 Sida Meng1†
Sida Meng1† Jing Jiang
Jing Jiang Yue Liang
Yue Liang Wenxing Pang
Wenxing Pang
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Plant Sci. , 06 June 2024
Sec. Functional and Applied Plant Genomics
Volume 15 - 2024 | https://doi.org/10.3389/fpls.2024.1391173
This article is part of the Research Topic Research on Brassicaceae Crops Genomics and Breeding, Volume II View all 10 articles
Clubroot disease, which is caused by the obligate biotrophic protist Plasmodiophora brassicae, leads to the formation of galls, commonly known as pathogen-induced tumors, on the roots of infected plants. The identification of crucial regulators of host tumor formation is essential to unravel the mechanisms underlying the proliferation and differentiation of P. brassicae within plant cells. To gain insight into this process, transcriptomic analysis was conducted to identify key genes associated with both primary and secondary infection of P. brassicae in Chinese cabbage. Our results demonstrate that the k-means clustering of subclass 1, which exhibited specific trends, was closely linked to the infection process of P. brassicae. Of the 1610 differentially expressed genes (DEGs) annotated in subclass 1, 782 were identified as transcription factors belonging to 49 transcription factor families, including bHLH, B3, NAC, MYB_related, WRKY, bZIP, C2H2, and ERF. In the primary infection, several genes, including the predicted Brassica rapa probable pectate lyase, RPM1-interacting protein 4-like, L-type lectin-domain-containing receptor kinase, G-type lectin S-receptor-like serine, B. rapa photosystem II 22 kDa protein, and MLP-like protein, showed significant upregulation. In the secondary infection stage, 45 of 50 overlapping DEGs were upregulated. These upregulated DEGs included the predicted B. rapa endoglucanase, long-chain acyl-CoA synthetase, WRKY transcription factor, NAC domain-containing protein, cell division control protein, auxin-induced protein, and protein variation in compound-triggered root growth response-like and xyloglucan glycosyltransferases. In both the primary and secondary infection stages, the DEGs were predicted to be Brassica rapa putative disease resistance proteins, L-type lectin domain-containing receptor kinases, ferredoxin-NADP reductases, 1-aminocyclopropane-1-carboxylate synthases, histone deacetylases, UDP-glycosyltransferases, putative glycerol-3-phosphate transporters, and chlorophyll a-binding proteins, which are closely associated with plant defense responses, biosynthetic processes, carbohydrate transport, and photosynthesis. This study revealed the pivotal role of transcription factors in the initiation of infection and establishment of intracellular parasitic relationships during the primary infection stage, as well as the proliferation and differentiation of the pathogen within the host cell during the secondary infection stage.
Clubroot disease, which is caused by the obligate biotrophic protist Plasmodiophora brassicae, affect the roots of cruciferous plants, leading to the formation of root galls that disrupt water and nutrient uptake (Voorrips et al., 2003). It is considered one of the most damaging diseases affecting crucifer crops worldwide, and can cause total yield loss under conducive conditions (Dixon, 2009a). The disease affects all cultivated Brassica species, including important vegetables and oilseed crops such as Chinese cabbage, cauliflower, cabbage, turnip, oilseed rape, and the model plant Arabidopsis thaliana (Dixon, 2009b).
A significant breakthrough in clubroot research was the unraveling of the pathogen life cycle and infection processes. Scientists have successfully identified the crucial stages in the development of this disease, including spore germination, primary infection, secondary infection, and gall formation. It is difficult to observe the clear time point between primary infection and secondary infection cause of the primary and secondary zoospores cannot be differentiated based on morphology (Kageyama and Asano, 2009). Feng et al. (2013a) reported that primary infections began to be noticed as early as 12 hours after inoculation (hai) with resting spores and secondary infections were observed at 72 hai. In recently, Liu et al. (2020) reported that the primary infection stage from 0 to 7 days post inoculation (DPI)and the secondary infection stage from 7 to the resting spore formation. Starch accumulation in infected hosts provides P. brassicae with carbon and energy during infection (Ikegami et al., 1984; Schuller et al., 2016; Ma et al., 2022). This understanding has paved the way for targeted interventions to disrupt these processes and reduce the disease severity.
Genetic studies have also played a crucial role in clubroot research, as researchers have identified and characterized multiple resistance genes in cruciferous crops that confer resistance to specific pathotypes of P. brassicae (Pang et al., 2020). Advancements in molecular techniques have facilitated the identification and characterization of genes involved in clubroot susceptibility and resistance (Ueno et al., 2012; Hatakeyama et al., 2013; Pang et al., 2022; Wang et al., 2023). Effective clubroot management relies heavily on the use of resistant cultivars because chemical and cultural controls have limited effectiveness against this soil-borne disease. However, the resistance conferred by major clubroot resistance (CR) genes is often quickly overcome by the prevalence of pathogenic strains of P. brassicae as a result of selection pressure (Kuginuki et al., 1999; Strelkov et al., 2016; Struck et al., 2022). Therefore, it is crucial to make efforts to understand the molecular mechanisms underlying clubroot pathogenesis and CR in order to develop durable clubroot resistance and improve management strategies.
Over the past decade, transcriptomic studies utilizing microarrays and RNA sequencing have generated extensive datasets that offer novel insights into the molecular basis of clubroot infection and defense responses in cruciferous hosts (Ciaghi et al., 2019; Zhou et al., 2020). Extensive transcriptomic reprogramming occurred during the secondary phase of clubroot development and coincided with gall formation. Galls develop abnormally enlarged cells with thin cell walls, large vacuoles, dense cytoplasmic bodies, and highly proliferating mitochondria and plastids (Ludwig-Müller et al., 2009; Dodueva et al., 2020). Feng et al. (2013b) reported that the differentially expressed gene expression patterns in primary and secondary zoospores investigated by dot-blot and qPCR. Microarray analysis of Arabidopsis has identified over 3,900 differentially expressed genes (>2-fold change) in infected gall tissues, whereas RNA-seq has identified over 4,500 differentially expressed transcripts (Agarwal et al., 2011; Jubault et al., 2013). The upregulated pathways in galls included JA, ET, auxin, CK, and brassinosteroid (BR) hormone signaling, which stimulate cell enlargement and division. Genes involved in cell wall modification, cytochrome P450s, transporters, and DNA replication/repair were also upregulated. Conversely, the genes associated with photosynthesis, sulfur/glucosinolate metabolism, defense responses, and cell death were strongly downregulated, indicating extensive metabolic reprogramming. Comparative analysis of Arabidopsis transcriptomes at 10, 14, 21, and 28 DPI revealed dynamic temporal changes, with the most significant alteration occurring at 21 DPI, coinciding with extensive gall formation (Agarwal et al., 2011). In late-stage infection (35 DPI), Shi et al. (2014) identified 515 Arabidopsis genes that were differentially expressed and shared between B. napus and B. rapa. These included genes involved in cell wall modification, hormone signaling, and secondary metabolism. Phenylpropanoid biosynthesis is exclusively induced in Brassica species, whereas photosynthetic genes are repressed solely in Arabidopsis. These analyses revealed the common induction of certain cellular processes, such as cell wall remodeling and hormone signaling, but also indicated divergence in specific responsive genes during clubroot infection across crucifer hosts. Additionally, they demonstrated a stronger correlation between transcriptional changes during the latter stages of disease progression than during the early stages.
Comparative transcriptomics of clubroot-resistant and clubroot-susceptible genotypes have facilitated the identification of candidate resistance genes and pathways. Chu et al. (2014) observed a higher upregulation of genes associated with ET signaling, glutathione-S-transferases, and trehalose biosynthesis in resistant B. rapa lines than in susceptible lines at 10 DPI. Hosts of crucifers and P. brassicae interactions in clubroot has been reviewed in which clubroot pathogenesis and host resistance were well discussed (Feng et al., 2014). Furthermore, RNA-seq analysis of two B. napus lines with varying resistance revealed 618 differentially expressed genes at 21 DPI, which were related to cell organization, biotic stress response, hormone signaling, and glucosinolate biosynthesis (Zhang et al., 2016). The more resistant line exhibited increased expression of the camalexin biosynthesis genes PAD3 and CYP71A13, which corresponded to higher levels of camalexin.
These findings contribute to a better understanding of the mechanisms involved in plant defense responses and susceptibility to clubroot. Over the past ten years, research on clubroot disease has made noteworthy progress in various areas, such as pathogen biology, genetics, and management strategies (Zhang et al., 2023). These advancements have not only improved our comprehension of the disease, but have also provided valuable tools and techniques for effective disease control. However, continuous research efforts are required to advance our understanding of pathogen evolution and adaptation, thereby ensuring sustainable management options for cruciferous crop production.
In this study, a highly susceptible Chinese cabbage inbred line ‘325’ was used. The ‘325’ line was inoculated with P. brassicae isolate ‘SCDY-57,’ identified as pathotype Pb1 according to the Sinitic clubroot differential set (Pang et al., 2020). The P. brassicae resting spore preparation and inoculation processes followed the methodology described by Pang et al. (2020). Briefly, the galls were ground in sterile distilled water using a homogenizer, and the resulting mixtures were filtered through eight layers of cheesecloth. Resting spores were collected by centrifugation at 2,500g and quantified using a hemocytometer (Neubauer improved, Marienfeld, Germany). The concentration of the resting spores was adjusted to 1 × 107/ml, and 1 ml of the suspension was inoculated into 10-day-old seedlings of each plant. Plants treated with 1 ml distilled water were used as mock controls.
Tissue sampling was performed at 3, 7, 14, 21, 28, and 35 DPI with P. brassicae isolate ‘SCDY-57’ or distilled water according to Ma et al. (2022). Roots of 20 individual plants were sampled at each time point and used for RNA isolation. Total RNA was extracted from infected and mock control root samples using TRIzol™ reagent (Invitrogen, Carlsbad, USA). Additional RNA quality was determined using a NanoDrop spectrophotometer (Thermo Scientific, Waltham, MA, USA). Sequencing libraries were generated using the NEBNext UltraTM RNA Library Prep Kit for Illumina [New England Biolabs (NEB), Ipswich, MA, USA], following the manufacturer’s recommendations, and index codes were added to attribute sequences for each sample. Sequencing was performed on an Illumina HiSeq 2500 platform (Illumina, San Diego, CA, USA) by Annoroad Gene Technology Co., Ltd. (Beijing, China).
Raw data were cleaned by removing adapter sequences, N-sequences, and low-quality reads. Reference genomes of Brassica rapa (Brapa_genome_v3.0) and P. brassicae downloaded from the Brassicaceae Database and NCBI were used in this study (Stjelja et al., 2019; Chen et al., 2022). Bowtie2 v2.2.3 was used to build the genome index, and Clean Data were aligned to the reference genome using HISAT2 v2.1.0. The read Count for each gene in each sample was determined using HTSeq v0.6.0, and fragments per kilobase million mapped reads (FPKM) were calculated to estimate the expression level of genes in each sample. DEGseq was used for differential gene expression analysis. Genes with q ≤ 0.05 and |log2_ratio|≥1 are identified as differentially expressed genes (DEGs). The TBtools software (Chen et al., 2020) was used for the Venn map and heat map analysis according to the RNA-Seq data in this study.
Functional and pathway enrichment of proteins encoded by the candidate genes was analyzed. Gene Ontology (GO, http://geneontology.org/) enrichment of DEGs was implemented using the hypergeometric test, in which the p-value was calculated and adjusted as a q-value, and the data background was genes in the whole genome. GO terms with q<0.05 were considered to be significantly enriched. GO enrichment analysis revealed the biological functions of DEGs. The KEGG enrichment of DEGs was implemented using the hypergeometric test, in which the p-value was adjusted by multiple comparisons as q-values. KEGG terms with q<0.05 were considered to be significantly enriched.
PrimeScript™ RT Reagent Kit (TaKaRa, Beijing, China) was used for cDNA synthesis according to the manufacturer’s instructions. The Primer 3.0 online program was used to design the primers for qRT–PCR; the primer information is listed in Supplementary Table 5. The qRT–PCR was conducted in TB Green® Premix Ex Taq™ II FAST qPCR (TaKaRa, Beijing, China) with the CFX96™ Real-Time System. All experiments were performed in triplicate. The relative expression levels of the genes were determined by the 2−ΔΔCT method (Livak and Schmittgen, 2001), and the sample from each time point for distilled water treatment was used as the control.
Data analyses were performed using the SPSS statistical package (SPSS, Chicago, IL, USA). Analysis of variance (ANOVA) was conducted to evaluate the treatments. If a significant treatment effect was identified (P = 0.05), Duncan’s Multiple Range Test was used to determine significant differences at P < 0.05.
A total of 12 root samples were collected from Chinese cabbage inbred line ‘325’ at 3, 7, 14, 21, 28, and 35 days post-treatment with P. brassicae isolate ‘SCDY-57’ as well as distilled water. The samples were named D3Pb, W1Pb, W2Pb, W3Pb, W4Pb, and W5Pb, and D3CK, W1CK, W2CK, W3CK, W4CK, and W5CK for 3, 7, 14, 21, 28, and 35 days after treated with P. brassicae and distilled water, respectively. Twelve libraries were constructed and analyzed, resulting in clean Q30 base-rate values ranging from 93.77%–94.31%. Overall, 516 million good-quality reads were obtained, with mapping rates to the B. rapa genome ranging from 35.61% to 90.51% (Table 1). The mapping rate greatly increased from 0.0078% to 58.37% in P. brassicae genome from 3 to 35 days after P. brassicae inoculation. In total, 30495 and 10518 genes were predicted from W5Pb for B. rapa and P. brassicae, respectively.
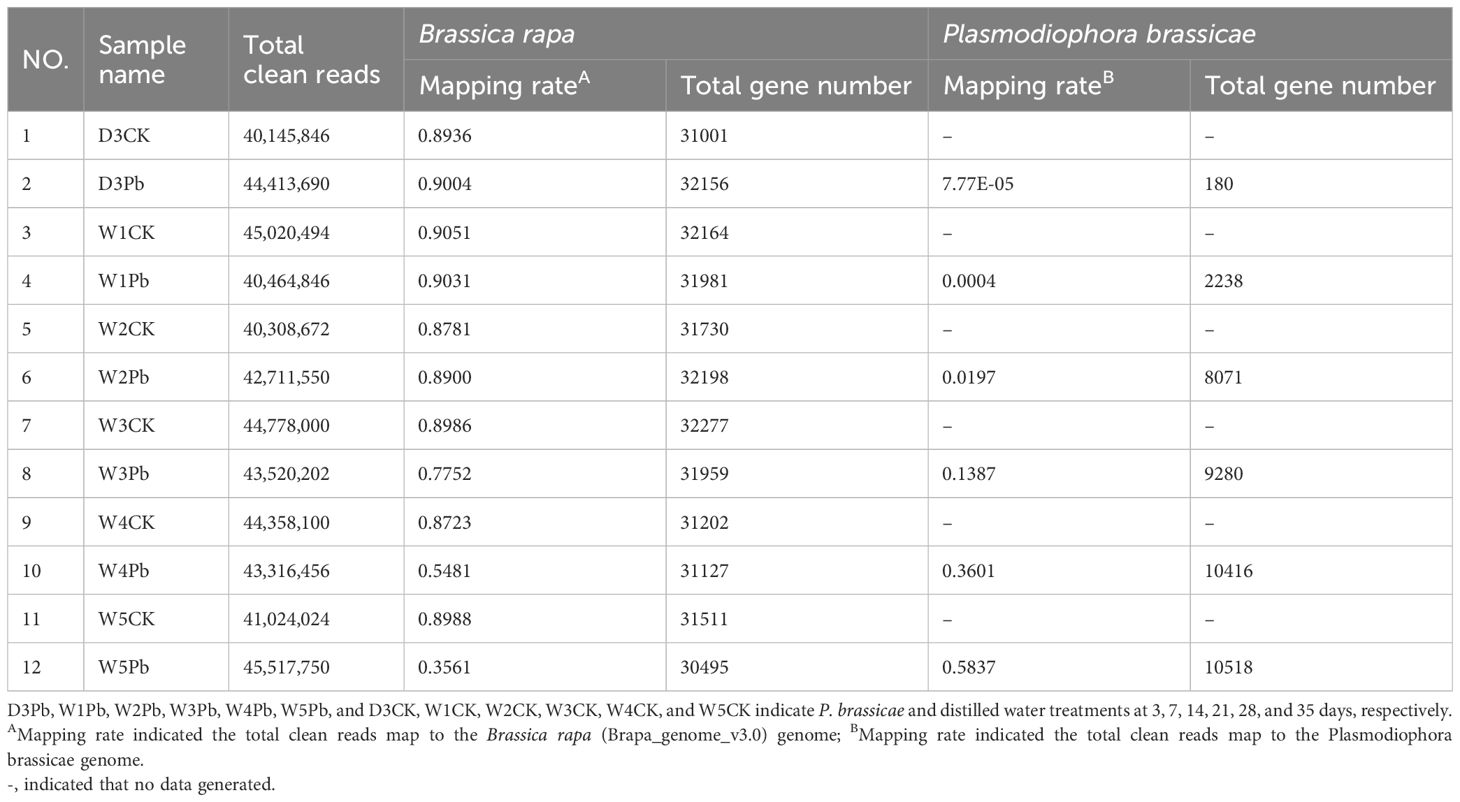
Table 1 The clean reads mapping rate and annotated gene number of the RNA-Seq data were compared to the reference genome of Brassica rapa and Plasmodiophora brassicae.
The sample cluster revealed that 12 samples were grouped into three distinct clusters (Figure 1). Samples D3CK and D3Pb formed a separate cluster that was distinct from that of the other samples. Furthermore, the remaining 10 samples were divided into two groups, with W1Pb, W2Pb, W3Pb, W4Pb, and W5Pb representing the samples treated with P. brassicae, and W1CK, W2CK, W3CK, W4CK, and W5CK representing the samples treated with distilled water.
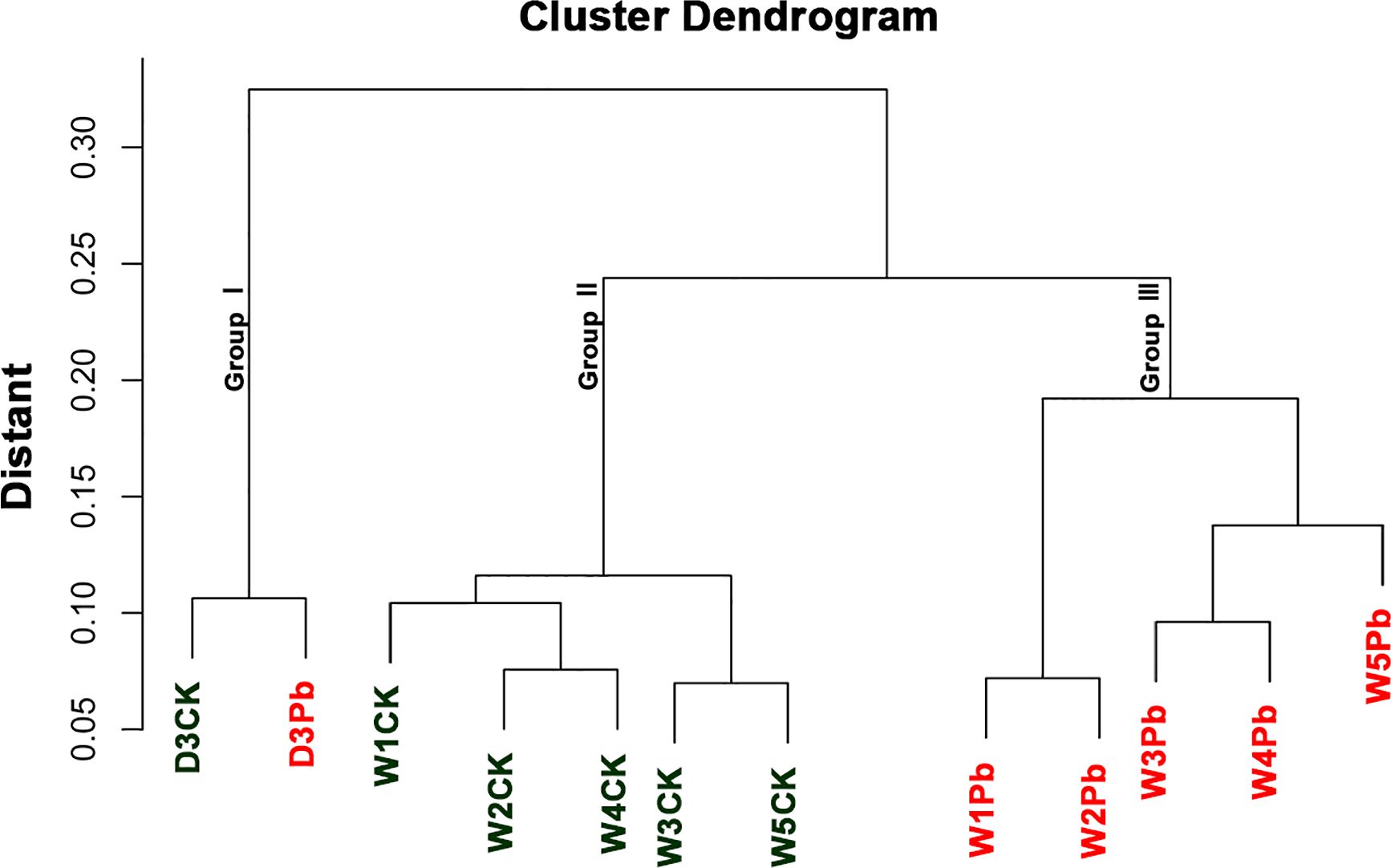
Figure 1 Cluster dendrogram and grouping information of 12 samples.
A total of 9025, 10997, 9620, 8999, 10658, and 10333 DEGs were identified in response to P. brassicae inoculation compared to distilled water at 3, 7, 14, 21, 28, and 35 days post-treatment (Figure 2). The downregulated genes were upregulated more in all P. brassicae treatments than in distilled water, except at 3 and 28 d after treatment.
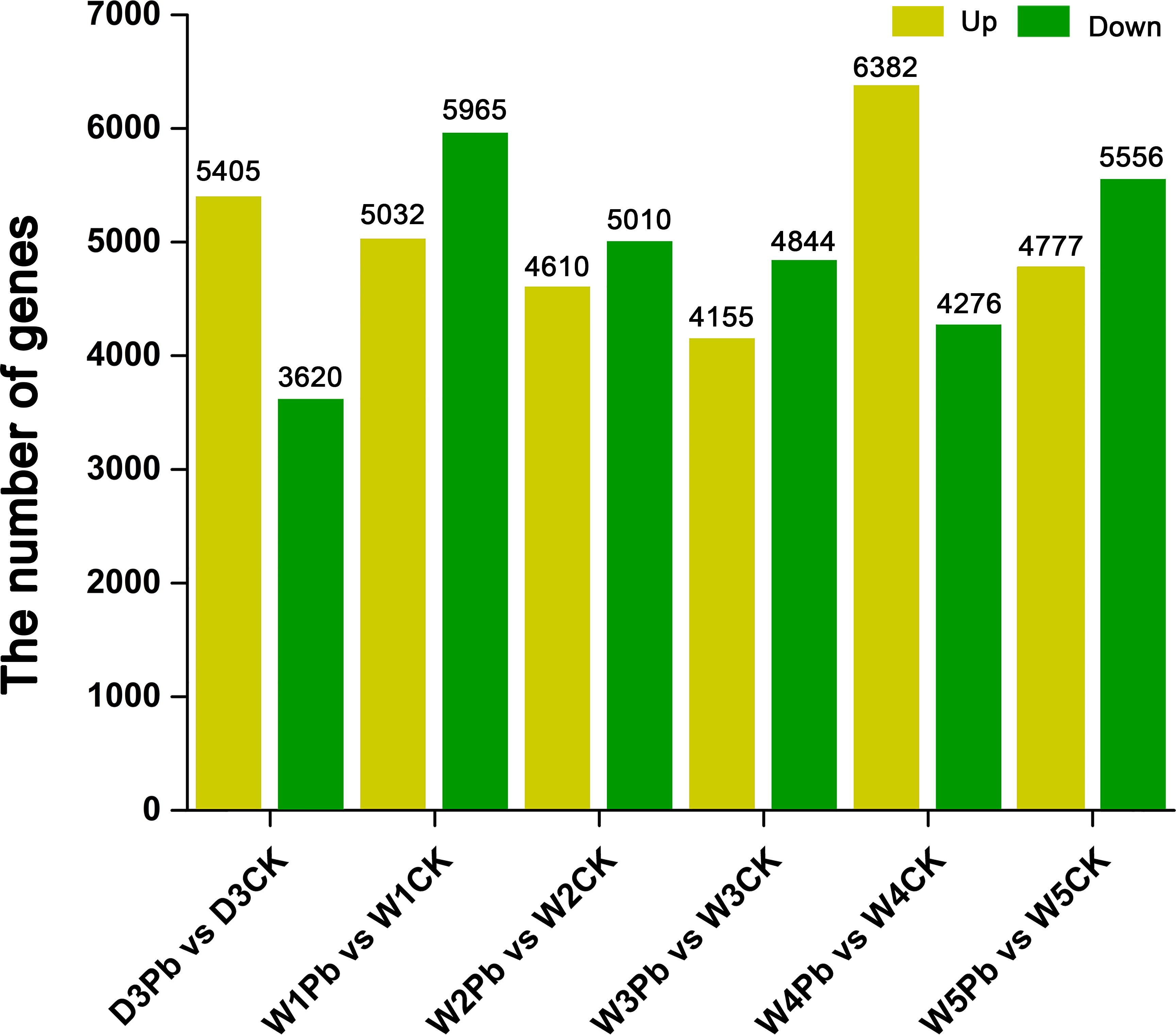
Figure 2 Numbers of differentially expressed genes (DEGs) obtained from P. brassicae inoculation compared to distilled water at 3, 7, 14, 21, 28, and 35 days after treatment.
K-means clustering analysis of the DEGs revealed that these genes were related to processes associated with the infection of P. brassicae (Figure 3). The results showed that the k-means clustering of sub-classes 1 and 3 with certain trends were associated with the infection of P. brassicae. However, sub-class 1 had a much closer relationship with the process of P. brassicae infection than sub-class 3. Of the 1610 DEGs annotated in subclass 1, 782 were characterized as transcription factors. These 782 genes belonged to 49 transcription factor families, including basic helix-loop-helix (bHLH), B3, NAC (no apical meristem (NAC), MYB_related, WRKY, bZIP (basic region/leucine zipper motif (bZIP), C2H2, and ERF(Ethylene response factors) etc. (Figure 4). Most genes were annotated to the transcription factor family bHLH, and 79 genes were detected.
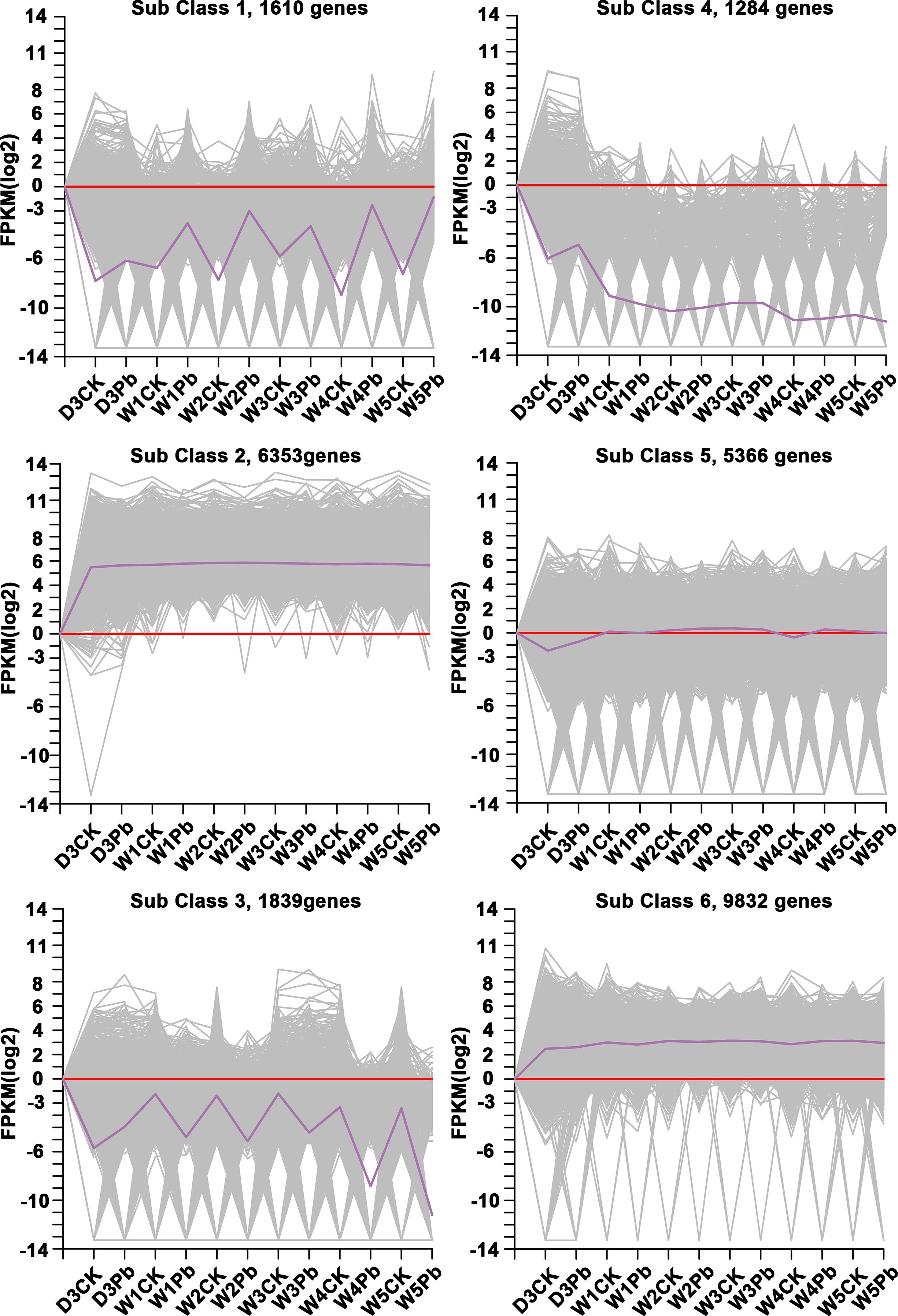
Figure 3 Clusters obtained by k-means cluster analysis.
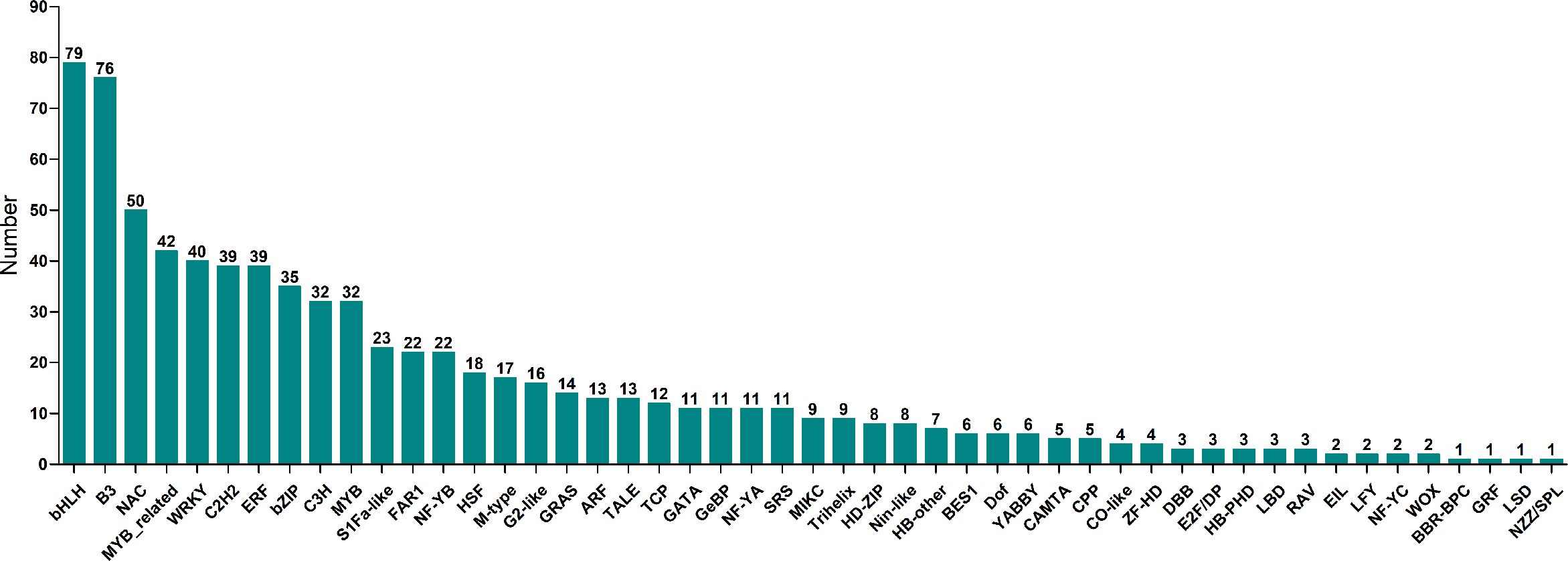
Figure 4 Transcription factors number of 49 transcription factor family annotated in sub class 1.
To explore the functional genes involved in P. brassicae infection, DEGs in subclass 1 were chosen for further analysis. Figure 5 shows the Venn map of the DEGs obtained from subclass 1. There were 29 overlapping genes at all-time points. Moreover, 17 and 50 genes overlapped at the primary (3 and 7 DAI, respectively) and secondary (14, 21, 28, and 35 DAI) infection stages (Figure 5).
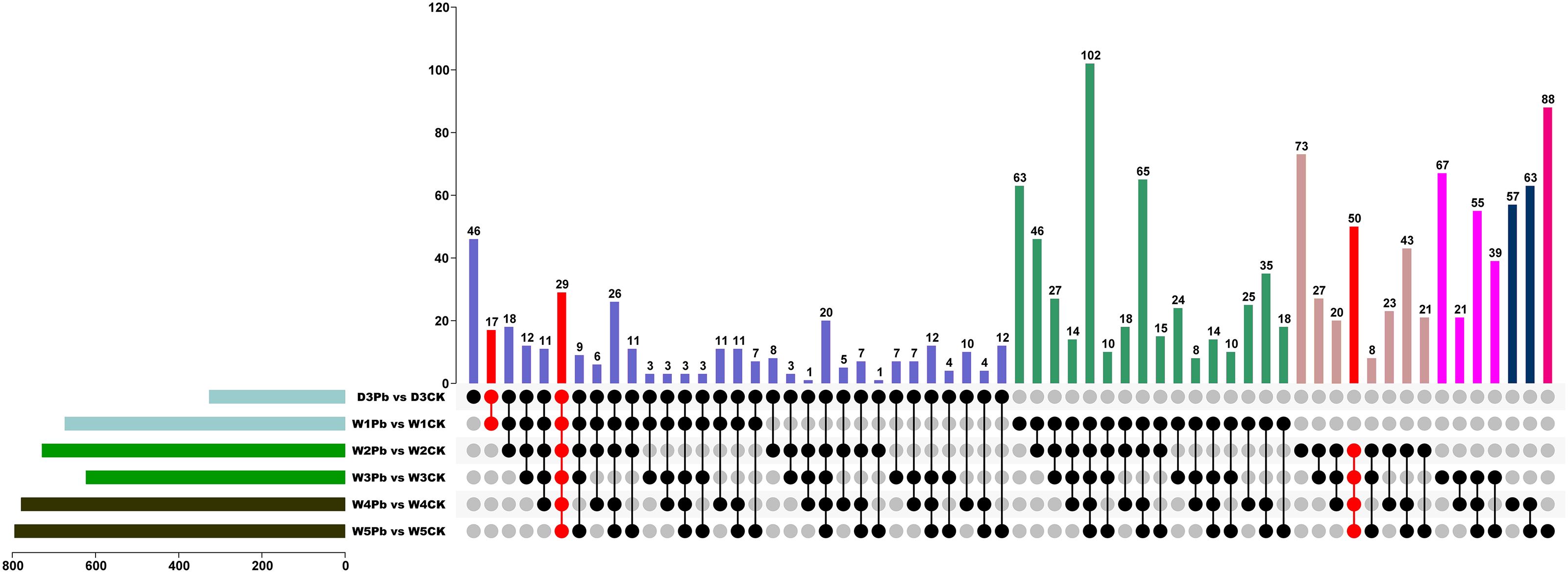
Figure 5 Venn diagram of differentially expressed genes (DEGs) from P. brassicae inoculation compared to distilled water at 3, 7, 14, 21, 28, and 35 days after treatment.
In both the primary and secondary infection stages, the 29 overlapping DEGs were predicted to be Brassica rapa putative disease resistance proteins, L-type lectin-domain-containing receptor kinases, ferredoxin–NADP reductases, 1-aminocyclopropane-1-carboxylate synthases, histone deacetylases, UDP-glycosyltransferases, putative glycerol-3-phosphate transporters, and chlorophyll a-b binding proteins, which are closely related to plant defense responses, biosynthetic processes, carbohydrate transport, and photosynthesis (Figure 6A; Supplementary Table 1).
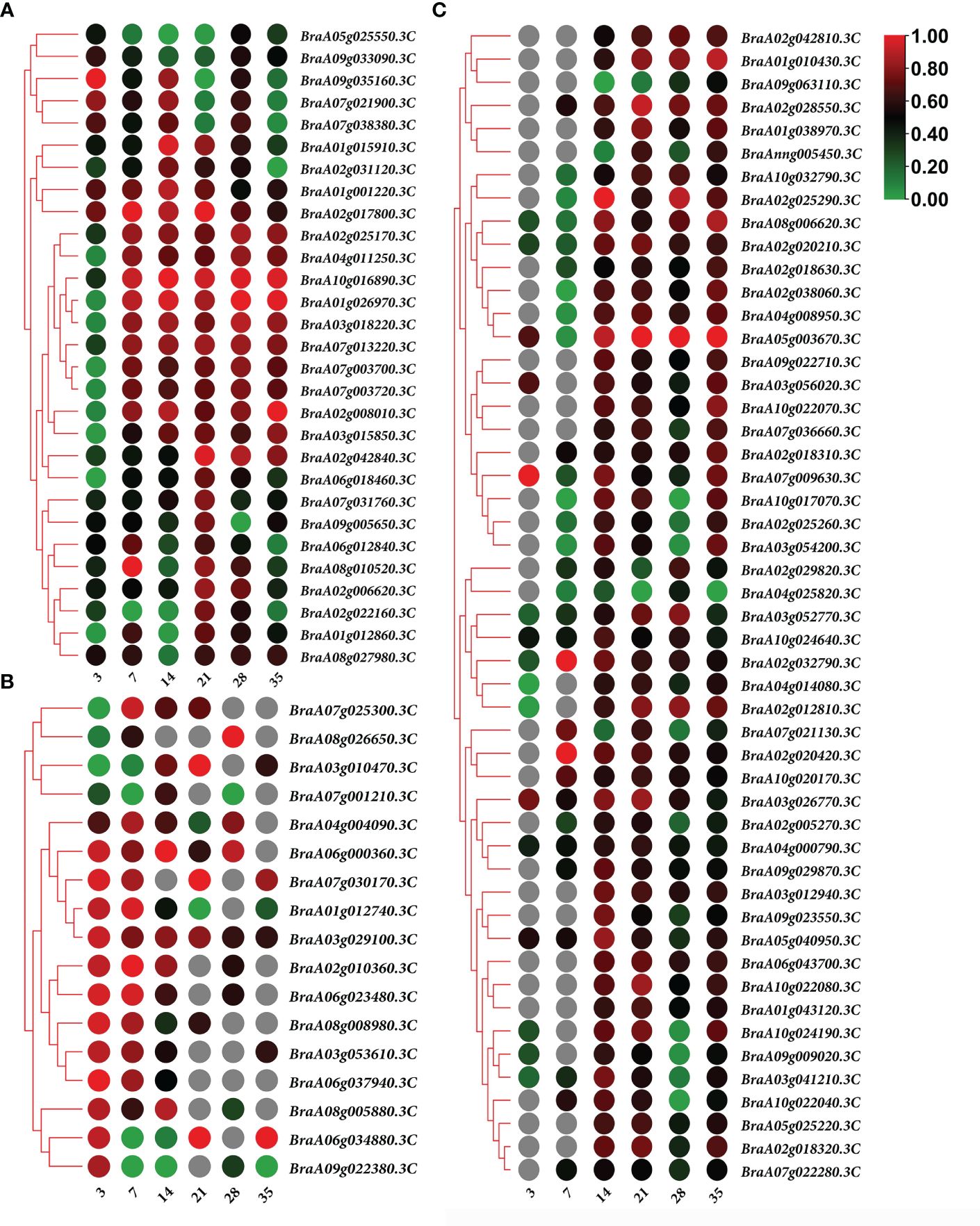
Figure 6 Heatmap of gene expression. (A) 29 overlapping differentially expressed genes (DEGs) expression heatmap in both of the primary and secondary infection stages. (B) 17 overlapping DEGs expression heatmap in primary infection stage. (C) 50 overlapping DEGs expression heatmap in the secondary infection stage. Notes: Numbers 3, 7, 14, 21, 28, and 35 indicate the different stages at 3, 7, 14, 21, 28, and 35 days after treatment, respectively.
The predicted Brassica rapa probable pectate lyase, RPM1-interacting protein 4-like, L-type lectin-domain-containing receptor kinase, Brassica napus G-type lectin S-receptor-like serine, Brassica rapa photosystem II 22 kDa protein and MLP-like protein were all upregulated during primary infection (Figure 6B; Supplementary Table 2). Meanwhile, the predicted Brassica rapa cation/H(+) antiporter and Brassica napus uncharacterized LOC106429977 were downregulated during primary infection (Figure 6B; Supplementary Table 2).
In the secondary infection stage, 45 out of 50 overlapping DEGs were upregulated, including predicted as Brassica rapa endoglucanase, long-chain acyl-CoA synthetase, probable WRKY transcription factor, NAC domain-containing protein, cell division control protein, auxin-induced protein, protein variation in compound triggered root growth response-like, and Brassica napus xyloglucan glycosyltransferase (Figure 6C; Supplementary Table 3). Only 5 DEGs BraA02g029820.3C, BraA07g021130.3C, BraA09g063110.3C, BraA04g025820.3C and BraAnng005450.3C, identified as Brassica rapa kinesin-like protein NACK1, cell division control protein, and uncharacterized, were downregulated at certain time points (Figure 6C; Supplementary Table 3).
Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway enrichment analysis, 131, 131, 132, 131, 132, and 133 KEGG pathways were associated with the infection of P. brassicae at 3, 7, 14, 21, 28, and 35 days after treatment. The top 10 enriched pathways are shown in Figure 7; Supplementary Table 4. KEGG analysis revealed significant enrichment in plant hormone signal transduction, plant-pathogen interactions, starch and sucrose metabolism, MAPK signaling, cysteine and methionine metabolism, carbon metabolism, and phenylpropanoid biosynthesis pathways, all of which are related to the infection process of P. brassicae.
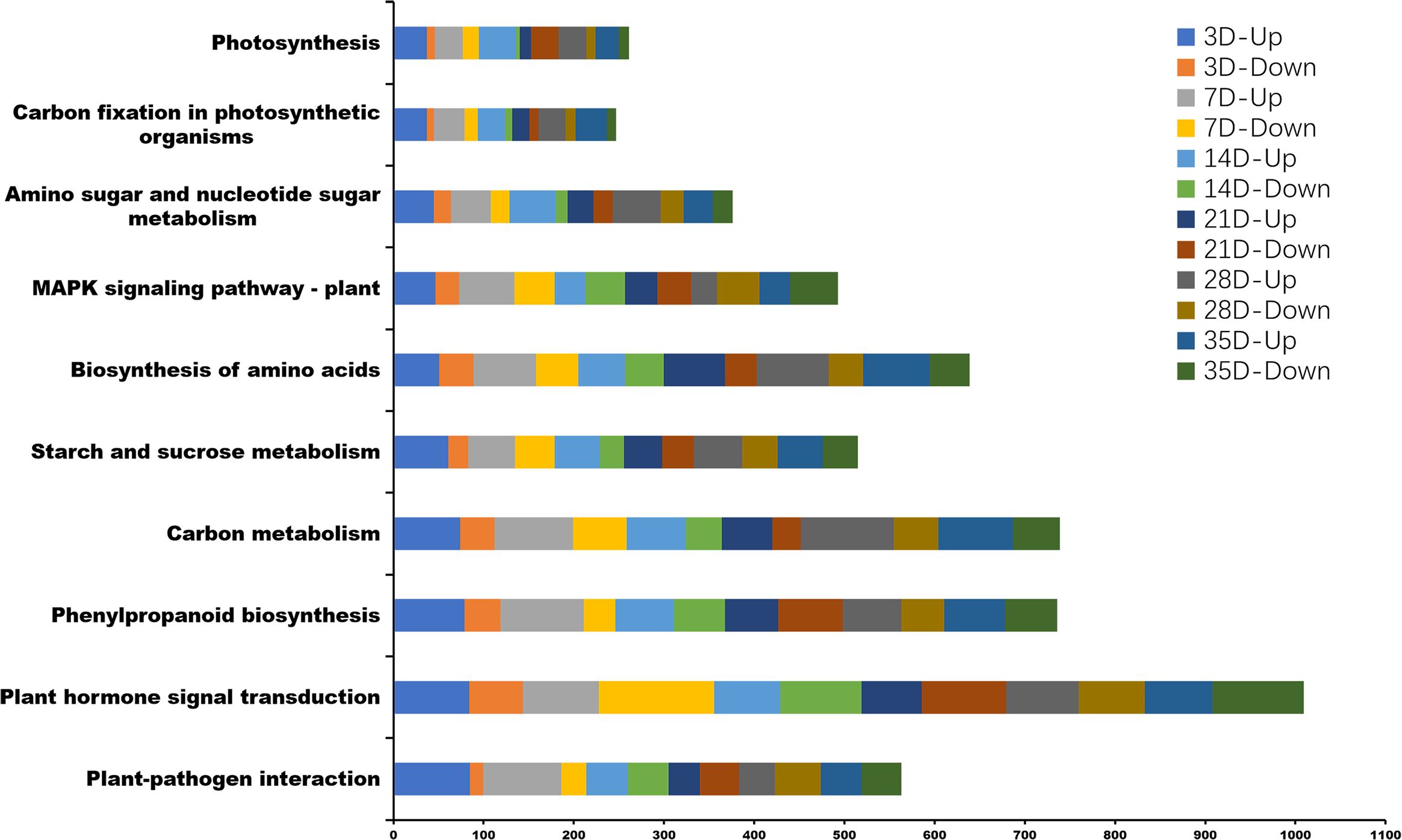
Figure 7 The top 10 Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway enrichment analysis between P. brassicae inoculation compared to distilled water at 3, 7, 14, 21, 28, and 35 days after treatment.
Differentially expressed transcription factors were selected from primary, secondary, and primary and secondary infection stages. Three genes were selected at each stage for quantitative real-time PCR analysis. The primers used for qRT-PCR are listed in Supplementary Table 5. The qRT-PCR results showed the same expression pattern as the RNA-Seq data (Figure 8). During the primary infection stage, BraA01g012740.3C (Brassica rapa probable pectate lyase 16) and BraA03g053610.3C (Brassica napus G-type lectin S-receptor-like serine/threonine-protein kinase) were significantly upregulated at 3 and 7 DPI. However, BraA03g010470.3C (Brassica rapa cation/H(+) antiporter 9-like) was significantly downregulated at 3 and 7 DPI (Figure 8A). During the secondary infection stage, the WRKY transcription factor BraA02g028550.3C (Brassica rapa long-chain acyl-CoA synthetase 5-like) and BraA02g042810.3C (Brassica rapa histone deacetylase 5-like) were significantly upregulated (Figure 8B). Both BraA01g012860.3C (Brassica rapa putative cysteine-rich receptor-like protein kinase) and BraA02g008010.3C (Brassica rapa putative disease resistance protein) were downregulated at 3DPI and then, upregulated, 7,14,21,28 and 35 DPI (Figure 8C).
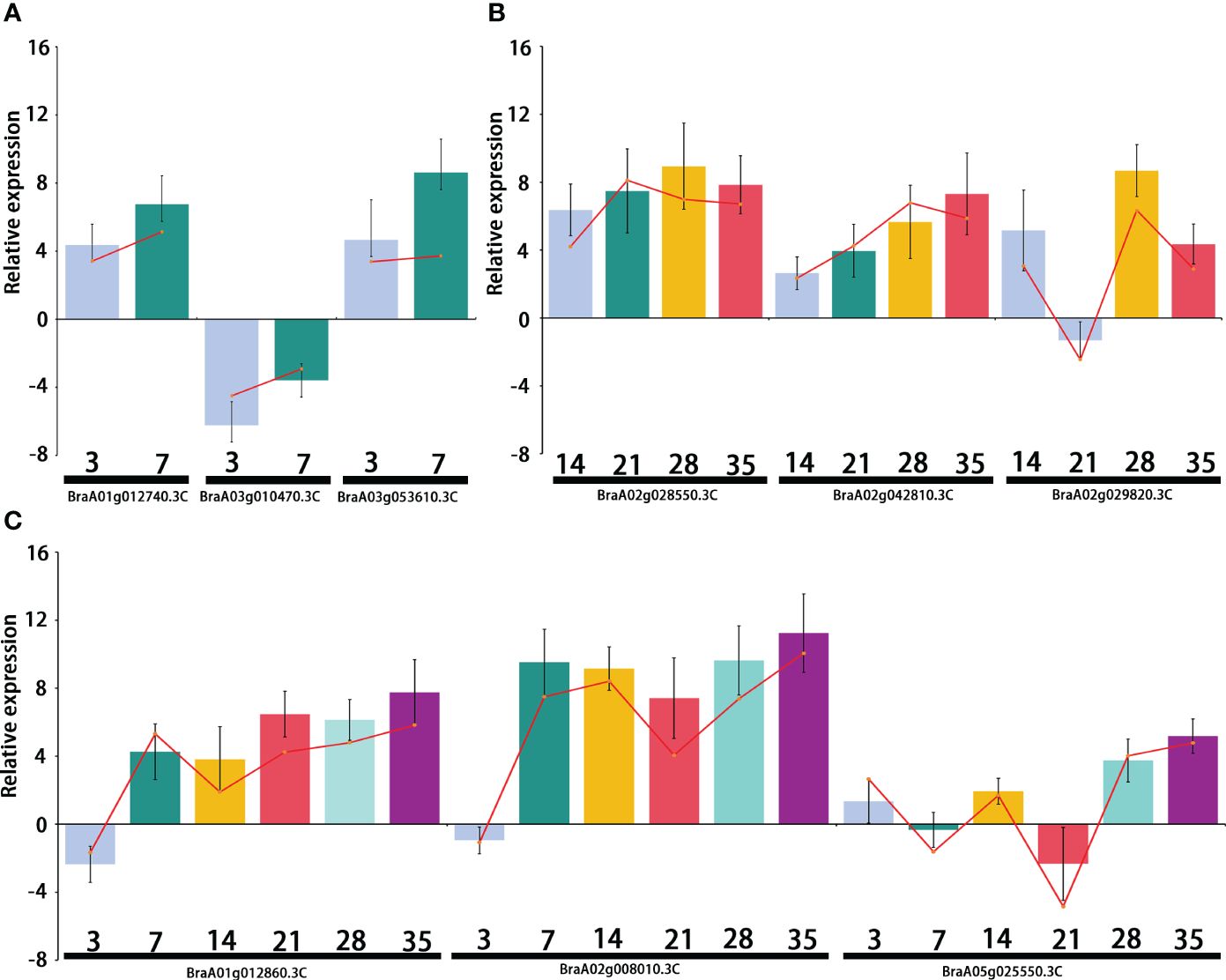
Figure 8 Relative expression of the differentially expressed genes (DEGs) selected from the primary infection, secondary infection, and primary and secondary infection stages, respectively. (A) Relative expression of DEGs from the primary infection stage; (B) Relative expression of DEGs from the secondary infection stage; (C) Relative expression of DEGs from the primary and secondary infection stages. Bar chat graph represents the gene expression by comparing P. brassicae inoculation to distilled water treatment from qRT-PCR at different stages, and the red line represents the gene expression from RNA-seq.
Infection with P. brassicae leads to increased cell division in both the cambium and the phloem parenchyma of Arabidopsis (Kobelt et al., 2000). Tumors in higher plants are abnormal tissue outgrowths resulting from the uncontrolled proliferation of a group of cells (Dodueva et al., 2020). Therefore, clubs that form on the roots of infected plants caused by the obligate biotrophic protist P. brassicae also called tumors, were described by Dodueva et al. (2020) in a review. Clubroot causes considerable economic damage owing to reduced crop yields in cultivated cruciferous crops. The prevention and management of clubroots has become a global challenge. Transcriptomic and genomic studies of the interactions between P. brassicae and cruciferous plants have provided valuable insights into the molecular mechanisms underlying these interactions (Zhao et al., 2017; Irani et al., 2018; Li et al., 2023). Studies have identified differentially expressed genes and signaling pathways associated with the response of cruciferous plants to P. brassicae infection (Grsic-Rausch et al., 2000; Devos et al., 2005; Ma et al., 2022).
The cell wall provides the first line of defense in plants and plays an important role in disease resistance (Wan et al., 2021). Microorganisms have evolved various strategies to break down cell walls. Pectin is a major component of primary cell walls and plays an important role in cell wall formation in higher plants. During the primary infection, the predicted B. rapa probable pectate lyase gene BraA01g012740.3C was upregulated by infection with P. brassicae (Figure 6B). BraA01g012740.3C encodes an HD-ZIP family protein that activates Poly(1,4-alpha-D-galacturonate)(n) via the unsaturated D-galacturonate pathway. A series of pectin enzymes are secreted by microorganisms to directly break down de-esterified HG, and infection attempts have been reported (Lionetti et al., 2012). Moreover, Pst DC3000 hijacks the host signaling pathway, which induces cell wall remodeling during plant development in Arabidopsis (Wang et al., 2017). IDL6 is upregulated upon infection with Pst DC3000, which then activates the HAE/HSL2 pathway, thereby promoting pectin degradation. These results indicated that Pst DC3000 can enhance infection by hijacking the IDL6-HAE/HSL2-ADPG2 signaling pathway (Wang et al., 2017). RPM1-interacting protein 4 (RIN4) is a conserved plant immunity regulator that can be modified by pathogenic effector proteins that plays an important role in both PAMP-triggered (PTI) and effector-triggered immunity (ETI) (Zhao et al., 2021). Kim et al. (2005) showed that RIN4 is a negative regulator of regulator of PAMP signaling, as the overexpression of RIN4 results in reduced defense responses. BraA02g010360.3C, predicted to be B. rapa RPM1-interacting protein 4-like, was highly upregulated by infection with P. brassicae during primary infection (Figure 6B). Moreover, L-type lectin domain-containing receptor kinase (BraA03g029100.3C, S1Fa-like), B. napus G-type lectin S-receptor-like serine (BraA03g053610.3C,TALE), and B. rapa photosystem II 22 kDa protein (BraA08g005880.3C) were upregulated at the primary infection stage. L-type lectin receptor kinases (LecRKs) recognize a variety of invasion patterns because of their large diversity. LecRKs are key players in plant immunity; however, their functions in plant defense are not well understood (Wang and Bouwmeester, 2017). The G-type lectin S-receptor-like serine/threonine protein kinase has been identified as a positive regulator of salt stress (Sun et al., 2013). Major latex-like proteins (MLP) confer resistance to pathogens by inducing pathogenesis-related protein genes. The MLP-like protein (BraA08g026650.3C) was downregulated at 3 DPI in P. brassicae. All of these DEGs were closely related to pathogen invasion induced by the infection of P. brassicae, offering insights for future investigations.
P. brassicae can manipulate Brassicaceae hosts by hijacking plant carbohydrate metabolism pathways to generate a strong physiological sink, such as accumulating abundant starch grains in infected roots (Malinowski et al., 2012; Ma et al., 2022). Our study showed that putative glycerol-3-phosphate transporter (BraA08g010520.3C), B. rapa endoglucanase 9-like (BraA02g020420.3C, WRKY), and long-chain acyl-CoA synthetase (BraA02g028550.3C, WRKY) were upregulated during P. brassicae secondary infection stage. A recent study reported that glucose transporters and glucose content significantly increase during the late stages of root infection (Kong et al., 2022). Moreover, previous studies have shown that the upregulation of the MEX1 maltose transporter and starch synthesis pathway (BrAGPS2 and BrISA2b) is activated by the growing P. brassicae plasmodia to mediate the energy supply from the host to the pathogen (Badstöber et al., 2020; Ma et al., 2022).
In the present study, transcription factors related to carbohydrate synthesis and transportation were found to play important roles in the invasion and proliferation of P. brassicae. Transcription factors such as BraA02g018320.3C, BraA03g056020.3C, BraA07g022280.3C, BraA10g022040.3C, BraA10g022070.3C, BraA10g022080.3C, BraA10g024190.3C and BraA10g024640.3C all belong to the bHLH family. These genes annotated as B. rapa protein variation in compound-triggered root growth response-like, B. rapa lectin domain-containing receptor kinase VI.3-like, B. rapa auxin-induced protein 15A-like, B. rapa transcription factor PRE2, B. rapa cytochrome P450 were upregulated during the secondary infection stage. Lectins serve as sugar code readers and reversibly bind to specific carbohydrates (Naithani et al., 2021). Evidence has shown that L-type lectin receptor kinase is involved in the resistance response to the pathogenic oomycetes P. infestans and P. capsici and fungus A. brassicicola (Wang et al., 2014). Auxins are also involved in invasion and gall formation during P. brassicae infection (Malinowski et al., 2016), and the auxin-induced protein-encoding gene 15A-like (BraA10g022040.3C, BraA10g022070.3C, BraA10g022080.3C) was upregulated, strengthening the result that auxin and cytokines play a key role during gall formation (Devos et al., 2006). The cytochrome P450 (CYP) superfamily catalyzes a wide range of reactions and plays important roles in several fundamental biological processes, such as steroid synthesis, fatty acid metabolism, and chemical defense (Pankov et al., 2021). A total of 258 non-redundant P450 genes have been identified, and P450 genes may play essential roles in pathogen-triggered immunity (PTI) in Chinese cabbage (Zhang et al., 2021).
In the context of Plasmodiophora brassicae infection, transcription factor families including bHLH, B3, NAC, MYB_related, WRKY, bZIP, C2H2, and ERF regulate the expression of genes related to defense responses, cell wall modifications, hormone signaling, and other processes involved in the plant’s defense mechanisms against the pathogen. Their coordination is essential for orchestrating an effective response to the infection and is likely crucial for the plant’s ability to combat the pathogen. Overall, transcript and genomic studies have provided valuable insights into the molecular mechanisms underlying interactions between P. brassicae and cruciferous plants. Understanding these molecular mechanisms can help to develop strategies for the management and control of clubroot diseases. In summary, this interaction involves complex changes in gene expression in both organisms, with the pathogen actively modulating plant defense and development through the secretion of effector proteins. Further research on the transcription factors related to carbohydrate metabolism pathways will provide more evidence for P. brassicae host manipulation in cruciferous crops.
The datasets generated and analyzed during this study are available on reasonable requests from the corresponding authors. The RNA-Seq data of this study has been deposited at NCBI under the BioProject ID PRJNA1060352.
SM: Data curation, Formal Analysis, Writing – original draft. XY: Data curation, Formal Analysis, Writing – original draft. YP: Data curation, Writing – review & editing. SL: Data curation, Writing – review & editing. XW: Data curation, Writing – review & editing. JJ: Data curation, Writing – review & editing. YL: Data curation, Writing – review & editing. WP: Data curation, Funding acquisition, Project administration, Resources, Writing – review & editing.
The author(s) declare financial support was received for the research, authorship, and/or publication of this article. This study was supported by grants from the National Natural Science Foundation of China (Project No. 32272720); the Liaoning Natural Science Foundation (2021-MS-229); the Fundamental Research Program for Liaoning Higher Education Institution (LJKMZ20221046) and the Shenyang Science and Technology Project (23–410-2–10).
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fpls.2024.1391173/full#supplementary-material
Agarwal, A., Kaul, V., Faggian, R., Rookes, J. E., Ludwig-Müller, J., Cahill, D. M. (2011). Analysis of global host gene expression during the primary phase of the Arabidopsis thaliana-Plasmodiophora brassicae interaction. Funct. Plant Biol. 38, 462–475. doi: 10.1071/FP11026
Badstöber, J., Gachon, C. M.M., Ludwig-Müller, J., Sandbichler, A. M., Neuhauser, S. (2020). Demystifying biotrophs: FISHing for mRNAs to decipher plant and algal pathogen–host interaction at the single cell level. Scientific Reports 10.
Chen, C., Chen, H., Zhang, Y., Thomas, H. R., Frank, M. H., He, Y., et al. (2020). TBtools: an integrative toolkit developed for interactive analyses of big biological data. Mol. Plant 13, 1194–1202. doi: 10.1016/j.molp.2020.06.009
Chen, H., Wang, T., He, X., Cai, X., Lin, R., Liang, J., et al. (2022). Brad v3.0: an upgraded brassicaceae database. Nucleic Acids Res. 50 (D1), D1432–D1441. doi: 10.1093/nar/gkab1057
Chu, M., Song, T., Falk, K. C., Zhang, X., Liu, X. (2014). Fine mapping of Rcr1 and analyses of its effect on transcriptome patterns during infection by Plasmodiophora brassicae. BMC Genomics 15, 1166. doi: 10.1186/1471-2164-15-1166
Ciaghi, S., Schwelm, A., Neuhauser, S. (2019). Transcriptomic response in symptomless roots of clubroot infected kohlrabi (Brassica oleracea var. gongylodes) mirrors resistant plants. BMC Plant Biol. 19. doi: 10.1186/s12870-019-1902-z
Devos, S., Laukens, K., Deckers, P., Van Der Straeten, D., Beeckman, T., Inzé, D., et al. (2006). A hormone and proteome approach to picturing the initial metabolic events during. Mol. Plant Microbe Interact. 19, 1431–1443. doi: 10.1094/MPMI-19-1431
Devos, S., Vissenberg, K., Verbelen, J.-P., Prinsen, E. (2005). Infection of Chinese cabbage by Plasmodiophora brassicae leads to a stimulation of plant growth: impacts on cell wall metabolism and hormone balance. New Phytol. 166, 241–250. doi: 10.1111/j.1469-8137.2004.01304.x
Dixon, G. R. (2009a). Plasmodiophora brassicae (Clubroot): A plant pathogen that alters host growth and productivity. J. Plant Growth Regul. 28, 193. doi: 10.1007/s00344-009-9087-6
Dixon, G. R. (2009b). The occurrence and economic impact of Plasmodiophora brassicae and clubroot disease. J. Plant Growth Regul. 28, 194–202. doi: 10.1007/s00344-009-9090-y
Dodueva, I. E., Lebedeva, M. A., Kuznetsova, K. A., Gancheva, M. S., Paponova, S. S, Lutova, L. L. (2020). Plant tumors: a hundred years of study. Planta 251, 82. doi: 10.1007/s00425-020-03375-5
Feng, J., Cao, T., Rennie, D. C., Strelkov, S. E., Hwang, S. F. (2014). Host-parasite interactions in clubroot of crucifers, Canadian. J. Plant Pathol. 36:sup1, 113–121. doi: 10.1080/07060661.2013.860192
Feng, J., Hwang, S. F., Strelkov, S. E. (2013a). Studies into primary and secondary infection processes by Plasmodiophora brassicae on canola. Plant Pathol. 62, 177–183. doi: 10.1111/j.1365-3059.2012.02612.x
Feng, J., Hwang, S. F., Strelkov, S. E. (2013b). Assessment of gene expression profiles in primary and secondary zoospores of Plasmodiophora brassicae by dot blot and real-time PCR. Microbiol. Res. 168, 518–524. doi: 10.1016/j.micres.2013.02.011
Grsic-Rausch, S., Kobelt, P., Siemens, J. M., Bischof, M., Ludwig-Müller, J. (2000). Expression and localization of nitrilase during symptom development of the clubroot disease in Arabidopsis. Plant Physiol. 122, 369–378. doi: 10.1104/pp.122.2.369
Hatakeyama, K., Suwabe, K., Tomita, R. N., Kato, T., Nunome, T., Fukuoka, H., et al. (2013). Identification and characterization of Crr1a, a gene for resistance to clubroot disease (Plasmodiophora brassicae Woronin) in Brassica rapa L. PloS One 8, e54745. doi: 10.1371/journal.pone.0054745
Ikegami, H., Watanabe, Y., Ito, T., Imuro, Y. (1984). Ultrastructures of starch grains in Chinese cabbage and turnip root cells infected with Plasmodiophora brassicae (Studies on the Clubroot of Cruciferous Plants 6). Res. Bull. Fac. Agr. Gifu. Univ. 49, 37–48.
Irani, S., Trost, B., Waldner, M., Nayidu, N., Tu, J., Kusalik, A. J., et al. (2018). Transcriptome analysis of response to Plasmodiophora brassicae infection in the Arabidopsis shoot and root. BMC Genomics 19, 23. doi: 10.1186/s12864-017-4426-7
Jubault, M., Hamon, C., Gravot, A., Lariagon, C., Delourme, R., Bouchereau, A., et al. (2013). Differential regulation of root arginine catabolism and polyamine metabolism in clubroot-susceptible and partially resistant Arabidopsis genotypes. Plant Physiol. Biochem. 70, 195–203. doi: 10.1104/pp.108.117432
Kageyama, K., Asano, T. (2009). Life cycle of Plasmodiophora brassicae. J. Plant Growth Regul. 28, 203–211. doi: 10.1007/s00344-009-9101-z
Kim, M. G., da Cunha, L., McFall, A. J., Belkhadir, Y., DebRoy, S., Dangl, J. L., et al. (2005). Two Pseudomonas syringae type III effectors inhibit RIN4-regulated basal defense in Arabidopsis. Cell 121, 749–759. doi: 10.1016/j.cell.2005.03.025
Kobelt, P., Siemens, J., Sacristan, M. D. (2000). Histological characterization of the incompatible interaction between Arabidopsis thaliana and the obligate biotrophic pathogen Plasmodiophora brassicae. Mycol. Res. 2, 220–225. doi: 10.1017/S0953756299001781
Kong, L., Li, X., Zhan, Z., Piao, Z. (2022). Sugar transporters in Plasmodiophora brassicae: genome-wide identification and functional verification. Int. J. Mol. Sci. 23, 5264. doi: 10.3390/ijms23095264
Kuginuki, Y., Yoshigawa, H., Hirai, M. (1999). Variation in virulence of Plasmodiophora brassicae in Japan tested with clubroot resistant cultivars of Chinese cabbage. Eur. J. Plant Pathol. 105, 327–332. doi: 10.1023/A:1008705413127
Li, P., Lv, S., Zhang, Z., Su, T., Wang, W., Xin, X., et al. (2023). Genome assembly of the plant pathogen Plasmodiophora brassicae reveals novel secreted proteins contributing to the infection of Brassica rapa. Hortic. Plant J. doi: 10.1016/j.hpj.2023.09.001
Lionetti, V., Cervone, F., Bellincampi, D. (2012). Methyl esterification of pectin plays a role during plant-pathogen interactions and affects plant resistance to diseases. J. Plant Physiol. 169, 1623–1630. doi: 10.1016/j.jplph.2012.05.006
Liu, L., Qin, L., Zhou, Z., Hendriks, W. G. H. M., Liu, S., Wei, Y. (2020). Refining the life cycle of Plasmodiophora brassicae. Phytopathology 110, 1704–1712. doi: 10.1094/PHYTO-02-20-0029-R
Livak, K. J., Schmittgen, T. D. (2001). Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 25, 402–408. doi: 10.1006/meth.2001.1262
Ludwig-Müller, J., Prinsen, E., Rolfe, S. A., Scholes, J. D. (2009). Metabolism and plant hormone action during clubroot disease. J. Plant Growth Regul. 28, 229–244. doi: 10.1007/s00344-009-9089-4
Ma, Y., Choi, S. R., Wang, Y., Chhapekar, S. S., Zhang, X., Wang, Y., et al. (2022). Starch content changes and metabolism related gene regulation of Chinese cabbage synergistically induced by plasmodiophora brassicae infection. Hortic. Res. 9. doi: 10.1093/hr/uhab071
Malinowski, R., Novák, O., Borhan, M. H., Spíchal, L., Strnad, M., Rolfe, S. A. (2016). The role of cytokinins in clubroot disease. Eur. J. Plant Pathol. 145, 543–557. doi: 10.1007/s10658-015-0845-y
Malinowski, R., Smith, J. A., Fleming, A. J., Scholes, J. D., Rolfe, S. A. (2012). Gall formation in clubroot-infected Arabidopsis results from an increase in existing meristematic activities of the host but is not essential for the completion of the pathogen life cycle. Plant J. 71, 226–238. doi: 10.1111/j.1365-313X.2012.04983.x
Naithani, S., Komath, S. S., Nonomura, A., Govindjee, G. (2021). Plant lectins and their many roles: Carbohydrate-binding and beyond. J. Plant Physiol. 266, Article 153531. doi: 10.1016/j.jplph.2021.153531
Pang, W., Liang, Y., Zhan, Z., Li, X., Piao, Z. (2020). Development of a sinitic clubroot diferential set for the pathotype classifcation of Plasmodiophora brassicae. Front. Plant Sci. 11, 568771. doi: 10.3389/fpls.2020.568771
Pang, W., Zhang, X., Ma, Y., Wang, Y., Zhan, Z., Piao, Z. (2022). Fine mapping and candidate gene analysis of CRA3.7 conferring clubroot resistance in Brassica rapa. Theor. Appl. Genet. 135, 4541–4548. doi: 10.1007/s00122-022-04237-2
Pankov, K. V., McArthur, A. G., Gold, D. A., Nelson, D. R., Goldstone, J. V., Wilson, J. Y. (2021). The cytochrome P450 (CYP) superfamily in cnidarians. Sci. Rep. 11, 9834. doi: 10.1038/s41598–021-88700-y
Schuller, A., Ludwig-Müller, J. (2016). Histological methods to detect the clubroot pathogen Plasmodiophora brassicae during its complex life cycle. Plant Pathol. 65, 1223–1237. doi: 10.1111/ppa.12520
Shi, Z., Liu, Y., Noor, N., Bao, Y., Ming, R., Liu, T., et al. (2014). Gene expression profiles deciphering clubroot infection and defense mechanisms in Brassica genomes. PloS One 9, e98660. doi: 10.1007/s10142-013-0312-9
Stjelja, S., Fogelqvist, J., Tellgren-Roth, C., Dixelius, C. (2019). The architecture of the Plasmodiophora brassicae nuclear and mitochondrial genomes. Sci. Rep. 9, 15753. doi: 10.1038/s41598-019-52274-7
Strelkov, S. E., Hwang, S., Manolii, V. P., Cao, T., Feindel, D. (2016). Emergence of new virulence phenotypes of Plasmodiophora brassicae on canola (Brassica napus) in Alberta. Can. Eur. J. Plant Pathol. 145, 517–529. doi: 10.1007/s10658-016-0888-8
Struck, C., Rüsch, S., Strehlow, B. (2022). Control strategies of clubroot disease caused by Plasmodiophora brassicae. Microorganisms 10, Art. 3. doi: 10.3390/microorganisms10030620
Sun, X. L., Yu, Q. Y., Tang, L. L., Ji, W., Bai, X., Cai, H., et al. (2013). GsSRK, a G-type lectin S-receptor-like serine/threonine protein kinase, is a positive regulator of plant tolerance to salt stress. J. Plant Physiol. 170, 505–515. doi: 10.1016/j.jplph.2012.11.017
Ueno, H., Matsumoto, E., Aruga, D., Kitagawa, S., Matsumura, H., Hayashida, N. (2012). Molecular characterization of the CRa gene conferring clubroot resistance in Brassica rapa. Plant Mol. Biol. 80, 621–629.
Voorrips, R. E., Jongerius, M. C., Kanne, H. J. (2003). Quantitative trait loci for clubroot resistance in Brassica oleracea. Biotechnol. Agric. For. 52, 87–104. doi: 10.1094/MPMI-06-14-0191-R
Wan, J., He, M., Hou, Q., Zou, L., Yang, Y., Wei, Y., et al. (2021). Cell wall associated immunity in plants. Stress Biol. 1, 3. doi: 10.1007/s44154-021-00003-4
Wang, Y., Bouwmeester, K. (2017). L-type lectin receptor kinases: New forces in plant immunity. PloS Pathog. 13, e1006433. doi: 10.1371/journal.ppat.1006433
Wang, Y., Bouwmeester, K., Beseh, P., Shan, W., Govers, F. (2014). Phenotypic analyses of Arabidopsis T-DNA insertion lines and expression profiling reveal that multiple L-type lectin receptor kinases are involved in plant immunity. Mol. Plant-Microbe Interactions.: MPMI. 27, 1390. doi: 10.1094/MPMI-06-14-0191-R
Wang, W., Qin, L., Zhang, W., Tang, L., Zhang, C., Dong, X., et al. (2023). WeiTsing, a pericycle-expressed ion channel, safeguards the stele to confer clubroot resistance. Cell 186, 2656–2671 e2618. doi: 10.1016/j.cell.2023.05.023
Wang, X., Hou, S., Wu, Q., Lin, M., Acharya, B. R., Wu, D., et al. (2017). IDL6-HAE/HSL2 impacts pectin degradation and resistance to Pseudomonas syringae pv tomato DC3000 in Arabidopsis leaves. Plant J. Cell Mol. Biol. 89, 250–263. doi: 10.1111/tpj.13380
Zhang, C., Du, C., Li, Y., Wang, H., Zhang, C., Chen, P. (2023). Advances in biological control and resistance genes of brassicaceae clubroot disease-the study case of China. Int. J. Mol. Sci. 24, 785. doi: 10.3390/ijms24010785
Zhang, S., Wu, Q. R., Zhang, H. M., Pei, Z. M., Gao, J. W. (2021). Genome-wide identification and transcriptomic data exploring of the cytochrome P450 family in Chinese cabbage (Brassica rapa L. ssp. pekinensis). J. Plant Interact. 16, 136–155. doi: 10.1080/17429145.2021.1909761
Zhang, F., Yu, J., Cui, X., Shen, X., Dong, X. (2016). Transcriptomic analysis of Brassica napus the highly resistant genotype infected with Plasmodiophora brassicae. BMC Genomics 17, 836.
Zhao, Y., Bi, K., Gao, Z., Chen, T., Liu, H., Xie, J., et al. (2017). Transcriptome Analysis of Arabidopsis thaliana in Response to Plasmodiophora brassicae during Early Infection. Front. Microbiol. 8, 673. doi: 10.3389/fmicb.2017.00673
Zhao, G., Guo, D., Wang, L., Li, H., Wang, C., Guo, X. (2021). Functions of RPM1-interacting protein 4 in plant immunity. Planta 253, 11. doi: 10.1007/s00425-020-03527-7
Keywords: Plasmodiophora brassicae, Chinese cabbage, clubroot, transcription factor, intracellular
Citation: Meng S, Yan X, Piao Y, Li S, Wang X, Jiang J, Liang Y and Pang W (2024) Multiple transcription factors involved in the response of Chinese cabbage against Plasmodiophora brassicae. Front. Plant Sci. 15:1391173. doi: 10.3389/fpls.2024.1391173
Received: 25 February 2024; Accepted: 20 May 2024;
Published: 06 June 2024.
Edited by:
Xiangshu Dong, Yunnan University, ChinaReviewed by:
Su Ryun Choi, Seeders, Daejeon, Republic of KoreaCopyright © 2024 Meng, Yan, Piao, Li, Wang, Jiang, Liang and Pang. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Wenxing Pang, cHd4c3lhdUBzeWF1LmVkdS5jbg==
†These authors have contributed equally to this work
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.