 Markus Göbel1*
Markus Göbel1* Samiksha Dulal1
Samiksha Dulal1 Lea Sommer1
Lea Sommer1 Markus Weinmann2
Markus Weinmann2 Abdullah Al Mamun2
Abdullah Al Mamun2 Aneesh Ahmed2
Aneesh Ahmed2 Neerakkal Sujeeth3Karin Mai4
Neerakkal Sujeeth3Karin Mai4 Günter Neumann2
Günter Neumann2 Torsten Müller1
Torsten Müller1 Klára Bradáčová1
Klára Bradáčová1- 1Institute of Crop Science, Fertilization and Soil Matter Dynamics, University of Hohenheim, Stuttgart, Germany
- 2Institute of Crop Science, Nutritional Crop Physiology, University of Hohenheim, Stuttgart, Germany
- 3BioAtlantis Ltd., Clash Industrial Estate, Tralee, County Kerry, Ireland
- 4SP Sourcon Padena GmbH, Research and Development, Tübingen, Germany
Introduction: The production of high-quality food for the growing world population on the one hand and the reduction of chemical-synthetic pesticides on the other hand represents a major challenge for agriculture worldwide. The effectiveness of a combination of microbial and non-microbial biostimulants (BSs) with various nitrogen (N) forms in pathogen defense is discussed as a promising, but still poorly understood bio-based alternative for crop protection.
Methods: For this reason, nitrate and stabilized ammonium fertilizer both combined with a consortium of Pseudomonas brassicacearum, Bacillus amyloliquefaciens, and Trichoderma harzianum as soil treatment or with a mixture of seaweed extract (Ascophyllum nodosum) together with chitosan-amended micronutrient fertilizer as foliar spray application were compared under controlled greenhouse conditions. Furthermore, a combination of microbial and different non-microbial BSs (seaweed extracts + chitosan) and micronutrients with nitrate or with stabilized ammonium fertilizer was tested under field conditions to improve nutrient availability, promote plant growth, and suppress Zymoseptoria tritici (Zt) in winter wheat.
Results and discussion: While plant-protective effects against Zt by the microbial consortium application could be observed particularly under ammonium fertilization, the application of seaweed extract–chitosan mixture expressed plant defense against Zt more strongly under nitrate fertilization. In the field trial, the combination of microbial consortium with the seaweed extract–chitosan mixture together with micronutrients zinc (Zn) and manganese (Mn) showed positive effects against Zt under ammonium fertilization, associated with increased levels of defense metabolites. Furthermore, the additional input of Zn and copper (Cu) from the chitosan application improved the micronutrient status by minimizing the risk of Zn and Cu deficiency under controlled and field conditions. The use of BSs and the inoculation of Zt did not show any effects on plant growth and yield neither under controlled greenhouse conditions nor in the field. Summarized, microbial and non-microbial BSs separately applied or even combined together as one treatment did not influence plant growth or yield but made a positive contribution to an N form-dependent promotion of pathogen defense.
1 Introduction
One of the most critical fungal pathogens in wheat is Zymoseptoria tritici (Zt) (Quaedvlieg et al., 2011), which leads to yield losses of 5%–10% and reduction of food quality worldwide (Fones and Gurr, 2015). Intensive farming with the application of pesticides against pathogens such as Zt and chemical fertilizers is often regarded as necessary to feed the world’s growing population and to guarantee food security, but at the same time, this industrialized style of agriculture has a negative impact on the environment. Many of those pesticides are assumed to be readily degraded and removed from the soil. It was later shown that they can form bound residues, formerly undetected, and are even found as pesticide residues in food (Pogacean and Gavrilescu, 2009; Raliya et al., 2018). This agricultural challenge points out the relevance of an improved and sustainable cropping system avoiding excessive use of chemical-synthetic pesticides for environmental protection and high-quality food supply (Zimmermann et al., 2021). Targeted mineral fertilization with the focus on nitrogen (N) is most common for high plant growth and crop yield, even though its pathogen-suppressing effect is not less relevant (Huber and Haneklaus, 2007) but has been neglected in the past. However, it has been confirmed that adequate mineral N fertilization can improve plant tolerance to biotic and abiotic stresses (Fernandes and Rossiello, 1995; Walters and Bingham, 2007; Geisseler and Scow, 2014; Ding et al., 2021). Other studies proved the dependence of the effect of N fertilization on plant tolerance to the disease susceptibility of the crop varieties (Huber and Watson, 1974) or the importance of the source of N and the rate of application (Baker and Martinson, 1970; Huber and Watson, 1974). On the one hand, the impact of nitrate fertilization on plant tolerance correlates with the applied amount. The higher the N application is, the higher the susceptibility to certain diseases. Nevertheless, it has to be mentioned that the effect of nitrogen depends on the biology of the pathogen and the response of the crop plant. For example, a dense canopy formation can create a microclimate favorable for fungal infections. Furthermore, the nitrogen effect depends on the difference between biotrophic or saprophytic fungi and the influence of the nitrogen form (Weinmann et al., 2023). On the other hand, Sun et al. (2020) mentioned in their research that both ammonium and nitrate fertilization can promote or suppress plant diseases equally. This may depend on the type of pathogen and whether the ammonium fertilizer was stabilized or not, because of fast nitrification in aerated soils ammonium fertilizer show almost the same effect as nitrate. To combine the different N forms with microbial biostimulants (BSs), a consortium of plant-growth-promoting microorganisms (PGPMs) is supposed to be more effective than single PGPMs to strengthen plants against biotic and abiotic stresses (Mamun et al., 2024). Furthermore, a microbial consortium offers a broader range of usage than single strains due to the versatile compounds which are present in the consortium (Sarma et al., 2015; Bradáčová et al., 2019). Bradáčová et al. (2020) and Mamun et al. (2024) additionally identified that the form of N supply can be decisive for the efficiency of microbial BSs. Stabilized ammonium nutrition in combination with microbial consortium of Pseudomonas spp., Bacillus spp., and Trichoderma spp. improved the uptake of ammonium-N and led to increased phosphorus (P) concentrations in maize shoot tissues compared to nitrate supply. Increased root length was also observed with stabilized ammonium supply compared to nitrate supply. In addition, Halpern et al. (2015) recorded many positive abilities of a diversity of microorganisms, such as increased root growth due to the production of phytohormones, higher phosphate availability through mineralization of organic P, solubilization of iron (Fe) through chelating siderophores produced by the bacteria, and enhanced accumulation of Zn and potassium (K) in the plant tissue by excreting organic acids resulting in higher plant uptake and yield. Non-microbial BSs such as chitin, chitosan, or seaweed extracts also perform more efficiently as combined treatments to improve disease tolerance according to several studies (Nanda et al., 2021). Apart from stress priming effects, seaweed extracts in higher concentrations can contribute to enhanced solubility of micronutrients chelating them by large organic molecules and affect the nutrient uptake (Halpern et al., 2015). Furthermore, it is reported that chitosan can reduce the water content in cells due to its hydrophilic nature to mitigate stress and increases root length by acting as an additional source of carbon, resulting in improved nutrient uptake (Shahrajabian et al., 2021). Regarding the application of the micronutrients Zn and Mn, a Mn-dependent superoxide dismutase (MnSOD) is produced as a defense reaction by plants due to biotic and abiotic stresses (Kumar et al., 2020). Furthermore, Mn is involved in the synthesis of toxic phenols and lignification by forming a physical barrier against fungal pathogens. Similar to Mn, Zn acts as a cofactor for enzymes such as superoxide dismutase, and can increase the production of total phenolics or total antioxidants resulting in protective plant responses (Silva et al., 2022). In this multicomponent approach, the effect of microbial (Pseudomonas brassicacearum, Bacillus amyloliquefaciens, and Trichoderma harzianum) and non-microbial (seaweed extracts + chitosan) BSs and micronutrients (Zn and Mn) on Zt affected winter wheat with different N supplies was investigated. To examine both, the plant growth promoting potential and the plant protecting potential of biostimulants (BSs) combined with ammonium or nitrate fertilization, nutrient analyses, and metabolic assessments were performed. As complex treatments were tested in this study, it is hypothesized that beneficial effects on plant performance, nutrient acquisition, and tolerance of wheat plants result from a complexity of interactions of these treatments with the plant and its environment. However, the experimental setups do not allow for an isolation of effects that could be clearly attributed to certain components of the complex treatments.
We hypothesized that:
1. the application of different N forms together with a microbial consortium or seaweed extract in combination with chitosan improves plant growth and development by enhanced root growth and nutrient uptake under biotic stress conditions in the greenhouse.
2. the combination of stabilized ammonium fertilizers with microbial BSs, non-microbial BSs, and micronutrients leads to increased plant growth and enhanced disease suppression of Zt-inoculated wheat plants in the field.
3. different N forms combined with various BSs alleviate the effects of Zt infestation by inducing increased production of defense metabolites under greenhouse and field conditions.
The overall objective of this study was to examine the effect of different N forms in combination with different BSs on the suppression of Zt.
1.1 Pot experiment
1.1.1 Experimental setup
Winter wheat Triticum aestivum L. Asory (SECOBRA Saatzucht GmbH, Unterschleißheim, Germany), a medium-late ripening variety and showing a moderately tolerance to Zt, was cultivated from 05/05/2022 to 29/06/2022 (55 days) in a greenhouse at a mean temperature of 28.7°C and a mean relative humidity of 45.7%. Plastic cylinders with a diameter of 95 mm (pot surface 71 cm2), and a height of 210 mm were used as pots. Each pot contained 1,173 g soil (Filderlehm 2015) from the experimental station Heidfeldhof of the University of Hohenheim) and 587 g quartz sand mixture (0.6 – 1.2 mm) (66.5:33.5 w:w). An overview on physical and chemical soil properties is shown in Supplementary Table 1. Per pot, 12 wheat seeds were sown. A 100-g layer of washed quartz sand (0.6 – 1.2 mm) was added on top of the soil surface to reduce evaporation and pest pressure and avoid siltation by irrigation. Each pot was fertilized with 120 mg P kg−1 DW in the form of Ca(H2PO4)2, 150 mg K kg−1 DW in the form of K2SO4, and 50 mg Mg kg−1 DW in the form of MgSO4 and sprayed on the soil–sand mixture while the substrate was mixed by hand before filling into the pots. The two different nitrogen treatments, calcium nitrate (YaraTera® CALCINIT®, YARA GmbH & Co. KG, Dülmen, Germany) and ammonium sulfate (NovaTec® Solub 21, COMPO EXPERT GmbH, Münster, Germany) with a nitrification inhibitor [3,4-dimethyl-1H-pyrazole phosphate (DMPP)], were fertilized each with 100 mg kg−1 DW as milled powder mixed with the soil–sand mixture before filling into the pots. The pots were regularly watered by weight up to 70% of the water holding capacity (WHC) when the water content in the soil/sand substrate fell below 50% of the WHC. The pots were arranged according to a split-plot design with two real repetitions and two main plots per repetition (Supplementary Figure 1). In total, 50 pots were prepared, 10 different variants with five repetitions per treatment (Supplementary Table 2). Within the main plots, treatments were arranged on two tables (table one with two replicates and table two with three replicates) as randomized complete block design (RCBD). The model for this design was
with μ as the overall effect, ti as the fixed/random effect of the ith table, rij as the fixed effect of the jth block on the ith table, τk as the effect of the kth treatment, gtli as the random effect of the tth main plot in the jth block on the ith table, and eijkl as the error of yijkl.
1.1.2 Application of biostimulants
As biostimulant products, a microbial consortium composed of different microorganisms and a mixture of seaweed extract with chitosan was tested. The consortium (Pseudomonas brassicacearum: 2 × 1010 cfu g−1, Bacillus amyloliquefaciens: 2 × 1010 cfu g−1, and Trichoderma harzianum: 1 × 108 cfu g−1, SP Sourcon Padena GmbH, Tübingen, Germany) formulated with milk powder that served as a carrier and nutrient medium was applied as a soil application on the sowing day shortly before sowing. The 10-g package of consortium powder (for the 107 cm2 soil surface) was dissolved into 500 ml of distilled water. From the stock solution (SL), 35.5 µl (0.05 µl cm−2) were pipetted and mixed in a beaker with 100 ml of distilled water using a magnetic stirrer. For each pot, 10 ml SL (0.14 ml cm−2) was dropped onto the top layer of the soil and then carefully mixed into the soil by hand. The seaweed extract (Ascophyllum nodosum extract, BioAtlantis Ltd., Tralee, Ireland) together with chitosan as a micronutrient formulation with surfactants containing the mixture of micronutrients together with copper (Cu), manganese (Mn), molybdenum (Mo), and zinc (Zn); 7.1 N (0.55 NH4-N+6.55 NH2N) + 11.8 K + 3.4 S + <0.1 Cl + 1.42 Cu + 2.84 Mn + 2.23 Zn + 0.028 Mo w/v [g 100 ml−1 chitosan formulation] (Wuxal® Micromix plus Chitosan; A7116, AGLUKON Spezialdünger GmbH & Co. KG, Düsseldorf, Germany), were applied as foliar spraying. The total amounts of mineral nutrients applied to the plants via chitosan are shown in Supplementary Table 3. In accordance with the BioAtlantis’ application recommendation, the seaweed + chitosan mixture was applied 2 days prior to a biotic stress event and every 5 days thereafter if the disease intensity was >25% of infested leaf area or every 7 days thereafter if the disease spread was <25% of infested leaf area with 2 l seaweed extract ha−1 and 2 l chitosan ha−1 (0.02 µl cm−2) in 250 l water ha−1 (2.5 µl water cm−2). In total, five applications have been performed during the pot experiment. An SL was prepared as follows: 15.62 µl of the seaweed extract was pipetted in one 2 ml Eppendorf tube and the same amount of chitosan in another 2 ml Eppendorf tube, both filled up with distilled water to 1 ml. Both tubes were properly mixed by use of an orbital shaker, then 968 µl of distilled water was added in each and stirred again. Finally, the contents of both Eppendorf tubes were combined with 200 ml of distilled water in a glass beaker and mixed with a magnetic stirrer. Of this application mixture, 18.2 ml was sprayed onto the leaves of the plants of each pot.
1.1.3 Zymoseptoria tritici cultivation, inoculation, and disease assessment
YMDA–agar YMB–liquid culture medium as the most efficient culture media for inoculum production according to Saidi et al. (2012) was selected for the Zt cultivation. 4 g Bacto™ Yeast Extract Technical (A288620; Becton, Dickinson and Company, Franklin Lakes, United States) 4 g Bacto™ Malt Extract (A218630; Becton, Dickinson and Company, Franklin Lakes, United States), 10 g (D(+)-glucose 1-hydrate), (A143140.1211; AppliChem GmbH, Darmstadt, Germany), and 15 g Bacter Agar (A0949, 1000; AppliChem GmbH, Darmstadt, Germany) were dissolved in 1 l of distilled water. The solution was mixed properly with a magnetic stirrer and autoclaved (121°C, 20 min). Afterwards, the solution was poured into sterile Petri dishes and solidified during cooling. With a flame-sterilized spatula, a piece of Zt fungal mycelium was gouged out and placed vice versa on the agar-medium, and the Petri dishes were locked with parafilm and stored in a light cabinet for 1–3 weeks at room temperature (18°C–20°C), with the culture medium facing the above. The YMB–liquid culture medium was prepared, consisting of 4 g YE (Bacto™ Yeast Extract Technical, A288620; Becton, Dickinson and Company, Franklin Lakes, United States), 4 g ME (Bacto™ Malt Extract, A218630; Becton, Dickinson and Company, Franklin Lakes, United States), and 10 g glucose (D(+)-glucose 1-hydrate) (A143140.1211; AppliChem GmbH, Darmstadt, Germany) filled up to 1 l with distilled water. The solution was mixed with a magnetic stirrer and autoclaved (121°C, 20 min). In a 1 l Erlenmeyer flask, two complete Zt plates were mixed with 500 ml of liquid YMB medium. In a 0.5 l Erlenmeyer flask, one complete Zt plate was mixed with 333 ml liquid YMB medium. The Zt fungi on the plates were gouged out with a flame-sterilized spatula and cut into pieces before adding them to the liquid medium. Everything together was mixed in a sterile cabinet and put on a shaker with a frequency of 100 - 125 rounds per minute (rpm) for 2 weeks at 18°C–20°C. Thereafter, the YMB medium was poured through an autoclaved, two-layered gauze bandages into an autoclaved vessel. The residue represented the mycelium, and the filtrate contained the Zt spores. The filtrate was centrifuged at 20°C for 10 min and 6,000 min-1 (17,307×g) (Sorvall RC 6 Plus centrifuge, Thermo Electron LED GmbH, Osterode, Germany), and the supernatant was discarded. The remaining Zt spores were then diluted with autoclaved water, filled into tubes, and mixed with an orbital shaker. The spore suspension was filled from the tubes into 0.5 l PET bottles which were stored at −40°C until they were used. The Zt spores were counted with a “Fuchs-Rosenthal” cell chamber. Three 2 ml Eppendorf tubes with a 1:1,000 concentration of Zt spores were used, and 3.2 µl was pipetted in the counting chamber. There were 16 group squares existing in the chamber; one group square included 16 small squares. For every sample, five group squares were counted and the mean value was determined. Lastly, the mean value from all three counts was 374.8 × 107 spores ml−1. Zt was applied with a small sprayer and a concentration of 1 × 107 spores ml−1 Zt suspension 15 days after sowing (DAS). 355 μl Zt suspension/pot (5 μl cm−2 soil surface) was applied. The pots were covered for 3 days with four different foil tunnels with wet towels inside to keep a high relative humidity. For keeping the humidity even higher, a transparent plastic foil was additionally placed over the tunnels. Visual assessment of Zt infestation was done 29, 34, 43, 49, and 55 DAS. The percentage of leaf area covered with Zt for six youngest, fully developed leaves from six different plants per pot were evaluated based on the method of James W.C. described by Eyal et al. (1987).
1.1.4 Plant analyses
The dry weight of the aboveground plant tissue and root samples per pot was determined after harvest at 55 DAS. Aliquots of washed root samples were stored in 70% (v/v) ethanol and analyzed for root length and morphological structure (i.e., root diameter classes) with an Epson Expression 10000XL scanner (Seiko Epson K.K., Japan) using WinRHIZO root analysis software package (Regent Instruments Inc., Quebec, Canada) (Moradtalab et al., 2020). For estimating the shoot and root dry weights, the plant tissue and root samples packed in paper bags were oven-dried at 60°C for 4 days and then weighed.
1.1.5 Determination of stress metabolites
Fresh leaf samples from 27 DAS and 55 DAS were used for the determination of selected physiological stress indicators such as hydrogen peroxide (H2O2), ascorbate peroxidase (APX; EC 1.11.1.11) activity, and guaiacol peroxidase (GPX; EC 1.11.1.7) activity after homogenization of 0.1 g of plant tissues shock-frozen in liquid nitrogen in 1.5 ml of 50 mM potassium phosphate extraction buffer, followed by centrifugation for 10 min at 14,000 min−1 (20,160×g) (Hettich centrifuge MIKRO 24-48 R, Tuttlingen, Germany). APX activity was recorded spectrophotometrically at 290 nm according to the method of Boominathan and Doran (2002). H2O2 levels were determined spectrophotometrically at 390 nm according to the method described by Moradtalab et al. (2018). GPX activity was performed spectrophotometrically at 470 nm using the tetra-guaiacol assay described by Moradtalab et al. (2020). For the determination of total antioxidants, leaf samples from 27 DAS were shock-frozen and homogenized in liquid nitrogen to use 0.1 g of fresh matter for methanolic extraction (80% v/v methanol) followed by centrifugation for 10 min at 14,000 min−1 (20,160×g) (Hettich MIKRO 24-48 R centrifuge, Tuttlingen, Germany). The 1,1-diphenyl-2-picrylhydrazyl radical (DPPH)-modified method was used to evaluate the free radical scavenging activity of antioxidants in the plant tissue (Moradtalab et al., 2020). A U-3300 spectrophotometer (Hitachi, Tokyo, Japan) was used for all spectrophotometric measurements.
1.1.6 Analysis of mineral nutrients
The concentrations of the essential nutrients N, P, K, Ca, Mg, S, Zn, Mn, Fe, and Cu were determined in the oven-dried and milled plant tissues of final harvest (55 DAS). The shoots were ground in a disc-oscillating agate stone mill (SIEBTECHNIK GmbH, Mülheim-Ruhr, Germany) for 3 min – 4 min to a fine powder. 0.2 g of powdered plant tissue per sample was weighed into a quartz glass beaker, and 1 ml ultrapure water and 2.5 ml nitric acid (ROTIPURAN® ≥65%, p.a., ISO; Carl Roth GmbH & Co. KG) were added. After microwave (“ultraCLAVE III”; Fa. MLS Leutkirch) digestion was accomplished, elemental concentrations were measured by inductive coupled plasma optical emission spectrometry (ICP-OES; “Agilent 5110”, Santa Clara, United States). Total nitrogen, carbon, and sulfur were determined with the Vario MAX CNS elemental analyzer (Elementar Analysensysteme GmbH, Langenselbold, Germany) (VDLUFA, 2012).
1.1.7 Statistical evaluation
Statistical analyses were performed using SAS/STAT software package of SAS® 9.4 (2016) (SAS Institute Inc., Cary, USA). A one-way ANOVA followed by a Tukey test (p < 0.05 significance level) was used to compare means for statistically significant differences. Data are presented as mean values. Normal distributions and variance homogeneities of the residuals were checked by the Shapiro–Wilk test and by Levene’s test, respectively, and graphically against the predicted values by QQ plots, histograms, and graphs of the residuals according to Kozak and Piepho (2017).
1.2 Field experiment
1.2.1 Experimental setup
Similar to the pot experiment, winter wheat of the variety Asory (SECOBRA Saatzucht GmbH, Unterschleißheim, Germany) was the test plant in the field experiment. This variety is described as moderately tolerant to Zt with medium plant length and standability, and medium-late ripening quality. It was cultivated from 20/10/2021 to 27/07/2022 (280 days) with a sowing density of 330 grains m-2 at the experimental station Heidfeldhof of the University of Hohenheim, Filderhauptstraße 201, 70599 Stuttgart, Germany (48°42′56.21 N, 9°11′15.64 E, 402 m above sea level, mean temperature of 8.8°C, mean relative humidity of 76.6%). A 3.28 ha field was used for the experiment with a plot size of 6 m × 8 m (48 m2). The sowing was performed with Amazone AD 303 (Amazonen-Werke H. Dreyer SE & Co. KG, Hasbergen, Germany) with a 3 m working width and front cultivator, a 3.5 cm sowing depth, and a 12.5 cm row spacing. The soil texture in the field was loamy clay, and the soil type was Cambisol. The field was tilled with a rotary harrow twice before sowing. Five days after sowing and during field emergence (BBCH 23), weeding/hoeing was undertaken to control the weeds, except for the negative control variant 27. Soil sampling for Nmin measurements was performed during the vegetation period for the whole field for fertilizer requirement calculation on 03/03/2022 (Nmin = 38.46 kg N ha−1). Furthermore, after harvesting, plot-specific sampling for Nmin analyses was performed on 11/08/2022. One sampling was performed to collect soil samples from 0–30-cm, 30–60-cm, and 60–90-cm depths according to VDLUFA (2023). The results of the soil analyses are shown in Supplementary Table 4. The fertilizer "P, K, Mg plus S – fertilizer" containig 4.4 P +12.5 K + 3 Mg + 12 S (w/w) [kg 100 kg−1] (Beiselen GmbH, Ulm, Germany) was applied, broadcast twice: once 380 kg fertilizer ha−1 before tillage in September 2021 and 324 kg fertilizer ha−1 at the beginning of the vegetation (BBCH 22) on 03/03/2022 with Rauch Aero 1110 pneumatic spreader (Rauch Landmaschinenfabrik GmbH, Rheinmünster, Germany). In total, 31 kg P ha−1, 88 kg K ha−1, 21 kg Mg ha−1, and 84 kg S ha−1 were fertilized. The two different nitrogen treatments, calcium nitrate (YaraTera® CALCINIT®, YARA GmbH & Co. KG, Dülmen, Germany) and ammonium sulfate (NovaTec® Solub 21, COMPO EXPERT GmbH, Münster, Germany) with nitrification inhibitor [3,4-dimethyl-1H- pyrazole phosphate (DMPP)], were fertilized each with application rates of 240 kg N ha−1 also with the Rauch Aero 1110 pneumatic spreader. The application of calcium nitrate was divided into three applications according to farming practice. The first one with 40% of the total amount of calcium nitrate (80.6 kg N ha−1) at BBCH 23 on 10/03/2022, the second one with 43% (86.7 kg N ha−1) at BBCH 30 on 22/04/2022, and the third one with 17% (34.3 kg N ha−1) at BBCH 33–37 on 20/05/2022.
The application of ammonium sulfate was divided into two applications: the first one with 60% of the total amount of ammonium sulfate (120.9 kg N ha−1) at BBCH 23 on 10/03/2022, and the second one with 40% (80.6 kg N ha−1) at BBCH 33–37 on 20/05/2022. The herbicide Artus® (Cheminova Deutschland GmbH & Co. KG, Stade, Germany) was sprayed with 50 g ha−1 at BBCH 23; the herbicides Biathlon® 4D + Dash® E.C. (70 g ha-1 + 1 l ha-1; BASF SE, Limburgerhof, Germany) were sprayed together at BBCH 30. The herbicides were mixed with 300 l of water using an Amazone UF 901 (Amazonen-Werke H. Dreyer SE & Co. KG, Hasbergen, Germany) field sprayer (nozzle type IDKN 120-03, 3–4 bar, application volume 300 l ha−1, driving speed 5 km/h; Lechler GmbH, Metzingen, Germany). Variants treated with herbicides were also treated with the calcium nitrate fertilizer and therefore performed as a positive control treatment. The experimental design was a row-column design latinized in columns and blocks (Richter et al., 2009) (Supplementary Figure 2). There were 28 different variants with four repetitions for each sharing 112 plots in total (Supplementary Table 5). Ten variants were investigated in this study.
1.2.2 Application of biostimulants
As BS products, a combined treatment of seaweed extract + chitosan mixture, micronutrients Zn and Mn, and a milk powder as a carrier medium for the microbial consortium of different microorganisms was used. The consortium (Pseudomonas brassicacearum: 2 × 1010 cfu g−1, Bacillus amyloliquefaciens: 2 × 1010 cfu g−1, and Trichoderma harzianum: 1 × 108 cfu g−1, SP Sourcon Padena GmbH, Tübingen, Germany) formulated with milk was applied as a soil application on the sowing day shortly before sowing. A 5 g package of pure milk powder (for 500 m2) as a blank control and a 14 g package of milk powder mixed with the consortium (for 1,400 m2) were each dissolved into 500 ml of distilled water and filled up to 20 l of water to prepare the SL. The quantities applied were 1,920 ml SL milk powder plot-1 and 685.7 ml SL milk powder + consortium plot-1 each with three watering cans/plot. Each watering can was filled with 640 ml SL milk powder or 228.6 ml SL milk powder + consortium both along with 10 l of water applied simultaneously per plot in the longitudinal direction at first, and then perpendicular to the first application in the transverse direction of the plot, to ensure an even distribution of the products. The seaweed extract (Ascophyllum nodosum extract, BioAtlantis Ltd., Tralee, Ireland) together with WUXAL® MICROMIX plus Chitosan (AGLUKON Spezialdünger GmbH & Co. KG, Düsseldorf, Germany) were applied as foliar application with an Amazone UF 901 (Amazonen-Werke H. Dreyer SE & Co. KG, Hasbergen, Germany) field sprayer (nozzle type IDKN 120-03, 3–4 bar, application volume 300 l ha−1, driving speed 5 km/h; Lechler GmbH, Metzingen, Germany). The total amounts of mineral nutrients applied to the plants via chitosan are shown in Supplementary Table 6. In accordance with AGLUKON’s application recommendation, the seaweed + chitosan mixture to be applied at shooting (BBCH 30) and at ear pushing (BBCH 51) was 2 l seaweed extract ha−1 and 2 l chitosan ha−1 (0.2 ml m−2) in 250 l water ha-1 (25 ml water m-2). An SL for 49 half plots was prepared as follows: 235.2 ml of seaweed extract and 235.2 ml of chitosan were filled up with distilled water to 1 l. The seaweed + chitosan solution got mixed with a magnetic stirrer and was filled in the tank of an Amazone UF 901 field sprayer with 29.4 l of water and 0.6 l seaweed + chitosan solution (25 ml m−2). Then, the SL was sprayed on the leaves of the plants per plot. Micronutrients Zn and Mn (Lebosol®-Zink700SC; 40% total Zn as zinc oxide 700 g Zn l−1; Lebosol®-Mangan500SC; 27.9% total Mn as manganese carbonate 500 g Mn l−1; Lebosol® Dünger GmbH, Elmstein, Germany) were used for foliar application with an Amazone UF 901 field sprayer (nozzle type IDKN 120-03, 3–4 bar, application volume 300 l ha−1, driving speed 5 km/h; Lechler GmbH, Metzingen, Germany). In accordance with the manufacturer’s application recommendation, the Zn and Mn to be applied about 10 days after the start of vegetation, at BBCH 14 (3–4 days before Zt inoculation) were each 0.3 l Zn ha−1 and 0.5 l Mn ha−1 in 200 l of water (1.4 ml Zn plot-1 and 2.4 ml Mn plot-1 in 1 l of water), at BBCH 16 and at shooting/extension (BBCH 31) each 0.5 l Zn ha−1 and 0.75 l Mn ha−1 in 200 l of water (2.4 ml Zn plot-1 and 3.6 ml Mn plot-1 in 1 l of water).
1.2.3 Zymoseptoria tritici cultivation, inoculation, and disease assessment
The process of Zt cultivation was performed as described in the pot experiment (see 1.1.3) based on Saidi et al. (2012). Zt was applied with a Hege 76 field sprayer (device carrier from Wintersteiger, Ried im Innkreis, Austria; field sprayer from Kubota, Osaka, Japan) at BBCH 23-24 (160 DAS) and a concentration of 1 × 107 spores ml−1 Zt suspension. 100 ml Zt suspension m−2 (4.8 l plot-1) was applied. The Zt disease incidence was determined visually by evaluating the percentage of infested plants inside a 60 × 40 cm rectangle formed with a meter stick at BBCH 30-31 (190 DAS), BBCH 31-33 (204 DAS), and BBCH 75 (259 DAS) modified according to Moll et al. (2000).
1.2.4 Plant analyses
The dry weight of five fully unfolded leaves from five different plants per plot at BBCH 33-37 (209 DAS) and three root samples per plot at BBCH 99 (278 DAS) were determined. To ensure the extraction of representative root samples, the shovel was placed carefully leaving sufficient space along the side and the puncture was created deep to the full height of the shovel blade (30 cm) to dig fresh roots out. The determination of the leaf and root dry weight as well as of the root length and morphological structure took place as in the pot experiment (see 1.1.4). Furthermore, for estimating grain nutritional analyses, grain samples (400 - 500 g per plot) from harvest at BBCH 99 (280 DAS) were oven-dried at 40°C for 4 days and then weighed.
1.2.5 Determination of stress metabolites
Five fully unfolded leaves without visible disease symptoms from five different plants per plot from BBCH 75 (245 DAS) were used for the determination of selected physiological stress indicators with the methods as described in the pot experiment (see 1.1.5).
1.2.6 Analysis of mineral nutrients
The concentrations of the essential nutrients N, P, K, Ca, Mg, S, Zn, Mn, Fe, and Cu were determined in oven-dried and milled leaf samples from BBCH 33-37 (209 DAS) and in 400 g–500 g oven-dried and milled grain samples per plot from harvest at BBCH 99 (280 DAS). Grinding of the leaf samples was the same process as for the pot experiment (see 1.1.6). 0.2 g of powdered leaf material and powdered grain material was weighed into a quartz glass beaker followed by the process of element analysis, which took place as described in the pot experiment (see 1.1.6).
1.2.7 Statistical evaluation
Statistical analyses were performed using SAS/STAT software package of SAS® 9.4 (2016) (SAS Institute Inc., Cary, USA). A one-way ANOVA followed by a Tukey test (p < 0.05 significance level) was used to compare means for statistically significant differences. Data are presented as mean values. Normal distributions and variance homogeneities of the residuals were checked by the Shapiro–Wilk test and by Levene’s test, as well as graphically against the predicted values by QQ plots, histograms, and graphs of the residuals according to Kozak and Piepho (2017).
2 Results
2.1 Pot experiment
2.1.1 Disease severity affected by different N supplies and microbial and non-microbial biostimulants
The Zt disease severity (DS) increased over time following a biphasic pattern with a slower relative increase of 25%–27% between 14 and 28 days after inoculation (DAI) and a steeper increase by 48%–53% over the next 12 days, finally reaching comparatively low absolute DS levels of 15% at the end of the culture period without N-form-dependent differences (Figure 1).
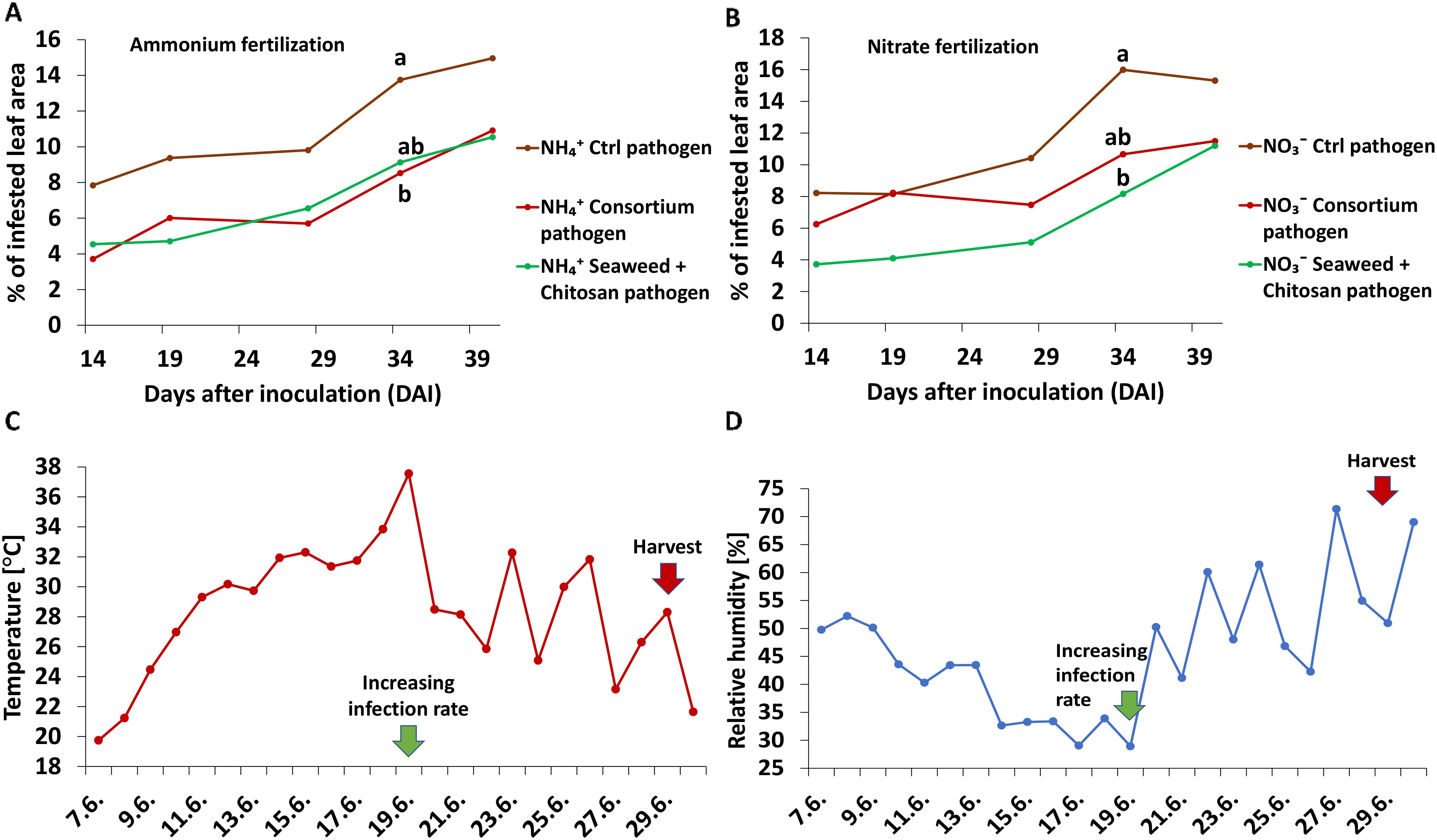
Figure 1. (A, B) Time course of disease spread of Zymoseptoria tritici (Zt) [% of infested leaf area] on winter wheat plants in the greenhouse treated with ammonium sulfate or calcium nitrate under control condition (brown lines) or with different biostimulants (microbial consortium, red lines; seaweed extract + chitosan, green lines) both inoculated with Zt. (C) Temperature course [°C] and (D) relative humidity course [%] over the plant growth period. Increasing infection rate from 19.06.2022 on. (A, B) represent mean values of five replicates per treatment. Mean values with at least one same or without lowercase letters within graph (A, B) are not significantly different according to Tukey test (α=0.05).
Protective effects of microbial consortium (MC) were particularly expressed under the ammonium nutrition [average DS suppression rate 40% (NH4+) vs. 22% (NO3−)] over the culture period. Furthermore, a clear trend although not significant at all time-points was detected (Figures 2A, B).
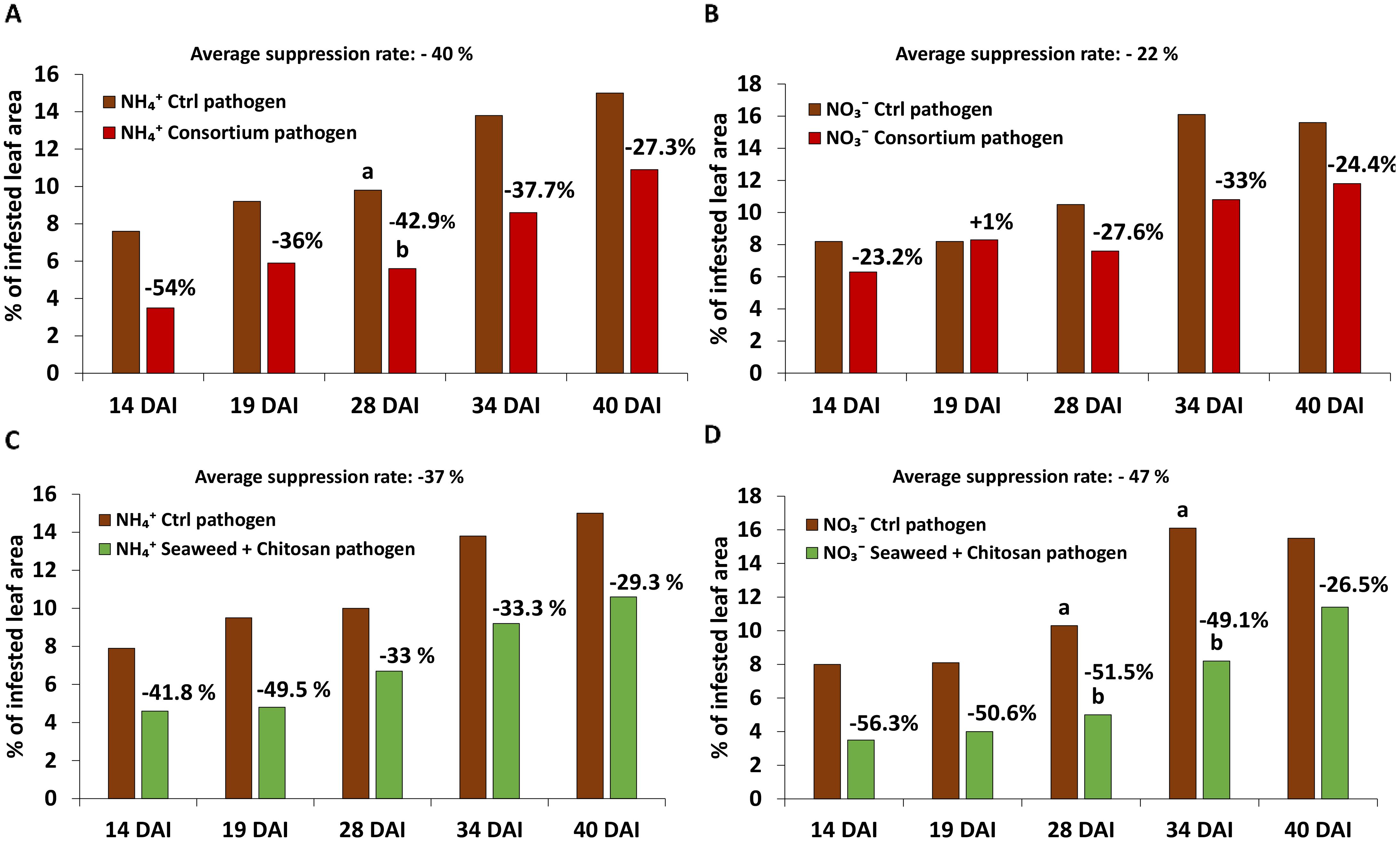
Figure 2. Disease severity [% of infested leaf area] of Zymoseptoria tritici (Zt) on winter wheat plants in the greenhouse 14, 19, 28, 34 and 40 days after inoculation (DAI) treated with (A, C) ammonium sulfate and (B, D) calcium nitrate, both under control condition (brown bars) and with different biostimulants (microbial consortium, red bars; seaweed extract + chitosan, green bars) inoculated with Zt. (A–D) represent mean values of five replicates per treatment. Mean values with the same or without lowercase letters within each graph are not significantly different according to Tukey test (α=0.05).
Protective effects of seaweed extract + chitosan (SC) appeared particularly under nitrate nutrition [average DS suppression rate 47% (NO3−) vs 37% (NH4+)]. Similar to MC, differences declined by the end of the culture period (Figures 2C, D).
2.1.2 Defense metabolites
MC: At final harvest (55 DAS, 40 DAI), a trend of increased leaf H2O2 concentrations by 47% (Figure 3A) was associated with a similar but significant increase in APX activity (47%, Figure 3B) mediating H2O2 detoxification in MC-inoculated plants with NH4+ fertilization. No stimulatory effects of MC inoculation on H2O2 accumulation and APX activity were recorded under NO3− supply (Figures 3A, B), but APX activity was generally significantly increased under NO3− supply compared with plants supplied with NH4+ fertilization (Figure 3B). No comparable effects were detected at earlier developmental stages (27 DAS, 12 DAI), for GPX activity or for total antioxidants mediating non-enzymatic detoxification of reactive oxygen species (ROS) (Supplementary Figure 3).
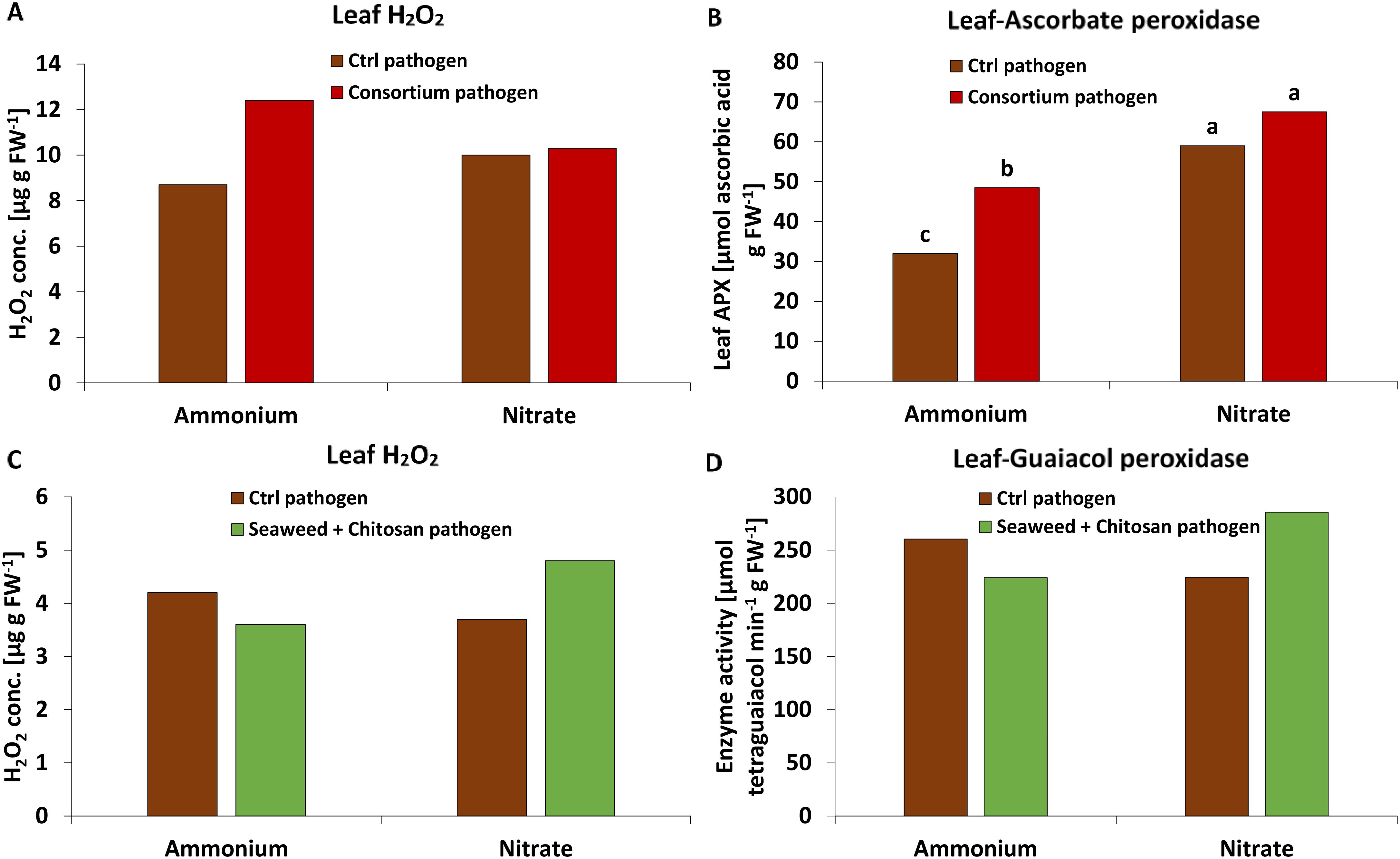
Figure 3. (A, C) Hydrogen peroxide (H2O2) concentration [ug g FW-1], (B) ascorbate peroxidase (APX) activity [μmol ascorbic acid g FW-1] and (D) guaiacol peroxidase (GPX) activity [μmol tetraguaiacol min-1 g FW-1] in the leaf tissue of winter wheat plants in the greenhouse treated with ammonium sulfate or calcium nitrate under control condition (brown bars) or with different biostimulants (microbial consortium, red bars (55 DAS); seaweed extract + chitosan, green bars (27 DAS)) both inoculated with Zymoseptoria tritici (Zt). A represents mean values of three replicates in the ammonium control and ammonium- consortium treatments, five replicates in the nitrate control and nitrate-consortium treatments. (B) represents mean values of three replicates in the ammonium control and ammonium-consortium treatment, five replicates in the nitrate control and four replicates in the nitrate-consortium treatment. (C, D) represent mean values of five replicates per treatment. Mean values with the same or without lowercase letters within each graph are not significantly different according to Tukey test (α=0.05).
SC: In contrast to MC-inoculated plants, earlier changes in defense metabolites were detectable already at 27 DAS (12 DAI). In plants with NO3− fertilization, H2O2 accumulation in the leaf tissue tended to increase by 30% (Figure 3C). This was associated with an increase (27%) in the activity of GPX (Figure 3D). No comparable effects were detectable under NH4+ fertilization (Figures 3C, D), at later developmental stages (55 DAS, 40 DAI) or for APX activity. Also, no significant differences were detectable for total antioxidants (Supplementary Figure 4).
2.1.3 Mineral nutritional status
For all treatments, the nutritional status of N, P, S, Mg, Fe, and Mn at final harvest was sufficient for adequate growth of wheat plants. Critical nutrient concentrations close or even below the reported deficiency threshold values (Bergmann, 1992) were recorded for K, Ca, Zn, and Cu. N form effects were detected for P, S, and Zn with increased leaf concentrations under NH4+ fertilization and for Ca, Mg, K, and Mn concentrations promoted under NO3− supply (Supplementary Table 7).
No nutritional benefits were recorded in response to inoculation with MC. By contrast, SC applications increased the concentrations of Zn and Cu (Figures 4A, C) above the deficiency thresholds (Bergmann, 1992). The plants with nitrate fertilization exhibited particularly high Mn shoot concentrations, which were further increased by SC application (Figure 4B).

Figure 4. (A) Zinc (Zn), (B) manganese (Mn), (C) copper (Cu) concentration [mg kg DW-1] in the shoot tissue of winter wheat plants in the greenhouse 55 days after sowing (DAS) treated with ammonium sulfate or calcium nitrate under control condition (brown bars) or with seaweed extract + chitosan (green bars) both inoculated with Zymoseptoria tritici (Zt). The dashed lines show the nutrient deficiency limits according to Bergmann (1992). (A–C) represent mean values of five replicates per treatment. Mean values with the same lowercase letters within each graph are not significantly different according to Tukey test (α=0.05).
2.1.4 Plant growth
Neither the pathogen inoculation nor the application of biostimulants affected shoot and root biomass production, total root length, or root diameter (Figure 5). A significant decline in shoot biomass (14%) was recorded in pathogen-infected plants with NO3− supply as compared with NH4+ fertilization (Figure 5A).
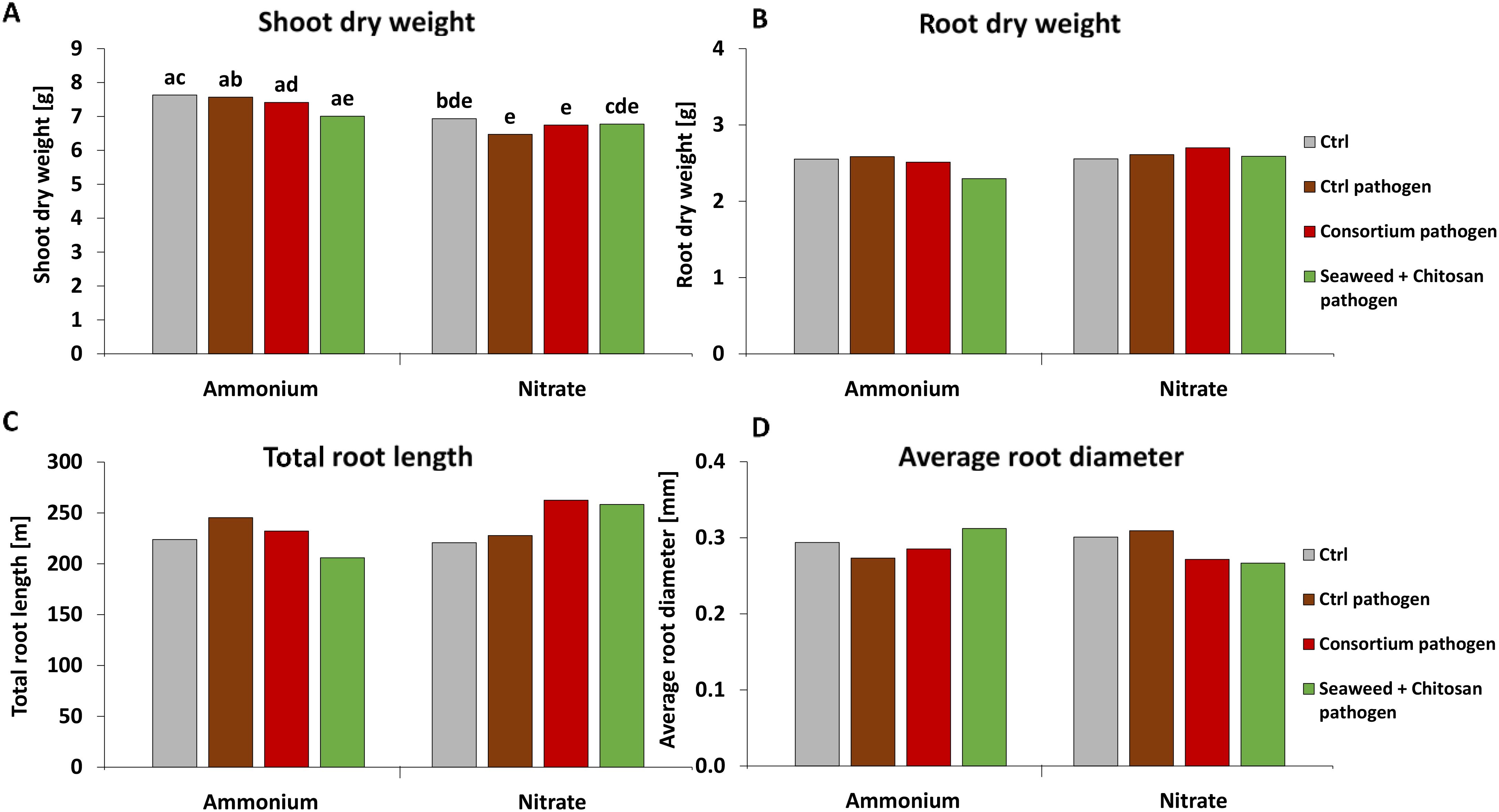
Figure 5. (A) Shoot and (B) root dry weight, (C) total root length and (D) average root diameter of winter wheat plants in the greenhouse 55 days after sowing (DAS) treated with ammonium sulfate or calcium nitrate under control condition with or without Zymoseptoria tritici (Zt) inoculum or with different biostimulants (microbial consortium, red bars; seaweed extract + chitosan, green bars) both inoculated with Zt. (A–D) represent mean values of five replicates per treatment. Mean values with at least one same or without lowercase letters within each graph are not significantly different according to Tukey test (α=0.05).
2.2 Field experiment
2.2.1 Disease incidence affected by combination of biostimulants
Due to the fact that no significant differences in Zt disease incidence (DI) between the Zt- and non-infected plants could be determined, all results with applied Zt inoculum and natural infestation were combined to a pool of Zt-inoculated and naturally infested variants in the following graphs. Independent of Zt inoculation, at 190 DAS (April, BBCH 30-31), first leaf blotch symptoms with a disease incidence of 5%–6% became detectable in all treatments with the exception of the control without N fertilization where DI reached already 13% (Figure 6A). A massive increase of DI was recorded within the next 10 weeks until 259 DAS (July, BBCH 75). Maximum DI values of approximately 65% were recorded in plants without N fertilization but similarly also under nitrate fertilization with or without application of biostimulants. Ammonium fertilization significantly reduced DI by 10% and the lowest DI values below 50% were recorded for BS-treated plants with NH4+ supply (Figure 6B).
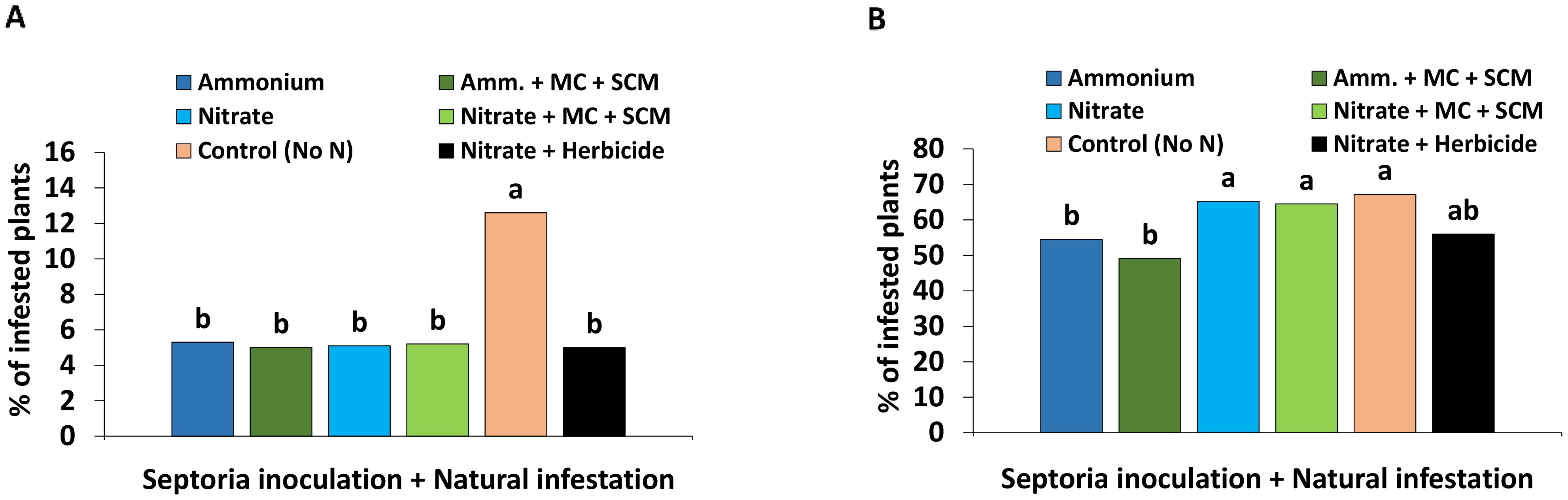
Figure 6. Disease incidence (A) 190 days after sowing (DAS) and (B) 259 days after sowing (DAS) [% of infested plants] of winter wheat plants in the field treated with either ammonium sulfate (Amm.) or calcium nitrate (Nitrate). In addition, both fertilizers were combined with a microbial consortium (MC) and seaweed extract + chitosan + micronutrients zinc + manganese (SCM). (A, B) depict a pool of Zymoseptoria tritici (Zt) inoculated and natural infested treatments. A negative control without nitrogen (N) (orange bars) and a positive control (black bars) both with natural infestation are included. (A, B) represent seaweed extract + chitosan not yet applied at the time of bonituring. (A, B) represent mean values of eight replicates per treatment. Mean values with at least one same lowercase letter within each graph are not significantly different according to Tukey test (α=0.05).
2.2.2 Defense-related metabolites
The leaf H2O2 accumulation significantly increased by approximately 50% in the BS-treated plants as compared with the remaining treatments associated also with the highest APX activity (Figures 7A, B). Compared with plants supplied with nitrate fertilization, ammonium fertilization tended to increase also GPX activity and accumulation of total antioxidants (Figures 7C, D).
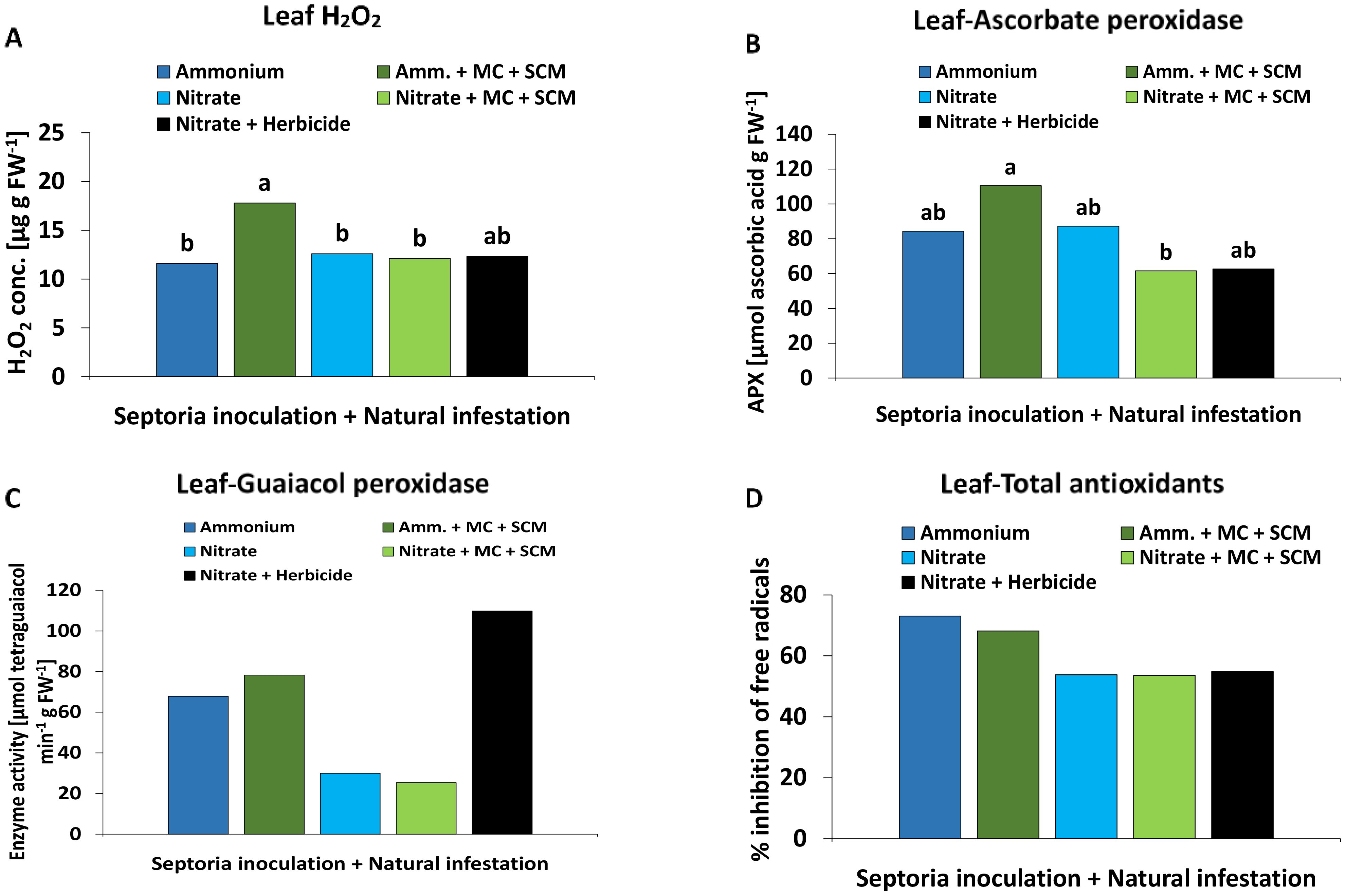
Figure 7. (A) Hydrogen peroxide (H2O2) concentration [μg g FW-1], (B) ascorbate peroxidase (APX) activity [μmol ascorbic acid g FW-1], (C) guaiacol peroxidase (GPX) activity [μmol tetraguaiacol min-1 g FW-1] and (D) total antioxidant potential [% inhibition of free radicals] in the leaf tissue of winter wheat plants in the field 245 days after sowing (DAS) treated with either ammonium sulfate (Amm.) or calcium nitrate (Nitrate). In addition, both fertilizers were combined with a microbial consortium (MC) and seaweed extract + chitosan + micronutrients zinc + manganese (SCM). (A–D) depict a pool of Zymoseptoria tritici (Zt) inoculated and natural infested treatments. A positive control (black bars) with natural infestation is included. (A–D) represent mean values of eight replicates per treatment. Mean values with at least one same or without lowercase letters within each graph are not significantly different according to Tukey test (α=0.05).
2.2.3 Mineral nutritional status
With the exception of Zn, the mineral nutritional status was sufficient for all investigated nutrients (N, P, K, S, Ca, Mg, Fe, Mn, Cu) in all plants which received N fertilization. However, negative controls without N supply showed multiple nutrient deficiencies with respect to N, K, Ca, Mg, and Zn but also decreased leaf concentrations of the remaining nutrients except Cu. Compared with NO3− fertilization, NH4+ fertilization decreased the leaf concentrations of N, K, Mg, and Ca (summarized in Supplementary Figure 5). The only recorded BS effect was reflected by a significantly increased Zn concentration in grains in the ammonium-treated variant. Additionally, NH4+ fertilization significantly increased grain concentrations of Mn (Figure 8).
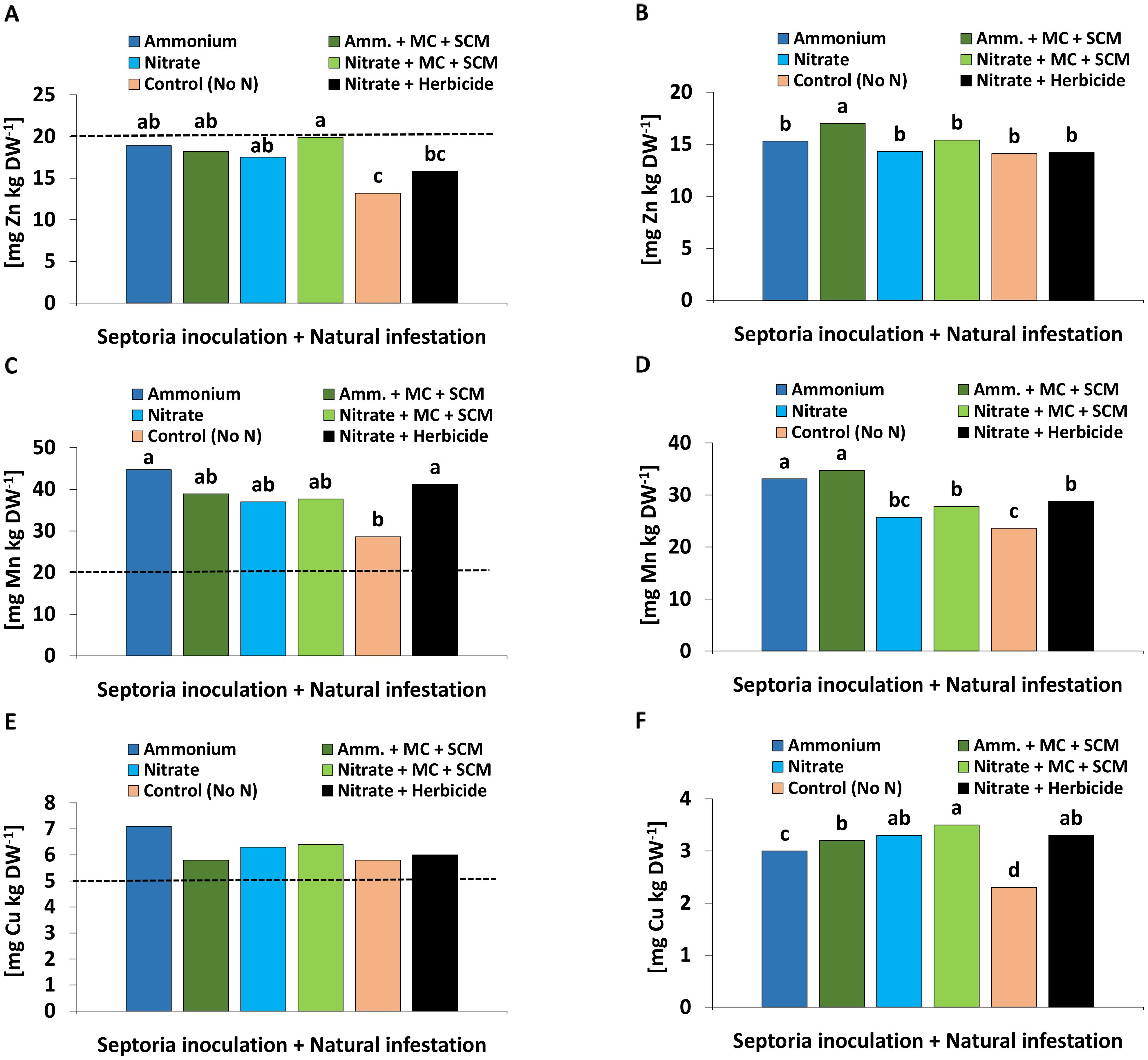
Figure 8. (A) Zinc (Zn), (C) manganese (Mn), (E) copper (Cu) concentration [mg kg DW-1] in the leaf tissue 209 days after sowing (DAS) and (B) Zn, (D) Mn, (F) Cu concentration [mg kg DW-1] in the grain 280 days after sowing (DAS) of winter wheat plants in the field treated with either ammonium sulfate (Amm.) or calcium nitrate (Nitrate). In addition, both fertilizers were combined with a microbial consortium (MC) and seaweed extract + chitosan + micronutrients zinc + manganese (SCM). A-F depict a pool of Zymoseptoria tritici (Zt) inoculated and natural infested treatments. A negative control without nitrogen (N) (orange bars) and a positive control (black bars) both with natural infestation are included. (A, C, E) represent seaweed extract + chitosan not yet applied at the time of bonituring. The dashed lines show the nutrient deficiency limits according to Bergmann and Neubert (1976). (A–F) represent mean values of eight replicates per treatment. Mean values with at least one same or without lowercase letters within each graph are not significantly different according to Tukey test (α=0.05).
2.2.4 Grain yield, grain protein, and root growth characteristics
A high grain yield with an average of 9.2 t ha−1 was achieved in all treatments except the negative control without N supply, which showed a 40% reduction (Figure 9A). In all treatments receiving N fertilization, approximately 12% grain protein content was recorded with the lowest values in BS-treated plants under NH4+ fertilization. In the negative control without N fertilization, the grain protein content reached only 8% (Figure 9B).
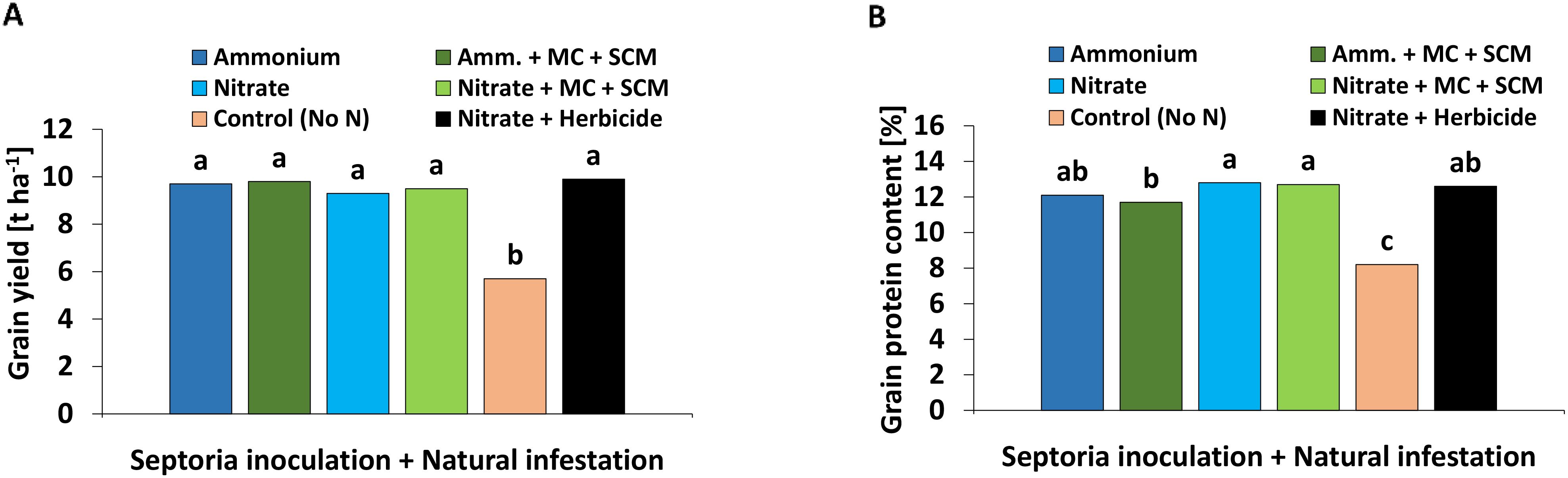
Figure 9. (A) Total grain yield [t ha-1] and (B) grain protein content [%] 280 days after sowing (DAS) of winter wheat plants in the field treated with either ammonium sulfate (Amm.) or calcium nitrate (Nitrate). In addition, both fertilizers were combined with a microbial consortium (MC) and seaweed extract + chitosan + micronutrients zinc + manganese (SCM). (A, B) depict a pool of Zymoseptoria tritici (Zt) inoculated and natural infested treatments. A negative control without nitrogen (N) (orange bars) and a positive control (black bars) both with natural infestation are included. (A, B) represent mean values of eight replicates per treatment. Mean values with at least one same lowercase letter within each graph are not significantly different according to Tukey test (α=0.05).
For the characterization of root morphological characteristics, root length in different root diameter classes was investigated from excavated root systems. For all treatments, the proportion of fine roots (0 - 0.2 mm diameter) was very similar and comprised between 30% and 40% of the total root length (Supplementary Figure 6).
3 Discussion
3.1 Disease spread affected by form and amount of N supply
In the greenhouse experiment, disease spreading showed a biphasic pattern, starting with a slow increase of approximately 25% during 14 DAI–28 DAI followed by a stronger spreading of approximately 50% during the next 12 days (Figures 1A, B). This may be attributed to unfavorable conditions with respect to temperature (25°C–36°C) and relative humidity (<50%) during the initial phase of pathogen infection (Eyal et al., 1987; Shaw, 1991; Fones et al., 2017). Accordingly, disease spread subsequently increased with declining temperature (<25°C) and increasing relative humidity (>50%) (Figures 1C, D), finally reaching a moderate disease severity (DS) of 15% independent of the N form supply (NO3− vs NH4+ during the first 8 weeks of plant development).
Similarly also under field conditions, a low disease incidence (DI) of 5%–6%, which was independent of the applied N form (Figure 6A), was observed during early growth of winter wheat in spring until BBCH 30-31, associated with generally suboptimal temperatures of <15°C for Zt infection during this time period (Eyal et al., 1987). During the next 10 weeks, a steep DI increase (Figure 6B) coincided with conductive conditions, characterized by increasing temperatures, relative humidity values of 60%–90%, and precipitation above the long-term average (164%–168%) in April and June (Supplementary Figure 7). However, in later stages of plant development (BBCH 75), a clear N form effect was detectable with a maximum DI of 65% and the highest leaf N concentrations in plants with NO3− fertilization. By contrast, a significantly lower DI (55%) and a lower leaf N status were recorded in plants with NH4+ supply (Figure 6B; Supplementary Figure 5A). High levels of N fertilization can improve the N-nutritional status and promote plant growth, but at the expense of reduced formation of lignin and waxy cuticles acting as physical barriers for pathogen penetration (Sun et al., 2020). Accordingly, also Simón et al. (2003) and Harrat and Bouznad (2014) reported increased disease spread for Zt in wheat, associated with increased N supply. In our experiment, the lower N status of plants supplied with stabilized NH4+ fertilization may be related to reduced N availability due to stronger adsorption and lower mobility of NH4+ in soils compared to NO3− fertilizers (Marschner and Rengel, 2023). Accordingly, a stimulation of oxidative stress defense, lignification, and accumulation of epicuticular waxes under NH4+ fertilization was reported by Wang et al. (2010) and Blanke et al. (1996).
Increased DI values were similarly recorded also in wheat plants without N fertilization both during early growth (BBCH 30-31, Figure 6A) and in later stages of plant development (BBCH 75, Figure 6B). A massive decline in grain yield (Figure 9A) associated with leaf concentrations of N, K, Mg, Ca, and Zn below the reported deficiency thresholds (Bergmann, 1992) suggests that the respective plants were obviously affected by multiple nutrient deficiencies, weakening the expression of defense responses against pathogens.
3.2 Disease spread affected by microbial and non-microbial biostimulants
A microbial PGPM consortium (MC) derived from a combination of bacterial and fungal PGPM strains (Pseudomonas, Bacillus, Trichoderma) and a non-microbial BS combination product (SC) based on Ascophyllum nodosum seaweed extract, chitosan, and stress-protective micronutrients (Zn, Mn, Cu) were used for the experiments. The selection was based on literature reports suggesting synergistic or complementary benefits of BS combinations to protect plants against various biotic and abiotic stress factors (Zaim et al., 2018; Bradáčová et al., 2019; Gunupuru et al., 2019; Karuppiah et al., 2019; Moradtalab et al., 2020) to cover a wider range of environmental conditions.
In the greenhouse experiment, protective effects against Zt leaf blotch were recorded for both MC and SC treatments (Figure 2). In contrast to the untreated controls (see. 4.1), disease severity was differentially affected by the form of N supply in MC- and SC-treated plants. While the MC formulation was more effective in plants with NH4+ supply, the non-microbial SC combination responded faster compared with MC and showed stronger suppressive effects under NO3− fertilization (Figure 2).
3.2.1 Oxidative burst
Similar to DS, also physiological stress defense responses to BS applications were differentially influenced depending on the form of N supply. Ammonium-fertilized plants specifically responded to MC application with a selective increase in leaf H2O2 accumulation by almost 50%. A similar trend was recorded after SC application in plants with NO3− supply (Figure 3). This may reflect the locally increased ROS (H2O2) production of pathogen-infected tissues (oxidative burst), frequently recorded as a first defense line against invading pathogens (Choudhary et al., 2017). Accordingly, the capacity for H2O2 production during the oxidative burst seems to be one factor determining Zt resistance in wheat genotypes (Shetty et al., 2003). Interestingly, after application of PGPMs or non-microbial biostimulants, the opposite response, characterized by improved ROS detoxification and declining tissue concentrations of H2O2, is frequently reported in plants affected by abiotic stress (Orzali et al., 2017; EL Mehdi EL Boukhari et al., 2020). However, in the presence of pathogens, also defense priming via increased PGPM-induced H2O2 production (mediated, e.g., by bacterial surfactants), has been described (Zhu et al., 2022). This scenario applies similarly also for the SC components chitosan (Vasil’ev et al., 2009) and Ascophyllum nodosum seaweed extract (Cook et al., 2018), suggesting selective effects of the investigated microbial and non-microbial BSs, depending on biotic vs abiotic stress factors.
3.2.2 Detoxification of reactive oxygen species
In parallel with the stimulation of H2O2 accumulation, MC inoculation also increased the activity of APX mediating the enzymatic degradation of H2O2 (Caverzan et al., 2012) preferentially in wheat plants with NH4+ fertilization (Figure 3B). This effect may protect non-infected tissues located close to the infection sites from oxidative stress associated with the oxidative burst. Systemic induction of APX and other enzymes involved in ROS detoxification is a well-known response not only to inoculation with PGPM strains of Pseudomonas, Bacillus, or Trichoderma (reviewed by Kumudini and Patil, 2021) but also to treatments with non-microbial BSs such as seaweed extracts (EL Mehdi EL Boukhari et al., 2020) or chitosan (Orzali et al., 2017). In contrast to MC-treated plants, no comparable increase in APX activity was recorded after SC application. However, under NO3− fertilization, the activity of GPX, known to play a central role in pathogen defense (Prakasha and Umesha, 2016), was increased in SC-treated plants. Similar to APX, also GPX activity might be expected to reduce the level of ROS by metabolizing H2O2. However, GPX is also capable of catalyzing various oxidase reactions leading to H2O2 generation and is involved in lignification, biosynthesis of ethylene, wound healing, and polysaccharide cross-linking (Sharma et al., 2012).
3.2.3 Micronutrient status
SC application increased the plant micronutrient status, thereby reducing Zn and Cu deficiencies (Bergmann, 1992) of the investigated plants (Figure 4). However, this effect was not detected in response to MC inoculation (Supplementary Figure 8). Deficiency of Zn and Cu was likely related to the neutral soil pH promoting the fixation of these micronutrients (Marschner, 1995). Supplementation by foliar micronutrient supply via the SC mixture/treatment obviously provided these micronutrients as important cofactors for enzymatic ROS detoxification and the GPX-mediated reactions described above (Datnoff et al., 2007).
An extraordinarily high plant-Mn status, far above the reported deficiency threshold (Bergmann, 1992), was recorded in the pot experiment, which was even further increased by SC application under NO3− fertilization (Figure 4B). In plants with NH4+ fertilization, this increase was likely counteracted by cation competition between uptake of NH4+ and Mn2+ (Marschner, 1995). The high Mn status may reflect an exceptionally high Mn availability in the respective soil, which was probably caused by long-time exposure to high greenhouse temperatures of 25°C–35°C (Figure 1C) reported to increase soil Mn availability (Reid and Racz, 1985). This may be mediated by stimulation of reductive processes such as increased respiratory oxygen consumption and modifications of microbial communities involved in Mn mobilization at higher soil temperatures especially when there is too much water and/or soil substrate compaction in the pot (Sparrow and Uren, 1987). A protective effect of Mn nutrition on controlling root or foliar diseases of plants (e.g., powdery mildew, downy mildew, take-al) is well documented (reviewed by Datnoff et al., 2007). Accordingly, also Eskandari et al. (2018) and Eskandari et al. (2020) reported improved resistance of cucumber to powdery mildew and anthracnose after foliar application of Mn, reaching similar Mn leaf concentrations >100 mg kg−1 DM as recorded in SC-treated plants in our study (Figure 4). The protective effects were attributed to improved lignification associated with increased activities of guaiacol peroxidase and phenol oxidase and increased callose production as mechanical barriers against fungal infection (Eskandari et al., 2018, 2020).
Taken together, the results suggest complementary protective effects of the MC and SC formulations against Zt leaf blotch, influenced by different forms of N fertilization. The defense responses, systemically induced by the MC formulation under NH4+ supply may be related to beneficial effects of NH4+ fertilizers on the establishment of plant microbial interactions. Therefore, PGPM inoculants, based on strains of Pseudomonas, Bacillus, and Trichoderma may promote pathogen suppression, e.g., via improved root colonization, increased auxin production or proliferation of root hairs as potential infection sites (Bradáčová et al., 2019; Mpanga et al., 2019; Moradtalab et al., 2020). The SC formulation additionally provides a source of micronutrients (Zn, Mn, Cu) with essential functions as cofactors for various physiological defense responses to pathogen infection. This is particularly important under conditions of limited micronutrient solubility at neutral to alkaline soil pH, further promoted by root-induced rhizosphere alkalization induced by NO3− fertilization (Marschner, 1995).
3.2.4 Field experiment
Under field conditions, a combined application of MC and SC with the addition of micronutrients zinc + manganese (MC-SCM application) was performed to exploit the potential benefits arising from complementary effects detected in the greenhouse experiment. At 259 DAS (BBCH 75), the highest DI of 65% was recorded under NO3− fertilization without any benefits by MC-SCM application (Figure 6). By contrast, MC-SCM-treated plants supplied with NH4+ fertilization showed the lowest DI (<50%) associated with increased H2O2 accumulation and APX activity in the leaf tissue (Figure 7), similar to the MC responses in the greenhouse experiment. However, compared with NO3− fertilization, the DI significantly declined to 55% even in untreated controls with NH4+ supply, demonstrating only a small additional non-significant effect of MC-SCM (Figure 6), at least during the investigated later stages of plant development. This applied similarly also for an improved micronutrient status (Zn, Mn, Cu) detectable in the leaf tissue and finally also in the grains (Figure 8).
Similarly, also in the greenhouse experiment, the protective effects of the MC/SC inoculants declined with increasing age of the plants and increasing DS (Figure 2). This may indicate that the applied BS products mainly promoted the early stages of pathogen defense. It may also be speculated that this reflects a downregulation of plant defense reactions induced by fungal effector proteins, characteristic for many biotrophic and hemibiotrophic pathogens including Zt (Yang et al., 2013; Brennan et al., 2019).
3.3 Plant growth and grain yield
Despite mitigation effects on DI and DS recorded in the experiments, plant growth indicators such as shoot and root biomass, root length, root diameter (Figure 5; Supplementary Figure 6), and also final grain yield (Figure 9) remained largely unaffected, both by pathogen inoculation or application of the microbial and non-microbial BSs. High grain yields of 9 t ha−1–10 t ha−1 reaching baking quality with grain protein contents around 12% in all treatments supplied with N fertilizers even in the presence of high DI values (50%–65%) may reflect a certain inherent disease tolerance of the investigated wheat cultivar (Asory). Asory is claimed to be a variety of medium-high resistance against Zt with a rating of 7.5 (Danko Saatzucht Deutschland GmbH, 2024). The high grain yields, far above the 2022 average of 7.5 t ha−1 in Baden-Württemberg (Statistisches Landesamt Baden-Württemberg, 2023), may indicate that there was no relevant impact of stress factors other than the Zt-related pathogen pressure. This could also be a reason for the lack of expression of relevant effects on plant growth in response to application of the investigated microbial and non-microbial BS products, of which benefits have been frequently proven under abiotic stress conditions (Bradáčová et al., 2016; Mpanga et al., 2019; Moradtalab et al., 2020; Rasul et al., 2021).
3.4 Concluding remarks
The BS-assisted fertilization strategies investigated in this study could not fully prevent but clearly slowed down Zt-induced disease spread, depending on the stages of plant development and the form of N fertilization. The applied BS products promoted early defense responses to pathogen attack with preferences for the microbial MC formulation if combined with NH4+ fertilization and the non-microbial SC formulation with NO3− supply. Benefits of NH4+-dominated N fertilization correlated with an improved micronutrient status but were detectable also in later stages of plant development under field conditions. Thus, the combined application of MC and SC with the addition of micronutrients zinc + manganese (MC-SCM application) reduced the pathogen pressure with NH4+ fertilization in the field experiment by inducing increased H2O2 and APX activities. Furthermore, plant growth remained largely unaffected, under both greenhouse and field conditions. It remains to be investigated to which extent these effects can be used to replace fungicide applications during the respective time periods as part of a strategy for integrated pest management. Additional benefits may arise from protective effects reported for the applied BS products against abiotic stress factors.
Data availability statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Author contributions
MG: Writing – original draft, Writing – review & editing. SD: Writing – review & editing. LS: Writing – review & editing. MW: Writing – review & editing. AM: Writing – review & editing. AA: Writing – review & editing. NS: Writing – review & editing. KM: Writing – review & editing. GN: Writing – review & editing. TM: Writing – review & editing. KB: Writing – review & editing.
Funding
The authors declare financial support was received for the research, authorship, and/or publication of this article. The research was conducted within the project “Agriculture 4.0 without Chemical-Synthetic Plant Protection” (NOcsPS), which is funded by the German Federal Ministry of Education and Research and by Agrarsysteme der Zukunft. This research was funded by Bundesministerium für Bildung und Forschung (BMBF), grant number 031B0731A. The APC was funded by Bundesministerium für Bildung und Forschung (BMBF), grant number 031B0731A.
Acknowledgments
We thank the Core Facility Hohenheim (CFH) for performing the nutrient analysis. In addition, we also thank the agricultural research station Heidfeldhof for providing the area for the field experiment. Furthermore, we thank the KeraSan Project for developing the chitosan product formulation provided by Aglukon (KeraSan Project partner). In addition, we thank that personal costs for Gabriel Modolell and Weinmann were covered by the KeraSan and BioFair Project.
Conflict of interest
Author NS was employed by BioAtlantis Ltd. Author KM was employed by Sourcon Padena GmbH.
The remaining authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fpls.2024.1407585/full#supplementary-material
References
Baker, R., Martinson, C. A. (1970). “Epidemiology of Diseases Caused by Rhizoctonia Solatìi,” in Rhizoctonia Solani, Biology and Pathology: Based on an American Phytopathological Society Symposium on Rhizoctonia solani held at the Miami meeting of the Society, October 1965. Ed. Parmeter, J. R. (University of California Press, Berkeley, CA), 172–188.
Bergmann, W. (1992). Nutritional disorders of plants. Development, visual and analytical diagnosis (Jena: Gustav Fischer).
Bergmann, W., Neubert, P. (1976). Pflanzendiagnose und Pflanzenanalyse zur Ermittlung von Ernährungsstörungen und des Ernährungszustandes der Kulturpflanzen (Jena: Gustav Fischer).
Blanke, M. M., Bacher, W., Pring, R. J., Baker, E. A. (1996). Ammonium nutrition enhances chlorophyll and glaucousness in Kohlrabi. Ann. Bot. 78, 599–604. doi: 10.1006/anbo.1996.0166
Boominathan, R., Doran, P. M. (2002). Ni-induced oxidative stress in roots of the Ni hyperaccumulator, Alyssum bertolonii. New Phytol. 156, 205–215. doi: 10.1046/j.1469-8137.2002.00506.x
Bradáčová, K., Kandeler, E., Berger, N., Ludewig, U., Neumann, G. (2020). Microbial consortia inoculants stimulate early growth of maize depending on nitrogen and phosphorus supply. Plant Soil Environ. 66, 105–112. doi: 10.17221/382/2019-PSE
Bradáčová, K., Sittinger, M., Tietz, K., Neuhäuser, B., Kandeler, E., Berger, N., et al. (2019). Maize inoculation with microbial consortia: contrasting effects on rhizosphere activities, nutrient acquisition and early growth in different soils. Microorganisms 7, 329. doi: 10.3390/microorganisms7090329
Bradáčová, K., Weber, N. F., Morad-Talab, N., Asim, M., Imran, M., Weinmann, M., et al. (2016). Micronutrients (Zn/Mn), seaweed extracts, and plant growth-promoting bacteria as cold-stress protectants in maize. Chem. Biol. Technol. Agric. 3, 1–10. doi: 10.1186/s40538-016-0069-1
Brennan, C. J., Benbow, H. R., Mullins, E., Doohan, F. M. (2019). A review of the known unknowns in the early stages of septoria tritici blotch disease of wheat. Plant Pathol. 68, 1427–1438. doi: 10.1111/ppa.13077
Caverzan, A., Passaia, G., Rosa, S. B., Ribeiro, C. W., Lazzarotto, F., Margis-Pinheiro, M. (2012). Plant responses to stresses: Role of ascorbate peroxidase in the antioxidant protection. Genet. Mol. Biol. 35, 1011–1019. doi: 10.1590/s1415-47572012000600016
Choudhary, R. C., Kumaraswamy, R. V., Kumari, S., Pal, A., Raliya, R., Biswas, P., et al. (2017). “Synthesis, Characterization, and Application of Chitosan Nanomaterials Loaded with Zinc and Copper for Plant Growth and Protection,” in Nanotechnology: An agricultural paradigm. Eds. Prasad, R. e., Kumar, e. M., Kumar, e. V. (Springer Singapore, Singapore), 227–247.
Cook, J., Zhang, J., Norrie, J., Blal, B., Cheng, Z. (2018). Seaweed Extract (Stella Maris®) Activates Innate Immune Responses in Arabidopsis thaliana and Protects Host against Bacterial Pathogens. Mar. Drugs 16, 221. doi: 10.3390/md16070221
Danko Saatzucht Deutschland GmbH (2024). Asory - Impresses with its yield and quality. Available online at: https://danko.pl/en/odmiany-type/asory-2/ (Accessed January 13, 2024).
Datnoff, L. E., Elmer, W. H., Huber, D. M. (Eds.) (2007). Mineral nutrition and plant disease (St. Paul, Minn: APS Press, American Phytopathological Soc).
Ding, S., Shao, X., Li, J., Ahammed, G. J., Yao, Y., Ding, J., et al. (2021). Nitrogen forms and metabolism affect plant defence to foliar and root pathogens in tomato. Plant Cell Environ. 44, 1596–1610. doi: 10.1111/pce.14019
EL Mehdi EL Boukhari, M., Barakate, M., Bouhia, Y., Lyamlouli, K. (2020). Trends in seaweed extract based biostimulants: manufacturing process and beneficial effect on soil-plant systems. Plants 9, 359. doi: 10.3390/plants9030359
Eskandari, S., Höfte, H., Zhang, T. (2020). Foliar manganese spray induces the resistance of cucumber to Colletotrichum lagenarium. J. Plant Physiol. 246-247, 153129. doi: 10.1016/j.jplph.2020.153129
Eskandari, S., Khoshgoftarmanesh, A. H., Sharifnabi, B. (2018). The effect of foliar-applied manganese in mineral and complex forms with amino acids on certain defense mechanisms of cucumber (Cucumis sativus L.) against powdery mildew. J. Plant Growth Regul. 37, 481–490. doi: 10.1007/s00344-017-9747-x
Eyal, Z., Scharen, A. L., Prescott, J. M., van Ginkel, M. (1987). The Septoria diseases of wheat: Concepts and methods of disease management (Mexico, D.F: Centro Internac. de Mejoramiento de Maïz y Trigo).
Fernandes, M. S., Rossiello, R. O. P. (1995). Mineral nitrogen in plant physiology and plant nutrition. Crit. Rev. Plant Sci. 14, 111–148. doi: 10.1080/07352689509701924
Fones, H., Gurr, S. (2015). The impact of Septoria tritici Blotch disease on wheat: An EU perspective. Fungal Genet. Biol. 79, 3–7. doi: 10.1016/j.fgb.2015.04.004
Fones, H. N., Eyles, C. J., Kay, W., Cowper, J., Gurr, S. J. (2017). A role for random, humidity-dependent epiphytic growth prior to invasion of wheat by Zymoseptoria tritici. Fungal Genet. Biol. 106, 51–60. doi: 10.1016/j.fgb.2017.07.002
Geisseler, D., Scow, K. M. (2014). Long-term effects of mineral fertilizers on soil microorganisms – A review. Soil Biol. Biochem. 75, 54–63. doi: 10.1016/j.soilbio.2014.03.023
Gunupuru, L. R., Patel, J. S., Sumarah, M. W., Renaud, J. B., Mantin, E. G., Prithiviraj, B. (2019). A plant biostimulant made from the marine brown algae Ascophyllum nodosum and chitosan reduce Fusarium head blight and mycotoxin contamination in wheat. PloS One 14, e0220562. doi: 10.1371/journal.pone.0220562
Halpern, M., Bar-Tal, A., Ofek, M., Minz, D., Muller, T., Yermiyahu, U. (2015). “The Use of Biostimulants for Enhancing Nutrient Uptake,” in Advances in agronomy. Eds. Sparks, D. L., Antanasovic, S. (Waltham, Massachusetts, Academic Press), 141–174. doi: 10.1016/bs.agron.2014.10.001
Harrat, W., Bouznad, Z. (2014). Effects of nitrogen and potassium fertilization on septoria tritici development. Adv. Environ. Biol. 21, 335–338.
Huber, D. M., Haneklaus, S. (2007). Managing nutrition to control plant disease. Landbauforschung Völkenrode 4, 313–322.
Huber, D. M., Watson, R. D. (1974). Nitrogen form and plant disease. Annu. Rev. Phytopathol. 12, 139–165. doi: 10.1146/annurev.py.12.090174.001035
Karuppiah, V., Sun, J., Li, T., Vallikkannu, M., Chen, J. (2019). Co-cultivation of Trichoderma asperellum GDFS1009 and Bacillus amyloliquefaciens 1841 Causes Differential Gene Expression and Improvement in the Wheat Growth and Biocontrol Activity. Front. Microbiol. 10. doi: 10.3389/fmicb.2019.01068
Kozak, M., Piepho, H.-P. (2017). What’s normal anyway? Residual plots are more telling than significance tests when checking ANOVA assumptions. J. Agron. Crop Sci. 204, 86–98. doi: 10.1111/jac.12220
Kumar, R. R., Dubey, K., Goswami, S., Hasija, S., Pandey, R., Singh, P. K., et al. (2020). Heterologous expression and characterization of novel manganese superoxide dismutase (Mn-SOD) - A potential biochemical marker for heat stress-tolerance in wheat (Triticum aestivum). Int. J. Biol. Macromol. 161, 1029–1039. doi: 10.1016/j.ijbiomac.2020.06.026
Kumudini, B. S., Patil, S. V. (2021). “Antioxidant-mediated defense in triggering resistance against biotic stress in plants,” in BIOCONTROL AGENTS AND SECONDARY METABOLITES: Application and immunization ([S.l.] (WOODHEAD PUBLISHING), 383–399. doi: 10.1016/B978-0-12-822919-4.00015-6
Mamun, A. A., Neumann, G., Moradtalab, N., Ahmed, A., Dupuis, B., Darbon, G., et al. (2024). Microbial consortia versus single-strain inoculants as drought stress protectants in potato affected by the form of N supply. Horticulturae 10, 102. doi: 10.3390/horticulturae10010102
Marschner, P., Rengel, Z. (2023). “Nutrient availability in soils,” in Marschner’s mineral nutrition of plants. Eds. Rengel, Z., Cakmak, I., White, P. J. (ELSEVIER ACADEMIC Press, London), 499–522.
Moll, E., Flath, K., Piepho, H.-P. (2000). Die Prüfung von Pflanzen auf ihre Widerstandsfähigkeit gegen SChadorganismen in der Biologischen Bundesanstalt; Testing of crop cultivars for resistance to noxious organisms at the Federal Biological Research Centre. Teil 3: Methodische Anleitung zur Bewertung der partiellen Resistenz von Getreidesortimenten und die SAS-Applikation RESI; Part 3: Methodological guidelines for the assessment of partial resistance in cereal cultivars and the SAS application RESI (Berlin: Parey).
Moradtalab, N., Ahmed, A., Geistlinger, J., Walker, F., Höglinger, B., Ludewig, U., et al. (2020). Synergisms of microbial consortia, N forms, and micronutrients alleviate oxidative damage and stimulate hormonal cold stress adaptations in maize. Front. Plant Sci. 11, 396. doi: 10.3389/fpls.2020.00396
Moradtalab, N., Weinmann, M., Walker, F., Höglinger, B., Ludewig, U., Neumann, G. (2018). Silicon improves chilling tolerance during early growth of maize by effects on micronutrient homeostasis and hormonal balances. Front. Plant Sci. 9, 420. doi: 10.3389/fpls.2018.00420
Mpanga, I. K., Gomez-Genao, N., Moradtalab, N., Wanke, D., Chrobaczek, V., Ahmed, A., et al. (2019). The role of N form supply for PGPM-host plant interactions in maize. J. Plant Nutr. Soil Sci. 182, 908–920. doi: 10.1002/jpln.201900133
Nanda, S., Kumar, G., Hussain, S. (2021). Utilization of seaweed-based biostimulants in improving plant and soil health: current updates and future prospective. Int. J. Environ. Sci. Technol. 19, 12839–12852. doi: 10.1007/s13762-021-03568-9
Orzali, L., Corsi, B., Forni, C., Riccioni, L. (2017). “Chitosan in Agriculture: A New Challenge for Managing Plant Disease,” in Biological Activities and Application of Marine Polysaccharides. Ed. Shalaby, E. A. (InTech). doi: 10.5772/66840
Pogacean, M. O., Gavrilescu, M. (2009). Plant protection products and their sustainable and environmentally friendly use. Environ. Eng. Manage. J. 8, 607–627. doi: 10.30638/eemj.2009.084
Prakasha, A., Umesha, S. (2016). Biochemical and molecular variations of guaiacol peroxidase and total phenols in bacterial wilt pathogenesis of solanum melongena. Biochem. Anal. Biochem. 5. doi: 10.4172/2161-1009.1000292
Quaedvlieg, W., Kema, G. H. J., Groenewald, J. Z., Verkley, G. J. M., Seifbarghi, S., Razavi, M., et al. (2011). Zymoseptoria gen. nov. a new genus to accommodate Septoria-like species occurring on graminicolous hosts. Persoonia 26, 57–69. doi: 10.3767/003158511X571841
Raliya, R., Saharan, V., Dimkpa, C., Biswas, P. (2018). Nanofertilizer for precision and sustainable agriculture: current state and future perspectives. J. Agric. Food Chem. 66, 6487–6503. doi: 10.1021/acs.jafc.7b02178
Rasul, F., Gupta, S., Olas, J. J., Gechev, T., Sujeeth, N., Mueller-Roeber, B. (2021). Priming with a seaweed extract strongly improves drought tolerance in arabidopsis. Int. J. Mol. Sci. 22, 1469. doi: 10.3390/ijms22031469
Reid, J. M., Racz, G. J. (1985). Effects of soil temperature on manganese availability to plants grown on an organic soil. Can. J. Soil Sci. 65, 769–775. doi: 10.4141/cjss85-081
Richter, C., Piepho, H.-P., Thöni, H. (2009). Das “Lateinische Rechteck” – seine Planung, Randomisation und Auswertung verbunden mit einer Begriffsrevision. Pflanzenbauwissenschaften: German J. Agron. 13, 1–14.
Saidi, A., Eslahi, M. R., Safaie, N. (2012). Efficiency of Septoria tritici sporulation on different culture media. TRAKIA J. OF Sci. 10, 15–18.
Sarma, B. K., Yadav, S. K., Singh, S., Singh, H. B. (2015). Microbial consortium-mediated plant defense against phytopathogens: Readdressing for enhancing efficacy. Soil Biol. Biochem. 87, 25–33. doi: 10.1016/j.soilbio.2015.04.001
Shahrajabian, M. H., Chaski, C., Polyzos, N., Tzortzakis, N., Petropoulos, S. A. (2021). Sustainable agriculture systems in vegetable production using chitin and chitosan as plant biostimulants. Biomolecules 11, 819. doi: 10.3390/biom11060819
Sharma, P., Jha, A. B., Dubey, R. S., Pessarakli, M. (2012). Reactive oxygen species, oxidative damage, and antioxidative defense mechanism in plants under stressful conditions. J. Bot. 2012, 1–26. doi: 10.1155/2012/217037
Shaw, M. W. (1991). Interacting effects of interrupted humid periods and light on infection of wheat leaves by Mycosphaerella graminicola (Septoria tritici). Plant Pathol. 40, 595–607. doi: 10.1111/j.1365-3059.1991.tb02424.x
Shetty, N. P., Kristensen, B. K., Newman, M.-A., Møller, K., Gregersen, P. L., Jørgensen, H. (2003). Association of hydrogen peroxide with restriction of Septoria tritici in resistant wheat. Physiol. Mol. Plant Pathol. 62, 333–346. doi: 10.1016/S0885-5765(03)00079-1
Silva, S., Dias, M. C., Silva, A. M. S. (2022). Titanium and zinc based nanomaterials in agriculture: A promising approach to deal with (A)biotic stresses? Toxics 10, 172. doi: 10.3390/toxics10040172
Simón, M. R., Cordo, C. A., Perelló, A. E., Struik, P. C. (2003). Influence of nitrogen supply on the susceptibility of wheat to septoria tritici. J. Phytopathol. 151, 283–289. doi: 10.1046/j.1439-0434.2003.00720.x
Sparrow, L. A., Uren, N. C. (1987). Oxidation and reduction of Mn in acidic soils: Effect of temperature and soil pH. Soil Biol. Biochem. 19, 143–148. doi: 10.1016/0038-0717(87)90073-3
Statistisches Landesamt Baden-Württemberg (2023). Ernteerträge 2022 in Baden-Württemberg. Available online at: https://www.statistik-bw.de/Service/Veroeff/Faltblatt/803823005.pdf (Accessed January 13, 2024).
Sun, Y., Wang, M., Mur, L. A. J., Shen, Q., Guo, S. (2020). Unravelling the roles of nitrogen nutrition in plant disease defences. Int. J. Mol. Sci. 21, 572. doi: 10.3390/ijms21020572
Vasil’ev, L. A., Dzyubinskaya, E. V., Zinovkin, R. A., Kiselevsky, D. B., Lobysheva, N. V., Samuilov, V. D. (2009). Chitosan-induced programmed cell death in plants. Biochem. Biokhimiia 74, 1035–1043. doi: 10.1134/s0006297909090120
VDLUFA. (2023). Probenahme für die Nmin-Methode. Verband Deutscher Landwirtschaftlicher Untersuchungs- und Forschungsanstalten (VDLUFA) Methodenbuch Band I, Böden, A 1. ENTNAHME, TRANSPORT UND AUFBEREITUNG VON PROBEN, A 1.2.2. (Darmstadt, Germany: VDLUFA-Verlag).
VDLUFA. (2012). Bestimmung von Gesamt-Stickstoff nach trockener Verbrennung (Elementaranalyse). Verband Deutscher Landwirtschaftlicher Untersuchungs- und Forschungsanstalten (VDLUFA) Methodenbuch Band I, 6. Teillieferung, A 2.2.5, (Darmstadt, Germany: VDLUFA-Verlag), pp. 1–6.
Walters, D. R., Bingham, I. J. (2007). Influence of nutrition on disease development caused by fungal pathogens: implications for plant disease control. Ann. Appl. Biol. 151, 307–324. doi: 10.1111/j.1744-7348.2007.00176.x
Wang, C., Zhang, S. H., Wang, P. F., Li, W., Lu, J. (2010). Effects of ammonium on the antioxidative response in Hydrilla verticillata (L.f.) Royle plants. Ecotoxicol. Environ. Saf. 73, 189–195. doi: 10.1016/j.ecoenv.2009.08.012
Weinmann, M., Bradáčová, K., Nikolic, M. (2023). “Relationship between mineral nutrition, plant diseases, and pests,” in Marschner’s mineral nutrition of plants. Eds. Rengel, Z., Cakmak, I., White, P. J. (ELSEVIER ACADEMIC Press, London), 445–476.
Yang, F., Li, W., Jørgensen, H. J. L. (2013). Transcriptional reprogramming of wheat and the hemibiotrophic pathogen Septoria tritici during two phases of the compatible interaction. PloS One 8, e81606. doi: 10.1371/journal.pone.0081606
Zaim, S., Bekkar, A. A., Belabid, L. (2018). Efficacy of Bacillus subtilis and Trichoderma harzianum combination on chickpea Fusarium wilt caused by F. oxysporum f. sp. ciceris. Arch. Phytopathol. Plant Prot. 51, 217–226. doi: 10.1080/03235408.2018.1447896
Zhu, L., Huang, J., Lu, X., Zhou, C. (2022). Development of plant systemic resistance by beneficial rhizobacteria: Recognition, initiation, elicitation and regulation. Front. Plant Sci. 13. doi: 10.3389/fpls.2022.952397
Keywords: biotic stress, Zymoseptoria tritici, winter wheat, seaweed extracts, microbial consortium, agriculture, nitrate, ammonium
Citation: Göbel M, Dulal S, Sommer L, Weinmann M, Mamun AA, Ahmed A, Sujeeth N, Mai K, Neumann G, Müller T and Bradáčová K (2024) Protective potential of selected microbial and non-microbial biostimulants against Zymoseptoria tritici leaf blotch in winter wheat as affected by the form of N supply. Front. Plant Sci. 15:1407585. doi: 10.3389/fpls.2024.1407585
Received: 26 March 2024; Accepted: 21 August 2024;
Published: 27 September 2024.
Edited by:
Stephen O. Amoo, Agricultural Research Council of South Africa (ARC-SA), South AfricaReviewed by:
Arup Ghosh, Council of Scientific and Industrial Research (CSIR), IndiaRoxana Vidican, University of Agricultural Sciences and Veterinary Medicine of Cluj-Napoca, Romania
Suresh Kaushik, Indian Agricultural Research Institute, India
Ali Baghdadi, University of Bologna, Italy
Copyright © 2024 Göbel, Dulal, Sommer, Weinmann, Mamun, Ahmed, Sujeeth, Mai, Neumann, Müller and Bradáčová. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Markus Göbel, bWFya3VzLmdvZWJlbEB1bmktaG9oZW5oZWltLmRl