 Zhao Liu
Zhao Liu Tao Xiong4
Tao Xiong4 Xiangyang Kang
Xiangyang Kang Jun Yang
Jun Yang
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Plant Sci. , 28 July 2022
Sec. Plant Breeding
Volume 13 - 2022 | https://doi.org/10.3389/fpls.2022.952877
This article is part of the Research Topic Artificial Polyploidy in Plants, Volume II View all 15 articles
Golden 2-Like (GLK) transcription factors play a crucial role in chloroplast development and chlorophyll synthesis in many plant taxa. To date, no systematic analysis of GLK transcription factors in tree species has been conducted. In this study, 40 EgrGLK genes in the Eucalyptus grandis genome were identified and divided into seven groups based on the gene structure and motif composition. The EgrGLK genes were mapped to 11 chromosomes and the distribution of genes on chromosome was uneven. Phylogenetic analysis of GLK proteins between E. grandis and other species provided information for the high evolutionary conservation of GLK genes among different species. Prediction of cis-regulatory elements indicated that the EgrGLK genes were involved in development, light response, and hormone response. Based on the finding that the content of chlorophyll in mature leaves was the highest, and leaf chlorophyll content of triploid Eucalyptus urophylla was higher than that of the diploid control, EgrGLK expression pattern in leaves of triploid and diploid E. urophylla was examined by means of transcriptome analysis. Differential expression of EgrGLK genes in leaves of E. urophylla of different ploidies was consistent with the trend in chlorophyll content. To further explore the relationship between EgrGLK expression and chlorophyll synthesis, co-expression networks were generated, which indicated that EgrGLK genes may have a positive regulatory relationship with chlorophyll synthesis. In addition, three EgrGLK genes that may play an important role in chlorophyll synthesis were identified in the co-expression networks. And the prediction of miRNAs targeting EgrGLK genes showed that miRNAs might play an important role in the regulation of EgrGLK gene expression. This research provides valuable information for further functional characterization of GLK genes in Eucalyptus.
Golden 2-Like (GLK) transcription factors are members of the GARP family (Riechmann et al., 2000; Xiao et al., 2019). Most GLK genes contain two highly conserved domains: a MYB DNA-binding domain and a C-terminal (GCT) box (Rossini et al., 2001). A GLK gene was first identified in maize (Zea mays L.) and, subsequently, numerous GLK genes were detected in Arabidopsis thaliana, rice (Oryza sativa L.), and tomato (Solanum lycopersicum; Fitter et al., 2002; Powell et al., 2012; Bhutia et al., 2020). The GLK transcription factors play crucial roles in chloroplast development and chlorophyll synthesis in many plant taxa (Bravo-Garcia et al., 2009; Chen et al., 2016).
Although the function of GLK genes is conserved, different genetic regulatory mechanisms may operate in different species. There are three types of chloroplasts in C4 plants: C4 bundle sheath cells, C4 and C3 mesophyll cells (Langdale and Nelson, 1991). The spatially tissue-specific expression of different GLK genes in maize might represent a specialization required for the development of distinct bundle sheath and mesophyll chloroplasts (Rossini et al., 2001). However, in Cleome gynandra, different GLK genes are both expressed in bundle sheath and mesophyll cells (Wang et al., 2013). In the C3 plant Arabidopsis, GLK genes play a redundant role in regulating chloroplast development. Single-insertion mutants showed normal phenotypes in most photosynthetic tissues, whereas in double mutants all photosynthetic tissues and chloroplasts were pale green (Fitter et al., 2002). In addition, two typical GLK genes were found in tomato, both of which are expressed in the leaves, but only one is predominantly expressed in fruit (Powell et al., 2012; Nguyen et al., 2014). In many plant species, GLK genes act as transcriptional regulators of chloroplast development. However, to date, no study of GLK genes in forest tree species has been conducted.
Eucalyptus is a genus of fast-growing tree species that are widely planted around the world (Booth et al., 2017; Deng et al., 2020). These trees provide raw materials for pulp and paper manufacturing, and have the advantage of fixing large amounts of atmospheric carbon (Pérez et al., 2006; Hii et al., 2017; Vilasboa et al., 2019). Compared with diploid individuals, polyploid plants usually exhibit superior growth and carbon absorption, which reflects improved photosynthetic efficiency after polyploidization (Liao et al., 2016; Li et al., 2019). Therefore, polyploid Eucalyptus is potentially important to improve plant biomass accumulation and to mitigate global warming. Photosynthesis occurs mainly in chloroplasts (Gan et al., 2019). GLK family genes are associated with chloroplast development and chlorophyll synthesis in many plant taxa (Chen et al., 2016). However, after whole-genome duplication, the gene dosage effect and epigenetic modification may affect gene expression and ultimately lead to trait variation (Allario et al., 2013; Zhang et al., 2015). For example, in triploid poplar, genes associated with chlorophyll synthesis are upregulated as a result of the gene dosage effect, which lead to increase in chlorophyll content (Du et al., 2020). The effect of GLK genes on chloroplast development and chlorophyll synthesis in Eucalyptus of different ploidies remains to be studied.
In this study, 40 GLK family genes were identified by genome-wide analysis of the genome of Eucalyptus grandis. The chromosomal distribution, phylogenetic relationships, conserved motifs, intron and exon structure, and cis-acting regulatory elements of GLK genes were analyzed. In combination with analysis of the chlorophyll content in E. urophylla of different ploidies, the effect of GLK family genes on chlorophyll synthesis was studied based on RNA-sequencing data and co-expression network analysis. In addition, putative miRNAs targeting EgrGLK genes were also been predicted. The results are important to enhance understanding of the GLK gene family and provide a reference for studying the molecular mechanism of the increase in chlorophyll content in polyploid plants.
The analysis done in this study was presented in Supplementary Figure S1 in the form of flow chart. To identify the GLK genes of E. grandis, genomic data were downloaded from the Phytozome database.1 Using published GLK protein sequences from Arabidopsis, maize, and tomato as query sequences (Liu et al., 2016; Alam et al., 2022; Wang et al., 2022), the GLK protein sequences in the E. grandis reference genome were identified with the BLASTP tool (E-value: 1e−5). The identified sequences were submitted to the SMART online tool2 and the NCBI Web CD-Search Tool3 for further confirmation of GLK proteins. The protein sequences that included a GLK domain (PF00249) were retained and designated EgrGLK. The physical parameters and subcellular localization of these proteins were predicted with the ExPASy4 and WoLF PSORT5 online tools.
The location of genes on chromosome and the analysis of gene duplication can provide us with more genetic information about the EgrGLK genes. Information on the chromosomal location of each EgrGLK gene was extracted from the Phytozome database and the identified EgrGLK genes were mapped to individual chromosomes using TBtools (Chen et al., 2020). The duplication landscape of EgrGLK genes and cross-species collinearity of GLK genes was confirmed with MCScanX software (Wang et al., 2012). The parameters non-synonymous mutations (Ka), synonymous mutations (Ks) and estimated evolutionary constraints (Ka/Ks) among the EgrGLK genes were calculated using TBtools (Chen et al., 2020).
To explore the evolutionary relationships of GLK proteins in plants, a phylogenetic tree was constructed derived from the EgrGLK protein sequences and published GLK protein sequences from Arabidopsis, maize, and tomato. A multiple sequence alignment of the sequences was generated with ClustalX (Thompson et al., 1997). A phylogenetic tree with 1,000 bootstrap replicates was generated using the neighbor-joining method with MEGAX software (Kumar et al., 2018). The tree was manipulated with the iTOL online tool.6
Structure, conserved motifs and cis-elements of genes can provide important information for understanding gene function. The structure of the EgrGLK genes was analyzed with the GSDS platform7 for prediction of introns and exons. The conserved motifs of the EgrGLK proteins were predicted using the MEME Suite online tool.8 The identified motifs were annotated using the NCBI Web CD-Search Tool. The nucleotide sequence 2000 bp upstream of the start codon for the EgrGLK genes was extracted from the E. grandis reference genome, and the sequences were submitted to the PlantCARE database9 for prediction of cis-acting regulatory elements. Conserved motifs, gene structure, and cis-element information were visualized using TBtools (Chen et al., 2020).
To explore the effect of polyploidization on chlorophyll content in plant leaves, triploid E. urophylla obtained by sexual polyploidization and its diploid control were used as materials for measurement of chlorophyll content (Yang et al., 2018). Five clones of triploid and diploid E. urophylla were selected. Young leaves at the shoot tips, fully expanded mature leaves, and senescent leaves were randomly selected. The chlorophyll content was determined following the method described by Du et al. (2020). Fresh leaf tissue (1 g), 5 ml of 95% ethanol, and a small amount of quartz sand and calcium carbonate were mixed and ground into a homogenate. An additional 5 ml of 95% ethanol was added and ground further. After standing for 3 min, the homogenate was filtered into a 50 ml brown volumetric flask and diluted to 50 ml with 95% ethanol. Absorbance (A) was measured at 645 and 663 nm using a spectrophotometer (Ultrospec 6300 Pro, Biochrom, Cambridge, United Kingdom). The chlorophyll content (mg/g) was calculated with the formula 8.02 × A663 + 20.20 × A645.
In order to reveal the reasons for the changes of chlorophyll content in plant leaves after polyploidization, terminal buds, young leaves, mature leaves, and senescent leaves were collected from the triploid and diploid E. urophylla clones. Total RNA was extracted using the TRIzol Kit (Invitrogen, Carlsbad, CA, United States). The cDNA libraries were prepared using the TruSeq Stranded Total RNA HT Sample Prep Kit (Illumina, San Diego, CA, United States). Following the manufacturer’s recommended protocol, transcriptome sequencing was performed on an Illumina HiSeq 4000 platform by Lc-bio technologies Co., Ltd. (Hangzhou, China). The abundance of transcripts was expressed as reads per kilobase per million mapped reads. The transcriptome data for EgrGLK genes was log2-transformed. The expression patterns and differential expression among E. urophylla clones of different ploidies were visualized by means of a heatmap with TBtools (Chen et al., 2020). In addition, EgrGLK genes in leaves were annotated based on the GO database10 to understand their functions.
Co-expression networks were generated to identify which EgrGLK genes might play an important role in chlorophyll synthesis. Transcriptome data for genes associated with chlorophyll synthesis and EgrGLK genes were subjected to a Pearson correlation analysis. Genes with a Pearson correlation coefficient within the appropriate range (r ≥ 0.6 or ≤ −0.6) were selected to generate a co-expression network using Cytoscape software (Kohl et al., 2011).
To determine the reliability of the RNA-seq data, 5 EgrGLK genes in leaves were selected for qRT-PCR analysis. Terminal buds, young leaves, mature leaves, and senescent leaves of triploid and diploid E. urophylla were used for qRT-PCR analysis. qPCR was subsequently performed using a TransStart® Tip Green qPCR SuperMix (TransGen Biotech, Beijing, China) in 25 μl volume on the 7500 Fast real-time PCR system (Thermo Fisher, Singapore) according to the manufacturer’s instructions. Three technical replicates and three biological replicates were performed on all reactions. The primers and reference gene used for qRT-PCR analysis are listed in Supplementary Table S1.
To understand the regulation of gene expression at the post-transcriptional level, we predicted the putative miRNAs targeting EgrGLK genes. The miRNA sequences of E. grandis were downloaded from a previous study, and the miRNAs found in leaves were used for analysis (Lin et al., 2018). The miRNAs targeting EgrGLK genes were predicted by submitting the miRNAs and EgrGLK genes to psRNATarget.11 Cytoscape was used to establish the regulatory network of miRNAs and EgrGLK genes (Kohl et al., 2011).
A total of 40 putative GLK proteins were identified in the E. grandis genome database (Table 1 and Supplementary Table S2). The online tools NCBI Web CD-Search and SMART were used to verify the identity of the proteins to ensure that they contained conserved GLK domains (Supplementary Table S3). The verified proteins were designated EgrGLK1 to EgrGLK40. The molecular weight and isoelectric point of each EgrGLK protein are listed in Table 1. The proteins ranged in size from 102 aa (EgrGLK16) to 689 aa (EgrGLK12). The molecular weight ranged from 11.69 kDa (EgrGLK16) to 74.96 kDa (EgrGLK12). The isoelectric point ranged from 4.77 (EgrGLK4) to 10.22 (EgrGLK16). In addition, 38 of the 40 EgrGLK proteins were predicted to be localized in the nucleus.
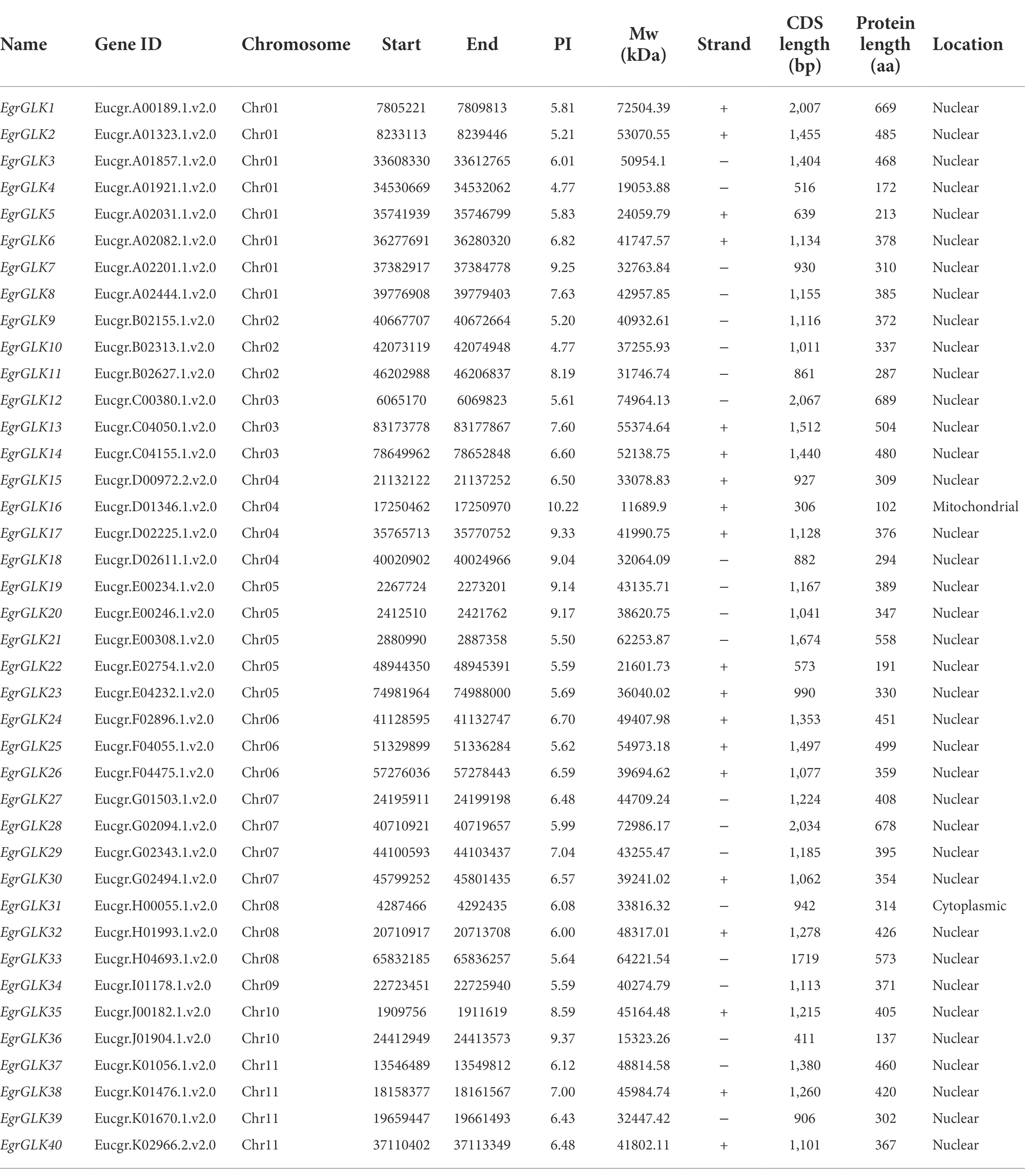
Table 1. Physical parameters of GLK transcription factors in Eucalyptus grandis.
Based on chromosomal position data, the 40 EgrGLK genes were mapped to 11 chromosomes (Figure 1). The distribution of EgrGLK genes on chromosome was uneven. Chr1 carried eight genes, whereas Chr9 contained only one gene. The longest chromosome, Chr3, contained three genes, only one gene more than the shortest chromosome (Chr10). The number of EgrGLK genes on the other chromosomes ranged from three to five. Thus, no correlation between chromosome length and EgrGLK gene number was apparent. Investigation of potential duplication events identified five duplicated pairs of EgrGLK genes as the products of segmental duplication (Figure 1 and Supplementary Table S4). In addition, the synteny relationships were displayed by comparing the genome of E. grandis with those of the other three species (Figure 2). These species include two dicotyledons (Arabidopsis and tomato) and one monocotyledon (maize). A total of 41, 39 and 9 homologous gene pairs were identified between E. grandis and the three species, respectively. To estimate the type of evolutionary selection on the duplicated EgrGLK genes, the Ka, Ks, and Ka/Ks ratio among the gene pairs were calculated (Supplementary Table S4), which indicated that all gene pairs were subject to purifying selection (Ka/Ks < 1).
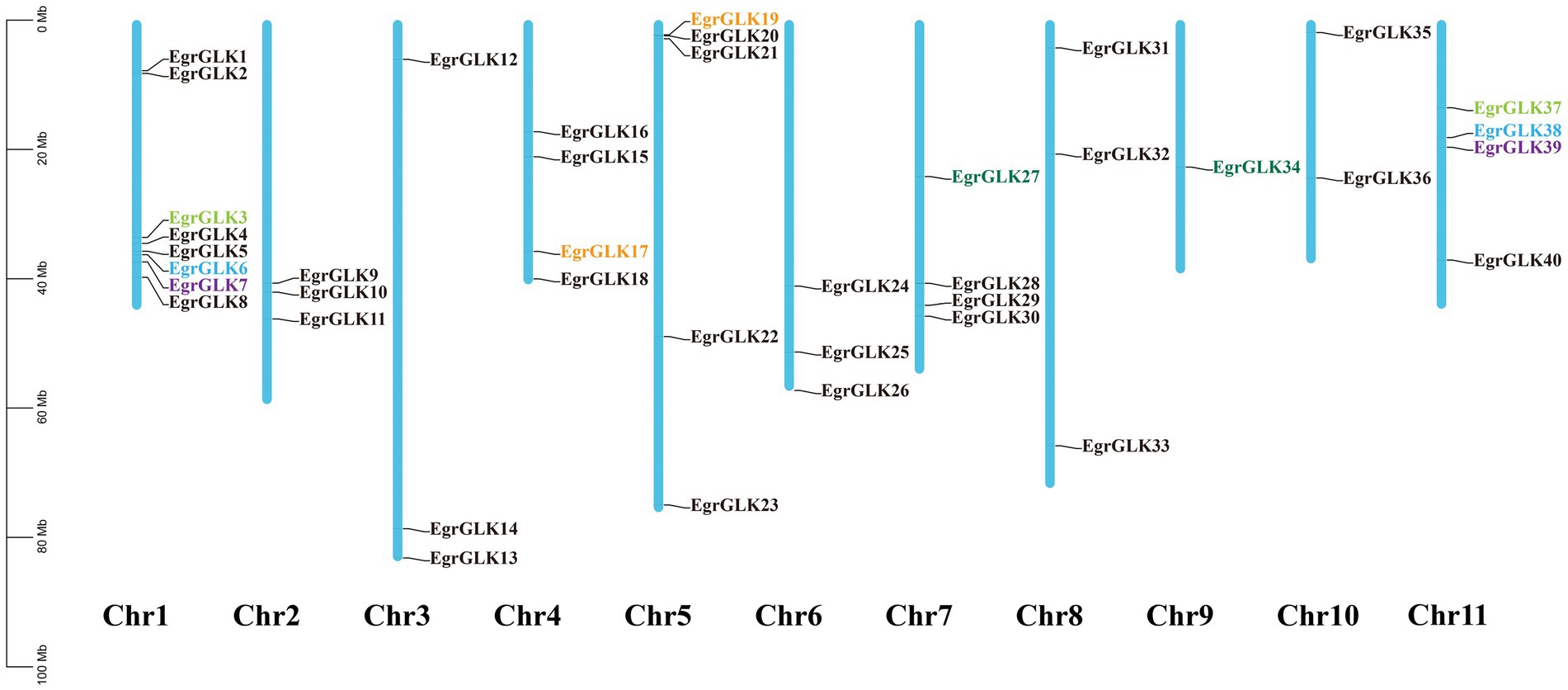
Figure 1. Chromosomal location of EgrGLK genes in Eucalyptus grandis. Genes of the same color represents a pair of segmented duplicated genes, and genes of different colors represent different gene pairs. No segmental duplication in black genes.
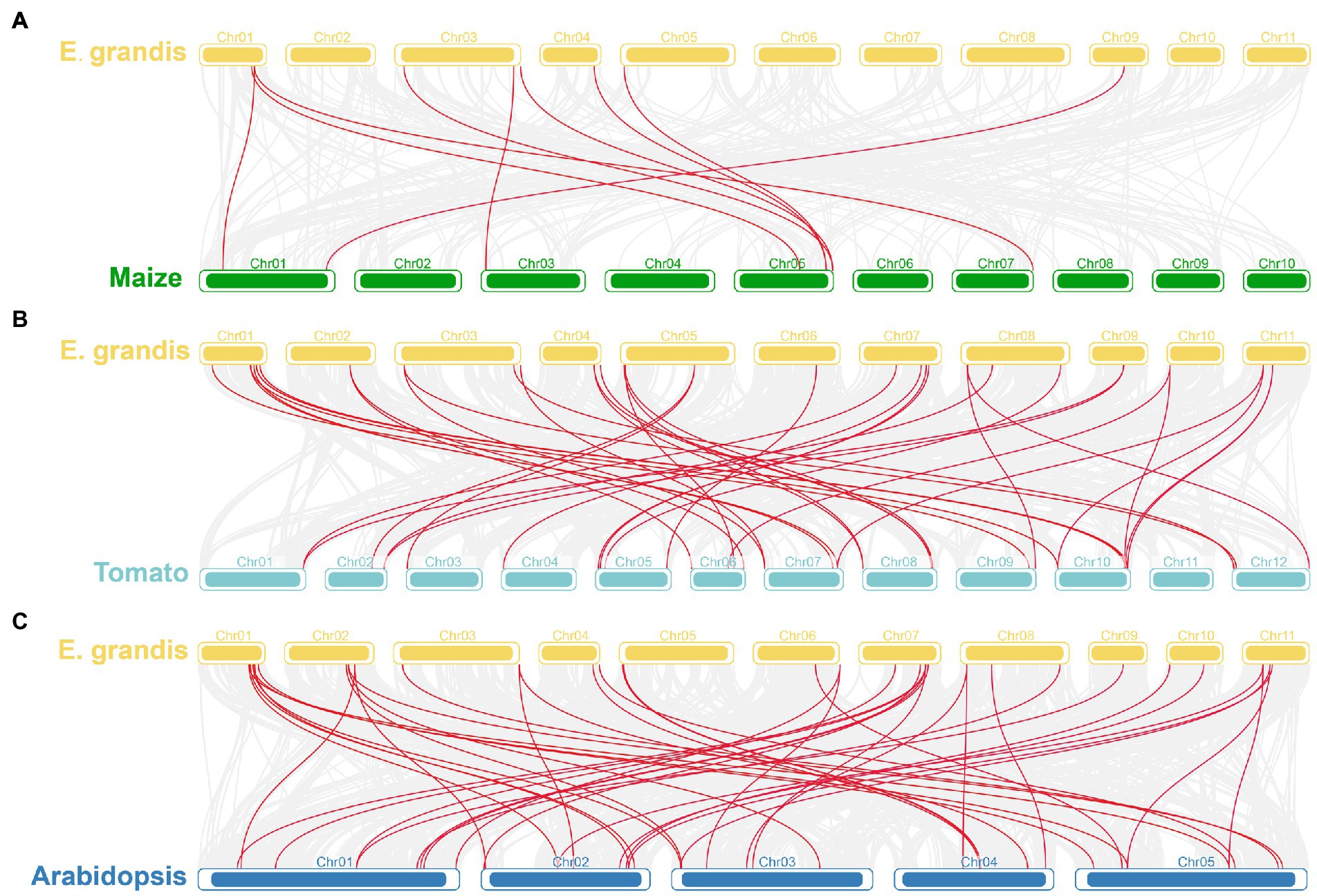
Figure 2. Synteny analysis of GLK genes in different plants. Synteny analyses of GLK genes between E. grandis and maize (A), tomato (B), and Arabidopsis (C). Red lines indicate the homologous GLK genes between E. grandis genome and other plant genomes.
To explore the evolutionary relationships of GLK proteins in plants and classify the identified EgrGLK proteins, a neighbor-joining tree was constructed. The EgrGLK proteins were clustered into seven groups based on their grouping with Arabidopsis, maize, and tomato GLK proteins (Figure 3). The GLK proteins from all four species were clustered in each group, but the 40 EgrGLK proteins were unevenly distributed. Eight EgrGLK proteins were clustered in group VII, which was double the number of EgrGLK proteins in groups IV and VI. Six, six, seven, and five EgrGLK proteins were included in groups I, II, III, and V, respectively. EgrGLK proteins were distributed in each group, which provided information on the orthologous relationships and strong evolutionary conservation among GLK proteins of different species.
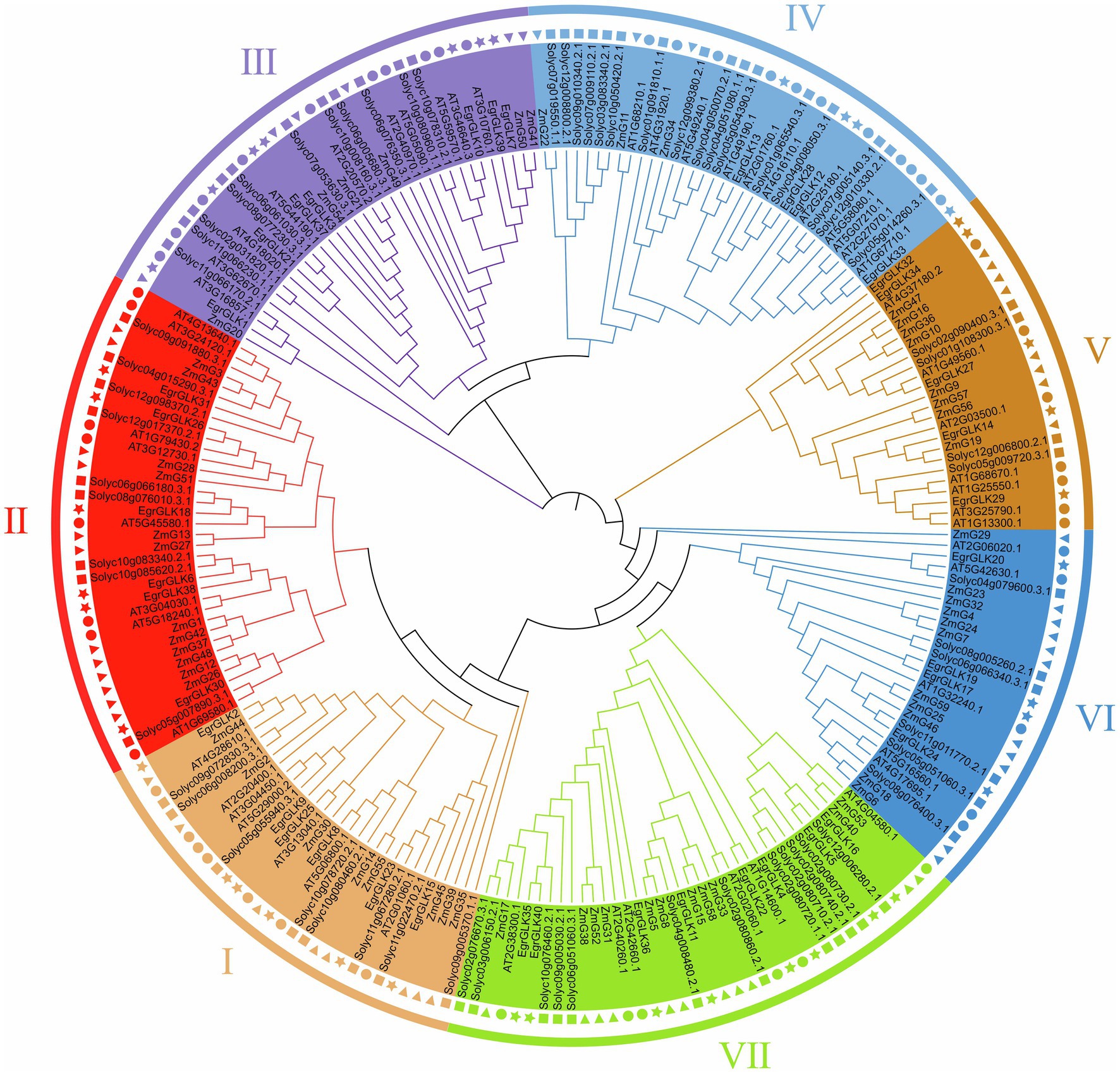
Figure 3. Phylogenetic analysis of GLK proteins from E. grandis, maize, Arabidopsis, and tomato. Different GLK protein groups are indicated by different colors. The star, triangle, circle, and square represent E. grandis, maize, Arabidopsis, and tomato GLK proteins, respectively.
To further explore evolutionary relationships among the EgrGLK genes, a phylogenetic tree was generated for the 40 EgrGLK protein sequences. The proteins were resolved into seven groups (Figure 4A). The conserved motifs of the proteins were analyzed using the online MEME tool, and the conserved sequences of each motif are shown in Supplementary Table S5. Seven putative motifs were functionally annotated, which were defined as MYB-SHAQKYF for motif 1, components of the conserved GLK domain for motifs 2 and 10, MYB-CC-LHEQLE for motif 3, and the REC superfamily for motifs 4, 5, and 7. No functional annotation was assigned for the remaining three putative motifs (Figure 4B). The MYB-SHAQKYF motif was observed to be a component of the conserved GLK domain. All EgrGLK proteins contained motifs 1 and 2, which indicated that these two motifs constituted the basic GLK domain associated with the typical function. The proteins contained different motifs in accordance with the phylogenetic grouping. Motif 3 was only detected in group I, motifs 4, 5, and 7 were coincident in group VII, and motifs 6 and 9 were mostly present in groups IV and III, respectively. Motif 8 was only detected in EgrGLK19, EgrGLK20, and EgrGLK28. Motif 10 was only identified in group VI, which was the only group to contain three conserved GLK motifs. In general, proteins in the same group contained basically the same conserved motifs, indicating that these proteins perform similar functions within a group.
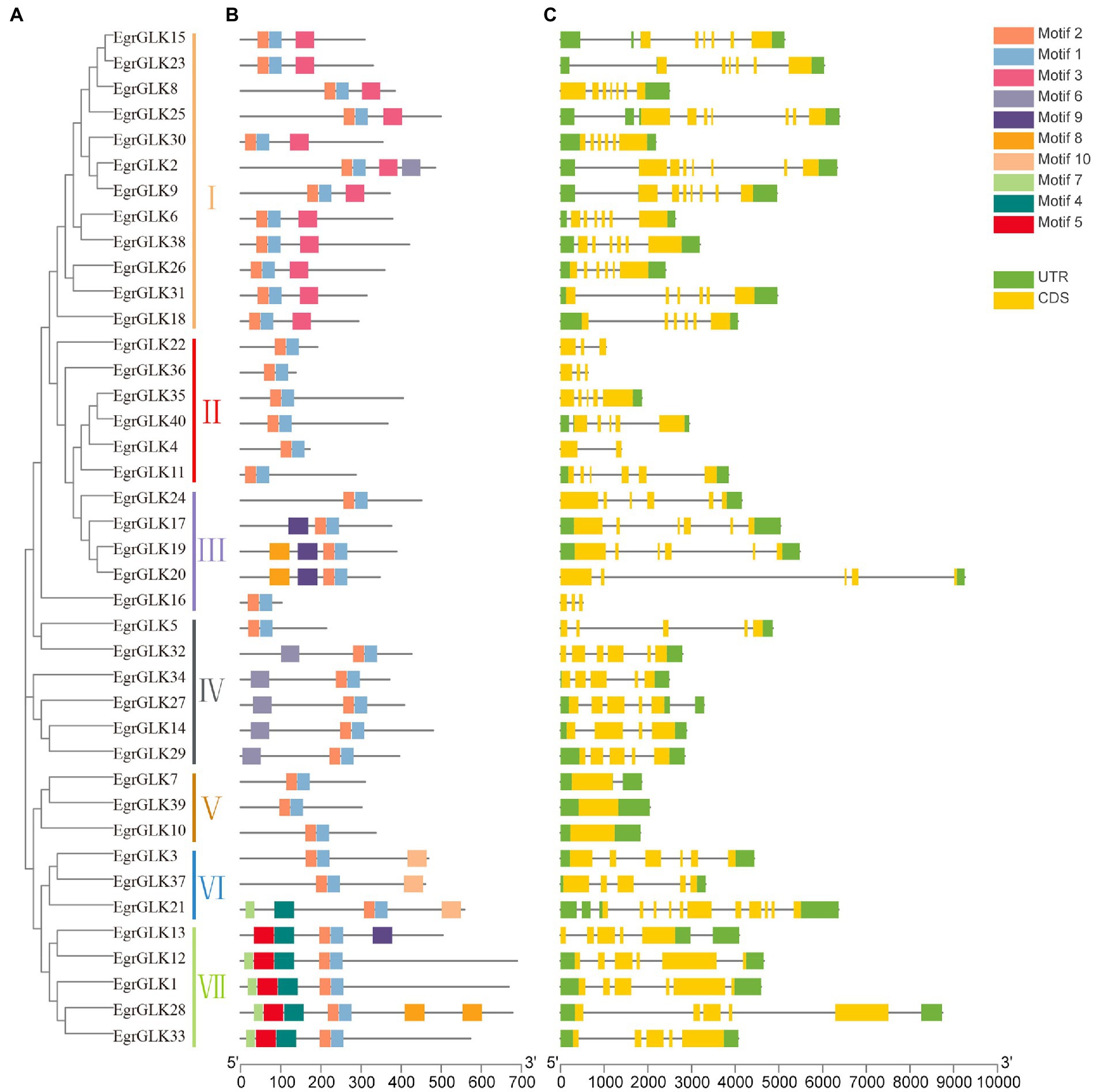
Figure 4. Phylogenetic tree, conserved motifs, and gene structure of EgrGLK genes of E. grandis. (A) Phylogenetic reconstruction for EgrGLK proteins. (B) Ten conserved motifs indicated by different colors. (C) Gene structure with exons and introns indicated.
To examine the structural variation among the EgrGLK genes, the exon–intron organization of each EgrGLK gene was assessed based on the phylogenetic classification (Figure 4C). Differences in the number of introns between genes were observed in different groups. No introns were detected in group V, and the gene (EgrGLK21) with the most introns was classified in group VI. In group II, the intron number ranged from one to five. In addition, most EgrGLK genes that were clustered in the same phylogenetic group showed similar exon–intron structures.
The presence of cis-acting regulatory elements in promoter regions is important for the expression of downstream target genes and the regulation of transcription factor interaction. Therefore, cis-regulatory elements related to development, light response, and hormone response in the promoter regions of the EgrGLK genes were investigated (Supplementary Table S6 and Figure 5). The most abundant putative cis-elements were involved in hormone response and comprised ABRE, CGTCA-motif, TGACG-motif, TCA-element, TATC-box, and AuxRR-core elements. The ABRE elements were distributed in the promoter regions of 33 EgrGLK genes and are involved in abscisic acid response. The CGTCA-motif and TGACG-motif are involved in methyl jasmonate response; thus, cis-regulatory elements responsive to methyl jasmonate were the most frequent. Three types of light-responsive elements were identified, namely ACE, G-box, and C-box. Among all cis-regulatory elements identified, G-box elements were the most widely distributed and were identified in 34 EgrGLK genes, which indicated that many EgrGLK genes may be sensitive to light. In addition, five cis-elements involved in development were identified, comprising CAT-box, circadian, GCN4-motif, RY-element, MSA-like elements and motif I elements. These elements are involved in plant growth, cell division, and diverse plant-specific tissues. Notably, many cis-regulatory elements consisted of two or more copies in the 2 kb upstream region, which may enhance their binding to the corresponding transcription factors.
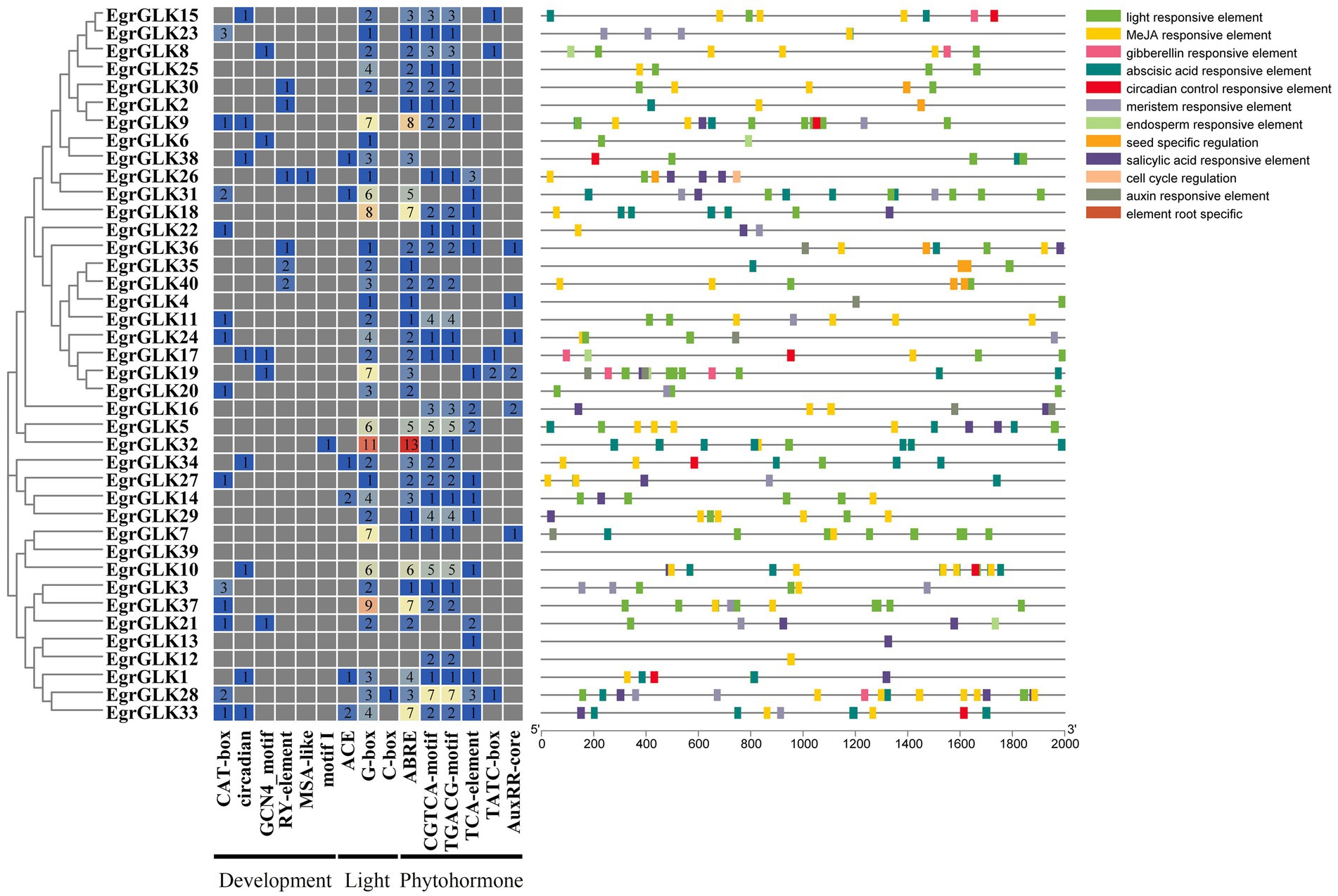
Figure 5. Analysis of cis-acting regulatory element in promoter regions of EgrGLK genes of E. grandis. Numerals in different color boxes indicate the number of the main cis-elements of EgrGLK genes. Cis-elements of different responsive types are indicated by different colors.
Triploid E. urophylla and its diploid control were used to measure the chlorophyll content in young, mature, and senescent leaves (Figure 6). The trend in chlorophyll content of the different leaves of the diploid and triploid clones was identical. With increase in leaf age, the chlorophyll content initially increased and then decreased, thus the chlorophyll content was highest in mature leaves. The leaf chlorophyll content was higher in the triploid than in the diploid, and in young and mature leaves the chlorophyll content of the triploid was significantly higher than that of the diploid.
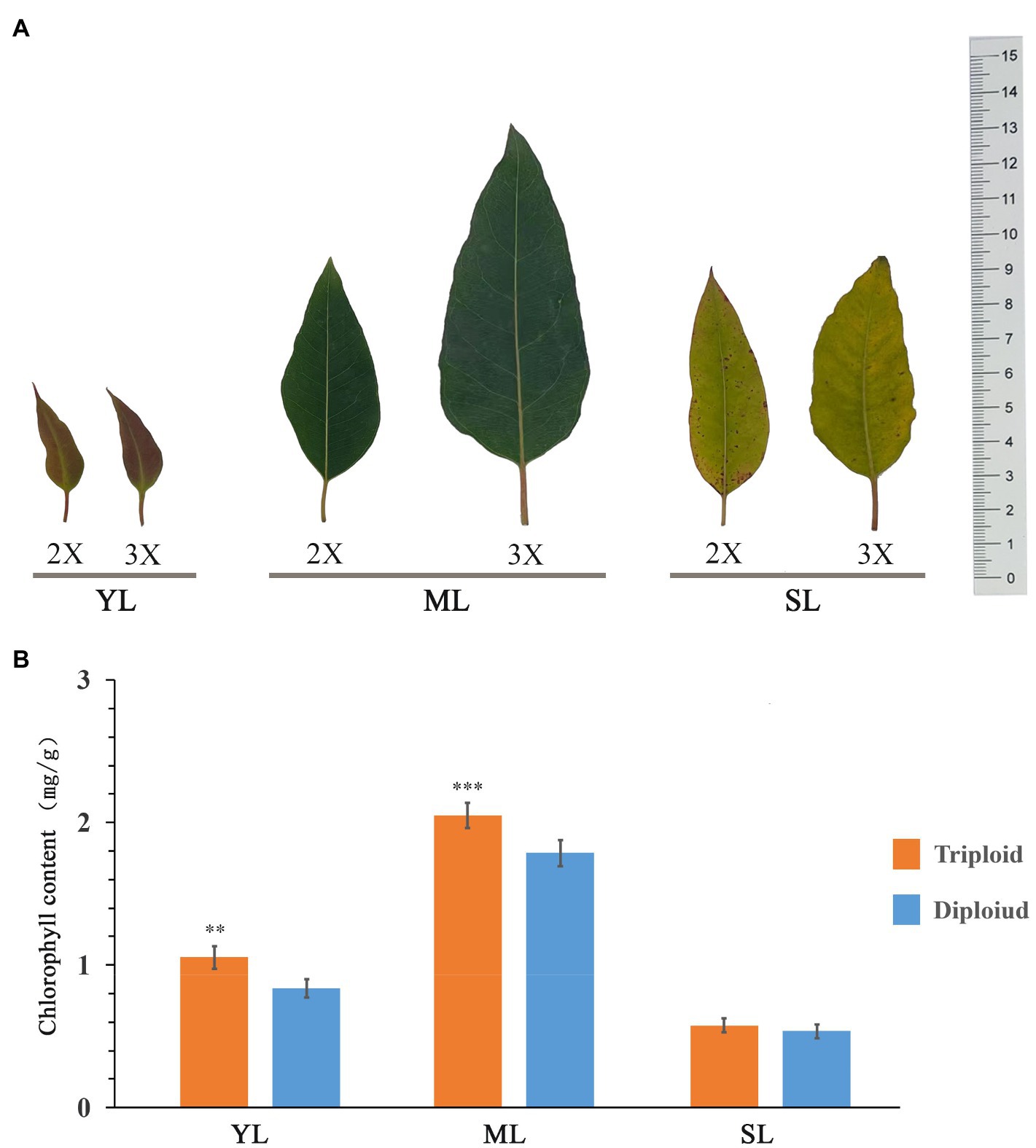
Figure 6. Measurement of chlorophyll content in leaves of E. urophylla. Phenotype (A) and chlorophyll content (B) of young leaves (YL), mature leaves (ML), and senescent leaves (SL) of diploid (2X) and triploid (3X) clones.
To explore the influence of EgrGLK gene expression on chlorophyll synthesis in diploid and triploid E. urophylla, transcriptome data from leaves of four developmental stages (terminal buds, young leaves, mature leaves, and senescent leaves) were used to analyze the transcript abundance. The transcripts of 36 EgrGLK genes were detected in the leaves. Then, the GO term classification and enrichment analysis of 36 EgrGLK genes were performed. The first twenty GO terms were shown in the Supplementary Figure S2 according to the significance of enrichment. And the most enriched five GO terms were ‘regulation of transcription’, ‘DNA-binding transcription factor activity’, ‘transcription’, ‘DNA binding’ and ‘nucleus’. Among them, there were 35 and 33 EgrGLK genes involved in ‘nucleus’ and ‘regulation of transcription’, respectively, indicating that EgrGLK genes mainly functions as transcription factors in the nucleus.
The expression data for these 36 EgrGLK genes were log2-transformed and used to generate a clustered heatmap to visualize the expression patterns at each leaf developmental stage (Figure 7). The 36 EgrGLK genes were divided into four and three groups in diploid and triploid E. urophylla, respectively. Ten EgrGLK genes were included in group I of the diploid and 15 EgrGLK genes were included in group I of the triploid, which exhibited low transcript levels at each developmental stage. Group IV of the diploid and group III of the triploid contained 15 and 10 EgrGLK genes, respectively, and exhibited relatively high transcription levels. Group III of the diploid and group II of the triploid contained eight and 11 EgrGLK genes, respectively, and these genes exhibited high transcript abundance in all analyzed leaves, hinting that these genes were essential in E. urophylla leaves. It was noteworthy that EgrGLK6, EgrGLK35, and EgrGLK40 were grouped in group II of the diploid. These three genes showed low transcript abundance in terminal buds and young leaves, and high expression levels in mature and senescent leaves. The same expression patterns of EgrGLK6, EgrGLK35, and EgrGLK40 were observed in leaves of the triploid; however, the differential expression of these three EgrGLK genes in the triploid was more moderate than in the diploid, hence they were not clustered into a separate group. In general, the expression patterns of most EgrGLK genes in leaves of the diploid and triploid were approximately identical.
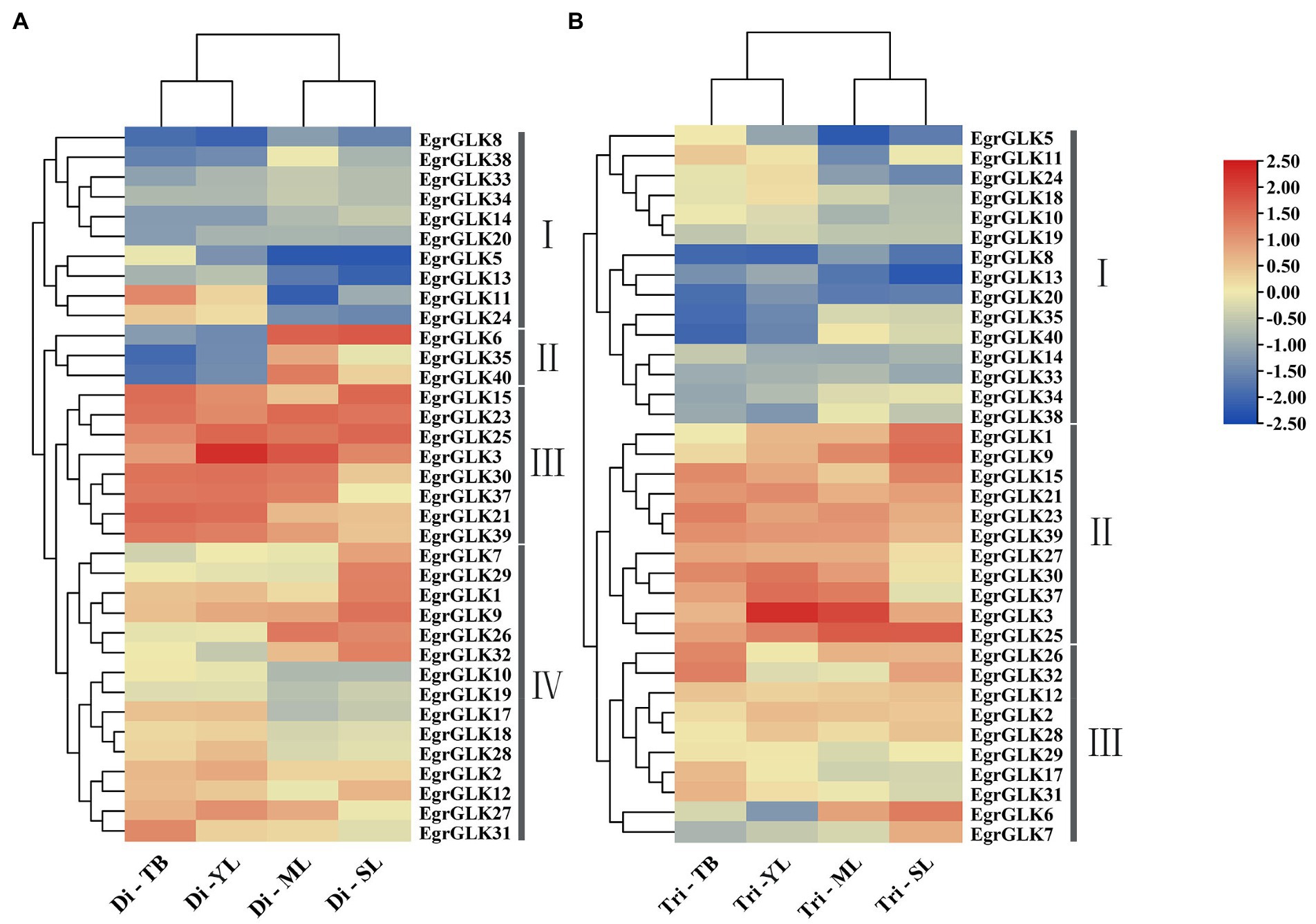
Figure 7. Expressions patterns of EgrGLK genes in leaves of E. urophylla. The expression patterns of EgrGLK genes in the terminal bud (TB), young leaves (YL), mature leaves (ML), and senescent leaves (SL) of diploid (A) and triploid (B) clones.
To further explore the effect of differential expression of EgrGLK genes on chlorophyll synthesis in leaves of E. urophylla of different ploidies, the expression data for the EgrGLK genes in diploid E. urophylla leaves were used as the control group, and the fold change in expression between triploid leaves and diploid leaves was used to generate heat maps (Figure 8). More than half of the EgrGLK genes were down-regulated in terminal buds of triploid E. urophylla. In contrast, 30, 24, and 28 EgrGLK genes were upregulated in young, mature, and senescent leaves of triploid E. urophylla (Supplementary Table S7). Among the genes highly expressed in leaves that were common to diploid and triploid E. urophylla (Figure 7, EgrGLK23, EgrGLK30, EgrGLK3, EgrGLK37, EgrGLK15, EgrGLK25, and EgrGLK39), EgrGLK3, EgrGLK15, EgrGLK25, EgrGLK37 and EgrGLK39 were upregulated in triploid young (1.03, 1.05, 1.10, 1.01 and 1.04 fold change, respectively), mature (1.77, 1.22, 2.03, 1.48 and 1.46 fold change, respectively), and senescent leaves (1.04, 1.13, 1.91, 1.24 and 1.84 fold change, respectively), and down-regulated in triploid terminal buds (Figure 8). Three genes were highly expressed only in the triploid, of which EgrGLK1 and EgrGLK9 showed the same differential expression pattern as the common highly expressed genes, and the other gene (EgrGLK27) was upregulated in all analyzed leaves. To further confirm the effect of polyploidization on gene expression, genes upregulated in triploid including EgrGLK3, EgrGLK15, EgrGLK25, EgrGLK37 and EgrGLK39 were selected for qRT-PCR analysis (Supplementary Figure S3). Correlation analysis showed that there was a high correlation coefficient between qRT-PCR and RNA-seq (R2 = 0.826, p < 0.01), indicating that the differential expression of EgrGLK genes among different ploidies was reliable. Differential expression of EgrGLK genes provided preliminary information for the study of chlorophyll synthesis and leaf development of E. urophylla with different ploidies.
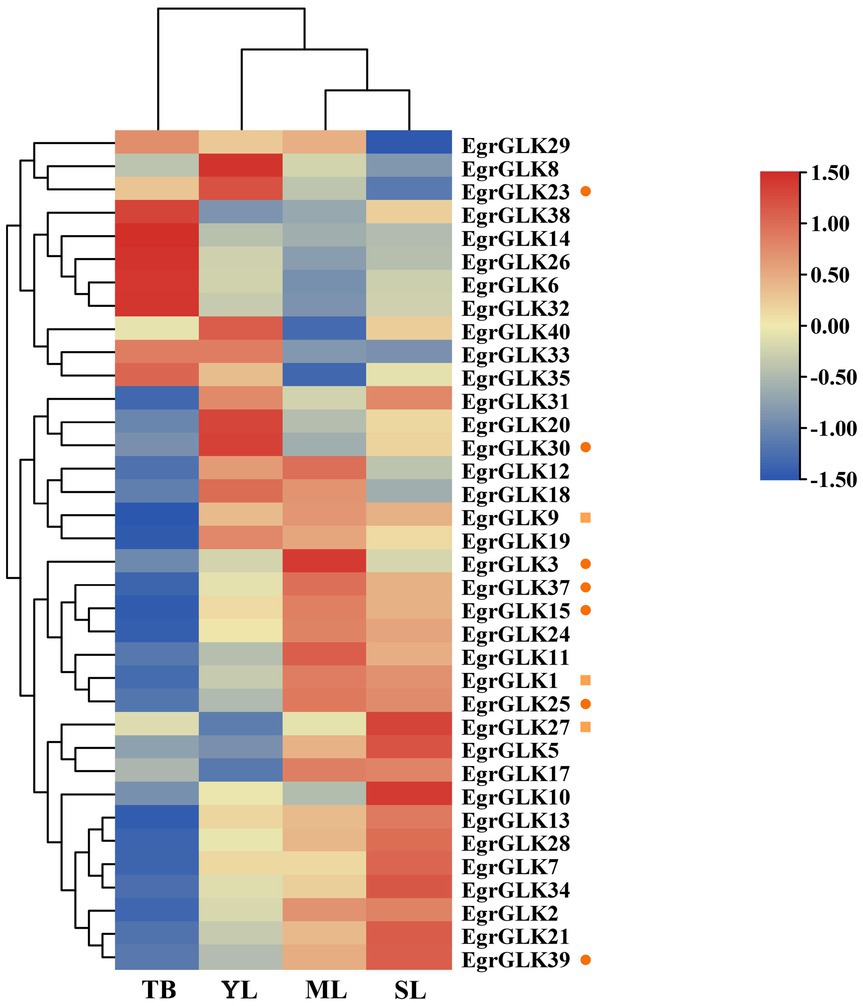
Figure 8. Differential expression of EgrGLK genes in the terminal bud (TB), young leaves (YL), mature leaves (ML), and senescent leaves (SL) of diploid and triploid clones. Circles represent highly expressed genes common to the diploid and triploid clones, and squares represent genes highly expressed only in the triploid clone.
Transcriptome data for genes associated with chlorophyll synthesis and the EgrGLK genes were used for a correlation analysis to explore the role of EgrGLK genes in chlorophyll synthesis (Supplementary Table S8). The transcriptome data for chlorophyll synthesis related genes and EgrGLK genes that were highly correlated (r ≥ 0.6 or ≤ −0.6) were used to generate co-expression networks (Figure 9). Twenty-five EgrGLK genes and 21 chlorophyll synthesis related genes were involved in the positive correlation co-expression network, forming a total of 115 correlation network lines. Among these genes, EgrGLK3 and EgrGLK37 were correlated with 17 and 16 chlorophyll synthesis related genes, respectively. The negative correlation co-expression network incorporated 14 EgrGLK genes and 16 chlorophyll synthesis related genes, forming a total of 36 correlation network lines. Among these genes, EgrGLK32 was correlated to 10 chlorophyll synthesis related genes. These results showed that EgrGLK genes and chlorophyll synthesis related genes were mainly positively correlated, and thus that EgrGLK genes may play a positive regulatory role in chlorophyll synthesis.
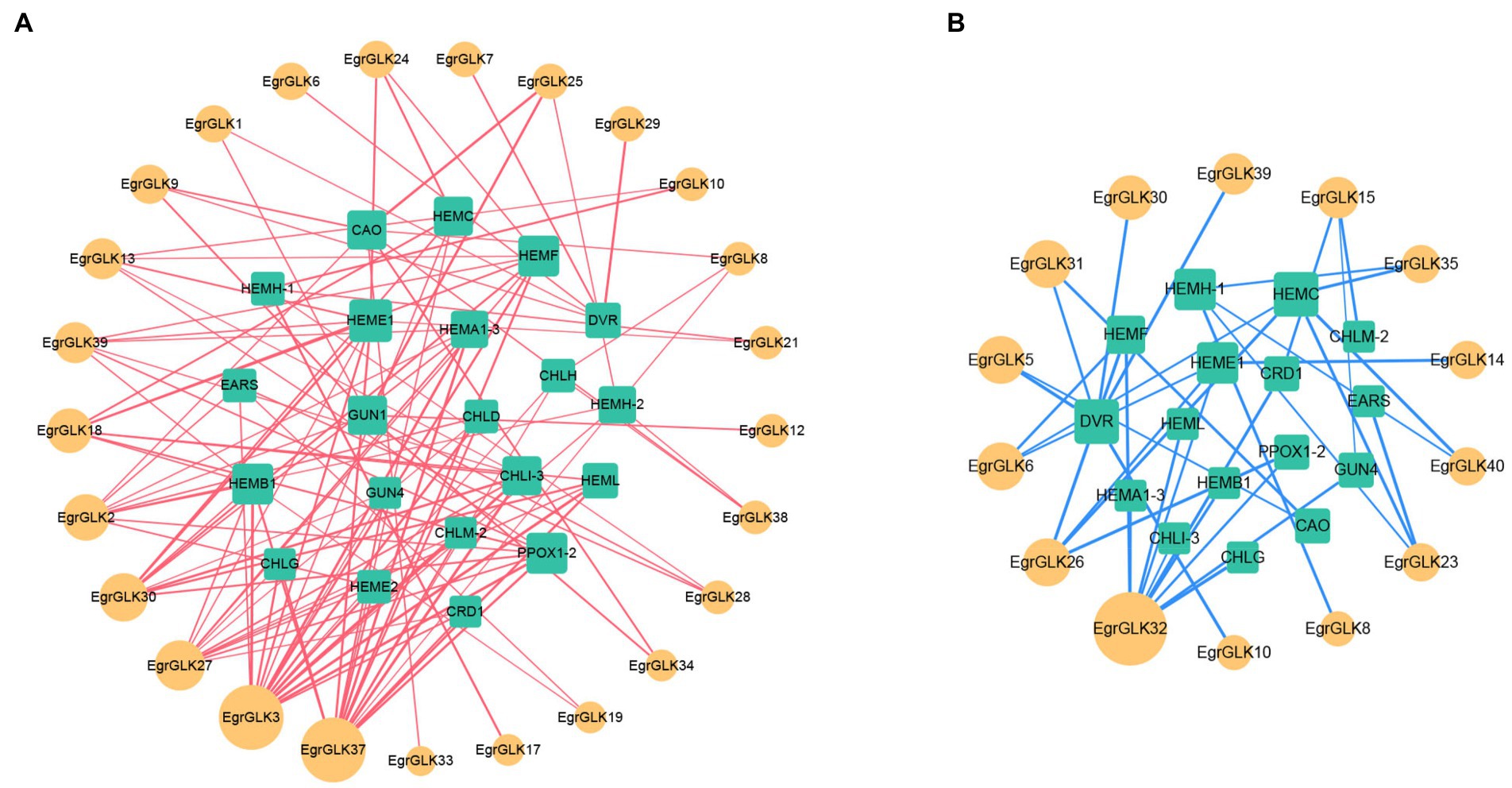
Figure 9. Co-expression network of EgrGLK genes and chlorophyll synthesis related genes. Positive correlation network (A) and negative correlation network (B) between EgrGLK genes and chlorophyll synthesis related genes. Circles represent EgrGLK genes, squares represent chlorophyll synthesis related genes, and the size of the circle or square represents the number of relationships between genes.
In plants, microRNAs (miRNAs) have been found to play important roles in the regulation of gene expression at the post-transcriptional level. We predicted the miRNAs targeting EgrGLK genes to further reveal the possible reasons for the differential expression of EgrGLK genes in different ploidies. In previous studies, 179 miRNAs were found in leaves (Lin et al., 2018), of which 85 miRNAs from 23 families were involved in the regulation of 31 EgrGLK genes (Supplementary Table S9). According to the targeting relationship between miRNAs and EgrGLK genes, 12 regulatory networks were generated and they were divided into 5 groups (Figure 10). Figure 10A showed the most complex regulatory network containing 52 miRNAs and 16 EgrGLK genes, while six miRNAs in Figure 10E showed the simplest regulatory network in the type of one-to-one target gene. EgrGLK3, EgrGLK32 and EgrGLK37 has been identified as the genes that play an important role in chlorophyll synthesis, and there were nine, four and one miRNAs targeting these three genes, respectively. In addition, five EgrGLK genes including EgrGLK10, EgrGLK27, EgrGLK35, EgrGLK29, EgrGLK31 with the most targeting relationships were found and they were regulated by 15, 11, 8, 8 and 8 miRNAs, respectively. These results suggest that miRNAs play an important role in the regulation of EgrGLK gene expression.
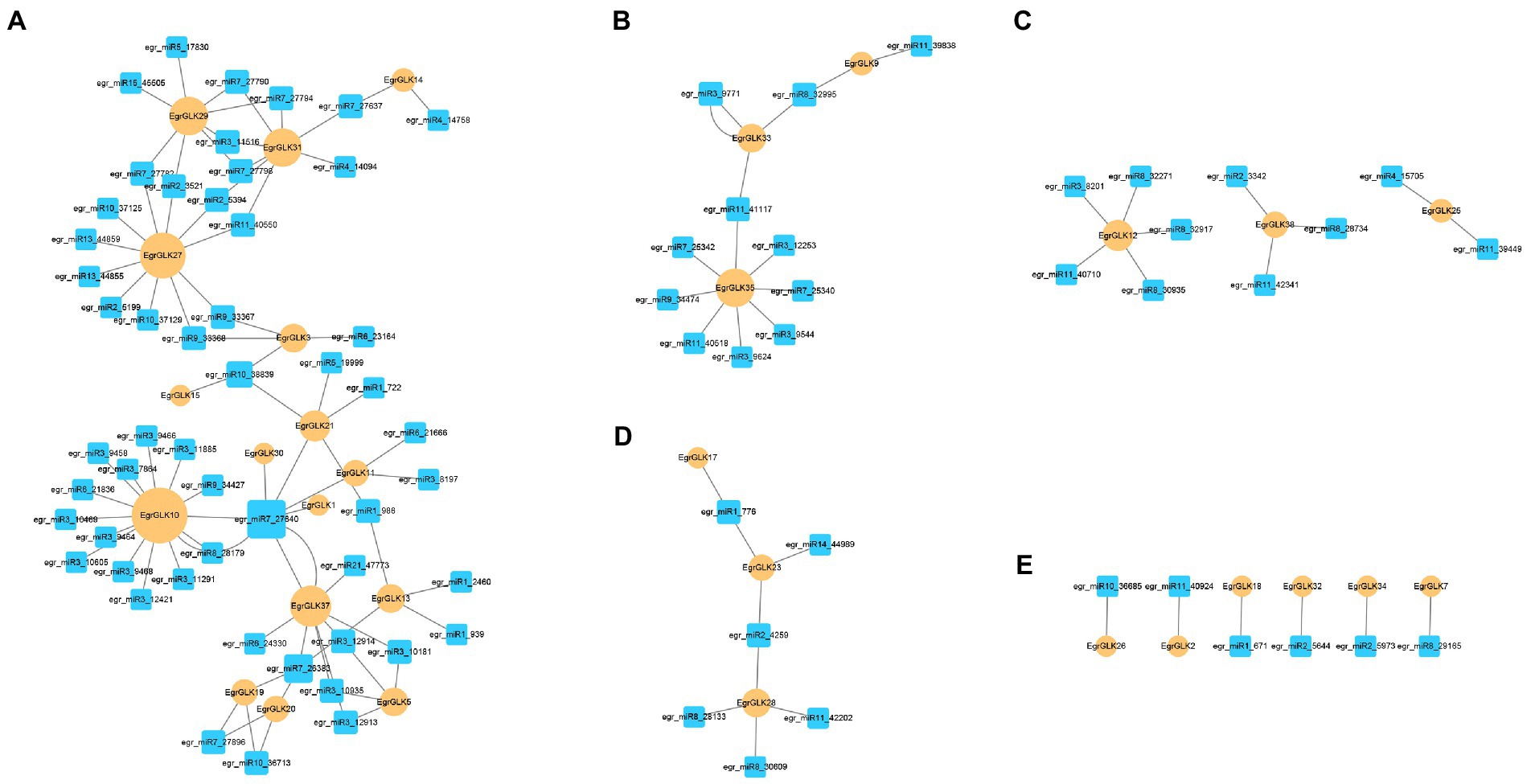
Figure 10. Regulatory networks were divided into (A-E) according to the number of targeting relationship between miRNAs and EgrGLK genes.
In this study, 40 EgrGLK genes were identified in the E. grandis genome (Supplementary Table S2). The number of EgrGLK genes was less than the number of GLK genes identified in Arabidopsis (55), maize (59), and tomato (66) (Liu et al., 2016; Alam et al., 2022; Wang et al., 2022). It was suspected that some EgrGLK genes were lost during evolution (Du et al., 2022). It was observed that the number of GLK family members was independent of genome size. The diversity of GLK family members in different species may be influenced by genome duplication events, such as whole-genome duplication, segmental duplication, or tandem duplication (Zhang, 2003; Chang and Duda., 2012). In the present study, five segmental duplication gene pairs were identified among the EgrGLK genes, indicating that segmental duplication was the main contributor to expansion of GLK genes in E. grandis (Figure 1). It has previously been reported that segmental duplication is more common than tandem duplication in the GLK gene family, thus the former might play an important role in chloroplast evolution (Song et al., 2016; Wu et al., 2016). The synteny relationships of GLK genes between E. grandis and other species showed that the number of homologous gene pairs between E. grandis and dicotyledons was much more than that between E. grandis and monocotyledons, indicating that GLK gene family has been amplified after differentiation between dicotyledon and monocotyledon (Figure 2). The EgrGLK genes were classified into seven groups based on a phylogenetic analysis (Figure 3), which was consistent with the classification of GLK genes in Arabidopsis (Alam et al., 2022). This result indicated that, although some EgrGLK genes were lost during evolution, the sufficient genetic diversity has been retained in E. grandis.
As previously reported, the structural characteristics of gene families may reflect their evolutionary trends (Haas et al., 2008; Nie et al., 2022), whereas the conserved motifs reflect their protein-specific functions (Lai et al., 1998; Yang et al., 2021). The 40 EgrGLK genes were divided into seven groups, and the gene structure and motif arrangement of the genes in the same group were similar (Figure 4). This finding indicated that the EgrGLK genes in the same group might have similar functions (Li et al., 2022; Liang et al., 2022). In the current study, seven motifs were functionally annotated using the NCBI Web CD-Search Tool, which have also been detected in the GLK family members in tobacco (Qin et al., 2021). Thus, excluding the GLK motifs, other motifs detected in the GLK proteins were also relatively conserved. Moreover, group VI was the only group that contained three conserved GLK motifs; this phenomenon may enhance the regulatory role of these three EgrGLK genes as transcription factors.
The cis-acting elements may be activated by trans-acting elements to regulate the activity of the target genes (Valli et al., 2022). Among the cis-acting elements detected in the promoter regions of EgrGLK genes (Figure 5), elements associated with development were only sporadically distributed in the promoter regions, whereas elements involved in light response and hormone response were observed in the promoter regions of almost all EgrGLK genes. The motifs involved in methyl jasmonate response comprised the CGTCA-motif and TGACG-motif, which were the most frequent cis-regulatory elements identified. These motifs play important roles in multiple physiological processes, including development, senescence, and response to diverse environmental stresses (Browse and Howe, 2008; Wang et al., 2011). The G-box was indicated to be the most widely distributed element. The G-box element may be unique to light regulation and is a ubiquitous element in functionally diverse genes (Menkens et al., 1995; Waters et al., 2009). These results suggested that EgrGLK genes are controlled by light and phytohormones (Nakamura et al., 2009; Lupi et al., 2019). The diversity of types, quantity, and distribution of cis-acting elements in the gene promoters reflects the complex response regulatory mechanism and complex evolutionary relationships of the EgrGLK genes.
The GLK genes play an important role in chloroplast development and chlorophyll synthesis (Fitter et al., 2002; Chen et al., 2016). The expression pattern of GLK family genes in E. urophylla leaves that differ in chlorophyll content was studied to explore the effect of the genes on chlorophyll synthesis. The chlorophyll content in young and mature leaves of triploid E. urophylla was significantly higher than that of the diploid clone. Such an increase in chlorophyll content has been observed in other polyploid plants, such as Populus and Rhododendron fortunei (Liao et al., 2016; Mo et al., 2020). Thirty-six EgrGLK genes were expressed in leaves of both diploid and triploid E. urophylla. The EgrGLK genes not expressed in the leaves might be tissue-specific genes (Deveaux et al., 2003; Lim et al., 2012). The 36 EgrGLK genes in the diploid and triploid clones were divided into four and three groups, respectively, based on transcriptome data. The genes EgrGLK6, EgrGLK35, and EgrGLK40 in leaves of the diploid were clustered in group II, but did not form a separate group in the triploid (Figure 7). The promoter regions of these EgrGLK genes in group II of the diploid contained cis-acting elements associated with the seed and endosperm (Figure 5). Triploids are characterized by sterility and thus whether the differential expression pattern of these three genes is associated with triploid sterility requires further study (Fujiwara and Beachy, 1994). Except for group II, other groups in the diploid showed the same expression pattern as in the triploid, which could be divided into low, relatively high, or high expression levels based on the expression data (Figure 7). Compared with the diploid, three additional genes in the triploid were clustered into the high-expression-level group. The high expression levels of a greater number of genes in polyploids might be caused by the participation of duplicated genes derived from whole-genome duplication (Jackson and Chen, 2010; Roulin et al., 2013). The high expression levels of EgrGLK1, EgrGLK9, and EgrGLK27 in the triploid may be one factor that promotes the increase in chlorophyll content in leaves of triploid E. urophylla.
The differential expression of GLK genes in leaves also affects chloroplast development and chlorophyll synthesis (Nguyen et al., 2014). In Arabidopsis, overexpression of GLK genes increases the chlorophyll content in leaves (Waters et al., 2009). In the present study, EgrGLK genes were differentially expressed in triploid and diploid E. urophylla leaves (Figure 8). In the terminal bud, most EgrGLK genes in the triploid were down-regulated compared with the diploid, whereas in young, mature, and senescent leaves, more than two-thirds of the EgrGLK genes were up-regulated in the triploid. The down-regulation of EgrGLK genes in the terminal bud of the triploid did not affect the chlorophyll content of mature leaves, and the chlorophyll content of mature leaves was significantly higher in the triploid than in the diploid (Figure 6). The chlorophyll content in leaves increases gradually with increase in leaf age before the leaves are fully developed and attain maturity (Bertamini and Nedunchezhian, 2002; Du et al., 2020). This is consistent with the present results for E. urophylla, indicating that the chlorophyll content might be associated with the expression of EgrGLK genes during leaf development. The chlorophyll content of young, mature, and senescent leaves of the triploid was higher than that of diploid. The differential expression of EgrGLK genes in leaves of E. urophylla of different ploidies was consistent with the trend in chlorophyll content. To further verify the relationship between EgrGLK gene expression and chlorophyll synthesis, co-expression networks of the EgrGLK genes and chlorophyll synthesis related genes were generated (Figure 9). The networks indicated that EgrGLK genes may positively regulate chlorophyll synthesis (Waters et al., 2008; Brand et al., 2014; Zubo et al., 2018). In addition, based on the number of correlations between EgrGLK genes and chlorophyll synthesis related genes, three important EgrGLK genes were identified. EgrGLK37 and EgrGLK3 were involved in the positive correlation network and EgrGLK32 was included in the negative correlation network (Figure 9). Among these genes, EgrGLK37 and EgrGLK3 showed high expression levels in leaves, whereas EgrGLK32 showed relatively high expression levels in leaves (Figure 7). The present results provide a reference for further studies of the relationship between EgrGLK genes and chlorophyll synthesis.
Mature miRNA combines with the RNA-induced silencing complex, which interacts with target genes to regulate the expression of genes by inhibiting gene translation or degrading targeted mRNAs (Baulcombe, 2004). In this study, 85 miRNAs targeting EgrGLK genes were predicted, which is almost half of the miRNAs in leaves found in previous study (Lin et al., 2018). EgrGLK10, EgrGLK27, EgrGLK29, EgrGLK31, EgrGLK35 and EgrGLK37 are all regulated by more than eight miRNAs, of which five EgrGLK genes do not belong to the group with high transcription level (Figure 7). The result confirmed the inhibitory effect of miRNA on gene expression (Pappas Mde et al., 2015; Unnikrishnan and Shankaranarayana, 2020). EgrGLK37 is a gene regulated by nine miRNAs but with high expression level. And it is speculated that EgrGLK37 may also be affected by other regulatory mechanisms, such as lncRNA (Lin et al., 2019; Chen et al., 2021). In addition, EgrGLK37 was also found to be associated with the most chlorophyll related genes in the co-expression network, indicating that EgrGLK37 may play an important role in the gene expression regulation network. In this study, three types of miRNAs were found, including one to multiple target genes, one to one target gene and multiple miRNAs to a common target gene, which is consistent with the results found in previous miRNA studies (Lin et al., 2018). Differential expression of EgrGLK genes among different ploidies have been proved in our study and the regulation of miRNA may also be one of the reasons for the change of gene expression after plant polyploidization.
In this study, GLK transcription factors of E. grandis were systematically analyzed using bioinformatic methods. Forty EgrGLK genes were identified in the E. grandis genome and were divided into seven groups according to the gene structure and motif composition. The number of EgrGLK genes is less than the number of GLK genes identified in other species, but the sufficient genetic diversity has been retained in E. grandis, which indicates that GLK proteins exhibit strong evolutionary conservation across diverse species. Analysis of phenotypic and transcriptome data for leaves at different developmental stages in diploid and triploid E. urophylla revealed a positive correlation between EgrGLK genes and chlorophyll synthesis. On the basis of a differential expression analysis, it was speculated that the increase in chlorophyll content in leaves of triploid E. urophylla may be caused by up-regulation of EgrGLK gene expression. In addition, three EgrGLK genes that may play an important role in chlorophyll synthesis were identified. The present research provides valuable information for further functional characterization of EgrGLK genes in Eucalyptus. In the future, increasing the expression of GLK gene in plants by polyploidy or other methods may promote photosynthesis and growth of plants, which is of great value to improve plant yield.
The data presented in the study are deposited in the GEO repository, accession number GSE207860.
JY and XK conceived and designed the research. ZL, TX, and YZ conducted the experiments. ZL, BQ, and HC collected and analyzed the data. ZL and JY wrote the manuscript. XK provided the valuable suggestions on the manuscript. All authors contributed to the article and approved the submitted version.
This research was supported by the National Natural Science Foundation of China (31901337) and the National Key R&D Program of China during the 14th Five-year Plan Period (2021YFD2200105).
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
The Supplementary materials for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fpls.2022.952877/full#supplementary-material
1. ^https://phytozome-next.jgi.doe.gov
2. ^http://smart.embl-heidelberg.de
3. ^https://www.ncbi.nlm.nih.gov/Structure/bwrpsb/bwrpsb.cgi
4. ^https://web.expasy.org/compute_pi
5. ^https://www.genscript.com/wolf-psort.html
8. ^http://meme-suite.org/meme/
9. ^http://bioinformatics.psb.ugent.be/webtools/plantcare/html/
Alam, I., Wu, X., Yu, Q., and Ge, L. (2022). Comprehensive genomic analysis of G2-Like transcription factor genes and their role in development and abiotic stresses in Arabidopsis. Diversity 14, 228. doi: 10.3390/d14030228
Allario, T., Brumos, J., Colmenero-Flores, J. M., Iglesias, D. J., Pina, J. A., Navarro, L., et al. (2013). Tetraploid Rangpur lime rootstock increases drought tolerance via enhanced constitutive root abscisic acid production. Plant Cell Environ. 36, 856–868. doi: 10.1111/pce.12021
Bertamini, M., and Nedunchezhian, N. (2002). Leaf age effects on chlorophyll, Rubisco, photosynthetic electron transport activities and thylakoid membrane protein in field grown grapevine leaves. J. Plant Physiol. 159, 799–803. doi: 10.1078/0176-1617-0597
Bhutia, K. L., Nongbri, E. L., Gympad, E., Rai, M., and Tyagi, W. (2020). In silico characterization, and expression analysis of rice Golden 2-Like (OsGLK) members in response to low phosphorous. Mol. Biol. Rep. 47, 2529–2549. doi: 10.1007/s11033-020-05337-2
Booth, T. H., Jovanovic, T., and Arnold, R. J. (2017). Planting domains under climate change for Eucalyptus pellita and Eucalyptus urograndis in parts of China and South East Asia. Aust. For. 80, 1–9. doi: 10.1080/00049158.2016.1275101
Brand, A., Borovsky, Y., Hill, T., Rahman, K. A., Bellalou, A., Van Deynze, A., et al. (2014). CaGLK2 regulates natural variation of chlorophyll content and fruit color in pepper fruit. Theor. Appl. Genet. 127, 2139–2148. doi: 10.1007/s00122-014-2367-y
Bravo-Garcia, A., Yasumura, Y., and Langdale, J. A. (2009). Specialization of the Golden2-like regulatory pathway during land plant evolution. New Phytol. 183, 133–141. doi: 10.1111/j.1469-8137.2009.02829.x
Browse, J., and Howe, G. A. (2008). New weapons and a rapid response against insect attack. Plant Physiol. 146, 832–838. doi: 10.1104/pp.107.115683
Chang, D., and Duda, T. F. Jr. (2012). Extensive and continuous duplication facilitates rapid evolution and diversification of gene families. Mol. Biol. Evol. 29, 2019–2029. doi: 10.1093/molbev/mss068
Chen, C., Chen, H., Zhang, Y., Thomas, H. R., Frank, M. H., He, Y., et al. (2020). TBtools: an integrative toolkit developed for interactive analyses of big biological data. Mol. Plant 13, 1194–1202. doi: 10.1016/j.molp.2020.06.009
Chen, M., Ji, M., Wen, B., Liu, L., Li, S., Chen, X., et al. (2016). Golden 2-Like transcription factors of plants. Front. Plant Sci. 7, 1509. doi: 10.3389/fpls.2016.01509
Chen, H., Li, J., Qiu, B., Zhao, Y., Liu, Z., Yang, J., et al. (2021). Long non-coding RNA and its regulatory network response to cold stress in Eucalyptus urophylla S.T.Blake. Forests 12, 836. doi: 10.3390/f12070836
Deng, X., Guo, S., Sun, L., and Chen, J. (2020). Identification of short-rotation Eucalyptus plantation at large scale using multi-satellite imageries and cloud computing platform. Remote Sens. 12, 2153. doi: 10.3390/rs12132153
Deveaux, Y., Peaucelle, A., Roberts, G. R., Coen, E., Simon, R., Mizukami, Y., et al. (2003). The ethanol switch: a tool for tissue-specific gene induction during plant development. Plant J. 36, 918–930. doi: 10.1046/j.1365-313X.2003.01922.x
Du, K., Liao, T., Ren, Y., Geng, X., and Kang, X. (2020). Molecular mechanism of vegetative growth advantage in alotriploid Populus. Int. J. Mol. Sci. 21, 441. doi: 10.3390/ijms21020441
Du, K., Xia, Y., Zhan, D., Xu, T., Lu, T., Yang, J., et al. (2022). Genome-wide identification of the Eucalyptus urophylla GATA gene family and its diverse roles in chlorophyll biosynthesis. Int. J. Mol. Sci. 23, 5251. doi: 10.3390/ijms23095251
Fitter, D. W., Martin, D. J., Copley, M. J., Scotland, R. W., and Langdale, J. A. (2002). GLK gene pairs regulate chloroplast development in diverse plant species. Plant J. 31, 713–727. doi: 10.1046/j.1365-313X.2002.01390.x
Fujiwara, T., and Beachy, R. N. (1994). Tissue-specific and temporal regulation of a β-conglycinin gene: roles of the RY repeat and other cis-acting elements. Plant Mol. Biol. 24, 261–272. doi: 10.1007/BF00020166
Gan, P., Liu, F., Li, R., Wang, S., and Luo, J. (2019). Chloroplasts-beyond energy capture and carbon fixation: tuning of photosynthesis in response to chilling stress. Int. J. Mol. Sci. 20, 5046. doi: 10.3390/ijms20205046
Haas, B. J., Salzberg, S. L., Zhu, W., Pertea, M., Allen, J. E., Orvis, J., et al. (2008). Automated eukaryotic gene structure annotation using EVidenceModeler and the program to assemble spliced alignments. Genome Biol. 9, R7–R21. doi: 10.1186/gb-2008-9-1-r7
Hii, S. Y., Ha, K. S., Ngui, M. L., Ak Penguang, S., Duju, A., Teng, X. Y., et al. (2017). Assessment of plantation-grown Eucalyptus pellita in Borneo, Malaysia for solid wood utilisation. Aust. For. 80, 26–33. doi: 10.1080/00049158.2016.1272526
Jackson, S., and Chen, Z. J. (2010). Genomic and expression plasticity of polyploidy. Curr. Opin. Plant Biol. 13, 153–159. doi: 10.1016/j.pbi.2009.11.004
Kohl, M., Wiese, S., and Warscheid, B. (2011). Cytoscape: software for visualization and analysis of biological networks. Methods Mol. Biol. 696, 291–303. doi: 10.1007/978-1-60761-987-1_18
Kumar, S., Stecher, G., Li, M., Knyaz, C., and Tamura, K. (2018). MEGA X: molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 35, 1547–1549. doi: 10.1093/molbev/msy096
Lai, E. C., Burks, C., and Posakony, J. W. (1998). The K box, a conserved 3′ UTR sequence motif, negatively regulates accumulation of enhancer of split complex transcripts. Development 125, 4077–4088. doi: 10.1242/dev.125.20.4077
Langdale, J. A., and Nelson, T. (1991). Spatial regulation of photosynthetic development in C4 plants. Trends Genet. 7, 191–196. doi: 10.1016/0168-9525(91)90435-S
Li, Y., Yang, J., Song, L., Qi, Q., Du, K., Han, Q., et al. (2019). Study of variation in the growth, photosynthesis, and content of secondary metabolites in Eucommia triploids. Trees 33, 817–826. doi: 10.1007/s00468-019-01818-5
Li, X., Zhao, L., Zhang, H., Liu, Q., Zhai, H., Zhao, N., et al. (2022). Genome-wide identification and characterization of CDPK family reveal their involvements in growth and development and abiotic stress in sweet potato and its two diploid relatives. Int. J. Mol. Sci. 23, 3088. doi: 10.3390/ijms23063088
Liang, Y., Xia, J., Jiang, Y., Bao, Y., Chen, H., Wang, D., et al. (2022). Genome-wide identification and analysis of bZIP gene family and resistance of TaABI5 (TabZIP96) under freezing stress in wheat (Triticum aestivum). Int. J. Mol. Sci. 23, 2351. doi: 10.3390/ijms23042351
Liao, T., Cheng, S., Zhu, X., Min, Y., and Kang, X. (2016). Effects of triploid status on growth, photosynthesis, and leaf area in Populus. Trees 30, 1137–1147. doi: 10.1007/s00468-016-1352-2
Lim, C. J., Lee, H. Y., Kim, W. B., Lee, B.-S., Kim, J., Ahmad, R., et al. (2012). Screening of tissue-specific genes and promoters in tomato by comparing genome wide expression profiles of Arabidopsis orthologues. Mol. Cells 34, 53–59. doi: 10.1007/s10059-012-0068-4
Lin, Z., Li, Q., Yin, Q., Wang, J., Zhang, B., Gan, S., et al. (2018). Identification of novel miRNAs and their target genes in Eucalyptus grandis. Tree Genet. Genomes 14, 1–9. doi: 10.1007/s11295-018-1273-x
Lin, Z., Long, J., Yin, Q., Wang, B., Li, H., Luo, J., et al. (2019). Identification of novel lncRNAs in Eucalyptus grandis. Ind. Crop. Prod. 129, 309–317. doi: 10.1016/j.indcrop.2018.12.016
Liu, F., Xu, Y., Han, G., Zhou, L., Ali, A., Zhu, S., et al. (2016). Molecular evolution and genetic variation of G2-Like transcription factor genes in maize. PLoS One 11:e0161763. doi: 10.1371/journal.pone.0161763
Lupi, A. C. D., Lira, B. S., Gramegna, G., Trench, B., Alves, F. R. R., Demarco, D., et al. (2019). Solanum lycopersicum Golden 2-Like transcription factor affects fruit quality in a light- and auxin-dependent manner. PLoS One 14:e0212224. doi: 10.1371/journal.pone.0212224
Menkens, A. E., Schindler, U., and Cashmore, A. R. (1995). The G-box: a ubiquitous regulatory DNA element in plants bound by the GBF family of bZIP proteins. Trends Biochem. Sci. 20, 506–510. doi: 10.1016/S0968-0004(00)89118-5
Mo, L., Chen, J., Lou, X., Xu, Q., Dong, R., Tong, Z., et al. (2020). Colchicine-induced polyploidy in Rhododendron fortunei Lindl. Plan. Theory 9, 424. doi: 10.3390/plants9040424
Nakamura, H., Muramatsu, M., Hakata, M., Ueno, O., Nagamura, Y., Hirochika, H., et al. (2009). Ectopic overexpression of the transcription factor OsGLK1 induces chloroplast development in non-green rice cells. Plant Cell Physiol. 50, 1933–1949. doi: 10.1093/pcp/pcp138
Nguyen, C. V., Vrebalov, J. T., Gapper, N. E., Zheng, Y., Zhong, S., Fei, Z., et al. (2014). Tomato Golden2-Like transcription factors reveal molecular gradients that function during fruit development and ripening. Plant Cell 26, 585–601. doi: 10.1105/tpc.113.118794
Nie, Y. M., Han, F. X., Ma, J. J., Chen, X., Song, Y. T., Niu, S. H., et al. (2022). Genome-wide TCP transcription factors analysis provides insight into their new functions in seasonal and diurnal growth rhythm in Pinus tabuliformis. BMC Plant Biol. 22, 167. doi: 10.1186/s12870-022-03554-4
Pappas Mde, C., Pappas, G. J. Jr., and Grattapaglia, D. (2015). Genome-wide discovery and validation of Eucalyptus small RNAs reveals variable patterns of conservation and diversity across species of Myrtaceae. BMC Genomics 16, 1113. doi: 10.1186/s12864-015-2322-6
Pérez, S., Renedo, C. J., Ortiz, A., Mañana, M., and Silió, D. (2006). Energy evaluation of the Eucalyptus globulus and the Eucalyptus nitens in the north of Spain (Cantabria). Thermochim. Acta 451, 57–64. doi: 10.1016/j.tca.2006.08.009
Powell, A. L. T., Nguyen, C. V., Hill, T., Cheng, K. L., Figueroa-Balderas, R., Aktas, H., et al. (2012). Uniform ripening encodes a Golden 2-Like transcription factor regulating tomato fruit chloroplast development. Science 336, 1711–1715. doi: 10.1126/science.1222218
Qin, M., Zhang, B., Gu, G., Yuan, J., Yang, X., Yang, J., et al. (2021). Genome-wide analysis of the G2-like transcription factor genes and their expression in different senescence stages of tobacco (Nicotiana tabacum L.). Front. Genet. 12:626352. doi: 10.3389/fgene.2021.626352
Riechmann, J. L., Heard, J., Martin, G., Reuber, L., Jiang, C.-Z., Keddie, J., et al. (2000). Arabidopsis transcription factors: genome-wide comparative analysis among eukaryotes. Science 290, 2105–2110. doi: 10.1126/science.290.5499.2105
Rossini, L., Cribb, L., Martin, D. J., and Langdale, J. A. (2001). The maize Golden2 gene defines a novel class of transcriptional regulators in plants. Plant Cell 13, 1231–1244. doi: 10.1105/tpc.13.5.1231
Roulin, A., Auer, P. L., Libault, M., Schlueter, J., Farmer, A., May, G., et al. (2013). The fate of duplicated genes in a polyploid plant genome. Plant J. 73, 143–153. doi: 10.1111/tpj.12026
Song, H., Wang, P., Lin, J.-Y., Zhao, C., Bi, Y., and Wang, X. (2016). Genome-wide identification and characterization of WRKY gene family in peanut. Front. Plant Sci. 7:534. doi: 10.3389/fpls.2016.00534
Thompson, J. D., Gibson, T. J., Plewniak, F., Jeanmougin, F., and Higgins, D. G. (1997). The CLUSTAL_X windows interface: flexible strategies for multiple sequence alignment aided by quality analysis tools. Nucleic Acids Res. 25, 4876–4882. doi: 10.1093/nar/25.24.4876
Unnikrishnan, B. V., and Shankaranarayana, G. D. (2020). Insights into microRNAs and their targets associated with lignin composition in Eucalyptus camaldulensis. Plant Gene 24:100248. doi: 10.1016/j.plgene.2020.100248
Valli, H., Prasanna, D., Rajput, V. S., Sridhar, W., Sakuntala, N. N. V., Harshavardhan, P., et al. (2022). Cis elements: added boost to the directed evolution of plant genes. J. Pure Appl. Microbiol. 16, 663–668. doi: 10.22207/jpam.16.1.68
Vilasboa, J., Da Costa, C. T., and Fett-Neto, A. G. (2019). Rooting of eucalypt cuttings as a problem-solving oriented model in plant biology. Prog. Biophys. Mol. Biol. 146, 85–97. doi: 10.1016/j.pbiomolbio.2018.12.007
Wang, P., Fouracre, J., Kelly, S., Karki, S., Gowik, U., Aubry, S., et al. (2013). Evolution of Golden 2-Like gene function in C3 and C4 plants. Planta 237, 481–495. doi: 10.1007/s00425-012-1754-3
Wang, Y., Liu, G. J., Yan, X. F., Wei, Z. G., and Xu, Z. R. (2011). MeJA-inducible expression of the heterologous JAZ2 promoter from Arabidopsis in Populus trichocarpa protoplasts. J. Plant Dis. Prot. 118, 69–74. doi: 10.1007/BF03356384
Wang, Y., Tang, H., Debarry, J. D., Tan, X., Li, J., Wang, X., et al. (2012). MCScanX: a toolkit for detection and evolutionary analysis of gene synteny and collinearity. Nucleic Acids Res. 40:e49. doi: 10.1093/nar/gkr1293
Wang, Z. Y., Zhao, S., Liu, J. F., Zhao, H. Y., Sun, X. Y., Wu, T. R., et al. (2022). Genome-wide identification of Tomato Golden 2-Like transcription factors and abiotic stress related members screening. BMC Plant Biol. 22, 82. doi: 10.1186/s12870-022-03460-9
Waters, M. T., Moylan, E. C., and Langdale, J. A. (2008). GLK transcription factors regulate chloroplast development in a cell-autonomous manner. Plant J. 56, 432–444. doi: 10.1111/j.1365-313X.2008.03616.x
Waters, M. T., Wang, P., Korkaric, M., Capper, R. G., Saunders, N. J., and Langdale, J. A. (2009). GLK transcription factors coordinate expression of the photosynthetic apparatus in Arabidopsis. Plant Cell 21, 1109–1128. doi: 10.1105/tpc.108.065250
Wu, S., Wu, M., Dong, Q., Jiang, H., Cai, R., and Xiang, Y. (2016). Genome-wide identification, classification and expression analysis of the PHD-finger protein family in Populus trichocarpa. Gene 575, 75–89. doi: 10.1016/j.gene.2015.08.042
Xiao, Y., You, S., Kong, W., Tang, Q., Bai, W., Cai, Y., et al. (2019). A GARP transcription factor anther dehiscence defected 1 (OsADD1) regulates rice anther dehiscence. Plant Mol. Biol. 101, 403–414. doi: 10.1007/s11103-019-00911-0
Yang, X., Guo, T., Li, J., Chen, Z., Guo, B., and An, X. (2021). Genome-wide analysis of the MYB-related transcription factor family and associated responses to abiotic stressors in Populus. Int. J. Biol. Macromol. 191, 359–376. doi: 10.1016/j.ijbiomac.2021.09.042
Yang, J., Wang, J., Liu, Z., Xiong, T., Lan, J., Han, Q., et al. (2018). Megaspore chromosome doubling in Eucalyptus urophylla S.T. Blake induced by colchicine treatment to produce triploids. Forests 9, 728. doi: 10.3390/f9110728
Zhang, J. (2003). Evolution by gene duplication: an update. Trends Ecol. Evol. 18, 292–298. doi: 10.1016/S0169-5347(03)00033-8
Zhang, J., Liu, Y., Xia, E. H., Yao, Q. Y., Liu, X. D., and Gao, L. Z. (2015). Autotetraploid rice methylome analysis reveals methylation variation of transposable elements and their effects on gene expression. Proc. Natl. Acad. Sci. U. S. A. 112, E7022–E7029. doi: 10.1073/pnas.1515170112
Keywords: chlorophyll synthesis, co-expression networks, EgrGLK, miRNA, polyploid, transcriptome analysis
Citation: Liu Z, Xiong T, Zhao Y, Qiu B, Chen H, Kang X and Yang J (2022) Genome-wide characterization and analysis of Golden 2-Like transcription factors related to leaf chlorophyll synthesis in diploid and triploid Eucalyptus urophylla. Front. Plant Sci. 13:952877. doi: 10.3389/fpls.2022.952877
Edited by:
Jen-Tsung Chen, National University of Kaohsiung, TaiwanReviewed by:
Phanikanth Jogam, Kakatiya University, IndiaCopyright © 2022 Liu, Xiong, Zhao, Qiu, Chen, Kang and Yang. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Jun Yang, eWFuZ19qdW5AYmpmdS5lZHUuY24=
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.