
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
REVIEW article
Front. Plant Sci. , 17 January 2024
Sec. Functional and Applied Plant Genomics
Volume 14 - 2023 | https://doi.org/10.3389/fpls.2023.1324176
 Abdul Rehman1,2
Abdul Rehman1,2 Sana Khan3
Sana Khan3 Fenlei Sun1
Fenlei Sun1 Zhen Peng1,2Keyun Feng4Ning Wang4Yinhua Jia1,2Zhaoe Pan1
Zhen Peng1,2Keyun Feng4Ning Wang4Yinhua Jia1,2Zhaoe Pan1 Shoupu He1,2,5
Shoupu He1,2,5 Lidong Wang1,2
Lidong Wang1,2 Abdul Qayyum6
Abdul Qayyum6 Xiongming Du1,2*
Xiongming Du1,2* Hongge Li1,2*
Hongge Li1,2*Plants experience diverse abiotic stresses, encompassing low or high temperature, drought, water logging and salinity. The challenge of maintaining worldwide crop cultivation and food sustenance becomes particularly serious due to drought and salinity stress. Sustainable agriculture has significant promise with the use of nano-biotechnology. Nanoparticles (NPs) have evolved into remarkable assets to improve agricultural productivity under the robust climate alteration and increasing drought and salinity stress severity. Drought and salinity stress adversely impact plant development, and physiological and metabolic pathways, leading to disturbances in cell membranes, antioxidant activities, photosynthetic system, and nutrient uptake. NPs protect the membrane and photosynthetic apparatus, enhance photosynthetic efficiency, optimize hormone and phenolic levels, boost nutrient intake and antioxidant activities, and regulate gene expression, thereby strengthening plant’s resilience to drought and salinity stress. In this paper, we explored the classification of NPs and their biological effects, nanoparticle absorption, plant toxicity, the relationship between NPs and genetic engineering, their molecular pathways, impact of NPs in salinity and drought stress tolerance because the effects of NPs vary with size, shape, structure, and concentration. We emphasized several areas of research that need to be addressed in future investigations. This comprehensive review will be a valuable resource for upcoming researchers who wish to embrace nanotechnology as an environmentally friendly approach for enhancing drought and salinity tolerance.
In the current situation, population expansion has become a serious barrier to maintaining sustainable food production to meet the increasing population’s requirements (Calicioglu et al., 2019). The global population is expected to extend 9.6 billion individuals by the year 2050, necessitating a significant 70–100% increase in food output to meet the demands of this expanding population (Alabdallah and Hasan, 2021). Revolutionizing conventional agricultural practices is urgently necessary to attain the United Nations sustainable growth objective of “Zero Hunger” by 2030. These changes may be implemented through eco-friendly and sustainable approaches (Rajput et al., 2021b; Ranjan et al., 2021). Crop yields are significantly impacted by several issues, including increased abiotic stresses, declining fertile land, excessive fertilizer, pesticide usage, climate change and global warming (Hasan et al., 2021a; Hasan et al., 2021b). The drastic decrease in agricultural output is a serious hazard to world food security and a major problem. The required steps must be taken to lessen the damaging effects of abiotic stressors on crops if global food security is to be maintained. Therefore, implementing appropriate measures becomes imperative to address this issue effectively (Hasan et al., 2021a). Plants are immobile; hence they cannot relocate physically to escape the effects of environmental challenges, especially abiotic stress. Abiotic stresses like soil salinity and drought can cause considerable reduction in crop yield and quality (Haider et al., 2021; Yadav et al., 2021). Salt stress causes the cytosol to accumulate with Na+ and chloride Cl- ions, severely damaging the cellular structure (Ranjan et al., 2021). The impacts of drought stress include the induction of stomatal closure, obstruction of photosynthesis, reduction of leaf area, inhibition of biomass and growth, reduction of water potential, elevation of osmolyte levels, and induction of reactive oxygen species (ROS) (Ibrahim et al., 2019). Hence, the modifications to plant metabolism under abiotic stress cause disturbances, resulting in the reorganization of the metabolic network to maintain crucial activities.
Over the last few decades, researchers have made great progress in creating a variety of stress management techniques. Among other strategies, nanotechnology is a highly effective strategy for significantly increasing crop yield (Shahid et al., 2020). However, most studies on NPs focused on examining their possible toxicity rather than their benefits (Ranjan et al., 2021). Nanotechnology has attracted much interest as an auspicious arena widely utilized in agriculture, food production, and medical (Alabdallah and Hasan, 2021). A tiny molecule cluster with an interfacial layer around it is called a nanoparticle with diameter 1 and 100 nanometers (Mohammadi et al., 2016). NPs have unique and uncommon features do not present in bulk materials because of their tiny size. Nanoparticle’s interesting scientific appeal arises from their capacity to link atomic or molecular structures to bulk materials (Khalid et al., 2022). NPs have been widely used in a variety of industries, including agricultural and allied ones as well as the chemical, optical, biomedical, pharmaceutical, culinary, and textile sectors (Verma et al., 2019). Due to their advantageous and safe uses, a number of NPs have recently attracted a lot of interest in the agriculture industry. These NPs comprise TiO2, Fe3O4, ZnO, SiO2, Cu-NPs, and Se-NPs (Alabdallah and Hasan, 2021; Hashem et al., 2021). There are precise methods to produce NPs, such as chemical, green, and physical synthesis (Akhtar et al., 2022). NPs can positively affect plant growth and development, although the precise impacts can vary depending on factors such as their time, origin, application, and size of crop administration (Rubilar et al., 2013). Current research exhibited that NPs protect plants by increasing the antioxidant metabolic activity to reduce oxidative damage (Ahmed et al., 2021). NPs regulate salt tolerance in diverse plant species by regulating hormone levels, antioxidant enzyme activities, ion balance, gene expression, and defense mechanisms (Zulfiqar and Ashraf, 2021). NPs can effects plant species and the environment in different ways like, NP shape, applications, and size, as well as environmental conditions, can influence their impact (Wahid et al., 2020). Nanopesticides and nano fertilizers are the two main use of nanotechnology in the agricultural industry.
Nanotechnology acts as a harbinger for a forthcoming industrial revolution. Agriculture’s productivity might be revolutionized by nanotechnology. According to a recent study, adding magnetic NPs through hydroponic system at seedling stage, significantly augmented the content of chlorophyll (a and b) and carotenoids in Hordeum vulgare. Additionally, the utilization of magnetic NPs favoured the genes related to photosystems (Tombuloglu et al., 2020). The utilization of NPs has been observed to mitigate the harmful effects of salinity and drought stress (Ali et al., 2021a; Singh et al., 2021). The effects of foliar spray of the NPs on plant growth and development are listed in Figure 1.
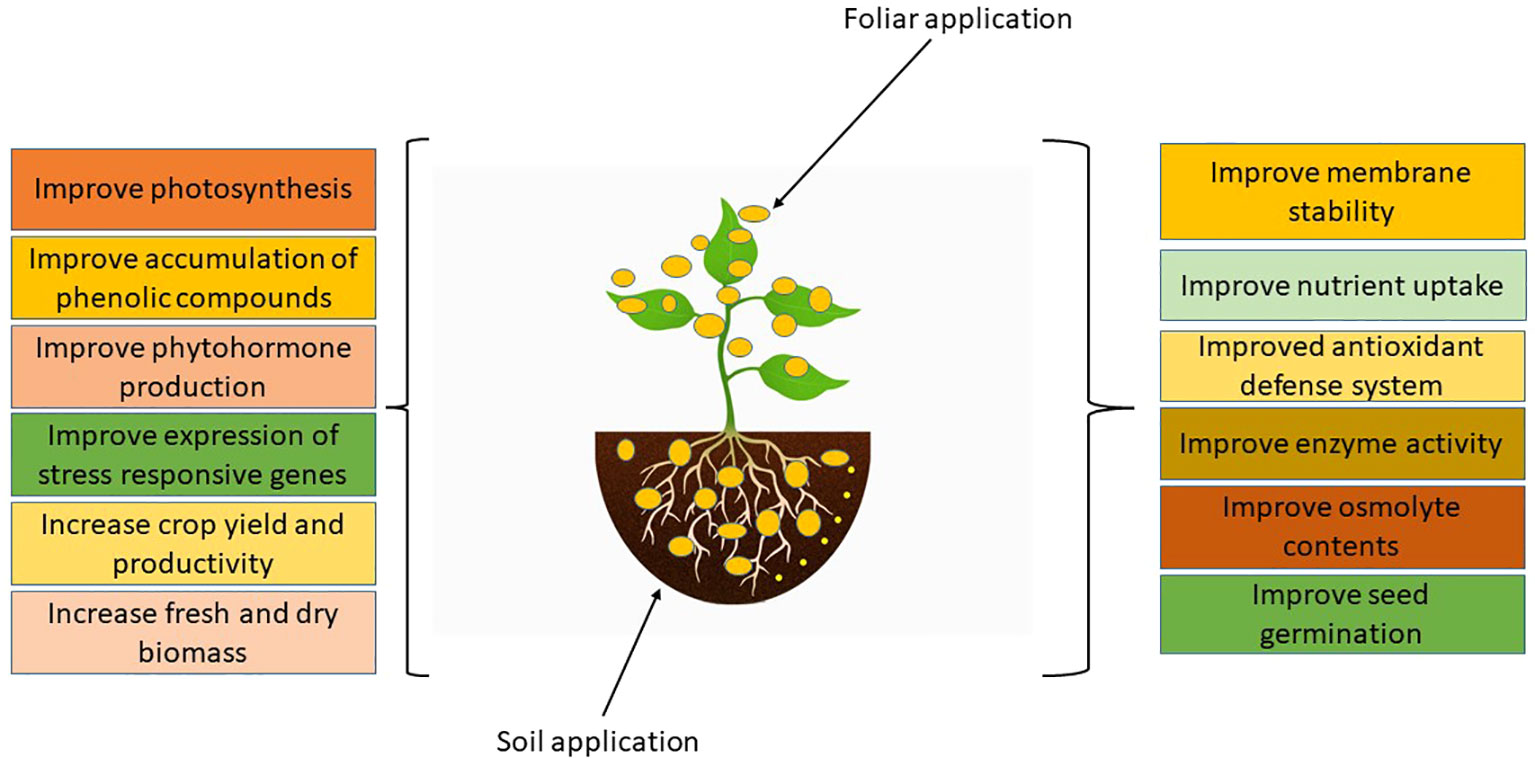
Figure 1 Impacts of NPs on plant growth and development.
The need for an appropriate categorization system for NPs eventually arose to advise scientists and engineers working on NPs studies and uses and encourage secure and more practical use of these resources. Studies have shown that the two most important factors in determining the classification of NPs are their dimensionality and composition. The need for an accurate categorization system to offer more convenience has grown during the last twenty years as the number of nanostructured materials has expanded (Prajitha et al., 2019). The “dimensionality” of the particles is the main criterion for categorizing NPs. According to Chung et al. (2013) NPs with zero dimension would effectively lack any observable dimension bigger than 100 nm in length. The production of zero-dimensional (0D) nanomaterials with precisely regulated dimensions has been made possible with the introduction of many physical and chemical manufacturing techniques (Figure 2).
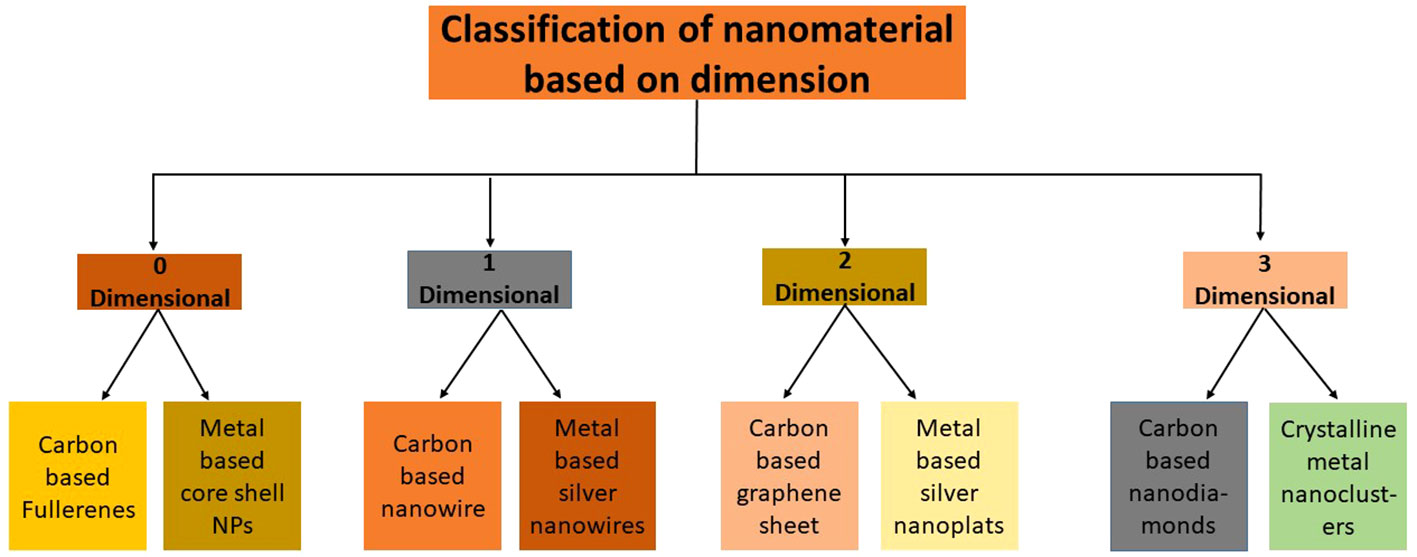
Figure 2 Classification of NPs based on dimension and composition.
Recently, numerous study groups have successfully improved various kinds of zero-dimensional (0D) nanomaterials, including but not limited to regular particle arrangements like quantum dots, varied particle arrangements, holy spheres, nano-lenses, and more (Kim et al., 2010). One dimension beyond the nanoscale distinguishes this class of nanomaterials, which we refer to as one-dimensional nanomaterials (1D). Nano horns, nanofibers, nanotubes nanowires, and nanorods are rare example. The two-dimensional (2-D) category of nanomaterials has two additional dimensions beyond the nanoscale. Nanolayers, nanofilms, and nanosheets are three prominent illustrations (Joudeh and Linke, 2022). Three-dimensional (3-D) materials, are not hindered by any dimension to the nanoscale. This category includes a variety of substances, including bulk powders, nanoparticle dispersions, arrays of nanowires, and nanotubes (Joudeh and Linke, 2022).
Next to dimensionality, the composition is also viewed appropriate criterion for nanoparticle categorization as it reliably reflects the chemical constitutes of the material. The categorization system includes NPs made entirely of carbon atoms as its initial category. This category contains substances like fullerenes, carbon nanotubes, graphene, and others. Carbon-based materials have several significant qualities, including exceptional strength and reactivity (Prajitha et al., 2019). The metal-based NPs, often known as metallic NPs, are categorized as the next group in this system. These NPs are made of metals with nanostructures, including titanium, gold, silver, and their equivalent oxides (Cataldo and Da Ros, 2008). This category also includes metal-decorated multi-walled nanotubes (MWNTs), metal nanoclusters, metal-filled single-walled nanotubes (SWNTs) and metallofullerenes. The optical characteristics of metallic NPs are vital in controlling their functions due to the peculiar surface plasmon resonance at visible wavelengths (Kumar et al., 2012). These polymeric NPs can contain the required therapeutic substance within their core due to their unique structural properties, or they can alternatively adsorb or be attached to their surface (Weir et al., 2012). The term “nanocomposites” refers to the next group in this categorization of NPs. Nanocomposites have been more popular over the past 25 years due to their wide variety of desired qualities that may be modified to meet particular needs (Kubiak, 2014). The distinctive physicochemical properties displayed by these materials result from the morphological and interstitial features of the component constituents.
Plants are the main producers in the environment, they are extremely important, and their interaction with NPs is a complicated process involving many variables. Plants are exposed to NPs found in soil and aerosols in their natural habitat (Azim et al., 2023). The uptake, translocation, and accumulation of NPs in plants are induced by various factors like the type, chemical composition, size, functionalization, root exudates, and microorganisms linked to the roots of the plant species (Etesami et al., 2021). The high mobility of NPs is governed by various factors, such as Brownian motion, Vander Waal forces, double-layer forces, and gravity. These elements are crucial for the adhesive properties of nanoparticles. NPs can enter into plants through air and soil when applied in liquid form (Azim et al., 2023). Aerosol NPs can reach plant aerial parts by various paths, including stomata, wounds, direct diffusion, or through aerial parts in contrast to soil NPs (Hussain et al., 2019). It is supposed that all the NPs behave differently in different plants and have different effects. Some researchers also explore that some NPs alter their shape and structure when exposed to plants like, silver, copper, and zinc oxide. It was observed that when ZnO NPs were applied on wheat they dissolve and release Zn ions in the root cells (Ali et al., 2021b). Likewise, CuO NPs transformed in to Cu(OH)2 and Cu20 in the roots of the soybean plants (Kim et al., 2017). Moreover, Sliver NPs converted into silver chloride and silver sulfide in the leaf of the Arabidopsis thaliana (Thounaojam et al., 2021). Ensuring the validity and reliability of determining the shape, size, structure, and composition of NPs before and after exposure to plant tissue is a great challenge (Zhang et al., 2021). So, there is dire need to identify the exact, precise, pertinent, trustworthy, and appropriate method to characterize the NPs.
NPs experience various modifications that are associated with their interactions, stability, and production in response to the environmental conditions. These modifications could be crucial in determining the properties and performance of NPs in different applications. The modifications carried out are of utmost importance to validate the safety, effectiveness, and overall influence of the NPs in plants.
NPs are synthesized through the alteration and transformation of precursor materials at the nanoscale level, resulting in the creation of distinct nanostructures. Diverse techniques are employed to synthesize NPs which include physical, biological, and chemical approaches. Various characterization methods play a crucial role in determining the physical properties of NPs, such as their size, shape, and surface properties. These factors have a significant impact on the performance and functionality of NPs, making their accurate determination essential for analyzing and optimizing their applications (Nel et al., 2006). NPs are often subject to surface modifications aimed at increasing their strength and enhancing their performance (Badawy et al., 2010).
NPs tend to either disperse or aggregate in the soil or water after their synthesis. The extent of dispersion or aggregation depends on multiple factors, such as the presence and concentration of organic substances, the pH level of the soil or water, and the concentration of various ions. Organic substances can either stabilize or destabilize the NPs, while the pH level and ion concentration can affect the electrostatic repulsion or attraction between the NPs. Understanding these factors is crucial for predicting and controlling the behavior of NPs in the environment (Azim et al., 2023). The stability, bioavailability, and movement of NPs are crucial factors that determine their interaction with roots. Prior to the interaction, these parameters play a pivotal role in determining the effectiveness and efficiency of the process (Baalousha et al., 2016).
NPs are often coated with a surface layer to improve their performance. These coatings can be modified using various factors such as ions, pH, and temperature. (Matzke et al., 2014). The process of surface coating plays a crucial role in the uptake of NPs, and also helps in controlling the germination and growth of plants (Biba et al., 2022). According to a recent study, it was found that the physiochemical properties of CRISPR Cas gene delivery are significantly influenced by the surface coating used. This highlights the importance of carefully selecting the coating material for effectiveness of NPs (Alallam et al., 2021).
The fate of NPs is significantly influenced by the oxidation and reduction reactions taking place in their surrounding environments. NPs composed of metals are particularly susceptible to oxidation, which can affect their interaction with plant tissues. Ultimately, the outcome of these chemical transformations plays a crucial role in determining the behavior and impact of NPs in different biological systems (Sotiriou and Pratsinis, 2010). The unique chemical properties of NPs can cause them to behave differently than bulk materials in terms of reactivity, conductivity, and strength. This is due to the high surface area to volume ratio of NPs, which can lead to increased surface reactivity and altered electronic properties (Verma et al., 2019; Staroń et al., 2020). As a result, it is important to consider the specific properties of NPs when designing and using them in various applications.
NPs display intricate interplay with the rhizosphere, encompassing a variety of factors such as microbes, minerals, organic matter, and other substances, which in turn, interact with the roots of plants (Ge et al., 2014). NPs are biologically inert and non-destructive, allowing them to persist in soil for extended periods. This can lead to alterations in soil microflora populations, soil fertility, and the metabolism and physiology of plant species (Pittol et al., 2017; Yang et al., 2017). The interactions demonstrate noteworthy impacts on the accessibility, transportation, and mobility of NPs in the soil system.
The behavior of NPs in transportation is significantly impacted by various factors, such as dissolved ions, water content, and soil structure. Due to their small size, NPs display increased mobility in soil water. (Klaine et al., 2008). NPs can penetrate plant cells either through the transport system or endocytosis. Once inside, they accumulate in plant tissues and interfere with plant molecules, leading to disruptions in the plant’s physiological, morphological, and anatomical features and activities (Burman and Kumar, 2018; García-Gómez et al., 2018).
NPs exhibited a significant role in increasing plant yield in drought and salinity. NPs demonstrated their potential to improve water loss by balancing water status ultimately abiotic stress tolerance (Rasheed et al., 2022). NPs also regulate stomatal conductance and transpiration rate through leaf anatomy and closing of stomata (Acosta-Motos et al., 2017). NPs demonstrated defensive impacts to protect photosynthetic machinery and increase photosynthesis in plants, also activate the antioxidant system to repair the damage caused by ROS in photosystems and chloroplasts. Moreover, NPs trigger electron transport chain and increase chlorophyll contents in plant cells (Forni et al., 2017; Manzoor et al., 2022). They also have influential effects on multiple physiological systems of plants including stress responsive mechanisms, hormone metabolism, biosynthesis of osmolytes, ethylene production, nitric oxide, ABA and calcium signaling. Additionally, regulate signal transduction pathways during salinity and drought stress and activate stress responsive genes, hence empowering the plant to cope and survive in stress conditions (Rasheed et al., 2022). Overwhelmingly, application of NPs has key role in plants systems to survive in drought and salinity and capable the plant to modulate its normal functionality, keep the plant and its environment healthy as well as maintain the plant yield (Figure 3). In-depth knowledge is still imperative to know the details functions of NPs to comprehend the stress tolerance in plants.
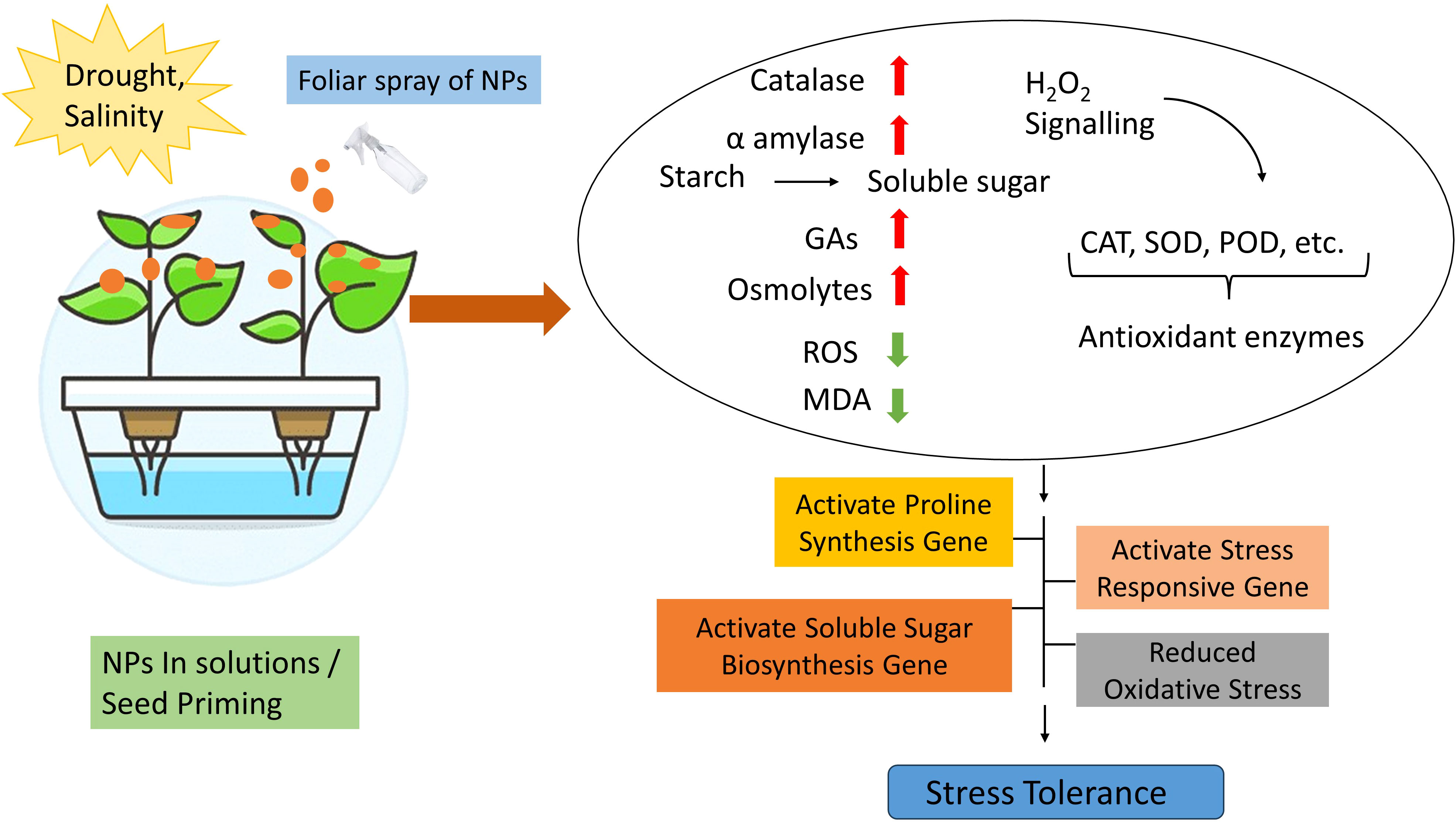
Figure 3 Molecular mechanism of NPs in drought and salinity stress tolerance.
Endocytosis offers a practical and effective method for internalizing extracellular substances like NPs. Endocytosis is a transmembrane process that occurs when the cell membrane is folded. The endocytosis of NPs can occur via clathrin-dependent or -independent routes. The number of parameters, such as size, charge, surface qualities, and other relevant parameters, substantially impact the uptake process of NPs (Azim et al., 2023). Metal-based NPs showed varied degrees of germination-unaffected seed penetration in wheat, maize, spinach, zucchini, rapeseed, and several desert plants (Chichiriccò and Poma, 2015). NPs may enter seeds and boost absorption, leading to better germination (Ahmadov et al., 2020). Multiwall carbon nanotubes (MWCNTs) were used to encourage the upregulation of tomato seedlings in terms of their seeds and root systems (Khodakovskaya et al., 2009). The beneficial influences of NPs on seed germination is also linked to their function in controlling aquaporins, tiny transmembrane water channels important for water permeability, seed germination, and plant development.
The research investigated the uptake and buildup of 8 nm ZnO nanoparticles in soybean seedlings across the 500-4000 mg range of exposure. At a dosage of 500 mg L-1, soybean seedlings showed a significantly higher uptake of Zn NPs in comparison to dosage 1000 mg L-1 or more. At lower dosage (500 mg L-1), NP aggregation seems to be less frequent. On the other hand, greater dosage (1000–4000 mg L-1) tend to lead to the formation of agglomerates (López-Moreno et al., 2010). The ability of gold NPs to translocate and concentrate within soybean plants after seed inoculation has been shown based on experimental findings (Maharramov et al., 2015). The thickness of seed coats makes it difficult for NPs to enter seeds when assessed to plant membranes and cell walls (Srinivasan and Saraswathi, 2010). Due to a mechanism known as enhanced water absorption, carbon nanotubes have demonstrated excellent penetration of the seed coat (Ganguly et al., 2014). It was discovered that nanotubes function as potential nano transporters, making it easier to carry DNA and tiny color molecules into whole plant cells (Savithramma et al., 2012).
According to studies, the rate at which NPs build up in plant roots may depend on environmental factors and NP characteristics (Ali et al., 2021a). It is believed that the small NPs can penetrate the roots of plants through capillary forces, osmotic pressure or by accurately passing through the epidermal cells of the roots (Ali et al., 2021a). Therefore, the semipermeable cell walls of the root epidermal cells have microscopic gaps that act as pores and prevent the passage of huge NPs. Furthermore, the cuticle performs as a key protective obstacle on leaves, successfully preventing NPs larger than 5 nm from penetrating the leaf. In general, determining the impacts of nanomaterials on the absorption, transport, and buildup of NPs within plants depends critically on their fundamental structure (Raliya et al., 2016). The term “apoplastic route” describes how holes in the root epidermal cell walls, generally between 5 and 20 nm in size, allow roots to absorb tiny NPs (Lin and Xing, 2007). In prior research, the content of Ag-NPs in both the shoot and roots of lettuce dramatically increased when silver sulphide (Ag2S) NPs were treated together with KCl and ammonium thiosulfate (Doolette et al., 2015). In another research, it was shown that organic matter in the soil reduced the amount of cerium dioxide (CeO2) NPs taken up by maize roots (Zhao et al., 2012). When applying NPs to soil or using them as a foliar application, findings have suggested that the mobility of NPs can be affected by bacteria associated with roots and leaves. Research has indicated that the use of NPs on soil or as a foliar application may affect the movement of NPs through the involvement of bacteria in the roots and leaves (Guo and Chi, 2014). The mucilage can also make the rhizosphere more acidic (Schaller et al., 2013), which encourages the disintegration of certain insoluble NPs (Schwab et al., 2016) and impacts on the extent to which plants absorb the NPs. Most research shows that the main restriction on entering NPs into plant cells is the size of the holes in the cell wall.
The stomatal holes offer a different route for NPs to enter plants when examining foliar absorption (Larue et al., 2014). NPs can be transported to various plant parts, including the roots, through leaf translocation. Several plant species, including rapeseed, wheat, beans, corn, lettuce, and cucumber, show evidence of NPs internalization through their leaves (Chichiriccò and Poma, 2015). Leaves can internalize NPs of various sizes, varying from a few nanometers to several hundred nanometers, and are made of diverse materials, such as ceria, titania, FeO, ZnO, and Ag (Chichiriccò and Poma, 2015). Airborne NPs attach to plant leaf surfaces before being absorbed and entering epidermal cells. When NPs reach the epidermal cells, they have the potential to travel through apoplastic or symplastic pathways to numerous plant organs (Ahmadov et al., 2020). NPs are frequently sprayed onto leaf surfaces in agricultural applications, where they accumulate and are then taken up by plants via stomata or cuticle on the leaf surface. The primary constituents of the leaf epidermis’ waxy cuticle are wax, cutin, and pectin. The waxy cuticle of plant leaves is a crucial protective layer, preventing water loss when growing and acting as a key obstacle to inhibiting NPs from penetrating the leaves (Pérez-De-Luque, 2017). There are two different channels, lipophilic and hydrophilic channels, found on the waxy cuticle’s surface. Diffusion and penetration of lipophilic NPs into leaves are made possible by lipophilic channels on the cuticle surface (Bussieres, 2014). Confocal fluorescence microscopy with excellent temporal and spatial resolution was utilized by Hu et al. to demonstrate the ability of 2 nm carbon dots to penetrate cotton leaves via the cuticular route, showing their remarkable potential as a plant penetration-enhancing agent (Hu et al., 2020). Recent research has shown that various elements, including the properties of the NPs themselves, the plant species involved, and the surrounding environmental conditions, impact the absorption of NPs in plants. The immersion behavior of NPs in plant leaves can be affected by numerous properties such as chemical makeup, surface charge, particle size, and surface alteration (Wang et al., 2023). The particle magnitude of NPs has become a key component in investigating their absorption in the blade due to the size restriction limit of NPs in the blade absorption route (Li et al., 2020). NPs mostly travelled through the stomatal channel in the epidermis of wheat leaves before accumulating inside the chloroplasts.
The entry points for NPs into the plant body are the roots and leaves, which can cause various biochemical, molecular, physiological and morphological changes in plants (Khan et al., 2019). The changes made have an important impact on the development of plants, which can differ based on the amount, method, and size of using the NPs. Plants’ physiological functions and overall health can be significantly impacted by the size, reactivity and chemical composition of NPs (Ahmed et al., 2021). According to the available research, various nanoparticle kinds may improve plant growth and development when subjected to salinity and drought stress (Ali et al., 2021a). Mechanism of NPs and their possible impacts under drought and salinity stress are depicted in Figure 4 and Table 1.
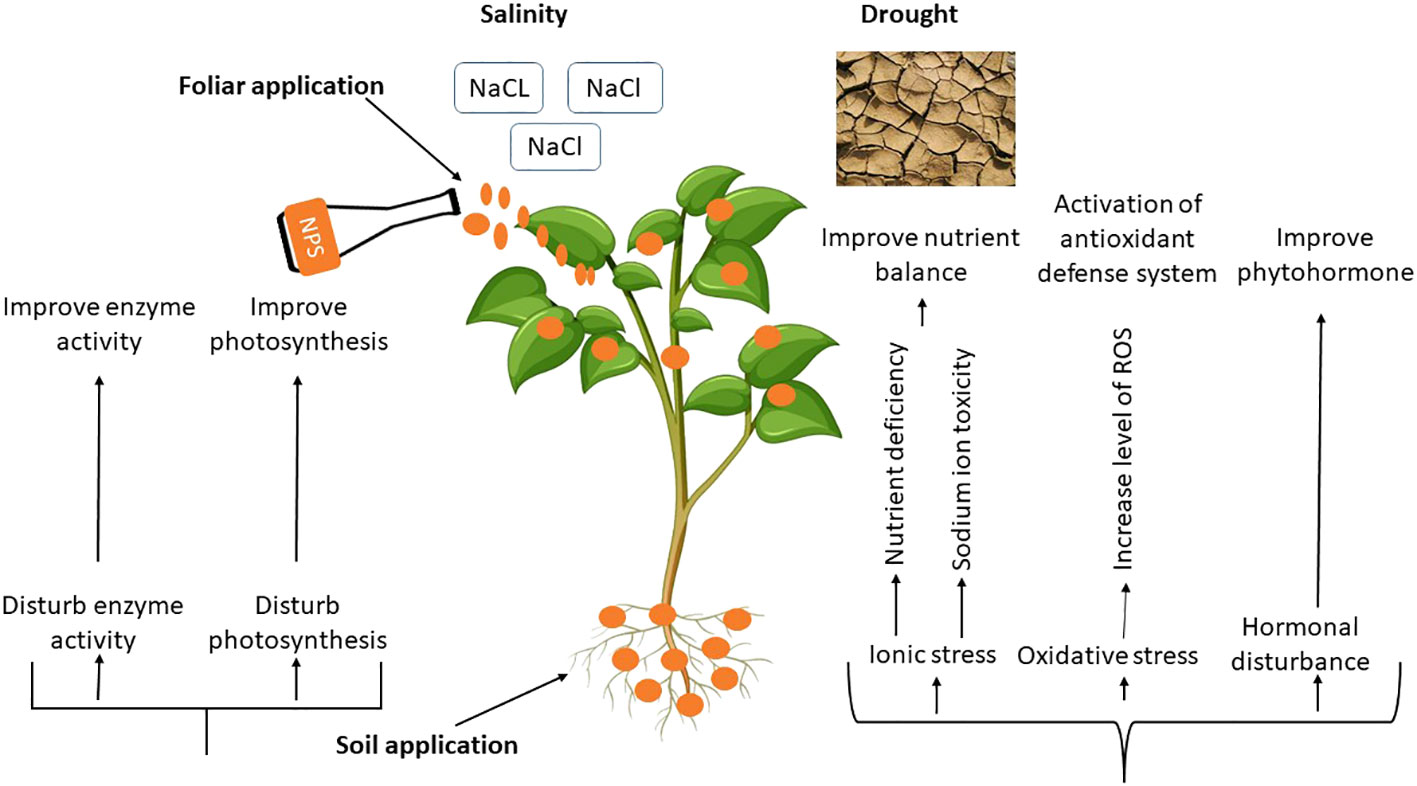
Figure 4 Physiological Mechanism of NPs induced drought and salinity tolerance in plants.
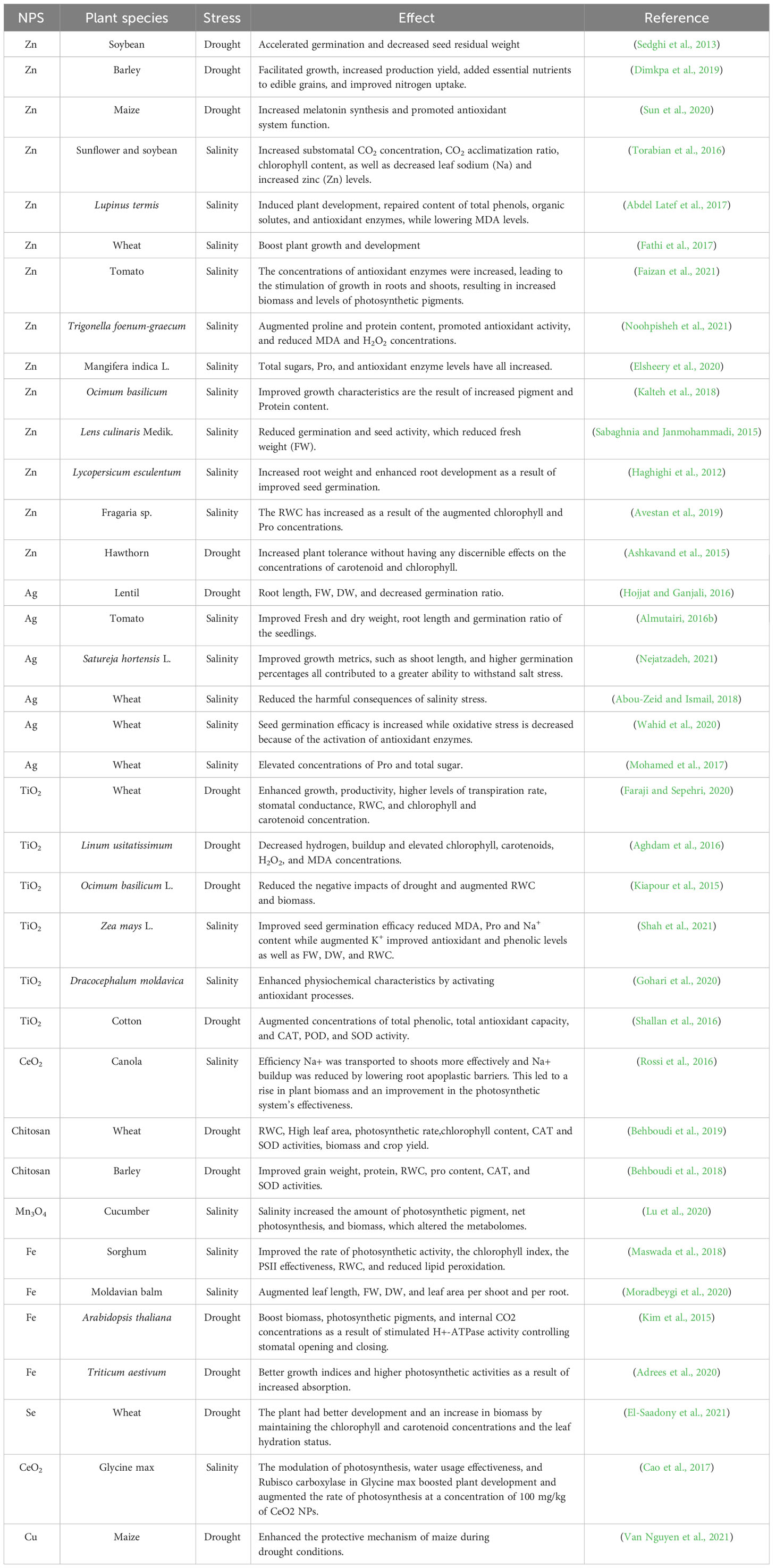
Table 1 Impact of Nanoparticles in enhancing crop tolerance under drought and salinity stress.
Plant development is negatively impacted by nutritional deficiencies and imbalances caused by drought stress, which disturbs the balance of nutrients inside plants (Umair Hassan et al., 2020). The nutrient absorption, transport, and distribution processes across different plant tissues are significantly improved by NPs, which also play a critical role in maintaining nutritional balance in plants (Kopittke et al., 2019). Plants with less water had lower N, K, Mn, and Zn levels due to reduced transpiration flux, nutrient uptake and compromised membrane stability (Semida et al., 2021). The absorption of N, phosphorous (P), K, and zinc was greatly enhanced by the treatment of NPs (ZnO) via foliar application and soil, and the negative impacts of drought stress were lessened (Akhtar et al., 2022). Maize plants that were cultivated under drought stress showed significant improvement in various parameters like photosynthesis, relative water content, antioxidant activity, and nutrient intake when they were co-treated with Si-NPs and plant growth-promoting rhizobacteria (Hafez et al., 2021). Adding zinc NPs (Zn-NPs) to wheat and sorghum plants increased productivity and improved nutrient absorption significantly (Dimkpa et al., 2019). NPs considerably increased nutrient intake, nitrate reductase activity, and nitrogen assimilation, enhancing protein and amino acid synthesis (Yuan et al., 2013). Using NPs boosts the appropriation of nutrients towards plant roots and improves plant nutrient absorption (Jaberzadeh et al., 2013).
Salinity stress is recognized to cause an imbalance in nutrient levels, resulting in excessive nutritional deficiencies in plants (Etesami and Maheshwari, 2018). It has been shown that Nano SiO2 increased the K+ amount in the leaves, which in turn improved the development of soybean seedlings under salt stress conditions (Farhangi-Abriz and Torabian, 2018). The exogenous spray of Cu-NPs was stated to reduce salinity stress in different research on tomato plants by promoting growth and preserving a balanced Na+/K+ ratio (Pérez-De-Luque, 2017). Abdoli et al. (2020) applying Fe2O3 NPs helped Trachyspermum ammi plants reduce salinity stress by rising the K+/Na+ ratio and Fe content. The research showed by Liu et al. (2021) augmentation of the K+-to-systolic sodium ratio in cotton plants was identified as one of the key processes underlying the improved plant growth observed upon the treatment of Ce NPs under salt stress. The results of this experiment show that the treatment of these NPs enhanced the amount of potassium in cells, which is a method to improve plant stress resistance. After using NPs, maintaining a balanced nutritional profile inside the plants is crucial to maximizing plant development under salt stress. For instance, research suggests that using Zn NPs can improve nutritional uptake of Zn. The transportation and intake of various nutrients, such as phosphorus, can be hampered when Zn absorption rises, causing an imbalance in the Zn-to-P ratio (Hussein and Abou-Baker, 2018).
Plant development is significantly reduced by drought stress, which has negative effects on cellular membranes and the interactions between plants and water (Umair Hassan et al., 2020). Drought stress results in the production of ROS, impacts lipid peroxidation damages cell membranes, and leads to a build-up of high levels of MDA (malondialdehyde) (Das and Roychoudhury, 2014). The foliar spray treatment of hydrogen peroxide (H2O2), ZnO NPs, and MDA accumulation was significantly decreased, and membrane stability was maintained, preventing the deficiency of vital osmolytes (El-Zohri et al., 2021). However, when NPs (ZnO) were added exogenously, they were critical in preserving membrane integrity and cell water status during drought stress, which led to better PS-II efficiency and metabolic activities (Semida et al., 2021). The treatment of NPs preserved cell integrity and membrane stability, which boosted water absorption by improving the anatomy of the plants (Hafez et al., 2020). The relative electrolyte leakages of the maize membrane were dramatically decreased by priming with TiO2 NPs. This demonstrated that the protective effect of TiO2 against membrane deterioration caused by salt stress (Shah et al., 2021). The foliar treatment with CsNPs and modified CsBMs improved defense-related genes, JA signaling, anthocyanins, membrane stability, and diterpene glycoside synthesis under salt stress (Balusamy et al., 2022). Hence, NPs improved membrane stability, chloroplast formation, and sugar accumulation, all of which contributed to the improvement overall.
It was discovered that the use of 50 mg/L Cu NPs caused a substantial elevation of the gene expression level of SOD rose six-fold (Mosa et al., 2018). In contrast, ZnO NPs reduced the SOD activity of Cicer arietinum (Burman et al., 2013). The increases in the levels of numerous antioxidant enzymes GR, (Catalase), APX, SOD, CAT and GPX within plants suggest that applying fullerene NPs (FNPs) to the leaves may have positive effects in lowering the oxidative stress induced by drought stress (Liu et al., 2016). According to Taran et al. (2017), ZnO NPs increased wheat antioxidant enzyme (SOD and CAT) activity, which improved drought resistance.
Additionally, it was showed that bulk zinc oxide NPs caused higher stress in horticultural crops (Rajput et al., 2021a). According to a different study, applying selenium NPs and copper NPs to S. lycopersicum demonstrated increased crop production, higher levels of vitamin C, glutathione, chlorophyll, and increased activity of antioxidant enzymes such GPX, SOD and PAL (Hernández-Hernández et al., 2019). The research revealed that the action of both POX and SOD enzymes were greater in plants treated with the substance’s nanoform than those preserved with the bulk form, except for CAT activity (Ghorbanpour et al., 2015).
Using nano-sized SiO2 and TiO2 together has triggered a similar protective mechanism that improves fertilizer and water consumption and increases nitrate reductase activity in soybean (Changmei et al., 2002). Numerous nitrogen metabolism-related enzymes, including glutamate dehydrogenase, NiR, GS, and GPT, are controlled by titanium dioxide NPs (TiO2 NPs) in different plant species. This control allows the transformation of inorganic nitrogen into organic nitrogen, which is then converted into proteins, chlorophyll, and amino acids, as well as the absorption of nitrate and increased plant biomass (Mishra et al., 2014). According to Shah et al. (2021), TiO2 NPs defend the chloroplast from intense light by boosting the activity of antioxidant enzymes such CAT, POD, and SOD.
It is well known that phytohormones play crucial roles in assisting plants in becoming acclimated to various environments through various processes. Studies frequently show that phytohormones improve a plant’s capacity to endure salt stress (Fahad et al., 2015). Some studies have suggested that plant’s improved stress resistance is responsible for the alteration in plant hormonal balance that NPs induce (Paramo et al., 2020). In particular research, the presence of silver NPs caused substantial changes in the concentrations of ethylene, gibberellin and abscisic acid (ABA) in rice plants (Manickavasagam et al., 2019). The treatment of Ag-NPs used as priming agent at 1 mg L-1 on wheat plants improved the amounts of α-napthaleneacetic acid (NAA), 6-benzyl aminopurine (BAP), and indole-3-butyric acid, while concurrently reducing the content of ABA. These modifications were considered a crucial mechanism by which silver NPs boost plant development in challenging environments (Abou-Zeid and Ismail, 2018). The sixth hormone in plants, brassinolide (BR), is important for increasing cell elongation and division and boosting resistance to salt, drought, and heat stressors (Hou et al., 2018).
NPs reduce oxidative stress by controlling the buildup of osmolytes and hormones by boosting the antioxidant machinery (Silva et al., 2022). The treatment of NPs increases plant performance under drought stress circumstances by up-regulating the production of proline and sugars, which in turn helps preserve the integrity of cellular membranes, proteins, and enzymes (Gohari et al., 2020). Likewise, the utilization of NPs, specifically TiO2, substantially increases the buildup of phenolic substances, proline, glycine betaine, soluble sugars, and total proteins and improves plant development under drought stress (Mustafa et al., 2021). According to research by (Khan et al., 2019), mesoporous silica NPs (MSNs) that responded to glutathione were used to successfully distribute abscisic acid (ABA) to plants. The AtGALK2 was upregulated due to the limited discharge of ABA from MSNs, which eventually improved Arabidopsis thaliana’s ability to withstand drought. The treatment of NPs promotes the development of plants under drought stress by improving the production of indole acetic acid and gibberellins (GA) (Li et al., 2021). Using Fe-NPs and salicylic acid, a crucial plant growth hormone, dramatically increased the yield of strawberries (Mozafari et al., 2018). Sun et al. (2020) claim that the increased melatonin production caused by NPs was responsible for the induction of greater drought tolerance, suggesting that NPs may be able to cope drought stress via modulating endogenous hormones. In conclusion, maintaining hormonal balance with the treatment of NPs can increase plant’s ability to withstand drought and salinity.
NPs can dramatically increase the accumulation of phenolic compounds in plants, whereas drought stress significantly reduces this accumulation. ZnO-NPs applied externally at 25 and 50 mg/L doses raised phenolic compound levels under drought stress, resulting in higher antioxidant activity and reduced MDA and H2O2 buildup (El-Zohri et al., 2021). The ZnO-NPs were investigated for their impact on the phenolic content of Stevia rebaudiana. The study exposed that the treatment of ZnO-NPs at concentrations of both 100 and 1000 mg/L resulted in significant reduction of the phenolic concentration (Javed et al., 2017). Additionally, plants treated with NPs have been shown to have a significant increase in non-enzymatic activities and total phenolic compounds, which lower lipid peroxidation and lessen oxidative damage (Ghani et al., 2022). The amounts of anthocyanin, phenolic, and antioxidant activity were increased after the treatment of Si- and Se-NPs (Zahedi et al., 2021). The growth and development of maize depend on the control of proteins and phenolic chemicals regulated by SiO2 NPs (Suriyaprabha et al., 2012).
The synthesis of phenolic compounds is known to rise in response to diverse abiotic stimuli because of its critical function in scavenging free radicals and antioxidants (Król et al., 2014). External application of TiO2 considerably increased the total phenolic content in Vigna radiata, which is consistent with earlier results that TiO2 seed priming efficiently controls phenolic compound synthesis in the maize hybrid under salt stress (Qi et al., 2013). The study carried out by Moradbeygi et al. (2020) aimed to investigate the effects of Fe NPs on Dracocephalum moldavica L. in salt stress circumstances. According to the experimental results, NPs enhanced plant development under salt stress by raising the abundance of flavonoid and phenolic chemicals, particularly in the roots, and lessening the activity of antioxidant enzymes. Additionally, compared to the control group, Cu NPs showed a considerable increase in glutathione, polyphenols, and vitamin C content under salt stress (Rajput et al., 2021a). The levels of total phenols, Osmo protectants, tannin, anthocyanins and flavonoids under salinity stress were also enhanced by the treatment of sulfur NPs to lettuce (Najafi et al., 2020). Additionally, AgNPs augmented the amounts of flavonoids and phenolic compounds while suppressing the leaf Na+/K+ ratio (Khan et al., 2020). For instance, foliar-applied Cu NPs in tomato plants reduced salinity stress by promoting growth and controlling the Na+/K+ ratio and growth (Pérez-Labrada et al., 2019). In contrast, CuNPs significantly amplified the amounts of phenols by 16%, vitamin C by 80%, glutathione by 81%, and phenols by 7.8%. Additionally, Helianthus annuus plants grown in a salty environment profited from the foliar treatment of Fe NPs because it improved the activities of polyphenol oxidase, CAT, and POD (Torabian et al., 2018).
Drought stress reduce the amount of chlorophyll produced, the effectiveness of PS-II, and the overall efficiency of photosynthesis in plants is adversely impacted (Semida et al., 2021). ZnO NPs enhanced chlorophyll synthesis, the activity of chlorophyll-synthesizing enzymes like chlorophyllase and fluorescence. As a result, photosynthetic efficiency increased under drought stress conditions (Abd El-Mageed et al., 2021). The external treatment of NPs aids in the stabilization of the ultrastructure of mitochondria and chloroplasts, which aids plants to sustain photosynthetic efficiency in drought stress (Rahmatpour et al., 2018). Titanium dioxide NPs are used to speed up the process of light-induced water hydrolysis, which releases oxygen, electrons, and protons (Silva et al., 2022). The efficiency of plants’ photosynthetic process subsequently improves significantly due to the faster entrance of protons and electrons into ETC (electron transport chain) (Alabdallah and Hasan, 2021). Titanium dioxide NPs foliar application increases the levels of photosynthetic pigments and boosts the gas exchange properties of plants by enhancing the activity of enzymes responsible for CO2 fixation and chlorophyll synthesis (Faraji and Sepehri, 2020). NPs further enhance the absorption of light within the chloroplasts, resulting in improved electron transport, enhanced efficiency of PS-II, increased O2 progression, and more efficient photo-phosphorylation, improved photosynthetic efficacy in plants under shortage of water (Shafea et al., 2017).
Photosynthesis is one of the critical activities that is strongly influenced by salinity stress, with its impacts varied based on elements including the plant species, the amount of salt, and other environmental conditions (Hnilickova et al., 2021). Several research has shown that adding NPs to plant leaves significantly increases the amount of chlorophyll present. According to studies, using manganese NPs (Mn-NPs) can help sustain a healthy rate of photosynthesis even when faced with severe abiotic stress conditions (Ye et al., 2020). In Vigna radiata plants under salt stress, manganese augmentation was found to improve several variables, including the membrane stability index, the amount of chlorophyll, and the activity of the enzyme nitrate reductase (Shahi and Srivastava, 2018). The exogenous treatment of Cu was observed to be valuable in reducing the negative effects of salinity on photosynthesis and water relations in maize plants (Iqbal et al., 2018). In Brassica, cerium NPs enhanced both biomass and photosynthetic efficiency when compared to plants that weren’t treated (Rossi et al., 2016). According to studies, Ag NPs to plants under salt stress can increase their chlorophyll levels and improve their fluorescence properties. Furthermore, Cao et al. (2017) showed that the incorporation of 100 mg/kg of CeO2 NPs in the growth medium accelerated photosynthesis and promoted plant development by controlling water usage efficiency, particularly in drought-stressed environments.
Insufficient water intake leads to the generation of ROS, H2O2, and MDA, which in turn triggers oxidative stress (Sutulienė et al., 2021). The remarkable ability of NPs to increase antioxidant activity helps to alleviate the negative consequences of water stress. The introduction of NPs (Si, ZnO, and Se) led to a substantial upsurge in the activity of APX, SOD, and CAT enzymes, which led to decreased oxidative harm triggered by shortage of water (Sun et al., 2020). The use of zinc oxide NPs has been found to improve the non-enzymatic actions of antioxidants like phenolic compounds and ascorbic acid (AsA). These compounds act in harmony with the antioxidant enzymes, including APX, CAT and SOD (El-Zohri et al., 2021). The foliar spray of SiO2 NPs at concentration 0, 12.5 ppm, 25 ppm, and 50 ppm considerably boosts non-enzymatic activities in plants under drought stress by increasing ferric reducing antioxidant power total phenolic content, 2,2-diphenyl-1-picrylhydrazyl scavenging activity, and total phenolic content, 2,2-diphenyl-1-picrylhydrazyl scavenging activity, ferric reducing antioxidant power (Sutulienė et al., 2021). Plants are protected from oxidative stress by NPs, which cause the buildup of antioxidant genes, osmolytes, minerals, and amino acids (Mittal et al., 2020).
Certain NPs exhibit characteristics comparable to some antioxidant enzymes, enabling them to help plants combat oxidative conditions. For instance, cesium, Mn, Cu, and Fe NPs exhibited POD-like capabilities, while cobalt, iron, and cesium NPs show CAT-like properties (Rico et al., 2015). Their investigation suggested that exposure to these NPs significantly improved the plant’s growth features. This improvement was associated with an upsurge in the activity of antioxidant enzymes, such as SOD, CAT and GPX as well as a reduction in the K+/Na+ ratio. Additionally, it was shown that using Ce-NPs augmented the activity of antioxidant enzymes in cotton plants, assisting in the elimination of ROS from within the cells. Additionally, the NPs supported plant development despite saline stress (Liu et al., 2021). A recent study examined the impact of applying iron nanoparticles (Fe-NPs) to Dracocephalum moldavica L. and found that it has the potential to enhance plant growth under salt stress conditions. The study showed that Fe-NPs increased the levels of flavonoids and phenolic compounds in the roots, which in turn contributed to improved plant development. In addition, it was shown that the NPs decreased the activity of antioxidant enzymes (Moradbeygi et al., 2020).
NPs treatment causes a considerable upregulation of genes that respond to drought, including GmRD20A, GmDREB2, GmMYB118, and GmMYB174 (Kandhol et al., 2022). According to Yang et al. (2018), applying ZnO-NPs and CuO-NPs considerably intensified the expression of genes associated with drought resilience of wheat plant roots. Additionally, Catharanthus roseus plants cultivated under drought stress circumstances improved their antioxidant capability and activation of genes linked to alkaloid production as a consequence of the administration of CS-NPs (Ali et al., 2021a). Additionally, using ceria-based NPs caused kidney beans to express more proteins linked with stress resistance while simultaneously downregulating the proteins in charge of nutrition storage and glucose metabolism (Majumdar et al., 2015). GmWRKY27, 118, and 174 was also showed increased expression by NPs, which promoted hormone signaling, the formation of seed germination, lignin, and secondary metabolites in response to shortage of water (Rushton et al., 2010). The enhanced GmWRKY27 gene expression in plants subjected to NPs suggests a link between NPs and the control of ABA production and stomatal function under drought stress circumstances (Linh et al., 2020). The utilization of NPs resulted in a widespread elevation of the expression of genes linked to the ability to withstand water stress. The higher concentrations of copper (Cu) and zinc (Zn) were connected with the increased expression of genes correlated with metal stress in the shoots of plants treated with ZnO or CuO NPs. This finding implies that plants exposed to CuO or ZnO NPs defended against various threats, including drought (Khan et al., 2019).
The foliar treatment of Zn-NPs at 0, 20 and 80 mgL−10 to the leaves of rapeseed plants under salt stress produced alterations in the expression of genes associated to stress response such as SKRD2, MYC, and MPK4 showed decreased expression, whereas ARP and MPK showed increased expression (Hezaveh et al., 2019). These genes influence a range of hormonal, developmental, and physiologic reactions. Notably, the transcription factor-related genes MYC and SKRD2 showed higher expression and supported improved tolerance to abiotic stress (Hezaveh et al., 2019). Silicon-NPs have positively impacted hemp under salt stress, stimulating improved development and causing molecular alterations in this plant species (Guerriero et al., 2021). Silicon NPs were found to affect genes associated with cytochrome b6f (Cytb6f), ATP-synthase complex and the light-harvesting complexes in tomato plants subjected to salt stress, as revealed by a proteomics study. The treatment of silicon NPS changed the expression of 29 genes, kinase/phosphatase genes comprising transcription factors, genes involved in photosynthetic processes, and genes associated with stress. The impact of silicon also extends to the control of genes involved in manufacturing nitric oxide and auxin (Tripathi et al., 2021a). Furthermore, silicon NPs reduce the impacts of salt stress by varying the expression of genes like OsNCED, and OsZEP, which are essential for manufacturing the hormone ABA. These genes include OsZEP, OsNCED, and OsZEP (Tripathi et al., 2021b). Under salinity stress, silicon NPs have been discovered to promote salt transport into the vacuole by upregulating the protein OsHMA3, which improves plant development. Furthermore, silicon shields plants from stress by boosting activity of antioxidant enzymes (Siddiqui et al., 2020). The downregulation of salt stress genes (DDF2, MAPK3 and RBOH1) upon exposure to S NPs led to an improvement in the salt resilience of tomato plants (Almutairi, 2016a).
The cutting-edge development in plant science, known as plant genetic engineering, is a key instrument to increase crop quality and production, boosting secondary metabolite levels in medicinal plants, and growing sustainable crops. Commonly, Agrobacterium and gene gun methods are used to transfer the gene of interest, although plant cell walls resist entering nucleus. Agrobacterium exhibited host specificity while gene gun method has potential to damage the plant tissues. In recent days, carbon nanotubes, magneto faction, DNA nanostructure, clay nanosheets, peptide NPs, CRISPER Cas9 and de novo transgenic plant production have accelerated progress in crop improvement (Lv et al., 2020). The usage of magnetic NPs has primarily been seen in medical research and animal science (Nasiri et al., 2020). However, research also exhibited the effective application of magnetoreception in plants (Zhao et al., 2017) established a quick and simple process utilizing magnetic NPs to produce transgenic seeds without needing tissue culture regeneration.
Additionally, SWCNTs exhibit effective membrane penetration via organelle walls (Hendler-Neumark and Bisker, 2019). The genetic transformation of chloroplasts and mitochondria efficiently gets around the restriction of gene migration, subsequent in increased herbicide tolerance in weeds because of their maternally transmitted nature (Lv et al., 2020). Kwak et al. (2019) developed SWCNTs complexed with chitosan specifically designed for the targeted delivery of DNA to the chloroplasts of numerous plant species, including spinach tobacco, watercress, and arugula. Similarly, Zhang et al. (2019) presented DNA NPs as a ground-breaking stage for siRNA delivery in plants. The researchers used the intensity of a fluorophore to gauge the internalization of these nanostructures into plant cells. This fluorophore was joined to the DNA strands at the connection sites of the DNA nanostructures through hybridization, enabling detection and analysis (Lv et al., 2020). NPs have great significance as materials for delivering biomolecules into cells due to their ability to cross biological membranes, protect and release a variety of payloads, and achieve varied targeting through chemical and physical changes. The first demonstration of the co-delivery of DNA and chemicals to Nicotiana tabacum plants in 2007 was done by Torney et al. employing biolistic delivery of gold-capped mesoporous silica NPs (MSNs) that vary in size from 100 to 200 nm (Torney et al., 2007). Mesoporous silica NPs (MSN), which were capped with gold NPs by covalent bonding, had a chemical expression inducer inserted into their pores (3 nm) (Cunningham et al., 2018). Additionally, recent studies have demonstrated the use of carbon nanotubes (CNTs) for the direct transport of plasmid DNA and siRNA into a variety of plant species, including model and non-model plants (Demirer et al., 2019). NPs have begun to play a crucial role in enabling and enhancing genome editing procedures by permitting effective and accurate transport of plasmids, RNA, and ribonucleoproteins (RNPs).
Genome editing is gaining popularity for precise modifications of specific sequences and studying biological processes and plant genetics. CRISPER Cas, ZFNs and TALENs are three essential components used to insert specific genetic mutations into plants in crop improvement (Doman et al., 2020). In contrast, CRISPER Cas technology is a faster and easier genome editing method which suggests various horizons for crop improvement against abiotic stresses (Gao et al., 2020). Sg-RNA initiate Cas9 nuclease to form complex of Cas9-sgRNA at targeted genomic DNA and spark the cleavage. Hence leads to the development of double stranded breaks with the help Cas9 nuclease in DNA of plant (Shan et al., 2013). The successful stories of CRISPER Cas9 technology were revealed by three research groups on rice, wheat, tobacco and Arabidopsis (Nekrasov et al., 2013). Afterwords, this technology immensely used in various project and objectives like powdery mildews in wheat to regulate TaMLO homologs (Wang et al., 2014). Similarly in maize, yield was improved by editing waxy alleles by CRISPERCas9 successfully (Gao et al., 2020). The utilization of CRISPR-Cas system has opened new avenues for crop breeding, genetics, and genomics. Its precise and efficient genome editing capabilities have enabled researchers to make targeted modifications in the genome of crops, leading to the development of improved crop varieties with desirable traits. This technology has revolutionized the field of crop improvement and holds great promise for the future of agriculture. This technology has the potential to improve specific traits in plants while minimizing the risk of unintended effects, making it useful in addressing both biotic and abiotic stress factors.
ROS are generated due to interactions of NPs with plants through numerous physical and chemical processes (Figure 5). Increased ROS generation carried by NPs can negatively impact plant cells, severely limiting plant growth and development (Xie et al., 2019). The shape, size, and characteristics of the NPs regulate their hazardous effects. Metal oxide NPs like Fe3O4, ZnO, and TiO2, which have applications in a variety of sectors, as well as metal NPs like gold, platinum, silver, and iron, have the potential to endanger human health. These NPs can harm proteins, DNA, and cell membranes when they come into contact with cells, significantly reducing plant development (Hsin et al., 2008).
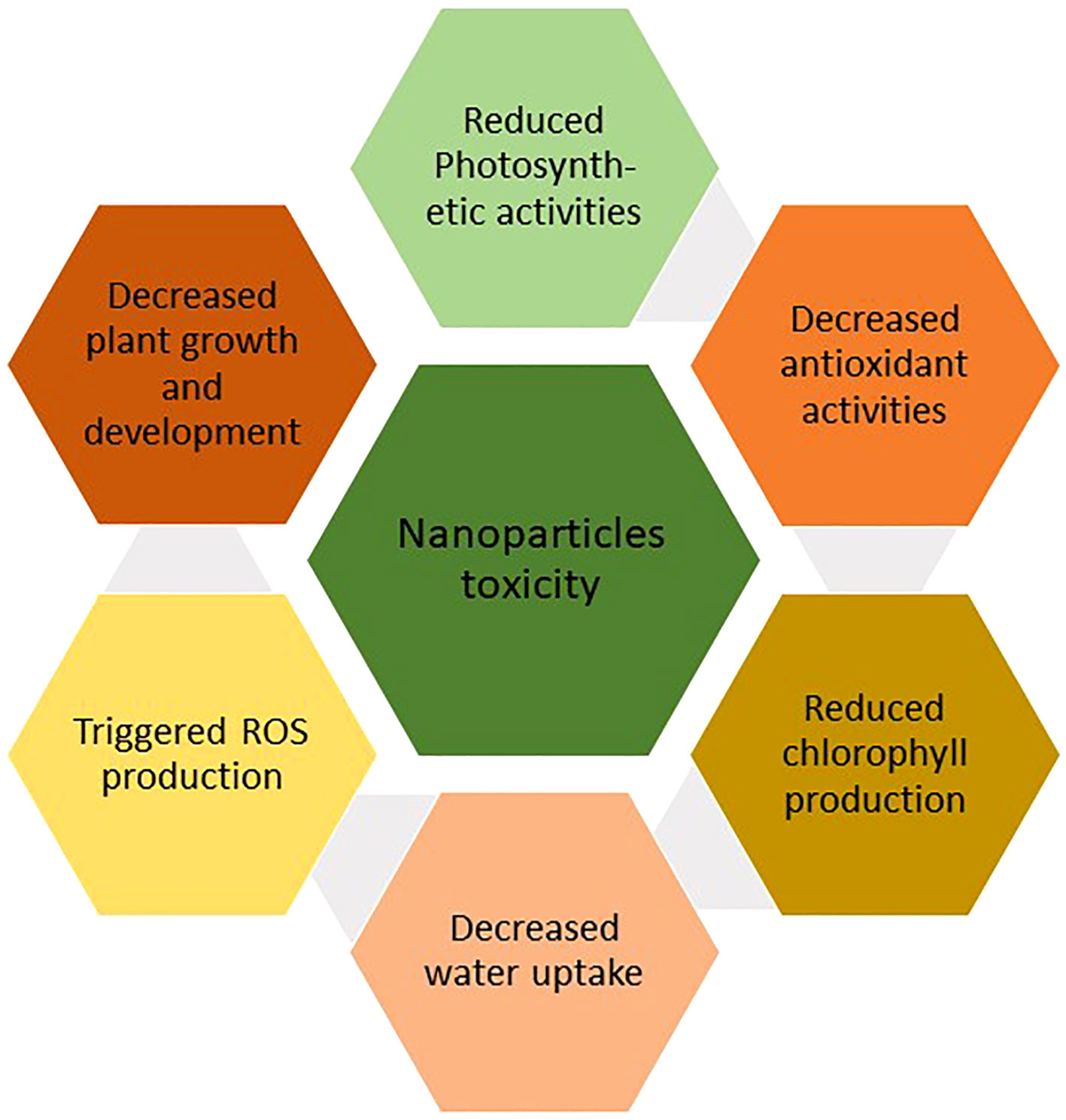
Figure 5 Impact of NPs toxicity on plant growth.
Soil is a significant source of NPs, and plants use active transport systems in their roots to take up both NPs and nutrients from the soil (Khan et al., 2021). NPs are uptake by the roots and pass through the root cortex and epidermis of cell walls before reaching the upper parts of the plant and causing adverse effects inside the plant (Rajput et al., 2018). NPs might negatively affect plant development at higher concentrations by diminishing antioxidant activities, decreasing photosynthetic effectiveness, and decreasing chlorophyll production. NPs have the benefit of being substantially smaller than normal bulky materials, enabling them to be immersed into biological systems near 15-20 times more quickly (Khan et al., 2021). Research suggests that the toxicity of NPs in various microorganisms such as plants and algae is linked to physical damage and the production of ROS, leading to oxidative stress (Hou et al., 2018). The colour structure of photosynthesis, the effectiveness of PS-II (photosystem II), and the development of aquatic plants like amphibian plants can all be impacted by NPs in aquatic environments (Jacobasch et al., 2014).
The agriculture industry cannot fully embrace and implement nanotechnologies due to increasing concerns about the bioavailability and toxicity of NPs and the shortcomings of the current regulatory framework (Ali et al., 2021a). According to Rao and Shekhawat (2016) improper use of NPs can negatively impact plant development, such as reducing the production of protein and pigment in plants. Plant cells also develop extra substances like metallothioneins and phytochelatins due to the stress produced by NPs. These substances defend against the damaging impacts of oxidative stress on plant cells (Dev et al., 2018). NPs in the soil environment go through a sequence of bio/geo transformations that ultimately regulate their toxicity and bioavailability (Ali et al., 2021a). NPs of various compositions adversely influence the roots and shoot elongation of seedlings, which is mostly associated with the uptake of NPs into the roots. Silver NPs (AgNPs) have cytotoxic effects on some plants and reduce germination, transpiration, shoot and root length, and involved changes in gene expression, oxidative stress and cell death (Thuesombat et al., 2014). The concentration of the NPs influences the extent that growth is inhibited in mung bean and sorghum (Lee et al., 2012). Inhibitory and toxic effects of NPs are listed in Table 2.
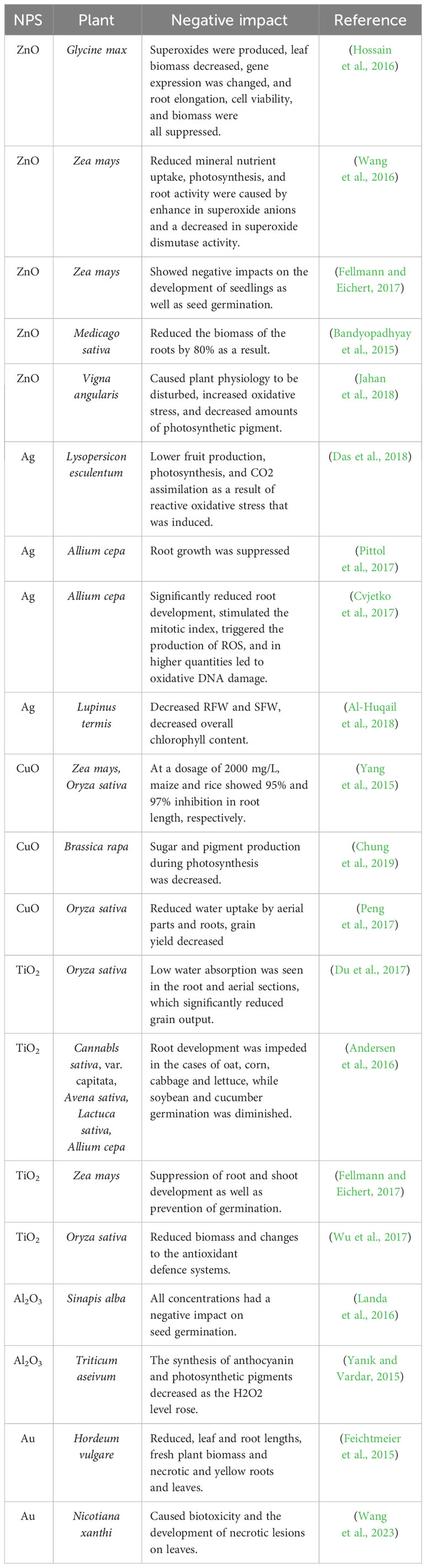
Table 2 Toxic and inhibitory effects of nanoparticles on plants.
The increasing antioxidant enzyme activity is an indirect indication of increased cellular ROS levels. The plants containing NPs can protect biological components from oxidative stress by using phenols and phenolic acids (Yasur and Rani, 2013). Wang et al. (2011) investigated the effects of Fe3O4 NPs on cushaw pumpkin and ryegrass plants. They investigated how these NPs affected the production of ROS, which affected membrane stability. According to other research, releasing copper ions or copper oxide NPs might cause oxidative stress (Nair and Chung, 2015).
Nevertheless, the solubilization of copper oxide NPs within plant cells leads to the subsequent release of copper ions. Consequently, a redox process involving both Cu2+ and Cu+ ions leads to oxidative damage (Shi et al., 2011). Lipid peroxidation was shown to be primarily caused by ROS, which is produced by the Fenton reaction due to the existence of polyvalent CuO NPs (Fubini et al., 2007). Panda et al. (2011) supervised a study on Allium cepa and examined the formation of certain ROS, including superoxide radicals (O2%-) and H2O2. They discovered that the presence of silver NPs caused these reactive species levels to rise, which caused an oxidative burst in the plant. According to Atha et al. (2012), copper oxide NPs were discovered to cause DNA harm and prevent growth in radishes, and annual ryegrass. The association between titanium oxide NPs and DNA and the underlying mechanism causing DNA damage in onion and tobacco have been shown through the use of atomic force microscope imaging (Ghosh et al., 2010). Mukherjee et al. (2014) examined the membrane damage produced by zinc oxide NPs in Pisum sativum and observed a comparatively higher level of lipid peroxidation in plants exposed to NPs evaluated to the control (non-treated) plants. The study also reported that Pisum sativum plants treated to zinc oxide NPs produced an excessive amount of hydrogen peroxide (H2O2).
Dimkpa et al. (2012) carried out research to evaluate the promising impacts of CuO and ZnO NPs. The study showed a rise in lipid peroxidation levels and reduced chlorophyll content, indicators of oxidative stress. Compared to the corresponding control groups, the incidence of copper oxide NPs in wheat induced a nearly four-fold increase in lipid peroxidation, whereas the existence of ZnO NPs resulted in a two-fold increase. It was shown that cerium oxide NPs had an adverse effect on asparagus lettuce, causing membrane degradation, lipid peroxidation, cell membrane impairment, and inhibiting root elongation (Cui et al., 2014). Marusenko et al. (2013) they stated a decrease in chlorophyll content in A. thaliana plants when subjected to Fe NPs. Faisal et al. (2013) examined that NiO nanoparticle treatment of tomato plants caused oxidative stress to be induced in their roots. Additionally, flow cytometry analysis found that the protoplasts had a greater level of oxidative stress production. Higher levels of enzymes like CAT and SOD were associated with increased reactive stress. Shaw and Hossain (2013) demonstrated that copper oxide NPs affected both root and shoot development as well as photosynthetic system in Hordeum vulgare.
NPs are being used at an exponential rate; however, it’s important to remember that there may be harmful and toxicological impacts on environment and human health. NPs have the potential to be released into the environment through several processes, including manufacture, use, recycling, or disposal. These NPs may linger in biological systems, soil, water, or the air (Roy et al., 2014). NPs can enter the body of a person or an animal by skin contact, oral ingestion, or inhalation through the respiratory system, then spread to other bodily compartments. The activation of pro-inflammatory cytokines and chemokines upon exposure to NPs was discovered to result in the recruitment of inflammatory cells, which affects the immune system’s homeostasis and can cause autoimmune, allergy, or malignant illnesses (Roy et al., 2014).
Additionally, breathing in or exposure to ultrafine particles has been linked to several respiratory, cardiovascular, and central nervous system problems (Joudeh and Linke, 2022). Similarly, NPs can penetrate plant cells and have negative effects (Stark, 2011). For example, zinc oxide (ZnO) and aluminum (Al) NPs have been discovered to hinder the development of plant roots (Lin and Xing, 2007). Numerous NPs have been studied for their potential to cause immunological and cellular damage. The effect of NPs on human health has been examined using in vitro toxicity assessment models, such as cell cultures that include cancer cell lines. However, a substantial body of research on the impacts of nanomaterials on human health and the environment reveals that metal NPs can cause cytotoxic effects, which are determined by their charge (Pande and Arora, 2019). NPs can increase their toxicity at the nanoscale level by interacting electrostatically with biological membranes and various cellular metabolites in the cytoplasm (Prasad et al., 2017). According to research, when exposed to light and molecular oxygen, certain photochemically active NPs, including fullerenes, ZnO, SiO2, and TiO2, can directly transfer electrons to produce reactive oxygen species such as superoxide radicals. The result might be oxidative stress (Nayan et al., 2016).
Currently, the use of NPs is constantly growing and becoming indispensable in many industries, including agriculture. For sustainable agriculture, we need ecologically acceptable solutions to crop yield decline caused by drought and salt stress. Intensive research is underway to explore the possible role of various NPs in mitigating damage caused by drought and salinity stress to enhance plant development and crop yield. Due to their tiny size, NPs may easily enter plant tissues and have a favorable effect on morphology, physiology, and biochemistry of plant, promoting plant growth and increasing agricultural output, especially under drought and salinity stress situations. Moreover, the application of NPs greatly increases plant functioning and offers tremendous resilience to plants, enabling them to tolerate drought and salt stress efficiently. The application of NPs enhances membrane integrity, nutrient absorption, and defends the plant’s photosynthetic system from injury produced by salt and drought stress, subsequent augmenting plant development in these challenging circumstances. The utilization of NPs also enhances the production of phenolic compounds and hormones that protect plants from stress. The expression of antioxidant and stress-responsive genes is also increased by NPs, which considerably strengthens the defense mechanism against salinity and drought stress. The function of NPs in aiding numerous processes to promote resilience to drought and salt stress has recently been the subject of substantial investigation. The use of NPs in crop development and sustainable agriculture is still nascent, and the existing research in this field lacks consistency and is insufficient. The impact of NPs on seed germination remains unexplored, highlighting the need to investigate their role in germination mechanisms, encompassing water uptake, radical protrusion, and activation of enzymes responsible for food mobilization. Furthermore, it is critical to investigate how NPs affect their metabolic functions because gibberellins and abscisic acids are essential for seed germination. NPs enhance nutrient absorption in plants under drought and salinity stress, but further research is required to fully understand their function in nutrient channels and ionic transporters in plants under such stresses. The treatment of NPs substantially protects the photosynthetic apparatus, but further research is needed to determine how they affect stomata motions, anion channel control, and intercellular signaling in guard cells of plants under salinity and drought stress. The investigations on the effects of NPs on proteome may be advantageous to understand better the numerous processes by which NPs promote drought and salinity tolerance. The impacts of NPs on genetic and proteomic parameters have not received enough attention, underlining the need to investigate these topics in future studies. The synergistic potential of combining microorganisms with NPs to promote drought and salinity tolerance also constitutes an interesting research direction. Additionally, understanding the interactions between NPs and plants would help us understand how plants cope with drought and salt. The timing and content of NPs under diverse climatic circumstances must be optimized to meet the unique needs of distinct crops. While the bulk of research focuses on the impacts of NPs on plants under salt and drought stress, it is critical to examine the influence of NPs on plants exposed to various stressors at altered times throughout their life cycle.
AR: Data curation, Writing – original draft. SK: Writing – original draft. FS: Data curation, Writing – review & editing. ZPe: Conceptualization, Data curation, Visualization, Writing – review & editing. KF: Writing – review & editing. NW: Writing – review & editing. YJ: Conceptualization, Writing – review & editing. ZPa: Conceptualization, Writing – review & editing. SH: Data curation, Writing – review & editing. LW: Data curation, Writing – review & editing. AQ: Conceptualization, Data curation, Writing – review & editing. XD: Conceptualization, Supervision, Writing – review & editing. HL: Supervision, Writing – review & editing.
The author(s) declare financial support was received for the research, authorship, and/or publication of this article. Grants from the National Key Research and Development Program of China (grant no: 2022YFD1200300), China Agriculture Research System (CARS-15-01) and National Natural Science Foundation of China (32060466) provided support for this work.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The author(s) declared that they were an editorial board member of Frontiers, at the time of submission. This had no impact on the peer review process and the final decision.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Abdel Latef, A., Abu Alhmad, M. F., Abdelfattah, K. E. (2017). The possible roles of priming with ZnO nanoparticles in mitigation of salinity stress in lupine (Lupinus termis) plants. J. Plant Growth Regul. 36, 60–70. doi: 10.1007/s00344-016-9618-x
Abd El-Mageed, T. A., Shaaban, A., Abd El-Mageed, S. A., Semida, W. M., Rady, M. O. (2021). Silicon defensive role in maize (Zea mays L.) against drought stress and metals-contaminated irrigation water. Silicon 13, 2165–2176. doi: 10.1007/s12633-020-00690-0
Abdoli, S., Ghassemi-Golezani, K., Alizadeh-Salteh, S. (2020). Responses of ajowan (Trachyspermum ammi L.) to exogenous salicylic acid and iron oxide nanoparticles under salt stress. Environ. Sci. pollut. Res. 27, 36939–36953. doi: 10.1007/s11356-020-09453-1
Abou-Zeid, H., Ismail, G. (2018). The role of priming with biosynthesized silver nanoparticles in the response of Triticum aestivum L to salt stress. Egyptian J. Bot. 58, 73–85. doi: 10.21608/EJBO.2017.1873.1128
Acosta-Motos, J. R., Ortuño, M. F., Bernal-Vicente, A., Diaz-Vivancos, P., Sanchez-Blanco, M. J., Hernandez, J. A. (2017). Plant responses to salt stress: adaptive mechanisms. Agronomy 7, 18. doi: 10.3390/agronomy7010018
Adrees, M., Khan, Z. S., Ali, S., Hafeez, M., Khalid, S., Ur Rehman, M. Z., et al. (2020). Simultaneous mitigation of cadmium and drought stress in wheat by soil application of iron nanoparticles. Chemosphere 238, 124681. doi: 10.1016/j.chemosphere.2019.124681
Aghdam, M. T. B., Mohammadi, H., Ghorbanpour, M. (2016). Effects of nanoparticulate anatase titanium dioxide on physiological and biochemical performance of Linum usitatissimum (Linaceae) under well-watered and drought stress conditions. Braz. J. Bot. 39, 139–146. doi: 10.1007/s40415-015-0227-x
Ahmadov, I. S., Ramazanov, M. A., Gasimov, E. K., Rzayev, F. H., Veliyeva, S. B. (2020). The migration study of nanoparticles from soil to the leaves of plants. Biointerface Res. Appl. Chem. 10, 6101–6111. doi: 10.33263/BRIAC105.61016111
Ahmed, T., Noman, M., Manzoor, N., Shahid, M., Abdullah, M., Ali, L., et al. (2021). Nanoparticle-based amelioration of drought stress and cadmium toxicity in rice via triggering the stress responsive genetic mechanisms and nutrient acquisition. Ecotoxicology Environ. Saf. 209, 111829. doi: 10.1016/j.ecoenv.2020.111829
Akhtar, N., Ilyas, N., Meraj, T. A., Pour-Aboughadareh, A., Sayyed, R., Mashwani, Z.-U.-R., et al. (2022). Improvement of plant responses by nanobiofertilizer: a step towards sustainable agriculture. Nanomaterials 12, 965. doi: 10.3390/nano12060965
Alabdallah, N. M., Hasan, M. M. (2021). Plant-based green synthesis of silver nanoparticles and its effective role in abiotic stress tolerance in crop plants. Saudi J. Biol. Sci. 28, 5631–5639. doi: 10.1016/j.sjbs.2021.05.081
Alallam, B., Doolaanea, A. A., Kyaw Oo, M., Mohd Nasir, M. H., Taher, M. (2021). Influence of nanoparticles surface coating on physicochemical properties for CRISPR gene delivery. J. Drug Delivery Sci. Technol. 66, 102910. doi: 10.1016/j.jddst.2021.102910
Al-Huqail, A. A., Hatata, M. M., Al-Huqail, A. A., Ibrahim, M. M. (2018). Preparation, characterization of silver phyto nanoparticles and their impact on growth potential of Lupinus termis L. seedlings. Saudi J. Biol. Sci. 25, 313–319. doi: 10.1016/j.sjbs.2017.08.013
Ali, E., El-Shehawi, A. M., Ibrahim, O., Abdul-Hafeez, E., Moussa, M., Hassan, F. (2021a). A vital role of chitosan nanoparticles in improvisation the drought stress tolerance in Catharanthus roseus (L.) through biochemical and gene expression modulation. Plant Physiol. Biochem. 161, 166–175. doi: 10.1016/j.plaphy.2021.02.008
Ali, S., Mehmood, A., Khan, N. (2021b). Uptake, translocation, and consequences of nanomaterials on plant growth and stress adaptation. J. Nanomaterials 2021, 6677616. doi: 10.1155/2021/6677616
Almutairi, Z. M. (2016a). Effect of nano-silicon application on the expression of salt tolerance genes in germinating tomato ('Solanum lycopersicum'L.) seedlings under salt stress. Plant Omics 9, 106–114. doi: 10.17957/IJAB/15.0114
Almutairi, Z. M. (2016b). Influence of silver nano-particles on the salt resistance of tomato (Solanum lycopersicum) during germination. Int. J. Agric. Biol. 18, 449–457. doi: 10.17957/IJAB/15.0114
Andersen, C. P., King, G., Plocher, M., Storm, M., Pokhrel, L. R., Johnson, M. G., et al. (2016). Germination and early plant development of ten plant species exposed to titanium dioxide and cerium oxide nanoparticles. Environ. Toxicol. Chem. 35, 2223–2229. doi: 10.1002/etc.3374
Ashkavand, P., Tabari, M., Zarafshar, M., Tomásková, I., Struve, D. (2015). Effect of SiO2 nanoparticles on drought resistance in hawthorn seedlings. Leśne Prace Badawcze 76, 350–359. doi: 10.1515/frp-2015-0034
Atha, D. H., Wang, H., Petersen, E. J., Cleveland, D., Holbrook, R. D., Jaruga, P., et al. (2012). Copper oxide nanoparticle mediated DNA damage in terrestrial plant models. Environ. Sci. Technol. 46, 1819–1827. doi: 10.1021/es202660k
Avestan, S., Ghasemnezhad, M., Esfahani, M., Byrt, C. S. (2019). Application of nano-silicon dioxide improves salt stress tolerance in strawberry plants. Agronomy 9, 246. doi: 10.3390/agronomy9050246
Azim, Z., Singh, N., Singh, A., Amist, N., Niharika, Khare, S., et al. (2023). A review summarizing uptake, translocation and accumulation of nanoparticles within the plants: current status and future prospectus. J. Plant Biochem. Biotechnol. 32, 211–224. doi: 10.1007/s13562-022-00800-6
Baalousha, M., Yang, Y., Vance, M. E., Colman, B. P., Mcneal, S., Xu, J., et al. (2016). Outdoor urban nanomaterials: the emergence of a new, integrated, and critical field of study. Sci. Total Environ. 557, 740–753. doi: 10.1016/j.scitotenv.2016.03.132
Badawy, A. M. E., Luxton, T. P., Silva, R. G., Scheckel, K. G., Suidan, M. T., Tolaymat, T. M. (2010). Impact of environmental conditions (pH, ionic strength, and electrolyte type) on the surface charge and aggregation of silver nanoparticles suspensions. Environ. Sci. Technol. 44, 1260–1266. doi: 10.1021/es902240k
Balusamy, S. R., Rahimi, S., Sukweenadhi, J., Sunderraj, S., Shanmugam, R., Thangavelu, L., et al. (2022). Chitosan, chitosan nanoparticles and modified chitosan biomaterials, a potential tool to combat salinity stress in plants. Carbohydr. Polymers 284, 119189. doi: 10.1016/j.carbpol.2022.119189
Bandyopadhyay, S., Plascencia-Villa, G., Mukherjee, A., Rico, C. M., José-Yacamán, M., Peralta-Videa, J. R., et al. (2015). Comparative phytotoxicity of ZnO NPs, bulk ZnO, and ionic zinc onto the alfalfa plants symbiotically associated with Sinorhizobium meliloti in soil. Sci. Total Environ. 515, 60–69. doi: 10.1016/j.scitotenv.2015.02.014
Behboudi, F., Tahmasebi Sarvestani, Z., Kassaee, M. Z., Modares Sanavi, S., Sorooshzadeh, A., Ahmadi, S. B. (2018). Evaluation of chitosan nanoparticles effects on yield and yield components of barley (Hordeum vulgare L.) under late season drought stress. J. Water Environ. Nanotechnology 3, 22–39. doi: 10.22090/jwent.2018.01.003
Behboudi, F., Tahmasebi-Sarvestani, Z., Kassaee, M. Z., Modarres-Sanavy, S., Sorooshzadeh, A., Mokhtassi-Bidgoli, A. (2019). Evaluation of chitosan nanoparticles effects with two application methods on wheat under drought stress. J. Plant Nutr. 42, 1439–1451. doi: 10.1080/01904167.2019.1617308
Biba, R., Košpić, K., Komazec, B., Markulin, D., Cvjetko, P., Pavoković, D., et al. (2022). Surface coating-modulated phytotoxic responses of silver nanoparticles in plants and freshwater green algae. Nanomaterials 12, 24. doi: 10.3390/nano12010024
Burman, U., Kumar, P. (2018). “Plant response to engineered nanoparticles,” in Nanomaterials In Plants, Algae, And Microorganisms (London: Elsevier), 103–118.
Burman, U., Saini, M., Kumar, P. (2013). Effect of zinc oxide nanoparticles on growth and antioxidant system of chickpea seedlings. Toxicological Environ. Chem. 95, 605–612. doi: 10.1080/02772248.2013.803796
Bussieres, P. (2014). Estimating the number and size of phloem sieve plate pores using longitudinal views and geometric reconstruction. Sci. Rep. 4, 4929. doi: 10.1038/srep04929
Calicioglu, O., Flammini, A., Bracco, S., Bellù, L., Sims, R. (2019). The future challenges of food and agriculture: An integrated analysis of trends and solutions. Sustainability 11, 222. doi: 10.3390/su11010222
Cao, Z., Stowers, C., Rossi, L., Zhang, W., Lombardini, L., Ma, X. (2017). Physiological effects of cerium oxide nanoparticles on the photosynthesis and water use efficiency of soybean (Glycine max (L.) Merr.). Environ. Science: Nano 4, 1086–1094. doi: 10.1039/C7EN00015D
Cataldo, F., Da Ros, T. (2008). Medicinal Chemistry And Pharmacological Potential Of Fullerenes And Carbon Nanotubes (Germany: Springer Science & Business Media).
Changmei, L., Chaoying, Z., Junqiang, W., Guorong, W., Mingxuan, T. (2002). Research of the effect of nanometer materials on germination and growth enhancement of Glycine max and its mechanism. Soybean Sci. 21, 168–171.
Chichiriccò, G., Poma, A. (2015). Penetration and toxicity of nanomaterials in higher plants. Nanomaterials 5, 851–873. doi: 10.3390/nano5020851
Chung, C., Kim, Y.-K., Shin, D., Ryoo, S.-R., Hong, B. H., Min, D.-H. (2013). Biomedical applications of graphene and graphene oxide. Accounts Chem. Res. 46, 2211–2224. doi: 10.1021/ar300159f
Chung, I.-M., Rekha, K., Venkidasamy, B., Thiruvengadam, M. (2019). Effect of copper oxide nanoparticles on the physiology, bioactive molecules, and transcriptional changes in Brassica rapa ssp. rapa seedlings. Water Air Soil pollut. 230, 1–14. doi: 10.1007/s11270-019-4084-2
Cui, D., Zhang, P., Ma, Y., He, X., Li, Y., Zhang, J., et al. (2014). Effect of cerium oxide nanoparticles on asparagus lettuce cultured in an agar medium. Environ. Science: Nano 1, 459–465. doi: 10.1039/C4EN00025K
Cunningham, F. J., Goh, N. S., Demirer, G. S., Matos, J. L., Landry, M. P. (2018). Nanoparticle-mediated delivery towards advancing plant genetic engineering. Trends Biotechnol. 36, 882–897. doi: 10.1016/j.tibtech.2018.03.009
Cvjetko, P., Milošić, A., Domijan, A.-M., Vrček, I. V., Tolić, S., Štefanić, P. P., et al. (2017). Toxicity of silver ions and differently coated silver nanoparticles in Allium cepa roots. Ecotoxicology Environ. Saf. 137, 18–28. doi: 10.1016/j.ecoenv.2016.11.009
Das, P., Barua, S., Sarkar, S., Chatterjee, S. K., Mukherjee, S., Goswami, L., et al. (2018). Mechanism of toxicity and transformation of silver nanoparticles: Inclusive assessment in earthworm-microbe-soil-plant system. Geoderma 314, 73–84. doi: 10.1016/j.geoderma.2017.11.008
Das, K., Roychoudhury, A. (2014). Reactive oxygen species (ROS) and response of antioxidants as ROS-scavengers during environmental stress in plants. Front. Environ. Sci. 2, 53. doi: 10.3389/fenvs.2014.00053
Demirer, G. S., Zhang, H., Goh, N. S., González-Grandío, E., Landry, M. P. (2019). Carbon nanotube–mediated DNA delivery without transgene integration in intact plants. Nat. Protoc. 14, 2954–2971. doi: 10.1038/s41596-019-0208-9
Dev, A., Srivastava, A. K., Karmakar, S. (2018). Nanomaterial toxicity for plants. Environ. Chem. Lett. 16, 85–100. doi: 10.1007/s10311-017-0667-6
Dimkpa, C. O., Mclean, J. E., Latta, D. E., Manangón, E., Britt, D. W., Johnson, W. P., et al. (2012). CuO and ZnO nanoparticles: phytotoxicity, metal speciation, and induction of oxidative stress in sand-grown wheat. J. Nanoparticle Res. 14, 1–15. doi: 10.1007/s11051-012-1125-9
Dimkpa, C. O., Singh, U., Bindraban, P. S., Elmer, W. H., Gardea-Torresdey, J. L., White, J. C. (2019). Zinc oxide nanoparticles alleviate drought-induced alterations in sorghum performance, nutrient acquisition, and grain fortification. Sci. Total Environ. 688, 926–934. doi: 10.1016/j.scitotenv.2019.06.392
Doman, J. L., Raguram, A., Newby, G. A., Liu, D. R. (2020). Evaluation and minimization of Cas9-independent off-target DNA editing by cytosine base editors. Nat. Biotechnol. 38, 620–628. doi: 10.1038/s41587-020-0414-6
Doolette, C. L., Mclaughlin, M. J., Kirby, J. K., Navarro, D. A. (2015). Bioavailability of silver and silver sulfide nanoparticles to lettuce (Lactuca sativa): Effect of agricultural amendments on plant uptake. J. hazardous materials 300, 788–795. doi: 10.1016/j.jhazmat.2015.08.012
Du, W., Gardea-Torresdey, J. L., Xie, Y., Yin, Y., Zhu, J., Zhang, X., et al. (2017). Elevated CO2 levels modify TiO2 nanoparticle effects on rice and soil microbial communities. Sci. Total Environ. 578, 408–416. doi: 10.1016/j.scitotenv.2016.10.197
El-Saadony, M. T., Saad, A. M., Najjar, A. A., Alzahrani, S. O., Alkhatib, F. M., Shafi, M. E., et al. (2021). The use of biological selenium nanoparticles to suppress Triticum aestivum L. crown and root rot diseases induced by Fusarium species and improve yield under drought and heat stress. Saudi J. Biol. Sci. 28, 4461–4471. doi: 10.1016/j.sjbs.2021.04.043
Elsheery, N. I., Helaly, M. N., El-Hoseiny, H. M., Alam-Eldein, S. M. (2020). Zinc oxide and silicone nanoparticles to improve the resistance mechanism and annual productivity of salt-stressed mango trees. Agronomy 10, 558. doi: 10.3390/agronomy10040558
El-Zohri, M., Al-Wadaani, N. A., Bafeel, S. O. (2021). Foliar sprayed green zinc oxide nanoparticles mitigate drought-induced oxidative stress in tomato. Plants 10, 2400. doi: 10.3390/plants10112400
Etesami, H., Fatemi, H., Rizwan, M. (2021). Interactions of nanoparticles and salinity stress at physiological, biochemical and molecular levels in plants: A review. Ecotoxicology Environ. Saf. 225, 112769. doi: 10.1016/j.ecoenv.2021.112769
Etesami, H., Maheshwari, D. K. (2018). Use of plant growth promoting rhizobacteria (PGPRs) with multiple plant growth promoting traits in stress agriculture: Action mechanisms and future prospects. Ecotoxicology Environ. Saf. 156, 225–246. doi: 10.1016/j.ecoenv.2018.03.013
Fahad, S., Hussain, S., Matloob, A., Khan, F. A., Khaliq, A., Saud, S., et al. (2015). Phytohormones and plant responses to salinity stress: a review. Plant Growth Regul. 75, 391–404. doi: 10.1007/s10725-014-0013-y
Faisal, M., Saquib, Q., Alatar, A. A., Al-Khedhairy, A. A., Hegazy, A. K., Musarrat, J. (2013). Phytotoxic hazards of NiO-nanoparticles in tomato: a study on mechanism of cell death. J. hazardous materials 250, 318–332. doi: 10.1016/j.jhazmat.2013.01.063
Faizan, M., Bhat, J. A., Chen, C., AlYemeni, M. N., Wijaya, L., Ahmad, P., et al. (2021). Zinc oxide nanoparticles (ZnO-NPs) induce salt tolerance by improving the antioxidant system and photosynthetic machinery in tomato. Plant Physiol. Biochem. 161, 122–130. doi: 10.1016/j.plaphy.2021.02.002
Faraji, J., Sepehri, A. (2020). Exogenous nitric oxide improves the protective effects of TiO 2 nanoparticles on growth, antioxidant system, and photosynthetic performance of wheat seedlings under drought stress. J. Soil Sci. Plant Nutr. 20, 703–714. doi: 10.1007/s42729-019-00158-0
Farhangi-Abriz, S., Torabian, S. (2018). Nano-silicon alters antioxidant activities of soybean seedlings under salt toxicity. Protoplasma 255, 953–962. doi: 10.1007/s00709-017-1202-0
Fathi, A., Zahedi, M., Torabian, S., Khoshgoftar, A. (2017). Response of wheat genotypes to foliar spray of ZnO and Fe2O3 nanoparticles under salt stress. J. Plant Nutr. 40, 1376–1385. doi: 10.1080/01904167.2016.1262418
Feichtmeier, N. S., Walther, P., Leopold, K. (2015). Uptake, effects, and regeneration of barley plants exposed to gold nanoparticles. Environ. Sci. pollut. Res. 22, 8549–8558. doi: 10.1007/s11356-014-4015-0
Fellmann, S., Eichert, T. (2017). Acute effects of engineered nanoparticles on the growth and gas exchange of Zea mays L.—What are the underlying causes? Water Air Soil pollut. 228, 1–13. doi: 10.1007/s11270-017-3364-y
Forni, C., Duca, D., Glick, B. R. (2017). Mechanisms of plant response to salt and drought stress and their alteration by rhizobacteria. Plant Soil 410, 335–356. doi: 10.1007/s11104-016-3007-x
Fubini, B., Fenoglio, I., Tomatis, M. (2007). Physicochemical characteristics of nanoparticles that determine potential toxicity. Nanotoxicology: characterization dosing Health effects. Informa Healthcare New York, 50–70.
Ganguly, S., Das, S., Dastidar, S. (2014). Effect of zinc sulphide nanoparticles on germination of seeds of vigna radiata and their subsequent acceleration of growth in presence of the nanoparticles. Eur. J. BioMed. Pharm. Sci. 1, 273–280.
Gao, H., Gadlage, M. J., Lafitte, H. R., Lenderts, B., Yang, M., Schroder, M., et al. (2020). Superior field performance of waxy corn engineered using CRISPR–Cas9. Nat. Biotechnol. 38, 579–581. doi: 10.1038/s41587-020-0444-0
García-Gómez, C., García, S., Obrador, A. F., González, D., Babín, M., Fernández, M. D. (2018). Effects of aged ZnO NPs and soil type on Zn availability, accumulation and toxicity to pea and beet in a greenhouse experiment. Ecotoxicology Environ. Saf. 160, 222–230. doi: 10.1016/j.ecoenv.2018.05.019
Ge, Y., Priester, J. H., Van De Werfhorst, L. C., Walker, S. L., Nisbet, R. M., An, Y.-J., et al. (2014). Soybean plants modify metal oxide nanoparticle effects on soil bacterial communities. Environ. Sci. Technol. 48, 13489–13496. doi: 10.1021/es5031646
Ghani, M. I., Saleem, S., Rather, S. A., Rehmani, M. S., Alamri, S., Rajput, V. D., et al. (2022). Foliar application of zinc oxide nanoparticles: An effective strategy to mitigate drought stress in cucumber seedling by modulating antioxidant defense system and osmolytes accumulation. Chemosphere 289, 133202. doi: 10.1016/j.chemosphere.2021.133202
Ghorbanpour, M., Hatami, M., Hatami, M. (2015). Activating antioxidant enzymes, hyoscyamine and scopolamine biosynthesis of Hyoscyamus Niger L. plants with nano-sized titanium dioxide and bulk application. Acta Agriculturae Slovenica 105, 23–32-23–32. doi: 10.14720/aas.2015.105.1.03
Ghosh, M., Bandyopadhyay, M., Mukherjee, A. (2010). Genotoxicity of titanium dioxide (TiO2) nanoparticles at two trophic levels: plant and human lymphocytes. Chemosphere 81, 1253–1262. doi: 10.1016/j.chemosphere.2010.09.022
Gohari, G., Mohammadi, A., Akbari, A., Panahirad, S., Dadpour, M. R., Fotopoulos, V., et al. (2020). Titanium dioxide nanoparticles (TiO2 NPs) promote growth and ameliorate salinity stress effects on essential oil profile and biochemical attributes of Dracocephalum moldavica. Sci. Rep. 10, 912. doi: 10.1038/s41598-020-57794-1
Guerriero, G., Sutera, F. M., Torabi-Pour, N., Renaut, J., Hausman, J.-F., Berni, R., et al. (2021). Phyto-courier, a silicon particle-based nano-biostimulant: evidence from Cannabis sativa exposed to salinity. ACS nano 15, 3061–3069. doi: 10.1021/acsnano.0c09488
Guo, J., Chi, J. (2014). Effect of Cd-tolerant plant growth-promoting rhizobium on plant growth and Cd uptake by Lolium multiflorum Lam. and Glycine max (L.) Merr. in Cd-contaminated soil. Plant Soil 375, 205–214. doi: 10.1007/s11104-013-1952-1
Hafez, Y., Attia, K., Alamery, S., Ghazy, A., Al-Doss, A., Ibrahim, E., et al. (2020). Beneficial effects of biochar and chitosan on antioxidative capacity, osmolytes accumulation, and anatomical characters of water-stressed barley plants. Agronomy 10, 630. doi: 10.3390/agronomy10050630
Hafez, E. M., Osman, H. S., Gowayed, S. M., Okasha, S. A., Omara, A. E.-D., Sami, R., et al. (2021). Minimizing the adversely impacts of water deficit and soil salinity on maize growth and productivity in response to the application of plant growth-promoting rhizobacteria and silica nanoparticles. Agronomy 11, 676. doi: 10.3390/agronomy11040676
Haghighi, M., Afifipour, Z., Mozafarian, M. (2012). The effect of N-Si on tomato seed germination under salinity levels. J. Biol. Environ. Sci. 6, 87–90.
Haider, F. U., Liqun, C., Coulter, J. A., Cheema, S. A., Wu, J., Zhang, R., et al. (2021). Cadmium toxicity in plants: Impacts and remediation strategies. Ecotoxicology Environ. Saf. 211, 111887. doi: 10.1016/j.ecoenv.2020.111887
Hasan, M. M., Gong, L., Nie, Z.-F., Li, F.-P., Ahammed, G. J., Fang, X.-W. (2021a). ABA-induced stomatal movements in vascular plants during dehydration and rehydration. Environ. Exp. Bot. 186, 104436. doi: 10.1016/j.envexpbot.2021.104436
Hasan, M. M., Skalicky, M., Jahan, M. S., Hossain, M. N., Anwar, Z., Nie, Z.-F., et al. (2021b). Spermine: its emerging role in regulating drought stress responses in plants. Cells 10, 261. doi: 10.3390/cells10020261
Hashem, A. H., Abdelaziz, A. M., Askar, A. A., Fouda, H. M., Khalil, A. M., Abd-Elsalam, K. A., et al. (2021). Bacillus megaterium-mediated synthesis of selenium nanoparticles and their antifungal activity against Rhizoctonia solani in faba bean plants. J. Fungi 7, 195. doi: 10.3390/jof7030195
Hendler-Neumark, A., Bisker, G. (2019). Fluorescent single-walled carbon nanotubes for protein detection. Sensors 19, 5403. doi: 10.3390/s19245403
Hernández-Hernández, H., Quiterio-Gutiérrez, T., Cadenas-Pliego, G., Ortega-Ortiz, H., Hernández-Fuentes, A. D., Cabrera de la Fuente, M., et al. (2019). Impact of selenium and copper nanoparticles on yield, antioxidant system, and fruit quality of tomato plants. Plants 8, 355. doi: 10.3390/plants8100355
Hezaveh, T. A., Pourakbar, L., Rahmani, F., Alipour, H. (2019). Interactive effects of salinity and ZnO nanoparticles on physiological and molecular parameters of rapeseed (Brassica napus L.). Commun. Soil Sci. Plant Anal. 50, 698–715. doi: 10.1080/00103624.2019.1589481
Hnilickova, H., Kraus, K., Vachova, P., Hnilicka, F. (2021). Salinity stress affects photosynthesis, malondialdehyde formation, and proline content in Portulaca oleracea L. Plants 10, 845. doi: 10.3390/plants10050845
Hojjat, S. S., Ganjali, A. (2016). The effect of silver nanoparticle on lentil seed germination under drought stress. Int. J. Farm Allied Sci. 5, 208–212.
Hossain, Z., Mustafa, G., Sakata, K., Komatsu, S. (2016). Insights into the proteomic response of soybean towards Al2O3, ZnO, and Ag nanoparticles stress. J. hazardous materials 304, 291–305. doi: 10.1016/j.jhazmat.2015.10.071
Hou, J., Wu, Y., Li, X., Wei, B., Li, S., Wang, X. (2018). Toxic effects of different types of zinc oxide nanoparticles on algae, plants, invertebrates, vertebrates and microorganisms. Chemosphere 193, 852–860. doi: 10.1016/j.chemosphere.2017.11.077
Hsin, Y.-H., Chen, C.-F., Huang, S., Shih, T.-S., Lai, P.-S., Chueh, P. J. (2008). The apoptotic effect of nanosilver is mediated by a ROS-and JNK-dependent mechanism involving the mitochondrial pathway in NIH3T3 cells. Toxicol. Lett. 179, 130–139. doi: 10.1016/j.toxlet.2008.04.015
Hu, P., An, J., Faulkner, M. M., Wu, H., Li, Z., Tian, X., et al. (2020). Nanoparticle charge and size control foliar delivery efficiency to plant cells and organelles. ACS nano 14, 7970–7986. doi: 10.1021/acsnano.9b09178
Hussain, I., Singh, A., Singh, N., Singh, P. (2019). Plant-nanoceria interaction: Toxicity, accumulation, translocation and biotransformation. South Afr. J. Bot. 121, 239–247. doi: 10.1016/j.sajb.2018.11.013
Hussein, M., Abou-Baker, N. (2018). The contribution of nano-zinc to alleviate salinity stress on cotton plants. R. Soc. Open Sci. 5, 171809. doi: 10.1098/rsos.171809
Ibrahim, W., Zhu, Y. M., Chen, Y., Qiu, C. W., Zhu, S., Wu, F. (2019). Genotypic differences in leaf secondary metabolism, plant hormones and yield under alone and combined stress of drought and salinity in cotton genotypes. Physiologia plantarum 165, 343–355. doi: 10.1111/ppl.12862
Iqbal, M. N., Rasheed, R., Ashraf, M. Y., Ashraf, M. A., Hussain, I. (2018). Exogenously applied zinc and copper mitigate salinity effect in maize (Zea mays L.) by improving key physiological and biochemical attributes. Environ. Sci. pollut. Res. 25, 23883–23896. doi: 10.1007/s11356-018-2383-6
Jaberzadeh, A., Moaveni, P., Moghadam, H. R. T., Zahedi, H. (2013). Influence of bulk and nanoparticles titanium foliar application on some agronomic traits, seed gluten and starch contents of wheat subjected to water deficit stress. Notulae botanicae horti agrobotanici cluj-napoca 41, 201–207. doi: 10.15835/nbha4119093
Jacobasch, C., Völker, C., Giebner, S., Völker, J., Alsenz, H., Potouridis, T., et al. (2014). Long-term effects of nanoscaled titanium dioxide on the cladoceran Daphnia magna over six generations. Environ. pollut. 186, 180–186. doi: 10.1016/j.envpol.2013.12.008
Jahan, S., Alias, Y. B., Bakar, A. F. B. A., Yusoff, I. B. (2018). Toxicity evaluation of ZnO and TiO2 nanomaterials in hydroponic red bean (Vigna angularis) plant: physiology, biochemistry and kinetic transport. J. Environ. Sci. 72, 140–152. doi: 10.1016/j.jes.2017.12.022
Javed, R., Usman, M., Yücesan, B., Zia, M., Gürel, E. (2017). Effect of zinc oxide (ZnO) nanoparticles on physiology and steviol glycosides production in micropropagated shoots of Stevia rebaudiana Bertoni. Plant Physiol. Biochem. 110, 94–99. doi: 10.1016/j.plaphy.2016.05.032
Joudeh, N., Linke, D. (2022). Nanoparticle classification, physicochemical properties, characterization, and applications: a comprehensive review for biologists. J. Nanobiotechnology 20, 262. doi: 10.1186/s12951-022-01477-8
Kalteh, M., Alipour, Z. T., Ashraf, S., Marashi Aliabadi, M., Falah Nosratabadi, A. (2018). Effect of silica nanoparticles on basil (Ocimum basilicum) under salinity stress. J. Chem. Health Risks 4, 49–55. doi: 10.22034/jchr.2018.544075
Kandhol, N., Jain, M., Tripathi, D. K. (2022). Nanoparticles as potential hallmarks of drought stress tolerance in plants. Physiologia Plantarum 174, e13665. doi: 10.1111/ppl.13665
Khalid, M. F., Iqbal Khan, R., Jawaid, M. Z., Shafqat, W., Hussain, S., Ahmed, T., et al. (2022). Nanoparticles: the plant saviour under abiotic stresses. Nanomaterials 12, 3915. doi: 10.3390/nano12213915
Khan, M., Khan, M. S. A., Borah, K. K., Goswami, Y., Hakeem, K. R., Chakrabartty, I. (2021). The potential exposure and hazards of metal-based nanoparticles on plants and environment, with special emphasis on ZnO NPs, TiO2 NPs, and AgNPs: a review. Environ. Adv. 6, 100128. doi: 10.1016/j.envadv.2021.100128
Khan, I., Raza, M. A., Awan, S. A., Shah, G. A., Rizwan, M., Ali, B., et al. (2020). Amelioration of salt induced toxicity in pearl millet by seed priming with silver nanoparticles (AgNPs): The oxidative damage, antioxidant enzymes and ions uptake are major determinants of salt tolerant capacity. Plant Physiol. Biochem. 156, 221–232. doi: 10.1016/j.plaphy.2020.09.018
Khan, I., Saeed, K., Khan, I. (2019). Nanoparticles: Properties, applications and toxicities. Arabian J. Chem. 12, 908–931. doi: 10.1016/j.arabjc.2017.05.011
Khodakovskaya, M., Dervishi, E., Mahmood, M., Xu, Y., Li, Z., Watanabe, F., et al. (2009). Carbon nanotubes are able to penetrate plant seed coat and dramatically affect seed germination and plant growth. ACS nano 3, 3221–3227. doi: 10.1021/nn900887m
Kiapour, H., Moaveni, P., Habibi, D. (2015). Evaluation of the application of gibbrellic acid and titanium dioxide nanoparticles under drought stress on some traits of basil (Ocimum basilicum L.)., 138–150.
Kim, D. H., Gopal, J., Sivanesan, I. (2017). Nanomaterials in plant tissue culture: the disclosed and undisclosed. RSC Adv. 7, 36492–36505. doi: 10.1039/C7RA07025J
Kim, Y. T., Han, J. H., Hong, B. H., Kwon, Y. U. (2010). Electrochemical synthesis of CdSe quantum-dot arrays on a graphene basal plane using mesoporous silica thin-film templates. Advanced Materials 22, 515–518. doi: 10.1002/adma.200902736
Kim, J.-H., Oh, Y., Yoon, H., Hwang, I., Chang, Y.-S. (2015). Iron nanoparticle-induced activation of plasma membrane H+-ATPase promotes stomatal opening in Arabidopsis thaliana. Environ. Sci. Technol. 49, 1113–1119. doi: 10.1021/es504375t
Klaine, S. J., Alvarez, P. J., Batley, G. E., Fernandes, T. F., Handy, R. D., Lyon, D. Y., et al. (2008). Nanomaterials in the environment: behavior, fate, bioavailability, and effects. Environ. Toxicol. Chemistry: Int. J. 27, 1825–1851. doi: 10.1897/08-090.1
Kopittke, P. M., Lombi, E., Wang, P., Schjoerring, J. K., Husted, S. (2019). Nanomaterials as fertilizers for improving plant mineral nutrition and environmental outcomes. Environ. Science: Nano 6, 3513–3524. doi: 10.1039/C9EN00971J
Król, A., Amarowicz, R., Weidner, S. (2014). Changes in the composition of phenolic compounds and antioxidant properties of grapevine roots and leaves (Vitis vinifera L.) under continuous of long-term drought stress. Acta Physiologiae Plantarum 36, 1491–1499. doi: 10.1007/s11738-014-1526-8
Kubiak, M. (2014). Dendrimers-fascinating nanoparticles in the application in medicine. Chemik 68, 146–150.
Kumar, S., Dilbaghi, N., Saharan, R., Bhanjana, G. (2012). Nanotechnology as emerging tool for enhancing solubility of poorly water-soluble drugs. Bionanoscience 2, 227–250. doi: 10.1007/s12668-012-0060-7
Kwak, S.-Y., Lew, T. T. S., Sweeney, C. J., Koman, V. B., Wong, M. H., Bohmert-Tatarev, K., et al. (2019). Chloroplast-selective gene delivery and expression in planta using chitosan-complexed single-walled carbon nanotube carriers. Nat. nanotechnology 14, 447–455. doi: 10.1038/s41565-019-0375-4
Landa, P., Cyrusova, T., Jerabkova, J., Drabek, O., Vanek, T., Podlipna, R. (2016). Effect of metal oxides on plant germination: phytotoxicity of nanoparticles, bulk materials, and metal ions. Water Air Soil pollut. 227, 1–10. doi: 10.1007/s11270-016-3156-9
Larue, C., Castillo-Michel, H., Sobanska, S., Cécillon, L., Bureau, S., Barthès, V., et al. (2014). Foliar exposure of the crop Lactuca sativa to silver nanoparticles: evidence for internalization and changes in Ag speciation. J. hazardous materials 264, 98–106. doi: 10.1016/j.jhazmat.2013.10.053
Lee, W.-M., Kwak, J. I., An, Y.-J. (2012). Effect of silver nanoparticles in crop plants Phaseolus radiatus and Sorghum bicolor: media effect on phytotoxicity. Chemosphere 86, 491–499. doi: 10.1016/j.chemosphere.2011.10.013
Li, L., Luo, Y., Li, R., Zhou, Q., Peijnenburg, W. J., Yin, N., et al. (2020). Effective uptake of submicrometre plastics by crop plants via a crack-entry mode. Nat. sustainability 3, 929–937. doi: 10.1038/s41893-020-0567-9
Li, M., Zhang, P., Adeel, M., Guo, Z., Chetwynd, A. J., Ma, C., et al. (2021). Physiological impacts of zero valent iron, Fe3O4 and Fe2O3 nanoparticles in rice plants and their potential as Fe fertilizers. Environ. pollut. 269, 116134. doi: 10.1016/j.envpol.2020.116134
Lin, D., Xing, B. (2007). Phytotoxicity of nanoparticles: inhibition of seed germination and root growth. Environ. pollut. 150, 243–250. doi: 10.1016/j.envpol.2007.01.016
Linh, T. M., Mai, N. C., Hoe, P. T., Lien, L. Q., Ban, N. K., Hien, L. T. T., et al. (2020). Metal-based nanoparticles enhance drought tolerance in soybean. J. Nanomaterials 2020, 1–13. doi: 10.1155/2020/4056563
Liu, J., Li, G., Chen, L., Gu, J., Wu, H., Li, Z. (2021). Cerium oxide nanoparticles improve cotton salt tolerance by enabling better ability to maintain cytosolic K+/Na+ ratio. J. Nanobiotechnology 19, 153. doi: 10.1186/s12951-021-00892-7
Liu, F.-Y., Xiong, F.-X., Fan, Y.-K., Li, J., Wang, H.-Z., Xing, G.-M., et al. (2016). Facile and scalable fabrication engineering of fullerenol nanoparticles by improved alkaline-oxidation approach and its antioxidant potential in maize. J. Nanoparticle Res. 18, 1–13. doi: 10.1007/s11051-016-3642-4
López-Moreno, M. L., de la Rosa, G., Hernández-Viezcas, J.Á., Castillo-Michel, H., Botez, C. E., Peralta-Videa, J. R., et al. (2010). Evidence of the differential biotransformation and genotoxicity of ZnO and CeO2 nanoparticles on soybean (Glycine max) plants. Environ. Sci. Technol. 44, 7315–7320. doi: 10.1021/es903891g
Lu, L., Huang, M., Huang, Y., Corvini, P. F.-X., Ji, R., Zhao, L. (2020). Mn 3 O 4 nanozymes boost endogenous antioxidant metabolites in cucumber (Cucumis sativus) plant and enhance resistance to salinity stress. Environ. Science: Nano 7, 1692–1703. doi: 10.1039/D0EN00214C
Lv, Z., Jiang, R., Chen, J., Chen, W. (2020). Nanoparticle-mediated gene transformation strategies for plant genetic engineering. Plant J. 104, 880–891. doi: 10.1111/tpj.14973
Maharramov, A., Ahmadov, I., Ramazanov, M., Aliyeva, S., Ramazanli, V. (2015). Fluorescence emission spectrum of elodea leaves exposed to nanoparticles. J. Biomaterials Nanobiotechnology 6, 135. doi: 10.4236/jbnb.2015.63013
Majumdar, S., Almeida, I. C., Arigi, E. A., Choi, H., Verberkmoes, N. C., Trujillo-Reyes, J., et al. (2015). Environmental effects of nanoceria on seed production of common bean (Phaseolus vulgaris): a proteomic analysis. Environ. Sci. Technol. 49, 13283–13293. doi: 10.1021/acs.est.5b03452
Manickavasagam, M., Pavan, G., Vasudevan, V. (2019). A comprehensive study of the hormetic influence of biosynthesized AgNPs on regenerating rice calli of indica cv. IR64. Sci. Rep. 9, 8821. doi: 10.1038/s41598-019-45214-y
Manzoor, N., Ali, L., Ahmed, T., Noman, M., Adrees, M., Shahid, M. S., et al. (2022). Recent advancements and development in nano-enabled agriculture for improving abiotic stress tolerance in plants. Front. Plant Sci. 13, 951752. doi: 10.3389/fpls.2022.951752
Marusenko, Y., Shipp, J., Hamilton, G. A., Morgan, J. L., Keebaugh, M., Hill, H., et al. (2013). Bioavailability of nanoparticulate hematite to Arabidopsis thaliana. Environ. pollut. 174, 150–156. doi: 10.1016/j.envpol.2012.11.020
Maswada, H., Djanaguiraman, M., Prasad, P. (2018). Seed treatment with nano-iron (III) oxide enhances germination, seeding growth and salinity tolerance of sorghum. J. Agron. Crop Sci. 204, 577–587. doi: 10.1111/jac.12280
Matzke, M., Jurkschat, K., Backhaus, T. (2014). Toxicity of differently sized and coated silver nanoparticles to the bacterium Pseudomonas putida: risks for the aquatic environment? Ecotoxicology 23, 818–829. doi: 10.1007/s10646-014-1222-x
Mishra, V., Mishra, R. K., Dikshit, A., Pandey, A. C. (2014). “Interactions of nanoparticles with plants: an emerging prospective in the agriculture industry,” in Emerging Technologies And Management Of Crop Stress Tolerance (Florida, United States: Elsevier), 159–180.
Mittal, D., Kaur, G., Singh, P., Yadav, K., Ali, S. A. (2020). Nanoparticle-based sustainable agriculture and food science: Recent advances and future outlook. Front. Nanotechnology 2, 579954. doi: 10.3389/fnano.2020.579954
Mohamed, A. K. S., Qayyum, M. F., Abdel-Hadi, A. M., Rehman, R. A., Ali, S., Rizwan, M. (2017). Interactive effect of salinity and silver nanoparticles on photosynthetic and biochemical parameters of wheat. Arch. Agron. Soil Sci. 63, 1736–1747. doi: 10.1080/03650340.2017.1300256
Mohammadi, H., Esmailpour, M., Gheranpaye, A. (2016). Effects of TiO2 nanoparticles and water-deficit stress on morpho-physiological characteristics of dragonhead (Dracocephalum moldavica L.) plants. Acta Agriculturae Slovenica 107, 385–396. doi: 10.14720/aas.2016.107.2.11
Moradbeygi, H., Jamei, R., Heidari, R., Darvishzadeh, R. (2020). Investigating the enzymatic and non-enzymatic antioxidant defense by applying iron oxide nanoparticles in Dracocephalum moldavica L. plant under salinity stress. Scientia Hortic. 272, 109537. doi: 10.1016/j.scienta.2020.109537
Mosa, K. A., El-Naggar, M., Ramamoorthy, K., Alawadhi, H., Elnaggar, A., Wartanian, S., et al. (2018). Copper nanoparticles induced genotoxicty, oxidative stress, and changes in superoxide dismutase (SOD) gene expression in cucumber (Cucumis sativus) plants. Front. Plant Sci. 9, 872. doi: 10.3389/fpls.2018.00872
Mozafari, A. A., Havas, F., Ghaderi, N. (2018). Application of iron nanoparticles and salicylic acid in in vitro culture of strawberries (Fragaria× ananassa Duch.) to cope with drought stress. Plant Cell Tissue Organ Culture (PCTOC) 132, 511–523. doi: 10.1007/s11240-017-1347-8
Mukherjee, A., Peralta-Videa, J. R., Bandyopadhyay, S., Rico, C. M., Zhao, L., Gardea-Torresdey, J. L. (2014). Physiological effects of nanoparticulate ZnO in green peas (Pisum sativum L.) cultivated in soil. Metallomics 6, 132–138. doi: 10.1039/C3MT00064H
Mustafa, H., Ilyas, N., Akhtar, N., Raja, N. I., Zainab, T., Shah, T., et al. (2021). Biosynthesis and characterization of titanium dioxide nanoparticles and its effects along with calcium phosphate on physicochemical attributes of wheat under drought stress. Ecotoxicology Environ. Saf. 223, 112519. doi: 10.1016/j.ecoenv.2021.112519
Nair, P. M. G., Chung, I. M. (2015). Changes in the growth, redox status and expression of oxidative stress related genes in chickpea (Cicer arietinum L.) in response to copper oxide nanoparticle exposure. J. Plant Growth Regul. 34, 350–361. doi: 10.1007/s00344-014-9468-3
Najafi, S., Razavi, S. M., Khoshkam, M., Asadi, A. (2020). Effects of green synthesis of sulfur nanoparticles from Cinnamomum zeylanicum barks on physiological and biochemical factors of Lettuce (Lactuca sativa). Physiol. Mol. Biol. Plants 26, 1055–1066. doi: 10.1007/s12298-020-00793-3
Nasiri, R., Dabagh, S., Meamar, R., Idris, A., Muhammad, I., Irfan, M., et al. (2020). Papain grafted into the silica coated iron-based magnetic nanoparticles ‘IONPs@ SiO2-PPN’as a new delivery vehicle to the HeLa cells. Nanotechnology 31, 195603. doi: 10.1088/1361-6528/ab6fd4
Nayan, R., Rawat, M., Negi, B., Pande, A., Arora, S. (2016). Zinc sulfide nanoparticle mediated alterations in growth and anti-oxidant status of Brassica juncea. Biologia 71, 896–902. doi: 10.1515/biolog-2016-0107
Nejatzadeh, F. (2021). Effect of silver nanoparticles on salt tolerance of Satureja hortensis l. during in vitro and in vivo germination tests. Heliyon 7. doi: 10.1016/j.heliyon.2021.e05981
Nekrasov, V., Staskawicz, B., Weigel, D., Jones, J. D., Kamoun, S. (2013). Targeted mutagenesis in the model plant Nicotiana benthamiana using Cas9 RNA-guided endonuclease. Nat. Biotechnol. 31, 691–693. doi: 10.1038/nbt.2655
Nel, A., Xia, T., Madler, L., Li, N. (2006). Toxic potential of materials at the nanolevel. science 311, 622–627. doi: 10.1126/science.1114397
Noohpisheh, Z., Amiri, H., Mohammadi, A., Farhadi, S. (2021). Effect of the foliar application of zinc oxide nanoparticles on some biochemical and physiological parameters of Trigonella foenum-graecum under salinity stress. Plant Biosystems-An Int. J. Dealing all Aspects Plant Biol. 155, 267–280. doi: 10.1080/11263504.2020.1739160
Panda, K. K., Achary, V. M. M., Krishnaveni, R., Padhi, B. K., Sarangi, S. N., Sahu, S. N., et al. (2011). In vitro biosynthesis and genotoxicity bioassay of silver nanoparticles using plants. Toxicol. Vitro 25, 1097–1105. doi: 10.1016/j.tiv.2011.03.008
Pande, A., Arora, S. (2019). Nanotechnological interventions for improving plant health and productivity. In: Khurana, S., Gaur, R. (eds) Plant Biotechnology: Progress in Genomic Era. Springer, Singapore. doi: 10.1007/978-981-13-8499-8_17
Paramo, L. A., Feregrino-Pérez, A. A., Guevara, R., Mendoza, S., Esquivel, K. (2020). Nanoparticles in agroindustry: Applications, toxicity, challenges, and trends. Nanomaterials 10, 1654. doi: 10.3390/nano10091654
Peng, C., Xu, C., Liu, Q., Sun, L., Luo, Y., Shi, J. (2017). Fate and transformation of CuO nanoparticles in the soil–rice system during the life cycle of rice plants. Environ. Sci. Technol. 51, 4907–4917. doi: 10.1021/acs.est.6b05882
Pérez-De-Luque, A. (2017). Interaction of nanomaterials with plants: what do we need for real applications in agriculture? Front. Environ. Sci. 5, 12. doi: 10.3389/fenvs.2017.00012
Pérez-Labrada, F., López-Vargas, E. R., Ortega-Ortiz, H., Cadenas-Pliego, G., Benavides-Mendoza, A., Juárez-Maldonado, A. (2019). Responses of tomato plants under saline stress to foliar application of copper nanoparticles. Plants 8, 151. doi: 10.3390/plants8060151
Pittol, M., Tomacheski, D., Simões, D. N., Ribeiro, V. F., Santana, R. M. C. (2017). Macroscopic effects of silver nanoparticles and titanium dioxide on edible plant growth. Environ. nanotechnology Monit. Manage. 8, 127–133. doi: 10.1016/j.enmm.2017.07.003
Prajitha, N., Athira, S., Mohanan, P. (2019). Bio-interactions and risks of engineered nanoparticles. Environ. Res. 172, 98–108. doi: 10.1016/j.envres.2019.02.003
Prasad, R., Bhattacharyya, A., Nguyen, Q. D. (2017). Nanotechnology in sustainable agriculture: recent developments, challenges, and perspectives. Front. Microbiol. 8, 1014. doi: 10.3389/fmicb.2017.01014
Qi, M., Liu, Y., Li, T. (2013). Nano-TiO 2 improve the photosynthesis of tomato leaves under mild heat stress. Biol. Trace element Res. 156, 323–328. doi: 10.1007/s12011-013-9833-2
Rahmatpour, S., Mosaddeghi, M. R., Shirvani, M., Šimůnek, J. (2018). Transport of silver nanoparticles in intact columns of calcareous soils: The role of flow conditions and soil texture. Geoderma 322, 89–100. doi: 10.1016/j.geoderma.2018.02.016
Rajput, V. D., Harish, Singh, R. K., Verma, K. K., Sharma, L., Quiroz-Figueroa, F. R., et al. (2021a). Recent developments in enzymatic antioxidant defence mechanism in plants with special reference to abiotic stress. Biology 10, 267. doi: 10.3390/biology10040267
Rajput, V. D., Minkina, T. M., Behal, A., Sushkova, S. N., Mandzhieva, S., Singh, R., et al. (2018). Effects of zinc-oxide nanoparticles on soil, plants, animals and soil organisms: A review. Environ. Nanotechnology Monit. Manage. 9, 76–84. doi: 10.1016/j.enmm.2017.12.006
Rajput, V. D., Minkina, T., Kumari, A., Harish, Singh, V. K., Verma, K. K., et al. (2021b). Coping with the challenges of abiotic stress in plants: New dimensions in the field application of nanoparticles. Plants 10, 1221. doi: 10.3390/plants10061221
Raliya, R., Franke, C., Chavalmane, S., Nair, R., Reed, N., Biswas, P. (2016). Quantitative understanding of nanoparticle uptake in watermelon plants. Front. Plant Sci. 7, 1288. doi: 10.3389/fpls.2016.01288
Ranjan, A., Rajput, V. D., Minkina, T., Bauer, T., Chauhan, A., Jindal, T. (2021). Nanoparticles induced stress and toxicity in plants. Environ. nanotechnology Monit. Manage. 15, 100457. doi: 10.1016/j.enmm.2021.100457
Rao, S., Shekhawat, G. S. (2016). Phytotoxicity and oxidative stress perspective of two selected nanoparticles in Brassica juncea. 3 Biotech. 6, 244. doi: 10.1007/s13205-016-0550-3
Rasheed, A., Li, H., Tahir, M. M., Mahmood, A., Nawaz, M., Shah, A. N., et al. (2022). The role of nanoparticles in plant biochemical, physiological, and molecular responses under drought stress: A review. Front. Plant Sci. 13. doi: 10.3389/fpls.2022.976179
Rico, C. M., Peralta-Videa, J., Gardea-Torresdey, J. (2015). “Chemistry, biochemistry of nanoparticles, and their role in antioxidant defense system in plants,” in Nanotechnology and Plant Sciences: Nanoparticles and Their Impact on Plants (Switzerland: Springer), 1–17.
Rossi, L., Zhang, W., Lombardini, L., Ma, X. (2016). The impact of cerium oxide nanoparticles on the salt stress responses of Brassica napus L. Environ. pollut. 219, 28–36. doi: 10.1016/j.envpol.2016.09.060
Roy, R., Kumar, S., Tripathi, A., Das, M., Dwivedi, P. D. (2014). Interactive threats of nanoparticles to the biological system. Immunol. Lett. 158, 79–87. doi: 10.1016/j.imlet.2013.11.019
Rubilar, O., Rai, M., Tortella, G., Diez, M. C., Seabra, A. B., Durán, N. (2013). Biogenic nanoparticles: copper, copper oxides, copper sulphides, complex copper nanostructures and their applications. Biotechnol. Lett. 35, 1365–1375. doi: 10.1007/s10529-013-1239-x
Rushton, P. J., Somssich, I. E., Ringler, P., Shen, Q. J. (2010). WRKY transcription factors. Trends Plant Sci. 15, 247–258. doi: 10.1016/j.tplants.2010.02.006
Sabaghnia, N., Janmohammadi, M. (2015). Effect of nano-silicon particles application on salinity tolerance in early growth of some lentil genotypes. Annales Universitatis Mariae Curie-Sklodowska, sectio C – Biologia 69 (2), 39. doi: 10.17951/c.2014.69.2.39
Savithramma, N., Ankanna, S., Bhumi, G. (2012). Effect of nanoparticles on seed germination and seedling growth of Boswellia ovalifoliolata an endemic and endangered medicinal tree taxon. Nano Vision 2, 2.
Schaller, J., Brackhage, C., Paasch, S., Brunner, E., Bäucker, E., Dudel, E. G. (2013). Silica uptake from nanoparticles and silica condensation state in different tissues of Phragmites australis. Sci. Total Environ. 442, 6–9. doi: 10.1016/j.scitotenv.2012.10.016
Schwab, F., Zhai, G., Kern, M., Turner, A., Schnoor, J. L., Wiesner, M. R. (2016). Barriers, pathways and processes for uptake, translocation and accumulation of nanomaterials in plants–Critical review. Nanotoxicology 10, 257–278. doi: 10.3109/17435390.2015.1048326
Sedghi, M., Hadi, M., Toluie, S. G. (2013). Effect of nano zinc oxide on the germination parameters of soybean seeds under drought stress. Ann. West Univ Timisoara Ser. Biol. 16, 73.
Semida, W. M., Abdelkhalik, A., Mohamed, G. F., Abd El-Mageed, T. A., Abd El-Mageed, S. A., Rady, M. M., et al. (2021). Foliar application of zinc oxide nanoparticles promotes drought stress tolerance in eggplant (Solanum melongena L.). Plants 10, 421. doi: 10.3390/plants10020421
Shafea, A. A., Dawood, M. F., Zidan, M. A. (2017). Wheat seedlings traits as affected by soaking at titanium dioxide nanoparticles. Environment Earth Ecol. 1, 102–111. doi: 10.24051/eee/68607
Shah, T., Latif, S., Saeed, F., Ali, I., Ullah, S., Alsahli, A. A., et al. (2021). Seed priming with titanium dioxide nanoparticles enhances seed vigor, leaf water status, and antioxidant enzyme activities in maize (Zea mays L.) under salinity stress. J. King Saud University-Science 33, 101207. doi: 10.1016/j.jksus.2020.10.004
Shahi, S., Srivastava, M. (2018). Influence of foliar application of manganese on growth, pigment content, and nitrate reductase activity of Vigna radiata (L.) R. Wilczek under salinity. J. Plant Nutr. 41, 1397–1404. doi: 10.1080/01904167.2018.1454470
Shahid, M. A., Sarkhosh, A., Khan, N., Balal, R. M., Ali, S., Rossi, L., et al. (2020). Insights into the physiological and biochemical impacts of salt stress on plant growth and development. Agronomy 10, 938. doi: 10.3390/agronomy10070938
Shallan, M. A., Hassan, H., Namich, A. A., Ibrahim, A. A. (2016). Biochemical and physiological effects of TiO2 and SiO2 nanoparticles on cotton plant under drought stress. Res. J. Pharmaceutical Biol. Chem. Sci. 7, 1540–1551.
Shan, Q., Wang, Y., Li, J., Zhang, Y., Chen, K., Liang, Z., et al. (2013). Targeted genome modification of crop plants using a CRISPR-Cas system. Nat. Biotechnol. 31, 686–688. doi: 10.1038/nbt.2650
Shaw, A. K., Hossain, Z. (2013). Impact of nano-CuO stress on rice (Oryza sativa L.) seedlings. Chemosphere 93, 906–915. doi: 10.1016/j.chemosphere.2013.05.044
Shi, J., Abid, A. D., Kennedy, I. M., Hristova, K. R., Silk, W. K. (2011). To duckweeds (Landoltia punctata), nanoparticulate copper oxide is more inhibitory than the soluble copper in the bulk solution. Environ. pollut. 159, 1277–1282. doi: 10.1016/j.envpol.2011.01.028
Siddiqui, H., Ahmed, K. B. M., Sami, F., Hayat, S. (2020). “Silicon nanoparticles and plants: Current knowledge and future perspectives,” in Sustainable Agriculture Reviews 41: Nanotechnology for Plant Growth and Development (Switzerland: Springer), 129–142.
Silva, S., Dias, M. C., Silva, A. M. (2022). Titanium and zinc based nanomaterials in agriculture: A promising approach to deal with (a) biotic stresses? Toxics 10, 172. doi: 10.3390/toxics10040172
Singh, D., Sillu, D., Kumar, A., Agnihotri, S. (2021). Dual nanozyme characteristics of iron oxide nanoparticles alleviate salinity stress and promote the growth of an agroforestry tree, Eucalyptus tereticornis Sm. Environ. Science: Nano 8, 1308–1325. doi: 10.1039/D1EN00040C
Sotiriou, G. A., Pratsinis, S. E. (2010). Antibacterial activity of nanosilver ions and particles. Environ. Sci. Technol. 44, 5649–5654. doi: 10.1021/es101072s
Srinivasan, C., Saraswathi, R. (2010). Nano-agriculture―carbon nanotubes enhance tomato seed germination and plant growth. Curr. Sci. 99, 274–275.
Stark, W. J. (2011). Nanoparticles in biological systems. Angewandte Chemie Int. Edition 50, 1242–1258. doi: 10.1002/anie.200906684
Staroń, A., Długosz, O., Pulit-Prociak, J., Banach, M. (2020). Analysis of the exposure of organisms to the action of nanomaterials. Materials 13, 349. doi: 10.3390/ma13020349
Sun, L., Song, F., Guo, J., Zhu, X., Liu, S., Liu, F., et al. (2020). Nano-ZnO-induced drought tolerance is associated with melatonin synthesis and metabolism in maize. Int. J. Mol. Sci. 21, 782. doi: 10.3390/ijms21030782
Suriyaprabha, R., Karunakaran, G., Yuvakkumar, R., Prabu, P., Rajendran, V., Kannan, N. (2012). Growth and physiological responses of maize (Zea mays L.) to porous silica nanoparticles in soil. J. Nanoparticle Res. 14, 1–14. doi: 10.1007/s11051-012-1294-6
Sutulienė, R., Ragelienė, L., Samuolienė, G., Brazaitytė, A., Urbutis, M., Miliauskienė, J. (2021). The response of antioxidant system of drought-stressed green pea (Pisum sativum L.) affected by watering and foliar spray with silica nanoparticles. Horticulturae 8, 35. doi: 10.3390/agriculture10090396
Taran, N., Storozhenko, V., Svietlova, N., Batsmanova, L., Shvartau, V., Kovalenko, M. (2017). Effect of zinc and copper nanoparticles on drought resistance of wheat seedlings. Nanoscale Res. Lett. 12, 1–6. doi: 10.1186/s11671-017-1839-9
Thounaojam, T. C., Meetei, T. T., Devi, Y. B., Panda, S. K., Upadhyaya, H. (2021). Zinc oxide nanoparticles (ZnO-NPs): a promising nanoparticle in renovating plant science. Acta Physiologiae Plantarum 43, 136. doi: 10.1007/s11738-021-03307-0
Thuesombat, P., Hannongbua, S., Akasit, S., Chadchawan, S. (2014). Effect of silver nanoparticles on rice (Oryza sativa L. cv. KDML 105) seed germination and seedling growth. Ecotoxicology Environ. Saf. 104, 302–309. doi: 10.1016/j.ecoenv.2014.03.022
Tombuloglu, H., Slimani, Y., Tombuloglu, G., Alshammari, T., Almessiere, M., Korkmaz, A. D., et al. (2020). Engineered magnetic nanoparticles enhance chlorophyll content and growth of barley through the induction of photosystem genes. Environ. Sci. pollut. Res. 27, 34311–34321. doi: 10.1007/s11356-020-09693-1
Torabian, S., Farhangi-Abriz, S., Zahedi, M. (2018). Efficacy of FeSO 4 nano formulations on osmolytes and antioxidative enzymes of sunflower under salt stress. Indian J. Plant Physiol. 23, 305–315. doi: 10.1007/s40502-018-0366-8
Torabian, S., Zahedi, M., Khoshgoftar, A. H. (2016). Effects of foliar spray of two kinds of zinc oxide on the growth and ion concentration of sunflower cultivars under salt stress. J. Plant Nutr. 39, 172–180. doi: 10.1080/01904167.2015.1009107
Torney, F., Trewyn, B. G., Lin, V. S.-Y., Wang, K. (2007). Mesoporous silica nanoparticles deliver DNA and chemicals into plants. Nat. nanotechnology 2, 295–300. doi: 10.1038/nnano.2007.108
Tripathi, D. K., Rai, P., Guerriero, G., Sharma, S., Corpas, F. J., Singh, V. P. (2021a). Silicon induces adventitious root formation in rice under arsenate stress with involvement of nitric oxide and indole-3-acetic acid. J. Exp. Bot. 72, 4457–4471. doi: 10.1093/jxb/eraa488
Tripathi, D. K., Vishwakarma, K., Singh, V. P., Prakash, V., Sharma, S., Muneer, S., et al. (2021b). Silicon crosstalk with reactive oxygen species, phytohormones and other signaling molecules. J. hazardous materials 408, 124820. doi: 10.1016/j.jhazmat.2020.124820
Umair Hassan, M., Aamer, M., Umer Chattha, M., Haiying, T., Shahzad, B., Barbanti, L., et al. (2020). The critical role of zinc in plants facing the drought stress. Agriculture 10, 396. doi: 10.3390/horticulturae8010035
Van Nguyen, D., Nguyen, H. M., Le, N. T., Nguyen, K. H., Nguyen, H. T., Le, H. M., et al. (2021). Copper nanoparticle application enhances plant growth and grain yield in maize under drought stress conditions. J. Plant Growth Regul. 41, 1–12. doi: 10.1007/s00344-021-10301-w
Verma, S. K., Das, A. K., Gantait, S., Kumar, V., Gurel, E. (2019). Applications of carbon nanomaterials in the plant system: A perspective view on the pros and cons. Sci. Total Environ. 667, 485–499. doi: 10.1016/j.scitotenv.2019.02.409
Wahid, I., Kumari, S., Ahmad, R., Hussain, S. J., Alamri, S., Siddiqui, M. H., et al. (2020). Silver nanoparticle regulates salt tolerance in wheat through changes in ABA concentration, ion homeostasis, and defense systems. Biomolecules 10, 1506. doi: 10.3390/biom10111506
Wang, Y., Cheng, X., Shan, Q., Zhang, Y., Liu, J., Gao, C., et al. (2014). Simultaneous editing of three homoeoalleles in hexaploid bread wheat confers heritable resistance to powdery mildew. Nat. Biotechnol. 32, 947–951. doi: 10.1038/nbt.2969
Wang, H., Kou, X., Pei, Z., Xiao, J. Q., Shan, X., Xing, B. (2011). Physiological effects of magnetite (Fe3O4) nanoparticles on perennial ryegrass (Lolium perenne L.) and pumpkin (Cucurbita mixta) plants. Nanotoxicology 5, 30–42. doi: 10.3109/17435390.2010.489206
Wang, P., Lombi, E., Zhao, F.-J., Kopittke, P. M. (2016). Nanotechnology: a new opportunity in plant sciences. Trends Plant Sci. 21, 699–712. doi: 10.1016/j.tplants.2016.04.005
Wang, X., Xie, H., Wang, P., Yin, H. (2023). Nanoparticles in plants: uptake, transport and physiological activity in leaf and root. Materials 16, 3097. doi: 10.3390/ma16083097
Weir, A., Westerhoff, P., Fabricius, L., Hristovski, K., Von Goetz, N. (2012). Titanium dioxide nanoparticles in food and personal care products. Environ. Sci. Technol. 46, 2242–2250. doi: 10.1021/es204168d
Wu, B., Zhu, L., Le, X. C. (2017). Metabolomics analysis of TiO2 nanoparticles induced toxicological effects on rice (Oryza sativa L.). Environ. pollut. 230, 302–310. doi: 10.1016/j.envpol.2017.06.062
Xie, X., He, Z., Chen, N., Tang, Z., Wang, Q., Cai, Y. (2019). The roles of environmental factors in regulation of oxidative stress in plant. BioMed. Res. Int. 2019. doi: 10.1155/2019/9732325
Yadav, B., Jogawat, A., Rahman, M. S., Narayan, O. P. (2021). Secondary metabolites in the drought stress tolerance of crop plants: A review. Gene Rep. 23, 101040. doi: 10.1016/j.genrep.2021.101040
Yang, J., Cao, W., Rui, Y. (2017). Interactions between nanoparticles and plants: phytotoxicity and defense mechanisms. J. Plant Interact. 12, 158–169. doi: 10.1080/17429145.2017.1310944
Yang, K.-Y., Doxey, S., Mclean, J. E., Britt, D., Watson, A., Al Qassy, D., et al. (2018). Remodeling of root morphology by CuO and ZnO nanoparticles: effects on drought tolerance for plants colonized by a beneficial pseudomonad. Botany 96, 175–186. doi: 10.1139/cjb-2017-0124
Yang, C., Powell, C. A., Duan, Y., Shatters, R., Zhang, M. (2015). Antimicrobial nanoemulsion formulation with improved penetration of foliar spray through citrus leaf cuticles to control citrus huanglongbing. PloS One 10, e0133826. doi: 10.1371/journal.pone.0133826
Yanık, F., Vardar, F. (2015). Toxic effects of aluminum oxide (Al 2 O 3) nanoparticles on root growth and development in Triticum aestivum. Water Air Soil pollut. 226, 1–13. doi: 10.1007/s11270-015-2566-4
Yasur, J., Rani, P. U. (2013). Environmental effects of nanosilver: impact on castor seed germination, seedling growth, and plant physiology. Environ. Sci. pollut. Res. 20, 8636–8648. doi: 10.1007/s11356-013-1798-3
Ye, Y., Cota-Ruiz, K., HernáNdez-Viezcas, J. A., Valdes, C., Medina-Velo, I. A., Turley, R. S., et al. (2020). Manganese nanoparticles control salinity-modulated molecular responses in Capsicum annuum L. through priming: A sustainable approach for agriculture. ACS Sustain. Chem. Eng. 8, 1427–1436. doi: 10.1021/acssuschemeng.9b05615
Yuan, Z., Ye, Y., Gao, F., Yuan, H., Lan, M., Lou, K., et al. (2013). Chitosan-graft-β-cyclodextrin nanoparticles as a carrier for controlled drug release. Int. J. pharmaceutics 446, 191–198. doi: 10.1016/j.ijpharm.2013.02.024
Zahedi, S. M., Hosseini, M. S., Daneshvar Hakimi Meybodi, N., Peijnenburg, W. (2021). Mitigation of the effect of drought on growth and yield of pomegranates by foliar spraying of different sizes of selenium nanoparticles. J. Sci. Food Agric. 101, 5202–5213. doi: 10.1002/jsfa.11167
Zhang, H., Demirer, G. S., Zhang, H., Ye, T., Goh, N. S., Aditham, A. J., et al. (2019). DNA nanostructures coordinate gene silencing in mature plants. Proc. Natl. Acad. Sci. 116, 7543–7548. doi: 10.1073/pnas.1818290116
Zhang, W., Wang, Q., Chen, H. (2021). Challenges in characterization of nanoplastics in the environment. Front. Environ. Sci. Eng. 16, 11. doi: 10.1007/s11783-021-1445-z
Zhao, L.-S., Li, K., Wang, Q.-M., Song, X.-Y., Su, H.-N., Xie, B.-B., et al. (2017). Nitrogen starvation impacts the photosynthetic performance of Porphyridium cruentum as revealed by chlorophyll a fluorescence. Sci. Rep. 7, 8542. doi: 10.1038/s41598-017-08428-6
Zhao, L., Peralta-Videa, J. R., Varela-Ramirez, A., Castillo-Michel, H., Li, C., Zhang, J., et al. (2012). Effect of surface coating and organic matter on the uptake of CeO2 NPs by corn plants grown in soil: Insight into the uptake mechanism. J. Hazardous Materials 225, 131–138. doi: 10.1016/j.jhazmat.2012.05.008
Keywords: drought, genetic engineering, nanoparticles, nanotoxicity, salinity
Citation: Rehman A, Khan S, Sun F, Peng Z, Feng K, Wang N, Jia Y, Pan Z, He S, Wang L, Qayyum A, Du X and Li H (2024) Exploring the nano-wonders: unveiling the role of Nanoparticles in enhancing salinity and drought tolerance in plants. Front. Plant Sci. 14:1324176. doi: 10.3389/fpls.2023.1324176
Received: 19 October 2023; Accepted: 26 December 2023;
Published: 17 January 2024.
Edited by:
Peng Wang, Jiangsu Province and Chinese Academy of Sciences, ChinaReviewed by:
Keni Cota-Ruiz, Utica University, United StatesCopyright © 2024 Rehman, Khan, Sun, Peng, Feng, Wang, Jia, Pan, He, Wang, Qayyum, Du and Li. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Hongge Li, bGlob25nZ2VAY2Fhcy5jbg==; Xiongming Du, ZHV4aW9uZ21pbmdAY2Fhcy5jbg==
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.