 Rui Song
Rui Song Ran Tong2
Ran Tong2 G. Geoff Wang
G. Geoff Wang Tonggui Wu
Tonggui Wu
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Plant Sci. , 11 May 2022
Sec. Functional Plant Ecology
Volume 13 - 2022 | https://doi.org/10.3389/fpls.2022.905358
This article is part of the Research Topic Physiological Ecology of Trees under Environmental Stresses View all 14 articles
The plant nutrient acquisition strategies are diverse, such as root nutrient acquisition and leaf nutrient resorption, playing important roles in driving soil processes, vegetation performance as well as ecosystem nutrient cycling. However, it is still in a debate whether there is a synergy or tradeoff between above- and below-ground nutrient acquisition strategy under nitrogen (N) and phosphorus (P) addition, or with stand age. Herein, this study investigated the responses of root-soil accumulation factor (RSAF) and leaf nutrient resorption efficiency (NuRE) to long-term N and P fertilization, and further explored the trade-off between them in Metasequoia glyptostroboides plantations with different stand age. Results showed that under N fertilization in young plantations, leaf N resorption efficiency (NRE) increased, and root-soil accumulation factor for P (RSAF-P) decreased. For young forests under P fertilization, the NRE increased whereas RSAF-P decreased. For middle-aged forests under P fertilization, the NRE and leaf P resorption efficiency (PRE) increased and the RSAF-P decreased. Under P fertilization in young and middle-aged plantations, PRE had a significant positive correlation with RSAF-P. Under N fertilization in young plantations, NRE was significantly positive correlated with root-soil accumulation factor for N (RSAF-N). The covariance-based structural equation modeling (CB-SEM) analysis indicated that stand age had positive effects on PRE whether under N or P fertilization, as well as on RSAF-P under N fertilization, whereas had no effects on the NRE or RSAF-N. Overall, our results can shed light on the nutrient acquisition strategies of M. glyptostroboides plantations under future environmental changes and the results could be applied to the nutrient management practices.
The plant nutrient acquisition strategies cannot be underestimated in forest ecosystems, given that the plants usually do not have sufficient amounts of biologically available nutrients to support growth, survival, and reproduction, especially in nutrient-impoverished habitats (Abrahão et al., 2018; Wang et al., 2020). It has been always considered that roots are the major tissue for the plant to acquire nutrients, and root nutrient acquisition strategies play a decisive role in the maintenance of forest ecosystem health and vitality (Andersen et al., 2017; Ma et al., 2018; Li et al., 2019). In recent decades, some other plant nutrient acquisition strategies have also been proposed, such as leaf nutrient resorption proficiency and efficiency (Reed et al., 2012; He et al., 2020; Xu et al., 2020). Naturally, whether these plant nutrient acquisition strategies co-vary or exhibit coordinated responses to a changing environment has drawn great attention from ecologists.
Allocation of effort toward nutrient capture and resorption depends on both the environment nutrient availability and the cost involved in these processes (Wright and Westoby, 2003; Wang et al., 2014a). In barren soils, plants should bear traits that prioritize conservation over active absorption of resources, whereas the opposite was expected in nutrient-rich soils. Specifically, since nutrients derived from capture become less expensive than those from resorption as soil nutrient availability increases, the root capture strategy would be more favored than leaf resorption (Wright and Westoby, 2003; Kou et al., 2017). Inversely, species from nutrient-impoverished habitats also reduce nutrient losses by remobilizing a large fraction of the nutrients before senesced organ shedding (high remobilization efficiency), and shedding leaves with very low final nutrient concentrations (Killingbeck, 1996; Kobe et al., 2005; Hayes et al., 2014). Nevertheless, in cases of consistent environmental nutrient availability, the costs of the different plant nutrient acquisition strategies have not been effectively evaluated.
Increasing anthropogenic nitrogen (N) and phosphorus (P) deposition has enhanced N and P availability in many forest ecosystems, respectively, providing ideal venues for the studies of the trade-off relationships of plant nutrient acquisition strategies. It is well known that plants can maintain high nutrients level under nutrient enrichment through increasing above-ground nutrient conservation or improving below-ground nutrient uptake (Deng et al., 2016; Hofmann et al., 2016). Several studies showed the trade-off relationships between root nutrient acquisition and leaf nutrient resorption while only considering these two mechanisms (Deng et al., 2016; Kou et al., 2017). In contrast, other studies found the synergy relationship between these two mechanisms, and emphasized that only adopting above-ground nutrient conservation of below-ground nutrient uptake may not be applicative (Ushio et al., 2015; Hofmann et al., 2016).
The corresponding alterations in tree nutrient capture capacity during forest stand development had been widely demonstrated (Brant and Chen, 2015; Chen and Chen, 2022). For instance, the NRE rose and then dropped, and the PRE increased with stand age in Medicago sativa, which was closely related to the nutrient limitation of plant growth (Wang et al., 2014b). As trees had different root physiology and exudation patterns at different ages, their root nutrient capture capacity exhibited significant divergence (Liu et al., 2018). Yet, whether the patterns of the tree nutrient capture with different stand ages would alter under nutrient addition remained unclear.
As each mechanism demands distinct levels of resource investments, as well as its potential consequences for ecosystem functions, it is critical to understand how plants adjust above-ground conservation and below-ground uptake to alleviate N and P deficiency as stand development. Taken together, it is generally believed that a trade-off mechanism generally existed between the leaf nutrient resorption and the root acquisition strategies whether in the cases of sufficient or deficient nutrients supplies. In this study, we conducted N and P fertilization experiments to explore the effects of different gradient N and P fertilization on leaf nutrient resorption efficiency (NuRE) and root-soil accumulation factor (RSAF) in M. glyptostroboides plantations with different stand ages. Specifically, we aimed to: (1) describe the change trends of NuRE and RSAF with the increase of nutrient availability among nutrient fertilization types and along stand development; (2) explore the potential relationships between NuRE and RSAF under environmental changes.
The study was conducted in Dongtai Forest Farm (121° 45′E, 33° 42′N), a site located in the central coast of the Jiangsu Province, China. The climate of the study site is that of the monsoon subtropical moist marine climate zone. The altitude is 0–4 m, the annual average temperature, frost-free period, precipitation, and sunshine duration is 14.5°c, 220 days, 1055.7 mm, and 2130.5 h. The soil is alkaline sandy soil, which was more infertile than yellow-earth soil in the natural range of M. glyptostroboides. The study site was the pioneer area to plant the M. glyptostroboides in coastal China (Figure 1).
Figure 1. Location of M. glyptostroboides forest experiment site in Dongtai forest farm, Jiangsu Province, China.
N and P experiment was initiated in 2014 of different ages (young forest: 6-year old; middle-age forests: 24-year old) with three 300 m × 100 m plots of M. glyptostroboides plantations in Dongtai forest farm, and ten 20 m × 30 m plots of each plot were selected for completely random block N and P addition experiments, of which 5 plots were used for N addition treatments and 5 plots for P addition treatments in each age class. Each plot was separated by a buffer zone. The basic information is shown in Table 1. N and P addition gradients setting: set the N addition treatments as CK, 0.8, 2.4, 4.0, and 6.0 mol⋅m–2, respectively, and the fertilization was added in the form of urea [CO(NH2)2]; P fertilization treatment was set as CK, 0.05, 0.2, 0.6, and 1.0 mol⋅m–2, respectively, and fertilization was added in the form of superphosphate [Ca(H2PO4)2]. The fertilization gradients were based on a pretest and a previous seedling pot study (Wang et al., 2018). Fertilization dosage and time: the first addition time is from the end of March to the beginning of April, 60% of the total amount of fertilization added; the second addition time is in the middle of June, 40% of the total amount of fertilization added. In addition, N and P were applied to the field directly every year.
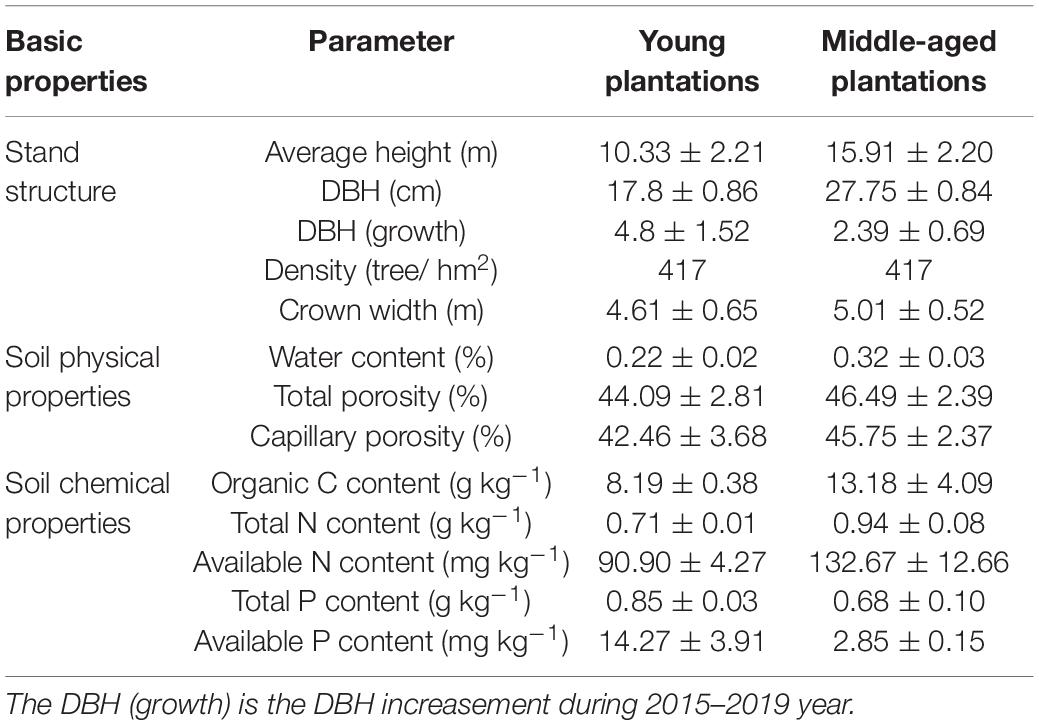
Table 1. General characteristics of the young and middle-aged M. glyptostroboides stands.
Three average trees in each plot were selected for collecting leaves, branches, and roots. Fully expanded leaves and branches from the upper and outer part of tree crowns were sampled. The fine roots (<2 mm) of each selected individual were sampled through the careful removal of the soil surrounding the roots. Five soil cores (2.5 cm in diameter) per plot were randomly collected from 0 to 10 cm depth following the removal of understory plants and surface litter, and thoroughly mixed to homogenize a sample. Leaves, branches, roots, and soils were sampled in August 2018 (the end of the growing season). Five litter traps (1.0 m2, made of nylon mesh) per plot were fixed 1.0 m above the ground. Leaf litter was collected in late November 2018.
All plant organs samples were dried in the oven at 105°C for 2 h, then dried at 75°C to constant weight, crushed with a mechanical grinder, and the samples were sieved to determine nutrient content. The C and N concentrations were determined for each sample using an autoanalyzer (Kjeltec 2300 Analyzer Unit, Foss, Sweden). P concentration was determined using the standard ammonium molybdate method (reference code GBW08513; General Administration of Quality Supervision, PRC).
Soil samples were air-dried after being sieved (2-mm mesh). Soil pH was determined by the potentiometric method, soil organic carbon (SOC) was determined with wet oxidation by sulfuric acid and potassium dichromate and back titration with ferrous sulfate. The total N concentration was determined by Kjeldahl method, Hydrolysable N concentration was determined by titration with a dilute solution of H2SO4 after extraction with a mixture of ferrous sulfate and sodium hydroxide. The total P and available P concentrations were determined with a molybdate blue colorimeter after extraction with 0.5 M sodium bicarbonate (Zhang et al., 2018).
Nutrient resorption efficiency (NuRE) was defined as the proportional withdrawal of a nutrient during senescence and was calculated as follows:
Where NuRE is N or P resorption efficiency, Nusenescenced and Nugreen represent N or P concentration (mass-based) in leaf and litter, respectively, and MLCF is mass loss correction factor with a value of 0.745 for deciduous coniferous species (Vergutz et al., 2012).
Root-soil accumulation factor (RSAF) for N and P were defined to describe the root N or P acquisition (Kou et al., 2017), as follows:
Where RSAF is root N or P accumulation factor, Nuroot and Nusoil represent N or P concentration (mass-based) in absorptive fine root and soil, respectively.
Data analysis using the single factor variance (One-way ANOVA) analysis of the different fertilization gradients of plant organs stoichiometric characteristics, nutrient acquisition, nutrient recycling, and nutrient absorption efficiency under different stand ages, respectively, the influence of significance level set as P = 0.05, with minimal process significantly difference (LSD) determined the level of significance in SPSS 23.0 (SPSS Inc., Chicago, IL, United States). The covariance-based structural equation modeling (CB-SEM) analysis was conducted in SPSS Amos 26 (SPSS Inc., Chicago, IL, United States) to explore the effects of stand age on NuRE and RSAF under N and P fertilization. All data were tested to fulfill the assumptions of normality and homogeneity of variance, and transformations were carried out when necessary. Figures were plotted using Origin 2018 (Origin Lab Corporation, Northampton, MA, United States).
Under N fertilization in young plantations, the NRE and PRE of CK treatment were 31.02 and 64.25%, respectively. The NRE remarkably increased while PRE kept relatively stable along N fertilization gradients (Figure 2A). The RSAF-N and RSAF-P of CK treatment were 131.75 and 198.49, respectively. The RSAF-P remarkably decreased while RSAF-N kept relatively stable along N fertilization gradients (Figure 2B).
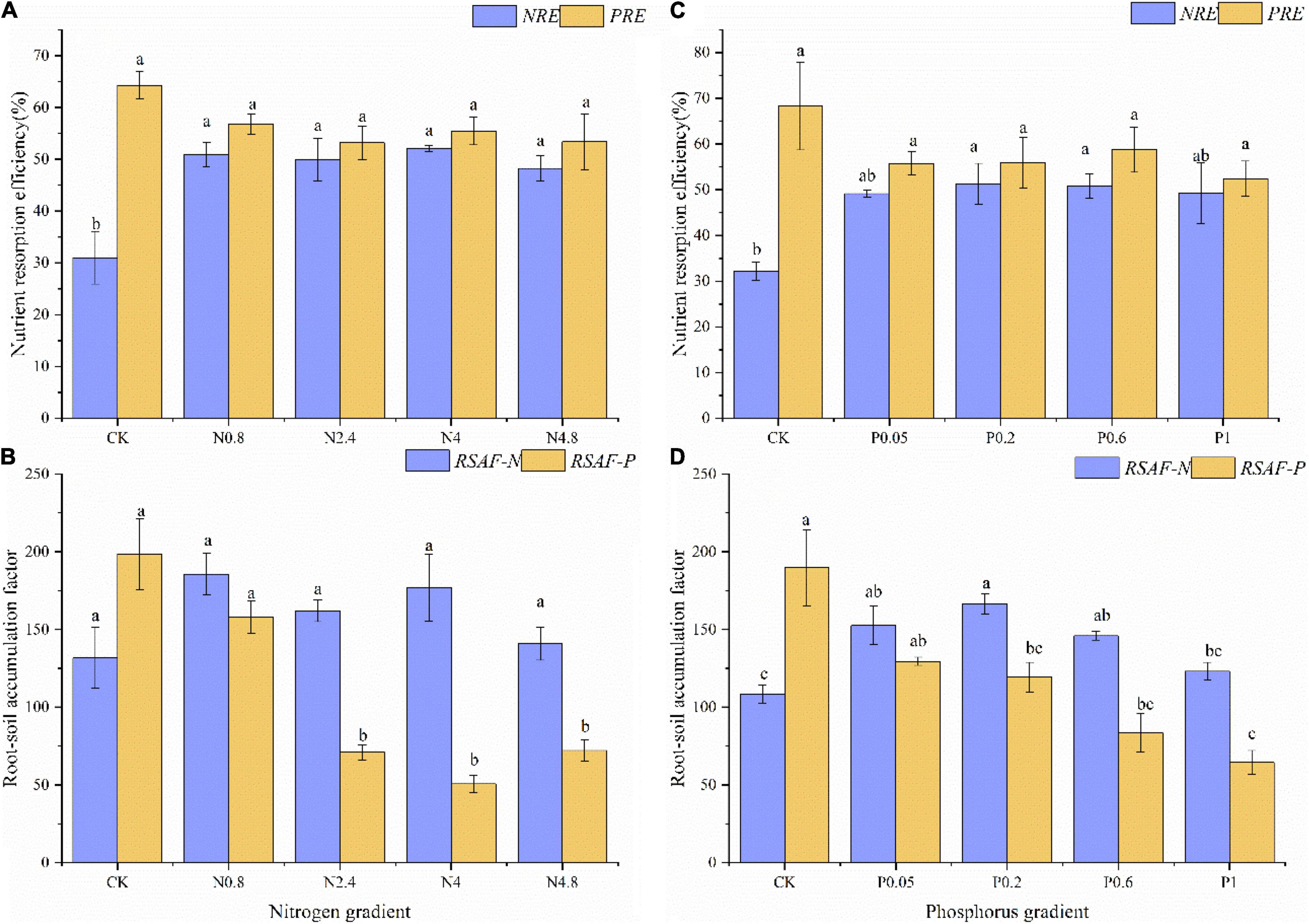
Figure 2. Effects of N fertilization (CK: control, N0.8: 0.8 mol N, N2.4: 2.4 mol N, N4: 4 mol N, N4.8: 4.8 mol N) on NuRE (A) and RSAF (B) in M. glyptostroboides forests of young forests. Values are presented as mean ± S.E. (n = 3). Effects of P fertilization (CK: control, P0.05: 0.05 mol P, P0.2: 0.2 mol P, P0.6: 0.6 mol P, P1: 1 mol P) on NuRE (C) and RSAF (D) in M. glyptostroboides forests of young forests. Values are presented as mean ± S.E. (n = 3). Different lowercase letters in the bars and above the bars indicated significant difference (P < 0.05), respectively.
Under P fertilization in young plantations, the NRE and PRE of CK treatment were 32.17 and 68.31%, respectively. The NRE remarkably increased, while PRE kept relatively stable along P fertilization gradients (Figure 2C). The RSAF-N and RSAF-P of CK treatment were 108.38 and 189.6, respectively. The RSAF-N and RSAF-P remarkably increased and decreased along P fertilization gradients (Figure 2D).
Under N fertilization in middle-aged plantations, the NRE and PRE of CK treatment were 45.02 and 54.24%, respectively. The NRE and PRE both kept relatively stable along N fertilization gradients (Figure 3A). The RSAF-N and RSAF-P of CK treatment were 82.59 and 126.71, respectively. The RSAF-N and RSAF-P both kept relatively stable along N fertilization gradients (Figure 3B).
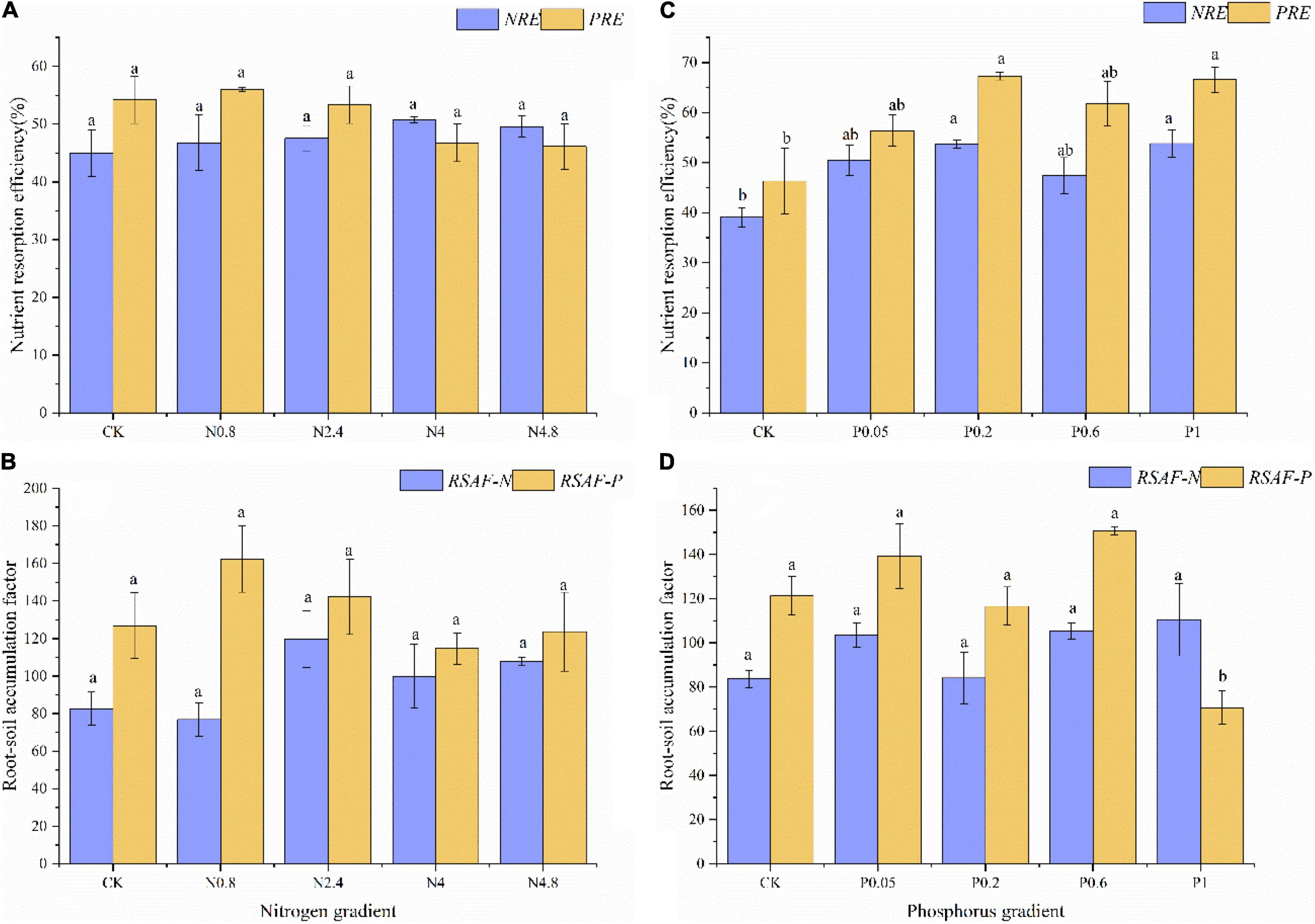
Figure 3. Effects of N fertilization (CK: control, N0.8: 0.8 mol N, N2.4: 2.4 mol N, N4: 4 mol N, N4.8: 4.8 mol N) on NuRE (A) and RSAF (B) in M. glyptostroboides forests of middle-aged forests. Values are presented as mean ± S.E. (n = 3). Effects of P fertilization (CK: control, P0.05: 0.05 mol P, P0.2: 0.2 mol P, P0.6: 0.6 mol P, P1: 1 mol P) on NuRE (C) and RSAF (D) in M. glyptostroboides forests of middle-aged forests. Values are presented as mean ± S.E. (n = 3). Different lowercase letters in the bars and above the bars indicated significant difference (P < 0.05), respectively.
Under P fertilization in middle-aged plantations, the NRE and PRE of CK treatment were 39.07 and 46.33%, respectively. The NRE and PRE both remarkably increased remarkably along P fertilization gradients (Figure 3C). The RSAF-N and RSAF-P of CK treatment were 83.64 and 121.24, respectively. The RSAF-N and RSAF-P both kept relatively stable along P fertilization gradients (Figure 3D).
The NRE/PRE of CK treatment was 0.49 and 0.83 in young and middle-aged plantations under N fertilization, respectively. The NRE/PRE for young plantation increased significantly under N fertilization, while the NRE/PRE for middle-aged plantations kept relatively stable along N fertilization gradients (Figure 4A). The RSAF-N/RSAF-P of CK treatment were 0.69 and 0.68 in young and middle-aged plantations under N fertilization, respectively. The RSAF-N/RSAF-P for young plantation increased significantly under N fertilization, while the RSAF-N/RSAF-P in middle-aged plantations kept relatively stable along N fertilization gradients (Figure 4B).
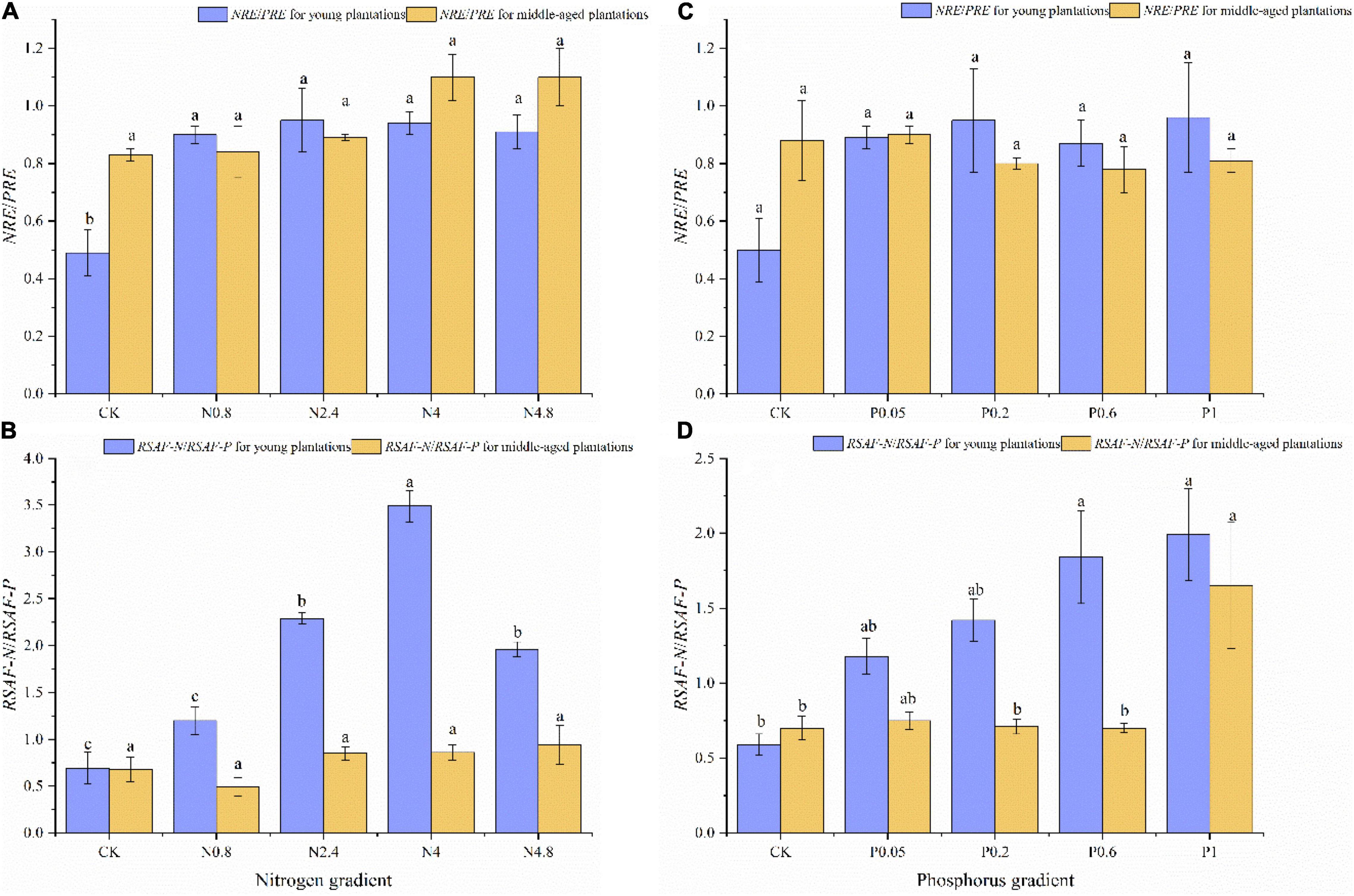
Figure 4. Effects of N fertilization (CK: control, N0.8: 0.8 mol N, N2.4: 2.4 mol N, N4: 4 mol N, N4.8: 4.8 mol N) on NRE/PRE (A) and RSAF-N/RSAF-P (B) in M. glyptostroboides plantations of different stand ages. Values are presented as mean ± S.E. (n = 3). Effects of P fertilization (CK: control, P0.05: 0.05 mol P, P0.2: 0.2 mol P, P0.6: 0.6 mol P, P1: 1 mol P) on NRE/PRE (C) and RSAF-N/RSAF/P (D) in M. glyptostroboides plantations of different stand ages. Different lowercase letters in the bars and above the bars indicated significant difference (P < 0.05), respectively.
The NRE/PRE of CK treatment was 0.50 and 0.88 in young and middle-aged plantations under P fertilization, respectively. The NRE/PRE in young and middle-aged plantations both kept relatively stable along P fertilization gradients (Figure 4C). The RSAF-N/RSAF-P of CK treatment were 0.59 and 0.70 in young and middle-aged plantations under P fertilization, respectively. The RSAF-N/RSAF-P remarkably increased under P fertilization, while the RSAF-N/RSAF-P in middle-aged plantations kept relatively stable along P fertilization gradients (Figure 4D).
Under N fertilization, NRE showed no association with RSAF-N, while PRE was significantly positive correlated with RSAF-P in young and middle-aged plantations (Figures 5A,C). Under P fertilization, NRE was significantly positively correlated with RSAF for N in young plantation, whereas no significant relationships were found between NRE and RSAF-N in middle-aged plantation, as well as PRE and RSAF-P in both young and middle-aged plantations (Figures 5B,D).
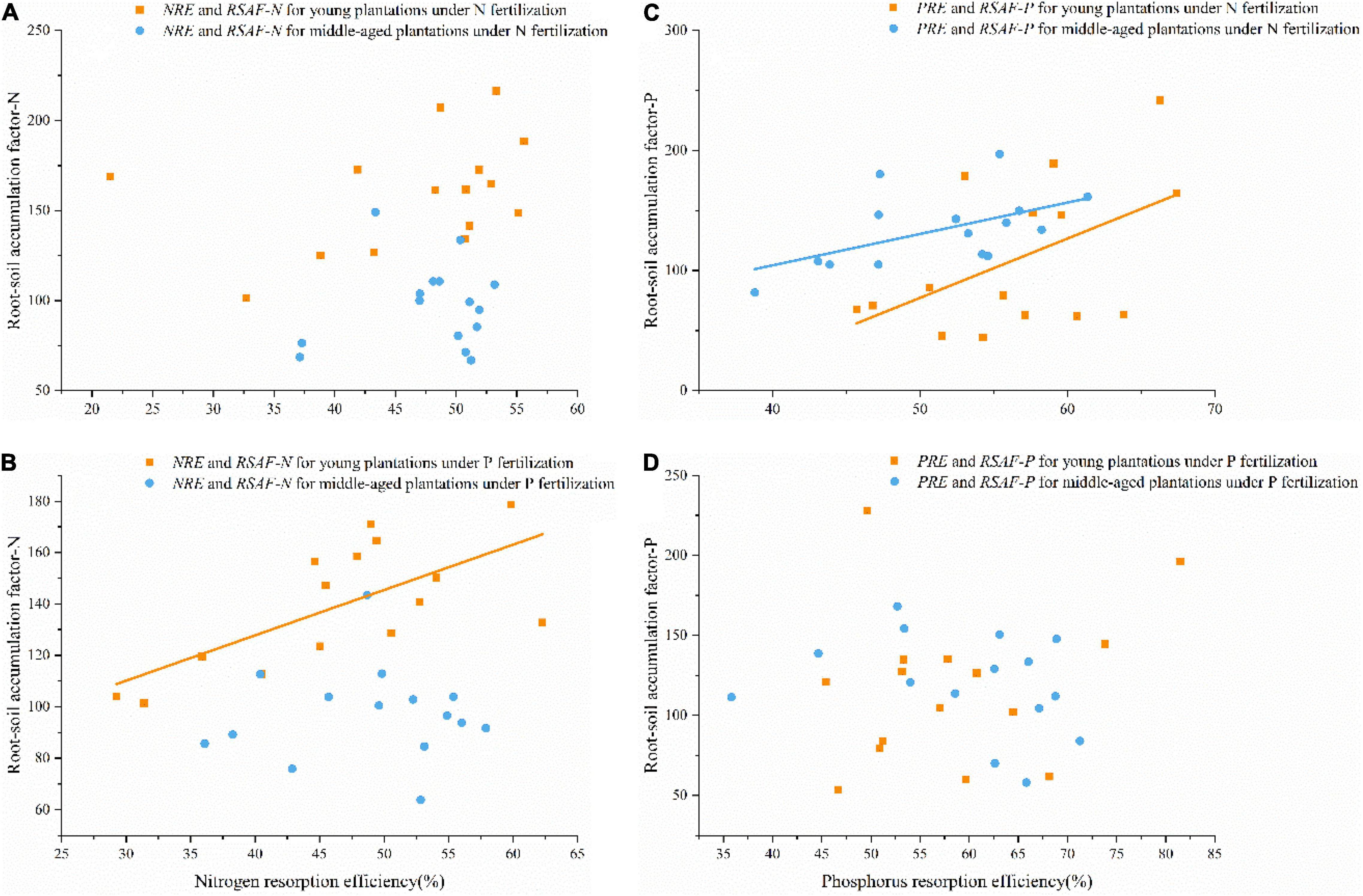
Figure 5. (A) Relationships between NRE and RSAF-N under N fertilization in young and middle-aged plantations (no significance). (B) Relationships between NRE and RSAF-N under P fertilization in young (R2 = 0.468, P < 0.01) and middle-aged (no significance) plantations. (C) Relationships between PRE and RSAF-P under N fertilization in young (R2 = 0.266, P < 0.05) and middle-aged plantations (R2 = 0.291, P < 0.05). (D) Relationships between PRE and RSAF-P under P fertilization in young and middle-aged plantations (no significance).
The covariance-based structural equation modeling (CB-SEM) analysis indicted that N fertilization and stand age together explained 84.3% of variations in NRE and 73.5% of variations in PRE (Figure 6A). N fertilization had a positive indirect effect on NRE via leaf N, and a negative indirect effect on PRE through leaf P and soil pH. Stand age had a positive direct effect on PRE via soil pH, and a negative indirect effect via soil total N and leaf P. N fertilization and stand age together explained 62.8% of variations in RSAF-N and 56.0% of variations in RSAF-P (Figure 6B). N fertilization and stand age had a negative and positive direct effect on RSAF-P, respectively. N fertilization had a positive indirect effect on RSAF-N via fine root N. Stand age had a negative indirect effect on RSAF-N via soil N:P.
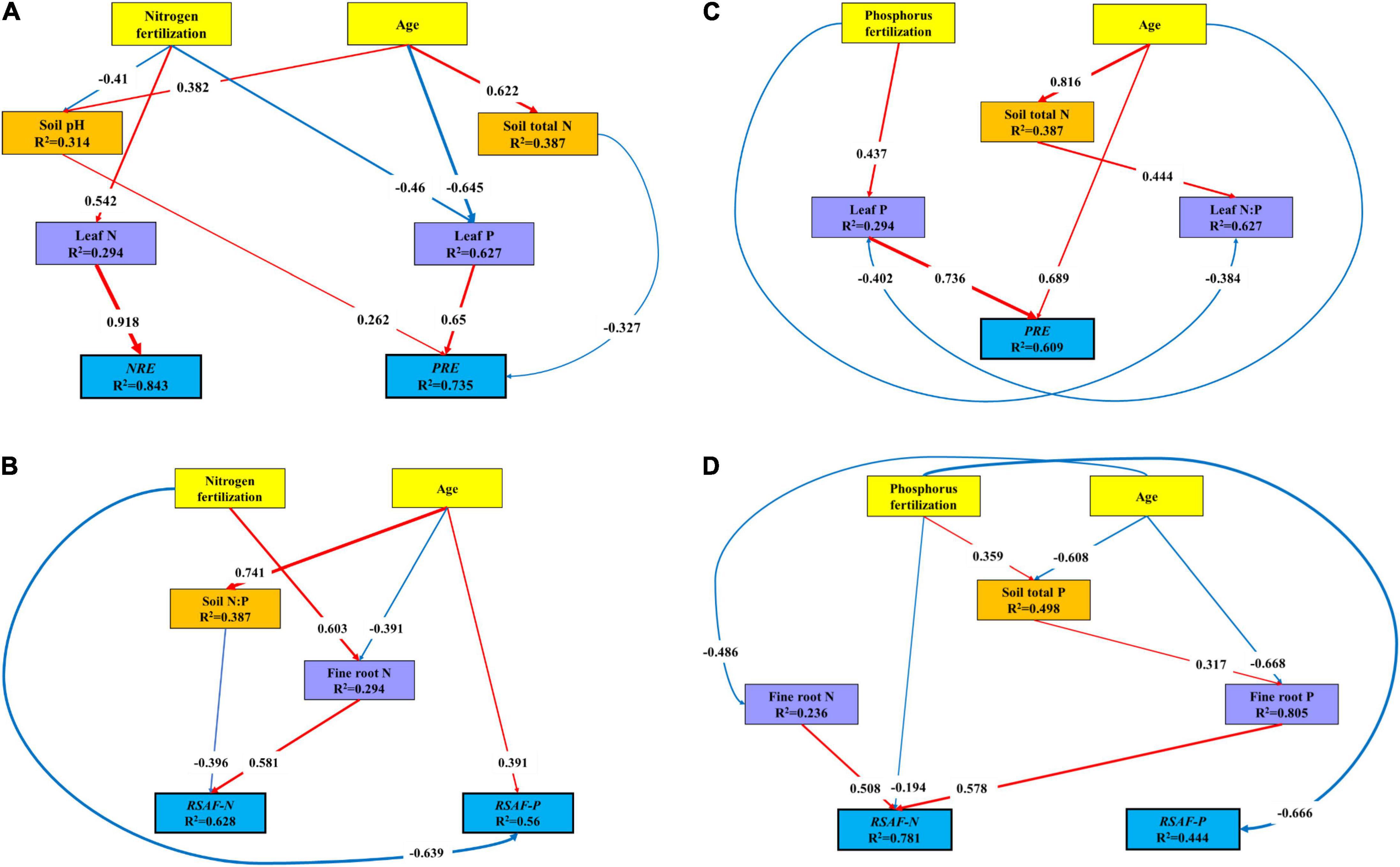
Figure 6. Covariance-based structural equation modeling (CB-SEM) analysis of the direct and indirect effects among variables. All the paths in the model were significant, and the standardized coefficients were listed on them. The thickness of the solid arrows reflected the magnitude of the standardized SEM coefficients, the red solid line represented the positive effect while the blue solid line represented the negative effect. (A) Results of model fitting: X2 = 13.51; Probability level = 0.563; df = 15; GFI = 0.9; RMSEA = 0.000; AIC = 55.51. (B) Results of model fitting: X2 = 7.116; Probability level = 0.524; df = 8; GFI = 0.927; RMSEA = 0.000; AIC = 33.116. (C) Results of model fitting: X2 = 5.915; Probability level = 0.55; df = 7; GFI = 0.938; RMSEA = 0.000; AIC = 33.915. (D) Results of model fitting: X2 = 10.519; Probability level = 0.484; df = 11; GFI = 0.918; RMSEA = 0.000; AIC = 44.519.
P fertilization and stand age together explained 60.9% of variations in PRE (Figure 6C). Stand age had a positive direct effect on PRE, and P fertilization had a positive indirect effect on PRE via leaf P. P fertilization and stand age together explained 78.1% of variations in RSAF-N, and 44.4% of variations in RSAF-P (Figure 6D). P fertilization had a negative direct effect on RSAF-P, and stand age had a negative direct effect on RSAF-N. Stand age had a negative indirect effect on RSAF-N via fine root N and P.
Bioavailable N is increasing due to human activity, which greatly alters nutrient availability for plant growth and affects fundamental ecological processes (Wang et al., 2019a). Nutrient resorption from senescing leaves is one of the plants’ essential nutrient conservation strategies, accounting for a large property of the nutrient demand for plants (Aerts, 1996). In this study, N fertilization increased leaf N concentration in young plantation, which was in line with the observations from the global-scale meta-analysis (Ostertag and DiManno, 2016; Sardans et al., 2017). However, in contrast to most previous studies, we found a significant increase in NRE for young plantation and no significant changes for middle-aged plantation under N fertilization (Yuan and Chen, 2015; Lu et al., 2019; Zhao et al., 2020). Our study also found that leaf litter N concentration kept relatively stable under N fertilization whether in young and middle-aged plantations (Wen et al., 2021). Thus, considering the violent strong natural and human-activities disturbances, we deemed that nutrient return of leaf litter to the soil, i.e., litter decomposition, might not be so essential, especially in the coastal man-made forests. Moreover, the RSAF-P decreased remarkably under N fertilization, which was not consistent with Kou et al. (2017), who reported that the RSAF-P kept relatively stable in Pinus elliottii plantations.
The NRE/PRE in leaf was recently developed to indicate the theoretical framework of N or P limitation for plant growth, which could be applied to quantitatively evaluate the spatial characteristics and key influencing factors of N and P limitation in the terrestrial ecosystems (Du et al., 2020). In this study, the NRE/PRE was always less than 1 under N fertilization, indicating that the plant growth was mostly limited by N. This result was consistent with one of our previous studies, which found N was the main limiting factor for M. glyptostroboides growth combining with the concept of stoichiometric homeostasis (Wang et al., 2019b). Moreover, the NRE/PRE of young plantation increased along the gradient of N fertilization, demonstrating that the N limitation got relieved to some extent (Li et al., 2016; Du et al., 2020).
It was always expected that plant tissues exhibited trade-offs relationships in water and nutrient acquisition, which is of great significance for plant stoichiometric homeostasis and the balance of matters and energy in the terrestrial ecosystems. While these trade-off relationships became unpredictable under the background of global change. In this study, we observed significant and positive associations between PRE and RSAF-P in both young and middle-aged plantations under N fertilization, which was not in line with Kou et al. (2017) for Pinus elliottii plantations. However, most of the trade-off relationships between leaf nutrient resorption and fine root nutrient acquisition had been lost under N fertilization. Therefore, we referred that the changes in environmental nutrient conditions might lead to the absence of trade-off relationships between under- and above-ground plant tissues.
Atmospheric P deposition brought by agricultural activities, dust transport, and combustion source emissions had greatly altered the soil P availability that greatly influenced plant growth (Widdig et al., 2019; Pan et al., 2021). Generally, the availability of one nutrient would inevitably affect relative availability of other nutrients, particularly in the status of nutrient deficiency or imbalance (Kou et al., 2017). In the current study, the NRE of young and middle-aged plantation increased along the gradient of P fertilization, which was also reported in Yan et al. (2015). Furthermore, the RSAF-N/RSAF-P increased remarkably whether under P fertilization in young plantation, and kept relatively stable in middle-aged plantation. This finding was also in line with the above observations for above-ground plant tissues. Therefore, fine root nutrient acquisition might play an important role in indicating the nutrient limitation of plant growth. Meanwhile, we observed that the RSAF-P decreased significantly under P fertilization whether in young or middle-aged plantation, which might be attributed to the fact that the fine root nutrient acquisition existed some delayed effects. Meanwhile, some limitations appeared in this study such as considering the fine roots with a diameter less than 2 mm, ignoring the high morphological and functional heterogeneity in fine roots (Pregitzer et al., 2002; Kou et al., 2018), which might lead to an underestimate of the RSAF.
Notably, the NRE and RSAF-N in young plantation under P fertilization, were significantly correlated with each other, even exhibiting synergy relationships to some extent. These findings were also observed in previous research, which emphasized that only adopting aboveground N and P conservation or belowground N and P uptake might not be accurate (Ushio et al., 2015; Hofmann et al., 2016). Therefore, we concluded that the synergistic relationship for P acquisition might be explained by the two following reasons. Firstly, the relationships between fine root nutrient acquisition and leaf nutrient resorption were indeed positively related, whereas there might exist some other mechanisms which could balance these two nutrient conversation mechanisms. Secondly, there was no trade-off evolutionary mechanism between leaves and roots, which might leading to the consistent responses of leaf nutrient resorption and fine root nutrient acquisition to environment changes by chance.
Trees at different growth stages have great differences in physiological processes and nutrition requirements, resulting in changes of nutrient acquisition strategies along an age sequence (Sun et al., 2016; Zhang et al., 2018). However, most previous studies considered the stand age effects on nutrient acquisition strategies in natural conditions, while little was known about its combination with environmental changes. In the current study, we used SEM analysis to explore the direct and indirect effects of stand age on NuRE and RSAF under N and P fertilization. Generally, stand age had positive effects on PRE whether under N or P fertilization, as well as on RSAF-P under N fertilization, which indicated that plant intended to resorb more P from senescing leaves and soil by fine root to deal with strengthening P-limitation during stand development (Zhang et al., 2018; Wang et al., 2019b). Thus, these findings confirmed that stand age mediated the fertilization effects on nutrient acquisition strategies.
This study evaluates how N and P fertilization affects nutrient acquisition strategies in M. glyptostroboides plantations with different stand ages of the coastal in China. We found that N fertilization increased NRE, while decreased RSAF-P in young plantations. P fertilization increased NRE and PRE, while decreased RSAF-P in middle-aged plantations. Little synergetic relationships were observed between NuRE and RSAF whether under N and P fertilization, as well as in young and middle-aged plantations. Stand age had positive effects on PRE and RSAF-P, whereas had no effects on the NRE or RSAF-N. These findings provided new insights into the predictions for the facts that how variations in nutrient availability induced by the global change will influence plant nutrient uptake and nutrient conservation strategies.
Please confirm that the Data Availability statement is accurate. Note that we have used the statement provided at Submission. If this is not the latest version, please let us know. Note that we have used the statement provided at Submission. If this is not the latest version, please let us know.
The original contributions presented in the study are included in the article/supplementary material, further inquiries can be directed to the corresponding author/s.
TW and XY designed the project and participated in the manuscript. RS, RT, and GW wrote and revised the manuscript. RS and HZ analyzed the data and constructed the database. RS collected the samples. All authors revised and approved the manuscript.
This work was supported by the Fundamental Research Funds for National Natural Science Foundation of China (31770756).
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Abrahão, A., Ryan, M. H., Laliberté, E., Oliveira, R. S., and Lambers, H. (2018). Phosphorus- and nitrogen-acquisition strategies in two Bossiaea species (Fabaceae) along retrogressive soil chronosequences in south-western Australia. Physiol. Plant. 163, 323–343. doi: 10.1111/ppl.12704
Aerts, R. (1996). Nutrient resorption from senescing leaves of perennials: are there general patterns? J. Ecol. 84, 597–608. doi: 10.2307/2261481
Andersen, K. M., Mayor, J. R., and Turner, B. L. (2017). Plasticity in nitrogen uptake among plant species with contrasting nutrient acquisition strategies in a tropical forest. Ecology 98, 1388–1398. doi: 10.1002/ecy.1793
Brant, A. N., and Chen, H. Y. (2015). Patterns and mechanisms of nutrient resorption in plants. Crit. Rev. Plant Sci. 34, 471–486.
Chen, X., and Chen, H. Y. (2022). Foliar nutrient resorption dynamics of trembling aspen and white birch during secondary succession in the boreal forest of central Canada. For. Ecol. Manag. 505:119876.
Deng, M., Liu, L., Sun, Z., Piao, S., Ma, Y., Chen, Y., et al. (2016). Increased phosphate uptake but not resorption alleviates phosphorus deficiency induced by nitrogen deposition in temperate Larix principis-rupprechtii plantations. New Phytol. 212, 1019–1029. doi: 10.1111/nph.14083
Du, E., Terrer, C., Pellegrini, A. F., Ahlström, A., van Lissa, C. J., Zhao, X., et al. (2020). Global patterns of terrestrial nitrogen and phosphorus limitation. Nat. Geosci. 13, 221–226.
Hayes, P., Turner, B. L., Lambers, H., and Laliberté, E. (2014). Foliar nutrient concentrations and resorption efficiency in plants of contrasting nutrient-acquisition strategies along a 2-million-year dune chronosequence. J. Ecol. 102, 396–410. doi: 10.1111/1365-2745.12196
He, M., Yan, Z., Cui, X., Gong, Y., Li, K., and Han, W. (2020). Scaling the leaf nutrient resorption efficiency: nitrogen vs phosphorus in global plants. Sci. Total Environ. 729:138920. doi: 10.1016/j.scitotenv.2020.138920
Hofmann, K., Heuck, C., and Spohn, M. (2016). Phosphorus resorption by young beech trees and soil phosphatase activity as dependent on phosphorus availability. Oecologia 181, 369–379. doi: 10.1007/s00442-016-3581-x
Killingbeck, K. T. (1996). Nutrients in senesced leaves: keys to the search for potential resorption and resorption proficiency. Ecology 77, 1716–1727.
Kobe, R. K., Lepczyk, C. A., and Iyer, M. (2005). Resorption efficiency decreases with increasing green leaf nutrients in a global data set. Ecology 86, 2780–2792.
Kou, L., Jiang, L., Fu, X., Dai, X., Wang, H., and Li, S. (2018). Nitrogen deposition increases root production and turnover but slows root decomposition in Pinus elliottii plantations. New Phytol. 218, 1450–1461.
Kou, L., Wang, H., Gao, W., Chen, W., Yang, H., and Li, S. (2017). Nitrogen addition regulates tradeoff between root capture and foliar resorption of nitrogen and phosphorus in a subtropical pine plantation. Trees 31, 77–91. doi: 10.1007/s00468-016-1457-7
Li, L., Gao, X., Li, X., Lin, L., Zeng, F., Gui, D., et al. (2016). Nitrogen (N) and phosphorus (P) resorption of two dominant alpine perennial grass species in response to contrasting N and P availability. Environ. Exp. Bot. 127, 37–44. doi: 10.1016/j.envexpbot.2016.03.008
Li, L., McCormack, M. L., Chen, F., Wang, H., Ma, Z., and Guo, D. (2019). Different responses of absorptive roots and arbuscular mycorrhizal fungi to fertilization provide diverse nutrient acquisition strategies in Chinese fir. For. Ecol. Manag. 433, 64–72. doi: 10.1016/j.foreco.2018.10.055
Liu, M., Xu, F., Xu, X., Wanek, W., and Yang, X. (2018). Age alters uptake pattern of organic and inorganic nitrogen by rubber trees. Tree Physiol. 38, 1685–1693. doi: 10.1093/treephys/tpy031
Lu, J., Yang, M., Liu, M., Lu, Y., and Yang, H. (2019). Nitrogen and phosphorus fertilizations alter nitrogen, phosphorus and potassium resorption of alfalfa in the Loess Plateau of China. J. Plant Nutr. 42, 2234–2246. doi: 10.1080/01904167.2019.1648668
Ma, Z., Guo, D., Xu, X., Lu, M., Bardgett, R. D., Eissenstat, D. M., et al. (2018). Evolutionary history resolves global organization of root functional traits. Nature 555, 94–97.
Ostertag, R., and DiManno, N. M. (2016). Detecting terrestrial nutrient limitation: a global meta-analysis of foliar nutrient concentrations after fertilization. Front. Earth Sci. 4:23. doi: 10.3389/feart.2016.00023
Pan, Y., Liu, B., Cao, J., Liu, J., Tian, S., and Du, E. (2021). Enhanced atmospheric phosphorus deposition in Asia and Europe in the past two decades. Atmos. Ocean. Sci. Lett. 14:100051. doi: 10.1016/j.aosl.2021.100051
Pregitzer, K. S., DeForest, J. L., Burton, A. J., Allen, M. F., Ruess, R. W., and Hendrick, R. L. (2002). Fine root architecture of nine North American trees. Ecol. Monogr. 72, 293–309. doi: 10.2307/3100029
Reed, S. C., Townsend, A. R., Davidson, E. A., and Cleveland, C. C. (2012). Stoichiometric patterns in foliar nutrient resorption across multiple scales. New Phytol. 196, 173–180. doi: 10.1111/j.1469-8137.2012.04249.x
Sardans, J., Grau, O., Chen, H. Y., Janssens, I. A., Ciais, P., Piao, S., et al. (2017). Changes in nutrient concentrations of leaves and roots in response to global change factors. Glob. Change Biol. 23, 3849–3856. doi: 10.1111/gcb.13721
Sun, Z., Liu, L., Peng, S., Peñuelas, J., Zeng, H., and Piao, S. (2016). Age-related modulation of the nitrogen tesorption efficiency response to growth requirements and soil nitrogen availability in a temperate pine plantation. Ecosystems 19, 698–709. doi: 10.1007/s10021-016-9962-5
Ushio, M., Fujiki, Y., Hidaka, A., and Kitayama, K. (2015). Linkage of root physiology and morphology as an adaptation to soil phosphorus impoverishment in tropical montane forests. Funct. Ecol. 29, 1235–1245. doi: 10.1111/1365-2435.12424
Vergutz, L., Manzoni, S., Porporato, A., Novai, R. F., and Jackson, R. B. (2012). Global resorption efficiencies and concentrations of carbon and nutrients in leaves of terrestrial plants. Ecol. Monogr. 82, 205–220. doi: 10.1890/11-0416.1
Wang, F., Fang, X., Wang, G. G., Mao, R., Lin, X., Wang, H., et al. (2019a). Effects of nutrient addition on foliar phosphorus fractions and their resorption in different-aged leaves of Chinese fir in subtropical China. Plant Soil 443, 41–54. doi: 10.1007/s11104-019-04221-8
Wang, J., Wang, J., Wang, L., Zhang, H., Guo, Z., Wang, G. G., et al. (2019b). Does stoichiometric homeostasis differ among tree organs and with tree age? For. Ecol. Manag. 453:117637. doi: 10.1016/j.foreco.2019.117637
Wang, J., Wang, J., Guo, W., Li, Y., Wang, G. G., and Wu, T. (2018). Stoichiometric homeostasis, physiology, and growth responses of three tree species to nitrogen and phosphorus addition. Trees 32, 1377–1386. doi: 10.1007/s00468-018-1719-7
Wang, L., Li, Y., Duan, Y., Lian, J., Luo, Y., Wang, X., et al. (2020). Effects of nitrogen addition and reproductive effort on nutrient resorption of a sand-fixing shrub. Front. Plant Sci. 11:588865. doi: 10.3389/fpls.2020.588865
Wang, M., Murphy, M. T., and Moore, T. R. (2014a). Nutrient resorption of two evergreen shrubs in response to long-term fertilization in a bog. Oecologia 174, 365–377. doi: 10.1007/s00442-013-2784-7
Wang, Z., Lu, J., Yang, H., Zhang, X., Luo, C., and Zhao, Y. (2014b). Resorption of nitrogen, phosphorus and potassium from leaves of lucerne stands of different ages. Plant Soil 383, 301–312. doi: 10.1007/s11104-014-2166-x
Wen, Y., Tong, R., Zhang, H., Feng, K., Song, R., Wang, G. G., et al. (2021). N addition decreased stand structure diversity in young but increased in middle-aged Metasequoia glyptostroboides plantations. Glob. Ecol. Conserv. 30:e01803. doi: 10.1016/j.gecco.2021.e01803
Widdig, M., Schleuss, P.-M., Weig, A. R., Guhr, A., Biederman, L. A., Borer, E. T., et al. (2019). Nitrogen and phosphorus additions alter the abundance of phosphorus-solubilizing bacteria and phosphatase activity in grassland soils. Front. Environ. Sci. 7:185. doi: 10.3389/fenvs.2019.00185
Wright, I. J., and Westoby, M. (2003). Nutrient concentration, resorption and lifespan: leaf traits of Australian sclerophyll species. Funct. Ecol. 17, 10–19. doi: 10.1046/j.1365-2435.2003.00694.x
Xu, M., Zhong, Z., Sun, Z., Han, X., Ren, C., and Yang, G. (2020). Soil available phosphorus and moisture drive nutrient resorption patterns in plantations on the Loess Plateau. For. Ecol. Manag. 461:117910. doi: 10.1016/j.foreco.2020.117910
Yan, Z., Kim, N., Han, W., Guo, Y., Han, T., Du, E., et al. (2015). Effects of nitrogen and phosphorus supply on growth rate, leaf stoichiometry, and nutrient resorption of Arabidopsis thaliana. Plant Soil 388, 147–155. doi: 10.1007/s11104-014-2316-1
Yuan, Z. Y., and Chen, H. Y. H. (2015). Negative effects of fertilization on plant nutrient resorption. Ecolgy 96, 373–380. doi: 10.1890/14-0140.1
Zhang, H., Wang, J., Wang, J., Guo, Z., Wang, G. G., Zeng, D., et al. (2018). Tree stoichiometry and nutrient resorption along a chronosequence of Metasequoia glyptostroboides forests in coastal China. For. Ecol. Manag. 430, 445–450. doi: 10.1016/j.foreco.2018.08.037
Keywords: nutrient acquisition strategies, N and P fertilization, stand age, leaf, root
Citation: Song R, Tong R, Zhang H, Wang GG, Wu T and Yang X (2022) Effects of Long-Term Fertilization and Stand Age on Root Nutrient Acquisition and Leaf Nutrient Resorption of Metasequoia glyptostroboides. Front. Plant Sci. 13:905358. doi: 10.3389/fpls.2022.905358
Received: 27 March 2022; Accepted: 25 April 2022;
Published: 11 May 2022.
Edited by:
Runguo Zang, Chinese Academy of Forestry, ChinaReviewed by:
Fu-Sheng Chen, Jiangxi Agricultural University, ChinaCopyright © 2022 Song, Tong, Zhang, Wang, Wu and Yang. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Tonggui Wu, d3V0b25nZ3VpQGNhZi5hYy5jbg==; Xiuqing Yang, eGl1cWluZ3kyMDAyQDEyNi5jb20=
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.