 Kai Dong
Kai Dong Wenjin Li
Wenjin Li Yulong Tang1
Yulong Tang1 Suhui Ma
Suhui Ma
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Plant Sci., 15 February 2023
Sec. Functional Plant Ecology
Volume 14 - 2023 | https://doi.org/10.3389/fpls.2023.1140462
This article is part of the Research TopicPatterns, Functions, and Processes of Alpine Grassland Ecosystems under Global Change.View all 76 articles
Introduction: Over the past three decades, the view of nutrient limitation has transferred from single-nutrient limitation to multiple-nutrient limitation. On the Qinghai-Tibetan Plateau (QTP), many nitrogen (N) and phosphorus (P) addition experiments have revealed different N- or P-limited patterns at many alpine grassland sites, whereas it is not clear what the general patterns of N and P limitation across the QTP grasslands.
Methods: We performed a meta-analysis, containing 107 publications, to assess how N and P constrained plant biomass and diversity in alpine grasslands across the QTP. We also tested how mean annual precipitation (MAP) and mean annual temperature (MAT) influence N and P limitations.
Results: The findings show that plant biomass in QTP grasslands is co-limited by N and P. Single N limitation is stronger than single P limitation, and the combined positive effect of N and P addition is stronger than that of single nutrient additions. The response of biomass to N fertilization rate shows an increase firstly and then declines, and peaks at approximately 25 g N·m-2·year-1. MAP promotes the effect of N limitation on plant aboveground biomass and diminishes the effect of N limitation on belowground biomass. Meanwhile, N and P addition generally decline plant diversity. Moreover, the negative response of plant diversity to N and P co-addition is strongest than that of single nutrient additions.
Discussion: Our results highlight that N and P co-limitation is more prevalent than N- or P-limitation alone in alpine grasslands on the QTP. Our findings provide a better understanding of nutrient limitation and management for alpine grasslands on the QTP.
Nutrient limitations, particularly those of nitrogen (N) and phosphorus (P) limitations, have profound influences on plant community biomass and composition (Elser et al., 2007). Typically, plant growth, development, reproduction, and changes in plant productivity and diversity are regulated by N and/or P supplies (Elser et al., 2007; Olde Venterink, 2011; Zhang et al., 2019). Some classical methods, such as nutrients addition experiments, plant stoichiometry and foliar nutrient resorption, have demonstrated that N and P limitations are extensive across terrestrial ecosystems (Elser et al., 2007; LeBauer and Treseder, 2008; Yan et al., 2017; Du et al., 2020). A recent global-scale study predicted that 18% of terrestrial natural lands are N limitation, P limitation accounts for 43% and the rest of 39% are N and P co-limitation area or slightly limited by single nutrient (Du et al., 2020), indicating that P is a more predominant limiting nutrient factor than N globally. The substrate soil-age hypothesis also predicts that nutrient limitation can be changing across the globe (Chadwick et al., 1999). Worldwide mapping of N and P limitation patterns shows that terrestrial ecosystems of boreal and temperate coniferous forests, mountainous grasslands and shrublands, and tundra are prevalently N-limited, while P limitation predominates in (sub-) tropical and temperate areas (Du et al., 2020). Therefore, nutrient limitation is spatially heterogeneous across various ecosystem types.
Spatial heterogeneity and climate factors can influence the magnitude of nutrient limitations. Not surprisingly, N and P limitations demonstrate obviously latitudinal and elevational patterns. Generally, N is more restrictive in high latitude and high altitude zones (i.e., low-temperature areas) (Du et al., 2020), whereas P limitation gradually increases from boreal and Arctic regions to tropical forests in the Northern Hemisphere (Chadwick et al., 1999). This is due to increasing precipitation and temperature could strongly diminish N limitation than P limitation (Du et al., 2020; Hou et al., 2020). Interestingly, there is mounting evidence that plant biomass and diversity are limited by multiple nutrients (i.e., co-limitation) in many terrestrial ecosystems (Olde Venterink, 2011; Fay et al., 2015). Therefore, the concept of nutrient limitation has extended from an earlier perception of single-nutrient limitation to the hypothesis of multiple nutrients co-limitation. The responses of plant biomass and diversity to simultaneously N and P fertilization could be synergistic, additive, and non-additive (Harpole et al., 2011; Olde Venterink, 2011; Fay et al., 2015; Wang et al., 2022a). However, previous studies on N and P limitation patterns are usually at the global scale, and specific knowledge on a local/regional scale (e.g., in alpine grasslands on the Qinghai-Tibetan Plateau) is often not clear.
The Qinghai-Tibetan Plateau (QTP), with a vast elevation gradient ranging from 2000 m to 5000 m, is undergoing huge environmental changes (e.g., global climate change and N deposition) (Piao et al., 2019). The alpine grassland, as the most widely distributed and key terrestrial ecosystems in the QTP (Zhang et al., 2014; Wang et al., 2022b), is an ideal experimental site to examine N and P limitation patterns due to low soil N and P contents resulting from low decomposition and mineralization rate (Ren et al., 2016). In the QTP throughout the past 5 decades, numerous N and P fertilization experiments were conducted. Differently, the alpine grasslands on the QTP appeared N limitation (Ren et al., 2016), P limitation (Yang et al., 2014), or N and P co-limitation (Ren et al., 2010). However, the general patterns of N and P limitation in alpine grasslands across the QTP scales remains unclear.
A clear understanding of general N and P limitation pattern in the QTP grasslands can guide the maintaining and restoration management of plant biomass and diversity of alpine grasslands. Here, we operated a meta-analysis to investigate whether plant biomass and diversity of alpine grasslands on the QTP are limited by N and P. Specifically, we mainly to answer: 1) What are N and P limitation patterns of alpine grasslands on the QTP, and which limitation pattern is more prevalent? 2) How does plant diversity respond to N and P addition across QTP? 3) How do mean annual precipitation (MAP, mm) and mean annual temperature (MAT, °C) influence N and P limitation patterns? We hypothesized that N limitation alone is more prevalent than N and P co-limitation in alpine grasslands because of high altitude zones (Du et al., 2020), and MAT is a key affecting factor for nutrient limitation due to low temperature in this region.
We gathered data from articles through searching in Web of Science (https://apps.webofknowledge.com) and the China National Knowledge Infrastructure (https://www.cnki.net) with the terms [(Qinghai-Tibet Plateau OR Qinghai-Tibet* Plateau) AND (nitrogen addition* OR N addition* OR nitrogen fertili* OR N fertili* OR nitrogen deposit* OR N deposit* OR nitrogen enrich* OR N enrich* OR nitrogen application OR N application OR nitrogen deposit* OR N deposit* OR phosph* addition* OR P addition* OR phosph* fertili* OR P fertili* OR phosph* deposit* OR P deposit* OR phosph* enrich* OR P enrich* OR phosph application OR P application OR fertilization) AND (biodivers* OR divers* OR species rich* OR biomass OR product*) AND (plant* OR grass* OR herb* OR forb* OR sedge* OR legume*)] through to March 1, 2022. We preliminary screened the titles and abstracts of all the publications that were retrieved from the two search platforms. Afterward, we evaluated each article and collected data with the following criteria: 1) Studies was based on fertilization experiments in natural alpine grasslands on the QTP; 2) Studies focused on the impacts of nutrient addition (N and/or P and/or N and P co-addition vs. Control) on alpine grasslands; 3) Articles reported at least one of the following variables: vascular plant aboveground biomass (AGB) and belowground biomass (BGB) of total and/or functional groups (grass, sedge, non-leguminous forb, legume) and biodiversity (plant richness, Shannon-Wiener index, Pielou’s evenness index); 4) Studies reported the selected variables of values, including means, standard deviations (SD), or standard error (SE), sample sizes. A total of 107 articles were retained, containing multiple pairwise experiments (i.e., nutrients addition vs. control). Details of the articles are listed in Supplementary Material.
We directly gathered data from the tables in the original main text and online data of each article, and used the WebPlotDigitizer (https://automeris.io/WebPlotDigitizer/) to extract the data presented in figures. For each study, we extracted the values of relative variables (mentioned in criteria 3) from the control and nutrient addition treatments. We also recorded citations, year, latitude, longitude, fertilizer type, fertilization rate, and experimental period of each study (Figure 1). MAP and MAT were acquired from the original publications, or, if not presented, from the WorldClim database (https://www.worldclim.org/data/index.html). For experiments excluding mammals physically, we retained those that prohibited grazing only in growing season.
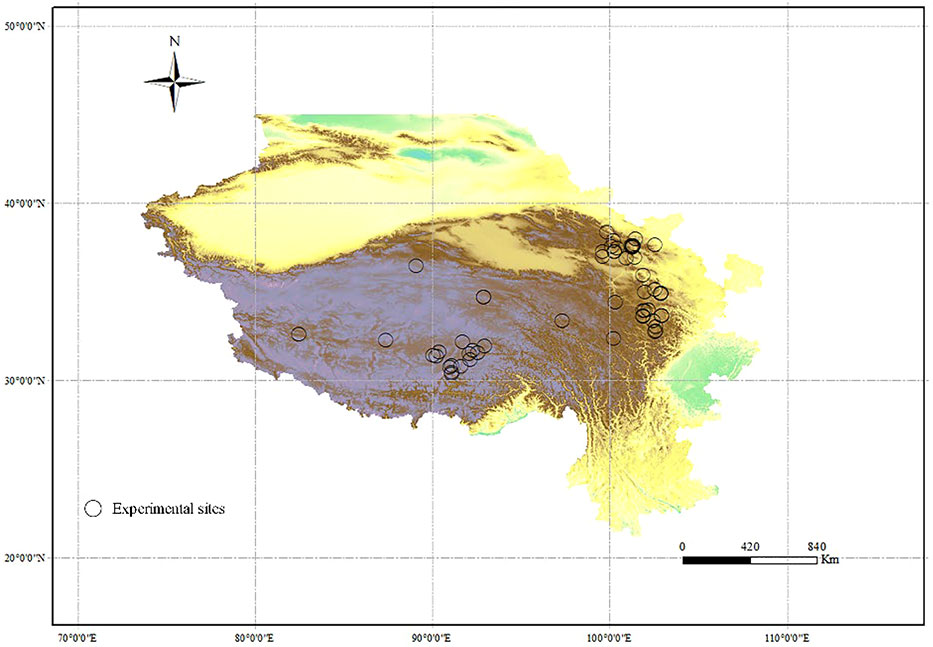
Figure 1 Experimental sites of each article in the meta-analysis in alpine grasslands on the QTP.
We calculated the log response ratio(lnRR) for each data pair (Hedges et al., 1999):
where and respectively represent the arithmetic mean values of each variable in nutrient addition and control manipulations.
We calculated the variance (v) of each lnRR as:
where Se and Sc represent the standard deviations of means, and Ne and Nc are the sample sizes in nutrient addition and control manipulations, respectively.
We used the escalc() and rma.mv() function from the R package “metafor” (Viechtbauer, 2010) to calculate the weighted effect sizes and 95% confidence intervals (CI), with each study nested in the paper as random effect to account for non-independence of data (i.e., multiple studies in a single paper). We considered the effects size (LnRR) to be significant once the 95% CI of mean value of each variable did not overlap with 0. We tested the overall effects of nutrient addition on plant biomass and diversity. Then, we used meta-regression (Borenstein et al., 2009) from the “metafor” package to test whether the influence of nutrient addition on plant biomass and diversity in alpine grasslands on the QTP depended on MAT and MAP. We estimated the moderator through the QM statistic and corresponding P-value to explain the amount of heterogeneity. Funnel plot asymmetry from Kendall’s rank test was used to assess to assess possible publication bias. Finally, we used linear regression analyses and fitted polynomial curves to explore the threshold of the effects of nutrient addition. All statistical analyses were performed in R platform [version 4.0.3] (R Core Team, 2020).
Our findings showed that the positive effects of N addition on plant aboveground biomass were larger than did P addition, with combined N and P addition showing the strongest positive impacts on plant AGB (Figures 2, 3A). Additionally, BGB significantly increased through single N or combined N and P additions, whereas P addition did not show significant effects on BGB (Figure 3B). These results demonstrate a prevalent N and P limitation pattern in alpine grassland on the QTP.
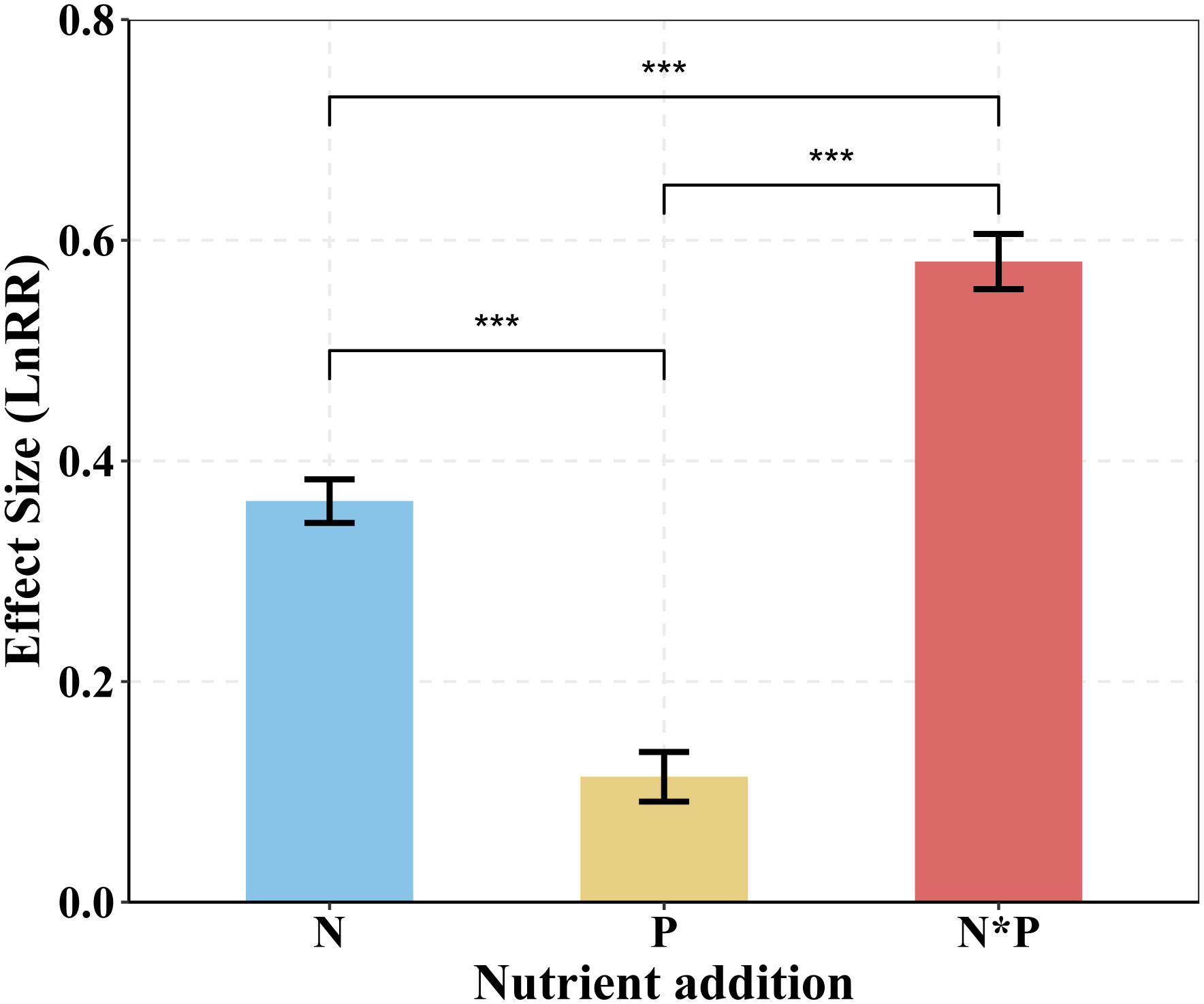
Figure 2 Responses of AGB to single N or P addition or to N*P addition in alpine grassland on the QTP. Mean ± SE are shown. AGB, aboveground biomass. N*P, N and P co-addition. ANOVA was used to analyze the response ratio (LnRR) of plant AGB for each nutrient treatment. The least significant difference (LSD) test was used to separate means at P= 0.05. The statistically significant effect denoted by *p < 0.05, **p < 0.01, ***p < 0.001.
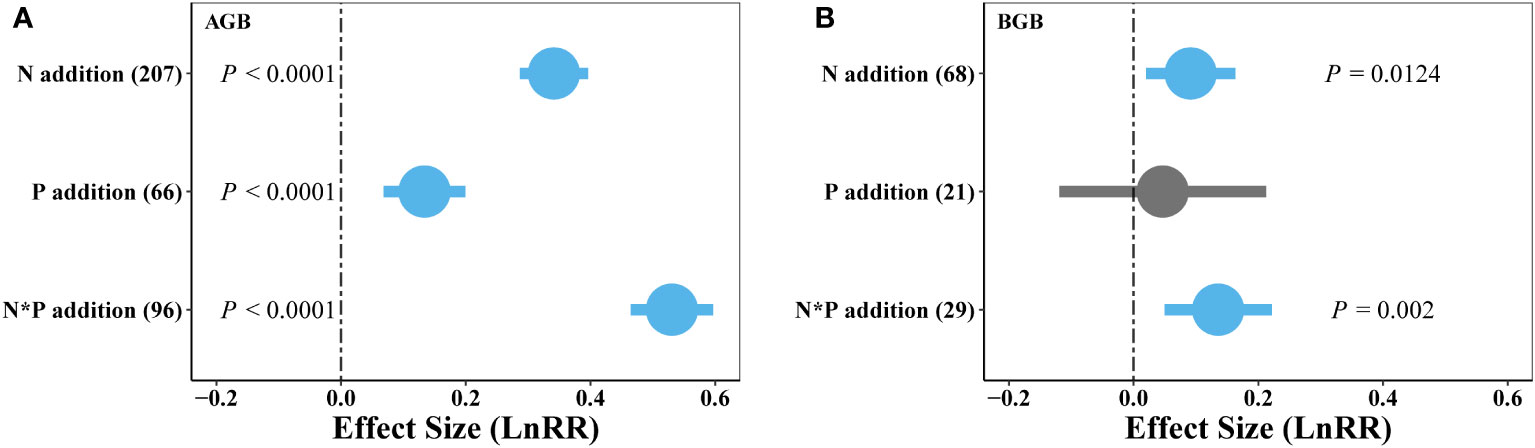
Figure 3 Effects of single N or P, and combined N and P (N*P) addition on weighted response ratio (LnRR) of AGB (A) and BGB (B) in alpine grassland on the QTP. The numbers in brackets indicate the number of studies for each treatment. Error bars represent 95% confidence intervals. The vertical dashed line represents the weighted effect size = 0. The effect of nutrient addition was considered statistically significant if the 95% CI did not overlap zero. Positive (blue) and neutral (grey) effects are shown.
N addition strongly increased AGB of grasses and sedges, but reduced legume AGB (Figure 4A). P addition promoted AGB of grasses but had weak effect on other functional groups (Figure 4B). Moreover, combined N and P addition significantly increased grass AGB, but decreased legume AGB (Figure 4C). The combined N and P addition had a similar effect size with single N addition.
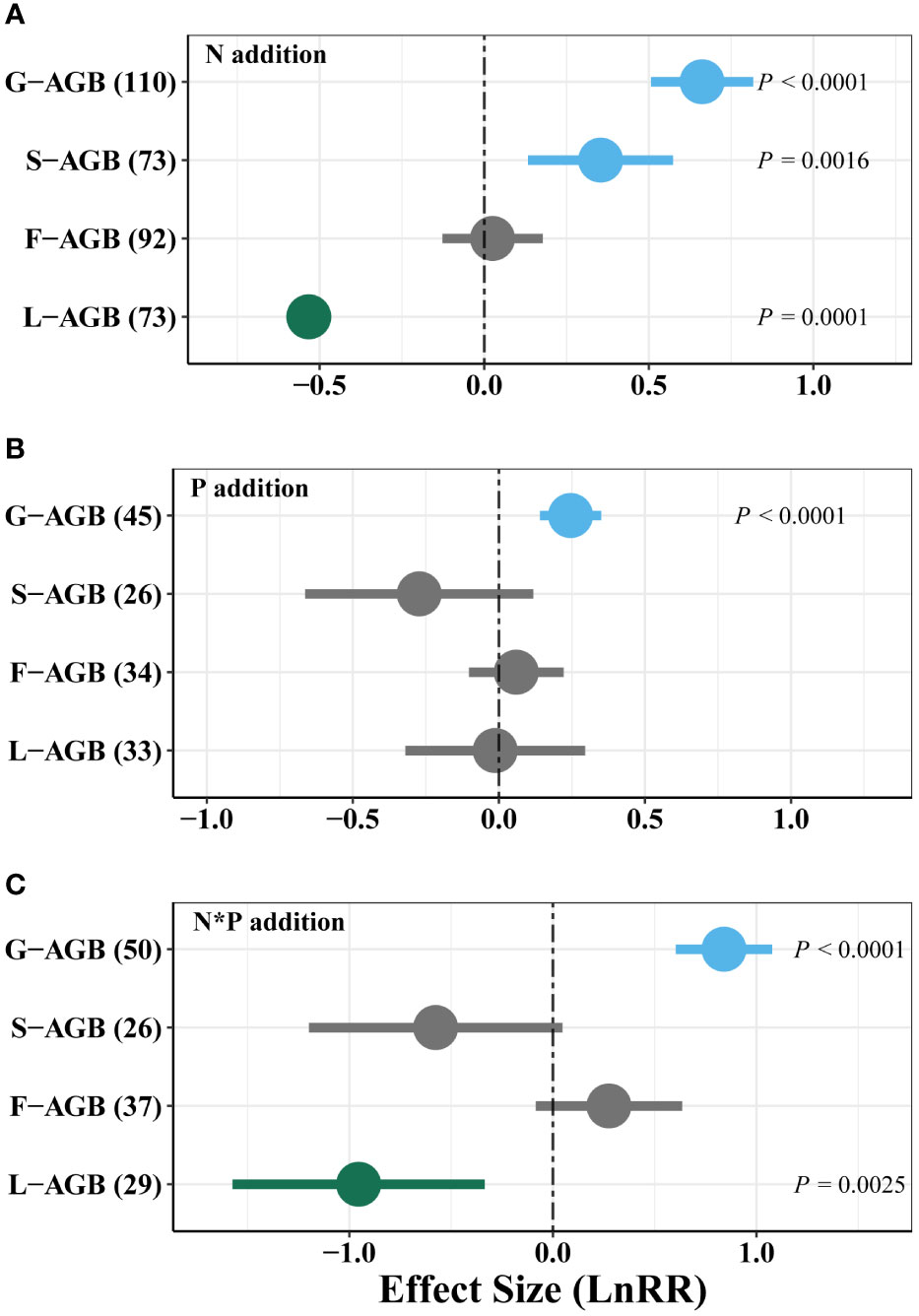
Figure 4 Effects of single N (A), P (B), and combined N and P (N*P, C) addition on weighted response ratio (LnRR) of functional groups’ AGB in alpine grassland on the QTP. G-AGB, grass aboveground biomass; S-AGB, sedge aboveground biomass; F-AGB, non-leguminous forb aboveground biomass; L-AGB, legume aboveground biomass. Positive (blue), negative (green) and neutral (grey) effects are shown.
The effect size of AGB to N addition raised firstly and then declined with increasing N fertilization rate, which peaked at approximately 25 g N·m-2·year-1 (Figure 5A). This pattern was mainly attributed to the response of grasses and sedges, which showed hump-shaped relationships between the effect size of AGB and N fertilization rate (Figure S1). Moreover, the effect size of AGB gradually decreased with increasing experimental period (Figure 5B).
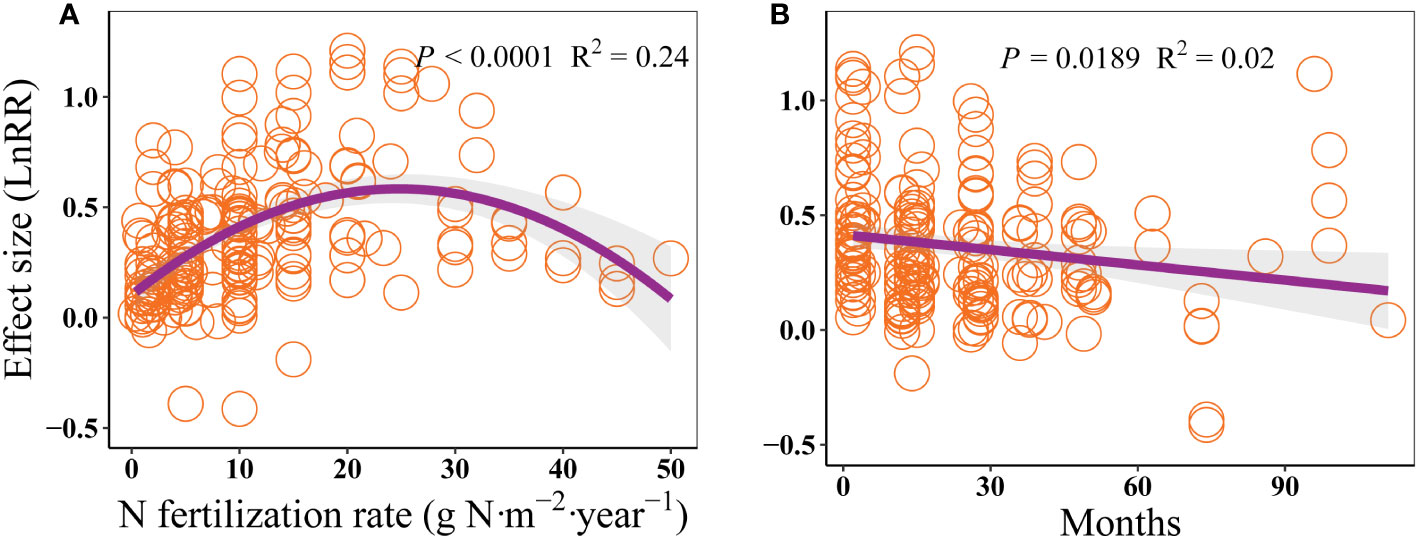
Figure 5 Effects of N fertilization rate (g N·m-2·year-1, A), and experimental period (month, B) on the response of AGB in alpine grassland on the QTP.
Nutrient addition showed negative effects on species richness, with N and P co-addition showing the strongest negative effects (Figure 6A). N and P co-addition also reduced Shannon diversity and Pielou’s evenness of plant communities (Figures 6B, C).
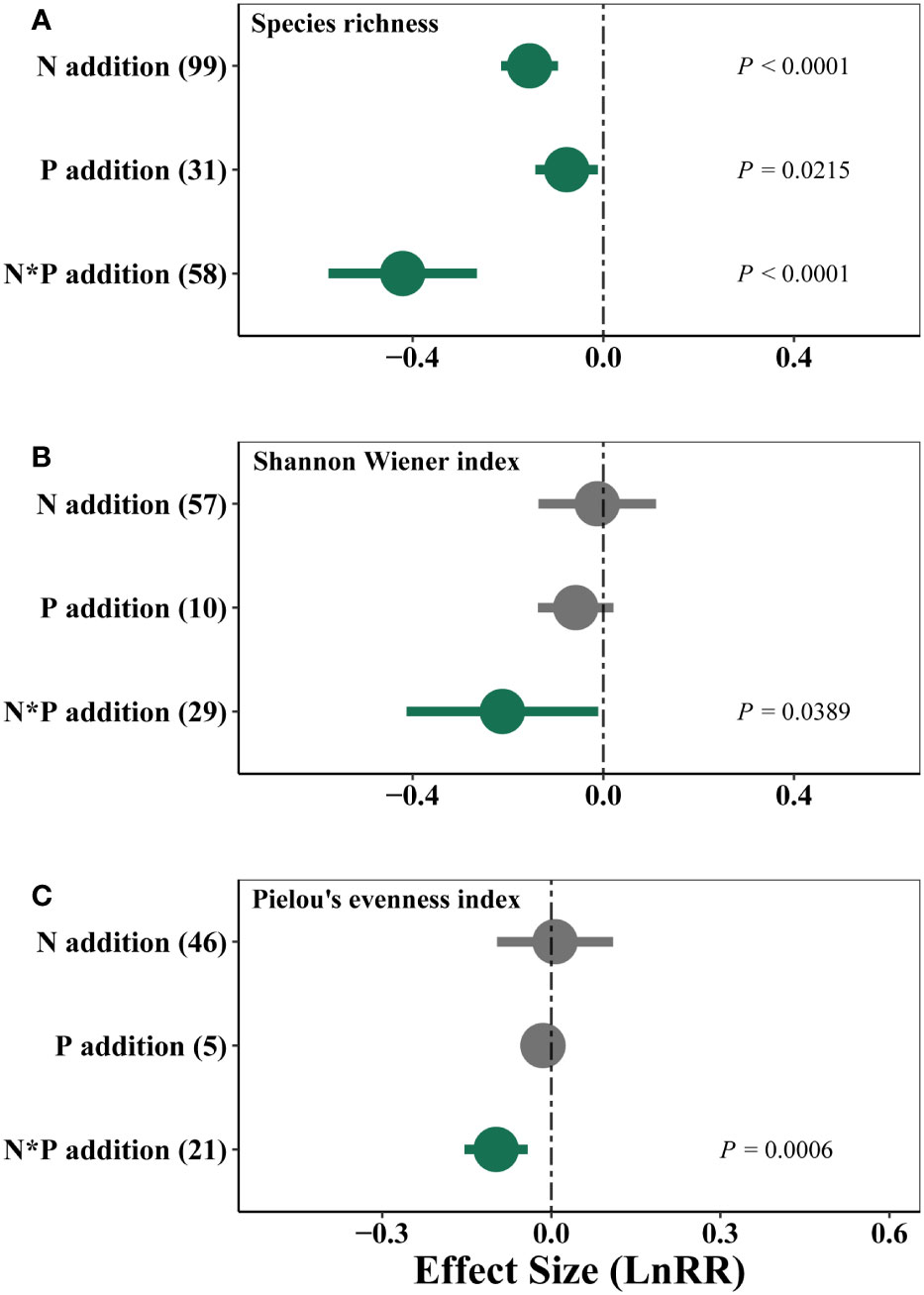
Figure 6 Effects of single N or P, and combined N and P (N*P) additions on weighted response ratio (LnRR) of species richness (A), Shannon Wiener index (B) and Pielou’s evenness index (C) in alpine grassland on the QTP.
The influence of N addition on AGB was positively related to MAP (Figure 7A), whereas the effect size of N addition on BGB showed the opposite (Figure 7B). The response to P addition did not show significant relationship with precipitation (Figures S2A, B). However, increasing MAT slightly decreased the positive effect of P and N addition on aboveground biomass (P = 0.068 in Figure S2C, P = 0.0676 in Figure S3A).
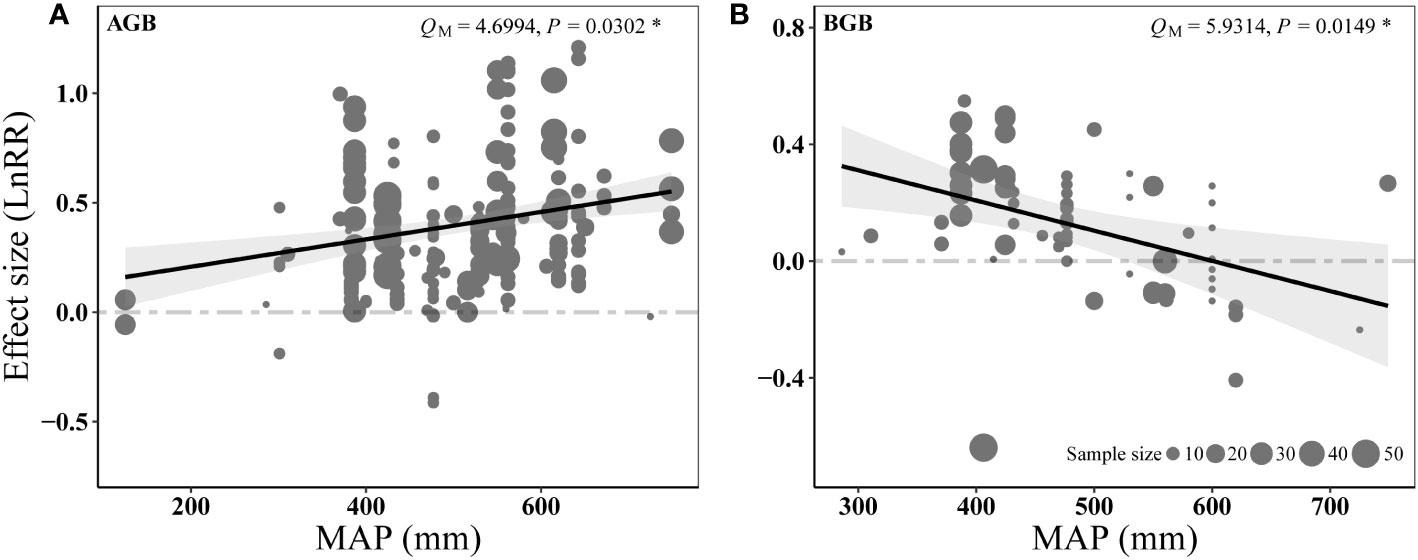
Figure 7 Results of meta-regressions relating the N addition effects on AGB (A) and BGB (B) (effect size, LnRR) to the MAP. The paralleled gray dashed lines represent an effect size (LnRR) of zero.
We demonstrated that N and P addition/co-addition considerably increased plant aboveground biomass, whereas reduced plant diversity in alpine grasslands on the QTP. AGB of plant responses to N addition are influenced by N fertilization rate and experimental duration. Additionally, we found that MAP, not MAT could change the magnitude of N limitation on plant growth, rather than P limitation or N and P co-limitation. This indicates that alpine grassland on the QTP not only is limited by N or P nutrient alone, more importantly, but also is strongly independent N and P co-limitation (Olde Venterink, 2016). This also supports that remarkable interactive effects of combined N and P enrichment are widespread in terrestrial ecosystems, which can be mediated by climatic conditions, such as MAP (Elser et al., 2007; Fay et al., 2015).
Single N addition promoted plant AGB and BGB, but decreased species richness, indicating N limitation on alpine grasslands of the QTP. It is well known that N, as an crucial component of chlorophyll and a variety of photosynthetic enzymes, can facilitate plant growth and reproduction (Kattge et al., 2009). Additionally, N supplies can encourage the growth of roots to meet other nutrient requirements (Yang et al., 2009; Peng et al., 2017). N limitation on plant AGB varied with plant functional groups. We found that N fertilization favored to increasing AGB of grasses and sedges, and decreased to the accumulation of legumes’ AGB, but did not change AGB of non-leguminous forbs, which suggests that N limited the growth of grass and sedge. Thus, in alpine grasslands on the QTP, the positive responses of total plant biomass to N addition mainly contribute to the increase in grass and sedge biomass. Contrarily, the response of higher aboveground biomass is accompanied by a decline of species richness through intense resource competition (e.g., nutrients and/or light competition) (Aerts et al., 2003; Olde Venterink, 2011). Due to light competition, presumably self-shading induced by rapid growth of grasses (e.g., Elymus nutans) as a result of N addition has a greater negative impact on short stature plants, leading to the loss of species diversity. N addition also might generate the toxic effects, when excessive nutrient addition to natural vegetation exceed the homeostasis regulation capability of plants (Güsewell, 2004). Thus, the productivity of plants declines as N fertilization rate exceeded threshold value. Our findings also demonstrated that the threshold of N fertilization rate is 25 g N·m-2·year-1, supporting the above-mentioned toxic effects’ prediction.
The impacts of N addition on AGB and species richness at QTP grasslands diminished over experimental duration. A robust explanation is long-term and continuous N addition increased P limitation. This is because N addition significantly reduced P concentration and enhanced plant N:P ratio, but plant rapid growth required more P, thereby mitigating the impacts of single N limitation (Li et al., 2016; Li et al., 2022).
N fixation and mineralization are biological processes that are hugely affected by climatic conditions (Houlton et al., 2008). In QTP, the N limitation of plant biomass was associated with MAP, while MAP had a positive relationship for AGB and negative relationship for BGB. The phenomenon reveals a shift of plant biomass allocation from belowground to aboveground with improved soil N and water condition. This might be due to the fact of that cold climate in alpine grasslands inhibits water evaporation, and the native plant species have evolved mechanisms to adapt to moisture condition (He et al., 2006). Therefore, more N and water allocation will enter the above-ground parts of the plant community. In addition, Eskelinen and Harrison (2015) have proposed the water and N co-limitation hypothesis, where emphasizes the increase of N-fixers caused by water addition. Accordingly, as a result of increased precipitation, enhanced N-fixers may promote decomposable litters, and change soil microorganism communities and nutrient cycling, further, alter the demands for nutrients by plants (Suttle et al., 2007; Eskelinen and Harrison, 2015; Zong et al., 2021). A recent study across the Tibetan permafrost region also suggests that the increased plant N requirement and loss of gaseous N caused by climate warming and wetting could strengthen the effects of N limitation on plant growth (Kou et al., 2020). Contrarily, LeBauer and Treseder (2008) have shown that N limitation is independent of temperature and precipitation in grassland. We did not find strong correlations between the effects of fertilization on plant biomass and MAT due to the prolonged cold temperature and narrow amplitudes in alpine grasslands on the QTP.
In general, N limitation is more severe than P limitation on the QTP since P weathering (a chemical process) is far less sensitive to temperature than N mineralization (a biological process) (Heerwaarden et al., 2003; Ren et al., 2010). Therefore, the low temperature in alpine grasslands on the QTP limits soil mineral nutrients availability, predominantly N availability (KÖener, 2003).
Although P effect was less than N effect in our meta-analysis, P limitation is also remarkable in alpine grasslands on the QTP. A main reason is that plant growth and reproduction can be limited once the supply of P for photosynthesis machinery (such as rRNA or proteins) is insufficient (Sterner and Elser, 2002). Additionally, we found that the response to P addition was not significantly correlated with MAP and MAT, suggesting that the response to P addition is not sensitive to precipitation and temperature, compared to the response of N in QTP.
Our findings also demonstrated that single P addition promoted AGB rather than BGB, but significantly reduced species richness, presumably because plant competition for nutrients shift into light competition (Borer et al., 2014). Evidently, Li et al. (2022) suggested that P addition significantly raised plant height by 18.8% in grasslands, while Ren et al. (2010) found a reduction in light penetration into the plant canopy. Moreover, soil acidification caused by excessive P addition may also be a reason for the reduction of plant species richness (Borer et al., 2014).
Our synthesis revealed that increase in plant biomass and decrease in plant diversity induced by N and P co-addition were greater than that of single N or P addition, suggesting that N and P co-limitation is more prevalent in alpine grasslands across the QTP. This is consistent with the results of Wang et al. (2020) in degraded grasslands of the QTP, they found that N and P co-addition promoted plant growth, but decreased plant diversity. The decomposition of soil organic matter and the release of nutrients in alpine grasslands on the QTP may be limited by low basic N and P contents as a result of lower temperatures and less rainfall. Harpole et al. (2011) proposed that independent co-limitation is more prevalent in areas with low-level total N and P. Apparently, as limited nutrients, N and P co-addition increase soil N and P contents, thereby enhance the N and P availability. Meanwhile, N and P addition could promote the synthesis of enzymes and mycorrhizal activity, which is advantageous for plants to absorb nutrients (Rowe et al., 2008). Multiple nutrients co-addition alters resource competition and community composition, thus co-limitation directly emerges at the community level (Danger et al., 2008).
Generally, N and P co-addition significantly decreased plant diversity, reflecting the decline of niche dimension due to surging nutrient limitation (Interlandi and Kilham, 2001; Ren et al., 2010). The homogenization of resources caused by nutrient supplies, which diminishes niche differentiation and increases the number of dominant species. In accordance with Tilman’s theory (Tilman, 1982), the dynamics of plant community are driven by resource competition (for limited resources). The general increase in plant biomass caused by nutrients co-addition intensifies resource competition in turn decreases species diversity as a result of competitive exclusion (Grime, 1973; Wang et al., 2022a). We found that N limitation has more negative effects on species richness than P limitation. According to the mechanism of resource pre-emption competition, the superior competitor will receive a resource from rhizosphere faster than its neighboring rivals (Craine et al., 2005). Belowground competition for the acquisition of nutrients might be more critical under N limitation as opposed to P limitation (Olde Venterink, 2011), because N limitation is more obvious in vegetation with a high biomass of roots, strong photosynthetic activity, rapid growth rate and high nitrogen productivity (Gusewell, 2005; Lambers et al., 2008; Lambers et al., 2011; Olde Venterink, 2011). Therefore, magnitude of increase in plant belowground biomass caused by different nutrients addition can reduce plant diversity in alpine grasslands on the QTP.
Studies on alpine grasslands also found that soil available N pools dramatically rose by 24% between the 1980s and 2010s, while available P and K pools declined by 3% and 23%, respectively (Tian et al., 2019), suggesting that Tibetan alpine grasslands could shift from N limitation to P or K-limited in the future (Tian et al., 2019). This was also supported by Zong et al. (2021), who revealed that the pattern of nutrient limitation in alpine grasslands on the QTP shifted from N limited to N and P co-limited, but depending on precipitation. A possible reason is that increasing soil N availability with the increase of N deposition, climate warming and wetting on the QTP may further exacerbate P deficiency (Aerts et al., 1992) and potentially increase other nutrients limitation, such as P, K, or micronutrients (Mahowald et al., 2008). Alternatively, because the uptake of one nutrient is closely correlated with that of other nutrients (Fay et al., 2015), single nutrient addition will rapidly induce limitation by the alternative nutrient (Elser et al., 2007), as a reason of P limitation induced by N addition could be compensated by N and P co-addition. Many studies have also demonstrated the positive effects of N and P co-addition on ANPP are greater than N or P addition alone on the QTP (Ren et al., 2010; Ren et al., 2016; Wang et al., 2022b). Therefore, N and P independent co-limitation is more widespread in alpine grassland on the QTP.
Our meta-analysis shows that N and P independent co-limitation is a more prevalent pattern than single N or P limitation in alpine grasslands on the Qinghai-Tibetan Plateau. This did not support our hypothesis that N limitation alone is more prevalent. Meanwhile, nutrients addition and MAP have stronger interactive effects on alpine grasslands than interactive effects of nutrients addition and MAT, suggesting that MAP has more important role in mediating nutrient limitation than MAT in this region. N and P co-addition results in a greater reduction of species richness than that of N or P addition alone, reconfirming the resource competition hypothesis and the compensatory effects between limited resources, and it also suggests that excessive fertilization and increased N and P deposition will result in rapid species loss. In summary, our findings will provide a scientific basis for helping to understand nutrients limitation patterns, which will be conductive to fertilization management in degraded alpine grasslands on the QTP under the global climate change.
The original contributions presented in the study are included in the article/Supplementary Material. Further inquiries can be directed to the corresponding author.
All authors conceived the project. KD, YT processed data and conducted the meta-analysis. WL, SM and MJ helped with data analyses and manuscript revision. All authors contributed to the article and approved the submitted version.
This research was supported by funding from National Natural Science Foundation of China (32271761) to WL and National Training Program for College Students’ Innovation and Entrepreneurship (202210730128) to ML.
We also thank Dr. Shan Luo from German Centre for Integrative Biodiversity Research (iDiv) Halle-Jena-Leipzig for providing constructive comments and improving language on an earlier version of the manuscript.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fpls.2023.1140462/full#supplementary-material
QTP, Qinghai-Tibetan Plateau; AGB, Aboveground biomass; ANPP, Aboveground net primary productivity; BGB, Belowground biomass; MAT, Mean annual temperature; MAP, Mean annual precipitation; G-AGB, Grass aboveground biomass; S-AGB, Sedge aboveground biomass; F-AGB, Non-leguminous forb aboveground biomass; L-AGB, Legume aboveground biomass.
Aerts, R., de Caluwe, H., Beltman, B. (2003). Is the relation between nutrient supply and biodiversity co-determined by the type of nutrient limitation? Oikos 101, 489–498. doi: 10.1034/j.1600-0706.2003.12223.x
Aerts, R., Wallen, B., Malmer, N. (1992). Growth-limiting nutrients in sphagnum-dominated bogs subject to low and high atmospheric nitrogen supply. J. Ecol. 80, 131–140. doi: 10.2307/2261070
Borenstein, M., Hedges, L., Higgins, J., Rothstein, H. (2009). Introduction to meta-analysis. (Wiley.).
Borer, E. T., Seabloom, E. W., Gruner, D. S., Harpole, W. S., Hillebrand, H., Lind, E. M., et al. (2014). Herbivores and nutrients control grassland plant diversity via light limitation. Nature 508, 517–520. doi: 10.1038/nature13144
Chadwick, O. A., Derry, L. A., Vitousek, P. M., Huebert, B. J., Hedin, L. O. (1999). Changing sources of nutrients during four million years of ecosystem development. Nature 397, 491–497. doi: 10.1038/17276
Craine, J. M., Fargione, J., Sugita, S. (2005). Supply pre-emption, not concentration reduction, is the mechanism of competition for nutrients. New Phytol. 166, 933–940. doi: 10.1111/j.1469-8137.2005.01386.x
Danger, M., Daufresne, T., Lucas, F., Pissard, S., Lacroix, G. (2008). Does liebig's law of the minimum scale up from species to communities? Oikos 117, 1741–1751. doi: 10.1111/j.1600-0706.2008.16793.x
Du, E., Terrer, C., Pellegrini, A. F. A., Ahlström, A., van Lissa, C. J., Zhao, X., et al. (2020). Global patterns of terrestrial nitrogen and phosphorus limitation. Nat. Geosci. 13, 221–226. doi: 10.1038/s41561-019-0530-4
Elser, J. J., Bracken, M. E. S., Cleland, E. E., Gruner, D. S., Harpole, W. S., Hillebrand, H., et al. (2007). Global analysis of nitrogen and phosphorus limitation of primary producers in freshwater, marine and terrestrial ecosystems. Ecol. Lett. 10, 1135–1142. doi: 10.1111/j.1461-0248.2007.01113.x
Eskelinen, A., Harrison, S. P. (2015). Resource colimitation governs plant community responses to altered precipitation. Proc. Natl. Acad. Sci. U.S.A. 112, 13009–13014. doi: 10.1073/pnas.1508170112
Fay, P. A., Prober, S. M., Harpole, W. S., Knops, J. M. H., Bakker, J. D., Borer, E. T., et al. (2015). Grassland productivity limited by multiple nutrients. Nat. Plants 1, 15080. doi: 10.1038/nplants.2015.80
Grime, J. (1973). Competitive exclusion in herbaceous vegetation. Nature 242, 344–347. doi: 10.1038/242344a0
Güsewell, S. (2004). N : P ratios in terrestrial plants: Variation and functional significance. New Phytol. 164, 243–266. doi: 10.1111/j.1469-8137.2004.01192.x
Gusewell, S. (2005). High nitrogen: Phosphorus ratios reduce nutrient retention and second-year growth of wetland sedges. New Phytol. 166, 537–550. doi: 10.1111/j.1469-8137.2005.01320.x
Harpole, W. S., Ngai, J. T., Cleland, E. E., Seabloom, E. W., Borer, E. T., Bracken, M. E., et al. (2011). Nutrient co-limitation of primary producer communities. Ecol. Lett. 14, 852–862. doi: 10.1111/j.1461-0248.2011.01651.x
He, J.-S., Fang, J., Wang, Z., Guo, D., Flynn, D. F. B., Geng, Z. (2006). Stoichiometry and large-scale patterns of leaf carbon and nitrogen in the grassland biomes of China. Oecologia 149, 115–122. doi: 10.1007/s00442-006-0425-0
Hedges, L. V., Gurevitch, J., Curtis, P. S. (1999). The meta-analysis of response ratios in experimental ecology. Ecology 80, 1150–1156. doi: 10.2307/177062
Heerwaarden, L., Toet, S., Aerts, R. (2003). Nitrogen and phosphorus resorption efficiency and proficiency in six sub-arctic bog species after 4 years of nitrogen fertilization. J. Ecol. 91, 1060–1070. doi: 10.2307/3599690
Hou, E., Luo, Y., Kuang, Y., Chen, C., Lu, X., Jiang, L., et al. (2020). Global meta-analysis shows pervasive phosphorus limitation of aboveground plant production in natural terrestrial ecosystems. Nat. Commun. 11, 637. doi: 10.1038/s41467-020-14492-w
Houlton, B. Z., Wang, Y.-P., Vitousek, P. M., Field, C. B. (2008). A unifying framework for dinitrogen fixation in the terrestrial biosphere. Nature 454, 327–330. doi: 10.1038/nature07028
Interlandi, S. J., Kilham, S. S. (2001). Limiting resources and regulation of diversity in phytoplankton communities. Ecology 82, 1270–1282. doi: 10.1890/0012-9658(2001)082[1270:lratro]2.0.co;2
Kattge, J., Knorr, W., Raddatz, T., Wirth, C. (2009). Quantifying photosynthetic capacity and its relationship to leaf nitrogen content for global-scale terrestrial biosphere models. Global Change Biol. 15, 976–991. doi: 10.1111/j.1365-2486.2008.01744.x
KÖener, C. (2003). Alpine plant life: Functional plant ecology of high mountain ecosystems (New York: Springer Berlin Heidelberg).
Kou, D., Yang, G., Li, F., Feng, X., Zhang, D., Mao, C., et al. (2020). Progressive nitrogen limitation across the Tibetan alpine permafrost region. Nat. Commun. 11, 3331. doi: 10.1038/s41467-020-17169-6
Lambers, H., Brundrett, M. C., Raven, J. A., Hopper, S. D. (2011). Plant mineral nutrition in ancient landscapes: high plant species diversity on infertile soils is linked to functional diversity for nutritional strategies. Plant Soil 348, 7–27. doi: 10.1007/s11104-011-0977-6
Lambers, H., Raven, J., Shaver, G., Smith, S. (2008). Plant nutrient-acquisition strategies change with soil age. Trends Ecol. Evol. 23, 95–103. doi: 10.1016/j.tree.2007.10.008
LeBauer, D. S., Treseder, K. K. (2008). Nitrogen limitation of net primary productivity in terrestrial ecosystems is globally distributed. Ecology 89, 371–379. doi: 10.1890/06-2057.1
Li, W., Gan, X., Jiang, Y., Cao, F., Lü, X.-T., Ceulemans, T., et al. (2022). Nitrogen effects on grassland biomass production and biodiversity are stronger than those of phosphorus. Environ. Pollut. 309, 119720. doi: 10.1016/j.envpol.2022.119720
Li, Y., Niu, S., Yu, G. (2016). Aggravated phosphorus limitation on biomass production under increasing nitrogen loading: A meta-analysis. Global Change Biol. 22, 934–943. doi: 10.1111/gcb.13125
Mahowald, N., Jickells, T. D., Baker, A. R., Artaxo, P., Benitez-Nelson, C. R., Bergametti, G., et al. (2008). Global distribution of atmospheric phosphorus sources, concentrations and deposition rates, and anthropogenic impacts. Global Biogeochemical Cycles 22, n/a–n/a. doi: 10.1029/2008gb003240
Olde Venterink, H. (2011). Does phosphorus limitation promote species-rich plant communities? Plant Soil 345, 1–9. doi: 10.1007/s11104-011-0796-9
Olde Venterink, H. (2016). Productivity increase upon supply of multiple nutrients in fertilization experiments; co-limitation or chemical facilitation? Plant Soil 408, 515–518. doi: 10.1007/s11104-016-3055-2
Peng, Y., Li, F., Zhou, G., Fang, K., Zhang, D., Li, C., et al. (2017). Nonlinear response of soil respiration to increasing nitrogen additions in a Tibetan alpine steppe. Environ. Res. Lett. 12, 024018. doi: 10.1088/1748-9326/aa5ba6
Piao, S. L., Zhang, X. Z., Wang, T., Liang, E. Y., Wang, S. P., Zhu, J. T., et al. (2019). Responses and feedback of the Tibetan Plateau's alpine ecosystem to climate change. Chin. Sci. Bull. 64, 2842–2855. doi: 10.1360/TB-2019-0074
R Core Team (2020). R: A language and environment for statistical computing. (Vienna, Austria: R Foundation for Statistical Computing). Available at: https://www.R-project.org/.
Ren, F., Song, W., Chen, L., Mi, Z., Zhang, Z., Zhu, W., et al. (2016). Phosphorus does not alleviate the negative effect of nitrogen enrichment on legume performance in an alpine grassland. J. Plant Ecol. 10, 822–830. doi: 10.1093/jpe/rtw089
Ren, Z., Li, Q., Chu, C., Zhao, L., Zhang, J., Dexiecuo, A., et al. (2010). Effects of resource additions on species richness and ANPP in an alpine meadow community. J. Plant Ecol. 3, 25–31. doi: 10.1093/jpe/rtp034
Rowe, E. C., Smart, S. M., Kennedy, V. H., Emmett, B. A., Evans, C. D. (2008). Nitrogen deposition increases the acquisition of phosphorus and potassium by heather calluna vulgaris. Environ. Pollut. 155, 201–207. doi: 10.1016/j.envpol.2007.12.008
Sterner, R. W., Elser, J. J. (2002). Ecological stoichiometry: The biology of elements from molecules to the biosphere (Princeton, NJ: Princeton University Press).
Suttle, K. B., Thomsen, M. A., Power, M. E. (2007). Species interactions reverse grassland responses to changing climate. Science 315, 640–642. doi: 10.1126/science.1136401
Tian, L., Zhao, L., Wu, X., Hu, G., Fang, H., Zhao, Y., et al. (2019). Variations in soil nutrient availability across Tibetan grassland from the 1980s to 2010s. Geoderma 338, 197–205. doi: 10.1016/j.geoderma.2018.12.009
Tilman, D. (1982). Resource competition and and community structure (Princeton, NJ: Princeton University Press).
Viechtbauer, W. (2010). Conducting meta-analyses in r with the metafor package. J. Stat. Software 36, 1–48. doi: 10.18637/jss.v036.i03
Wang, D., Zhou, H., Yao, B., Wang, W., Dong, S., Shang, Z., et al. (2020). Effects of nutrient addition on degraded alpine grasslands of the qinghai-Tibetan plateau: A meta-analysis. Agriculture Ecosyst. Environ. 301, 106970. doi: 10.1016/j.agee.2020.106970
Wang, Y., Du, J., Pang, Z., Liu, Y., Xue, K., Hautier, Y., et al. (2022a). Unimodal productivity–biodiversity relationship along the gradient of multidimensional resources across Chinese grasslands. Natl. Sci. Rev. 9, nwac165. doi: 10.1093/nsr/nwac165
Wang, Y., Lv, W., Xue, K., Wang, S., Zhang, L., Hu, R., et al. (2022b). Grassland changes and adaptive management on the qinghai–Tibetan plateau. Nat. Rev. Earth Environ. 3, 668–683. doi: 10.1038/s43017-022-00330-8
Yan, Z., Tian, D., Han, W., Tang, Z., Fang, J. (2017). An assessment on the uncertainty of the nitrogen to phosphorus ratio as a threshold for nutrient limitation in plants. Ann. Bot. 120, 937–942. doi: 10.1093/aob/mcx106
Yang, Y., Fang, J., Ji, C., Han, W. (2009). Above- and belowground biomass allocation in Tibetan grasslands. J. Vegetation Sci. 20, 177–184. doi: 10.1111/j.1654-1103.2009.05566.x
Yang, Z., Su, F., Miao, Y., Zhong, M., Xiao, R. (2014). Effects of fertilization and grazing on species richness in an alpine meadow of Qinghai-Xizang Plateau. Chin. J. Plant Ecol. 38, 1074–1081. doi: 10.3724/sp.J.1258.2014.00101
Zhang, C., Willis, C. G., Ma, Z., Ma, M., Csontos, P., Baskin, C. C., et al. (2019). Direct and indirect effects of long-term fertilization on the stability of the persistent seed bank. Plant Soil 438, 239–250. doi: 10.1007/s11104-019-04024-x
Zhang, Y., Qi, W., Zhou, C., Ding, M., Liu, L., Gao, J., et al. (2014). Spatial and temporal variability in the net primary production of alpine grassland on the Tibetan Plateau since 1982. J. Geographical Sci. 24, 269–287. doi: 10.1007/s11442-014-1087-1
Keywords: meta-analysis, nutrient limitation, nitrogen, phosphorus, plant biomass and diversity, alpine grassland, Qinghai-Tibetan Plateau
Citation: Dong K, Li W, Tang Y, Ma S and Jiang M (2023) Co-limitation of N and P is more prevalent in the Qinghai-Tibetan Plateau grasslands. Front. Plant Sci. 14:1140462. doi: 10.3389/fpls.2023.1140462
Received: 09 January 2023; Accepted: 30 January 2023;
Published: 15 February 2023.
Edited by:
Huakun Zhou, Northwest Institute of Plateau Biology (CAS), ChinaReviewed by:
Zhenchao Zhang, Qingdao Agricultural University, ChinaCopyright © 2023 Dong, Li, Tang, Ma and Jiang. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Wenjin Li, bGl3akBsenUuZWR1LmNu
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.