 Freddy Ibanez1*†
Freddy Ibanez1*† Sâmara Vieira Rocha2*†
Sâmara Vieira Rocha2*† William O. Dawson3
William O. Dawson3 Choaa El-Mohtar3
Choaa El-Mohtar3 Cecile Robertson3
Cecile Robertson3 Lukasz L. Stelinski4
Lukasz L. Stelinski4 Andrea Soares-Costa3*
Andrea Soares-Costa3*- 1Department of Entomology, Texas A&M AgriLife Research, Weslaco, TX, ;United States
- 2Department of Genetics and Evolution, Federal University of São Carlos, São Carlos, SP, ;Brazil
- 3Plant Pathology Department, Citrus Research and Education Center, University of Florida, Lake Alfred, FL, ;United States
- 4Department of Entomology and Nematology, Citrus Research and Education Center, University of Florida, Lake Alfred, FL, ;United States
Diaphorina citri Kuwayama (Hemiptera: Liviidae) is a vector of the bacteria Candidatus Liberibacter americanus (CLam) and Candidatus Liberibacter asiaticus (CLas), which are phloem-restricted and associated with the most important and destructive worldwide citrus disease, Huanglongbing (HLB). Currently, no cure for HLB has been described. Therefore, measures have focused on reducing D. citri populations. In these insects, cathepsin B (DCcathB) and L (DCcathL) enzymes play an important role in digestion, and are involved in embryogenesis, immune defense, and ecdysis. In this study, we used a CTV-based vector to deliver dsRNA (CTV-dsRNA) into Citrus macrophylla plants targeting DCcathB and DCcathL genes in D. citri that fed on the phloem of these CTV-RNAi infected plants. Subsequently, we evaluated expression of DCcathB and DCcathL genes as well as the Vitellogenin (Vg) gene by RT-qPCR in D. citri fed on CTV-dsRNA occurring in plant phloem. It was found that a defective phenotype in D. citri females as a result of knockdown of DCcathB and DCcathL genes mediated by CTV dsRNA. These results showed that Psyllids fed on plants treated with the CTV-dsRNA exhibited downregulation of the Vg gene, one of the most important genes associated with embryogenic and female development, which was associated with dsRNA-mediated silencing of the two cathepsin genes. Based on our findings, a CTV-based strategy for delivering RNAi via plants that targets DCcathB and DCcathL genes may represent a suitable avenue for development of dsRNA-based tools to manage D. citri that limits the spread of HLB.
1 Introduction
The Asian Citrus Psyllid (ACP), Diaphorina citri Kuwayama (Hemiptera: Liviidae) is a vector of the phytopathogens Candidatus Liberibacter americanus (Clam) (Coletta-Filho et al., 2005; Teixeira et al., 2005a; Teixeira et al., 2005b; Teixeira et al., 2005c) and Candidatus Liberibacter asiaticus (CLas) (Garnier and Bové, 1996; Bové, 2006). Both are phloem-restricted, belong to the alpha-proteobacteria group (Jagoueix et al., 1994), and are associated with the most serious citrus disease in the world, Huanglongbing (HLB) (McClean and Schwartz, 1970; Bové, 2006). The symptoms of HLB are well characterized, including reduced vigor (Etxeberria et al., 2009), a decline in fruit production, reduced fruit size and quality, aborted seeds, fruit asymmetry, an increase in juice acidity and, in more severe stages, death of the trees (Bové, 2006; Etxeberria et al., 2009) resulting in a significant economic loss. The life cycle of the D. citri vector includes an egg stage, 5 nymphal instars, and an adult stage; both immature and adult stages are classified as phloem feeding and sap-sucking insects and the duration of each developmental stage is dependent of the host-species and temperature (Tsai and Liu, 2000). D. citri population abundance is closely linked to the occurrence of young citrus leaf flush, which is the only suitable site for egg laying and nymph development (Hall and Albrigo, 2007). Given the high osmotic pressure and sugar content present in plant phloem-sap, hemipterans possess sucrose-transglucosidases in the alimentary canal that allow them to tolerate and transform the excessive sugars ingested into long chains of oligosaccharides that are subsequently excreted as honeydew (Douglas, 2006).
Currently, there is no known cure for HLB, and most measures associated with management of the disease focus on reducing D. citri populations. Over the past decade, most D. citri management programs have heavily relied on insecticide sprays (Qureshi et al., 2014; Boina and Bloomquist, 2011). However, this has resulted in significantly higher production costs, and unintended ecological consequences, such as decreased populations of beneficial insects and development of insecticide resistance among D. citri populations (Tiwari et al., 2011; Farnsworthn et al., 2014). Therefore, alternatives to insecticides must be developed and integrated into vector suppression.
One strategy that has received significant attention in recent years as a possible alternative to traditional insecticides is RNA interference (RNAi) (Huvenne and Smagghe, 2010; Mamta and Rajam, 2017; Yan et al., 2020). The purpose of RNAi is to silence specifically targeted genes involved in critical biological processes in the pest, but which are sufficiently specific to minimize non-target consequences. For example, knockdown of arginine kinase, abnormal wing disc, carboxylesterases, muscle protein 20, cathepsin D, chitin synthase and sucrose hydrolase genes cause various physical deformities and mortality in D. citri and could theoretically reduce D. citri populations (El-Shesheny et al., 2013; Hajeri et al., 2014; Andrade and Hunter, 2017; Galdeano et al., 2017; Kishk et al., 2017; Yu et al., 2017; Santos-Ortega and Killiny, 2018). The RNAi mechanism is triggered by a double-stranded RNA molecule (dsRNA) precursor, of variable length and origin, which is excised by RNase-III-like endonuclease (Dicer) into short ~20-30 nucleotides RNAs duplexes, called short interfering RNAs (siRNAs). The siRNAs are recognized by the RNA-induced silencing complex (RISC) and then this complex recognizes the expected target mRNA (single-strand), cleaving the mRNA molecule and thereby hindering the normal translational process (Elbashir et al., 2001; Hammond et al., 2001; Meister and Tuschl, 2004; Whyard et al., 2009). In agricultural systems, the potential implementation of RNAi delivery is based on transgenic and non-transgenic strategies. Among non-transgenic strategies to deliver RNAi, the use of Virus-induced gene silencing (VIGS) has been explored (Bachan and Dinesh-Kumar, 2012; Wuriyanghan and Falk, 2013; Hajeri et al., 2014; Khan et al., 2015). This strategy uses an engineered plant virus to deliver a targeted insect gene-sequence (dsRNA/siRNAs) during virus replication and/or transcription in the plant hosts (Bachan and Dinesh-Kumar, 2012). Upon feeding on a host colonized with the virus vector delivering the dsRNA/siRNA, the target pest ingests them leading to an inhibition of translation of the targeted genes within the insect (Hajeri et al., 2014; Khan et al., 2018).
In citrus, virus-based vectors have been created through the manipulation of Citrus tristeza virus (CTV) for heterologous gene expression (Folimonov et al., 2007; El-Mohtar and Dawson, 2014). CTV is a member of the Closterovirus genus within the family Closteroviridae; it possesses a single stranded RNA genome of ~19.3 kb (Bar-Joseph et al., 1979; Karasev et al., 1995), and it is considered the largest and most complex plant RNA virus with phloem limited tissue tropism where ACP feeds. The CTV-RNAi vectors were successfully used to target citrus and ACP genes (Hajeri et al., 2014). For example, engineering the CTV virus vector to deliver a 400 base, truncated RNA has been successfully demonstrated in citrus, by targeting the phytoene desaturase (PDS) gene, which resulted in photo-bleaching of citrus leaves upon infection with the CTV vector. Also, a CTV-RNAi vector delivering a truncated version of the Awd gene of D. citri resulted in reduced expression of the Awd gene upon ACP feeding on infected plants causing malformation of D. citri wings and high mortality of adults (Hajeri et al., 2014). This validates the use of CTV vectors for delivery of dsRNAs/siRNAs via citrus to target D. citri genes.
In our previous studies, two cathepsin genes were identified in D. citri, cathepsin B-like (DcCathB) (Ferrara et al., 2015) and cathepsin L-like (DcCathL) (Ferrara et al., 2020). Cathepsins are proteolytic cysteine-peptidase type enzymes (Storer and Ménard, 1994; Polgár, 2013; Rawlings et al., 2016), belonging to the family C1 and subfamily papain; cathepsins are proteases that occur broadly among animals and play important roles in immunity, metabolism, and development (Sajid and McKerrow, 2002; Wiederanders, 2003). In insects, cathepsins B and L play a key role in digestion and are involved in embryogenesis, immune defense, and ecdysis (Terra and Ferreira, 1994). In D. citri, DcCathB and DcCathL were highly expressed in the alimentary canal suggesting a role in digestion and were expressed during the egg stage suggesting involvement in embryonic development (Ferrara et al., 2020). We hypothesized that blocking activity of these enzymes using the recombinant CTV-RNAi vectors should disturb critical biological processes in D. citri. In this study, we evaluated the effects of RNAi targeting DcCathB and DcCathL genes in D. citri. The RNAi were constructed and delivered via modified CTV vectors. Our results showed a significant downregulation of both Cathepsin genes in ACP adults. Additionally, we examined the effects of these RNAi on ACP female reproduction, and our results indicated coincident downregulation of the Vitellogenin A1-like gene, which was associated with significant reduction in ovarial development among D. citri females feeding on RNAi-treated plants infected with modified CTV vectors.
2 Materials and methods
2.1 Plants and insects
Nicotiana benthamiana plants were maintained in a growth-room under controlled conditions set at 22–24°C, 16:8 h photoperiod 60% relative humidity (RH). One year old Citrus macrophylla seedlings were maintained under controlled greenhouse conditions of 23 ± 3°C, 60 RH, and a 16:8 h (Light: Dark) photoperiod with a maximum photosynthetic radiation of 215 µmol s-1 m-2. Plants were watered twice per week and fertilized twice per month with an alternating schedule of a 24-8-16 NPK solution at 4 g L-1 (Miracle-Gro All Purpose Plant Food; Scotts Miracle-Gro Products, Marysville, OH) and a 6-4-6 (N–P–K) granular fertilizer at 1 g per pot (Expert gardener Gro Tec. Inc. Madison, GA).
A laboratory population of D. citri with known susceptibility to insecticides was reared in a greenhouse at the Citrus Research and Education Center, University of Florida, Lake Alfred, FL. This population originated from adults collected in 2000 from citrus in Polk City, Polk County, FL and has been reared without exposure to insecticides. The culture was maintained on sweet orange (Citrus sinensis Osbeck) in a greenhouse at approximately 27 °C, with 60% relative humidity, and a 14:10 (light: dark) photoperiod.
2.2 Citrus tristeza virus (CTV) based vectors
The infectious cDNA clone of Citrus tristeza virus (CTV isolate T36; GenBank accession no. AY170468) in the binary vector pCAMBIA-1380 was used for engineering all the constructs used in this study (Satyanarayana et al., 1999; Satyanarayana et al., 2001; Gowda et al., 2005; El-Mohtar and Dawson, 2014). To clone Cathepsin B and Cathepsin L fragments and generate the corresponding CTV vector, primers were designed based on D. citri genes (GenBank accession no. NM_001329180 and MN166228.1) (Table 1). The fragments corresponding to each D. citri gene were amplified using 2 µg of total RNA as template using the Super Script®III One-Step RT-PCR System with Platinum®Taq DNA Polymerase (Life Technologies Corp.), 100 nM of each primer (Table 1) in a final volume of 100 µL adjusted with nuclease-free water (Thermo fisher). The PCR products of each gene were digested with PacI and StuI restriction enzymes and ligated into their corresponding digested CTV vector. CTV binary plasmid vectors were screened by restriction digestion and confirmed by sanger sequencing (Psomagen, Inc, USA).

Table 1 Sequence of specific primers for the DcCathL and DcCathB genes of D. citri.
2.3 Agroinfiltration and CTV virion isolation and inoculation to citrus
The CTV constructs dsRNA-DcCathB and dsRNA-DcCathL were used to transform Agrobacterium tumefaciens cells (EHA 105) and then, the transformant colonies were selected on LB plates containing the antibiotics Rifampicin (50 µg/ml) and Kanamycin (50 µg/ml). The transformant colonies were grown in LB liquid medium with Kanamycin (50 µg/ml), supplemented with 10 mM MES (pH 5.8) and 20 mM acetosyringone. This culture was used to prepare a agrobacteria cellular suspension containing the target constructs and infiltrated in the abaxial side of N. benthamiana leaves following the protocol described previously (Gowda et al., 2005; Ambrós et al., 2011). Systemic leaves from N. benthamiana that assessed positive for CTV by ELISA were harvested after 8 weeks post infiltration and used to isolate recombinant CTV virions by concentration over a sucrose cushion (Robertson et al., 2005). Thus, the solution containing the purified recombinant virions were used for bark-flap inoculation of C. macrophylla as described previously (Robertson et al., 2005).
2.4 Insect bioassays
Adult D. citri were harvested from the main culture described above and isolated as group of 50 females and 50 males in cages with four 1-year old M. paniculata plants for 10 days to allow egg laying. Then, adult insects were removed, and development of nymphs was tracked daily. On the day adults emerged, insects were separated by sex and color morph (only green/blue color morph insects were used in this study) and then transferred to 2-year-old C. macrophylla plants for 24 hours. On the second day after adult emergence, forty green/blue females and males per biological replicate were relocated onto 1-year old C. macrophylla seedlings (approximately two feet tall and stem 1 cm of diameter) colonized independently with CTV-DcCathB and -DcCathL RNAi vectors. The negative control used for experiments was Citrus macrophylla seedling infected with CTV-wt vector. For gene expression analysis, groups of 10 female and 10 males D. citri adults were collected 5 and 10 days after assays were initiated per biological replicate (N= 4 replicates). Similarly for analysis of ovary morphology, ten females per replicate were collected at day 10 and ovaries were dissected as previously described in Ibanez et al. (2019). In total four independent replicates were conducted per assay.
2.5 RNA extraction and cDNA synthesis
The total RNA for each pool of 10 insects per biological replicate was extracted after sample homogenization at room temperature with a plastic pestle using 500 µL of TRIzol reagent (Thermo Fisher scientific, MA) following the manufacturer’s instructions. Traces of genomic DNA were removed by DNase treatment using the TURBO DNA-free kit (Thermo Fisher scientific). The total RNA quantity and purity was evaluated on a Nanodrop Spectrophotometer (Thermo fisher Scientific), and the integrity of the RNA was visualized in 1.2% agarose gel electrophoresis. The synthesis of cDNA was performed using the Verso cDNA Synthesis kit (Thermo fisher Scientific). Each retro-transcription synthesis was conducted using five hundred ng of total RNA as template and adding anchored-Oligo (dT) primers, following manufacturer recommendations (Thermo Fisher scientific).
2.6 Gene expression analyses
Quantitative PCR (qPCR) reactions were performed using PowerUp SYBR Green Master Mix (Applied Biosystems). Each qPCR reaction contained 10 ng of cDNA as template, 300 nM of each primer (Table 2) and 1x of PowerUp SYBR Green Master Mix; in a final volume of 10 µL adjusted with nuclease-free water (Thermo fisher). The PCR program consisted of 2 min 50°C for UDG activation, 2 min 95°C for Dual-Lock DNA polymerase followed by 40 cycles at 95°C for 15 sec and 60°C for 1 min. Primers for each target gene were designed using Primer3 web (Untergasser et al., 2012). Each qPCR reaction was performed in duplicates with a negative control in each run using an Applied Biosystems ABI 7500 real-time PCR thermocycler (Applied Biosystems). The amplification specificity of each PCR product was monitored using the melting curve analysis in Sequence detection system (SDS) version 1.4.0.27 (Applied Biosystems) and visualizing the PCR products in a 2% agarose gel. The relative gene expression was estimated by the 2-ΔΔCT method (Livak and Schmittgen, 2001), using β-actin as a reference gene (GenBank XM_026821238.1) as was previously described (Hajeri et al., 2014).
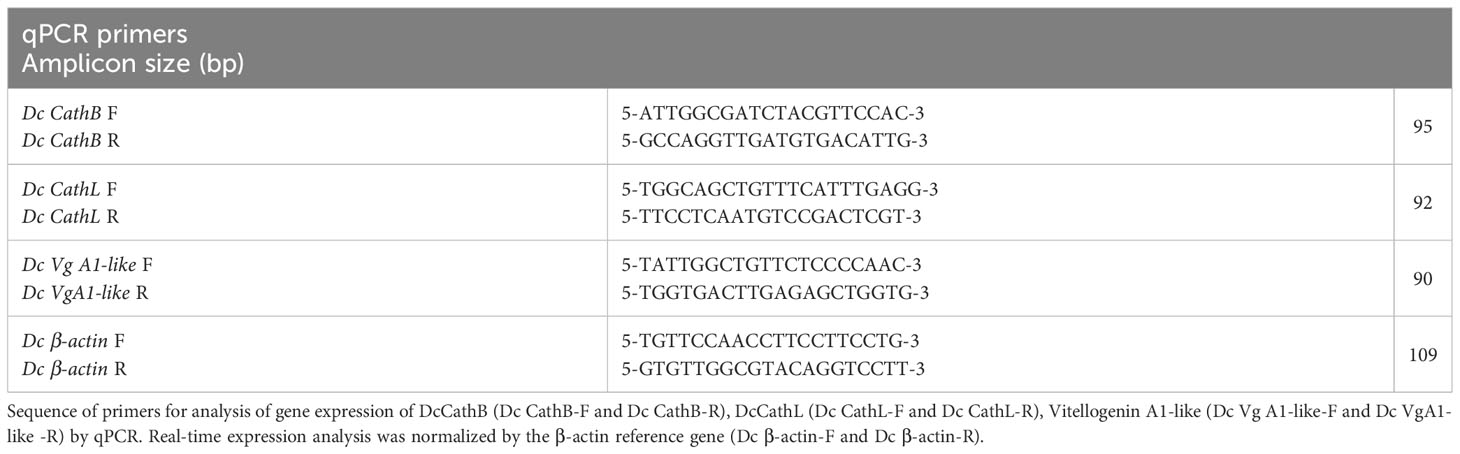
Table 2 Sequence of qPCR primers for D. citri genes.
2.7 Statistical analysis
To examine the effects of each CTV construct in D. citri, one-way analysis of variance (ANOVA) with Tukey’s post hoc test was used to compare mean gene expression and numbers of mature oocytes between treatments. All statistical analyses were performed using RStudio environment (RStudio Team, 2015).
3 Results
3.1 CTV-based citrus plant-mediated RNAi suppressed expression of DcCathB and DcCathL in D. citri adult
Expression of the DcCathB transcript in female D. citri was significantly reduced (F = 19.325, df = 4, P < 0.001) after 5 and 10 days of feeding on plants colonized with CTV RNAi vectors targeting DcCathB. DcCathB expression was reduced by 79% at day 5 (P = 0.043), and 95% after 10 days (P < 0.001) of feeding on plants expressing dsRNA- DcCathB, compared to control females at the same time points (Figure 1A). In D. citri males, the DcCathB transcript was also significantly reduced (F = 15.618, df = 4, P < 0.001). Pairwise comparisons at day 5 showed a 52% reduction in DcCathB expression (P = 0.008), while at day 10, there was no significant difference (P = 0.98) compared to male controls at the same time points (Figure 1B).
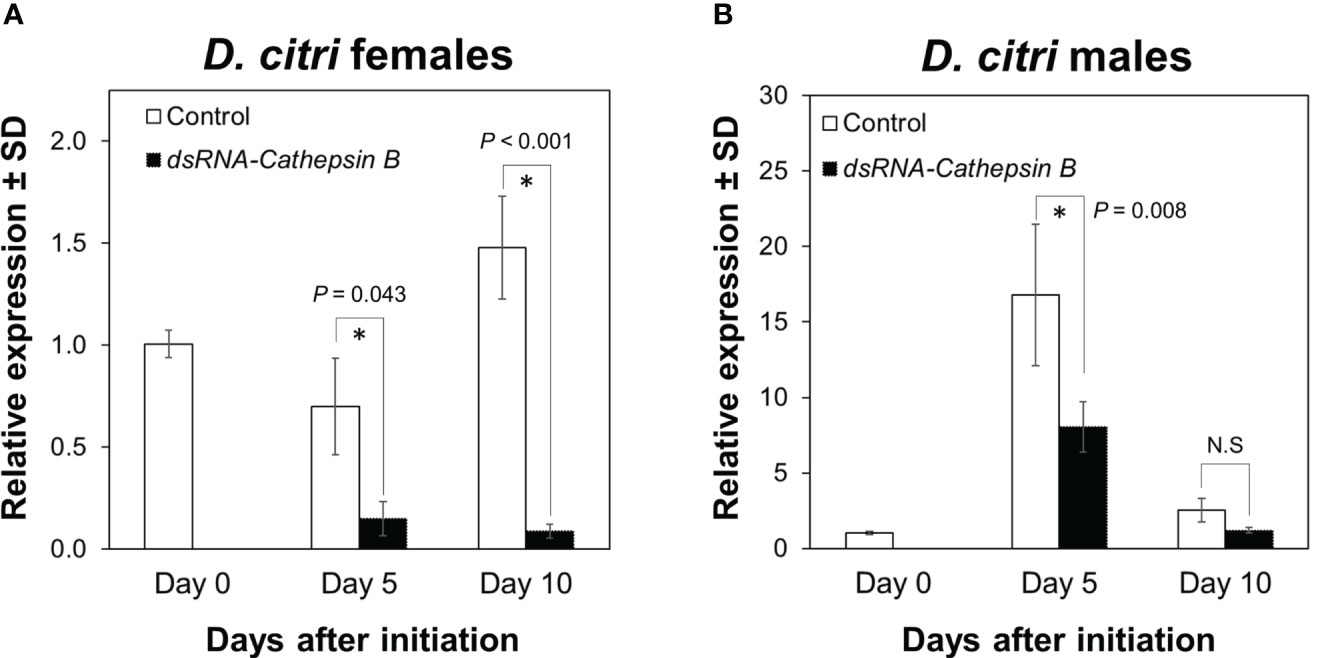
Figure 1 Expression of Cathepsin B in Diaphorina citri adults. The temporal pattern of Cathepsin B transcript relative expression in D. citri females (A) and males (B) was normalized to the expression value of the β-actin transcript. Data represent means ± standard deviations (SD) of 4 independent replicates. Asterisks indicate statistical differences between gene expression in psyllids fed on dsRNA-DCcathB or control plants using one-way ANOVA with Tukey’s post hoc test. N.S denotes no significance.
Similarly, exposure of female and male D. citri for 5 and 10 days of feeding on plants colonized by CTV RNAi vectors targeting DcCathL led to significantly reduced expression of the DcCathL transcript (F = 181.585, df = 4, P < 0.001). DcCathL expression was significantly reduced by 83% (P = 0.005) and 77% (P < 0.001) at 5 and 10 days after feeding was initiated, respectively, compared with control females at the same time points (Figure 2A). In D. citri males, expression of the DcCathL transcript was also significantly reduced (F = 9.510, df = 4, P < 0.001). Pairwise comparisons between controls and RNAi treatments at day 5 showed no significant change in DcCathL expression (P = 0.43), while at day 10, the DcCathL transcript was significantly reduced (P = 0.0005) by 96% compared to male controls at the same time point (Figure 2B).
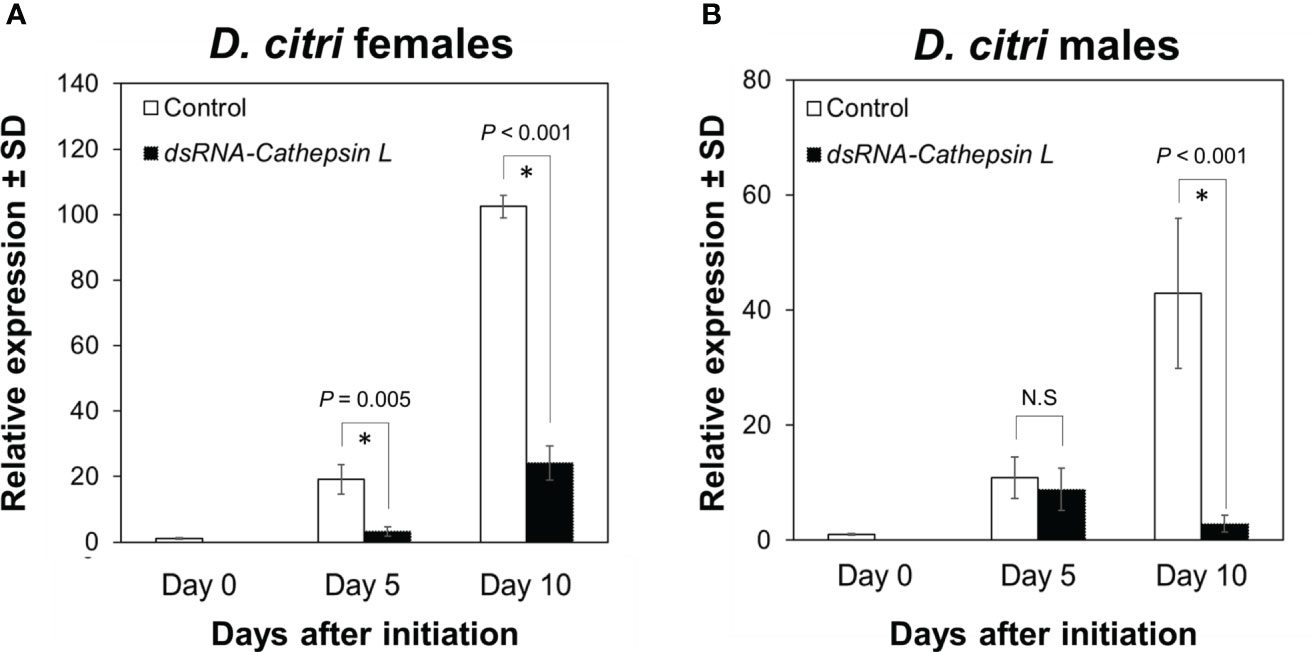
Figure 2 Expression of Cathepsin L in Diaphorina citri adults. The temporal pattern of Cathepsin L transcript relative expression in D. citri females (A) and males (B) was normalized to the expression value of the β-actin transcript. Data represent means ± standard deviations (SD) of 4 independent replicates. Asterisks indicate statistical differences between gene expression in psyllids fed on dsRNA- DcCathL or control plants using one-way ANOVA with Tukey’s post hoc test. N.S denotes no significance.
3.2 CTV-based citrus plant-mediated RNAi (DCcathB and DCcathL) suppresses expression of Vitellogenin A1-like and reduces ovary maturation in D. citri females
To evaluate if knockdown of Cathepsin expression interferes with D. citri reproduction, the expression of Vg A1-like and number of developing oocytes in females feeding on citrus plants colonized with CTV-based RNAi vectors independently targeting DcCathB or DcCathL were analyzed using pairwise comparisons. The relative expression of Vg A1-like was significantly (F = 51.542, df = 6, P < 0.001) downregulated in both treatments as compared with expression observed in controls after 10, but not 5 (P > 0.05), days of feeding (Figure 3A). The significant difference in Vg A1-like expression at day 10 prompted us to dissect the reproductive organs of female D. citri feeding on CTV-based RNAi colonized citrus (DcCathB or DcCathL) and compare them versus those feeding on control plants (Figures 3B, C). Significantly more developing oocytes were counted in the ovaries of control D. citri females than those fed on plants colonized with CTV-RNAi vectors independently targeting DcCathB (P = 0.002) or DcCathL (P = 0.005) (Figure 3B).
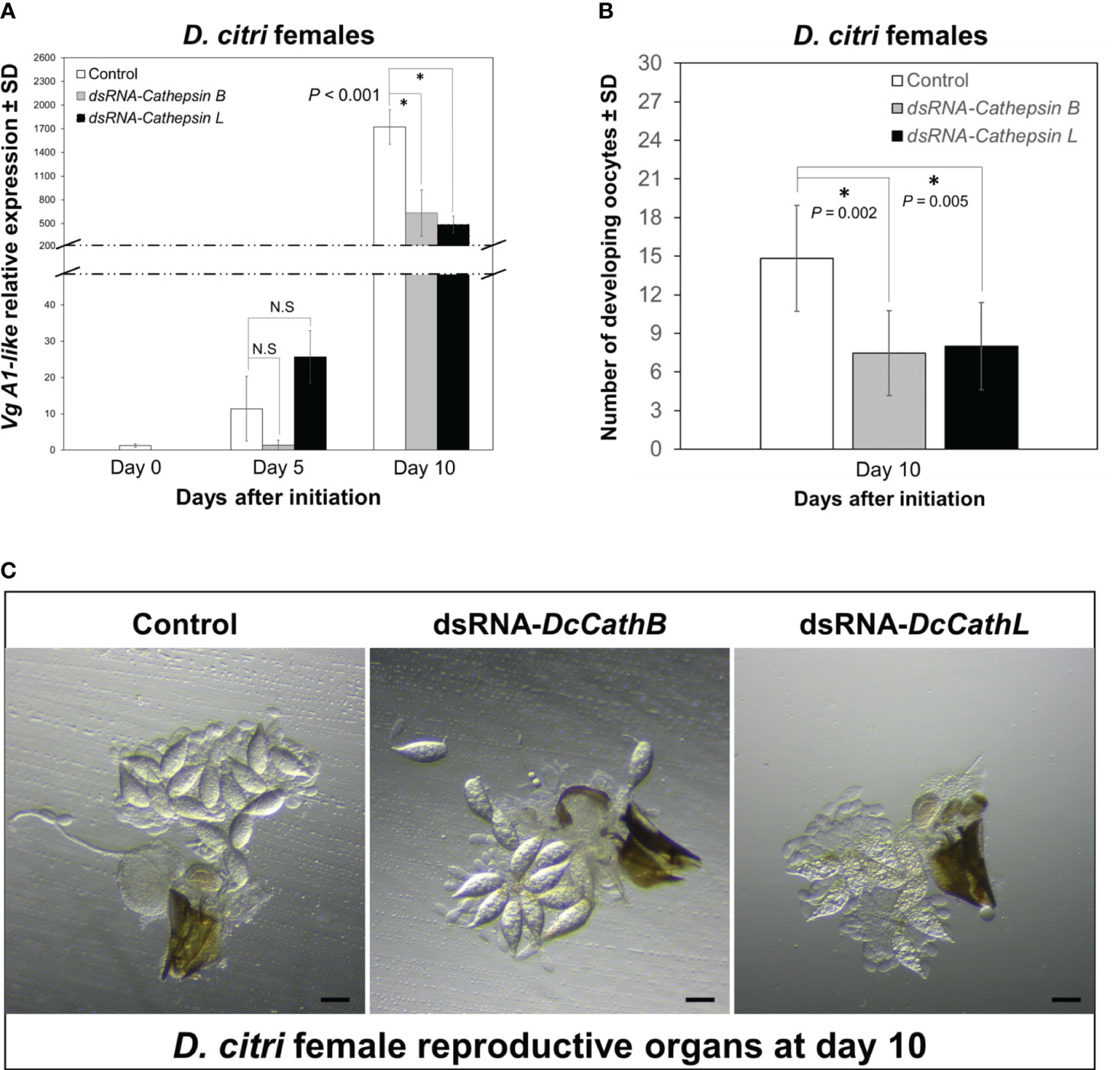
Figure 3 Effects of CTV-based RNAi (DCcathB and DCcathL) via expression in citrus plants on D. citri female reproductive development. (A) Temporal pattern of Vg1-like transcript relative expression levels in D. citri females normalized to the expression value of the β-actin transcript. Data represent means ± standard deviations (SD) of 4 independent replicates. Asterisks indicate statistical differences between gene expression in psyllids fed on plants expressing CTV-based dsRNAs (DcCathB and DcCathL) or control plants using one-way ANOVA with Tukey’s post hoc test. N.S denotes no significance. (B) Number of developing oocytes detected in D citri females after 10 days of challenge to plants expressing CTV-based dsRNAs or control plants. Data represents means ± SD (standard deviation) of at least thirty dissected females per group. (C) Representative images of ovaries from D. citri females. Scale bar is equal to 100 µm.
4 Discussion
Cathepsin B and cathepsin L enzymes in insects are associated with various biological processes including protein turnover, embryogenesis, mobilization, degradation of yolk proteins (Medina et al., 1988; Izumi et al., 1994; Cho et al., 1999; Zhao et al., 2002), metamorphosis (Takahashi et al., 1993; Shiba et al., 2001; Lee et al., 2009), and programmed cell death (Yang et al., 2020). We hypothesized that cathepsin enzymes should be useful targets for RNAi given their broad importance for D. citri development. Given the current lack of therapeutic treatments targeting the CLas pathogen, emphasis has been placed on reducing populations of the D. citri vector (Hajeri et al., 2014).
RNAi has been demonstrated on target insects for cathepsin genes using transgenic plants (Huang et al., 2006; Galdeano et al., 2017). In the present investigation, we utilized CTV-RNAi to target the cathepsin genes in D. citri. Delivery of dsRNA in citrus plants against a phloem feeding insect, such as D. citri, using CTV-based vectors offers advantages over the traditional transgenic approach in citrus. Engineering the virus is relatively easier and faster than producing transgenic plants. The engineered virus could colonize the mother plants from which budwood is used to generate citrus trees for establishing new grooves. Further, the CTV virus vector could be graft transmitted to current plants at any stage, including fully grown plants in the ground, obviating the need to remove the current generation of plants. However, despite the stability of CTV RNAi vectors for many years, at some stage, the engineered virus will recombine back into the wild type of the virus (El-Mohtar and Dawson, 2014). During this time, we expect that ongoing research may provide the ultimate solution for HLB via engineering a resistant/tolerant plant via biotechnology or traditional breeding.
DcCathB gene expression occurs in the D. citri gut of both nymph and adult stages, suggesting a possible role in digestion (Ferrara et al., 2015). Additionally, downregulation of DcCathB occurs in the presence of the CLas bacterium of both nymphs and adults, suggesting a possible role in immune response (Vyas et al., 2015). A different pattern of DcCathL expression has been reported compared to DcCathB, with expression observed in the head, thorax, and gut tissues of D. citri, suggesting function in more processes than digestion. Greater expression of the DcCathL gene in eggs than nymph and adult stages also suggests a role of in embryonic development (Ferrara et al., 2020). Additionally, DcCathL has been implicated in the innate immune response, like DcCathB (Yu et al., 2019). The preponderance of evidence collected to date would suggest that both DcCathB and DcCathL play roles associated with development in D. citri.
Our results provide evidence that DcCathB and DcCathL are associated with female reproductive development in D. citri (Figure 3). Following knockdown of either DcCathB (Figure 1) or DcCathL (Figure 2), oocyte development was significantly compromised (Figure 3B) and resulted in an abnormal ovary phenotype (Figure 3C). Furthermore, maturation of ovaries was delayed, which appeared related to interference with vitellogenesis (Figure 3). It is possible cathepsin gene regulation plays a role in vitellogenesis at some stage given the observed downregulation of the Vitellogenin A1-like gene coincident with downregulation of DcCathB or DcCathL (Figure 3).
Vitellogenesis is crucial for insect embryonic development and egg production (Wu et al., 2021), and alterations in Vitellogenin (Vg) and associated genes can significantly interfere with normal development. For example, interference with Vg and the Vitellogenin receptor (VgR) hinder vitellogenin accumulation and resultant egg maturation (Ibanez et al., 2017; Peng et al., 2020). Similarly, in locusts, reduced expression of Vg gene resulted in arrested oocyte maturation and impaired egg production (Guo et al., 2014; Wu et al., 2016; Wu et al., 2018). In D. citri, knockdown of Vitellogenin-4 like (Vg-4 like) and VgR genes decreases the number of mature eggs in the ovary and causes phenotypic abnormalities in eggs and nymphs (Li et al., 2022). Also, RNAi knockdown of the VgR gene in Cadra cautella resulted in low fecundity and fertility (Husain et al., 2020). Cathepsin L (Mn-CTS L1) has been reported to play an important role in the ovarian development of the oriental river prawn, Macrobrachium nipponense. Interestingly, it was hypothesized that Mn-CTS L1 silencing can effectively inhibit Vg hydrolysis, thereby reducing the availability of energy required for ovarian development (Zhu et al., 2021). Cathepsin B has been shown to play a role in embryogenesis and egg development in the hard tick, Haemaphysalis longicornis (Zhang et al., 2019). Accordingly, we could observe a downregulation of Vg A1-like in those insects that fed on plants expressing dsRNA-DcCaths (Figure 4). In addition, Cathepsins have been related to embryogenesis in insects as well the Vg hydrolysis (Moura et al., 2015; Zhang et al., 2019), for instance resulting in a reduction in developing oocytes and compromising ovarian maturation (Figure 3).
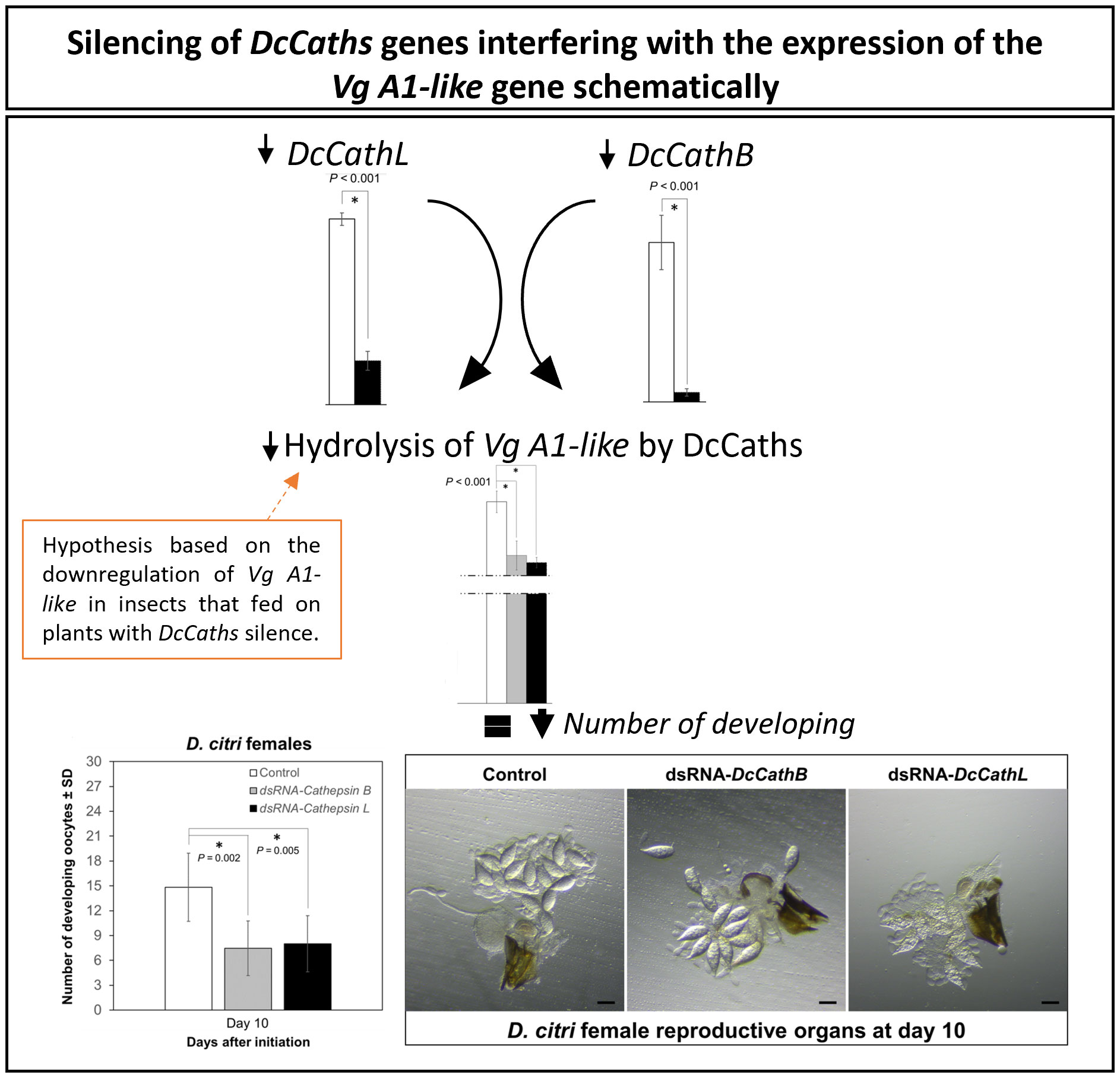
Figure 4 Hypothesis of Vitellogenin (Vg) and Cathepsins B and L-like (DcCathB and DcCathL) relationship. Vitellogenin (Vg) downregulated regarding DcCathB and DcCathL gene silencing resulting in decreased number of developing oocytes as well an ovarian development compromised.
However, how cathepsin and vitellogenin genes interact and why knockdown of DcCathB or DcCathL both appeared to coincidentally reduce vitellogenesis remain open questions.
Novel methodologies such as RNAi must be considered to mitigate economic and environmental damage of pathogen vectors (Zhang et al., 2013). We demonstrate here the silencing of cathepsin genes in D. citri by delivery of RNAi inducers via an engineered CTV vector colonizing citrus phloem tissue. Given that CTV accumulates and produces large amounts of dsRNA in the phloem, which is the sole source of nutrition for developing D. citri (Hajeri et al., 2014), use of CTV-based RNAi vectors is being explored as possible management tools for this phytopathogen vector. Our results indicate that reducing cathepsin gene expression causes visible phenotypic changes in D. citri, delaying and reducing ovarial development which appears related to a downregulation of vitellogenin expression. It will be important to determine whether reduced ovarial development is associated with changes to the fecundity and fertility of D citri (Bueno et al., 2020; Koji et al., 2020). We speculate that decreased ovarial development caused by silencing of cathepsin genes may result in reduced population growth of D. citri. This hypothesis will require further testing. The remarkable stability of the CTV vector, its effectiveness in delivering dsRNA into the plant phloem, and relatively fast activity further support its use for validation of biological targets and development into practical therapeutic tools. Our findings also indicate that Cathepsin B and L-like are promising target sites that should be further explored as a means of managing the D. citri vector. This research represents an initial step that will require scaling up to population-wide treatment of psyllids under authentic field conditions. Further biological testing of the consequences of cathepsin silencing in D. citri are needed to determine potential practical utility for population suppression of this phytopathogen vector, including effects on female fecundity and fertility.
Data availability statement
The datasets presented in this study can be found in online repositories. The names of the repository/repositories and accession number(s) can be found in the article/supplementary material.
Author contributions
AS-C, FI-C, CEM, CR, WD and LS contributed to conception and design of the study. AS-C, SVR and FI-C wrote sections of the manuscript. All authors contributed to the article and approved the submitted version.
Funding
This work was supported by the grants #2016/04305-9 and #2018/20823-5, São Paulo Research Foundation (FAPESP).
Acknowledgments
This paper and the research behind it would not have been possible without the exceptional support of Citrus Research and Education Center (CREC), at University of Florida and Department of Genetics and Evolution, at Federal University of Sao Carlos (UFSCar).
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Ambrós, S., El-Mohtar, C., Ruiz-Ruiz, S., Peña, L., Guerri, J., Dawson, W. O., et al. (2011). Agroinoculation of Citrus tristeza virus causes systemic infection and symptoms in the presumed nonhost Nicotiana benthamiana. Mol. Plant-Microbe Interact. 24 (10), 1119. doi: 10.1094/MPMI-05-11-0110
Andrade, E. C., Hunter, W. B. (2017). RNAi feeding bioassay: development of a non- transgenic approach to control Asian citrus psyllid and other hemipterans. Entomol. Exp. Appl. 162, 389–396. doi: 10.1111/eea.12544
Bachan, S., Dinesh-Kumar, S. P. (2012). “Tobacco Rattle Virus (TRV)-Based Virus-Induced Gene Silencing,” in Antiviral Resistance in Plants. Methods in Molecular Biology (Methods and Protocols). Eds. Watson, J., Wang, M. B. (Totowa, NJ: Humana Press), 894. doi: 10.1007/978-1-61779-882-5_6
Bar-Joseph, M., Garnsey, S. M., Gonsalves, D. (1979). The closteroviruses. A distinct group of elongated plant viruses. Adv. Virus Res. 25, 93–168. doi: 10.1016/S0065-3527(08)60569-2
Boina, D. R., Bloomquist, J. R. (2011). Chemical control of the Asian citrus psyllid and of huanglongbing disease in citrus. Pest Manage. Sci. 71, 808–823. doi: 10.1371/journal.pone.0112331
Bové, J. M. (2006). Huanglongbing: A destructive, newly-emerging, century-old disease of citrus. J. Plant Pathol. 88 (1), 7–37.
Bueno, F. G. B., Gloag, R., Latty, T., Ronai, I. (2020). Irreversible sterility of 392 workers and high-volume egg production by queens in the stingless bee 393 Tetragonula carbonaria. J. Exp. Biol. 223, jeb230599. doi: 10.1242/jeb.230599
Cho, W. L., Tsao, S. M., Hays, A. R., Walter, R., Chen, J. S., Snigirevskaya, E. S., et al. (1999). Mosquito cathepsin B-like protease involved in embryonic degradation of vitellin is produced as a latent extraovarian precursor. J. Biol. Chem. 274 (19), 13311–13321. doi: 10.1074/jbc.274.19.13311
Coletta-Filho, H. D., Targon, M. L. P. N., Takita, M. A., De Negri, J. D., Pompeu, J., Jr, MaChado, M. A. (2005). Analysis of 16S rDNA sequences from citrus huanglongbing bacteria reveal a different “Ca. Liberibacter” Strain associated with citrus disease in São Paulo. Plant Dis. 89 (8), 848–852. doi: 10.1094/PD-89-0848
Douglas, A. E. (2006). Phloem-sap feeding by animals: problems and solutions. J. Exp. Bot. 57 (4), 747–754. doi: 10.1093/jxb/erj067
Elbashir, S. M., Lendeckel, W., Tuschl, T. (2001). RNA interference is mediated by 21- and 22-nucleotide RNAs. Genes Develop. 12, 188–200. doi: 10.1101/gad.862301
El-Mohtar, C., Dawson, W. O. (2014). Exploring the limits of vector construction based on Citrus Tristeza virus. Virology 448, 274–283. doi: 10.1016/j.virol.2013.10.017
El-Shesheny, I., Hajeri, S., El-Hawary, I., Gowda, S., Killiny, N. (2013). Silencing abnormal wing disc gene of the Asian Citrus Psyllid, Diaphorina citri disrupts adult wing development and increases nymph mortality. PloS One 8, 5. doi: 10.1371/journal.pone.0065392
Etxeberria, E., Gonzalez, P., Achor, D., Albrigo, G. (2009). Anatomical distribution of abnormally high levels of starch in HLB-affected Valencia orange trees. Physiol. Mol. Plant Pathol. 74, 76–83. doi: 10.1016/j.pmpp.2009.09.004
Farnsworthn, D., Grogan, K. A., van Bruggen, A. H. C., Moss, C. B. (2014). The potential economic cost and response to Greening in Florida citrus. Choices 29, 1–6.
Ferrara, T. F., Schneider, V. K., Kishi, L. T., Carmona, A. K., Alves, M. F. M., Belasque-Júnior, J., et al. (2015). Cathepsin B-like cysteine peptidase from Diaphorina citri Kuwayama (Hemiptera: Liviidae): a putative target for control of citrus huanglongbing. PloS One 10, 12. doi: 10.1371/journal.pone.0145132
Ferrara, T. F. D., Shneider, V. K., Lima, P. S., Bronze, F. S., MaChado, M. F. M., Henrique-Silva, F., et al. (2020). Gene expression. Studies and molecular characterization of a cathepsin L-like from the Asian citrus psyllid Diaphorina citri, vector of Huanglongbing. Int. J. Biol. Macromol. 158, 375–383. doi: 10.1016/j.ijbiomac.2020.04.070
Folimonov, A. S., Folimonova, S. Y., Bar-Joseph, M., Dawson, W. O. (2007). A stable virus-based vector for citrus trees. Virology 368 (1), 205–216. doi: 10.1016/j.virol.2007.06.038
Galdeano, D. M., Breton, M. C., Lopes, J. R. S., Falk, B. W., MaChado, M. A. (2017). Oral delivery of double-stranded RNAs induces mortality in nymphs and adults of the Asian citrus psyllid, Diaphorina citri. PloS One 12 (3), e0171847. doi: 10.1371/journal.pone.0171847
Garnier, M., Bové, J. M. (1996). “Distribution of the Huanglongbing (Greening) Liberibacter species in fifteen African and Asian countries,” in Conference of the International Organization of Citrus Virologists. Proceedings of 13th Conference IOCV, Riverside: University of California. 388–391. doi: 10.5070/C58v467619
Gowda, S., Satyanarayana, T., Robertson, C. J., Garsey, S. M., Dawson, W. O. (2005). “Infection of citrus plants with virions generated in Nicotiana benthamiana plants agroinfiltrated with a binary vector based Citrus tristeza virus.,” in Sixteenth IOCV Conference. doi: 10.5070/C59cw9m5cd
Guo, W., Wu, Z., Song, J., Jiang, F., Wang, Z., Deng, S., et al. (2014). Juvenile hormone-receptor complex acts on mcm4 and mcm7 to promote polyploidy and vitellogenesis in the migratory locust. PloS Genet. 10, e1004702. doi: 10.1371/journal.pgen.1004702
Hajeri, S., Killiny, N., El-Mohtar, C., Dawson, W. O., Gowda, S. (2014). Citrus tristeza virus-based RNAi in citrus plants induces gene silencing in Diaphorina citri, a phloem-sap sucking insect vector of citrus greening disease (Huanglongbing). J. Biotechnol. 176, 42–49. doi: 10.1016/j.jbiotec.2014.02.010
Hall, D. G., Albrigo, L. G. (2007). Estimating the relative abundance of flush shoots in citrus, with implications on monitoring insects associated with flush. HortScience 42, 364–368. doi: 10.21273/HORTSCI.42.2.364
Hammond, S. M., Boettcher, S., Caudy, A. M. A., Kobayashi, R., Hannon, G. J. (2001). Argonaute2, a link between genetic and biochemical analyses of RNAi. Science 293, 1146. doi: 10.1126/science.1064023
Huang, G., Allen, R., Davis, E. L., Baum, T. J., Hussey, R. S. (2006). Engineering broad root-knot resistance in transgenic plants by RNAi silencing of a conserved and essential root-knot nematode parasitism gene. PNAS 103 (39), 1403–1406. doi: 10.1073/pnas.0604698103
Husain, M., Rasool, K. G., Tufail, M., Aldawood, A. S. (2020). Molecular characterization, expression pattern and RNAi-mediated silencing of vitellogenin receptor gene in almond moth, Cadra cautella. Insect Mol. Biol. 29, 417–430. doi: 10.1111/imb.12646
Huvenne, H., Smagghe, G. (2010). Mechanisms of dsRNA uptake in insects and potential of RNAi for pest control: A review. J. Insect Physiol. 56, 227–235. doi: 10.1016/j.jinsphys.2009.10.004
Ibanez, F., Levy, J., Tamborindeguy, C. (2017). Identification and expression analyses of vitellogenin in Bactericera cockerelli (Šulc). J. Insect. Physiol. 98, 205–213. doi: 10.1016/j.jinsphys.2016.11.009
Ibanez, F., Racine, K., Hoyte, A., Stelinski, L. L. (2019). Reproductive performance among color morphs of Diaphorina citri Kuwayama, vector of citrus greening pathogens. J. Insect Physiol. 117, 103904. doi: 10.1016/j.jinsphys.2019.103904
Izumi, S., Yano, K., Yamamoto, Y., Takahashi, S. Y. (1994). Yolk proteins from insect eggs: Structure, biosynthesis and programmed degradation during embryogenesis. J. Insect Physiol. 40 (9), 735–746. doi: 10.1016/0022-1910(94)90001-9
Jagoueix, S., Bové, J. M., Garnier, M. (1994). The phloem-limited bacterium of greening disease of citrus is a member of the α subdivision of the proteobacteria. Int. J. Sys. Bacteriol. 44 (3), 379–386. doi: 10.1099/00207713-44-3-379
Karasev, A. V., Boyko, V. P., Gowda, S., Nikolaeva, O. V., Hilf, M. E., Koonin, E. V., et al. (1995). Complete sequence of the citrus Tristeza virus RNA genome. Virology 208, 511–520. doi: 10.1006/viro.1995.1182
Khan, A. M., Ashfaq, M., Khan, A. A., Naseem, M. T., Mansoor, S. (2018). Evaluation of potential RNA-interference-target genes to control cotton mealybug, Phenacoccus solenopsis (Hemiptera: Pseudococcuidae). Insect Sci. 25, 778–786. doi: 10.1111/1744-7917.12455
Khan, A. M., Ashfaq, M., Khan, A. A., Rasool, A., Iqbal, J., Mansoor, S. (2015). Inoculation of Nicotiana tabacum with recombinant Potato virus X induces RNA interference in the solenopsis mealybug, Phenacoccus solenopsis Tinsley (Hemiptera: Pseudococcidae). Biotechnol. Letter. 37, 2083–2090. doi: 10.1007/s10529-015-1880-7
Kishk, A., Anber, H. A. I., AbdEl-Raof, T. K., El-Sherbeni, A. E. H. D., Hamed, S., Gowda, S., et al. (2017). RNA interference of carboxylesterases causes nymph mortality in the Asian citrus psyllid, Diaphorina citri. Arch. Insect Biochem. Physiol. 94. doi: 10.1002/arch.21377
Koji, T., Jun, Y., Takashi, A. Y. (2020). Morphological and developmental traits of the binucleation of male accessory gland cells in the benthic water bug, Aphelocheirus vittatus (Hemiptera: Aphelochiridae). J. Insect. Sci. 20, 1–8. doi: 10.1093/jisesa/ieaa082
Lee, K. S., Kim, B. Y., Choo, Y. M., Yoon, H. J., Kang, P. D., Woo, S. D., et al. (2009). Expression profile of cathepsin B in the fat body of Bombyx mori during metamorphosis. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 154 (2), 188–194. doi: 10.1016/j.cbpb.2009.06.002
Li, H., Wang, X., Pan, B., Xheng, Z., Lu, W. (2022). Study on the interference of DicdsVg4 and DicdsVgR in egg formation in the ovary of Diaphorina citri (Kuwayama). BioRxiv. doi: 10.1101/2022.04.15.488471
Livak, K. J., Schmittgen, T. D. (2001). Analysis of relative gene expression data using real-time quantitative PCR and the 2–ΔΔCT method. Methods 25 (4), 402–408. doi: 10.1006/meth.2001.1262
Mamta, B., Rajam, M. V. (2017). RNAi technology: a new platform for crop pest control. Physiol. Mol. Biol. Plants. 23, 487–501. doi: 10.1007/s12298-017-0443-x
McClean, A. P. D., Schwartz, R. E. (1970). Greening of blotchy-mottle disease in citrus. Phytophylactica 2, 177–194.
Medina, M., León, P., Vallejo, C. G. (1988). Drosophila cathepsin B-like proteinase: A suggested role in yolk degradation. Arch. Biochem. Biophys. 263, 355–363. doi: 10.1016/0003-9861(88)90646-7
Meister, G., Tuschl, T. (2004). Mechanisms of gene silencing by double-stranded RNA. Nature 431, 343–349. doi: 10.1038/nature02873
Moura, A. S., Cardoso, A. F., Costa-da-Silva, A. L., Winter, C. E., Bijovsky, A. T. (2015). Two cathepsins B are responsible for the yolk protein hydrolysis in culex quinquefasciatus. PloS One 10 (2), e0118736. doi: 10.1371/journal.pone.0118736
Peng, L., Wang, Q., Zou, M. M., Qin, Y. D., Vasseur, L., Chu, L. N., et al. (2020). CRISPR/Cas9-mediated vitellogenin receptor knockout leads to functional deficiency in the reproductive development of Plutella xylostella. Front. Physiol. 10. doi: 10.3389/fphys.2019.01585
Polgár, L. A. A. (2013). Catalytic mechanisms of cysteine peptidases. Handb. Proteol. Enzymes. 2, 1773–1784. doi: 10.1016/B978-0-12-382219-2.00405-1
Qureshi, J. A., Kostyk, B. C., Stansly, P. A. (2014). Insecticidal suppression of Asian citrus psyllid Diaphorina citri (Hemiptera: Liviidae) vector of huanglongbing pathogens. PloS One 9, 12. doi: 10.1371/journal.pone.0112331
Rawlings, N. D., Barrett, A. J., Finn, R. D. (2016). Twenty years of the MEROPS database of proteolytic enzymes, their substrates and inhibitors. Nucleic Acids Res. 44, D343–D350. doi: 10.1093/nar/gkv1118
Robertson, C. J., Garnsey, S. M., Satyanarayana, T., Folimonova, S., Dawson, W. O. (2005). “Efficient Infection of Citrus Plants with Different Cloned Constructs of Citrus tristeza virus Amplified in Nicotiana benthamiana Protoplasts,” in . International Organization of Citrus Virologists Conference Proceedings, Vol. 1957-2010). 16(16. doi: 10.5070/C50vs6c0m9
RStudio Team (2015). RStudio: Integrated Development Environment for R (Boston, MA). Available at: http://www.rstudio.com/.
Sajid, M., McKerrow, J. H. (2002). Cysteine proteases parasitic organisms. Mol. Biochem. Parasitol. 120, 1–21. doi: 10.1016/S0166-6851(01)00438-8
Santos-Ortega, Y., Killiny, N. (2018). Silencing of sucrose hydrolase causes nymph mortality and disturbs adult osmotic homeostasis in Diaphorina citri (Hemiptera: Liviidae). Insect Biochem. Mol. Biol. 101, 131–143. doi: 10.1016/j.ibmb.2018.09.003
Satyanarayana, T., Bar-Joseph, M., Mawassi, M., Albiach-Martí, M. R., Ayllón, M. A., Gowda, S., et al. (2001). Amplification of Citrus Tristeza Virus from a cDNA clone and infection of Citrus trees. Virology . 280, 87–96. doi: 10.1006/viro.2000.0759
Satyanarayana, T., Gowda, S., Boyko, V. P., Albiach-Marti, M. R., Mawassi, M., Navas-Castillo, J., et al. (1999). An engineered closterovirus RNA replicon and analysis of heterologous terminal sequences for replication. Proc. Natl. Acad. Sci. 96, 7433–7438. doi: 10.1073/pnas.96.13.7433
Shiba, H., Uchida, D., Kobayashi, H., Natori, M. (2001). Involvement of cathepsin B- and L-like proteinases in silk gland histolysis during metamorphosis of Bombyx mori. Arch. Biochem. Biophys. 390 (1), 28–34. doi: 10.1006/abbi.2001.2343
Storer, A. C., Ménard, R. (1994). Catalytic mechanism in papain family of cysteine peptidases. Methods Enzymol. 244, 486–500. doi: 10.1016/0076-6879(94)44035-2
Takahashi, N., Kurata, S., Natori, S. (1993). Molecular cloning of cDNA for the 29 kDa proteinase participating in decomposition of the larval fat body during metamorphosis of Sarcophaga peregrina (flesh fly). FEBS Lett. 334 (2), 153–157. doi: 10.1016/0014-5793(93)81702-2
Teixeira, D. C., Ayres, A. J., Kitajima, E. W., Tanaka, F. A. O., Danet, J. L., Jagoueix-Eveillard, S., et al. (2005a). First report of a huanglongbing-like disease of citrus in Sao Paulo State, Brazil, and association of a new liberibacter species, Candidatus Liberibacter americanus, with the disease. Plant Dis. 89, 107. doi: 10.1094/PD-89-0107A
Teixeira, D. C., Danet, J. L., Eveillard, S., Martins, E. C., Jesus Junior, W. C., Yamamoto, P. T., et al. (2005b). Citrus huanglongbing in São Paulo state, Brazil: PCR detection of the “Candidatus” Liberibacter species associated with the disease. Mol. Cell. Probes. 19, 173–179. doi: 10.1016/j.mcp.2004.11.002
Teixeira, D. C., Saillard, C., Eveillard, S., Danet, J. L., da Costa, P. I., Ayres, A. J., et al. (2005c). “Candidatus Liberibacter americanus”, associated with citrus huanglongbing (greening disease) in São Paulo State, Brazil. Int. J. Sys. Evol. Microbiol. 55, 1857–1862. doi: 10.1099/ijs.0.63677-0
Terra, W., Ferreira, R. C. (1994). Insect digestive enzymes: properties, compartmentalization and function. Comp. Biochem. Physiol. 109B, 1–62. doi: 10.1016/0305-0491(94)90141-4
Tiwari, S., Mann, R. S., Rogers, M. E., Stelinski, L. L. (2011). Insecticide resistance in field populations of Asian citrus psyllid in Florida. Pest Manage. Sci. 67, 1258–1268. doi: 10.1002/ps.2181
Tsai, J. H., Liu, Y. H. (2000). Biology of Diaphorina citri (Homoptera: Psyllidae) on four host plants. J. Econ. Entomol. 6, 1721–1725. doi: 10.1603/0022-0493-93.6.1721
Untergasser, A., Cutcutache, I., Koressaar, T., Ye, J., Faircloth, B. C., Remm, M., et al. (2012). Primer3 - new capabilities and interfaces. Nucleic Acids Res. 40 (15), e115.
Vyas, M., Fisher, T. W., He, R., Nelson, W., Yin, G., Cicero, J. M., et al. (2015). Asian citrus psyllid expression profiles suggest candidatus liberibacter asiaticus-mediated alteration of adult nutrition and metabolism, and of nymphal development and immunity. PloS One 10, 6. doi: 10.1371/journal.pone.0130328
Whyard, S., Singh, A. D., Wong, S. (2009). Ingested doubleçe-stranded RNAs can act species-specific insecticides. Insect Biochem. Mol. Biol. 39, 824–832. doi: 10.1016/j.ibmb.2009.09.007
Wiederanders, B. (2003). Structure-function relationships in class CA1 cysteine peptidase propeptides. Acta Biochim. Polonica. 50, 3. doi: 10.18388/abp.2003_3661
Wu, Z., Guo, W., Xie, Y., Zhou, S. (2016). Juvenile hormone activates the transcription of cell-division-cycle 6 (Cdc6) for polyploidy-dependent insect vitellogenesis and oogenesis. J. Biol. Chem. 291, 5418–5427. doi: 10.1074/jbc.M115.698936
Wu, Z., Guo, W., Yang, L., He, Q., Zhou, S. (2018). Juvenile hormone promotes locust fat body cell polyploidization and vitellogenesis by activating the transcription of Cdk6 and E2f1. Insect Biochem. Mol. Biol. 102, 1–10. doi: 10.1016/j.ibmb.2018.09.002
Wu, Z., Yang, L., He, Q., Zhou, S. (2021). Regulatory mechanisms of vitellogenesis in insects. Front. Cell Dev. Biol. 8. doi: 10.3389/fcell.2020.593613
Wuriyanghan, H., Falk, B. W. (2013). RNA interference towards the potato psyllid, Bactericera cockerelli, is induced in plants infected with recombinant Tobacco mosaic virus (TMV). PloS One 8, e66050. doi: 10.1371/journal.pone.0066050
Yan, S., Ren, B., Zeng, B., Shen, J. (2020). Improving RNAi efficiency for pest control in crop species. BioTechniques 68 (5), 283–290. doi: 10.2144/btn-2019-0171
Yang, H., Zhang, R., Zhang, Y., Liu, Q., Li, Y., Gong, J., et al. (2020). Cathepsin-L is involved in degradation of fat body and programmed cell death in Bombyx mori. Gene 760, (144998). doi: 10.1016/j.gene.2020.144998
Yu, X., Gowda, S., Killiny, N. (2017). Double-stranded RNA delivery through soaking mediates silencing of the muscle protein 20 and increases mortality to the Asian citrus psyllid, Diaphorina citri. Pest Manage. Sci. 73 (9), 1846–1853. doi: 10.1002/ps.4549
Yu, H. Z., Huang, Y. L., Li, N. Y., Xie, Y. X., Zhou, C. H., Lu, Z. J. (2019). Potential roles of two Cathepsin genes, DcCath-L and DcCath-O in the innate immune response of Diaphorina citri. J. Asia-Pacific Entomol. 22, 1060–1069. doi: 10.1016/j.aspen.2019.05.010
Zhang, H., Li, H. C., Miao, X. X. (2013). Feasibility, limitation and possible solutions of RNAi-based technology for insect pest control. Insect Sci. 20, 15–30. doi: 10.1111/j.1744-7917.2012.01513.x
Zhang, T. T., Qiu, Z. X., Li, Y., Wang, W. Y., Li, M. M., Guo, P., et al. (2019). The mRNA expression and enzymatic activity of three enzymes during embryonic development of the hard tich Haemaphysalis longicornis. Parasites Vectors. 12, 96. doi: 10.1186/s13071-019-3360-8
Zhao, X. F., Wang, J. X., Xu, X. L., Schmid, R., Wieczorek, H. (2002). Molecular cloning and characterization of the cathepsin B-like proteinase from the cotton boll worm, Helicoverpa armigera. Insect Mol. Biol. 11 (6), 567–575. doi: 10.1046/j.1365-2583.2002.00366.x
Keywords: Diaphorina citri, Huanglongbing, CTV, RNAi, Cathepsin B, Cathepsin L
Citation: Ibanez F, Vieira Rocha S, Dawson WO, El-Mohtar C, Robertson C, Stelinski LL and Soares-Costa A (2023) Gene silencing of cathepsins B and L using CTV-based, plant-mediated RNAi interferes with ovarial development in Asian citrus psyllid (ACP), Diaphorina citri. Front. Plant Sci. 14:1219319. doi: 10.3389/fpls.2023.1219319
Received: 08 May 2023; Accepted: 12 September 2023;
Published: 29 September 2023.
Edited by:
Vinay Panwar, Chaudhary Charan Singh University, IndiaReviewed by:
Subhas Hajeri, Citrus Pest Detection Program, United StatesWayne B. Hunter, United States Department of Agriculture (USDA), United States
Copyright © 2023 Ibanez, Vieira Rocha, Dawson, El-Mohtar, Robertson, Stelinski and Soares-Costa. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Freddy Ibanez, ZnJlZGR5LmliYW5lei1jYXJyYXNAYWcudGFtdS5lZHU=; Sâmara Vieira Rocha, c2FtYXJhdjE3QGdtYWlsLmNvbQ==; Andrea Soares-Costa, YW5kcmVhc2NAdWZzY2FyLmJy
†These authors have contributed equally to this work and share first authorship