical and chemical quality responses to soil physicochemical properties and the environment. Agronomy 9, 283. doi: 10.3390/agronomy9060283
Alejandro, S., Höller, S., Meier, B., Peiter, E. (2020). Manganese in plants: from acquisition to subcellular allocation. Front. Plant Sci. 11. doi: 10.3389/fpls.2020.00300
Alemu, M. D., Barak, V., Shenhar, I., Batat, D., Saranga, Y. (2024a). Dynamic physiological response of tef to contrasting water availabilities. Front. Plant Sci. 15. doi: 10.3389/fpls.2024.1406173
Alemu, M. D., Ben-Zeev, S., Hellwig, T., Barak, V., Shoshani, G., Chen, A., et al. (2024b). Genomic dissection of productivity, lodging, and morpho-physiological traits in Eragrostis tef under contrasting water availabilities. Plants People Planet 3, 10505. doi: 10.1002/ppp3.10505
Alemu, M. D., Ben-Zeev, S., Barak, V., Tutus, Y., Cakmak, I., Saranga, Y., et al. (2024c). Genomic loci associated with grain protein and mineral nutrients concentrations in Eragrostis tef under contrasting water regimes. bioRxiv, 2024.08.06.606859. doi: 10.1101/2024.08.06.606859
Allaby, R. (2021). First come, first served for ancient crops. Nat. Plants 7, 542–543. doi: 10.1038/s41477-021-00926-w
Allen, M., Poggiali, D., Whitaker, K., Marshall, T. R., Van Langen, J., Kievit, R. A. (2021). Raincloud plots: a multi-platform tool for robust data visualization. Wellcome Open Res. 4, 63. doi: 10.12688/wellcomeopenres.15191.2
Asefa, B. G., Tsige, F., Mehdi, M., Kore, T., Lakew, A. (2023). Rapid classification of tef [Eragrostis tef (Zucc.) Trotter] grain varieties using digital images in combination with multivariate technique. Smart Agric. Technol. 3, 100097. doi: 10.1016/j.atech.2022.100097
Assefa, K., Chanyalew, S., Tadele, Z. (2017). “Tef, Eragrostis tef (Zucc.) trotter,” in Millets and sorghum. Ed. Patil, J. V., 226–266, (Ames, IA, USA: Wiley-Blackwell). doi: 10.1002/9781119130765.ch9
Assefa, K., Yu, J., Zeid, M., Belay, G., Tefera, H., Sorrells, M. E. (2011). Breeding tef [ Eragrostis tef (Zucc.) trotter]: conventional and molecular approaches. Plant Breed. 130, 1–9. doi: 10.1111/j.1439-0523.2010.01782.x
Atwell, S., Huang, Y. S., Vilhjálmsson, B. J., Willems, G., Horton, M., Li, Y., et al. (2010). Genome-wide association study of 107 phenotypes in Arabidopsis thaliana inbred lines. Nature 465, 627–631. doi: 10.1038/nature08800
Baye, K., Mouquet-Rivier, C., Icard-Vernière, C., Picq, C., Guyot, J. (2014). Changes in mineral absorption inhibitors consequent to fermentation of E thiopian injera : implications for predicted iron bioavailability and bioaccessibility. Int. J. Food Sci. Technol. 49, 174–180. doi: 10.1111/ijfs.12295
Bekkering, C. S., Tian, L. (2019). Thinking outside of the cereal box: breeding underutilized (Pseudo)Cereals for improved human nutrition. Front. Genet. 10. doi: 10.3389/fgene.2019.01289
Ben Mariem, S., Soba, D., Zhou, B., Loladze, I., Morales, F., Aranjuelo, I. (2021). Climate change, crop yields, and grain quality of C3 cereals: A meta-analysis of [CO2], temperature, and drought effects. Plants 10, 1052. doi: 10.3390/plants10061052
Bonferroni, C. (1936). Teoria statistica delle classi e calcolo delle probabilita. Pubblicazioni R Ist. Super. Sci. Econ. E Commericiali Firenze 8, 3–62.
Brouder, S. M., Valenec, J. J. (2023). Nutrition of plants in a changing climate. In: Rengel, Z., Cakmak, I., White, P. J. (eds) Marschner’s Mineral Nutrition of Plants, 4th edn. Academic Press Elsevier Ltd, pp 723–750. doi: 10.1016/B978-0-12-819773-8.00023-X
Cakmak, I., Kutman, U. B. (2018). Agronomic biofortification of cereals with zinc: a review. Eur. J. Soil Sci. 69, 172–180. doi: 10.1111/ejss.12437
Cakmak, I., Pfeiffer, W. H., McClafferty, B. (2010). Review: biofortification of durum wheat with zinc and iron. Cereal Chem. 87, 10–20. doi: 10.1094/CCHEM-87-1-0010
Chadalavada, K., Guna, K., Kumari, B. R., Kumar, T. S. (2022). “Drought stress in sorghum: impact on grain quality,” in Climate change and crop stress. 113–134, (Amsterdam, Netherlands: Elsevier).
Chandra, A. K., Pandey, D., Sood, S., Joshi, D. C., Tiwari, A., Sharma, D., et al. (2024). Uncovering the genomic regions underlying grain iron and zinc content using genome-wide association mapping in finger millet. 3 Biotech. 14, 47. doi: 10.1007/s13205-023-03889-1
Chanyalew, S., Ferede, S., Damte, T., Fikre, T., Genet, Y., Kebede, W., et al. (2019). Significance and prospects of an orphan crop tef. Planta 250, 753–767. doi: 10.1007/s00425-019-03209-z
Chapman, M. A., He, Y., Zhou, M. (2022). Beyond a reference genome: pangenomes and population genomics of underutilized and orphan crops for future food and nutrition security. New Phytol. 234, 1583–1597. doi: 10.1111/nph.18021
Chebib, J., Guillaume, F. (2021). Pleiotropy or linkage? Their relative contributions to the genetic correlation of quantitative traits and detection by multitrait GWA studies. Genetics 219, iyab159. doi: 10.1093/genetics/iyab159
Cheng, A. (2018). Review: Shaping a sustainable food future by rediscovering long-forgotten ancient grains. Plant Sci. 269, 136–142. doi: 10.1016/j.plantsci.2018.01.018
Copeland, C., Woloshen, V., Huang, Y., Li, X. (2016). AtCDC48A is involved in the turnover of an NLR immune receptor. Plant J. 88, 294–305. doi: 10.1111/tpj.13251
Coskun, D., White, P. J. (2023). Ion-uptake mechanisms of individual cells and roots: short-distance transport. In: Rengel, Z., Cakmak, I., White, P. J. (eds) Marschner’s Mineral Nutrition of Plants, 4th edn. Academic Press Elsevier Ltd, pp 723–750. doi: 10.1016/B978-0-12-819773-8.00023-X
D’Andrea, A. C. (2008). T’ef (Eragrostis tef) in ancient agricultural systems of highland Ethiopia. Econ. Bot. 62, 547–566. doi: 10.1007/s12231-008-9053-4
Daba, T. (2017). Nutritional and soio-cultural values of teff (Eragrostis tef) in Ethiopia. Int. J. Food Sci. Nutr. 2, 50–57. https://api.semanticscholar.org/CorpusID:103115414
Dame, Z. T. (2020). Analysis of major and trace elements in teff (Eragrostis tef). J. King Saud Univ. - Sci. 32, 145–148. doi: 10.1016/j.jksus.2018.03.020
Dang, P. M., Lamb, M. C., Bowen, K. L., Chen, C. Y. (2019). Identification of expressed R-genes associated with leaf spot diseases in cultivated peanut. Mol. Biol. Rep. 46, 225–239. doi: 10.1007/s11033-018-4464-5
De Lorenzo, G., Ferrari, S. (2002). Polygalacturonase-inhibiting proteins in defense against phytopathogenic fungi. Curr. Opin. Plant Biol. 5, 295–299. doi: 10.1016/S1369-5266(02)00271-6
Dietz, K., Zörb, C., C., -M. (2021). Drought and crop yield. Plant Biol. 23, 881–893. doi: 10.1111/plb.13304
El Sabagh, A., Hossain, A., Barutçular, C., Islam, M. S., Ahmad, Z., Wasaya, A., et al. (2020). “Adverse effect of drought on quality of major cereal crops: implications and their possible mitigation strategies,” in Agronomic crops. Ed. Hasanuzzaman, M. (Springer Singapore, Singapore), 635–658. doi: 10.1007/978-981-15-0025-1_31
Ereful, N. C., Jones, H., Fradgley, N., Boyd, L., Cherie, H. A., Milner, M. J. (2022). Nutritional and genetic variation in a core set of Ethiopian Tef (Eragrostis tef) varieties. BMC Plant Biol. 22, 220. doi: 10.1186/s12870-022-03595-9
Fang, C., Ma, Y., Wu, S., Liu, Z., Wang, Z., Yang, R., et al. (2017). Genome-wide association studies dissect the genetic networks underlying agronomical traits in soybean. Genome Biol. 18, 161. doi: 10.1186/s13059-017-1289-9
FAO. (2020). The state of food security and nutrition in the world 2020. Rome, Italy: FAO. doi: 10.4060/ca9692en
FAO. (2023). The state of food security and nutrition in the world 2023. Rome, Italy: FAO. doi: 10.4060/cc3017en
Francavilla, A., Joye, I. J. (2020). Anthocyanins in whole grain cereals and their potential effect on health. Nutrients 12, 2922. doi: 10.3390/nu12102922
Galani, Y. J. H., Hansen, E.M.Ø., Droutsas, I., Holmes, M., Challinor, A. J., Mikkelsen, T. N., et al. (2022). Effects of combined abiotic stresses on nutrient content of European wheat and implications for nutritional security under climate change. Sci. Rep. 12, 5700. doi: 10.1038/s41598-022-09538-6
Gebru, Y. A., Hyun-II, J., Young-Soo, K., Myung-Kon, K., Kwang-Pyo, K. (2019). Variations in amino acid and protein profiles in white versus brown teff (Eragrostis tef) seeds, and effect of extraction methods on protein yields. Foods 8, 202. doi: 10.3390/foods8060202
Gomez-Becerra, H. F., Erdem, H., Yazici, A., Tutus, Y., Torun, B., Ozturk, L., et al. (2010). Grain concentrations of protein and mineral nutrients in a large collection of spelt wheat grown under different environments. J. Cereal Sci. 52, 342–349. doi: 10.1016/j.jcs.2010.05.003
Hager, A.-S., Wolter, A., Jacob, F., Zannini, E., Arendt, E. K. (2012). Nutritional properties and ultra-structure of commercial gluten free flours from different botanical sources compared to wheat flours. J. Cereal Sci. 56, 239–247. doi: 10.1016/j.jcs.2012.06.005
Huang, X., Han, B. (2014). Natural variations and genome-wide association studies in crop plants. Annu. Rev. Plant Biol. 65, 531–551. doi: 10.1146/annurev-arplant-050213-035715
Huang, M., Liu, X., Zhou, Y., Summers, R. M., Zhang, Z. (2019). BLINK: a package for the next level of genome-wide association studies with both individuals and markers in the millions. GigaScience 8:154. doi: 10.1093/gigascience/giy154
Hunter, D., Borelli, T., Beltrame, D. M. O., Oliveira, C. N. S., Coradin, L., Wasike, V. W., et al. (2019). The potential of neglected and underutilized species for improving diets and nutrition. Planta 250, 709–729. doi: 10.1007/s00425-019-03169-4
International Food Policy Research Institute (IFPRI). Global food policy report 2024: Food systems for healthy diets and nutrition. Washington, DC, USA: International Food Policy Research Institute. Available at: https://hdl.handle.net/10568/141760
Islam, A. S. M. F., Mustahsan, W., Tabien, R., Awika, J. M., Septiningsih, E. M., Thomson, M. J. (2022). Identifying the genetic basis of mineral elements in rice grain using genome-wide association mapping. Genes 13, 2330. doi: 10.3390/genes13122330
Jaiswal, V., Bandyopadhyay, T., Gahlaut, V., Gupta, S., Dhaka, A., Ramchiary, N., et al. (2019). Genome-wide association study (GWAS) delineates genomic loci for ten nutritional elements in foxtail millet (Setaria italica L.). J. Cereal Sci. 85, 48–55. doi: 10.1016/j.jcs.2018.11.006
Jifar, H., Assefa, K., Tadele, Z. (2015). Grain yield variation and association of major traits in brown-seeded genotypes of tef [Eragrostis tef (Zucc.)Trotter. Agric. Food Secur. 4, 7. doi: 10.1186/s40066-015-0027-3
Kamal, N. M., Gorafi, Y. S. A., Tomemori, H., Kim, J.-S., Elhadi, G. M. I., Tsujimoto, H. (2023). Genetic variation for grain nutritional profile and yield potential in sorghum and the possibility of selection for drought tolerance under irrigated conditions. BMC Genomics 24, 515. doi: 10.1186/s12864-023-09613-w
Kimani, W., Zhang, L.-M., Wu, X.-Y., Hao, H.-Q., Jing, H.-C. (2020). Genome-wide association study reveals that different pathways contribute to grain quality variation in sorghum (Sorghum bicolor). BMC Genomics 21, 112. doi: 10.1186/s12864-020-6538-8
Kjeldahl, J. (1883). Neue methode zur bestimmung des stickstoffs in organischen körpern. Z. Für Anal. Chem. 22, 366–382. doi: 10.1007/BF01338151
Lee, C. H., Shi, H., Pasaniuc, B., Eskin, E., Han, B. (2021). PLEIO: a method to map and interpret pleiotropic loci with GWAS summary statistics. Am. J. Hum. Genet. 108, 36–48. doi: 10.1016/j.ajhg.2020.11.017
Li, L., Zhang, H., Liu, J., Huang, T., Zhang, X., Xie, H., et al. (2023). Grain color formation and analysis of correlated genes by metabolome and transcriptome in different wheat lines at maturity. Front. Nutr. 10. doi: 10.3389/fnut.2023.1112497
Ligaba-Osena, A., Mengistu, M., Beyene, G., Cushman, J., Glahn, R., Piñeros, M. (2021). Grain mineral nutrient profiling and iron bioavailability of an ancient crop tef (Eragrostis tef). Aust. J. Crop Sci. 15, 1314–1324. doi: 10.21475/ajcs.21.15.10.p3264
Liu, J., Huang, L., Li, T., Liu, Y., Yan, Z., Tang, G., et al. (2021). Genome-wide association study for grain micronutrient concentrations in wheat advanced lines derived from wild emmer. Front. Plant Sci. 12. doi: 10.3389/fpls.2021.651283
Mabhaudhi, T., Chimonyo, V. G. P., Hlahla, S., Massawe, F., Mayes, S., Nhamo, L., et al. (2019). Prospects of orphan crops in climate change. Planta 250, 695–708. doi: 10.1007/s00425-019-03129-y
Marcos-Barbero, E. L., Pérez, P., Martínez-Carrasco, R., Arellano, J. B., Morcuende, R. (2021). Genotypic variability on grain yield and grain nutritional quality characteristics of wheat grown under elevated CO2 and high temperature. Plants 10, 1043. doi: 10.3390/plants10061043
Martínez-Martínez, R., Chávez-Servia, J. L., Vera-Guzmán, A. M., Aquino-Bolaños, E. N., Carrillo-Rodríguez, J. C., Pérez-Herrera, A. (2019). Phenotypic variation in grain mineral compositions of pigmented maize conserved in indigenous communities of Mexico. Maydica 64, 1–13.
Mathew, I., Shimelis, H., Shayanowako, A. I. T., Laing, M., Chaplot, V. (2019). Genome-wide association study of drought tolerance and biomass allocation in wheat. PloS One 14, e0225383. doi: 10.1371/journal.pone.0225383
McLeod, L., Barchi, L., Tumino, G., Tripodi, P., Salinier, J., Gros, C., et al. (2023). Multi-environment association study highlights candidate genes for robust agronomic quantitative trait loci in a novel worldwide Capsicum core collection. Plant J. 116, 1508–1528. doi: 10.1111/tpj.16425
Milla, R., Osborne, C. P. (2021). Crop origins explain variation in global agricultural relevance. Nat. Plants 7, 598–607. doi: 10.1038/s41477-021-00905-1
Nyiraguhirwa, S., Grana, Z., Ouabbou, H., Iraqi, D., Ibriz, M., Mamidi, S., et al. (2022). A genome-wide association study identifying single-nucleotide polymorphisms for iron and zinc biofortification in a worldwide barley collection. Plants 11, 1349. doi: 10.3390/plants11101349
Peleg, Z., Saranga, Y., Yazici, A., Fahima, T., Ozturk, L., Cakmak, I. (2008). Grain zinc, iron and protein concentrations and zinc-efficiency in wild emmer wheat under contrasting irrigation regimes. Plant Soil 306, 57–67. doi: 10.1007/s11104-007-9417-z
Peterson, B. G., Carl, P., Boudt, K., Bennett, R., Ulrich, J., Zivot, E., et al. (2014). PerformanceAnalytics: Econometric tools for performance and risk analysis. R Package Version 1.
Puranik, S., Sahu, P. P., Beynon, S., Srivastava, R. K., Sehgal, D., Ojulong, H., et al. (2020). Genome-wide association mapping and comparative genomics identifies genomic regions governing grain nutritional traits in finger millet (Eleusine coracana L. Gaertn.). Plants People Planet 2, 649–662. doi: 10.1002/ppp3.10120
R Core Team (2020). R: A language and environment for statistical computing. R Foundation for statistical Computing, Viena, Austria. Retrieved June 1, 2024, from https://www.r-project.org.
Remington, D. L., Thornsberry, J. M., Matsuoka, Y., Wilson, L. M., Whitt, S. R., Doebley, J., et al. (2001). Structure of linkage disequilibrium and phenotypic associations in the maize genome. Proc. Natl. Acad. Sci. 98, 11479–11484. doi: 10.1073/pnas.201394398
Saturni, L., Ferretti, G., Bacchetti, T. (2010). The gluten-free diet: safety and nutritional quality. Nutrients 2, 16–34. doi: 10.3390/nu2010016
Schmidt, S. B., Jensen, P. E., Husted, S. (2016). Manganese deficiency in plants: the impact on photosystem II. Trends Plant Sci. 21, 622–632. doi: 10.1016/j.tplants.2016.03.001
Sehgal, A., Sita, K., Siddique, K. H. M., Kumar, R., Bhogireddy, S., Varshney, R. K., et al. (2018). Drought or/and heat-stress effects on seed filling in food crops: impacts on functional biochemistry, seed yields, and nutritional quality. Front. Plant Sci. 9. doi: 10.3389/fpls.2018.01705
Shumoy, H., Pattyn, S., Raes, K. (2018). Tef protein: Solubility characterization, in-vitro digestibility and its suitability as a gluten free ingredient. LWT 89, 697–703. doi: 10.1016/j.lwt.2017.11.053
Siddique, K. H. M., Li, X., Gruber, K. (2021). Rediscovering Asia’s forgotten crops to fight chronic and hidden hunger. Nat. Plants 7, 116–122. doi: 10.1038/s41477-021-00850-z
Smith, P. M., Fox, J. L., Winge, D. R. (2012). Biogenesis of the cytochrome bc1 complex and role of assembly factors. Biochim. Biophys. Acta BBA - Bioenerg. 1817, 276–286. doi: 10.1016/j.bbabio.2011.11.009
Stagnari, F., Galieni, A., Pisante, M. (2016). Drought stress effects on crop quality. Water Stress Crop Plants Sustain. Approach 2, 375–392. doi: 10.1002/9781119054450.ch23
Stangoulis, J., Sison, C. (2008). Crop sampling protocols for micronutrient analysis. Harvest Plus Tech Monogr. Ser. 7, 1–20.
Stefano, G., Renna, L., Rossi, M., Azzarello, E., Pollastri, S., Brandizzi, F., et al. (2010). AGD5 is a GTPase-activating protein at the trans-Golgi network: AGD5’s role at the TGN in vivo. Plant J. 64, 790–799. doi: 10.1111/j.1365-313X.2010.04369.x
Stevens, G. A., Beal, T., Mbuya, M. N. N., Luo, H., Neufeld, L. M., Addo, O. Y., et al. (2022). Micronutrient deficiencies among preschool-aged children and women of reproductive age worldwide: a pooled analysis of individual-level data from population-representative surveys. Lancet Glob. Health 10, e1590–e1599. doi: 10.1016/S2214-109X(22)00367-9
Tadele, Z. (2019). Orphan crops: their importance and the urgency of improvement. Planta 250, 677–694. doi: 10.1007/s00425-019-03210-6
Tao, H., Dittert, K., Zhang, L., Lin, S., Römheld, V., Sattelmacher, B. (2007). Effects of soil water content on growth, tillering, and manganese uptake of lowland rice grown in the water-saving ground-cover rice-production system (GCRPS). J. Plant Nutr. Soil Sci. 170, 7–13. doi: 10.1002/jpln.200625033
Tietel, Z., Simhon, E., Gashu, K., Ananth, D. A., Schwartz, B., Saranga, Y., et al. (2020). Nitrogen availability and genotype affect major nutritional quality parameters of tef grain grown under irrigation. Sci. Rep. 10, 14339. doi: 10.1038/s41598-020-71299-x
Vadez, V., Grondin, A., Chenu, K., Henry, A., Laplaze, L., Millet, E. J., et al. (2024). Crop traits and production under drought. Nat. Rev. Earth Environ. 5, 211–225. doi: 10.1038/s43017-023-00514-w
VanBuren, R., Man Wai, C., Wang, X., Pardo, J., Yocca, A. E., Wang, H., et al. (2020). Exceptional subgenome stability and functional divergence in the allotetraploid Ethiopian cereal teff. Nat. Commun. 11, 884. doi: 10.1038/s41467-020-14724-z
VanRaden, P. M. (2008). Efficient methods to compute genomic predictions. J. Dairy Sci. 91, 4414–4423. doi: 10.3168/jds.2007-0980
Vavilov, N. I. (1951). The origin, variation, immunity and breeding of cultivated plants, Translated from the Russian by Chester KS. Soil Science. 72 (6), 482. doi: 10.1097/00010694-195112000-00018
Villanueva, M., Abebe, W., Pérez-Quirce, S., Ronda, F. (2022). Impact of the variety of tef [Eragrostis tef (Zucc.) trotter] on physical, sensorial and nutritional properties of gluten-free breads. Foods 11, 1017. doi: 10.3390/foods11071017
Von Berg, J., Ten Dam, M., van der Laan, S. W., De Ridder, J. (2022). PolarMorphism enables discovery of shared genetic variants across multiple traits from GWAS summary statistics. Bioinformatics 38, i212–i219. doi: 10.1093/bioinformatics/btac228
Wang, J., Zhang, Z. (2021). GAPIT version 3: boosting power and accuracy for genomic association and prediction. Genomics Proteomics Bioinf. 19, 629–640. doi: 10.1016/j.gpb.2021.08.005
Wickham, H., Chang, W., Wickham, M. H. (2016). Package ‘ggplot2.’. Create Elegant Data Vis. Using Gramm. Graph. Version 2, 1–189.
Woldeyohannes, A. B., Iohannes, S. D., Miculan, M., Caproni, L., Ahmed, J. S., De Sousa, K., et al. (2022). Data-driven, participatory characterization of farmer varieties discloses teff breeding potential under current and future climates. eLife 11, e80009. doi: 10.7554/eLife.80009
Xu, Z., Lai, T., Li, S., Si, D., Zhang, C., Cui, Z., et al. (2018). Promoting potassium allocation to stalk enhances stalk bending resistance of maize (Zea mays L.). Field Crops Res. 215, 200–206. doi: 10.1016/j.fcr.2017.10.020
Yazici, M. A., Asif, M., Tutus, Y., Ortas, I., Ozturk, L., Lambers, H., et al. (2021). Reduced root mycorrhizal colonization as affected by phosphorus fertilization is responsible for high cadmium accumulation in wheat. Plant Soil 468, 19–35. doi: 10.1007/s11104-021-05041-5
Zhu, F. (2018). Chemical composition and food uses of teff (Eragrostis tef). Food Chem. 239, 402–415. doi: 10.1016/j.foodchem.2017.06.101
Keywords: drought stress, GWAS, micronutrient, macronutrient, protein, seed color, tef
Citation: Alemu MD, Ben-Zeev S, Barak V, Tutus Y, Cakmak I and Saranga Y (2024) Genomic loci associated with grain protein and mineral nutrients concentrations in Eragrostis tef under contrasting water regimes. Front. Plant Sci. 15:1458408. doi: 10.3389/fpls.2024.1458408
Received: 08 July 2024; Accepted: 11 November 2024;
Published: 20 December 2024.
Edited by:
Ayalew Ligaba-Osena, University of North Carolina at Greensboro, United StatesReviewed by:
Fei Liu, Henan University, ChinaBullo Erena Mamo, University of California, Riverside, United States
Copyright © 2024 Alemu, Ben-Zeev, Barak, Tutus, Cakmak and Saranga. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Yehoshua Saranga, c2h1a2kuc2FyYW5nYUBtYWlsLmh1amkuYWMuaWw=
†ORCID: Muluken Demelie Alemu, orcid.org/0000-0003-1656-0996
Shiran Ben-Zeev, orcid.org/0000-0002-1133-5549
Yusuf Tutus, orcid.org/0000-0003-2438-2739
Ismail Cakmak, orcid.org/0000-0002-3183-5524
Yehoshua Saranga, orcid.org/0000-0002-1182-8730