
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Plant Sci. , 14 June 2022
Sec. Plant Pathogen Interactions
Volume 13 - 2022 | https://doi.org/10.3389/fpls.2022.908308
This article is part of the Research Topic Global Occurrence of Pine Wilt Disease: Biological Interactions and Integrated Management View all 14 articles
 Cláudia S. L. Vicente1,2
Cláudia S. L. Vicente1,2 Miguel Soares3Jorge M. S. Faria2Margarida Espada1
Miguel Soares3Jorge M. S. Faria2Margarida Espada1 Manuel Mota4
Manuel Mota4 Filomena Nóbrega2Ana P. Ramos3
Filomena Nóbrega2Ana P. Ramos3 Maria L. Inácio2,5*
Maria L. Inácio2,5*Considered one of the most devastating plant–parasitic nematodes worldwide, Bursaphelenchus xylophilus (commonly known as pinewood nematode, PWN) is the causal agent of the pine wilt disease in the Eurasian coniferous forests. This migratory parasitic nematode is carried by an insect vector (Monochamus spp.) into the host tree (Pinus species), where it can feed on parenchymal cells and reproduce massively, resulting in the tree wilting. In declining trees, PWN populations are strongly dependent on fungal communities colonizing the host (predominantly ophiostomatoid fungi known to cause sapwood blue-staining, the blue-stain fungi), which not only influence their development and life cycle but also the number of individuals carried by the insect vector into a new host. Our main aim is to understand if PWN-associated mycobiota plays a key role in the development of PWD, in interaction with the PWN and the insect vector, and to what extent it can be targeted to disrupt the disease cycle. For this purpose, we characterized the fungal communities of Pinus pinaster trees infected and non-infected with PWN in three collection sites in Continental Portugal with different PWD temporal incidences. Our results showed that non-infected P. pinaster mycoflora is more diverse (in terms of abundance and fungal richness) than PWN-infected pine trees in the most recent PWD foci, as opposed to the fungal communities of long-term PWD history sites. Then, due to their ecological importance for PWN survival, representatives of the main ophiostomatoid fungi isolated (Ophiostoma, Leptographium, and Graphilbum) were characterized for their adaptative response to temperature, competition in-between taxa, and as food source for PWN. Under the conditions studied, Leptographium isolates showed promising results for PWN control. They could outcompete the other species, especially O. ips, and significantly reduce the development of PWN populations when compared to Botrytis cinerea (routinely used for PWN lab culturing), suggesting this to be a natural antagonist not only for the other blue-stain species but also for the PWN.
Pine wilt disease (PWD) is a devastating disease that affects mainly coniferous trees of the Pinus genus. The plant parasitic nematode Bursaphelenchus xylophilus (Steiner & Buhrer) Nickle, commonly known as the pinewood nematode (PWN), is responsible for the onset of PWD symptoms, namely, the characteristic needle yellowing and wilting as a result of shoot desiccation and chlorosis, that can lead to tree death in under 2 months (Futai, 2013). The PWN was first identified in North America, where it is believed to be an endemic. In its native range, the PWN only seldom causes host disease, generally in weakened native species or on exotic introduced pine species (Dwinell, 1997). There, the disease has a low expression because of the co-evolution of the PWN with native pines. In the beginning of the 20th century, in the Japanese islands, extensive ranges of pine forest began developing PWD symptoms, and the PWN was identified soon after (Mamiya, 1983). Since then, PWD has spread throughout the Asian continent causing strong ecological and economic impacts in Japan, China, Korea, and Taiwan (Vicente et al., 2012). In 1999, Portugal became its entry point into the European continent leading to the enforcement of strong containment strategies by national authorities and the EU and was set under quarantine (Mota et al., 1999). However, by 2008, the Portuguese continental area and Madeira Island (Fonseca et al., 2012) were considered affected and even more restringing measures were applied (Rodrigues et al., 2015). Several outbreaks have been reported in border Spanish forests (Abelleira et al., 2011). The PWN is now considered an A2 quarantine pest in the EU and a strong yearly investment is made in its containment and eradication for fear of a European pandemic in its susceptible pine forests (Robertson et al., 2011; Fonseca et al., 2012; de la Fuente and Saura, 2021).
The development of PWD and the intensity of its symptomatology are highly dependent on environmental and biological factors (Liu et al., 2021). Besides its causal agent, the PWN, other organisms are known to influence the development of PWD. Fungi are of great importance for the PWN. For dispersal, the PWN depends on an insect vector, the Monochamus genus, that carries and transmits the parasite to the new susceptible healthy host pines. Once inside the new host, the PWN enters the resin canals, attacks the epithelial cells, and causes extensive damage as it moves through the canal system and rapidly reproduces. During this reproductive phase on dead or decaying wood, the PWN depends necessarily on local fungal flora as a source of food for rapid multiplication (Futai, 2013). A weakened pine tree is more easily attacked by the fungi transmitted by bark beetles (Coleoptera: Scolytidae) that further increase the symptomatology of wilting and function as a necessary food source for the PWN. Thus, these fungi are essential for nematode survival and reproduction (Vicente et al., 2021). Generally called blue-stain fungi, referring to the damage they cause, namely blue or even black discoloration of the sapwood of trees, mostly on conifers (Harrington, 1993; Kim et al., 2003; Zhou et al., 2006; Six, 2012), these bark beetle vectored fungi are also known as “ophiostomatoid fungi” (Kirisits, 2007). Within this group, Ophiostomatales, comprising six genera, Ophiostoma sensu stricto (s. str.), Raffaelea s. str., Ceratocystiopsis, Fragospaeria, Graphilbum, and Leptographium senso lato (s. l.) are clearly separated from fungi of the Microascales order, namely Ceratocystis sp. (de Beer et al., 2013). Ceratocystis, Ceratocystiopsis, and Ophiostoma fungi and their related anamorphs could be distinguished based on the chemical composition of their cell walls. Besides chitin, cell walls of Ophiostoma also contain cellulose and rhamnose, which is quite unusual for ascomycete fungi. In contrast, the cell walls of Ceratocystis and Ceratocystiopsis spp. consist mainly of chitin. The cell walls of Ophiostoma fungi make them highly tolerant to cycloheximide that inhibits the protein synthesis of most eukaryotic organisms (Harrington, 1981), while species of Ceratocystis and Ceratocystiopsis are very sensitive to even low concentrations of this antibiotic (Kirisits et al., 2002; Plattner et al., 2009; Reid et al., 2010).
The hypothesis of PWN nutritional dependency in relation to some species of ophiostomatoid fungi inhabiting pine trees is supported by significant observations: (1) PWN propagates at higher rates in fungus-inoculated wood than in uninoculated wood (Maehara and Futai, 2000); and the number of PWN individuals transferred to a beetle is much higher in fungus-colonized wood than in fungal-free wood (Maehara and Futai, 1996). Maehara and Futai (1996, 1997) showed that O. minus-inoculated wood-blocks of P. densiflora presented a higher percentage of PWN transferred by beetles than when inoculated with Trichoderma spp. or in uninoculated blocks. Beetles emerging from pupal chambers having an intense colonization by blue-stain fungi vectored higher numbers of dauer juveniles (JIV), highlighting the relation between the PWN population and the fungal species present in the infected tree (Maehara and Futai, 2005). These results and further work (to Maehara et al., 2006; Maehara, 2008; Niu et al., 2012; Futai, 2013; Pimentel et al., 2021) undoubtedly suggest a mutualistic symbiosis between the nematode and certain fungi, possibly because of a long-term co-evolution. This effect is not surprising considering the mycetophagous status of Bursaphelenchus nematodes and might represent the trace of an evolutionary transition from this fungal-feeding behavior to the plant parasitic activity of B. xylophilus (Kanzaki et al., 2014).
Although there is strong evidence of fungal involvement in the expression of PWD, this subject is controversial, and to understand the complex dynamics of PWD it is crucial to identify the fungal community in declining trees, ascertaining which species have a role in PWN feeding, reproduction, and dispersal. The comprehensive scenario of the fungal community in the PWN life cycle, through a spatio-temporal analysis of the mycobiota associated with the nematode and the susceptible host tree, constitutes a pioneering approach in Portuguese maritime pine stands with a high prevalence of PWD. The present study characterizes the: (i) fungal communities in non-infected and PWN-infected P. pinaster trees from three contrasting PWD focus on Continental Portugal and (ii) adaptative responses of ophiostomatoid fungi (genus Ophiostoma, Leptographium and Graphilbum) inhabiting Portuguese pine stands to temperature, antagonism, and PWN interaction due to their well-recognized history with PWN.
Three surveys were conducted in mainland Portugal during the period of October 2019 in Seia (Guarda District, Central Portugal) in January 2020 in Companhia das Lezírias (Santarém, Central Portugal) and June 2020 in Tróia (Setúbal District, Coast of Portugal) (Table 1). The first location (Seia) is considered a transitional PWD focus, where the disease was identified in 2018–2019. The other two locations, Companhia das Lezírias and Tróia, are known as long-term PWD foci, nearby the first PWN detection site in Portugal (Mota et al., 1999). At each site, 10 PWD-symptomatic trees and 10 non-symptomatic trees were randomly selected according to the wilting/defoliation classification: 0, no symptoms; 1, first branch with yellow needles; 2, yellow tree canopy; 3, yellow to reddish tree canopy; and 4, brown canopy to dead tree (Cadahia et al., 1991). Wood disk were collected and stored in plastic bags for transportation to INIAV I.P. laboratories (Oeiras, Portugal). Concerning wood processing, cutting was conducted thoroughly with disinfection of the mechanical woodworking band saw with 70% (v/v) ethanol in-between cuttings and beginning with non-symptomatic trees. Approximately 100 g of wood pieces were used for nematode extraction by the modified tray method (Whitehead and Hemming, 1965). The rest of the material was stored at 4°C. After 48 h of incubation, samples were observed under a stereoscopic microscope (Olympus SZX12, Tokyo, Japan) to detect PWN. The following classes were considered to score nematode abundance in 100 g of wood: class 0, no nematodes recovered; class I, <50 nematodes; class II, between 51 and 200 nematodes; class III, between 201 and 1,000 nematodes; class IV, with 1,001 to 5,000 nematodes; and class V, more than 5,000 nematodes. The suspension of PWN was collected and stored in sterile 50 mL Falcon tubes. All material (wood pieces, and nematode suspensions) were stored at 4°C until further processing. The selection of P. pinaster trees for fungal isolation was based on: PWD symptomology and presence of PWN in higher proportion to other tree-inhabiting nematodes.
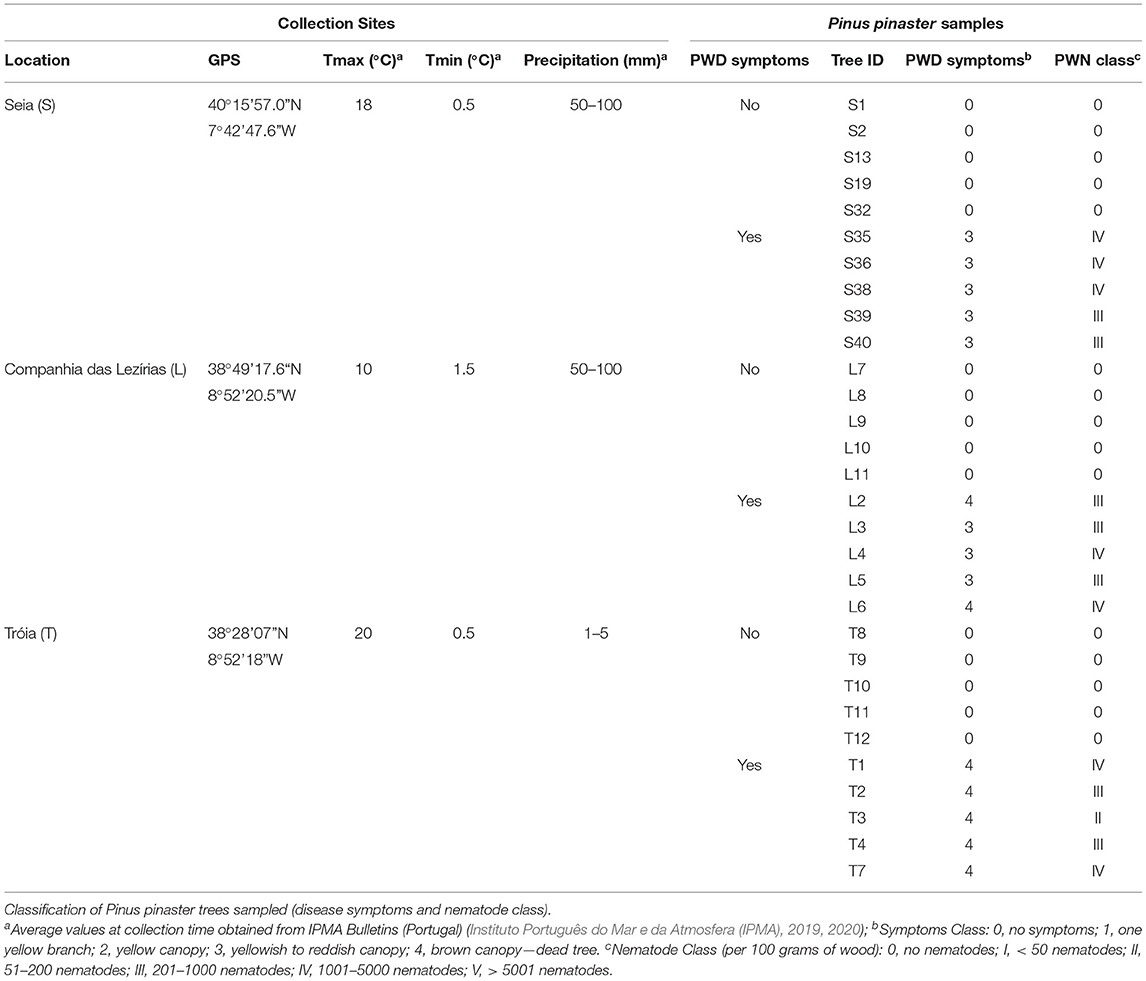
Table 1. Characterization of geographic and climate conditions for each collection site (Seia, Tróia, Companhia das Lezírias).
Under aseptic conditions, wood pieces from the selected trees were firstly cut into small fragments, surface sterilized with 70% ethanol (v/v) for 15 s, followed by serial washes with sterile distilled water, and dried in a sterilized filter paper. Following, surface-sterilized fragments were transferred to 2% (w/v) malt extract agar (MEA, Difco) supplemented with 200 mg/l of cycloheximide and 100 mg/l streptomycin. Plates were incubated at 25°C in the dark and monitored daily for fungal growth for at least 1 week. Hyphal tips of emerging colonies were transferred to fresh MEA plates. Pure cultures of the fungal isolates were grouped according to culture morphology. Representatives from each group were selected for further identification and characterization and deposited in the culture collection of INIAV institute (Micoteca da Estação Agronómica Nacional (MEAN).
For each fungal representative, fresh mycelium was grown in 2% MEA and incubated for 7 days at 25°C. Total DNA was extracted using the DNeasy Plant mini kit (Qiagen GmbH, Hilden, Germany) following the protocol provided by the manufacturer. Partial gene sequences were determined for the internal transcribed spacer region (ITS), β-tubulin (TUB), elongation factor 1-α (TEF), and calmodulin (CAL) using primers listed in Supplementary Table 1 (White et al., 1990; Gardes and Brunts, 1993; Glass and Donaldson, 1995; Carbone and Kohn, 1999; Duong et al., 2012; Marincowitz et al., 2015). For a final volume of 25 μl, the PCR reaction mixture consisted of 2.5 μl of 10x reaction Buffer, 0.5U Supreme NZYTaq II (NZYTECH, Portugal), 1.0 μl dNTP mix (10 mM), 1.25 μl of each primer (10 uM), 1.25 μl Mg2+ (50 mM), and 2 μl template DNA. The PCR conditions were: (i) initial denaturation at 95°C for 5 min; (ii) 34 cycles of denaturation at 94°C for 30 s, annealing at 48–52°C (depending on the marker gene), and extension at 72°C for 15–30 s; and (iii) final extension at 72°C for 10 min. PCR products were cleaned using EXO-SAP (Exonuclease I and FastAP™ Thermosensitive Alkaline Phosphatase, ThermoScientific, CA, USA) following the manufacturer's recommendations. The amplified PCR products were sequenced at Stabvida (Costa da Caparica, Portugal) using the same primers used for their amplification. All sequences from selected fungal isolates were deposited in NCBI GenBank.
Sequences were used as the query at NCBI GenBank for preliminary identification using BLAST tool (https://blast.ncbi.nlm.nih.gov/). Published sequences from closely related species were retrieved from the database (Supplementary Table 2). Different datasets of ITS, TUB, and CAL gene sequences were compiled based on the species complex or genera. Multiple sequence alignment was conducted using MAFFT v7.0 (Katoh and Standley, 2013) and trimmed with trimAl v.1.4.1 (Capella-Gutiérrez et al., 2009). Phylogenetic trees were reconstructed for each dataset using Maximum likelihood (ML) method, and the best fitting substitution model selected using the Akaike Information Criterion (AIC) estimated in the Model Testing plugin from CLC Main Workbench 21.0.5 software (QIAGEN, Aarthus A/S). Final adjustments were made in iTOL (https://itol.embl.de) (Letunic and Bork, 2021). Phylogenetic trees were supported by 1,000 bootstrap to assess node support and overall robustness of the tree topology.
Representatives of the ophiostomatoid isolates were selected for growth studies and antagonistic trials. For the growth studies, mycelium of each fungal representative was cultured on three 9-cm (Ø) Petri dishes containing 2% MEA (Difco®). Plates were inoculated with a 7 mm diameter mycelial plug taken from the margin of 5 days actively growing cultures and incubated in the dark at seven different temperatures: 5, 10, 20, 25, 30, 35, and 40°C. The radial growth of each plate was measured every 24 h, until the colonies reached 7 cm in diameter.
To assess the direct inhibition antagonism between ophiostomatoid-like isolates, different combinations were tested by dual culture assay (Royse and Ries, 1977; Almeida et al., 2020). A mycelium plug from pure cultures of the selected fungal isolates were placed on opposite sides of a 9-cm potato dextrose agar (PDA, Difco) plate, ~1 cm to the edge of the plate. Each combination was repeated three times and incubated at 25°C in dark conditions for 5 days. Measurements of the radial growth (from the edge of the plug until the end of the colony) were taken daily. The inhibition percentage was calculated by I (%) = [(R1-R2)/R1]x100, where R1 was the radial growth of the potential antagonist and R2 was the radial growth of the test organism.
The reference culture B. xylophilus BX013.003 (N 39°43'338”, W 9°01'557”) from INIAV Nematology collection (INIAV I.P., Oeiras) was used in this study. Nematodes were routinely cultured in the non-sporulating strain Botrytis cinerea grown on autoclaved barley seeds at 25°C. Prior to the experiments, the nematodes were extracted using the modified Baermann funnel technique (Whitehead and Hemming, 1965) and counted under an Olympus SZX12 (Tokyo, Japan) stereomicroscope for concentration adjustment.
The selected ophiostomatoid isolates were tested on feeding trials with PWN. Cultures of B. cinerea were used as a positive control for PWN population growth. For each isolate and control tested, six PDA plates were previously prepared and grown for 7 days at 25°C. Freshly extracted nematodes, mixed life-stages, were adjusted for a concentration of 1,000 nematodes per ml of distilled water, and 500 μl (ca. 500 nematodes) was inoculated into PDA plates containing the chosen isolates. Seven days after PWN inoculation, the nematodes were extracted by the modified Baermann method for 24 h and counted under an Olympus SZX12 stereomicroscope. The approximate total number of PWN was calculated for the total volume of the nematode suspension (final volume adjusted to 20 mL). Six replicates were conducted for each isolate. Each counting was repeated three times to correctly access the number of nematodes of each sample according to Aikawa and Kikuchi (2007).
The relative frequency of isolation (%) was estimated as the number of isolates of each order divided by the total number of species isolated and multiplied by 100 (Chang et al., 2017). To characterize the species diversity and richness of fungal communities from the different sites, the following indexes were calculated in PAST 4.02 software (Hammer et al., 2001): Simpson diversity index (S1-D), Shannon diversity index (H), dominance (D), evenness (E), and Sørensen index. Statistical analyses were performed using R in jamovi software 1.6.23 (R Core Team, 2020; The jamovi project, 2021). Kruskal–Wallis test (non-parametric one-way analysis of variance, ANOVA) was used to infer differences in the total number of PWN grown in the different ophiostomatoid isolates; the data was also analyzed using DSCF pairwise comparison (p < 0.05) for multiple mean comparisons.
Three surveys were conducted before the maturation feeding phase of the insect vector, when the insect vector is not yet flying (and carrying PWN) for new host trees. The three locations were sorted by long-term PWD foci (Tróia; and Companhia das Lezírias) and transitional or recent PWD focus (Seia), aiming the description of shifts in the fungal communities as the result of disease history in these locations. In each survey (Table 1), a total of 10 P. pinaster trees were considered for fungal isolation, namely, 5 non-symptomatic trees with symptoms and nematode class 0; and 5 PWD symptomatic trees with symptoms between class 3 and 4, and nematode concentration ranging from class III (201–1,000 nematodes per 100 g of wood) and class IV (1001-5000 nematodes per 100 g of wood). The presence of PWN was only detected and identified in symptomatic P. pinaster trees. Hereafter, symptomatic trees will be designated as PWN-infected, and non-symptomatic trees will be referred to as non-infected.
In terms of the total number of fungal isolations, 242 isolates were obtained from P. pinaster trees (45 from Companhia das Lezírias; 94 from Tróia; and 103 from Seia). Culture morphology and color supported by molecular marker sequencing were used for the taxonomic identification of each fungal isolate. Overall, the most abundant orders were Ophiostomatales (58% in Tróia, 60% in Companhia das Lezírias, and 90% in Seia) mostly detected in PWN-infected P. pinaster, followed by orders Pleosporales (2% in Companhia das Lezírias and 33% in Seia) and Eurotiales (64% in Companhia das Lezírias and 66% in Tróia) only in non-infected trees (Figure 1). The representative genera of Ophiostomatales were Ophiostoma with 34% (17 isolates in non-infected trees in Seia; and 65 isolates in all PWN-infected trees), Leptographium with 10% (24 isolates only in PWN-infected P. pinaster), Graphilbum with 2.1% (5 isolates in PWN-infected P. pinaster), and Sporothrix with 0.1% (one isolate detected in PWN-infected trees). In the orders Pleosporales and Eurotiales, the most predominant genera were Alternaria with 6.2% (15 isolates in non-infected trees) and Penicillium with 23.1% (56 isolates in non-infected trees). Only 2.5% of the fungal isolates were not identified as being classified as unidentified. Based on this general identification, and since the number of isolates was uneven between collection sites (Supplementary Figure 1), diversity indexes (Simpson 1-D and Shannon-H) varied widely among locations (Table 2). The fungal communities in the non-infected P. pinaster were more diverse (S1-D = 0.71; SH = 1.43) than in PWN-infected P. pinaster (S1-D = 0.46; SH = 0.87) in Seia. However, for the case of Companhia das Lezírias and Tróia, the results were different being PWN-infected more diverse in what concerns fungal composition than non-infected P. pinaster. For each site, similarity Sørensen index indicated that fungal communities from non-infected and PWN-infected trees were dissimilar, this observation being clearer in Tróia (S = 0.182) and Companhia das Lezírias (S = 0.2) than in Seia (S = 0.6). The PCoA analysis clearly separated PWN-infected fungal communities from non-infected P. pinaster fungal communities in Seia and Tróia (Supplementary Figure 2). Fungal communities from both PWN-infected and non-infected P. pinaster in Companhias das Lezirias clustered together, denoting no separation.
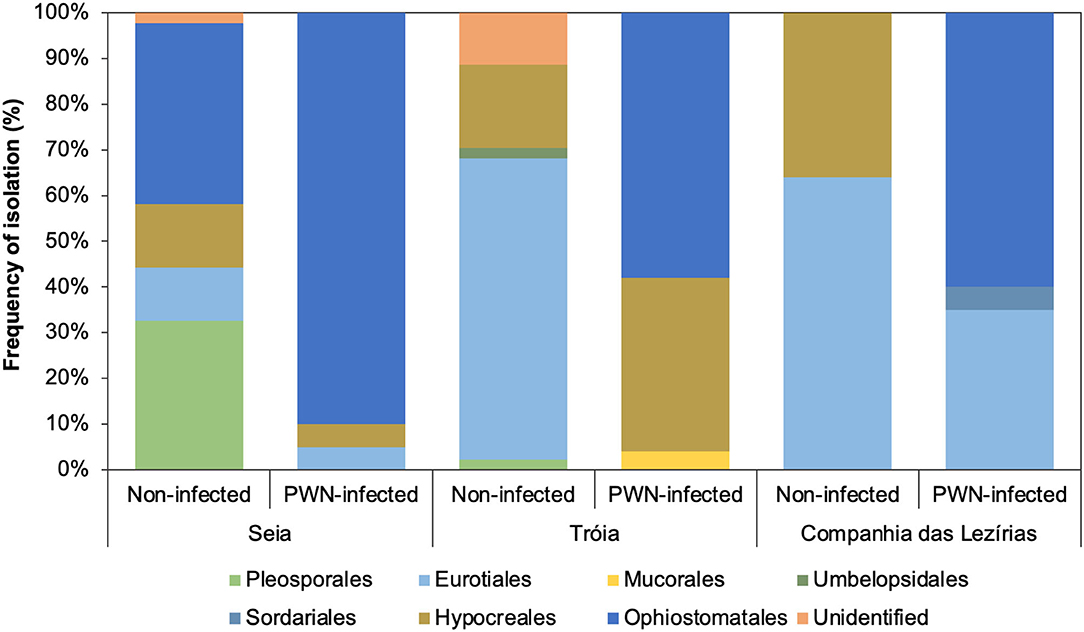
Figure 1. Frequency of isolation (%) of the main Ophiostomatales orders isolated in this study.
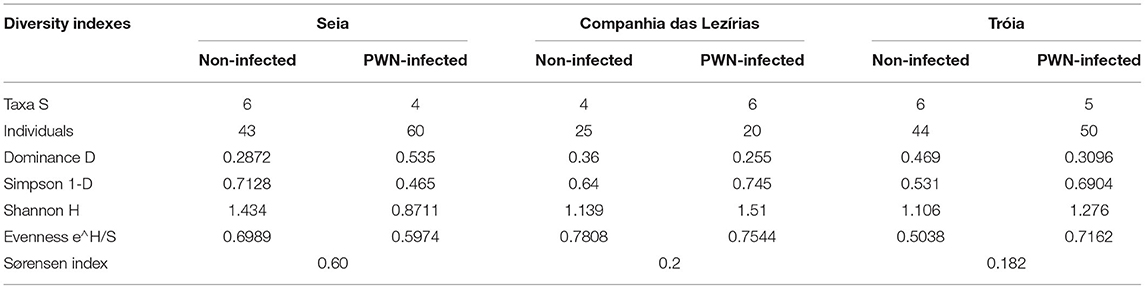
Table 2. Ecological diversity indexes (taxa = order) of fungal communities from non-infected and PWN-infected Pinus pinaster trees sampled.
A total of 366 DNA sequences (126 ITS; 84 TUB; 89 TEF; 67 CAL) were obtained to fully describe fungal communities in the non-infected and PWN-infected P. pinaster trees. All representative sequences for the four loci were BLAST against the NCBI GenBank database to retrieve the closest accessions (Supplementary Tables 2–5). From these sequences, only Ophiostomatales were considered for further phylogenetic analyses due to their close relation with B. xylophilus (Futai, 2013) (Table 3). Two datasets were considered based on preliminary BLAST results from ITS data: Ophiostoma s. l. and Leptographium s. l. as defined by de Beer et al. (2013). The amplification of ITS sequences for Leptographium s. l. species is often problematic (de Beer et al., 2013; Chang et al., 2017), and for some isolates were not obtained. In these cases, their taxonomic identification was supported by the other molecular markers. Additionally, the presence and absence of introns for TUB, TEF, and CAL sequences in both complexes, forced their separate analyses. Reference sequences for complexes within each dataset (Ophiostoma s. l. and Leptographium s. l.) were retrieved from GenBank (Supplementary Table 3).

Table 3. Fungal isolates representative of the main Ophiostomatales genus isolated in this study.
Within Leptographium s. l., four taxa were described: taxon 1, Graphilbum sp. 1-4; taxon 2, Leptographium terebrantis; taxon 3, Leptographium sp. 1-4; and taxon 4, Leptographium sosnaicola (Table 3). Multiple sequence alignment contained for each marker, respectively: 39 sequences with 665 bp, with gaps, for ITS; 31 sequences with 414 characters (with gaps) for TUB; 37 sequences with 528 bp (with gaps) for TEF; and 13 sequences with 597 bp (with gaps) for CAL (Figure 2, Supplementary Figures 3, 5). The best evolutionary substitution models for both trees were general time reversible (GTR) +G (rate variation) + T (topology variation). The Leptographium s. l. is well-represented by L. clavigera, L. lundbergii, L. penicillate, and the L. procerum complexes (Figure 2). The group taxon 1, Graphilbum sp., was considered the outgroup of all trees of Leptographium s. l. Within this group, Graphilbum sp. L 2.2 (TUB, TEF), Graphilbum sp. L4.2, L5.1, and S38.7 (TEF) clustered together with other Graphilbum species; however, their percentage of identity was lower than 98% (Supplementary Table 2), not allowing their species-level assignment. Taxon 2 and Taxon 3 belonged to L. clavigera complex (in both markers) and were grouped in-between L. wingfieldii and L. terebrantis with high support. In fact, isolates S38.3/S38.9 (isolated from the same tree) and S40.4b shared 100% similarity with L. terebrantis, while the other isolates (T3.1, T3.6, S40.11) were 100% identical in TUB but 99% in TEF (Supplementary Table 2). In CAL sequences, these isolates were identified as Grosmannia clavigera (100%) by the lack of Leptographium sequences in the database (Supplementary Table 2). Taxon 4 L. sosnaicola L2.6 formed a monophyletic group with L. sosnaicola MT210359 (TUB) and L. sosnaicola MW540787 (TEF), with good support, and clustering in between L. koreana and L. conjunctum within the L. lundbergii complex.
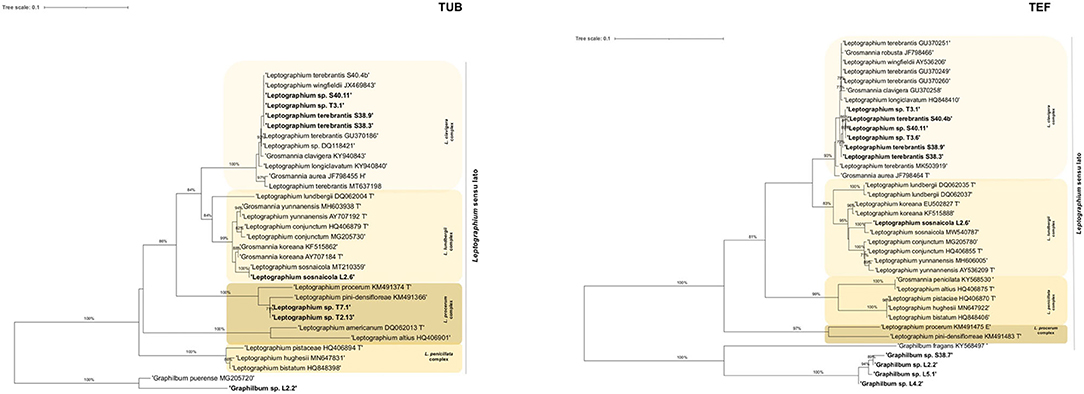
Figure 2. Maximum likelihood trees of Leptographium sensu lato generated by β-tubulin (TUB) and elongation factor 1-α (TEF) DNA sequence data. Sequences generated from this study are presented in bold (Table 2). Bootstrap values ≥70% are presented above the respective branch. Branch length is a measure of the number of substitutions per site (scale bar). T = ex-type isolates.
Within this lineage, only two taxa were described: taxon 5, Ophiostoma ips and taxon 6, Sporothrix sp.1 (Table 3). The alignment matrixes contained, respectively, 48 sequences (Table 2, Supplementary Table 3) with 648 characters, including gaps, for ITS marker; 43 sequences with 220 bp for TUB marker; 40 sequences with 379 bp for TEF; and 29 sequences with 339 bp for CAL (Figure 3, Supplementary Figures 4, 5). The best evolutionary substitution models for ITS and TEF phylograms were, respectively, Hasegawa-Kishino-Yano (HKY) +G+T and GTR+G+T. The well-defined phylogenetic complexes with Ophiostoma s. str. (O. ips complex, O. piceae complex, O. ulmi complex, and O. clavatum complex) were maintained, when possible, for all phylogenetic trees of Ophiostoma s. l. Fungal isolates from all locations and tree status, previously identified as O. ips, clustered together with good support with the type strain O. ips AY546704 and O. ips KU319013 (described in P. sylvestris in Poland) within the O. ips complex (Figure 3). Similar topologies were obtained in ML phylogenetic trees with the other molecular markers (BT, TEF and CAL; Figure 3, Supplementary Figures 4, 5), which sustained the identification of O. ips fungal isolates. Belonging to Ophiostoma s. l., the isolate L5.3 was identified as Sporothrix sp. 1 (99% similarity with Sporothrix sp. MW540762.1 with ITS; 100% S. pseudoabietina MH583598 with TUB; and 99.39% Sporothrix sp. KY568667 with TEF; Supplementary Table 2). In Figure 3, this isolate clustered with good support with S. schenckii NR_147566, being considered as the outgroup of this phylogenetic tree. Its position is also corroborated in Supplementary Figures 4, 5.
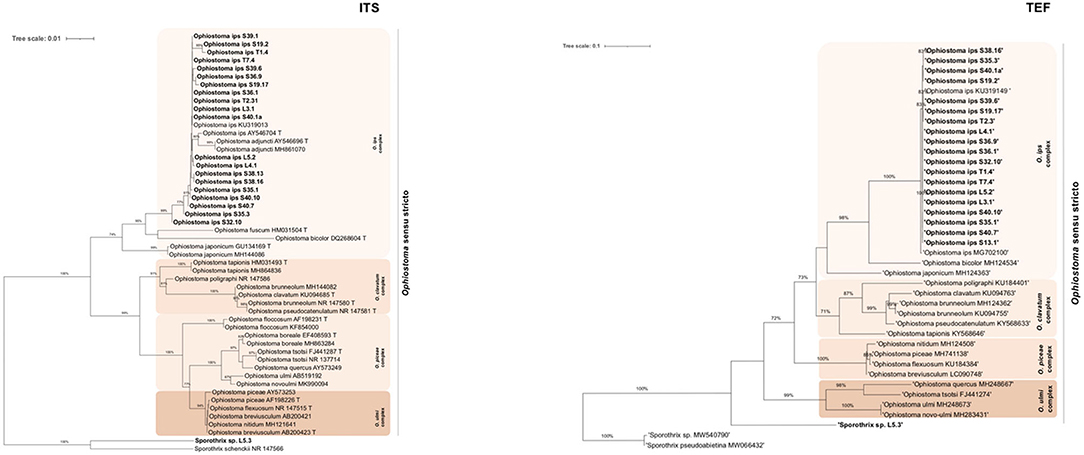
Figure 3. Maximum likelihood trees of Ophiostoma sensu stricto generated by internal-transcribed spacer (ITS2) and elongation factor 1-α (TEF) DNA sequence data. Sequences generated from this study are presented in Bold (Table 2). Bootstrap values ≥70% are presented above the respective branch. Branch length is a measure of the number of substitutions per site (scale bar). T = ex-type isolates.
To characterize the adaptative response (in terms of growth and antagonistic performance and PWN food source) of the main Ophiostomatales taxon (genus Ophiostoma, Leptographium, and Graphilbum) isolated from PWN-infected P. pinaster, ten ophiostomatoid fungal isolates (S36.1; S39.1; S38.16; S40.4b; S40.10; T2.13; T3.1; T3.6; L4.2; and L5.1) were selected as representatives. Only one isolate of Sporothrix was obtained and therefore was not included in these trials. Since the collection sites presented different geographic and climate conditions (in terms of average temperature and precipitation; Table 1), the selected fungal isolates were tested in a wide amplitude of temperatures between 5 and 40°C during 15 days of incubation (Figure 4). Overall, almost all fungal isolates could grow between 20 and 35°C; however with different optimal temperatures. All taxa failed to grow at 40°C. For Ophiostoma group (Figure 4A), the optimal growth temperature varied between 20°C (for S40.10) and 30°C (for S36.1, S39.1, S38.16). At lower temperatures (5–10°C), only S40.10 exhibited discrete growth but failed to grow at 35°C. For Leptographium group (Figure 4B), the optimal growth temperature ranged between 25°C (S40.4b, T3.1, and T3.6) and 30°C (T2.13). For Graphilbum sp. L4.2 and L5.1, the optimal growth temperature was 35°C (Figure 4C), yet these isolates were unable to grow at 5 or 10°C.
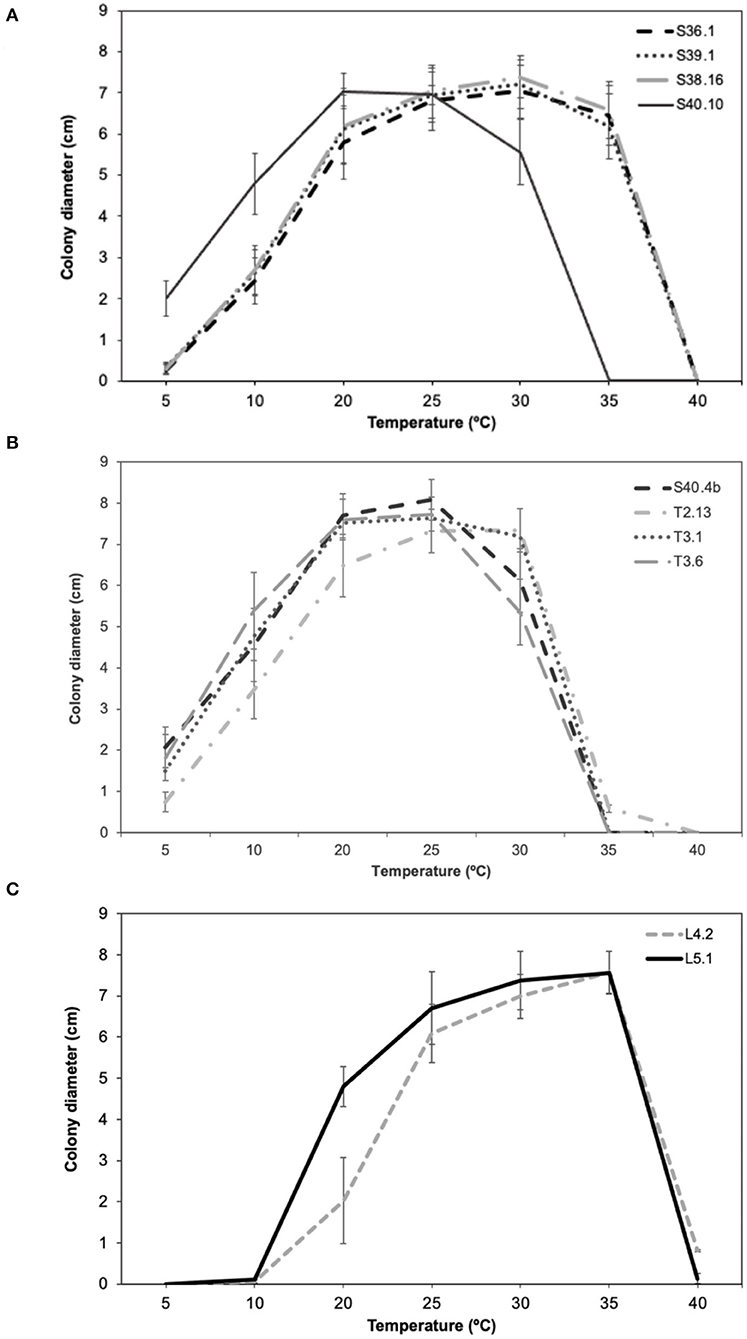
Figure 4. Comparison of mean growth per day (± standard error) of (A) Ophiostoma ips (S36.1, S39.1, S38.16, S40.10), (B) Leptographium sp. (S40.4b, T2.13, T3.1, T3.6), and (C) Graphilbum sp. (L4.2, L5.1).
For antagonistic performance of ophiostomatoid isolates, different combinations between representatives of the main taxa were tested considering a potential antagonist (R1) and a test organism (R2) (Supplementary Figure 6). Overall, the fungal isolate Leptographium sp. T3.6 inhibited the growth of the other 3 isolates [O. ips S36.1; Graphilbum sp. L4.2; L. terebrantis S40.4b (Figure 5)]. The percentage of inhibition of Leptographium sp. T3.6 and Ophiostoma ips S36.1 was 75%, while for Graphilbum sp. L4.2 and L. terebrantis S40.4b, they were 44 and 46%, respectively. No differences were seen between Leptographium sp. T3.6 and L. terebrantis S40.4b or Graphilbum sp. L4.2. The dual culture of Graphilbum sp. L4.2 x O. ips S36.1 and O. ips S36.1 x L. terebrantis S40.4b recorded the lowest inhibition, respectively, 33 and 15%.
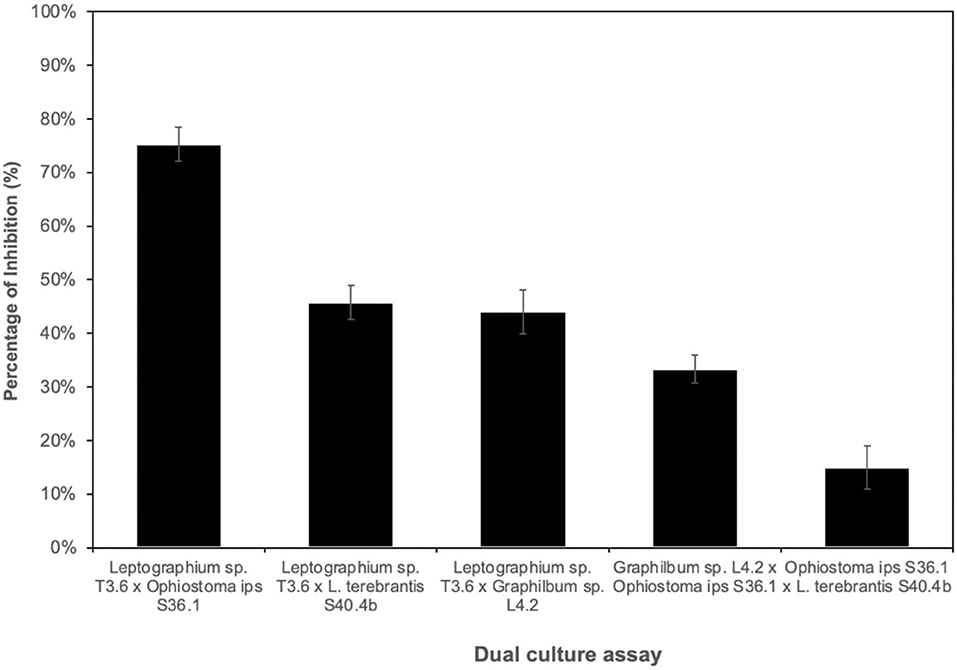
Figure 5. Percentage of inhibition of the potential antagonist x tested organism (grown at 25°C for 5 days).
Not all Ophiostomatales could support PWN growth. Except for O. ips S38.16 and S39.1, all fungal isolates tested were unsuitable for PWN as the food source (p < 0.001), when comparing with B. cinerea (total number of PWN after 7 days was 11,733 ± 546) (Figure 6). Within the group of O. ips, isolate S40.10 recorded a population increase of 27.4% when compared with the control, while S36.1 and S39.1 were, respectively, 54.8 and 80%. Ophiostoma ips S38.16 showed a close result with B. cinerea, with 94.6% of PWN growth (p > 0.001). For the Leptographium group, PWN growth was relatively low (<13%). In fact, PWN could not grow completely in S40.4b. The total number of PWN recovered from Leptographium sp. T2.13 and T3.1 were, respectively, 649 ± 160 and 636 ± 251 nematodes (~5.5% in comparison with B. cinerea). The two isolates of Graphilbum tested, L4.2 and L5.1, were also not suitable for PWN, with 12 and 4.5%, respectively, of PWN growth in comparison with the control fungus.
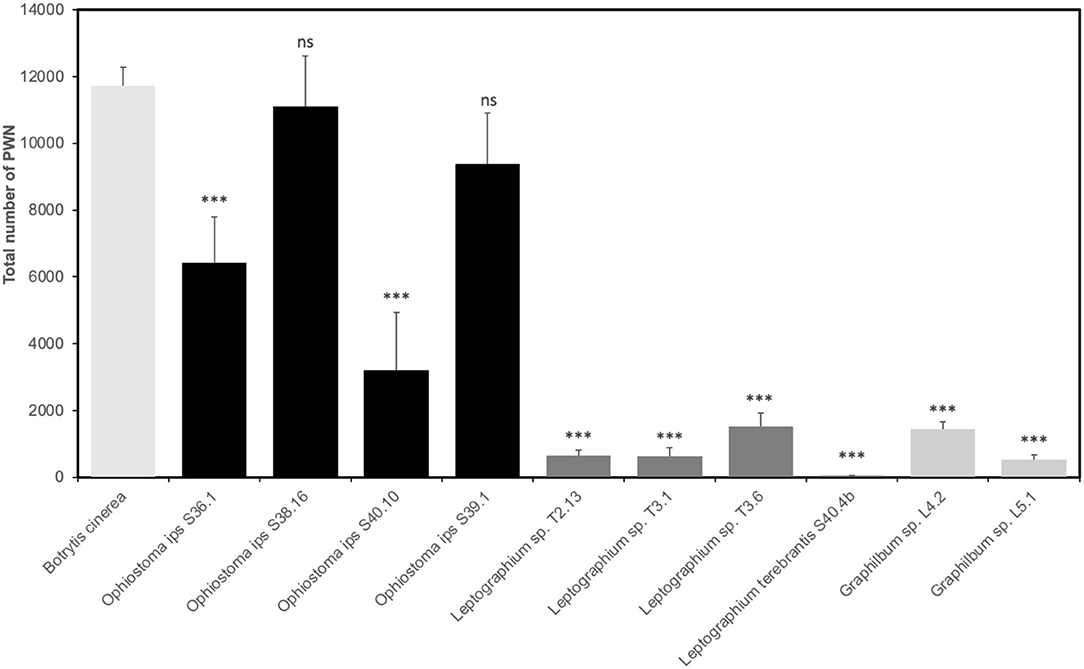
Figure 6. Reproduction of Bursaphelenchus xylophilus population in each ophiostomatoid fungi and Botrytis cinerea (control) (mean total no. nematodes ± standard error). Asterisks (***) indicate statistical differences at 99% of confidence in relation to B. cinerea.
Fungal communities seem to dominate different ecological niches in forest tree habitats due to their easy adaptation to wood colonization (de Boer et al., 2005). In this sense, these communities are more likely to undergo more severe shifts in abundance and composition due to biotic and abiotic stresses (Baldrian, 2017). The study of the fungal communities in the PWD complex, and their intermediates, are still underrated (Zhao et al., 2014; Nascimento et al., 2015) in spite of their strong influence on PWN populations (Vicente et al., 2021). In this study, we present a detailed characterization of fungal communities from non-infected and PWN-infected P. pinaster trees from three contrasting PWD foci (long-term PWN presence, Companhia das Lezírias and Tróia, and transitional/recent PWN detection, Seia). This is the first study focused exclusively on the fungal diversity in PWD affected areas, in Europe. In addition, we also characterized the adaptative response of a specific group of Ophiostomatales fungi (genus Ophiostoma, Leptographium, Graphilbum) due to their well-recognized history with PWN (Vicente et al., 2021), as means to uncover their resilience in the disease complex.
The surveys were conducted during early winter 2019 and spring 2020 before the maturation feeding of the insect-vector, which could justify the high PWN numbers extracted from the sampled wood. Despite the discrepancy in the total number of fungal isolates between sampling locations, overall, we observed clear differences in the fungal communities between collection sites, being long-term PWD loci (Companhia das Lezírias and Tróia) closest between themselves than each with the most recent PWD area (Seia). Several factors may influence this result, e.g., their PWD history or biogeographical conditions of the locations, which are completely distinct in terms of temperature/precipitation/altitude, as well as seasonality. In Seia, the fungal communities of non-infected and PWN-infected P. pinaster showed substantial differences in terms of composition and species richness, the non-infected pine trees being more biodiverse than PWN-infected trees. This observation suggests that the presence of the nematode could explain the community shifts which is corroborated by recent works with other Pinus species (Liu et al., 2021; Zhang et al., 2021). Still, this result is not evident for fungal communities in Companhia das Lezírias or Tróia, according to the diversity indexes. The fact that these locations have a long-term history of PWN presence could be an indication of communities' stability in the presence of the nematode invader. Kuroda and Ito (1992) also reported similar species in healthy and wilted P. thunbergii, which varied vaguely among seasons. Zhao et al. (2013) also suggested a link between disease history and microbial communities' diversity; however only infected trees were used in this study. In our work, most of the taxa observed were already reported for other Pinus species, as well as PWN and insect-vector Monochamus sp. (Hyun et al., 2007; Zhao et al., 2013, 2018). However, with the exception of two P. pinaster trees from Seia, only PWN-infected P. pinaster trees were colonized by Ophiostomatales, the causal agents of blue-stain (Six, 2012) commonly present in pine trees infected by the PWN (Futai, 2013). The most dominant ophiostomatoid species was O. ips. This species was firstly reported in association with the infection of bark beetles in south-eastern North America (Rumbold, 1931), and nowadays is distributed worldwide. In PWD complex, O. ips was described as associated with PWN-infected P. thunbergii, PWN, and M. alternatus in Korea (Hyun et al., 2007) in P. massoniana and P. thunbergii in China (Zhao et al., 2013, 2018; Wang et al., 2018), and M. galloprovincialis and P. pinaster in Portugal (Inácio et al., 2015; Trindade, 2019). As suggested by Wang et al. (2018), the dominance of O. ips can have either an indigenous origin or be an effective adaptation to local pine forests. In fact, the presence of O. ips in non-infected trees in Seia could be explained by natural infection with various species of bark beetles as is common in European pine forests (Chang et al., 2017; Jankowiak et al., 2021). The other ophiostomatoid isolates were Leptographium sp., Graphilbum sp., and Sporothrix sp., which were also described in other studies (Zhao et al., 2013, 2018; Wang et al., 2018). For the first time, we report Leptographium species associated with PWN-infected P. pinaster trees in Portugal, namely L. sosnaicola and L. terebrantis. Leptographium sosnaicola was described in logs of P. sylvestris in Poland and probably vectored by Ips sexdentatus (Jankowiak et al., 2021). The authors showed that L. sosnaicola formed a monophyletic line within other members of Leptographium, and its morphology did not resemble any known Leptographium species. Our results also show L. sosnaicola L2.6 within L. lundburgii complex and totally separated from the other sister species, corroborating the previous observations (Jankowiak et al., 2021). The other Leptographium species identified L. terebrantis, firstly described as associated with the black turpentine beetle Dendroctonus terebrans (Barras and Perry, 1971), was reported as pathogen of woody roots and recently related to crown symptoms and tree mortality in P. taeda (Mensah et al., 2021). Hausner et al. (2005) showed that ex-type culture of L. terebrantis is very similar morphologically to L. wingfieldii, which explains their clustering and non-resolved placement in the phylogenetic trees of this study. Although only a few isolates were recovered from PWN-infected P. pinaster, Graphilbum and Sporothrix genus are common in pine trees infected with the nematode. Zhao et al. (2013) showed that Sporothrix sp. 1 could sustain PWN growth with female-biased sex ratio offspring and suggested that the prevalence of this species was correlated with higher tree mortality that possibly influenced the nematode and insect-vector populations in the area studied. This ophiostomatoid fungi was also isolated in South Korea (Hyun et al., 2007) and other regions of China (Wang et al., 2018). As for other Leptographium isolates in this study, neither morphological characterization nor molecular identification was resolved enough to assign it into species level, which will be explored in future studies for the description of new species.
The presence of the PWN induces a disturbance in microbial (bacteria and fungi) community stability, which has been already analyzed using metagenomics approaches (Alves et al., 2016; Liu et al., 2021). Ecologically, this is often related with community insensitivity (resistance) or the rate at which community returns to a pre-disturbance condition, intrinsically related to their functional role in the ecosystem (Shade et al., 2012). In this study, the isolates of Ophiostoma, Leptographium, and Graphilbum groups showed differences in temperature adaptation, which may reflect the environmental conditions of their collection sites. The O. ips group grew in lower temperatures such as 5–10°C, which is characteristic of the Northern areas. Leptographium sp. isolates and Graphilbum sp. isolates showed a preference for higher temperatures, with an optimal growth at 25 and 35°C, respectively. Zhao et al. (2018) surveyed distinct regions of China forests and detected O. ips mainly in the Southern sites, with higher annual temperatures (16°C) and rainfall (1,020–1,408 mm). Cobian et al. (2019) demonstrated that plant-microbe specificity varies with different abiotic parameters such as temperature or elevation. They also showed a strong interaction between host and fungal endophyte, and that the environment plays a secondary role in fungal communities, mostly by modulating host specialization (Cobian et al., 2019). Overall, it seems that associations between ophiostomatoid fungi and the pine species in the context of PWD may be variable, suggesting a co-evolution history, however further studies are needed to address this issue.
Competitive interactions among different fungal symbionts of bark-beetles (such as O. minus, C. ranaculosis, and Entomocorticium sp.) have been reported in artificial and natural conditions (Klepzig and Hofstetter, 2011). Klepzig (1998) showed that on an artificial medium the biological control fungus O. piliferum outcompete all three fungi (O. minus, C. ranaculosis, and Entomocorticium sp.); however, in inoculations of natural substrate, O. piliferum compete successfully with the two mutualistic fungi C. ranaculosis and Entomocorticium sp., but not with O. minus. Here, we evaluated pairwise competition between the three main taxa and found that Leptographium sp. could outcompete neither with Ophiostoma nor with Graphilbum, suggesting potential antagonism between Ophiostomatales. Considering our sampled trees, this result could also explain why only few O. ips were collected in the trees mainly dominated with Leptographium. So far, we could not find any other study suggesting this kind of ecological interactions between ophiostomatoid fungi in the context of PWD.
Fukushige (1991a,b) showed that B. xylophilus propagation on fungus could differ in agar media and pine wood. The colonization of O. minus in pine-branch segments of P. densiflora resulted in a quick increase of PWN population (Maehara and Futai, 2000). Zhao et al. (2018) showed that L. pini-densiflorae could promote the highest population density, followed by Sporothrix sp. 1, while O. ips or O. minus resulted in significantly lower PWN populations. Pimentel et al. (2021) evaluated nematode propagation in 10 different species of Ascomycota fungi, including different Ophiostomales species (O. ips, O. piliferum, O. minus) and Leptographium (L. procerum, L. terebrantis). After 20 days of incubation, the nematode propagation in these fungal isolates was unexcepted, since all were unsuitable for PWN growth (Pimentel et al., 2021). In this sense, not all ophiostomatoid fungi have been considered suitable for PWN growth in declining trees (Zhao et al., 2014). In this study, the highest PWN multiplication in comparison with their B. cinerea culture fungi was obtained in the O. ips group, while Leptographium and Graphilbum group resulted in significantly lower PWN numbers in the conditions tested. Intraspecific variability within the O. ips group was also seen. With the exception of O. ips S40.10, all the other isolates had a positive impact on the PWN population, which seems to be in line with other studies (Maehara and Futai, 1997; Maehara, 2008). Indeed, this pattern seemed to be sustained by the initial quantification of PWN-infected trees. In Seia site, where O. ips was the most prevalent ophiostomatoid fungi (almost 70% of community composition, 3 out of the 5 pine trees presented a population of PWN between 1,001 and 5,000 nematodes per 100 g of wood (class IV). The Tróia site evidenced low to medium levels of PWN infection. In the specific case of one tree (T3), classified as class II (51-200 nematodes per 100 g of wood), the low concentration of PWN could be explained by the mycoflora present, given that all isolated fungi belonged to the Leptographium genus, which could not support PWN multiplication in the feeding trials. As for the two Graphilbum sp. isolates, the defoliation class of the trees sampled were class III and IV, not supporting the previous observations. Still, this seemingly relation between the host defoliation class and the impact of their isolated fungi in PWN multiplication is worth pursuing, especially confirming their effect in the native host P. pinaster. The present research constitutes a milestone in the study of the functional role of fungal communities inhabiting wilted trees, their ecological interactions, and their suitability for PWN growth, as well as their impact on PWD expression. This knowledge can lead to the identification of key points in PWN life cycle that can be targeted to disrupt the disease cycle using more natural alternatives.
The datasets presented in this study can be found in online repositories. The names of the repository/repositories and accession number(s) can be found in the article/Supplementary Material.
CV and MI: conceptualization. CV, MS, AR, and ME: research and data analysis. CV and JF: writing—original draft preparation. ME, AR, MM, FN, and MI: writing—review and editing. AR, MM, and MI: resources. All authors contributed to the article and approved the submitted version.
This research was funded by the Fundação para a Ciência e Tecnologia (FCT) and Fundo Europeu de Desenvolvimento Regional (FEDER) within the Programa Operacional Regional de Lisboa and Programa Operacional Regional do Alentejo through the national project LISBOA-01-0145-FEDER028724 - PTDC/ASP-PLA/28724/2017 (ALT20-03-0145-FEDER-028724) PineENEMY, Exploring the NEmatode-MYcobiota interactions in Pine Wilt Disease, through the R&D Unit, UIB/04551/2020 (GREEN-IT - Bioresources for Sustainability), and the CEECIND/00040/2018 (to CV).
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
The authors would like to thank the team of PineENEMY project (Laboratory of Nematology, Mycology and Entomology of INIAV). A special thanks to Helena Bragança (Mycology Lab) for supporting this work, to Luis Bonifácio (Entomology Lab, INIAV) for his dedication and expertise given in the selection of the collection sites.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fpls.2022.908308/full#supplementary-material
Abelleira, A., Picoaga, A., Mansilla, J. P., and Aguin, O. (2011). Detection of Bursaphelenchus xylophilus, causal agent of pine wilt disease on Pinus pinaster in northwestern Spain. Plant Dis. 95, 776. doi: 10.1094/PDIS-12-10-0902
Aikawa, T., and Kikuchi, T. K. (2007). Estimation of virulence of Bursaphelenchus xylophilus (Nematoda: Aphelenchoididae) based on its reproductive ability. Nematology 9, 371–377. doi: 10.1163/156854107781352007
Almeida, A. B., Concas, J., Campos, M. D., Materatski, P., Varanda, C., Patanita, M., et al. (2020). Endophytic fungi as potential biological control agents against grapevine trunk diseases in Alentejo region. Biology 9,1–23. doi: 10.3390/biology9120420
Alves, M., Pereira, A., Matos, P., Henriques, J., Vicente, C., Aikawa, T., et al. (2016). Bacterial community associated to the pine wilt disease insect vectors Monochamus galloprovincialis and Monochamus alternatus. Sci. Rep. 6, 23908. doi: 10.1038/srep23908
Baldrian, P. (2017). Forest microbiome: diversity, complexity, and dynamics. FEMS Microbiol. Rev. 41, 109–130. doi: 10.1093/femsre/fuw040
Barras, S. J., and Perry, T. (1971). Leptographium terebrantis sp. nov. associated with Dendroctonus terebrans in loblolly pine. Mycopathol. Mycol. Appl. 43, 1–10 doi: 10.1007/BF02051496
Cadahia, D., Cobos, J. M., Soria, S., Clauser, F., Gellini, R., Grossoni, P., et al. (1991). “Observation of damaged to Mediterranean forest species. ministry of agriculture, fisheries and food,” in Commission of the European Communities (Madrid), 96.
Capella-Gutiérrez, S., Silla-Martínez, J. M., and Gabaldón, T. (2009). trimAl: a tool for automated alignment trimming in large-scale phylogenetic analyses. Bioinformatics 25, 1972–1973. doi: 10.1093/bioinformatics/btp348
Carbone, I., and Kohn, L. M. (1999). A method for designing primer sets for speciation studies in filamentous ascomycetes. Mycologia 91, 553–556. doi: 10.1080/00275514.1999.12061051
Chang, R., Duong, T. A., Taerum, S. J., Wingfield, M. J., Zhou, X., and de Beer, Z. W. (2017). Ophiostomatoid fungi associated with conifer-infesting beetles and their phoretic mites in Yunnan, China. MycoKeys 28, 19–64. doi: 10.3897/mycokeys.28.21758
Cobian, G., CP, E., and Amend, A. (2019). Plant-microbe specificity varies as a function of elevation. ISME J. 13, 2778–2788. doi: 10.1038/s41396-019-0470-4
de Beer, Z. W., Seifert, K. A., and Wingfield, M. J. (2013). A nomenclator for ophiostomatoid genera and species in the Ophiostomatales and Microascales. Biodiversity Series 12, 245–322.
de Boer, W., Folman, L. B., Summerbell, R. C., and Boddy, L. (2005). Living in a fungal world: impact of fungi on soil bacterial niche development. FEMS Microbiol. Rev. 29, 795–811. doi: 10.1016/j.femsre.2004.11.005
de la Fuente, B., and Saura, S. (2021). Long-term projections of the natural expansion of the pine wood nematode in the Iberian Peninsula. Forests 12, 849. doi: 10.3390/f12070849
Duong, T. A., de Beer, Z. W., Wingfield, B. D., and Wingfield, M. J. (2012). Phylogeny and taxonomy of species in the Grosmannia serpens complex. Mycologia 104, 715–732. doi: 10.3852/11-109
Dwinell, L. D. (1997). The Pinewood Nematode: regulation and mitigation. Annu. Rev. Phytopathol. 35, 153–166. doi: 10.1146/annurev.phyto.35.1.153
Fonseca, L., Cardoso, J. M. S., Lopes, A., Pestana, M., Abreu, F., Nunes, N., et al. (2012). The pinewood nematode, Bursaphelenchus xylophilus, in Madeira Island. Helminthologia 49, 96–103. doi: 10.2478/s11687-012-0020-3
Fukushige (1991a). Propagation of Bursaphelenchus xylophilus (Nematoda: Aphelenchoididae) on fungi growing in pine-shoot segments. Appl. Entomol. Zool. 371–376. doi: 10.1303/aez.26.371
Fukushige (1991b). Effects of fungi coexisting with Ceratocystis sp. on propagation of Bursaphelenchus xylophilus (Nematoda: Aphelenchoididae). Appl. Entomol. Zool. 377–380. doi: 10.1303/aez.26.377
Futai, K. (2013). Pine wood nematode, Bursaphelenchus xylophilus. Ann. Rev. Phytopathol. 51, 61–83. doi: 10.1146/annurev-phyto-081211-172910
Gardes, M., and Brunts, T. D. (1993). ITS primers with enhanced specificity for basidiomycetes - application to the identification of mycorrhizae and rusts. Mol. Ecol. 2, 113–118. doi: 10.1111/j.1365-294X.1993.tb00005.x
Glass, N. L., and Donaldson, G. C. (1995). Development of primer sets designed for use with the PCR to amplify conserved genes from filamentous ascomycetes. Appl. Environ. Microbiol. 61, 1323–1330. doi: 10.1128/aem.61.4.1323-1330.1995
Hammer,Ø, Harper, D. A. T., and Ryan, P. D. (2001). PAST: paleontological statistics software package for education and data analysis. Palaeontol. Electron. 4, 1–9.
Harrington, T. (1993). “Diseases of conifers caused by species of Ophiostoma and Leptographium,” in Ceratocystis and Ophiostoma: Taxonomy, ecology and pathogenicity, eds M. J. Wingfield, K. A. Siefert, and J. F. Webber (The American Phytopathological Society Press), 161–172.
Harrington, T. C. (1981). Cycloheximide sensitivity as a taxonomic character in Ceratocystis. Mycologia 73, 1123–1129. doi: 10.1080/00275514.1981.12021447
Hausner, G., Iranpour, M., Kim, J., Breuil, C., Davis, C., Gibb, E. A., et al. (2005). Fungi vectored by the introduced bark beetle Tomicus piniperda in Ontario, Canada, and comments on the taxonomy of Leptographium lundbergii, Leptographium terebrantis, Leptographium truncatum, and Leptographium wingfieldii. Canad. J. Bot. 83, 1222–1237. doi: 10.1139/b05-095
Hyun, M., Kim, J., Suh, D., Lee, S., and Kim, S. (2007). Fungi isolated from pine wood nematode, its vector Japanese pine sawyer, and the nematode-infected Japanese black pine wood in Korea. Mycobiology 35, 159. doi: 10.4489/MYCO.2007.35.3.159
Inácio, M. L., Nóbrega, F., Trindade, J., Bonifácio, L., Naves, P., Sousa, E., et al. (2015). “Fungi associated with the vector of the pinewood nematode and their influence on Pine Wilt Disease,” in XVII Congress of European Mycologists (Madeira, Portugal), 21–25.
Jankowiak, R., Szewczyk, G., Bilański, P., Jazłowiecka, D., Harabin, B., and Linnakoski, R. (2021). Blue-stain fungi isolated from freshly felled Scots pine logs in Poland, including Leptographium sosnaicola sp. nov. For. Pathol. 51, 1–16. doi: 10.1111/efp.12672
Kanzaki, N., Tanaka, R., Giblin-Davis, R. M., and Davies, K. (2014). New plant-parasitic nematode from the mostly mycophagous genus Bursaphelenchus discovered inside figs in Japan. PLoS ONE 9, e99241. doi: 10.1371/journal.pone.0099241
Katoh, K., and Standley, D. (2013). MAFFT multiple sequence alignment software version 7: improvements in performance and usability. Mol. Biol. Evol. 30, 772–780. doi: 10.1093/molbev/mst010
Kim, J. J., Kim, S. H., Lee, S., and Breuil, C. (2003). Distinguishing Ophiostoma ips and Ophiostoma montium, two bark beetle-associated sapstain fungi. FEMS Microbiol. Lett. 222, 187–192. doi: 10.1016/S0378-1097(03)00304-5
Kirisits, T. (2007). “Fungal associates of European bark beetles with special emphasis on the ophiostomatoid fungi,” in Bark and Wood Boring Insects in Living Trees in Europe, a Synthesis, eds F. Lieutier, K. R. Day, A. Battisti, J. C. Grégoire, and H. F. Evans (Dordrecht: Springer), 181–235. doi: 10.1007/978-1-4020-2241-8_10
Kirisits, T., Wingfield, M. J., and Chhetri, D. B. (2002). Studies on the Association of Blue-Stain Fungi With the Eastern Hymalayan Spruce Bark Beetle (Ips schmutzenhoferi) and With Other Bark Betles in Bhutan. Yusipang Report (Vienna), 55.
Klepzig, K. (1998). Competition between a biological control fungus, Ophiostoma piliferum, and symbionts of the southern pine beetle. Mycologia 90, 69–75. doi: 10.1080/00275514.1998.12026880
Klepzig, K. D., and Hofstetter, R. W. (2011). “From attack to emergence: interactions between southern pine beetle, mites, microbes, and trees,” in 2011 Southern Pine Beetle II. Gen. Tech. Rep. SRS-140, eds R. N. Coulson, and K. D. Klepzig (Asheville, NC: U.S. Department of Agriculture Forest Service, Southern Research Station), 141–152.
Kuroda, K., and Ito, S. (1992). Migration speed of pine wood nematodes and activities of other microbes during the development of pine wilt disease in Pinus thunbergii. Kansai Res. Center Prod. Res. Inst. 74, 383–389.
Letunic, I., and Bork, P. (2021). Interactive Tree Of Life (iTOL) v5: an online tool for phylogenetic tree display and annotation. Nucleic Acids Res. 49, W293–W296, doi: 10.1093/nar/gkab301
Liu, Y., Qu, Z. L., Liu, B., Ma, Y., Xu, J., Shen, W. X., et al. (2021). The impact of pine wood nematode infection on the host fungal community. Microorganisms 9, 1–17. doi: 10.3390/microorganisms9050896
Maehara, N. (2008). Reduction of Bursaphelenchus xylophilus (Nematoda: Parasitaphelenchidae) population by inoculating Trichoderma spp. into pine wilt-killed trees. Biol. Control 44, 61–66. doi: 10.1016/j.biocontrol.2007.09.005
Maehara, N., and Futai, K. (1996). Factors affecting both the numbers of the pinewood nematode, Bursaphelenchus xylophilus (Nematoda: Aphelenchoididae), carried by the Japanese Pine Sawyer, Monochamus alternatus (Coleoptera: Cerambycidae), and the Nematode's Life History. Appl. Entomol. Zool. 31, 443–452. doi: 10.1303/aez.31.443
Maehara, N., and Futai, K. (1997). Effect of fungal interactions on the numbers of the pinewood nematode, Bursaphelenchus xylophilus (Nematoda: Aphelenchoididae), carried by the Japanese pine sawyer, Monochamus alternatus (Coleoptera: Cerambycidae). Fundam. Appl. Nematol. 20, 611–617.
Maehara, N., and Futai, K. (2000). Population changes of the pinewood nematode, Bursaphelenchus xylophilus (Nematoda: Aphelenchoididae), on fungi growing in pine-branch segments. Appl. Entomol. Zool. 35, 413–417. doi: 10.1303/aez.2000.413
Maehara, N., and Futai, K. (2005). Effect of blue-stain fungi on the number of Bursaphelenchus xylophilus (Nematoda: Aphelenchoididae) carried by Monochamus alternatus (Coleoptera: Cerambycidae). Nematology 7: 161–167. doi: 10.1163/1568541054879557
Maehara, N., Tsuda, K., Yamasaki, M., Shirakikawa, S., and Futai, K. (2006). Effect of fungus inoculation on the number of Bursaphelenchus xylophilus (Nematoda: Aphelenchoididae) carried by Monochamus alternatus (Coleoptera: Cerambycidae). Nematology 8, 59–67. doi: 10.1163/156854106776179944
Mamiya, Y. (1983). Pathology of the pine wilt disease caused by Bursaphelenchus xylophilus. Ann. Rev. Phytopathol. 21, 201–220. doi: 10.1146/annurev.py.21.090183.001221
Marincowitz, S., Duong, T., Wilhelm de Beer, Z., and Wingfield, M. (2015). Cornuvesica: a little known mycophilic genus with a unique biology and unexpected new species. Fungal Biol. 119, 615–630. doi: 10.1016/j.funbio.2015.03.007
Mensah, J. K., Sayer, M. A. S., Nadel, R. L., Matusick, G., Fan, Z., Carter, E. A., et al. (2021). Leptographium terebrantis inoculation and associated crown symptoms and tree mortality in Pinus taeda. Fungal Ecol. 51, 101057. doi: 10.1016/j.funeco.2021.101057
Mota, M. M., Braasch, H., Bravo, M. A., Penas, A. C., Burgermeister, W., Metge, K., et al. (1999). First report of Bursaphelenchus xylophilus in Portugal and in Europe. Nematology 1, 727–734. doi: 10.1163/156854199508757
Nascimento, F. X., Hasegawa, K., Mota, M., and Vicente, C. S. L. (2015). Bacterial role in pine wilt disease development - review and future perspectives. Environ. Microbiol. Rep. 7, 51–63. doi: 10.1111/1758-2229.12202
Niu, H. T., Zhao, L. L., Lu, M., Zhang, S., and Sun, J. H. (2012). The ratio and concentration of two monoterpenes mediate fecundity of the pinewood nematode and growth of its associated fungi. PLoS ONE 7, e31716. doi: 10.1371/journal.pone.0031716
Pimentel, C. S., Firmino, P. N., and Ayres, M. P. (2021). Interactions between pinewood nematodes and the fungal community of pine trees. Fungal Ecol. 51, 101046. doi: 10.1016/j.funeco.2021.101046
Plattner, A., Kim, J.-J., Reid, J., Hausner, G., Lim, Y. W., Yamaoka, Y., and Breuil, C. (2009). Resolving taxonomic and phylogenetic incongruence within species Ceratocystiopsis minuta. Mycologia 101, 878–887. doi: 10.3852/08-132
R Core Team (2020). R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing. Available online at: https://www.R-project.org/ (accessed March 29, 2022).
Reid, J., Iranpour, M., Rudski, S. M., Loewen, P. C., and Hausner, G. (2010). A new conifer-inhabiting species of Ceratocystis from Norway. Botany 88, 971–983. doi: 10.1139/B10-069
Robertson, L., Arcos, S. C., Escuer, M., Merino, R. S., Esparrago, G., Abelleira, A., et al. (2011). Incidence of the pinewood nematode Bursaphelenchus xylophlius (Steiner & Buhrer, 1934) Nickle, 1970 in Spain. Nematology 13, 755–757. doi: 10.1163/138855411X578888
Rodrigues, J. M., Sousa, E., and Abrantes, I. (2015). “Pine Wilt Disease historical overview,” in Pine Wilt Disease in Europe – Biological Interactions and Integrated Management. 1st ed, E. Sousa, F. Vale, and I. Abrantes (Lisbon: Federação Nacional das Associações de Proprietários Florestais), 13–32
Royse, D. J., and Ries, S. M. (1977). The influence of fungi isolated from peach twigs on the pathogenicity of Cytospora cincta. Ecol. Epidemiol. 68, 603–607. doi: 10.1094/Phyto-68-603
Rumbold, C. T. (1931). Two blue-staining fungi associated with bark beetle infestation of pines. J. Agric. Res. 43, 847–873.
Shade, A., Peter, H., Allison, S. D., Baho, D. L., Berga, M., Bürgmann, H., et al. (2012). Fundamentals of microbial community resistance and resilience. Front. Microbiol. 3, 1–19. doi: 10.3389/fmicb.2012.00417
Six, D. L. (2012). Ecological and evolutionary determinants of bark beetle —fungus symbioses. Insects 3, 339–366. doi: 10.3390/insects3010339
The jamovi project (2021). jamovi (Version 1.6) [Computer Software]. Retrieved from: https://www.jamovi.org (accessed March 29, 2022)
Trindade, M. J. F. (2019). Estudo da população de fungos em Pinus pinaster em P ortugal. Master Thesis, Instituto Superior de Agronomia, Lisbon.
Vicente, C., Espada, M., Vieira, P., and Mota, M. (2012). Pine Wilt Disease: a threat to European forestry. Eur. J. Plant Pathol. 133, 89–99. doi: 10.1007/s10658-011-9924-x
Vicente, C. S. L., Soares, M., Faria, J. M. S., Ramos, A. P., and Inácio, M. L. (2021). Insights into the role of fungi in pine wilt disease. J. Fungi 7, 1–16. doi: 10.3390/jof7090780
Wang, H. M., Lun, Y. Y., Lu, Q., Liu, H. X., Decock, C., and Zhang, X. Y. (2018). Ophiostomatoid fungi associated with pines infected by Bursaphelenchus xylophilus and Monochamus alternatus in China, including three new species. MycoKeys 39, 1–27. doi: 10.3897/mycokeys.39.27014
White, T., Bruns, T., Lee, S., and Taylor, J. (1990). “Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics,” in PCR protocols: A Guide to Methods and Applications, eds M. A. Innis, D. H. Gelfand, J. J. Snisky, and T. J. White (Academic Press), 315–322. doi: 10.1016/B978-0-12-372180-8.50042-1
Whitehead, A. G., and Hemming, J. R. (1965). A comparison of some quantitative methods of extracting small vermiform nematodes from soil. Ann. Appl. Biol. 55, 25–38. doi: 10.1111/j.1744-7348.1965.tb07864.x
Zhang, W., Wang, X., Li, Y., Wei, P., Sun, N., Wen, X., et al. (2021). Differences between microbial communities of pine species having differing level of resistance to the pinewood nematode. Microl. Ecol. doi: 10.1007/s00248-021-01907-4
Zhao, L., Lu, M., Niu, H., Fang, G., Zhang, S., and Sun, J. (2013). A native fungal symbiont facilitates the prevalence and development of an invasive pathogen–native vector symbiosis. Ecology 94, 2817–2826. doi: 10.1890/12-2229.1
Zhao, L., Mota, M., Vieira, P., Butcher, R. A., and Sun, J. (2014). Interspecific communication between pinewood nematode, its insect vector, and associated microbes. Trends Parasitol. 30, 299–308. doi: 10.1016/j.pt.2014.04.007
Zhao, L.;, Ahmad, F., Lu, M., Zhang, W., Wickham, J. D., Sun, J., et al. (2018). Ascarosides promote the prevalence of ophiostomatoid fungi and an invasive pathogenic nematode, Bursaphelenchus xylophilus. J. Chem. 44, 701–710. doi: 10.1007/s10886-018-0996-3
Keywords: biocontrol, blue-stain fungi, diversity, ecological interactions, mycoflora, pinewood nematode
Citation: Vicente CSL, Soares M, Faria JMS, Espada M, Mota M, Nóbrega F, Ramos AP and Inácio ML (2022) Fungal Communities of the Pine Wilt Disease Complex: Studying the Interaction of Ophiostomatales With Bursaphelenchus xylophilus. Front. Plant Sci. 13:908308. doi: 10.3389/fpls.2022.908308
Received: 30 March 2022; Accepted: 02 May 2022;
Published: 14 June 2022.
Edited by:
Xiangming Xu, National Institute of Agricultural Botany (NIAB), United KingdomReviewed by:
Noritoshi Maehara, Forestry and Forest Products Research Institute, JapanCopyright © 2022 Vicente, Soares, Faria, Espada, Mota, Nóbrega, Ramos and Inácio. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Maria L. Inácio, bHVyZGVzLmluYWNpb0Bpbmlhdi5wdA==
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.