 Shanshan Chen
Shanshan Chen Chun-Bo Zhao
Chun-Bo Zhao
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
REVIEW article
Front. Plant Sci. , 16 February 2023
Sec. Plant Abiotic Stress
Volume 14 - 2023 | https://doi.org/10.3389/fpls.2023.1141918
This article is part of the Research Topic Abiotic and Biotic Stress in Horticultural Crops: Insight into Recent Advances in the Underlying Tolerance Mechanism View all 46 articles
Horticultural crops are greatly disturbed by severe abiotic stress conditions. This is considered one of the major threats to the healthy lives of the human population. Salicylic acid (SA) is famous as one of the multifunctional phytohormones that are widely found in plants. It is also an important bio-stimulator involved in the regulation of growth and the developmental stages of horticultural crops. The productivity of horticultural crops has been improved with the supplemental use of even small amounts of SA. It has good capability to reduce oxidative injuries that occur from the over-production of reactive oxygen species (ROS), potentially elevated photosynthesis, chlorophyll pigments, and stomatal regulation. Physiological and biochemical processes have revealed that SA enhances signaling molecules, enzymatic and non-enzymatic antioxidants, osmolytes, and secondary metabolites activities within the cell compartments of plants. Numerous genomic approaches have also explored that SA regulates transcriptions profiling, transcriptional apprehensions, genomic expression, and metabolism of stress-related genes. Many plant biologists have been working on SA and its functioning in plants; however, its involvement in the enhancement of tolerance against abiotic stress in horticultural crops is still unidentified and needs more attention. Therefore, the current review is focused on a detailed exploration of SA in physiological and biochemical processes in horticultural crops subjected to abiotic stress. The current information is comprehensive and aims to be more supportive of the development of higher-yielding germplasm against abiotic stress.
Agriculture production is decreasing because of the rapid human population growth and recent global climate change. It is necessary to improve and boost the required agricultural outputs by nearly 70% by the mid-century to feed huge populations in the future (Bulgari et al., 2019). Moreover, the production of horticultural crops is badly affected by climate change. Crop growth and yield is reducing gradually due to elevated environmental extremities occurring from industrialization and urbanization. Several biotic (e.g., insect pests and diseases) and abiotic stresses (e.g., drought, salinity, heavy metals, and temperature extremities) are major constraints restricting the productivity of horticultural crops (Devi et al., 2022). One of the most challenging aspects for plant researchers is plant response under diverse environmental stress conditions. Water deficiency, an excess of salts, and temperature extremities are major abiotic constraints disturbing plant health at growth, yield, and quality stages (Mangal et al., 2022). The development, characterization, and evaluation of tolerant germplasm is a recent need against abiotic stresses. However, while some traditional strategies have been adopted for the development of tolerant germplasm, they are laborious, time-consuming, and not capable of eliminating multiple stresses (Ahmad and Anjum, 2018; Ahmad et al., 2023). Therefore, appropriate measures are being developed to fulfill the food needs of the world’s population. Moreover, it is imperative to develop suitable and appropriate strategies in the current era to alleviate the problems of intolerance in horticultural crops (Mahpara et al., 2019). Hence, the exogenous application of phytohormones (including salicylic acid, abscisic acid, ascorbic acid, melatonin, brassinosteroids, strigolactones, jasmonates, auxins, ethylene, cytokinins, and gibberellins) may possibly be an accurate and concise option for the production of climate-resilient crops with higher yields and excellent quality.
Among phytohormones, SA has been considered an alternative eco-friendly and suitable chemical regulator for the alleviation of abiotic stress tolerance issues in many horticultural crops. SA is effective even in very small amounts in numerous ways for the regulation of plant growth and developmental stages in response to stressful conditions, as reported by Jiang and Asami (2018). Wani et al. found SA to be more effective for the regulation of different signal transduction trails in response to plants growing under abiotic stress (2016). Regulation of external and internal stimuli is encouraged due to SA, which further contributes to major changes occurring in the developmental processes of horticultural crops (Kazan, 2013). SA is widely known as one of the signaling molecules in horticultural crops subjected to abiotic stress (Jahan et al., 2019). Therefore, it has been assumed that SA is appropriate and effective for the mitigation of the adversities of abiotic stress. SA is also beneficial for the quality production of horticultural crops (Li et al., 2021). Numerous physiological, biochemical, and photosynthetic pigments and molecular mechanisms were regulated by the supplemental use of SA against abiotic stress. SA is also famous as one of the promising phytohormones with an excellent capability to cope with the adversities that occur from abiotic stress. SA has the potential to enhance the generation of bioactive compounds and activate plant defense systems against stressful conditions (Mahpara et al., 2019). Therefore, SA supplemental use is effective and supportive for the excellent production of horticultural crops growing under abiotic stress conditions.
The supplemental application of SA is necessary for the sustainable production of horticultural crops subjected to abiotic stress conditions, as it improves plant defense systems. Therefore, the exogenous use of phytohormones (SA) is the most effective and promising method for the alleviation of abiotic stress intolerance in horticultural crops. Thus, there is a need to explore the physiological and biochemical responses of plants with the exogenous use of SA against abiotic stress for sustainable yields.
Horticultural crops are rich source of nutritional contents necessary for healthy life because of the higher nutritional value and capability of staple food for the biosphere. Horticultural crop production is disturbed globally due to abiotic stresses, i.e., water deficiencies, salinity, cold, heat, heavy metals, minerals deficiencies, UV light, and pesticides. Abiotic stresses are altering the morphological, physiological, anatomical, and biochemical processes in plants and have an adverse effect on production or plant senescence (Rao et al., 2016). It has been shown that plant response to stressful conditions is chiefly based on type, duration, level of stress, plant stage, and the genetic make-up of species/cultivars (Feller and Vaseva, 2014); this can result in restrictions in growth, poor and low yield, disturbances in developmental phases, the rupturing of photosynthetic pigments, membrane injuries, disruptions in photosynthesis and stomatal regulation, and a low water potential in leaves, as indicated by Wani et al. (2016).
Harsh climatic conditions are becoming more noticeable because of the rapid increase in pollution and extensive variations due to global climate change. For instance, water deficit conditions around the globe may possibly be raised due to persistent exposure to temperature extremities in rain-fed places. Higher temperature conditions in the future are likely to cause water shortages and excess salts in soils, as predicted by Raftery et al. (2017). Furthermore, they also revealed thattemperature is going to be increased due to global warming. However, production and quality decline in horticultural crops in the near future are likely to be due to multiple stresses like drought, salinity, cold, heat, mineral deficiencies, and heavy metals. Furthermore, it has been noted that 90% of agricultural regions are becoming more susceptible to single or multiple environmental stresses.
Transcriptional activities have a greater contribution to the tolerance to oxidative stress. Different researchers have revealed the significance of transcriptomic activities in the regulation of ABA production (Cao et al., 2014). Moreover, the ASC-GSH cycle is regulated at the maximum level in the tolerant-germplasm as compared to sensitive-germplasm, as reported by Mahajan and Sanejouand (2015). Moreover, it can also be used for the characterization of either tolerant or sensitive landraces for sustainable production (Gallardo et al., 2014).
Different management practices have been employed to cope with the adverse effects of abiotic stress in horticultural crops (Kazan, 2013). Agronomic practices, molecular approaches, and supplemental use of phytohormones are more appropriate practices for the alleviation of intolerance in horticultural crops against abiotic stress (Jiang and Asami, 2018; Jahan et al., 2019). Among phytohormones, SA is well known for its mitigation of adverse effects of abiotic stress. Previous plant researchers have revealed that SA has a good capability to regulate the uptake of minerals, stomata regulation, the excellent working of photosynthetic contents, the generation of metabolites, and the defense system by activating ROS scavengers against abiotic stress (Li et al., 2021). Therefore, appraising the associations of these environment-related extremities with plant morpho-physiological and biochemical responses is imperative for the improvement of numerous horticultural practices.
SA is a well-known molecule that can protect against abiotic stresses by acting as a signaling compound. Abiotic stress indication is necessary to cope with its adverse effects in a timely manner for higher yields with quality produce. Horticultural crops are rich in minerals and vitamins necessary for a healthy life. Therefore, management practices are imperative for higher yielding-germplasm of horticultural crops. Climate change and continuous cropping are threats to the productivity of horticultural crops through abiotic stress (Connor, 2002). Moreover, SA is not only important for the regulation and activation of the defense system of plants against biotic stress but also more effective and helpful for the improvement of abiotic stress tolerance in horticultural crops, as discussed by Horváth et al. (2007).
The fundamental appliances of SA-enhanced abiotic stress resistance comprise SA-mediated osmolytes generation and their accumulation. Osmolytes generation could be helpful for the maintenance of osmotic homeostasis, minerals and nutrients uptake and up-regulation, increasing the scavenging of ROS activities, and improving the productivity of secondary metabolites (such as glutathione, phenolics, phytoalexins, alkaloids, allicin, terpenes, thionins, defensins, and glucosinolates). SA signaling activates the osmolytes production necessary for the maintenance of numerous hormone pathways (Khan et al., 2015). Therefore, it has been assumed that SA is found to be an important signaling molecule, and also acts as a safeguard for the alleviation of intolerance in horticultural crops growing under climate-related extremities globally (Tables 1, 2).
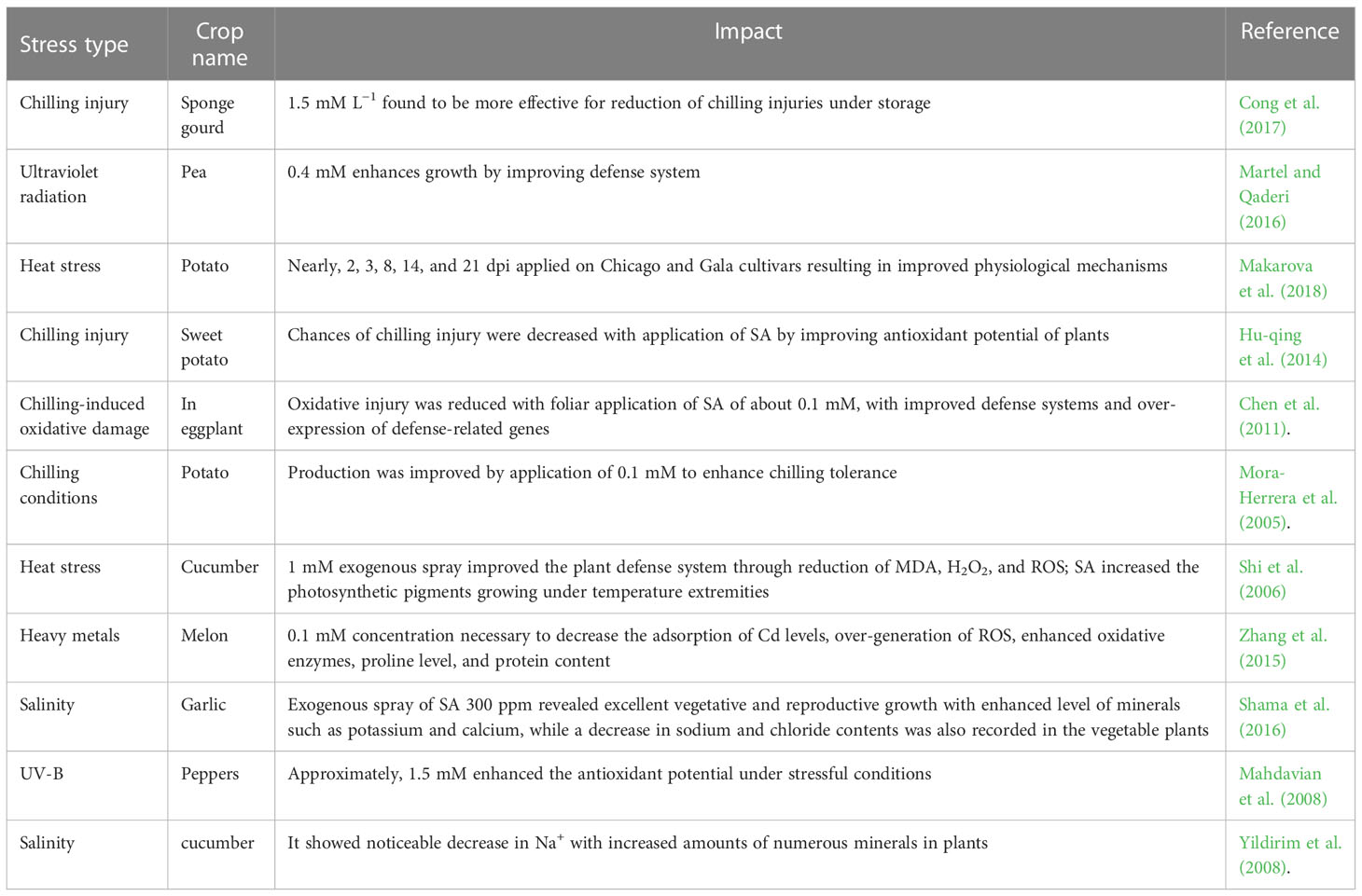
Table 1 Exogenous SA involvement against abiotic stress in vegetable production.
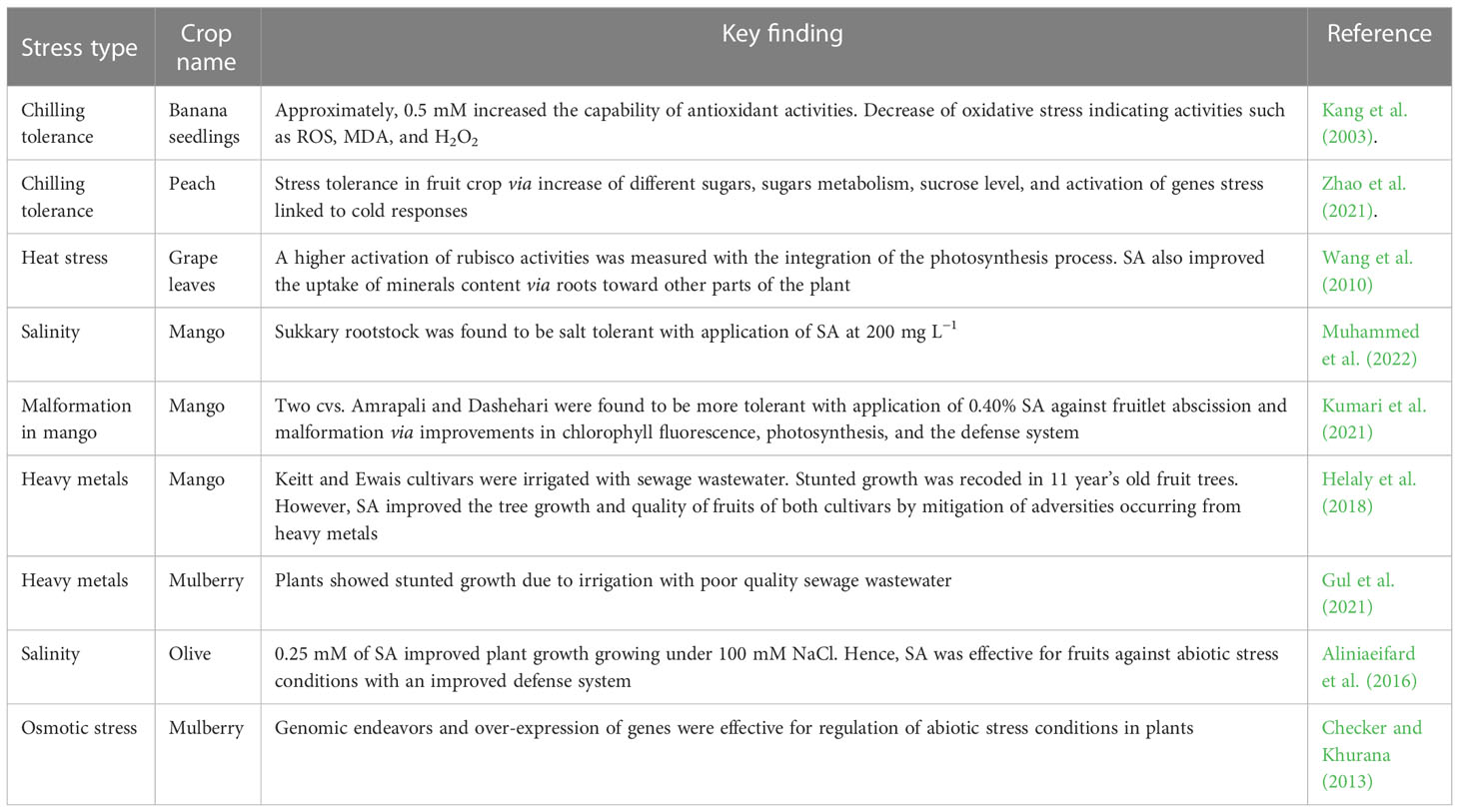
Table 2 Exogenous SA involvement against abiotic stress in fruit production.
Interestingly, SA has a large contribution to the expression of pathogens-related genes (PR) containing PR1, PR2, and PR5, as revealed by Ali et al. (2018). PR genes are fascinating due to their capability to reduce pathogen attacks. Furthermore, these genes are also involved in the reduction of abiotic stresses. Similarly, another study by Wu et al. (2016) also revealed that PR gene expression is also interesting and helpful for the mitigation of the negative effects of abiotic stress. Overexpression of transgenic tobacco PR-1 is also effective for tolerance against heavy metals in peppers, as revealed by Sarowar et al. (2005). In another study by Hong and Hwang (2005), it has been shown that pepper PR-1 overexpression is also involved in the improvement of tolerance in the Arabidopsis against salinity and drought stresses. However, more molecular approaches need to be investigated, and much is still necessary for the enhancement of tolerance against abiotic stress.
Plant microbes interaction with plant health is attracting more attention from plant researchers. However, research into the increased potential of microbes against abiotic stress is still limited and needs further attention from plant researchers. Some plant researchers have observed that SA is involved in the increase of SA levels in microbes, which is further necessary for plant health by increasing soil porosity and maximizing nutrient uptake chances in plants (Lundberg et al., 2012). Microbes enriched with endogenous SA levels had greater potential to increase plant tolerance against harsh climatic conditions either due to climate change or continuous cropping patterns (Lebeis et al., 2015).
Extremities of abiotic stress conditions result in excess toxic ROS. These potentially increase oxidative stress (Figure 1). However, ROS disturbs photosynthetic mechanisms, and the chance of osmotic stress occurs. Moreover, water stress conditions also majorly cause osmotic stress conditions in horticultural crops (Figure 2). Oxidative stress reduces plant growth, development, and yield. Cell membranes and electrolyte leakage are enhanced due to oxidative stress conditions. These conditions can damage parts or the whole of the plant (Muhammad et al., 2022).
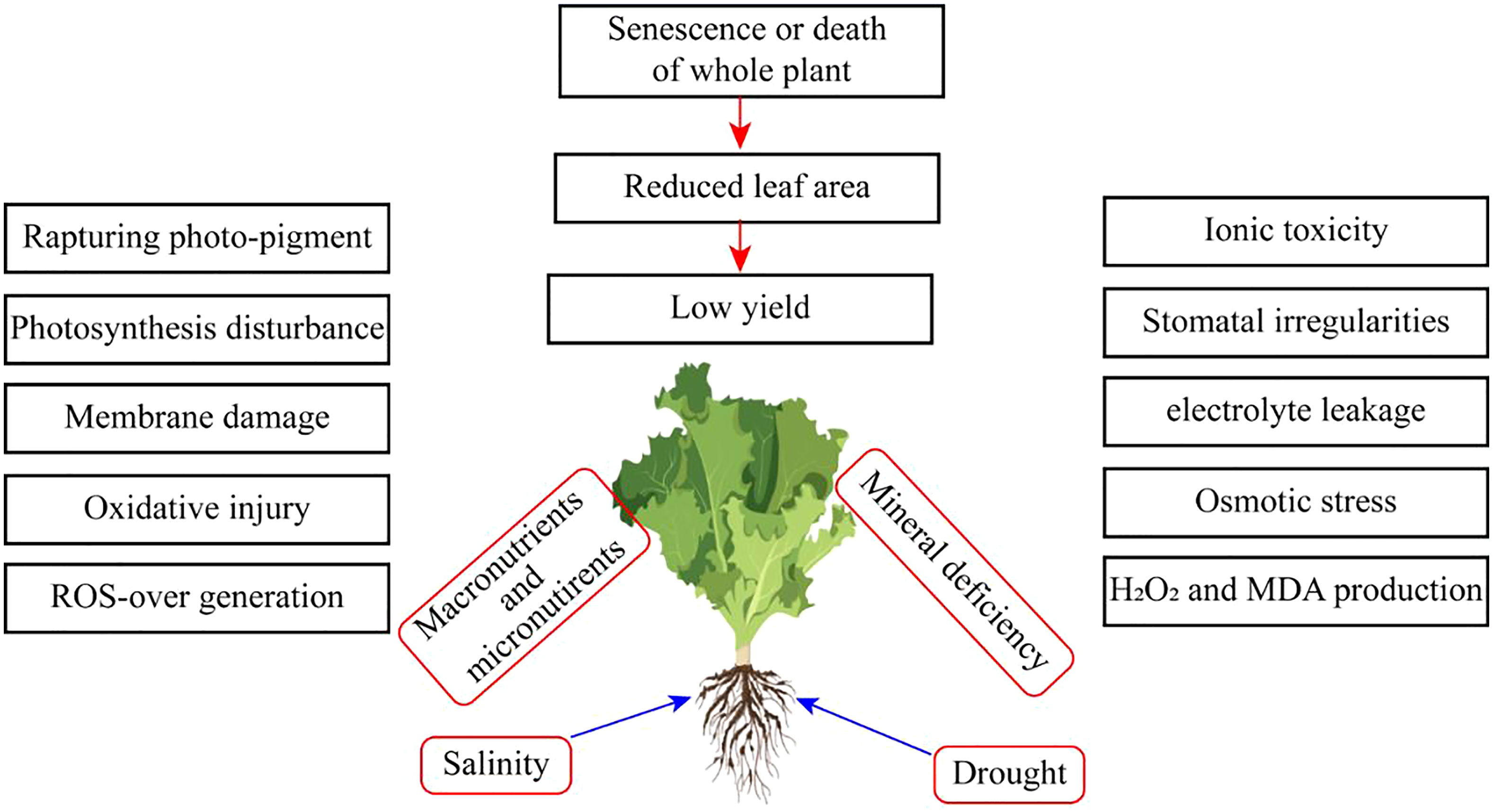
Figure 1 Effect of salinity and drought stress on horticultural crops.
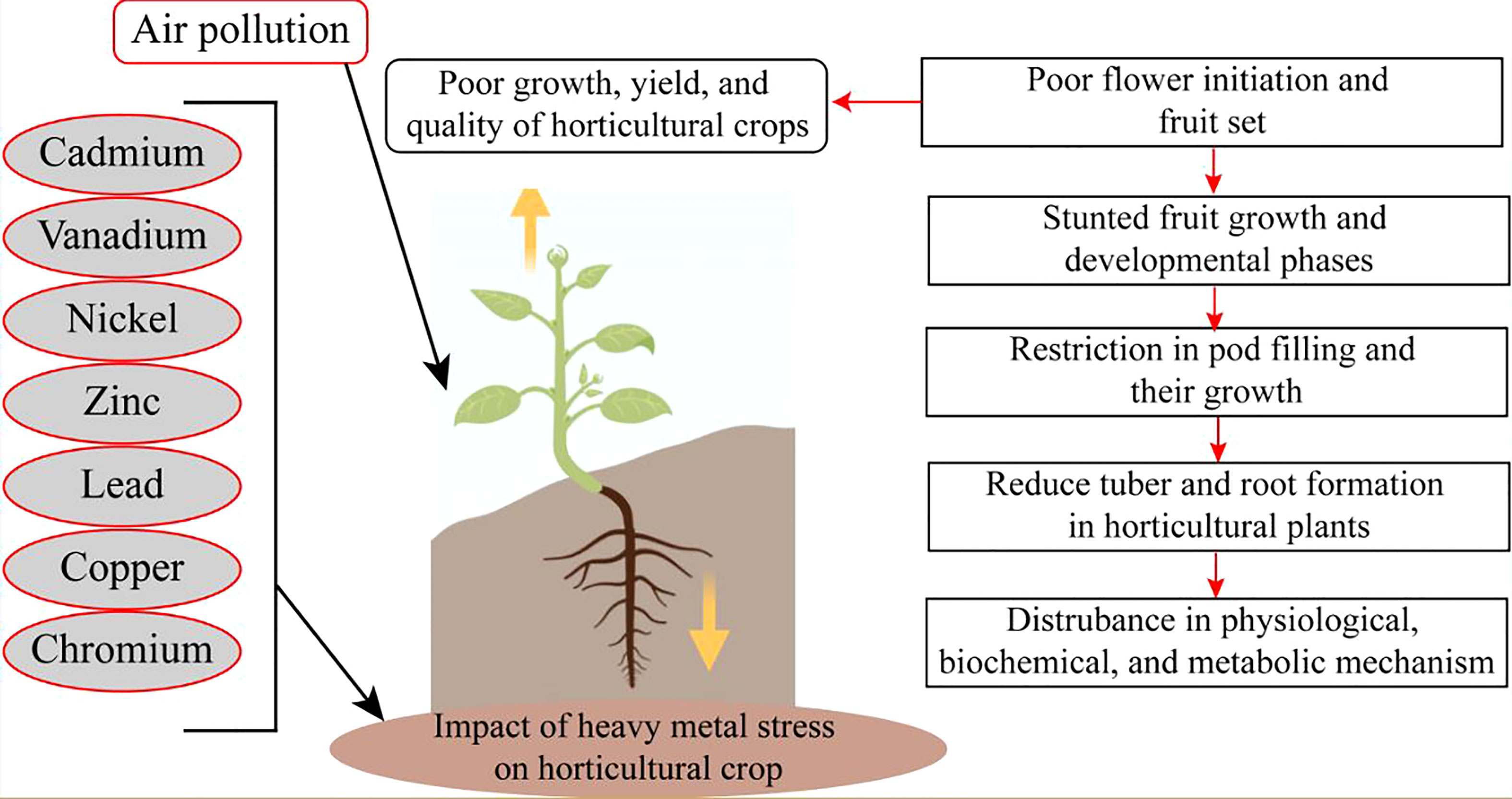
Figure 2 Heavy metals toxicity adversely affect the growth of horticultural crops.
Roots are considered as the first plant organ subjected to abiotic stress. Therefore, when exposed to abiotic stress, a higher reduction in growth and yield is observed in roots compared to other plant parts, like shoots. The elevated production of H2O2 reduces glutathione, and MDA indicates the extremities of oxidative stress conditions in the root zone, as studied by Ghosh et al. (2015). Elevated abiotic stress levels enhance the oxidative stress conditions in cells, compartments, and organelles. The increase of the accumulated photo-reducing effect is also due to oxidative stress. Furthermore, irregularities in the movement of electrons are mainly due to oxidative injuries in plants, as revealed by Sharma et al. (2012). Moreover, the Mehler reaction that occurs under abiotic stress deteriorates higher electrochemical energy and is considered one of the major causes of ROS, H2O2, and MDA activities at the extreme level.
The activation of enzymatic and non-enzymatic systems have the capability to trigger oxidative defense system under environmental stresses. However, it has been reported these responses to defense activities are chiefly organ specific. Polyphenols are extensively found in the root and young mature leaves. Therefore, these respond rapidly in roots and leaves compared to other plant parts. Polyphenol is famous as a non-enzymatic bioactive molecule involved in the defense system of plants (Tanou et al., 2009). However, the accumulation of tocopherols is not observed in roots because these are specific scavengers of singlet oxygen radicals in the photosystem II (Samira et al., 2015). In another study by Hussain et al. (2018), it was recorded that tocopherol accumulation was reduced in rice leaves due to excess salinity. Moreover, it has been found that plant roots have the potential to produce abscisic acid (ABA) under stressful conditions. ABA production is an indication of stress conditions in the root biosphere because it acts as a signaling molecule in plants under stress conditions.
Metabolic disturbances also occur due to oxidative extremities, as reported by Gallardo et al. (2014). Tolerant germplasm has the capability to produce an excess of tocopherols to cope with the adversities of stress conditions, as revealed by Sytykiewicz (2016). Furthermore, tocopherols have the potential to mitigate the negative effects of salinity even in seedling-stage plants. The exogenous application of SA is helpful to cope with the adversities that occur from abiotic stress. It is a multifunctional phytohormone with a diverse nature involved in the improvement of defense systems of numerous horticultural crops, i.e., bell pepper (Zhang et al., 2020), spinach (Gilani et al., 2020), peppermint (Ahmad et al., 2018), and potato (Li et al., 2019). Elevated growth, yield, photosynthesis, stomatal regulation, and protection from oxidative injury were due to the supplemental application of SA. Therefore, it can be assumed that SA is more effective for the alleviation of abiotic stress intolerance in horticultural crops focusing on higher yields with quality production (Figure 3).
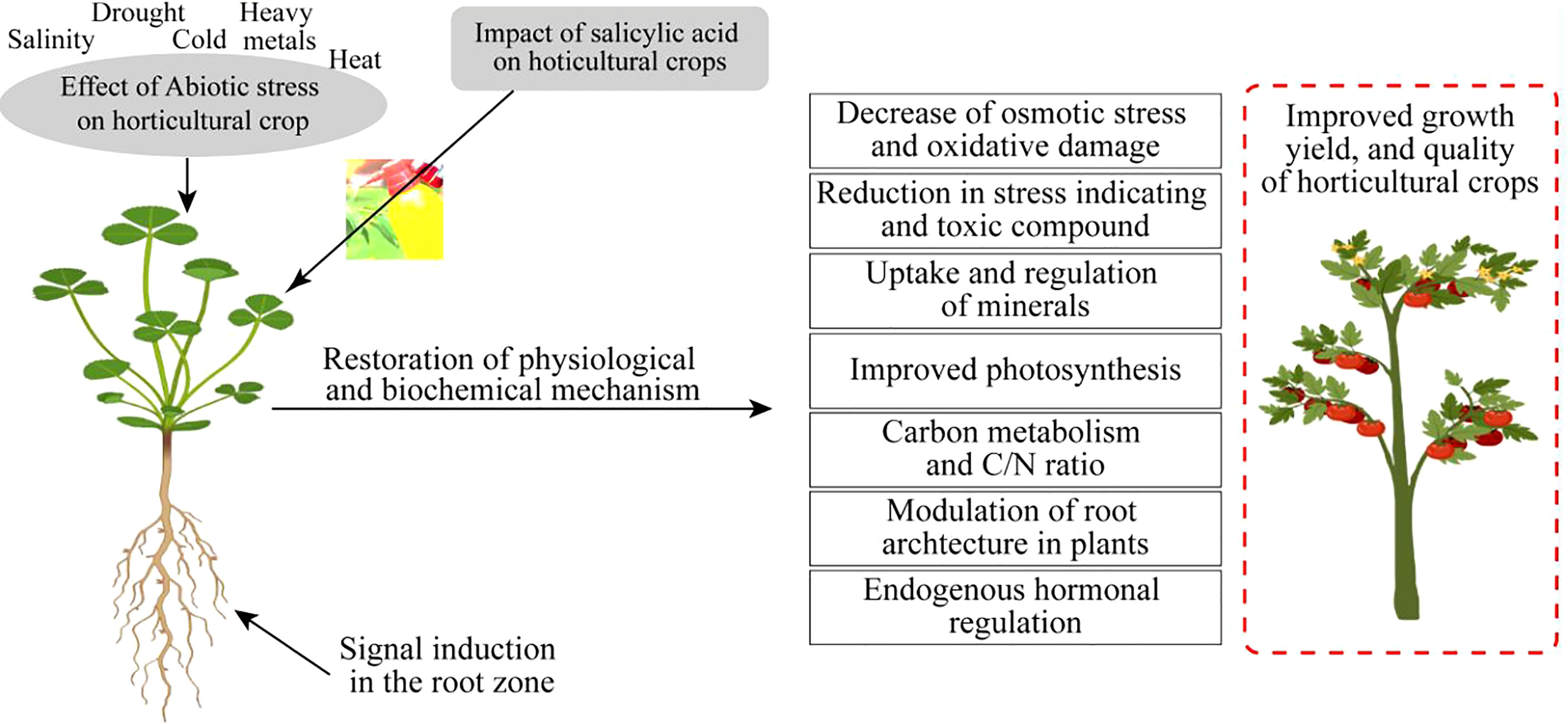
Figure 3 Impact of salicylic acid on horticultural crops under abiotic stress conditions.
SA is a multifaceted phenolic compound and naturally occurring molecule that signals against abiotic stress, as evaluated by Khan et al. (2015). SA contributes to the regular functioning of growth, photosynthetic adeptness, germination of seeds, roots enlargement, maturity and ripening, SA-microbes interaction, gravitropism maintenance, and responses to climate-related extremities, as described by Martel and Qaderi (2016). Supplemental use of about 1 mM of SA greatly increases water use efficiency (WUE), gas exchange mechanisms, activation of enzymatic activities, proline activation, and the reduction in oxidative injuries under extremities of temperature (heat stress) in tomatoes (Jahan et al., 2019). Biomass production is reduced due to drought stress. The disturbances in photosynthetic pigments and irregular stomatal functioning enhances the chances of osmotic stress. In another study, Galviz et al. (2021) demonstrated that 0.1 mM of SA improved biomass production, photosynthetic mechanism, enzymatic and non-enzymatic activities, and WUE, decreased cell-membrane damage, and provided better anatomical response under drought stress conditions. Similarly, Kaya (2021) revealed that SA had a better ability to induce drought tolerance in peppers by reducing oxidative injuries. Furthermore, photosynthetic behavior, enhanced proline levels, glyoxalase system, and regulation of stomata functioning were also reported with the utilization of SA in the peppers.
SA is effective for the mitigation of climatic adversities in temperate fruit crops by reducing oxidative and osmotic stress and improving defense systems. Chilling injury damage is most common in fruit crops grown in temperate regions. SA potentially provides tolerance to fruit crops through an increase in different sugars, sugars metabolism, sucrose levels, and the activation of genes related to cold responses, as described by Zhao et al. (2021). Chen et al. (2011) found that in eggplants growing under abiotic stress, oxidative injuries were reduced with a foliar spray of nearly 0.1 mM of SA (chilling injury), with improved defense systems and expression of defense-related genes. Potato production in chilling conditions was improved by the use of SA. Hence, 0.1 mM of SA was most effective to increase the chilling tolerance in plants, as demonstrated by Mora-Herrera et al. (2005). Similarly, another study by Shi et al. (2006) also evaluated that 1 mM of SA exogenously sprayed on cucumbers significantly improved the plant defense systems against heat stress through the reduction in MDA, H2O2, electrolyte leakage, and ROS. Moreover, SA improved the photosynthetic behavior of cucumbers against heat stress. Regarding heavy metals, Cd toxicity was reduced with 0.1 mM of SA in melons through the decreased adsorption of Cd, over-generation of ROS, and enhanced oxidative enzymes, proline levels, and protein content, as observed by Zhang et al. (2015). A better defense system indicated the tolerance level of a horticultural crop (potato) under stressful environments (heat stress) (Lopez-Delgado et al., 1998). Under heat stress, banana production improved with the exogenous use of SA, which improved the antioxidant pool, as examined by Kang et al. (2003). Furthermore, ROS, MDA, and H2O2 were reduced with SA application as a foliar spray. SA has the potential to mitigate adversities that occur due to abiotic stress by improving minerals uptake and ions homeostasis (Yalpani et al., 1994; Santisree et al., 2020).
The use of SA is an imperative way to lessen the adversities of abiotic stress and significantly enhance tolerance in horticultural crops of tropical and subtropical regions. A SA dose of 0.5 mM significantly enhances the activating potential of antioxidant activities in banana seedlings. However, the reduction in oxidative stress indicates activities such as ROS, MDA, and H2O2 (Kang et al., 2003). Moreover, higher activation of rubisco was recorded in grape leaves via the integration of photosynthesis under heat stress (Wang et al., 2010). SA improves mineral uptake from the roots toward other parts of the plant. Shama et al. (2016) demonstrated that an exogenous spray of SA 300 ppm revealed excellent vegetative and reproductive growth with enhanced level of minerals, such as potassium, while a decrease in sodium levels was recorded in garlic growing under salinity. SA is favorable for the growth of plants under normal or even stressful conditions. It produced a noticeable decrease in Na+ with increased amounts of numerous minerals in cucumbers against salinity stress, as described by Yildirim et al. (2008). A SA dose of 1.5 mM enhanced the antioxidant potential in peppers (Mahdavian et al., 2008), cucumbers (Lei et al., 2010), and peas (Martel and Qaderi, 2016) under UV-B stress.
Abiotic stress may be detected by measuring signaling molecules generated within the plant’s body. Osmolytes are important signaling molecules for abiotic stress in horticultural crops, especially fruit crops (Rabey et al., 2016). Disturbances in the osmotic conditions in the root’s biosphere are due to higher salt levels and water deficiency. Osmoprotectants are also naturally produced for the activation of the plant defense system against stressful conditions. Over-production of toxic ROS and H2O2 were regulated by the generation of osmolytes like proline, glycine betaine (GB), and ascorbates, and their regulation was an indication of tolerance to abiotic stress, as reported by Munns (2002).
Elevated production of osmolytes showed a major involvement in the regulation of stomata conductance, transpiration, and respiration rates (Fu et al., 2019). Production of ascorbates within the plant cells could decrease the generation of H2O2 and numerous other derivatives, as evaluated by Fatemi et al. (2021). Moreover, stress intolerance can be alleviated by the production of osmolytes within the plant compartments. Osmolytes production is effective for the removal of the toxic effects of MDA content in plants under abiotic stress. Furthermore, ROS reduction indicators were also observed in plants due to the production of osmolytes (Racchi, 2013). Tolerant germplasm had less MDA and H2O2 generation compared to sensitive germplasm (Abbas et al., 2015). SA can be effective in the reduction of MDA and H2O2 activities.
Proline contributes to the stabilization of numerous protein structures and membrane protection against the adversities of ROS activity. These are excellent scavengers of over-generated ROS within the plant cells (Mittler et al., 2004). Elevated proline concentration has been revealed in tolerant germplasm; however, sensitive germplasm has a very poor generation of proline content within the cells and compartments. Therefore, proline concentration supports coping with the negative effects of abiotic stress. Numerous plant biologists have revealed that proline is a stress-signaling molecule (Rahneshan et al., 2018). Osmolytes GB were generated in maximum amounts for plants growing under climate-related extremities examined by Karimi and Kuhbanani (2015). Similarly, in another study by Abdollahi and Takhti (2013), the stability of photosynthetic pigments and protein structures was found to be regulated by the productivity of GB content. The characterization, evaluation, and cultivation of tolerant germplasm is vital for higher yields.
Phytohormones are important for increasing the antioxidant potential that promotes the defense systems of plants. These contribute to the scavengers of over-generated ROS and H2O2 (Haneklaus et al., 2007). Supplemental application of SA in plants also improved the antioxidant profiling and regulation of stomata conductance and photosynthesis, as observed by Molassiotis et al. (2016). Hence, SA is necessary for the improvement of defense processes of plants cultivated under abiotic stress for excellent growth and elevated yields. The exogenous use of SA on horticultural crops is still limited, and little information currently exists. Hence, the present review inspires the utilization of SA on horticultural crops for enhanced defense systems.
Endogenous hormonal regulation is necessary in cucumbers to increase tolerance against abiotic stress, as described by Sharif et al. (2022). The excellent potential of hormonal regulation was reported in two Zizyphus species when compared to others. The regulation of hormones could be involved in the maintenance of stomata conductance and photosynthetic activities, as reported in a study by Meena et al. (2003). Among supplemental phytohormones, SA is vital for enhancing resistance against stressful plant conditions. SA spray improves the antioxidant potential and also improves the scavenging potential for ROS, H2O2, and MDA, as reported by Mansour (2000). Similarly, Verma et al. (2018) reported that a reduction in stress indicators was due to hormonal regulation in fruit crops. Moreover, hormonal regulation and molecular programming are effective for abiotic stress tolerance in plants (Raza et al., 2022a; Raza et al., 2022b). Therefore, the use of SA and its exploration is important for the mitigation of adversities in stressful conditions focusing on crop yields. The application of SA at 100 mg L-1 has been evaluated by Rehman et al. (2011) to have significant and positive effects on the improvement of seed germination and the overall production of cucumbers growing in stressful conditions. The production of eggplants was increased with the improvement of endogenous hormonal levels by the application of 0.3% SA under stressful conditions, as evaluated by Chen et al. (2011).
The adversities of biotic and abiotic stresses are increasing due to the fluctuations in climate change in the present era. Different management practices can be used for the reduction of the negative impacts of abiotic stress on horticultural crops. Among these, supplemental use of phytohormones is effective for the sustainable production of horticultural crops against climate-related extremities. Among phytohormones, SA is more helpful and supportive in nature, and involved in the alleviation of stress intolerance in plants. It has been recommended that a small amount of SA has an excellent capability to cope with adversities that occur due to abiotic stress, and this is a good way to ensure the sustainable production of agricultural crops globally.
SC and C-BZ: conceptualization, literature survey, writing original draft. SC and R-MR: writing – review and editing, figure designing. R-MR and J-HJ: figure designing, writing – review and editing. All authors contributed to the article and approved the submitted version.
This work was sponsored in part by Jilin Province Science and Technology Development Plan Project (20220202098NC),Jilin Province Modern Industrial Technology System Project (201619).
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Abbas, M. F., Jasim, A. M., Shareef, H. J. (2015). Role of sulphur in salinity tolerance of date palm (Phoenix dactylifera l.) offshoots cvs. berhi and sayer. Int. J. Agric. Food Sci. 5, 92–97.
Abdollahi, F., Takhti, G. S. (2013). Effect of GA3 on growth and chemical composition of jujube leaf (Ziziphus spina-christi) under salinity condition. J. Plant Proc. Fun. 2, 53–64.
Ahmad, R., Anjum, M. A. (2018). Applications of molecular markers to assess genetic diversity in vegetable and ornamental crops-a review. J. Hortic. Technol. 1, 1–7. doi: 10.46653/jhst180101001
Ahmad, I., Ercisli, S., Anjum, M. A., Ahmad, R. (2023). Progress in the methods of jujube breeding. Erwerbs-Obstbau. doi: 10.1007/s10341-022-00796-2
Ahmad, B., Jaleel, H., Sadiq, Y., Khan, M. M., Shabbir, A. (2018). Response of exogenous salicylic acid on cadmium induced photosynthetic damage, antioxidant metabolism and essential oil production in peppermint. Plant Growth Regul. 86, 273–286. doi: 10.1007/s10725-018-0427-z
Ali, S., Ganai, B. A., Kamili, A. N., Bhat, A. A., Mir, Z. A., Bhat, J. A., et al. (2018). Pathogenesis-related proteins and peptides as promising tools for engineering plants with multiple stress tolerance. Microbiol. Res. 212, 29–37. doi: 10.1016/j.micres.2018.04.008
Aliniaeifard, S., Hajilou, J., Tabatabaei, S. J. (2016). Photosynthetic and growth responses of olive to proline and salicylic acid under salinity condition. Not. Bot. Hortic. Agrobotanic. Cluj-Nap. 44, 579–585. doi: 10.15835/nbha44210413
Bulgari, R., Franzoni, G., Ferrante, A. (2019). Biostimulants application in horticultural crops under abiotic stress conditions. Agron. 9, 306. doi: 10.3390/agronomy9060306
Cao, M. J., Wang, Z., Zhao, Q., Mao, J. L., Speiser, A., Wirtz, M., et al. (2014). Sulfate availability affects ABA levels and germination response to ABA and salt stress in Arabidopsis thaliana. Plant J. 77, 604–615. doi: 10.1111/tpj.12407
Checker, V. G., Khurana, P. (2013). Molecular and functional characterization of mulberry EST encoding remorin (MiREM) involved in abiotic stress. Plant Cell Rep. 32, 1729–1741. doi: 10.1007/s00299-013-1483-5
Chen, S., Zimei, L., Cui, J., Jiangang, D., Xia, X., Liu, D., et al. (2011). Alleviation of chilling-induced oxidative damage by salicylic acid pretreatment and related gene expression in eggplant seedlings. Plant Growth Regul. 65, 101–108. doi: 10.1007/s10725-011-9579-9
Cong, H. A. N., Zuo, J. H., Qing, W., Dong, H. Z., Gao, L. P. (2017). Salicylic acid alleviates postharvest chilling injury of sponge gourd (Luffa cylindrica). J. Int. Agric. 16, 735–741. doi: 10.1016/S2095-3119(16)61390-4
Devi, R., Behera, B., Raza, M. B., Mangal, V., Altaf, M. A., Kumar, R., et al. (2022). An insight into microbes mediated heavy metal detoxification in plants: A review. J. Soil Sci. Plant Nutr. 22, 914–936. doi: 10.1007/s42729-021-00702-x
Fatemi, H., Esmaiel Pour, B., Rizwan, M. (2021). Foliar application of silicon nanoparticles affected the growth, vitamin c, flavonoid, and antioxidant enzyme activities of coriander (Coriandrum sativum l.) plants grown in lead (Pb)-spiked soil. Environ. Sci. Poll. Res. 28 (2), 1417–1425. doi: 10.1007/s11356-020-10549-x
Feller, U., Vaseva, I. I. (2014). Extreme climatic events: impacts of drought and high temperature on physiological processes in agronomically important plants. Front. Environ. Sci. 2. doi: 10.3389/fenvs.2014.00039
Fu, Q. Q., Tan, Y. Z., Zhai, H., Du, Y. P. (2019). Evaluation of salt resistance mechanisms of grapevine hybrid rootstocks. Sci. Hortic. 243, 148–158. doi: 10.1016/j.scienta.2018.07.034
Gallardo, K., Courty, P. E., Le Signor, C., Wipf, D., Vernoud, V. (2014). Sulfate transporters in the plant’s response to drought and salinity: regulation and possible functions. Front. Plant Sci. 5. doi: 10.3389/fpls.2014.00580
Galviz, Y. C., Bortolin, G. S., Guidorizi, K. A., Deuner, S., Reolon, F., de Moraes, D. M. (2021). Effectiveness of seed priming and soil drench with salicylic acid on tomato growth, physiological and biochemical responses to severe water deficit. J. Soil Sci. Plant Nutr. 21, 2364–2377. doi: 10.1007/s42729-021-00528-7
Gilani, M., Danish, S., Ahmed, N., Rahi, A. A., Akrem, A., Younis, U., et al. (2020). Mitigation of drought stress in spinach using individual and combined applications of salicylic acid and potassium. Pak. J. Bot. 52, 1505–1513. doi: 10.30848/PJB2020-5(18
Gul, A. S., Naveed, F., Ali, M., Ahmad, R., Saqib, M. (2021). Effect of different wastewater irrigation regimes on growth of mulberry (Morus macroura miq.). Erwerbs-Obstbau 63, 331–337. doi: 10.1007/s10341-021-00577-3
Haneklaus, S., Bloem, E., Schnug, E., Kok, L. J., de Stulen, I. (2007). “Sulfur,” in Handbook of plant nutrition. Eds. Barker, A. V., Pilbeam, D. J. (New York: Taylor and Francis), 183–223.
Helaly, M. N., El-Sheery, N. I., El-Hoseiny, H., Rastogi, A., Kalaji, H. M., Zabochnicka-Świątek, M. (2018). Impact of treated wastewater and salicylic acid on physiological performance, malformation and yield of two mango cultivars. Sci. Hortic. 233, 159–177. doi: 10.1016/j.scienta.2018.01.001
Hong, J. K., Hwang, B. K. (2005). Induction of enhanced disease resistance and oxidative stress tolerance by overexpression of pepper basic PR-1 gene in arabidopsis. Physiol. Plant 124, 267–277. doi: 10.1111/j.1399-3054.2005.00515.x
Horváth, E., Szalai, G., Janda, T. (2007). Induction of abiotic stress tolerance by salicylic acid signaling. J. Plant Growth Regul. 26, 290–300. doi: 10.1007/s00344-007-9017-4
Hu-qing, Y., Xiao-fei, Z., Cheng-qian, H., Guo-quan, L. U., Jian-nai, H. (2014). Effects of different treatments on chilling injury and antioxidative metabolism in sweet potato. J. Nuc. Agric. Sci. 28, 1407. doi: 10.11869/j.issn.100-8551.2014.08.1407
Hussain, M., Ahmad, S., Hussain, S., Lal, R., Ul-Allah, S., Nawaz, A. (2018). Rice in saline soils: physiology, biochemistry, genetics, and management. Adv. Agron. 148, 231–287. doi: 10.1016/bs.agron.2017.11.002
Jahan, M. S., Wang, Y., Shu, S., Zhong, M., Chen, Z., Wu, J., et al. (2019). Exogenous salicylic acid increases the heat tolerance in tomato (Solanum lycopersicum l) by enhancing photosynthesis efficiency and improving antioxidant defense system through scavenging of reactive oxygen species. Sci. Hortic. 247, 421–429. doi: 10.1016/j.scienta.2018.12.047
Jiang, K., Asami, T. (2018). Chemical regulators of plant hormones and their applications in basic research and agriculture. Biosci. Biotechnol. Biochem. 82, 1265–1300. doi: 10.1080/09168451.2018.1462693
Kang, G., Wang, C., Sun, G., Wang, Z. (2003). Salicylic acid changes activities of H2O2-metabolizing enzymes and increases the chilling tolerance of banana seedlings. Environ. Exp. Bot. 50, 9–15. doi: 10.1016/S0098-8472(02)00109-0
Karimi, H. R., Kuhbanani, A. M. (2015). The evaluation of inter-specific hybrid of P. atlantica p. vera cv. ‘Badami zarand’ as a pistachio rootstock to salinity stress. J. Nutr. 6, 113–122. doi: 10.22034/JON.2015.516319
Kaya, C. (2021). Nitrate reductase is required for salicylic acid-induced water stress tolerance of pepper by upraising the AsA-GSH pathway and glyoxalase system. Physiol. Plant 172, 351–370. doi: 10.1111/ppl.13153
Kazan, K. (2013). Auxin and the integration of environmental signals into plant root development. Annal. Bot. 112, 1655–1665. doi: 10.1093/aob/mct229
Khan, M. I. R., Fatma, M., Per, T. S., Anjum, N. A., Khan, N. A. (2015). Salicylic acid-induced abiotic stress tolerance and underlying mechanisms in plants. Front. Plant Sci. 6. doi: 10.3389/fpls.2015.00462
Kumari, S., Singh, A. K., Kumar, A., Singh, K. P., Bains, G. (2021). Evaluating the efficacy of chitosan and salicylic acid on photosynthetic pigments and antioxidant enzymes towards resistance of mango malformation. Sci. Hortic. 285, 110160. doi: 10.1016/j.scienta.2021.110160
Lebeis, S. L., Paredes, S. H., Lundberg, D. S., Breakfield, N., Gehring, J., McDonald, M., et al. (2015). Salicylic acid modulates colonization of the root microbiome by specific bacterial taxa. Sci. 349, 860–864. doi: 10.1126/science.aaa8764
Lei, T., Feng, H., Sun, X., Dai, Q. L., Zhang, F., Liang, H. G., et al. (2010). The alternative pathway in cucumber seedlings under low temperature stress was enhanced by salicylic acid. Plant Growth Regul. 603, 5–42. doi: 10.1007/s10725-009-
Li, Q., Wang, G., Wang, Y., Yang, D., Guan, C., Ji, J. (2019). Foliar application of salicylic acid alleviate the cadmium toxicity by modulation the reactive oxygen species in potato. Ecotoxicol. Environ. Saf. 172, 317–325. doi: 10.1016/j.ecoenv.2019.01.078
Li, Z. G., Xiang, R. H., Wang, J. Q. (2021). Hydrogen sulfide–phytohormone interaction in plants under physiological and stress conditions. J. Plant Growth Regul. 40, 1–9. doi: 10.1007/s00344-021-10350-1
Lopez-Delgado, H., Dat, J. F., Foyer, C. H., Scott, I. M. (1998). Induction of thermotolerance in potato microplants by acetylsalicylic acid and H2O2. J. Exp. Bot. 49, 713–720. doi: 10.1093/jxb/49.321.713
Lundberg, D. S., Lebeis, S. L., Paredes, S. H., Yourstone, S., Gehring, J., Malfatti, S., et al. (2012). Defining the core arabidopsis thaliana root microbiome. Nat. 488, 86–90. doi: 10.1038/nature11237
Mahajan, S., Sanejouand, Y. H. (2015). On the relationship between low-frequency normal modes and the large-scale conformational changes of proteins. Arch. Biochem. Biophysic. 567, 59–65. doi: 10.1016/j.abb.2014.12.020
Mahdavian, K., Ghorbanli, M., Kalantari, K. M. (2008). Role of salicylic acid in regulating ultraviolet radiation-induced oxidative stress in pepper leaves. Russ. J. Plant Physiol. 55, 560–563. doi: 10.1134/S1021443708040195
Mahpara, S., Shahnawaz, M., Rehman, K., Ahmad, R., Khan, F. U. (2019). Nitrogen fertilization induced drought tolerance in sunflower: A review. Pure Appl. Biol. 8, 1675–1683. doi: 10.19045/bspab.2019.80110
Makarova, S., Makhotenko, A., Spechenkova, N., Love, A. J., Kalinina, N. O., Taliansky, M. (2018). Interactive responses of potato (Solanum tuberosum l.) plants to heat stress and infection with potato virus y. Front. Microbiol. 9. doi: 10.3389/fmicb.2018.02582
Mangal, V., Lal, M. K., Tiwari, R. K., Altaf, M. A., Sood, S., Kumar, D., et al. (2022). Molecular insights into the role of reactive oxygen, nitrogen and sulphur species in conferring salinity stress tolerance in plants. J. Plant Growth Regul., 1–21. doi: 10.1007/s00344-022-10591-8
Mansour, M. M. F. (2000). Nitrogen containing compounds and adaptation of plants to salinity stress. Biol. Plant 43, 491–500. doi: 10.1023/A:1002873531707
Martel, A. B., Qaderi, M. M. (2016). Does salicylic acid mitigate the adverse effects of temperature and ultraviolet-b radiation on pea (Pisum sativum) plants? Environ. Exp. Bot. 122, 39–48. doi: 10.1016/j.envexpbot.2015.09.002
Meena, S. K., Gupta, N. K., Gupta, S., Khandelwal, S. K., Sastry, E. V. D. (2003). Effect of sodium chloride on the growth and gas exchange of young Ziziphus seedling rootstocks. J. Hortic. Sci. Biotechnol. 78, 454–457. doi: 10.1080/14620316.2003.11511649
Mittler, R., Vanderauwera, S., Gollery, M., Van Breusegem, F. (2004). Reactive oxygen gene network of plants. Trend. Plant Sci. 9, 490–498. doi: 10.1016/j.tplants.2004.08.009
Molassiotis, A., Job, D., Ziogas, V., Tanou, G. (2016). Citrus plants: a model system for unlocking the secrets of NO and ROS-inspired priming against salinity and drought. Front. Plant Sci. 7. doi: 10.3389/fpls.2016.00229
Mora-Herrera, M. E., López-Delgado, H., Castillo-Morales, A., Foyer, C. H. (2005). Salicylic acid and H2O2 function by independent pathways in the induction of freezing tolerance in potato. Physiol. Plant 125, 430–440. doi: 10.1111/j.1399-3054.2005.00572.x
Muhammad, H. M. D., Abbas, A., Ahmad, R. (2022). Fascinating role of silicon nanoparticles to mitigate adverse effects of salinity in fruit trees: A mechanistic approach. Silicon 14, 8319–8326. doi: 10.1007/s12633-021-01604-4
Muhammed, M. A., Mohamed, A. K. S., Qayyum, M. F., Haider, G., Ali, H. A. (2022). Physiological response of mango transplants to phytohormones under salinity stress. Sci. Hortic. 296, 110918. doi: 10.1016/j.scienta.2022.110918
Munns, R. (2002). Comparative physiology of salt and water stress. Plant Cell. Environ. 25, 239–250. doi: 10.1046/j.0016-8025.2001.00808.x
Rabey, E. H. A., Al-Malki, A. L., Abulnaja, K. O. (2016). Proteome analysis of date palm (Phoenix dactylifera l.) under severe drought and salt stress. Int. J. Genom. 2016, 1–8. doi: 10.1155/2016/7840759
Racchi, M. L. (2013). Antioxidant defenses in plants with attention to Prunus and. Citrus Antioxidants spp, 2. doi: 10.3390/antiox2040340
Raftery, A. E., Zimmer, A., Frierson, D. M. W., Startz, R., Liu, P. (2017). Less than 2 °C warming by 2100 unlikely. Nat. Clim. Change 7, 637. doi: 10.1038/nclimate3352
Rahneshan, Z., Nasibi, F., Moghadam, A. A. (2018). Effects of salinity stress on some growth, physiological, biochemical parameters and nutrients in two pistachio (Pistacia vera l.) rootstocks. J Plant Interact. 13, 73–82. doi: 10.1080/17429145.2018.1424355
Rao, N. S., Shivashankara, K. S., Laxman, R. H. (2016). Abiotic stress physiology of horticultural crops, vol 311 (India: Springer). doi: 10.1007/978-81-322-2725-0
Raza, A., Salehi, H., Rahman, M. A., Zahid, Z., Madadkar, H. M. (2022b). Plant hormones and neurotransmitter interactions mediate antioxidant defenses under induced oxidative stress in plants. Front. Plant Sci. 13. doi: 10.3389/fpls.2022.961872
Raza, A., Tabassum, J., Fakhar, A. Z., Sharif, R., Chen, H. (2022a). Smart reprograming of plants against salinity stress using modern biotechnological tools. Critic. Rev. Biotechnol., 1–28. doi: 10.1080/07388551.2022.2093695
Rehman, H., Farooq, M., Basra, S. M. A., Afzal, I. (2011). Hormonal priming with salicylic acid improves the emergence and early seedling growth in cucumber. J. Agric. Soc Sci. 7, 109–113.
Samira, M., Hichem, H., Boughalleb, F., Mounir, D. (2015). Effect of light-salinity interaction on photosystem activity of excised corn leaves. Afr. Crop Sci. J. 23, 343–354. doi: 10.4314/acsj.v23i4.4
Santisree, P., Jalli, L. C. L., Bhatnagar-Mathur, P., Sharma, K. K. (2020). Emerging roles of salicylic acid and jasmonates in plant abiotic stress responses. Protective chemical agents in: The amelioration of plant abiotic stress: Biochemical and molecular perspectives. Wiley Online Library. pp. doi: 10.1002/9781119552154.ch17
Sarowar, S., Kim, Y. J., Kim, E. N., Kim, K. D., Hwang, B. K., Islam, R., et al. (2005). Overexpression of a pepper basic pathogenesis-related protein 1 gene in tobacco plants enhances resistance to heavy metal and pathogen stresses. Plant Cell Rep. 24, 216–224. doi: 10.1007/s00299-005-0928-x
Shama, A. M., Moussa, S., Abo-El-Fadel, N. I. (2016). Salicylic acid efficacy on resistance of garlic plants (Allium sativum l.) to water salinity stress on growth, yield and its quality. Alexand. Sci. Exch. J. 37, 165–174. doi: 10.21608/asejaiqjsae.2016.2238
Sharif, R., Su, L., Chen, X., Qi, X. (2022). Involvement of auxin in growth and stress response of cucumber. Veget. Res. 2, 1–8. doi: 10.48130/VR-2022-0013
Sharma, P., Jha, A. B., Dubey, R. S., Pessarakli, M. (2012). Reactive oxygen species, oxidative damage, and antioxidative defense mechanism in plants under stressful conditions. J. Bot. 2012, 110–110. doi: 10.1155/2012/217037
Shi, Q., Bao, Z., Zhu, Z., Ying, Q., Qian, Q. (2006). Effects of different treatments of salicylic acid on heat tolerance, chlorophyll fluorescence, and antioxidant enzyme activity in seedlings of Cucumis sativa l. Plant Growth Regul. 48, 127–135. doi: 10.1007/s10725-005-5482-6
Sytykiewicz, H. (2016). Expression patterns of genes involved in ascorbate-glutathione cycle in aphid-infested maize (Zea mays l.) seedlings. Int. J. Mol. Sci. 17, 268. doi: 10.3390/ijms17030268
Tanou, G., Molassiotis, A., Diamantidis, G. (2009). Induction of reactive oxygen species and necrotic death-like destruction in strawberry leaves by salinity. Environ. Exp. Bot. 65, 270–281. doi: 10.1016/j.envexpbot.2008.09.005
Verma, S. S., Verma, R. S., Verma, S. K., Yadav, A. L., Verma, A. K. (2018). Impact of salt stress on plant establishment, chlorophyll and total free amino acid content of ber (Zizyphus mauritiana lamk.) cultivars. J. Pharm. Phytochem. 7, 556–559.
Wang, L. J., Fan, L., Loescher, W., Duan, W., Liu, G. J., Cheng, J. S., et al. (2010). Salicylic acid alleviates decreases in photosynthesis under heat stress and accelerates recovery in grapevine leaves. BMC Plant Biol. 10, 1–10. doi: 10.1186/1471-2229-10-34
Wani, S. H., Kumar, V., Shriram, V., Sah, S. K. (2016). Phytohormones and their metabolic engineering for abiotic stress tolerance in crop plants. Crop J. 4, 162–176. doi: 10.1016/j.cj.2016.01.010
Wu, J., Kim, S. G., Kang, K. Y., Kim, J. G., Park, S. R., Gupta, R., et al. (2016). Overexpression of a pathogenesis-related protein 10 enhances biotic and abiotic stress tolerance in rice. Plant Pathol. J. 32, 552. doi: 10.5423/PPJ.OA.06.2016.0141
Yalpani, N., Enyedi, A. J., León, J., Raskin, I. (1994). Ultraviolet light and ozone stimulate accumulation of salicylic acid, pathogenesis-related proteins and virus resistance in tobacco. Plant. 193 (3), 372–376. doi: 10.1007/BF00201815
Yildirim, E., Turan, M., Guvenc, I. (2008). Effect of foliar salicylic acid applications on growth, chlorophyll, and mineral content of cucumber grown under salt stress. J. Plant Nutrit. 31, 593–612. doi: 10.1080/01904160801895118
Zhang, Z., Lan, M., Han, X., Wu, J., Wang-Pruski, G. (2020). Response of ornamental pepper to high-temperature stress and role of exogenous salicylic acid in mitigating high temperature. J. Plant Growth Regul. 39, 133–146. doi: 10.1007/s00344-019-09969-y
Zhang, Y., Xu, S., Yang, S., Chen, Y. (2015). Salicylic acid alleviates cadmium-induced inhibition of growth and photosynthesis through upregulating antioxidant defense system in two melon cultivars (Cucumis melo l.). Protoplasm. 252, 911–924. doi: 10.1007/s00709-014-0732-y
Keywords: abiotic stress, osmotic stress, poor yield, plant metabolism, plant defense activities
Citation: Chen S, Zhao C-B, Ren R-M and Jiang J-H (2023) Salicylic acid had the potential to enhance tolerance in horticultural crops against abiotic stress. Front. Plant Sci. 14:1141918. doi: 10.3389/fpls.2023.1141918
Received: 11 January 2023; Accepted: 02 February 2023;
Published: 16 February 2023.
Edited by:
Muhammad Ahsan Altaf, Hainan University, ChinaReviewed by:
Imran Khan, Lanzhou University, ChinaCopyright © 2023 Chen, Zhao, Ren and Jiang. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Shanshan Chen, Y3NzamxhdUBzaW5hLmNvbQ==
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.