 Kun Ma
Kun Ma Caochuang Fang
Caochuang Fang- Shanghai Key Laboratory of Protected Horticultural Technology, Horticultural Research Institute, Shanghai Academy of Agricultural Sciences, Shanghai, China
Modern megacities offer convenient lifestyles to their citizens. However, agriculture is becoming increasingly vulnerable, especially during unexpected public health emergencies such as pandemics. Fortunately, the adaptability of terrace vegetables cultivation presents an opportunity to grow horticultural crops in residential spaces, bringing numerous benefits to citizens, including enhanced nutrition and recreational engagement in the cultivation process. Although certain planting skills and equipment have been developed, the citizens tend to sow some seeds with unknown pedigree, it is rare to find new plant varieties specifically bred for cultivation as terrace vegetables. To expand the genetic basis of new breeding materials, elite parents, and varieties (pre-breeding) for terrace cultivation, this review not only discusses the molecular breeding strategy for the identification, creation, and application of rational alleles for improving horticultural characteristics including plant architecture, flavor quality, and ornamental character, but also assesses the potential for terrace cultivation of some representative vegetable crops. We conclude that the process of pre-breeding specifically for terrace cultivation environments is vital for generating a genetic basis for urban terrace vegetable crops.
1 Introduction
Terrace cultivation had existed since the days of Babylon in 2300 BC (Udayan and Sreedaya, 2018). While in modern societies, terraces/balconies are the only places that citizens could cultivate their own vegetables with the expanding of megacities (Chitra, 2021), so local nutrient sources are rapidly depleting, consequently hindering food supplies. Moreover, as the COVID-19 pandemic subsides, challenges to food supplies induced by public health disturbances suggest that urban nutrient resilience may be crucial for meeting the needs of megacities (Langemeyer et al., 2021). Urban nutrient resilience reflects the sustainability of cities, which is based on resource availability and the need to enhance food supplies and quality of life (Zeng et al., 2022). Fortunately, terrace cultivation of vegetable crops plays a crucial role in urban nutrient resilience as it not only strives to provide food daily or emergently, but also enhances recreation in high-density cities. Additionally, in many countries such as USA, India, China, and Korea, terrace cultivation is becoming more and more popular among the citizens, and many cultivations equipment or materials are sales good.
The terrace cultivation of vegetable crops is characterized by utilizing edge/corner vacancies or installing box/shelf-shaped equipment on terraces, balconies or roofs of high-density residential buildings for the purpose of planting and growing vegetables. For this, complete commercialized cultivation substrates have been matched adequately for cultivation of vegetables (Kader et al., 2022). Many planting modes can be utilized on balconies or roofs, including placing pots along terrace handrails, utilizing compact versions of vertical farming equipment, employing light-emitting diode (LED) photon boxes, utilizing hollowed walls, creating extended grooves, and using slope shelves with aerosol cans. Thus, it is a half-open, half-protected, half-controlled, flexible, and relatively cramped cultivating environment. To adapt to this environment, three prerequisites are required for the cultivated plants and surely for the germplasm: First, adaptation to the limited planting space, which is a major limiting factor. The plant architecture should be reduced to a size that can be accommodated by terraces, balconies or roofs. Second, to enhance nutrient supply to citizens, it is necessary to ensure high levels of nutrient content and flavor quality. Terrace vegetables have the potential to independently produce rare, fresh, more nutritional, more flavorful, and higher value-added vegetable products. This eliminates the need for logistics processes and helps address urgent disruptions in nutrient supply caused by delays in vegetable transportation due to a pandemic or other social reasons, i.e., terrace vegetable cultivation contributes to increasing urban nutrient resilience. Third, being connected to the human environment, the ornamental value of vegetable germplasms should also be considered to complement the existing urban landscape ecology and satisfy the preferences of citizens engaged in terrace vegetable cultivation.
To obtain germplasms with the specialized attributes required for terrace vegetable systems, we propose the use of pre-breeding strategy as a bridge to connect genetic variations with breeding programs (Akkenapally and Kumar, 2022). Pre-breeding refers to all activities designed to identify desirable characteristics and/or genes from unadopted (exotic or semi-exotic) materials, including those that, although adapted have been subjected to any kind of selection for improvement (Akkenapally and Kumar, 2022). This strategy allows for the combination of valuable artificial or natural alleles into the recipient material, resulting in the development of highly beneficial germplasms. The aim of this review is to summarize an integrated pre-breeding strategy for expanding terrace vegetable germplasms by means of showing some examples of vegetable crops.
2 General breeding traits and breeding methods for terrace vegetables
Compact plant architecture is the highest priority breeding target for terrace vegetables. Under this premise, lots of vegetable crops have the potential for use in terrace vegetable systems, while certain plants do not possess traits that align with the three prerequisites. For instance, the size of most fruit vegetable crops exceeds the available space on a terrace. Meanwhile, although some germplasms exhibit a compact plant architecture, they may not yield as highly or adapt well to continuous harvest which might be eliminated. While for most leafy vegetables and root vegetables, their plant heights naturally suitable for terrace environments, so the flavor quality, nutrient quality, and ornamental value should be more concerned in pre-breeding programs. The evaluation of adaptability and deficiency of different vegetables for terrace cultivation were listed in Supplementary Table 1 as examples for reference.
The 5G breeding is an approach that fully reflects the newest modern molecular breeding spirits; these 5Gs are 1st G Genome assembly, 2nd G Germplasm characterization, 3rd G Gene function identification, 4th G Genomic breeding (GB), and 5th G Gene editing (GE) (Varshney et al., 2020). Combining the concepts of pre-breeding, the clustered regularly interspaced short palindromic repeats (CRISPR) is included by the 5th G, the quantitative trait locus (QTL) mapping is included by the 2nd, 3rd and 4th G, the genome-wide associated study (GWAS) is included by the 2nd G, and the marker assisted selection (MAS) is included by the 4th G, which are all commonly used molecular breeding technologies that can be employed to improve specific characteristics of crops, according to the purpose of pre-breeding.
3 Tomatoes genetic improvement exhibits a representative of pre-breeding strategy for terrace cultivation
Tomato is one of the most popular fruit vegetable crops around the world in a long history (Razifard et al., 2020). However, the plant architecture of most tomato cultivars is relatively taller for terrace cultivation. In tomatoes, three main effective genes are useful for molding compactness in the plant architecture (Kwon et al., 2019): SISP, SISP5G, and SLER. The gene SlSP has been found to delay flowering time and promote indeterminate growth, SlSP5G delays flowering time especially under long-day conditions, and SLER promotes the elongation of internodes (Torii et al., 1996; Pnueli et al., 1998; Xu et al., 2015; Soyk et al., 2016). Mutations in SlSP, whether naturally occurring or induced by CRISPR, result in a determinate growth habit without yield loss, while mutating of SlSP5G accelerates flowering in long-day conditions (Pnueli et al., 1998; Soyk et al., 2016). Additionally, the natural Sler, mutant shows shortened internodes and extremely compact inflorescences, forming tight fruit clusters (Kwon et al., 2019). The double-mutated genotype Slsp/Slsp5g exhibits rapid cycling and compacts the plants without affecting fruit number or yield when cultivated in high-density planting spaces (Soyk et al., 2016). The triple mutant Slsp/Sp5g/Sler is the most compact and exhibits significantly decreased fruit weight and yield than the sp mutant in the Mo82 background. Fortunately, the triple mutant has similar fruit numbers per plant, Brix content, and yield when planted in high-density LED-assisted photon greenhouse fields compared with those exhibited by the sp mutant in the Sweet100 background, indicating that the triple mutated genotype is most suitable for tomato terrace planting (Kwon et al., 2019).
Flavor quality, being the foundation of edibility, represents crucial targets for pre-breeding efforts (Bomgardner, 2017). The utilization of CRISPR mutants of SlINVINH1 and SlVPE5 resulted in increased glucose, fructose, and Brix values, raising levels of 40.82%, 42.76%, and 32.76, as well as 35.83%, 43.0%, and 32.43% than the parental lines, respectively (Wang B et al., 2021). These improvements were achieved without any significant alteration in fruit weight (Wang B et al., 2021; Kawaguchi et al., 2021). A genome-wide association study (GWAS) identified a main effect quantitative trait locus (QTL) located on chromosome 9 at position 62.64 M, which was found to explain 28.73% of the phenotypical variation observed during a 3-year test (Kim et al., 2021). Additionally, conditional QTL for taste quality should be explored and used as modern terrace -cropping often uses photon suppliers such as HPS and LED lamps. In a QTL mapping analysis of Brix values, two QTL regions have been identified; the first QTL was located on chromosome 2 at 43.5 - 50.5 Mb under both HPS and LED conditions, explaining 20% of the phenotypic variance; and the second QTL was found on chromosome 6 at 43.7 - 47.1 Mb only under the LED condition, explaining 26% of the phenotypic variance (Prinzenberg et al., 2021). The additive effect of these QTLs suggests that stacking high flavor quality alleles could lead to continuous improvement in Brix values in elite breeding lines. For example, a triple Brix-value-QTL pyramided isogenic introgression line showed a 145% higher Brix yield compared to the original M82 tomato line, providing valuable material for breeding (Gur and Zamir, 2015; Prinzenberg et al., 2021; Wang Z et al., 2022).
Aroma is also an essential component of flavor quality. In a previous study, QTL mapping indicated that SlFLORAL4 was the candidate gene for the phenylalanine-derived volatile locus on chromosome 4 in tomato, and the contents of 2-phenylethanol, phenylacetaldehyde, and volatile 1-nitro-2-phenylethane were significantly reduced in the CRISPR mutant of SlFLORAL4 (Tikunov et al., 2020). This finding highlights the importance of utilizing natural variations for high aroma quality. Due to the relatively rich genetic basis of aroma, many elite lines with aroma have been developed (Tzin et al., 2015); thus, reverse genetic strategies, such as CRISPR, may offer a faster approach than forward genetic strategies (e.g., QTL mapping) for the production of high-quality breeding materials.
Regarding ornamental value, color is an important component of the traits which contributes to the ornamental value of terrace mini-horticultural landscapes. Since some painting pigments are extracted from natural plants, the potential exists to match colors artificially in living plants. The synthesis of some plant pigments has been genetically dissected and can be used in breeding procedures. As reported for tomatoes, CRISPR interruption of SlPSY1, SlMYB12, and SlSGR1 interrupts the synthesis of carotenoids, naringenin chalcone, and chlorophyll, respectively, resulting in fruit color changes from red to yellow, pink, and brown (Yang et al., 2022). Triple mutants, with mutations in all three genes, exhibit a light green color; while double mutants, depending on the genes mutated, display light yellow, pink-brown, and yellow-green colors (Yang et al., 2022), thus demonstrating the ability to match new colors using basic pigments and genes (allelic variations) extracted from plant organs (Ye et al., 2022).
4 Pre-breeding for other types of terrace vegetables
4.1 Liana and fruit vegetables
For most liana and fruit vegetables, relatively taller plant architecture is the limitation factor which need to be improved for terrace cultivation.
For example, the ER orthologs CmER, CsER, and CmoER regulate internode length in melon, cucumber, and pumpkin, respectively (Torii et al., 1996; Xin et al., 2022). As predicted, the internode lengths of CRISPR-mutated genotypes Cmer, Cser, and Cmoer were 40%, 34%, and 60% shorter, respectively, compared to their parallel wild genotypes (Xin et al., 2022). The rare natural variation in the 5’ UTR of CmoYABBY1, known as the genetic essence of the Bu locus, results in a bushy architecture with clustered leaves and highly compressed internodes in the CRISPR mutant genotype Cmoyabby1/bu. This mutant genotype exhibits similar yield per plant to CmoYABBY1 but significantly higher yield per square meter under high plant density conditions (Wang S et al., 2022). The suppression of stem length by CmoYABBY1 variation is dose-dependent (Wang S et al., 2022), suggesting the potential value of Cmoyabby1/bu in adapting to different types of terraces structures. In the case of CsTFL1 in cucumber, a non-synonymous SNP disrupts the interaction between CsTFL1 and CsNOT2a, resulting in a loss of its ability to delay flowering (Wen et al., 2019). CsTFL1 is expressed in the subapical regions of the shoot apical meristem, lateral meristem, and young stems, indicating its multiple effects on plant architecture. Knockdown of CsTFL1 through RNAi leads to determinate growth and the formation of terminal flowers, resulting in a significant reduction in plant size, while flowering time remains unaffected (Wen et al., 2019). Although CsTFL1 does not possess a CRISPR mutation, the natural non-synonymous allele can be utilized for breeding dwarf cucumbers using marker-assisted selection (MAS). Recent advancements in Agrobacterium-mediated transgenic and CRISPR technologies have enabled the genetic modification of an increasing number of Cucurbitaceae crops to achieve a compact plant size.
Kiwifruit, a newly domesticated climbing woody perennial liana crop, utilizes CEN-like genes as flowering time repressors (Varkonyi-Gasic et al., 2013; Voogd et al., 2017). CRISPR-induced bi-allelic mutations of AcCEN4 and AcCEN in kiwifruit result in a compact annual plant with axillary inflorescences and rapid terminal flower and fruit development, making it suitable for terrace vegetable cultivation (Varkonyi-Gasic et al., 2018). Furthermore, a QTL on chromosome 26 increases vitamin C content in kiwifruit (McCallum et al., 2019), which help kiwifruit to a high nutrient food source for citizens.
For groundcherry, a related fruit vegetable crop of tomato, PgER is a member of the ER gene family that regulates stem length (Zhang et al., 2018; Kwon et al., 2019). The phenotype of the mutant Pger is more severely condensed compared to that of the tomato Sler mutant and resembles the Slsp/Sp5g/Sler triple mutant, while maintaining a similar fruit number and Brix content (Kwon et al., 2019). While, as for fruit crops like capsicum, the so-called chilli, the plant architecture is very suitable for terrace environment, and its flavor quality and nutrient quality has also been fit the scope of consumption of citizens, they could be adopted directly, the genetic improvement of capsicum in other traits is a kind of “add flowers to the brocade” for terrace cultivation.
4.2 Leafy vegetables
Leafy vegetables are very suitable for terrace cultivation for their dwarf or cramped architecture which fit the volume of terrace and cultivating facilities accordantly, including but not limited to lettuce, kale, broccoli, and bolt used rapeseed. The ornamental value and nutrient quality will become bonuses for these species if they could.
Rll1 is a gene that induce the synthesis of anthocyanins, resulting in the red color of lettuce leaves (Su et al., 2019). Chlorophyll can also influence external traits, as observed in lettuce, where LsVAR2 induces the formation of green speckles in the albino cotyledon (Nguyen et al., 2021). In kale, the CRISPR knockdown of BoaCRTISO results in the simultaneous reduction of chlorophyll and carotenoid concentrations. As a result, the leaf color changes from green to yellow, which weakens the color-masking effect of chlorophyll (Sun et al., 2020). In broccoli, a QTL mapping study indicates that two QTLs named Pur7.1 and Pur9.1 facilitate the biosynthesis of anthocyanin, and induce the purple cauliflower phenotype (Liu et al., 2022). Further, the CRISPR mutation of BolMYB28 increases glucoraphanin content in the leaves (Kim et al., 2022), which induces a healthcare usage of broccoli. For rapeseed, the edible value is reflected through bolting, and the ornamental value is reflected through flowering. PAP2 induces the synthesis of anthocyanins, resulting in the pink color of rapeseed petals, and CCD4 induces carotenoid synthesis, resulting in yellow coloration in rapeseed petals (Ye et al., 2022). The combination of anthocyanins and carotenoids forms a new color, i.e., apricot flower petal in rapeseed via the co-expression of CCD4 and PAP2 (Ye et al., 2022). By the way, early bolting could be accomplished by MAS (Fang et al., 2022), which makes the citizens could harvest the bolts earlier.
Compared to other types of vegetable crops, the current deficiency is the lack of reported genetic research of flavor quality for leafy vegetables, and most researchers use cultivation skills, fertilizers, and equipment to enhance flavor quality (Silva and Zorzeto, 2019; Gangathilaka et al., 2022). However, the anticipated carbon peak in 2050 (Zheng et al., 2022) means that enhanced genetic strategies may be a more efficient means of saving energy and resources, thereby improving megacity sustainability. We believe that the development of pan-genomes will allow the discovery of more alleles for pre-breeding using CRISPR or MAS for terrace vegetables (Fei, 2019; Gao et al., 2019).
4.3 Root vegetables
Root vegetables are also very suitable for terrace cultivation, including but not limited to carrots, onions, or turnip. Taking carrots as an example, the plant height and width of carrot is perfect for terrace environment, and its nutrient quality is also excellent, and its ornamentally proper for terraces. In spite of this, carrots could be genetically improved for higher nutrient content and ornamental value. For instance, DcMYB7 induces the synthesis of anthocyanin, resulting in the purple color of taproot (Xu et al., 2019). While the heterologous expression of CYP76AD1, DODA1, and DOPA5GT induces betalain in carrot, resulting in a red-violet color in the taproot (Deng et al., 2023). These genes not only increase the ornamental value of carrots, but also enhance medicinal and edible homologous functions. Generally, root vegetables expand the range of options available for terrace cultivation. Although some marker-free alleles of the mentioned genes above have yet to be identified, they hold the potential to serve as valuable pre-breeding resources.
4.4 Cereal crops
Flexibly consider, some cereal crops also suitable for terrace cultivation for their special identities of flavor or nutrient.
For instance, fresh maize can be eaten as vegetable in diets. In maize, the flavor compound 2-acetyl-1-pyrroline (2AP) is regulated by the activity of betaine aldehyde dehydrogenase 2 (BADH2). Natural maize varieties do not produce 2AP, but CRISPR-generated double mutants of ZmBADH2a and ZmBADH2b can produce 2AP in fresh and dry maize seeds, enhancing the aroma profile (Wang Y. et al., 2021).
Natural disasters, sudden epidemics, or uncertainties in human society can disrupt vegetable supply, leading to potential nutrient deficiencies. In such situations, nutrient enhancement or “biofortification” becomes crucial to ensure an adequate supply of important nutrients such as chlorophyll, cellulose, and vitamins from plants (Vlčko and Ohnoutková, 2019; Bhambhani et al., 2021), which is an important part of the resilience of megacities (Junior, 2017; Langemeyer et al., 2021). Just as the CRISPR mutation of OsHOL1 increases iodine content (Carlessi et al., 2021), the CRISPR-mediated marker-free double insertion of SSU-crtI and ZmPsy has been shown to increase carotene content in rice (Dong et al., 2020). Further, as the CRISPR mutation of TaIPK1 improves iron and zinc accumulation in wheat (Ibrahim et al., 2021), and the CRISPR mutation of FtMYB45 promotes flavonoid biosynthesis in buckwheat (Wen et al., 2022).
5 Hybrid vegetable seeds are encouraged for terrace cultivation
Although the basic three prerequisites are the basic requirements of the cultivars, sufficient yield is also need to be noted, especially for the smaller architecture plants. Sometime, it is not realistic to expect smaller plants modified for cramped conditions to produce comparable yields to those of stronger, taller plants grown in larger spaces. To address this issue, on one hand, the existed heterotic patterns should be insisted; on the other hand, we propose a single-gene advantageous stacking strategy to increase fruit yield in cramped plants (Fang et al., 2022). For instance, several gene families, including IAA7, FLC, TFL, SFT, and SSP, have been shown to have significant effects on plant architecture and exhibit strong single-gene heterosis, leading to increased yield (Krieger et al., 2010; Guo et al., 2014; Park et al., 2014; Li et al., 2019; Fang et al., 2022). Heterozygous genotypes of these genes can be stacked using polycistronic CRISPR or QTL pyramiding in hybrid breeding systems (Fang et al., 2022). Conversely, some high-yield-related alleles are effective in the homozygous state and can be stacked in elite lines to compensate for the reduced yields of smaller plants (Gur and Zamir, 2015; Song et al., 2021; Wang S et al., 2022).
6 Summary and expectation
In conclusion, the terrace vegetable system is a broad concept, and traditional food and oil crops, vegetables, and fruits are likely to become specialized commercial varieties for terrace vegetable systems after genetic improvement, provided that the plant materials meet the three main requirements described in this review (Figure 1; Supplementary Table 2) i.e., the breeders and citizens could choose their own vegetable crops following the information including but not limited to Supplementary Table 1. Furthermore, we recognize that the terrace vegetable system can provide several benefits. It offers flexibility in the supply of fresh vegetables, ensuring the availability of high-quality and nutritious food for urban populations. Additionally, it contributes to improving the microecological environment of urban residents, enhancing their well-being. Moreover, it increases the ornamental value of fruits and vegetables, adding to the aesthetic appeal of urban landscapes. Finally, it provides opportunities for citizen recreation, which can be considered a luxury in megacities.
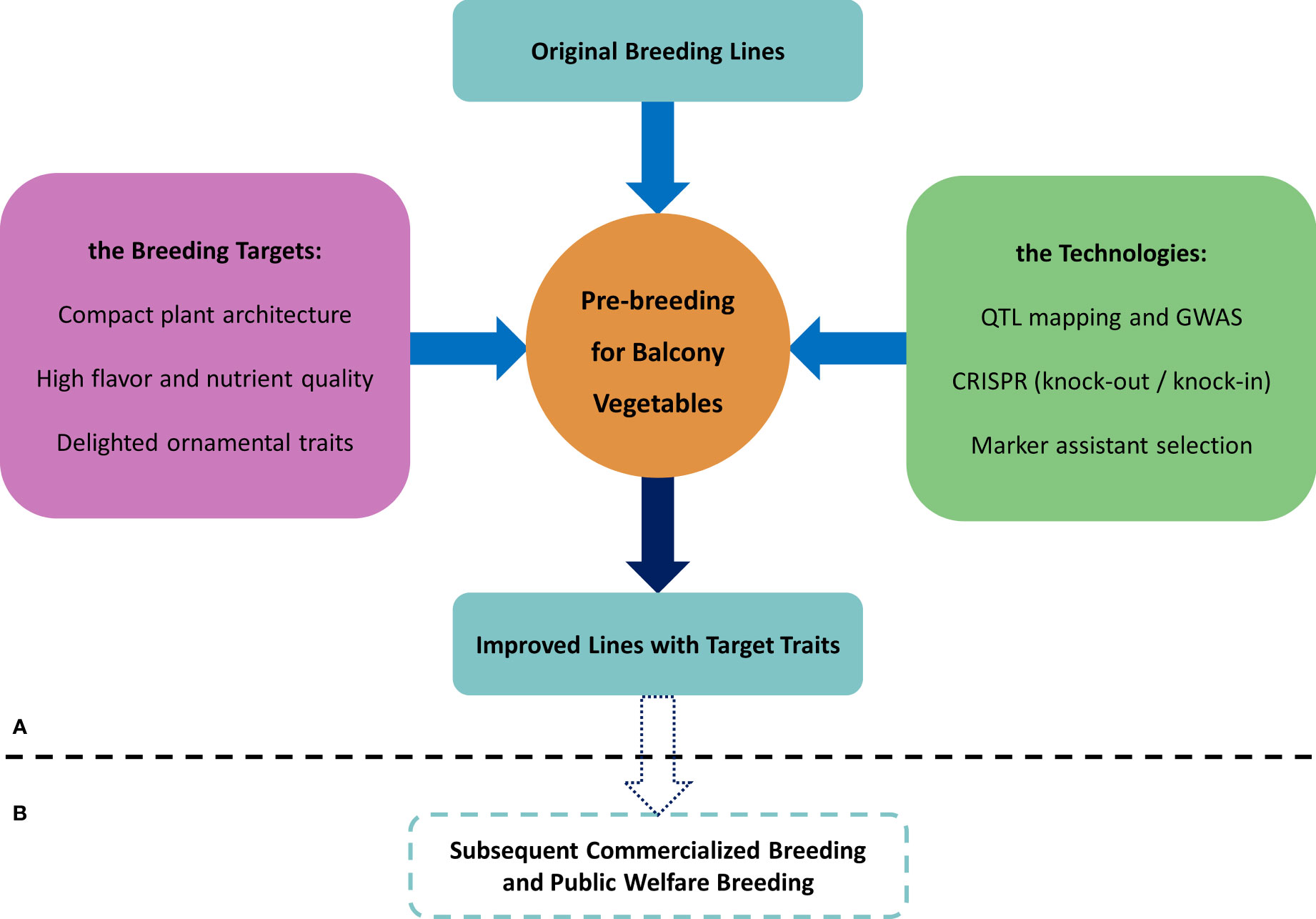
Figure 1 The thumbnail of pre-breeding. (A) Mind mapping of pre-breeding for terrace vegetables. (B) Subsequent steps after pre-breeding.
A major constraint is that parts of the citizens tend to get access to the seeds free of cost, even when the seeds are unknown pedigrees or segregating individuals of hybrids. To solve this problem which might hinder the development of the seed industry for terrace cultivation, on one hand, the breeders could enhance the level of breeding procedures and produce high performance seeds to attract citizens to buy; on the other hand, breeders could host some public activities to donate some seeds to the citizens as welfares. Our pre-breeding work strategy is a pivotal step to produce elite parental lines not only for commercialized breeding but also for welfare breeding.
In spring 2021, the vegetable garden owned by the first author of this review received attention from the public and media at the 10th China Flower EXPO and won a prize for scientific and technological innovation. This highlights that the selection and breeding of vegetable varieties suitable for special urban planting environments meet the current demand for urban green sustainable development and the needs of people living in cities. Thus, pre-breeding of terrace vegetables have forward-thinking and commercial application value. Only a few vegetable types have been genetically dissected, and no commercial varieties have been specifically bred for terrace vegetables. However, pre-breeding programs provide several breeding lines for subsequent steps, showing potential for yielding the most suitable vegetable varieties in the near future and informing important future research on gene function. With the development of 5G breeding (Varshney et al., 2020), more horticultural crops and even field crops could be genetically improved to fulfill the scope of terrace cultivation of vegetable crops. Thus, genetic improvement and the use of pre-breeding will not only support a promising industry but also enhance the nutrient resilience of megacities (Supplementary Figure 1).
Author contributions
KM and CF wrote the manuscript. YY helped to revise the manuscript. CF and KM designed the study. KM shared the experience. CF prepared the figures. All authors contributed to the article and approved the submitted version.
Funding
This work was supported by “Research and demonstration of key technologies for innovation and cultivation of vegetable germplasm resources for urban families and communities to ornamental and edible vegetables (2112216)” from Science and Technology Commission of Shanghai Municipality (Shanghai Administration of Foreign Experts Affairs) , and "Evaluation and identification of characteristic melon-vegetable germplasm resources of the 2023 special plan for research in the support field of agricultural science and technology innovation (Nong-Ke-Ying-Ji 2023(03))" from Shanghai Academy of Agricultural Sciences.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fpls.2023.1237099/full#supplementary-material
References
Bhambhani, S., Kondhare, K. R., Giri, A. (2021). Advanced genome editing strategies for manipulation of plant specialized metabolites pertaining to biofortification. Phytochem. Rev. 21, 81–99. doi: 10.1007/s11101-021-09749-1
Bomgardner, M. M. (2017). These four foods are ripe for CRISPR gene editing. Chem. Eng. News 95 (24). doi: 10.1021/cen-09524-cover2
Carlessi, M., Mariotti, L., Giaume, F., Fornara, F., Perata, P., Gonzali, S. (2021). Targeted knockout of the gene OsHOL1 removes methyl iodide emissions from rice plants. Sci. Rep. 11, 17010.
Chitra, K. (2021). Establishment of organic terrace garden and cultivation of nutritious vegetables. J. Univ. Shanghai Sci. Technol. 23 (12). doi: 10.51201/jusst/21/121048
Deng, Y., Duan, A., Liu, H., Wang, Y., Zhang, R., Xu, Z., et al. (2023). Generating colorful carrot germplasm through metabolic engineering of betalains pigments. Horticult. Res. 10 (4). doi: 10.1093/hr/uhad024
Dong, O. X., Yu, S., Jain, R., Zhang, N., Duong, P. Q., Butler, C., et al. (2020). Marker-free carotenoid-enriched rice generated through targeted gene insertion using CRISPR-Cas9. Nat. Commun. 11, 1178. doi: 10.1038/s41467-020-14981-y
Fang, C., Wang, Z., Wang, P., Song, Y., Ahmad, A., Dong, F., et al. (2022). Heterosis derived from nonadditive effects of the BnFLC homologs coordinates early flowering and high yield in rapeseed (Brassica napus L.). Front. Plant Sci. 12. doi: 10.3389/fpls.2021.798371
Fei, Z. (2019). The tomato pan-genome uncovers new genes and a rare allele regulating fruit flavor. Nat. Genet. 51, 1044–1051. doi: 10.1038/s41588-019-0410-2
Gangathilaka, K., Premarathna, M., Madawala, H., Seneviratne, G. (2022). Can Biofilm biofertilizer cut down chemical fertilizers in leafy vegetable cultivation? A case study with Centella asiatica (Gotukola). J. Agricul. Sci. – Sri Lanka. 17(3), 370–378. doi: 10.4038/jas.v17i3.9918
Gao, L., Gonda, I., Sun, H., Ma, Q., Bao, K., Tieman, D. M., et al. (2019). The tomato pan-genome 334 uncovers new genes and a rare allele regulating fruit flavor. Nature Genetics 51, 1044–1051. doi: 10.5061/dryad.m463f7k
Guo, Y., Hans, H. H., Christian, J., Molina, C. (2014). Mutations in single FT- and TFL1- paralogs of rapeseed (Brassica napus L.) and their impact on flowering time and yield components. Front. Plant Sci. 5. doi: 10.3389/fpls.2014.00282
Gur, A., Zamir, D. (2015). Mendelizing all components of a pyramid of three yield QTL in tomato. Front. Plant Sci. 6. doi: 10.3389/fpls.2015.01096
Ibrahim, S., Saleem, B., Rehman, N., Zafar, S. A., Naeem, M. K., Khan, M. R. (2021). CRISPR/Cas9 mediated disruption of Inositol Pentakisphosphate 2-Kinase 1 (TaIPK1) reduces phytic acid and improves iron and zinc accumulation in wheat grains. J. Adv. Res. 37, 33–41. doi: 10.1016/j.jare.2021.07.006
Junior, R. M. (2017). Vegetable breeding as a strategy of biofortification in carotenoids and prevention of vitamin A deficiency. Afr. J. Agric. Res. 12, 1059–1066. doi: 10.5897/AJAR2016.11895
Kader, S., Chadalavada, S., Jaufer, L., Spalević, V., Dudić, B. (2022). Green roof substrates—A literature review. Front. Built Environ. 8, 1019362. doi: 10.3389/fbuil.2022.1019362
Kawaguchi, K., Takei-Hoshi, R., Yoshikawa, I., Nishida, K., Kobayashi, M., Kusano, M., et al. (2021). Functional disruption of cell wall invertase inhibitor by genome editing increases sugar content of tomato fruit without decrease fruit weight. Sci. Rep. 11, 21534.
Kim, M., Nguyen, T., Ahn, J. H., Kim, G., Sim, S. C. (2021). Genome-wide association study identifies QTL for eight fruit traits in cultivated tomato (Solanum lycopersicum L.). Horticult. Res. 8, 203. doi: 10.1038/s41438-021-00638-4
Kim, Y., Ahn, W. S., Cha, A., Jie, E. Y., Kim, S., Hwang, B., et al. (2022). Development of glucoraphanin-rich broccoli (Brassica oleracea var. italica) by CRISPR/Cas9-mediated DNA-free BolMYB28 editing. Plant Biotechnol. Rep. 16, 123–132. doi: 10.1007/s11816-021-00732-y
Krieger, U., Lippman, Z. B., Zamir, D. (2010). The flowering gene SINGLE FLOWER TRUSS drives heterosis for yield in tomato. Nat. Genet. 42, 459–463. doi: 10.1038/ng.550
Kwon, C., Heo, J., Lemmon, Z. H., Capua, Y., Hutton, S. F., Van Eck, J., et al. (2019). Rapid customization of Solanaceae fruit crops for urban agriculture. Nat. Biotechnol. 38, 182–188. doi: 10.1038/s41587-019-0361-2
Langemeyer, J., Madrid-López, C., Mendoza Beltran, A., Villalba Méndez, G. (2021). Urban agriculture — A necessary pathway towards urban resilience and global sustainability? Landscape Urban Plann. 210, 104055.
Li, H., Li, J., Song, J., Zhao, B., Guo, C., Wang, B., et al. (2019). An auxin signaling gene BnaA3.IAA7 contributes to improved plant architecture and yield heterosis in rapeseed. New Phytol. 222 (2), 837–851.
Liu, C. Q., Yao, X., Li, G., Huang, L., Liu, C., Xie, Z. (2022). Development of Novel Markers and Creation of Non-Anthocyanin and Anthocyanin-Rich Broccoli (Brassica oleracea var. italica) Cultivars. Appl. Sci. 12 (12), 6267. doi: 10.3390/app12126267
McCallum, J., Laing, W. A., Bulley, S. M., Thomson, S. J., Catanach, A., Shaw, M. L., et al. (2019). Molecular characterisation of a supergene conditioning super-high vitamin C in kiwifruit hybrids. Plants 8, 237. doi: 10.3390/plants8070237
Nguyen, C. D., Li, J., Mou, B., Gong, H., Huo, H., Ars (2021). A case study of using an efficient CRISPR/Cas9 system to develop variegated lettuce. Vegetable Res. 1 (4). doi: 10.48130/VR-2021-0004
Park, S. J., Jiang, K., Tal, L., Yichie, Y., Gar, O., Zamir, D., et al. (2014). Optimization of crop productivity in tomato using induced mutations in the florigen pathway. Nat. Genet. 46, 1337–1342. doi: 10.1038/ng.3131
Pnueli, L., Carmel-Goren, L., Hareven, D., Gutfinger, T., Alvarez, J., Ganal, M. W., et al. (1998). The SELF-PRUNING gene of tomato regulates vegetative to reproductive switching of sympodial meristems and is the ortholog of CEN and TFL1. Development 125 11, 1979–1989. doi: 10.1242/dev.125.11.1979
Prinzenberg, A. E., Schoot, H. V., Visser, R., Marcelis, L. F., Heuvelink, E., Schouten, H. J. (2021). Genetic mapping of the tomato quality traits brix and blossom-end rot under supplemental LED and HPS lighting conditions. Euphytica 217, 213.
Razifard, H., Ramos, A., Della Valle, A. L., Bodary, C., Goetz, E., Manser, E. J., et al. (2020). Genomic evidence for complex domestication history of the cultivated tomato in latin America. Mol. Biol. Evol. 37, 1118–1132. doi: 10.1093/molbev/msz297
Silva, V. D., Zorzeto, T. Q. (2019). Lettuce quality in greenhouse with different technological levels.
Song, Y., Duan, X., Wang, P., Li, X., Yuan, X., Wang, Z., et al. (2021). Comprehensive speed breeding: a high-throughput and rapid generation system for long-day crops. Plant Biotechnol. J. 20, 13–15. doi: 10.1111/pbi.13726
Soyk, S., Müller, N. A., Park, S. J., Schmalenbach, I., Jiang, K., Hayama, R., et al. (2016). Variation in the flowering gene SELF PRUNING 5G promotes day-neutrality and early yield in tomato. Nat. Genet. 49, 162–168. doi: 10.1038/ng.3733
Su, W., Tao, R., Liu, W., Yu, C., Yue, Z., He, S., et al. (2019). Characterization of four polymorphic genes controlling red leaf colour in lettuce that have undergone disruptive selection since domestication. Plant Biotechnol. J. 18, 479–490. doi: 10.1111/pbi.13213
Sun, B., Jiang, M., Zheng, H., Jian, Y., Huang, W., Yuan, Q., et al. (2020). Color-related chlorophyll and carotenoid concentrations of Chinese kale can be altered through CRISPR/Cas9 targeted editing of the carotenoid isomerase gene BoaCRTISO. Horticult. Res. 7, 161. doi: 10.1038/s41438-020-00379-w
Tikunov, Y., Meijer-Dekens, F. R., Molthoff, J. W., Paulo, J., Finkers, R., Capel, I. E., et al. (2020). The genetic and functional analysis of flavor in commercial tomato: the FLORAL4 gene underlies a QTL for floral aroma volatiles in tomato fruit. Plant J. 103, 1189–1204. doi: 10.1111/tpj.14795
Torii, K. U., Mitsukawa, N., Oosumi, T., Matsuura, Y., Yokoyama, R., Whittier, R. F., et al. (1996). The Arabidopsis ERECTA gene encodes a putative receptor protein kinase with extracellular leucine-rich repeats. Plant Cell 8, 735–746. doi: 10.1105/tpc.8.4.735
Tzin, V., Rogachev, I., Meir, S., Zvi, M. M., Masci, T., Vainstein, A., et al. (2015). Altered levels of aroma and volatiles by metabolic engineering of shikimate pathway genes in tomato fruits. AIMS Bioengin. 2 (2), 75–92. doi: 10.3934/bioeng.2015.2.75
Udayan, G., Sreedaya, G. S. (2018). Extent of Adoption of House terrace Cultivation of Urban dwellers of Thiruvananthapuram Corporation, Kerala. J. Ext. Educ. 30, 6008. doi: 10.26725/JEE.2018.1.30.6008-6013
Varkonyi-Gasic, E., Moss, S. M., Voogd, C., Wang, T., Putterill, J., Hellens, R. P. (2013). Homologs of FT, CEN and FD respond to developmental and environmental signals affecting growth and flowering in the perennial vine kiwifruit. New Phytol. 198 3, 732–746. doi: 10.1111/nph.12162
Varkonyi-Gasic, E., Wang, T., Voogd, C., Jeon, S., Drummond, R. S., Gleave, A. P., et al. (2018). Mutagenesis of kiwifruit CENTRORADIALIS-like genes transforms a climbing woody perennial with long juvenility and axillary flowering into a compact plant with rapid terminal flowering. Plant Biotechnol. J. 17, 869–880. doi: 10.1111/pbi.13021
Varshney, R. K., Sinha, P., Singh, V. K., Kumar, A., Zhang, Q., Bennetzen, J. L. (2020). 5Gs for crop genetic improvement. Curr. Opin. Plant Biol. 56, 190–196. doi: 10.1016/j.pbi.2019.12.004
Vlčko, T., Ohnoutková, L. (2019). Recent advances and perspectives in crop biofortification. Biol. Plant 63, 586–593. doi: 10.32615/bp.2019.056
Voogd, C., Brian, L. A., Wang, T., Allan, A. C., Varkonyi-Gasic, E. (2017). Three FT and multiple CEN and BFT genes regulate maturity, flowering, and vegetative phenology in kiwifruit. J. Exp. Bot. 68, 1539–1553. doi: 10.1093/jxb/erx044
Wang, B., Li, N., Huang, S., Hu, J., Wang, Q., Tang, Y., et al. (2021). Enhanced soluble sugar content in tomato fruit using CRISPR/Cas9-mediated SlINVINH1 and SlVPE5 gene editing. PeerJ 9:e12478. doi: 10.7717/peerj.12478
Wang, Y., Liu, X., Zheng, X., Wang, W., Yin, X., Liu, H., et al. (2021). Creation of aromatic maize by CRISPR/Cas. J. Integr. Plant Biol 63(9), 1664–1670. doi: 10.1111/jipb.13105
Wang, S., Wang, K., Li, Z., Li, Y., He, J., Li, H., et al. (2022). Architecture design of cucurbit crops for enhanced productivity by a natural allele. Nat. Plants 8, 1394–1407. doi: 10.1038/s41477-022-01297-6
Wang, Z., Wang, F., Yu, Z., Shi, X., Zhou, X., Wang, P., et al. (2022). Pyramiding of multiple genes generates rapeseed introgression lines with clubroot and herbicide resistance, high oleic acid content, and early maturity. Crop J 11(3), 895–903. doi: 10.1016/j.cj.2022.10.009
Wen, D., Wu, L., Wang, M., Yang, W., Wang, X., Ma, W. J., et al. (2022). CRISPR/cas9-mediated targeted mutagenesis of ftMYB45 promotes flavonoid biosynthesis in tartary buckwheat (Fagopyrum tataricum). Front. Plant Sci. 13. doi: 10.3389/fpls.2022.879390
Wen, C., Zhao, W., Liu, W., Yang, L., Wang, Y., Liu, X., et al. (2019). CsTFL1 inhibits determinate growth and terminal flower formation through interaction with CsNOT2a in cucumber. Development 146 (14), dev180166. doi: 10.1242/dev.180166
Xin, T., Tian, H., Ma, Y., Wang, S., Yang, L., Li, X., et al. (2022). Targeted creation of new mutants with compact plant architecture using CRISPR/Cas9 genome editing by an optimized genetic transformation procedure in cucurbit plants. Horticult. Res. 9, uhab086. doi: 10.1093/hr/uhab086
Xu, C., Liberatore, K. L., MacAlister, C. A., Huang, Z., Chu, Y. H., Jiang, K., et al. (2015). A cascade of arabinosyltransferases controls shoot meristem size in tomato. Nat. Genet. 47, 784–792. doi: 10.1038/ng.3309
Xu, Z., Yang, Q., Feng, K., Xiong, A. (2019). Changing carrot color: insertions in dcMYB7 alter the regulation of anthocyanin biosynthesis and modification1. Plant Physiol. 181, 195–207. doi: 10.1104/pp.19.00523
Yang, T., Ali, M., Lin, L., Li, P., He, H., Zhu, Q., et al. (2022). Recoloring tomato fruit by CRISPR/Cas9-mediated multiplex gene editing. Horticult. Res. 10, uhac214. doi: 10.1093/hr/uhac214
Ye, S., Hua, S., Ma, T., Ma, X., Chen, Y., Wu, L., et al. (2022). Genetic and multi-omics analysis reveal bnaA07.PAP2In-184-317 as the key gene conferring anthocyanin-based color in brassica napus flowers. J. Exp. Bot. 3 (19), 6630–6645. doi: 10.1093/jxb/erac312
Zeng, X., Yu, Y., Yang, S., Lv, Y., Sarker, M. N. (2022). Urban resilience for urban sustainability: concepts, dimensions, and perspectives. Sustainability 14 (5), 2481. doi: 10.3390/su14052481
Zhang, Y., Li, S., Xue, S., Yang, S., Huang, J., Wang, L. (2018). Phylogenetic and CRISPR/cas9 studies in deciphering the evolutionary trajectory and phenotypic impacts of rice ERECTA genes. Front. Plant Sci. 9. doi: 10.3389/fpls.2018.00473
Keywords: pre-breeding, genetic improvement, allele, terrace vegetables, resilience
Citation: Ma K, Yuan Y and Fang C (2023) Mainstreaming production and nutrient resilience of vegetable crops in megacities: pre-breeding for terrace cultivation. Front. Plant Sci. 14:1237099. doi: 10.3389/fpls.2023.1237099
Received: 08 June 2023; Accepted: 30 October 2023;
Published: 17 November 2023.
Edited by:
Bingyu Zhao, Virginia Tech, United StatesReviewed by:
S. R. Pandravada, Indian Council of Agricultural Research (ICAR), IndiaCopyright © 2023 Ma, Yuan and Fang. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Caochuang Fang, YmlvY2h6ZXVAZm94bWFpbC5jb20=
†ORCID: Caochuang Fang, orcid.org/0000-0001-5169-5672