 Sergio Saia
Sergio Saia Emanuele Radicetti
Emanuele Radicetti Katharina Pawlowski
Katharina Pawlowski Sabine Dagmar Zimmermann
Sabine Dagmar Zimmermann Andrea Genre
Andrea Genre
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
EDITORIAL article
Front. Plant Sci., 09 April 2024
Sec. Plant Symbiotic Interactions
Volume 15 - 2024 | https://doi.org/10.3389/fpls.2024.1399420
This article is part of the Research TopicMechanisms and Practices for the Management of Plant-Soil Biota InteractionView all 9 articles
Editorial on the Research Topic
Mechanisms and practices for the management of plant-soil biota interaction
The perception of the importance of the soil biota for agricultural production, land management, and ecosystem management and functioning increased in the last decades. Plenty of local, national and international initiatives and research projects are addressing the relationship between plant and soil biota, using both holistic metabarcoding/metagenomics-based studies and mechanistic approaches addressing bipartite plant-microbe interactions. In most of these studies attention has been focussed on associations and symbioses, which may simultaneously promote plant health, crop yield, microbial spread, and microbiota-associated ecosystem services, with special emphasis on soil health and preservation. Indeed, the soil microbiota is the main component in soil fertility and can directly influence major soil traits at both the short and the long term. So far, the management of the soil biota, especially for those components that improve crop production, is scarcely explored even in studies that aim at introducing sustainable cropping systems and natural soil management. In particular, the benefits deriving from individual microbial taxa have been demonstrated under diverse conditions, but scientific literature is far from providing general guidelines to achieve the strongest benefits from the management of soil microbiota.
This Research Topic (RT) was launched in January 2023 on the trail of two previous RTs, namely Arbuscular Mycorrhizal Fungi: The Bridge between Plants, Soils, and Humans and Insights in Plant Symbiotic Interactions: 2021, both deepening insight in beneficial plant-symbiotic interactions and aiming to elucidate the functioning and applicative potential of such interactions. The present RT focussed on the management of these and other symbioses for improving plant growth, soil traits and food safety for both livestock and humans. The call for papers attracted several contributions, among which 7 original papers and 1 perspective article were accepted (Table 1).
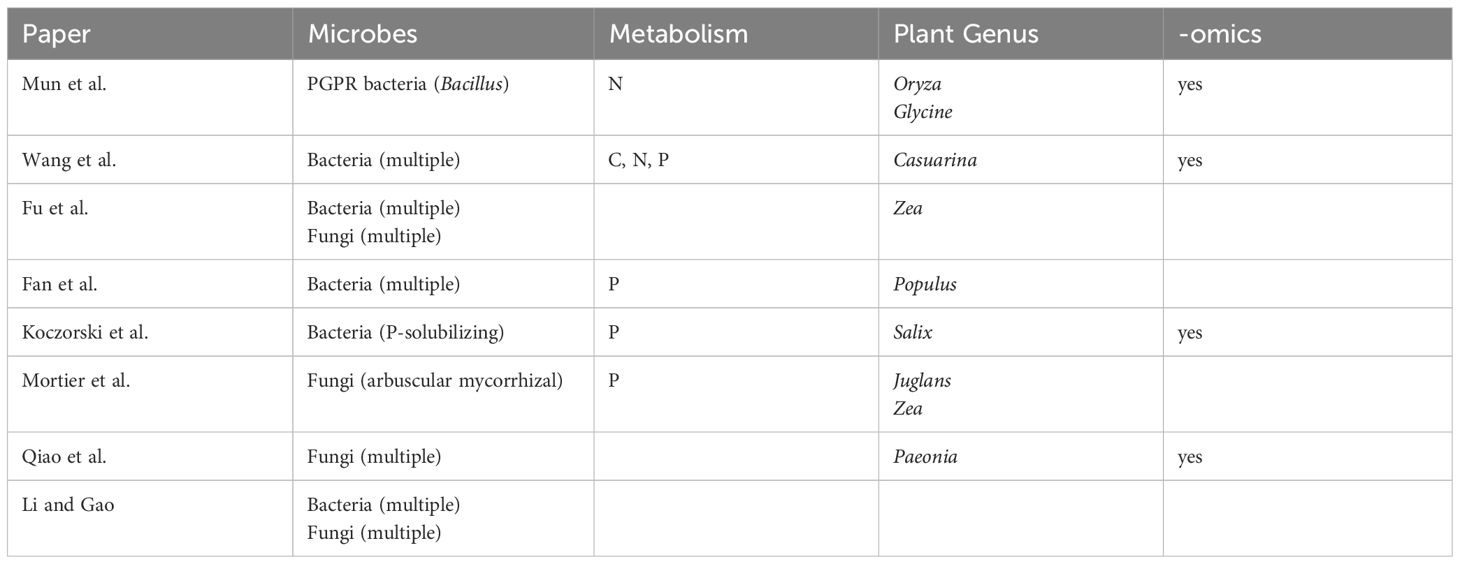
Table 1 Summary of the focus and approaches presented in each contribution of the RT.
Phosphorus is one of the most essential elements for plant growth and development. However, phosphorus is also one of the hardest nutrients for plants to get because of its low availability in the soil due to the rapid formation of poorly soluble complexes (Bindraban et al., 2020). Koczorski et al. have investigated the effect of phosphate-solubilizing bacteria inoculation on the development and transcriptomic scenario of two willow species under different phosphate concentrations. Regarding agricultural practices, Fan et al. studied phosphorus availability for growth of seedlings from the endangered species Populus euphratica (Ling et al., 2015). The authors analysed within a field experiment with two-year old seedlings the effect of cow dung return methods on soil microbiota and phosphorus within the root zone. In addition to soil properties, abundance of soil bacteria and genes involved in phosphorus solubilization and transformation have been analysed. Findings showing that use of biochar (carbonised cow manure; Kurniawan et al., 2023) increased the biodiversity of the microbial community as well as the availability of phosphorus indicate clearly a way to reduce phosphate fertilisation thus improving agricultural management.
Also related to phosphate nutrition (Plassard et al., 2019; Zhang et al., 2024), but in a rather different context, Mortier et al. present the results of their study of a simulated agroforestry system, where they demonstrated the role of a common mycorrhizal network (Walder et al., 2015) in phosphate distribution between intercropped walnuts and maize plants. Fungal communities have been studied also in the context of cultivation of a rare and endangered plant species in China, the shrub Paeonia ludlowii. A previous study described the rhizosphere microbial diversity and community structure in the introduction area (Gao et al., 2022). Here, growth behaviour of wild and cultivated species was compared and related to rhizosphere soil fungi (Qiao et al.). Network analyses revealed that Ascomycota were the key fungi in wild species but were absent in the cultivated ones. The authors concluded that a disturbed soil fungi community promoting pathogenic fungi might hinder the cultivation of the studied plant species. Similar network analyses for domesticated plants have already shown such negative plant-soil feedback (Carrillo et al., 2019; Martı́n-Robles et al., 2020; Chang et al., 2021). These results point clearly to the fact that the introduction of plant species to another environment can cause a shifted soil-microbiome-plant equilibrium and must be taken into account for plant cultivation.
According to the above-cited studies, plant roots throughout the release of different exudates, i.e. low-molecular weight organic acids and metabolites, and their interaction with water and nutrient cycles contribute to the modification of soil physicochemical characteristics determining a selective environment for the microorganisms’ establishment and development (Kuzyakov and Razavi, 2019). It is necessary to develop models able to enhance our understanding of the plant-driving microbial community distribution patterns in rhizospheric soil. Fu et al. suggest that the evaluation of the genetic structure of the soil microbiome of the plant rhizosphere could be adopted for the evaluation of these aspects. In particular, the study of Fu et al. aims to examine the role of microdiversity in influencing the distribution of rhizosphere-associated microbial species across environmental gradients from root surface to bulk soil at the operational taxonomic units (OTUs) and amplicon sequence variant (ASVs) levels. The finding suggests that ASVs exhibited microdiversity within OTUs in the root-associated soil compartments of mature maize plants growing under field conditions. In addition, environmental factors in root-associated soil compartments of maize may drive the microdiversity in microbial community structure.
Another key topic in this RT is the interaction between soil microbes and cultivated plants. Under this approach, the adoption of plant growth-promoting rhizobacteria (PGPR) may be of interest for their benefits on the agro-physiological characteristics of crops. The study of Mun et al. aims at the evaluation of rhizobacterium Bacillus aryabhattai on soybean and rice. The findings showed how B. aryabhattai affects amino acids and protein metabolism in terms of quantity and quality, thus indicating B. aryabhattai as a promising PGPR for field application not only for soybean and rice but also for various other crops.
Different groups focused on the assembly of rhizosphere microbiomes and their development over time. In 2001, Batish et al. (2001) had concluded that Casuarina equisetifolia, a tree with high resistance against abiotic stress and important in coast line protection, had allelopathic properties in that phenolics, in particular, 2,4-di-tert-butylphenol (2,4-DTBP), released from the photosynthetic branchlets and litter of the tree adversely affected the understorey vegetation. Recently, Xu et al. (2022) suggested that actually, litter fungi with higher abundance in young and mature plantations were involved in the synthesis of 2,4-DTBP. Based on rhizobiome metagenomics, Wang et al. conclude that a decrease of actinobacteria from 10 key genera led to reduced metabolic capacity of the soil microbiome for nutrient cycling together with reduced ion exchange capacity of the rhizosphere soil. This result offers options for ameliorating the negative effects of continuous planting of C. equisetifolia that not only affect the understorey vegetation but also C. equisetifolia itself. The conclusions also fit the hypothesis that host filtering effects are the main deterministic force in the assembly of microbiota, a process otherwise controlled by stochastic processes. However, Li and Gao, focussing on the root endophytome, propose in their intriguing Perspective article that dispersal limitation represents a further deterministic process, in particular with regard to the bacterial microbiome, which has to be considered when attempting microbiome engineering.
In summary, the majority of the studies included in this RT applied -omic techniques to investigate the relationship between bacteria and fungi and different crops. For agricultural applications, it will now be interesting to delve deeper into mechanisms and best practices for managing plant-soil biota on the field-scale, with more attention on whole microbiome interactions and herbaceous crops, in particular legumes and C3 cereals.
SS: Writing – original draft, Writing – review & editing. ER: Writing – original draft, Writing – review & editing. KP: Writing – original draft, Writing – review & editing. SZ: Writing – original draft, Writing – review & editing. AG: Writing – original draft, Writing – review & editing.
We are grateful to all Authors who contributed to this Research Topic, to the Reviewers who evaluated their work and to the Frontiers editorial staff for their assistance.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The author(s) declared that they were an editorial board member of Frontiers, at the time of submission. This had no impact on the peer review process and the final decision.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Batish, D., Singh, H. P., Kohli, R. (2001). Vegetation exclusion under Casuarina equisetifolia L.: Does allelopathy play a role? Community Ecol. 2, 93–100. doi: 10.1556/ComEc.2.2001.1.10
Bindraban, P. S., Dimkpa, C. O., Pandey, R. (2020). Exploring phosphorus fertilizers and fertilization strategies for improved human and environmental health. Biol. Fertil. Soils 56, 299–317. doi: 10.1007/s00374-019-01430-2
Carrillo, J., Ingwell, L. L., Li, X., Kaplan, L. (2019). Domesticated tomatoes are more vulnerable to negative plant-soil feedbacks than their wild relatives. J. Ecol. 107, 1753–1766. doi: 10.1111/1365-2745.13157
Chang, J., Sun, Y., Tian, L., Ji, L., Luo, S., Nasir, F., et al. (2021). The structure of rhizosphere fungal communities of wild and domesticated rice: changes in diversity and co-occurrence patterns. Front. Microbiol. 12. doi: 10.3389/fmicb.2021.610823
Gao, D., Wu, L., Meng, F., Yuan, T. (2022). Rhizosphere microbial diversity and community structure of Paeonia ludlowii in introduction area. J. Northeast Agric. Univ. 53, 19–29. doi: 10.19720/j.cnki.issn.1005-9369.2022.03.003
Kurniawan, T. A., Othman, M. H. D., Liang, X., Goh, H. H., Gikas, P., Chong, K.-K., et al. (2023). Challenges and opportunities for biochar to promote circular economy and carbon neutrality. J. Environ. Manage. 332, 117429. doi: 10.1016/j.jenvman.2023.117429
Kuzyakov, Y., Razavi, B. S. (2019). Rhizosphere size and shape: Temporal dynamics and spatial stationarity. Soil Biol. Biochem. 135, 343–360. doi: 10.1016/j.soilbio.2019.05.011
Ling, H., Zhang, P., Xu, H., Zhao, X. (2015). How to regenerate and protect desert riparian Populus euphratica forest in arid areas. Sci. Rep. 5, 1–12. doi: 10.1038/srep15418
Martı́n-Robles, N., Garcı́a-Palacios, P., Rodrı́guez, M., Rodrı́guez, D., Vigo, R., Sánchez-Moreno, S., et al. (2020). Crops and their wild progenitors recruit beneficial and detrimental soil biota in opposing ways. Plant Soil. 456, 159–173. doi: 10.1007/s11104-020-04703-0
Plassard, C., Becquer, A., Garcia, K. (2019). Phosphorus transport in mycorrhiza: How far are we? Trends Plant Sci. 24, 794–801. doi: 10.1016/j.tplants.2019.06.004
Walder, F., Brulé, D., Koegel, S., Wiemken, A., Boller, T., Courty, P. E. (2015). Plant phosphorus acquisition in a common mycorrhizal network: regulation of phosphate transporter genes of the Pht1 family in sorghum and flax. New Phytol. 205, 1632–1645. doi: 10.1111/nph.13292
Xu, Z., Zuo, L., Zhang, Y., Huang, R., Li, L. (2022). Is allelochemical synthesis in Casuarina equisetifolia plantation related to litter microorganisms? Front. Plant Sci. 13. doi: 10.3389/fpls.2022.1022984
Keywords: plant-soil biota management, ecosystem, agro-ecosystem, genotypes, beneficial fungi, viruses, bacteria
Citation: Saia S, Radicetti E, Pawlowski K, Zimmermann SD and Genre A (2024) Editorial: Mechanisms and practices for the management of plant-soil biota interaction. Front. Plant Sci. 15:1399420. doi: 10.3389/fpls.2024.1399420
Received: 11 March 2024; Accepted: 28 March 2024;
Published: 09 April 2024.
Edited and Reviewed by:
Febri Doni, Padjadjaran University, IndonesiaCopyright © 2024 Saia, Radicetti, Pawlowski, Zimmermann and Genre. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Sergio Saia, c2VyZ2lvLnNhaWFAdW5pcGkuaXQ=; Emanuele Radicetti, ZW1hbnVlbGUucmFkaWNldHRpQHVuaWZlLml0; Katharina Pawlowski, a2F0aGFyaW5hLnBhd2xvd3NraUBzdS5zZQ==; Sabine Dagmar Zimmermann, c2FiaW5lLnppbW1lcm1hbm5AY25ycy5mcg==; Andrea Genre, YW5kcmVhLmdlbnJlQHVuaXRvLml0
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.