
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
OPINION article
Front. Plant Sci. , 20 June 2023
Sec. Crop and Product Physiology
Volume 14 - 2023 | https://doi.org/10.3389/fpls.2023.1216337
This article is part of the Research Topic Advances in Environmental Stress Biology and Important Agronomic Traits Improvement in Non-staple Crops View all 11 articles
 Dominik K. Großkinsky1*†
Dominik K. Großkinsky1*† Jean-Denis Faure2†Yves Gibon3,4
Jean-Denis Faure2†Yves Gibon3,4 Richard P. Haslam5†Björn Usadel6,7†
Richard P. Haslam5†Björn Usadel6,7† Federica Zanetti8†
Federica Zanetti8† Claudia Jonak1*†
Claudia Jonak1*†The current agricultural and food system faces diverse and increasing challenges. These include feeding an ever-growing human population, expected to reach about 10 billion by 2050 combined with societal disruption, and the need to cope with the impact of climate change (FAO, 2022). Given that future environmental conditions will limit crop productivity (Zhao et al., 2017; Cooper et al., 2021) and the limited potential to continually increase the performance of staple crops by conventional breeding (Hickey et al., 2019), there is an urgent need to transform agricultural systems. Central to this transformation is the application of alternative, accelerated, and sustainable approaches for the improvement and development of underutilized crops (Hickey et al., 2019). Modern breeding strategies for major crops have widely integrated novel technologies, such as advanced phenotyping or genome-wide interactions, and even epigenomics within “beyond the gene” strategies (Crisp et al., 2022) to speed up crop/genotype selection (Hickey et al., 2019; Kumar et al., 2023). Deploying phenotyping at different scales has the potential to identify novel trait(s) components that can be targeted to accelerate crop improvement (Araus and Cairns, 2014; Großkinsky et al., 2015b; Zhao et al., 2019; Varshney et al., 2021). There is even greater potential for these technologies when used to improve underutilized crops and support the agricultural transformation, as underutilized crops typically lack a biased breeding/selection history, i.e., they often exhibit a high genetic diversity and potential, and are usually better adapted to challenging environments (Kumar et al., 2021; Kumar et al., 2023). To illustrate the application of an integrative phenomics approach we discuss how combining multi-omics and advanced phenotyping is being applied to the underutilized oilseed crop Camelina sativa (camelina, gold-of-pleasure, false flax) to facilitate the generation of climate-smart crops for future agricultural systems.
The time needed to select new varieties or species, but also to set up new agronomic practices (e.g., reduced input management) is currently too long in view of the rapidly changing climate1. While traditionally varieties have often been selected in a context of high input mechanized monoculture, it is now necessary to develop predictive strategies to formulate the genotypes and agronomic practices adapted to low input, sustainable and diversified cultures. That said, plant biologists have long been studying adaptation or acclimation to stresses associated with climate change (e.g., drought, heat, attacks from pests and pathogens) to unravel the involved mechanisms and identify targets to improve crop performance.
Reverse genetics usually starts from a gene and characterizes associated phenotypes. Here, the development of “omics” has revolutionized biology over the past 20 years and led to a dramatic increase in the number of candidate genes and networks (e.g., Kajala et al., 2021). Great progress was initially achieved in Arabidopsis but has increasingly been translated to major crops (Fernie and Tohge, 2017). Bottom-up systems biology goes one step further by assembling reductionist knowledge using models to explain phenotypes (Keurentjes et al., 2011). One consequence of this is that more complex mechanisms can be analysed, and more sophisticated improvement strategies proposed (Chen et al., 2021). This ensemble of approaches is being carried out in a growing number of species (Stevens et al., 2018; Hawkins et al., 2021). However, when going beyond model plants and major established crops, challenges remain. Whilst resolving homozygous genomes (van Rengs et al., 2021) and pangenomes (Bayer et al., 2020) of crops with a medium genome size is now achievable with the advent of novel sequencing technologies (Mascher et al., 2021), challenges remain with consistent gene annotation and ontology developments. Rapid progress is being made with more mature tools, facilitating structural (Holst et al., 2023) and functional genome annotation (Bolger et al., 2018; Schwacke et al., 2019), whilst plant genome data continues to be gathered in comprehensive databases (Vandepoele, 2017). However, several problems remain outstanding, including the accurate description of phenotypes together with necessary metadata (Papoutsoglou et al., 2020) and simple data management approaches (Pommier et al., 2019; Jacob et al., 2020). This is especially true for minor or even orphan crops where, without the necessary resources, these strategies have not yet reached the same level of maturity as for major field crops (Watt et al., 2020). To address this, remote sensing, deep phenotyping, and machine learning approaches are continually being developed for crops (Fernie et al., 2021; Wong et al., 2023).
Forward genetics starts from the phenotype and tries to identify the underlying mechanisms, genes and/or markers. Traditionally, crop improvement has been based on traits of interest, especially yield. The most promising lines are retained, which results in an enrichment of the germplasm with performance-promoting alleles. More recently, proxies such as plant height have started to be used to speed up the process (Madec et al., 2017). The aim being to find phenotype-based parameters that are easy and cheap to measure. Classical selection in the field using plots is very expensive and generally subject to seasons and environmental hazards. It is in this context that multiple phenotyping facilities have been established2. By enabling plant phenotyping under controlled conditions, their purpose is to generate field performance proxies at affordable cost (Reynolds et al., 2019), although the confirmation of agronomic trait evaluations carried out under controlled conditions in the field is challenging (Hohmann et al., 2016). Indeed, when transferring phenotypic information from controlled environments to agronomic field conditions, phenotypes are not always confirmed (Shi et al., 2017; Zhang et al., 2019; Metje-Sprink et al., 2020). For example, evaluations of edited plants that have targeted mutations for specific and well-characterized traits have shown limitations, as only 13 out of 22 publications confirmed yield effects of edited plants identified in controlled environments in subsequent field trials (Metje-Sprink et al., 2020). Therefore, field trials remain essential for the study of traits interacting with complex environments. Phenotyping under controlled conditions requires continued evaluation of growth scenarios to determine the best predictive variables. Over the past decade more and more sophisticated sensors and platforms have emerged. Whether in fields or greenhouses, they use an ever-increasing number of imaging techniques (Großkinsky et al., 2015a; Brichet et al., 2017; Yang et al., 2020; Dodig et al., 2021) as non-destructive tools that allow the generation of performance proxies throughout plant growth (e.g., Millet et al., 2019). Genomic selection (e.g., R2D2 Consortium et al., 2021), increasingly used by breeders, consists of predicting the genetic value of candidates for selection through the development of models linking phenotype and genotype. The advantage of this approach is that the phenotyping step can be done with a small subset of the germplasm and the search for potentially interesting genotypes (pre-breeding) can be done with young plants and therefore at low cost. Paradoxically, sampling of plant material is still largely manual and the development of robots for destructive-type observations is still in its infancy (Foix et al., 2018). This may be one of the reasons why “omics” are not yet fully considered as selection tools, in particular metabolomics, whose costs have fallen sharply (López-Ruiz et al., 2019). Evidence continues to emerge demonstrating that transcriptomics or metabolomics (Westhues et al., 2017) can carry more condensed information than the genome, and these approaches can be used to build top-down models capable of predicting traits, particularly in integrative approaches combining omics technologies with advanced phenotyping.
Despite a huge variety of cultivated plants, historical developments in global agriculture have resulted in a very narrow diversity of crops contributing to our current food system, with only few species accounting for over 90% of world food production, e.g., the staple cereals rice, wheat, and maize (Milla and Osborne, 2021). In contrast, the vast majority of more than 5000 known edible crops are only cultivated on a small scale, if at all, and are largely neglected in modern agricultural settings due to their limited competitiveness (Padulosi, 2017). These wild, cultivated, or semi-domesticated crops are not yet fully integrated into typical agricultural systems worldwide (Padulosi, 2017). These crops are typically underutilized in agriculture due to a lack of improvement and in-depth agronomic knowledge and/or experience, moreover, they are poorly represented in fundamental research, which is mostly focused on model plants or staple crops. Thus, there is often a lack of complementary knowledge, technologies, and resources from basic research to support use and integration of such crops. Underutilised crops (for examples see Table 1) have recently gained attention as many of them hold untapped potential for coping with the challenges of current and future agriculture (Padulosi, 2017; Kumar et al., 2023), e.g., increasing agro-biodiversity, improving the overall resilience of agricultural systems against climate change and sourcing alternative (high-quality) food ingredients. This is evident in research today e.g., EU-funded projects, aiming to promote the use of underutilized crops, such as INCREASE3 aiming at improving the sustainable use of the pulses chickpea, common bean, lentil and lupin, PROTEIN2FOOD4 addressing the development and sustainable use of quinoa, amaranth, buckwheat and pulses like lupins, and UNTWIST5 aiming at the promotion of the sustainable use of camelina as a promising climate-resilient oilseed crop. All these projects focus on crop phenotypes and environmental plasticity.
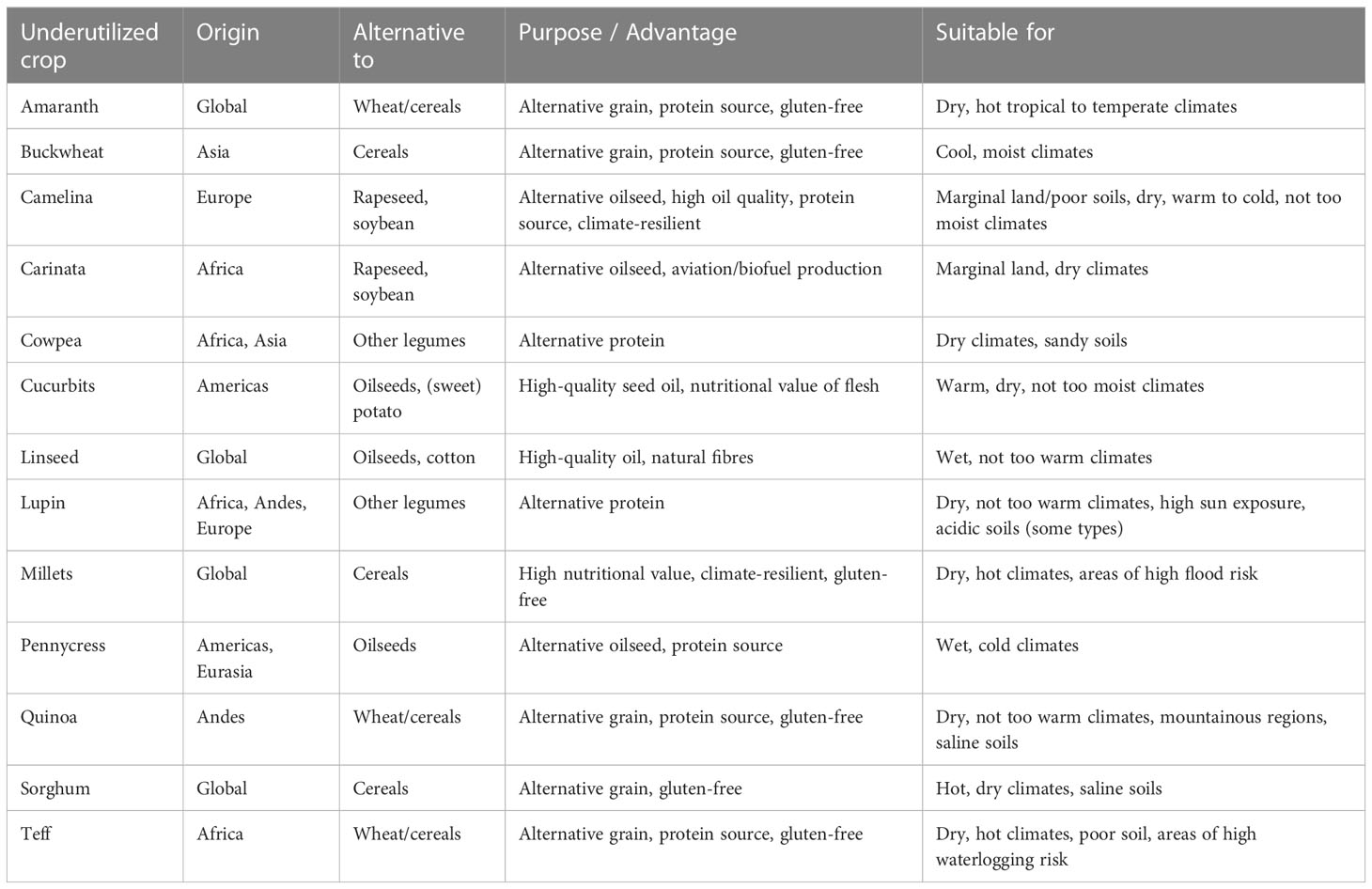
Table 1 Examples of promising underutilized crops exploitable for agricultural use.
Among underutilized crops with significant potential, camelina has recently attracted interest as a promising alternative to other oilseeds (e.g., rapeseed). From a scientific perspective, this is also due to its genetic similarity with the model plant Arabidopsis (Kagale et al., 2014). Camelina can be easily integrated into existing agricultural systems (Zanetti et al., 2021). It has a natural capacity to withstand harsh environmental conditions, and tolerance to pests and diseases represent additional features that make it a suitable crop for many environments worldwide (Berti et al., 2016; Zanetti et al., 2021), except for too wet conditions (Stasnik et al., 2022). This particularity also makes it a potential source of resistance mechanisms that would have disappeared or have become insufficient in species that have been selected above all for their yield in optimal conditions. So, the aim here is not to rediscover mechanisms that are already known. The emerging interest in camelina has spurred genomic and genetic investigations, including the description of a reference genome (Kagale et al., 2014). The development of Recombinant Inbred Lines (RILs) has further allowed the identification of important QTLs e.g., 1000-seed weight, seed number per pod, days to flowering or seed oil content (King et al., 2019; Li et al., 2021). Nevertheless, the throughput of these approaches is limited particularly at the phenotype level, as for other (underutilized) crops. Hence, further focused improvement of camelina is required and the use of novel approaches and technologies, specifically integrating the crop phenotype, is necessary and achievable as respective protocols for high-throughput phenotyping are established (Vello et al., 2022).
Several projects are emerging that take advantage of the availability and affordability of modern technologies (e.g., multi-omics, phenomics, network discovery) to investigate plant plasticity in novel crops (e.g., Girija et al., 2021; Marks et al., 2021). Understanding plant plasticity is key for the development of novel crops for resilient food production systems in increasingly variable climates. As an exemplary project, UNTWIST5 targets camelina to develop a systems-based understanding of the key mechanisms underlying crop stress tolerance using a phenomics approach. The project is based on the premise that the unravelling of stress adaptation mechanisms in naturally resilient camelina (Zanetti et al., 2021), which has not yet undergone intensive breeding, will reveal successful stress adaptation strategies. Research is centred on agronomically relevant stress scenarios (linking experiments in controlled environments and field conditions) to improve our understanding of plant responses to challenging growth conditions. Therefore, diverse germplasm is analysed at the genomic, epigenetic, transcriptional, physiological, proteomic, and metabolomic level (incl. lipidome, redox status) complemented by agronomic parameters and advanced non-invasive multi-sensor phenotyping. Despite limited basic knowledge on camelina compared to well-studied plant species, this integrative multi-omics approach deploying phenotyping from cell to whole plant level (Dhont et al., 2013; Ghanem et al., 2015; Großkinsky et al., 2015b) allows the identification and dissection of mechanisms contributing to camelina performance under challenging environments representative of future conditions. Integration of the multi-layered data serves modelling approaches and supports potential marker development for crop improvement under adverse climate conditions. Furthermore, the results will underpin databases, protocols, and advice for downstream application. The new knowledge can be utilized for further improvements of camelina and other (underutilized) crops in combination with modern strategies in breeding and engineering crops (Jiang et al., 2017; Morineau et al., 2017; Lemmon et al., 2018; Fernie and Yan, 2020; Rönspies et al., 2021; Yu et al., 2021; Bellec et al., 2022; Han et al., 2022).
Global agriculture urgently requires alternative strategies in crop development and improvement. So far, the great potential of underutilized crops to contribute to the agricultural transition in our food systems remains untapped. This is due to a lack of knowledge and appropriate technologies; shortfalls that are now being addressed. Recognizing that phenotypes are as or even more important than genotypic information has driven recent advances in crop phenotyping. These advances greatly contribute to the development of multi-omics systems-based phenomics approaches, which have shown their potential for accessing previously untapped resources and can be applied more directly to underutilized non-staple crops in the future, significantly reducing the dependency on information from staple crops and model plants as the only information source. This allows the effective improvement and utilization of such crops to support the re-configuration of agricultural and food systems in a sustainable manner. Most notably, phenomics and multi-omics approaches will enable agriculture to harness the potential of underutilized crops. Projects like UNTWIST, which apply these approaches, are needed to prove, and validate the potential of these novel tools. Ultimately, such projects can serve as a blueprint for improving underutilized crops and provide a platform for establishing more climate-resilient food production systems.
All authors jointly defined the outline of the manuscript. DKG drafted the manuscript with input from all authors. All authors contributed to the article and approved the submitted version.
This work was performed within the UNTWIST project (www.untwist.eu) funded by the European Commission Horizon 2020 Research & Innovation Programme (Grant Agreement No 862524). Rothamsted Research (UK) receives grant-aided support from the BBSRC (UK) and RH was partially supported by the BBSRC(UK) Institute Strategic Programme Green Engineering (BB/X010988/1). BU was partially supported by DFG grant 456082119.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Araus, J. L., Cairns, J. E. (2014). Field high-throughput phenotyping: the new crop breeding frontier. Trends Plant Sci. 19, 52–61. doi: 10.1016/j.tplants.2013.09.008
Bayer, P. E., Golicz, A. A., Scheben, A., Batley, J., Edwards, D. (2020). Plant pan-genomes are the new reference. Nat. Plants 6, 914–920. doi: 10.1038/s41477-020-0733-0
Bellec, Y., Guyon-Debast, A., François, T., Gissot, L., Biot, E., Nogué, F., et al. (2022). New flowering and architecture traits mediated by multiplex CRISPR-Cas9 gene editing in hexaploid Camelina sativa. Agronomy 12, 1873. doi: 10.3390/agronomy12081873
Berti, M., Gesch, R. W., Eynck, C., Anderson, J., Cermak, S. (2016). Camelina uses, genetics, genomics, production, and management. Ind. Crops Products 94, 690–710. doi: 10.1016/j.indcrop.2016.09.034
Bolger, M. E., Arsova, B., Usadel, B. (2018). Plant genome and transcriptome annotations: from misconceptions to simple solutions. Briefings Bioinf. 19, 437–449. doi: 10.1093/bib/bbw135
Brichet, N., Fournier, C., Turc, O., Strauss, O., Artzet, S., Pradal, C., et al. (2017). A robot-assisted imaging pipeline for tracking the growths of maize ear and silks in a high-throughput phenotyping platform. Plant Methods 13, 96. doi: 10.1186/s13007-017-0246-7
Chen, J., Beauvoit, B., Génard, M., Colombié, S., Moing, A., Vercambre, G., et al. (2021). Modelling predicts tomatoes can be bigger and sweeter if biophysical factors and transmembrane transporters are fine-tuned during fruit development. New Phytol. 230, 1489–1502. doi: 10.1111/nph.17260
Cooper, M., Voss-Fels, K. P., Messina, C. D., Tang, T., Hammer, G. L. (2021). Tackling G × e × m interactions to close on-farm yield-gaps: creating novel pathways for crop improvement by predicting contributions of genetics and management to crop productivity. Theor. Appl. Genet. 134, 1625–1644. doi: 10.1007/s00122-021-03812-3
Crisp, P. A., Bhatnagar-Mathur, P., Hundleby, P., Godwin, I. D., Waterhouse, P. M., Hickey, L. T. (2022). Beyond the gene: epigenetic and cis-regulatory targets offer new breeding potential for the future. Curr. Opin. Biotechnol. 73, 88–94. doi: 10.1016/j.copbio.2021.07.008
Dhont, S., Wuyts, N., Inzé, D. (2013). Cell to whole-plant phenotyping: the best is yet to come. Trends Plant Sci. 18, 428–439. doi: 10.1016/j.tplants.2013.04.008
Dodig, D., Božinović, S., Nikolić, A., Zorić, M., Vančetović, J., Ignjatović-Micić, D., et al. (2021). Dynamics of maize vegetative growth and drought adaptability using image-based phenotyping under controlled conditions. Front. Plant Sci. 12. doi: 10.3389/fpls.2021.652116
FAO (2022). The future of food and agriculture – drivers and triggers for transformation Vol. 3 (Rome: The Future of Food and Agriculture). doi: 10.4060/cc0959en
Fernie, A. R., Alseekh, S., Liu, J., Yan, J. (2021). Using precision phenotyping to inform de novo domestication. Plant Physiol. 186, 1397–1411. doi: 10.1093/plphys/kiab160
Fernie, A. R., Tohge, T. (2017). The genetics of plant metabolism. Annu. Rev. Genet. 51, 287–310. doi: 10.1146/annurev-genet-120116-024640
Fernie, A. R., Yan, J. (2020). Targeting key genes to tailor old and new crops for a greener agriculture. Mol. Plant 13, 354–356. doi: 10.1016/j.molp.2020.02.007
Foix, S., Alenyà, G., Torras, C. (2018). Task-driven active sensing framework applied to leaf probing. Comput. Electron. Agric. 147, 166–175. doi: 10.1016/j.compag.2018.01.020
Ghanem, M. E., Marrou, H., Sinclair, T. R. (2015). Physiological phenotyping of plants for crop improvement. Trend Plant Sci. 20, 139–144. doi: 10.1016/j.tplants.2014.11.006
Girija, A., Han, J., Corke, F., Brook, J., Doonan, J., Yadav, R., et al. (2021). Elucidating drought responsive networks in tef (Eragrostis tef) using phenomic and metabolomic approaches. Physiol. Plant. 174, e13597. doi: 10.1111/ppl.13597
Großkinsky, D. K., Pieruschka, R., Svensgaard, J., Rascher, U., Christensen, S., Schurr, U., et al. (2015a). Phenotyping in the fields: dissecting the genetics of quantitative traits and digital farming. New Phytol. 207, 950–952. doi: 10.1111/nph.13529
Großkinsky, D. K., Svensgaard, J., Christensen, S., Roitsch, T. (2015b). Plant phenomics and the need for physiological phenotyping across scales to narrow the genotype-to-phenotype knowledge gap. J. Exp. Bot. 66, 5429–5440. doi: 10.1093/jxb/erv345
Han, L., Silvestre, S., Sayanova, O., Haslam, R. P., Napier, J. A. (2022). Using field evaluation and systematic iteration to rationalize the accumulation of omega-3 long-chain polyunsaturated fatty acids in transgenic Camelina sativa. Plant Biotechnol. J. 20, 1833–1852. doi: 10.1111/pbi.13867
Hawkins, C., Ginzburg, D., Zhao, K., Dwyer, W., Xue, B., Xu, A., et al. (2021). Plant metabolic network 15: a resource of genome-wide metabolism databases for 126 plants and algae. J. Integr. Plant Biol. 63, 1888–1905. doi: 10.1111/jipb.13163
Hickey, L. T., Hafeez, A. N., Robinson, H., Jackson, S. A., Leal-Bertioli, S. C. M., Tester, M., et al. (2019). Breeding crops to feed 10 billion. Nat. Biotechnol. 37, 744–754. doi: 10.1038/s41587-019-0152-9
Hohmann, M., Stahl, A., Rudloff, J., Wittkop, B., Snowdon, R. J. (2016). Not a load of rubbish: simulated field trials in large-scale containers. Plant Cell Environ. 39, 2064–2073. doi: 10.1111/pce.12737
Holst, F., Bolger, A., Günther, C., Maß, J., Triesch, S., Kindel, F., et al. (2023). Helixer–de novo prediction of primary eukaryotic gene models combining deep learning and a hidden Markov model. bioRxiv. doi: 10.1101/2023.02.06.527280
Jacob, D., David, R., Aubin, S., Gibon, Y. (2020). Making experimental data tables in the life sciences more FAIR: a pragmatic approach. GigaScience 9, giaa144. doi: 10.1093/gigascience/giaa144
Jiang, W. Z., Henry, I. M., Lynagh, P. G., Comai, L., Cahoon, E. B., Weeks, D. P. (2017). Significant enhancement of fatty acid composition in seeds of the allohexaploid, Camelina sativa, using CRISPR/Cas9 gene editing. Plant Biotechnol. J. 15, 648–657. doi: 10.1111/pbi.12663
Kagale, S., Koh, C., Nixon, J., Bolina, V., Clarke, W. E., Tuteja, R., et al. (2014). The emerging biofuel crop Camelina sativa retains a highly undifferentiated hexaploid genome structure. Nat. Commun. 5, 3706. doi: 10.1038/ncomms4706
Kajala, K., Gouran, M., Shaar-Moshe, L., Mason, G. A., Rodriguez-Medina, J., Kawa, D., et al. (2021). Innovation, conservation, and repurposing of gene function in root cell type development. Cell 184, 3333–33348. doi: 10.1016/j.cell.2021.04.024
Keurentjes, J. J. B., Angenent, G. C., Dicke, M., Martins Dos Santos, V. A. P., Molenaar, J., van der Putten, W. H., et al. (2011). Redefining plant systems biology: from cell to ecosystem. Trends Plant Sci. 16, 183–190. doi: 10.1016/j.tplants.2010.12.002
King, K., Li, H., Kang, J., Lu, C. (2019). Mapping quantitative trait loci for seed traits in Camelina sativa. Theor. Appl. Genet. 132, 2567–2577. doi: 10.1007/s00122-019-03371-8
Kumar, A., Anju, T., Kumar, S., Chhapekar, S. S., Sreedharan, S., Singh, A., et al. (2021). Integrating omics and gene editing tools for rapid improvement of traditional food plants for diversified and sustainable food security. Int. J. Mol. Sci. 22, 8093. doi: 10.3390/ijms22158093
Kumar, B., Singh, A. K., Bahuguna, R. N., Pareek, A., Singla-Pareek, S. L. (2023). Orphan crops: a genetic treasure trove for hunting stress tolerance genes. Food Energy Secur. 12, e436. doi: 10.1002/fes3.436
Lemmon, Z. H., Reem, N. T., Dalrymple, J., Soyk, S., Swartwood, K. E., Rodriguez-Leal, D., et al. (2018). Rapid improvement of domestication traits in an orphan crop by genome editing. Nat. Plants 4, 766–770. doi: 10.1038/s41477-018-0259-x
Li, H., Hu, X., Lovell, J. T., Grabowski, P. P., Mamidi, S., Chen, C., et al. (2021). Genetic dissection of natural variation in oilseed traits of camelina by whole-genome resequencing and QTL mapping. Plant Genome 14, e20110. doi: 10.1002/tpg2.20110
López-Ruiz, R., Romero-González, R., Garrido Frenich, A. (2019). Ultrahigh-pressure liquid chromatography-mass spectrometry: an overview of the last decade. TrAC Trends Analytical Chem. 118, 170–181. doi: 10.1016/j.trac.2019.05.044
Madec, S., Baret, F., de Solan, B., Thomas, S., Dutartre, D., Jezequel, S., et al. (2017). High-throughput phenotyping of plant height: comparing unmanned aerial vehicles and ground LiDAR estimates. Front. Plant Sci. 8. doi: 10.3389/fpls.2017.02002
Marks, M. D., Chopra, R., Sedbrook, J. C. (2021). Technologies enabling rapid crop improvements for sustainable agriculture: example pennycress (Thlaspi arvense l.). Emerging Topics Life Sci. 5, 325–335. doi: 10.1042/ETLS20200330
Mascher, M., Wicker, T., Jenkins, J., Plott, C., Lux, T., Koh, C. S., et al. (2021). Long-read sequence assembly: a technical evaluation in barley. Plant Cell 33, 1888–1906. doi: 10.1093/plcell/koab077
Metje-Sprink, J., Sprink, T., Hartung, F. (2020). Genome-edited plants in the field. Curr. Opin. Biotechnol. 61, 1–6. doi: 10.1016/j.copbio.2019.08.007
Milla, R., Osborne, C. P. (2021). Crop origins explain variation in global agricultural relevance. Nat. Plants 7, 598–607. doi: 10.1038/s41477-021-00905-1
Millet, E. J., Kruijer, W., Couple-Ledru, A., Alvarez Prado, S., Cabrera-Bosquet, L., Lacube, S., et al. (2019). Genomic prediction of maize yield across European environmental conditions. Nat. Genet. 51, 952–956. doi: 10.1038/s41588-019-0414-y
Morineau, C., Bellec, Y., Tellier, F., Gissot, L., Kelemen, Z., Nogué, F., et al. (2017). Selective gene dosage by CRISPR-Cas9 genome editing in hexaploid Camelina sativa. Plant Biotechnol. J. 15, 729–739. doi: 10.1111/pbi.12671
Papoutsoglou, E. A., Faria, D., Arend, D., Arnaud, E., Athanasiadis, I. N., Chaves, I., et al. (2020). Enabling reusability of plant phenomic datasets with MIAPPE 1.1. New Phytol. 227, 260–273. doi: 10.1111/nph.16544
Pommier, C., Michotey, C., Cornut, G., Roumet P Duchêne, E., Flores, R., Lebreton, A., et al. (2019). Applying FAIR principles to plant phenotypic data management in GnpIS. Plant Phenomics 2019, 1671403. doi: 10.34133/2019/1671403
R2D2 Consortium, Fugeray-Scarbel, A., Bastien, C., Dupont-Nivet, M., Lemarié, S. (2021). Why and how to switch to genomic selection: lessons from plant and animal breeding experience. Front. Genet. 12. doi: 10.3389/fgene.2021.629737
Reynolds, D., Baret, F., Welcker, C., Bostrom, A., Ball, J., Cellini, F., et al. (2019). What is cost-efficient phenotyping? Optimizing costs for different scenarios. Plant Sci. 282, 14–22. doi: 10.1016/j.plantsci.2018.06.015
Rönspies, M., Dorn, A., Schindele, P., Puchta, H. (2021). CRISPR-cas-mediated chromosome engineering for crop improvement and synthetic biology. Nat. Plants 7, 566–573. doi: 10.1038/s41477-021-00910-4
Schwacke, R., Ponce-Soto, G. Y., Krause, K., Bolger, A. M., Arsova, B., Hallab, A., et al. (2019). MapMan4: a redefined protein classification and annotation framework applicable to multi-omics data analysis. Mol. Plant 12, 879–892. doi: 10.1016/j.molp.2019.01.003
Shi, J., Gao, H., Wang, H., Lafitte, H. R., Archibald, R. L., Yang, M., et al. (2017). ARGOS8 variants generated by CRISPR-Cas9 improve maize grain yield under field drought stress conditions. Plant Biotechnol. J. 15, 207–216. doi: 10.1111/pbi.12603
Stasnik, P., Großkinsky, D. K., Jonak, C. (2022). Physiological and phenotypic characterization of diverse Camelina sativa lines in response to waterlogging. Plant Physiol. Biochem. 183, 120–127. doi: 10.1016/j.plaphy.2022.05.007
Stevens, R. G., Baldet, P., Bouchet, J.-P., Causse, M., Deborde, C., Deschodt, C., et al. (2018). A systems biology study in tomato fruit reveals correlations between the ascorbate pool and genes involved in ribosome biogenesis, translation, and the heat-shock response. Front. Plant Sci. 9. doi: 10.3389/fpls.2018.00137
Vandepoele, K. (2017). “A guide to the PLAZA 3.0 plant comparative genomic database,” in Plant genomics databases. methods in molecular biology, vol. 1533 . Ed. van Dijk, A. (New York, NY: Humana Press). doi: 10.1007/978-1-4939-6658-5_10
van Rengs, W. M. J., Schmidt, M. H.-W., Effgen, S., Wang, Y., Zaidan, M. W. A. M., Huettel, B., et al. (2021). A gap-free tomato genome built from complementary PacBio and nanopore long DNA sequences reveals extensive linkage drag during breeding. BioRxiv. doi: 10.1101/2021.08.30.456472
Varshney, R. K., Bohra, A., Roorkiwal, M., Barmukh, R., Cowling, W. A., Chitikineni, A., et al. (2021). Fast-forward breeding for a food-secure world. Trends Genet. 37, 1124–1136. doi: 10.1016/j.tig.2021.08.002
Vello, E., Aguirre, J., Shao, Y., Bureau, T. (2022). “Camelina sativa high-throughput phenotyping under normal and salt conditions using a plant phenomics platform,” in High-throughput plant phenotyping: methods and protocols. methods in molecular biology. Eds. Lorence, A., Medina-Jimenez, K. (New York, NY: Humana Press), 2539. doi: 10.1007/978-1-0716-2537-8_4
Watt, M., Fiorani, F., Usadel, B., Rascher, U., Muller, O., Schurr, U. (2020). Phenotyping: new windows into the plant for breeders. Annular Rev. Plant Biol. 71, 689–712. doi: 10.1146/annurev-arplant-042916-041124
Westhues, M., Schrag, T. A., Heuer, C., Thaller, G., Utz, H. F., Schipprack, W., et al. (2017). Omics-based hybrid prediction in maize. Theor. Appl. Genet. 130, 1927–1939. doi: 10.1007/s00122-017-2934-0
Wong, C. Y., Gilbert, M. E., Pierce, M. A., Parker, T. A., Palkovic, A., Gepts, P., et al. (2023). Hyperspectral remote sensing for phenotyping the physiological drought response of common and tepary bean. Plant Phenomics 5, 21. doi: 10.34133/plantphenomics.0021
Yang, W., Feng, H., Zhang, X., Zhang, J., Doonan, J. H., Batchelor, W. D., et al. (2020). Crop phenomics and high-throughput phenotyping: past decades, current challenges, and future perspectives. Mol. Plant 13, 187–214. doi: 10.1016/j.molp.2020.01.008
Yu, H., Lin, T., Meng, X., Du, H., Zhang, J., Liu, G., et al. (2021). A route to de novo domestication of wild allotetraploid rice. Cell 184, 1156–1170. doi: 10.1016/j.cell.2021.01.013
Zanetti, F., Alberghini, B., Marjanović Jeromela, A., Grahovac, N., Rajković, D., Kiprovski, B., et al. (2021). Camelina, an ancient oilseed crop actively contributing to the rural renaissance in europe. a review. Agron. Sustain. Dev. 41, 2. doi: 10.1007/s13593-020-00663-y
Zhang, A., Liu, Y., Wang, F., Li, T., Chen, Z., Kong, D., et al. (2019). Enhanced rice salinity tolerance via CRISPR/Cas9-targeted mutagenesis of the OsRR22 gene. Mol. Breed. 39, 47. doi: 10.1007/s11032-019-0954-y
Zhao, C., Liu, B., Piao, S., Wang, X., Lobell, D. B., Huang, Y., et al. (2017). Temperature increase reduces global yields of major crops in four independent estimates. Proc. Natl. Acad. Sci. U. S. A. 114, 9326–9331. doi: 10.1073/pnas.1701762114
Keywords: agriculture, breeding, climate change resilience, crop improvement, neglected crops, phenomics, sustainability
Citation: Großkinsky DK, Faure J-D, Gibon Y, Haslam RP, Usadel B, Zanetti F and Jonak C (2023) The potential of integrative phenomics to harness underutilized crops for improving stress resilience. Front. Plant Sci. 14:1216337. doi: 10.3389/fpls.2023.1216337
Received: 03 May 2023; Accepted: 08 June 2023;
Published: 20 June 2023.
Edited by:
Kaixuan Zhang, Chinese Academy of Agricultural Sciences, ChinaReviewed by:
Federico Scossa, Research Center for Genomics and Bioinformatics (CREA-GB), ItalyCopyright © 2023 Großkinsky, Faure, Gibon, Haslam, Usadel, Zanetti and Jonak. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Dominik K. Großkinsky, ZG9taW5pay5ncm9zc2tpbnNreUBhaXQuYWMuYXQ=; Claudia Jonak, Y2xhdWRpYS5qb25ha0BhaXQuYWMuYXQ=
†ORCID: Dominik K. Großkinsky, orcid.org/0000-0001-8976-1057
Jean-Denis Faure, orcid.org/0000-0001-6867-6021
Richard P. Haslam, orcid.org/0000-0001-6226-5643
Björn Usadel, orcid.org/0000-0003-0921-8041
Federica Zanetti, orcid.org/0000-0003-4729-2082
Claudia Jonak, orcid.org/0000-0001-9605-6751
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.