
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
REVIEW article
Front. Plant Sci. , 16 June 2022
Sec. Plant Breeding
Volume 13 - 2022 | https://doi.org/10.3389/fpls.2022.886162
This article is part of the Research Topic Insights in Plant Breeding: 2021 View all 6 articles
 Juan Pablo Renzi1,2*
Juan Pablo Renzi1,2* Clarice J. Coyne3
Clarice J. Coyne3 Jens Berger4
Jens Berger4 Eric von Wettberg5,6
Eric von Wettberg5,6 Matthew Nelson4,7Soledad Ureta2
Matthew Nelson4,7Soledad Ureta2 Fernando Hernández2
Fernando Hernández2 Petr Smýkal8*
Petr Smýkal8* Jan Brus9*
Jan Brus9*Alongside the use of fertilizer and chemical control of weeds, pests, and diseases modern breeding has been very successful in generating cultivars that have increased agricultural production several fold in favorable environments. These typically homogeneous cultivars (either homozygous inbreds or hybrids derived from inbred parents) are bred under optimal field conditions and perform well when there is sufficient water and nutrients. However, such optimal conditions are rare globally; indeed, a large proportion of arable land could be considered marginal for agricultural production. Marginal agricultural land typically has poor fertility and/or shallow soil depth, is subject to soil erosion, and often occurs in semi-arid or saline environments. Moreover, these marginal environments are expected to expand with ongoing climate change and progressive degradation of soil and water resources globally. Crop wild relatives (CWRs), most often used in breeding as sources of biotic resistance, often also possess traits adapting them to marginal environments. Wild progenitors have been selected over the course of their evolutionary history to maintain their fitness under a diverse range of stresses. Conversely, modern breeding for broad adaptation has reduced genetic diversity and increased genetic vulnerability to biotic and abiotic challenges. There is potential to exploit genetic heterogeneity, as opposed to genetic uniformity, in breeding for the utilization of marginal lands. This review discusses the adaptive traits that could improve the performance of cultivars in marginal environments and breeding strategies to deploy them.
When coupled with the use of irrigation, fertilizers and pesticides, modern breeding has increased agricultural production several fold in favorable environments (Evenson and Gollin, 2003). However, such high-input agriculture also has a high environmental impact. High performing cultivars are designed under good field conditions and perform well when there is sufficient water and nutrients, e.g., fertile soil. However, such conditions are often not available because there is a large proportion of arable land in marginal environments globally (Sebby, 2010; Pingali, 2012). Marginal land exists worldwide (e.g., Kang et al., 2013), and typically, it is of poor soil fertility. In many areas these soils are prone to erosion, with shallow soil layers, and may be prone to drought, salinity, or other abiotic challenges. Moreover, these environments are expected to expand with ongoing climate change and increasing degradation of soil and water resources (El-Beltagy and Madkour, 2012).
Throughout evolutionary history, plants have had to adapt to a range of environmental conditions including adverse habitats. As a consequence, plant species including crop wild relatives typically exhibit a high level of genetic and phenotypic variation within their distribution range (Anderson et al., 2011; Russell et al., 2014; Smýkal et al., 2017; Hellwig et al., 2021). Understanding the genetic, biological and ecological bases of local adaptation is relevant to manage the impact of climate change on crop and animal production. In widely distributed plant species inhabiting different environments, ecotypic differentiation has been detected for populations along altitudinal, latitudinal or environmental gradients (e.g., Polechová and Barton, 2015; Berger et al., 2017). Understanding the extent to which such distinct ecotypes are formed will help not only to reveal and recognize their evolutionary adaptive patterns, but also will help breeders to use their diversity to maximize productivity under target environments.
It is widely accepted that past domestication processes have resulted in a narrower genetic base of cultivated germplasm that is prone to pests and diseases (Tanksley and McCouch, 1997; McCouch et al., 2020; Khoury et al., 2022). Unlike their domesticated crops, crop wild relatives (CWRs) continued to evolve to adapt to their environments and thus provide a key resource to counteract the effects of climate change on the world’s food supply. To fully understand and utilize this process, studies in the geographical centers of origin are needed combining ecology, physiology and genetics. This approach was pioneered by N.I. Vavilov in the 1920s–1930s starting with surveys, collecting missions and subsequent evaluation of germplasm collections in diverse environmental settings (Vavilov, 1957). Today, with the available tools of genomics, geospatial analysis and modeling combined with improved phenotyping methods, it has become all the more important to analyze wild accessions and use them as sources of stress tolerance for breeding purposes (Warschefsky et al., 2014; Coyne et al., 2020). Wild species have been used mainly for the introgression of disease and insect resistance into crops (e.g., Hajjar and Hodgkin, 2007); drought, cold, heat and salinity tolerance have also been addressed in some staple crops (Prohens et al., 2017). Biotic and abiotic stress resistances have been explored in wild relatives of many crops including chickpea, barley and maize (Brozynska et al., 2016; Fustier et al., 2017; von Wettberg et al., 2018; Liu et al., 2020). A complementary approach is the de novo domestication of new crops (Smýkal et al., 2018; Fernie and Yan, 2019).
Environmental fluctuations such as soil salinity, cold and drought stress represent a major constraint on agricultural productivity. Water availability and temperature extremes are the major factors controlling the distribution of vegetation over the earth’s surface. Crop yields are more dependent on an adequate supply of water than on any other single factor; environmental stress represents the primary cause of crop losses. With current climate change projections, extremely hot weather will become more frequent and rainfall will be more erratic in many regions of the world (Foyer et al., 2016). As many elite cultivars tend to be drought sensitive, ensuring food security will require development of more drought resistant varieties. Dissection of adaptation mechanisms to harsh conditions naturally occurring in wild relatives of domesticated crops may provide a solid foundation for crop improvement under the changing climate. Drought and heat stresses impact every plant developmental stage, starting from seed germination, through vegetative growth. However, the most adverse effects (from a crop production perspective) are on crop establishment (germination and young seedling phase) and reproduction, which are very sensitive to suboptimal conditions. Small seedlings and pollen grains happen to be particularly sensitive to lack of water and excess heat (Dürr et al., 2015; Yu et al., 2019; Chaturvedi et al., 2021; Chen et al., 2021).
The term “marginal” land originated in the field of agricultural economics during the 19th century (Kang et al., 2013) by Ricardo (2005) using the categorization from his land rent theory which was the basis of marginal productivity theory. Marginal lands are subject to socio-economic and biophysical constraints, and lower productivity. Nevertheless, it is difficult to define “productivity” within this concept because this varies depending on land use. For example, land that is too marginal for crop production might be suitable for grazing, and “fragile” land can be susceptible to soil degradation, but it may still be suitable for sustainable forestry. Many dry-land countries have not assessed the extent or characteristics of these lands, nor their sustainability as biofuels or food crops production. These lands are often difficult to exploit economically and sustainably for agriculture, and can lead to abandonment (Campbell et al., 2008). Land can also be degraded due to an intensive and unsustainable use (Campbell et al., 2008; Dauber et al., 2010). Marginal lands are areas with low rainfall, high temperatures, low quality soils, steep terrain, shallow soil depth (less than 50 cm), poor fertility, coarse texture, stony, heavy cracked clays, salt-affected lands, waterlogged, marshy land, barren and rocky soils, and other problems. If a land is considered marginal it means that it does not have enough capacity to produce food, or other agricultural activities.
Nearly one billion people live in poverty today, a high percentage living in these marginal lands in mostly developing countries (Ahmadzai et al., 2021). These areas with low soil fertility and limited agricultural value are also highly vulnerable to climate change (Castro and Castro, 2019). The research concluded that in total 29% of the agricultural area is marginal in European Union. The most common reason is the rooting limitations; 12% of Europe’s agricultural area required intervention to improve soil structure and depth to allow farming. This is followed by adverse climate and excessive soil moisture occurring in respectively 11 and 8% of the agricultural land (Elbersen et al., 2018). Within the global scale marginal lands in developed world covers about 9% of terrestrial ecosystems (Václavík et al., 2013). These lands occur predominantly in Western United States, Australia, Argentina, but also in North and South Africa (Figure 1). A more precise estimation of area covered by marginal lands is limited by uncertainties and problems of land cover classification (Brus et al., 2018). Marginal lands are most vulnerable to climate events like drought and floods, thus, most likely to be affected by climate change. Climate change has made it difficult to maintain established farming practices in recent decades. Within the US the prevalence of local yield deficits confirms that individual farmers looking to expand their operations are generally confined to cultivating increasingly marginal land, though widespread variation exists from location to location (Lark et al., 2020).
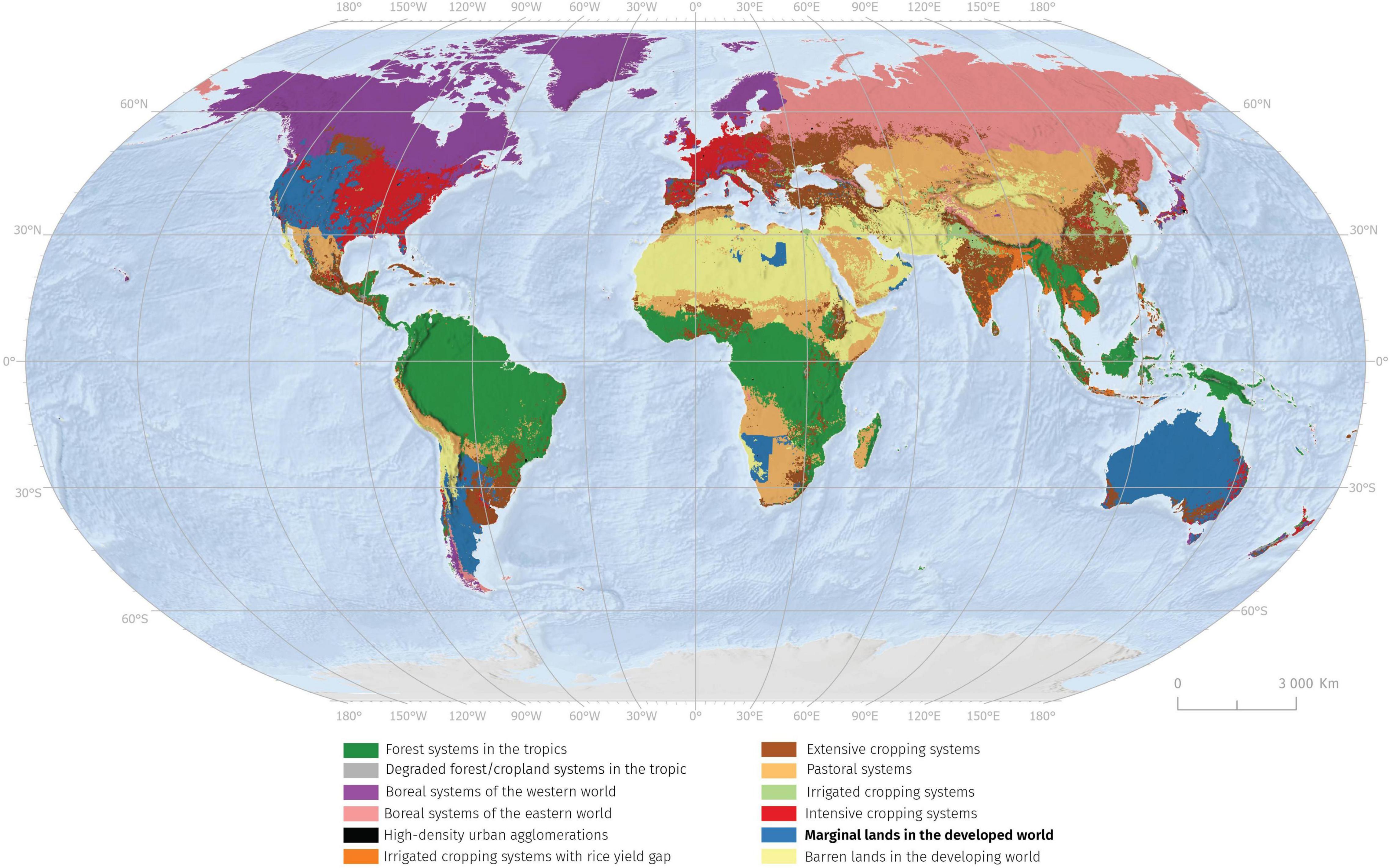
Figure 1. Map shows global land system archetypes classification by Václavík et al. (2013). Within the global scale, marginal lands in the developed world cover about 9% of terrestrial ecosystems.
The effects of climate change are manifested as erratic precipitation and weather changes, which have led to rapid fluctuations and unpredictable temperatures. Adaptation to this requires new approaches to agriculture. For example, research on agro-climatic conditions has shown that farmers in dryland areas are more aware of climate change impacts than those in wetland areas. Similar findings have also been reported for Ecuador (Skarbø and VanderMolen, 2014). It is unclear, however, if agricultural producers can keep up with climate change at the incredible pace it is predicted to occur in coming years (Jones et al., 2012). Adaptation to climate change is difficult and impacts are based on various assumptions. In addition to estimating future impacts, researchers need to make subjective value judgments. This is especially true of social and economic development in marginal regions. As a result, the effects of climate change on these areas are difficult to forecast. Although the impacts of climate change are not always measurable, they are still a huge concern for governments and local communities alike. Some areas of the world are particularly vulnerable, and these risks can be mitigated by developing new approaches and technologies in crop production. One such approach is the implementation of local and regional adaptation plans. However, it is crucial to consider the broader context of marginal environments when developing policies and adaptation strategies for agricultural producers. This research is particularly important in Latin America, where smallholder farmers are heavily impacted by climate change. Moreover, rural and subsistence farming are often location-specific systems, which are highly vulnerable to the stressors of the climate. As a result, the research needs to take this into account. Agro-ecosystems are critical for food production and sustainability, but they are also prone to pests. Consequently, many current crop varieties will have to be replaced as a result of climate change. Failure to address these challenges will have a profound impact on the global economy and on social well-being. If we fail to protect and restore natural ecosystems, it will become impossible to grow crops in these marginal areas in the future. One of the possible way how to cope with this situation is implementation of Climate-Smart Agriculture (CSA) (Lipper et al., 2014).
Crop wild relatives typically harbor far greater genetic diversity than modern elite crops as a result of primary and secondary domestication bottlenecks; the founder effect(s) and subsequent fixation of domestication syndrome and post-domestication traits including cultural selection for local preference (Tanksley and McCouch, 1997; Purugganan and Fuller, 2009). Moreover, the evolutionary trajectory and resultant selection pressures encountered by wild progenitors and elite cultivars are very different. While we can now deploy an impressive array of technical and analytical approaches for handling complexity in the search for adaptive genes (Cortes and Lopez-Hernandez, 2021), here we argue that understanding the divergent selection pressures encountered by wild progenitor and elite cultivar adds invaluable context for the use of CWR in improving crop adaptation.
Wild progenitors are subject to natural selection in response to environmental stressors, herbivory, pathogen attack and inter-plant competition from the same or other species occupying similar ecological niches in a variable climate. While viable seed production is the ultimate measure of ‘fitness,’ reproductive investment in CWR is usually lower than in their domesticated counterparts (Berger et al., 2017; Berger J. et al., 2020; Garibaldi et al., 2021). This implies that competition for resources such as light, nutrients or water is selecting for greater vegetative investment (roots, shoots, leaves) in CWR relative to the domesticated crop, akin to the competitors in Grime’s triangle (Grime, 1974). In some examples the genomic consequences of this divergent selection for reproductive versus vegetative investment have been identified, such as the selection for increased apical dominance (e.g., reduced branching) in the domestication of maize (Clark et al., 2004). Seed dormancy, a key domestication syndrome trait, can both exacerbate and ameliorate seasonal variability in CWR. In many Mediterranean CWR seed physical dormancy has been selected as bet-hedging against false breaks (heavy rainfall that precedes the growing season start and which would expose the recently germinated seedling to extreme heat and drought) and other disturbances such as catastrophic herbivory (Piano et al., 1996; Norman et al., 2002; Smýkal et al., 2014; Berger et al., 2017). Typically, there is wide ranging dormancy within populations (Berger et al., 2017; Hradilová et al., 2019; Renzi et al., 2019), that in combination with variable seed placement will drive staggered germination across the CWR population over the growing season, exposing individuals to different micro-habitats in space and time.
Conversely, crop plants are subject to a combination of natural and artificial selection giving rise to the domestication syndrome (large seed size, loss of dormancy, and seed dehiscence etc.) and subsequent selection for local adaptation and food preferences (Fuller and Allaby, 2009; Purugganan and Fuller, 2009; Smýkal et al., 2018). Compared to CWR, modern crops lead a strictly regulated life in a tightly defined growing season. Non-dormant seeds are accurately positioned with respect to soil depth and neighbor distance in a prepared seedbed during an optimal window, matching phenology to target environment to minimize frost, drought, and other stress. Ripening is both uniform and timely because of the use of determinate cultivars (e.g., cereals), selection for earliness (e.g., chickpea, lupin, canola), or the application of desiccants (e.g., canola, chickpea). Herbivory and diseases are minimized by management interventions. Importantly, crops are selected on the basis of communal, rather than individual yield, whereas the opposite is the case for CWR. As a result, modern crops tend to be weak-moderate competitors (Donald, 1981; Weiner et al., 2017) with a greater reproductive investment than their wild progenitors (Berger et al., 2017; Berger J. et al., 2020; Garibaldi et al., 2021).
Finally, domestication and subsequent global dispersal further widens the evolutionary dichotomy between crop and CWR (Purugganan and Fuller, 2009). Chickpea represents an extreme exemplar, having been domesticated from a very narrowly distributed southeastern Anatolian wild progenitor with a Mediterranean winter annual lifecycle into a global crop grown on all continents except Antarctica. It has been hypothesized to have returned to the Mediterranean as a spring-sown late season crop after a 2000 years archeological gap (Abbo et al., 2003). The much more recently domesticated narrow-leafed lupin (L. angustifolius) and yellow lupin (L. luteus) represent the opposite extreme. Here the wild progenitors range widely around the Mediterranean basin (particularly L. angustifolius), being domesticated in the 18th century as spring-sown crops for limited sandy acid soil regions in central and eastern Europe (Hondelmann, 1984), and then returning to Mediterranean climates in the southern hemisphere in the last 50 years (Gladstones, 1994). The combination of divergent genetic diversity and evolutionary trajectory/selection pressures leads to different adaptive traits in crops and their wild relatives.
The regulation of phenology in Mediterranean crops and wild relatives provides clear examples of divergent evolutionary paths. For example, vernalization plays an important role in regulating phenology in both wild Cicer and Lupinus, presumably as a consequence of the cool-cold winters and rapidly warming spring temperatures experienced in their Mediterranean distribution range. Wild narrow-leafed and yellow lupins are widespread species in which flowering time and associated traits including water use, biomass production and reproductive investment are strongly linked to rainfall at origin (Berger and Ludwig, 2014; Berger et al., 2017; Berger J. D. et al., 2020). Despite the importance of phenology in ecotypic adaptation of wild lupins, there is very little variation in vernalization response (Taylor et al., 2019). Similarly, most wild Cicer species also appear to be consistently vernalization responsive (Berger et al., 2005), suggesting that other regulatory mechanisms such as photoperiod or temperature responses or earliness per se are responsible for the phenological differences observed across wild germplasm collections. Indeed, wild Cicer species appear to be extremely photoperiod responsive (Sharma and Upadhyaya, 2019) and investigations are ongoing to reveal interactions between all three phenology regulators (vernalization, photoperiod, temperature) in order to understand the regulation of flowering time and what CWRs might bring to the crop.
Conversely, downregulation of the vernalization response has played a key role in the domestication of chickpea and both lupin species as these crops moved away from their cool-cold winter Mediterranean origins. Selection for earliness and high early vigor were key breeding criteria in both lupin species in their domestication as spring-sown crops because timely maturity in the cooling late central-eastern European summer was essential to reliable seed production (Hondelmann, 1984). When the crops moved to relatively warm-winter Mediterranean climates in Australia, where vernalization is a mostly unreliable regulator of phenology, it was necessary to identify unresponsive mutants to start the industry effectively (Gladstones and Hill, 1969). Now, both Australian and European Lupin breeding is dominated by the vernalization unresponsive FT alleles (Taylor et al., 2019, 2021). In chickpea vernalization was presumably downregulated in the Bronze Age introduction of the crop to South Asia and its hypothesized return as a Mediterranean spring-sown crop (Abbo et al., 2002, 2003; Redden and Berger, 2007). Unlike lupin, the much older chickpea crop has formed distinct ecotypes with different flowering regulatory mechanisms appropriate to each region (Berger et al., 2011). Thus, chickpea landraces become increasingly temperature responsive from the Mediterranean through north, central and southern India, temperature response being strongly correlated to collection site temperature during the vegetative phase (Berger et al., 2011). Among Eastern Mediterranean germplasm photoperiod and temperature response were negatively correlated, a relationship that is likely to have been essential in the colonization of warmer climates in South Asia where a strong photoperiod responsive is maladaptive, particularly in Southern India where chickpea flowers under declining day length.
Divergence in adaptive strategies between crops and wild relatives provides opportunities to modify crop adaptation to new and marginal environments, while understanding which traits are selected for what regions adds useful context to the problem. For example, genes from chickpea wild relatives would be invaluable in the creation of a winter cultivar because these appear to have been lost in evolution of the crop. Indeed, chickpea wild relatives are more winter hardy than domesticated chickpea (Singh et al., 1990, 1994) and appear to have greater reproductive chilling tolerance as well (Berger et al., 2012b). Efforts are currently underway to improve cold tolerance in chickpea using wild relatives. In narrow-leafed lupin the strong selection for earliness has created a bottleneck which is limiting yield potential in higher rainfall environments where longer season cultivars will be better adapted (Berger et al., 2012a; Chen et al., 2017). As a result, considerable effort is being invested in the search for other regulatory levers with which to delay flowering in lupin without re-introducing an obligatory vernalization requirement (Taylor et al., 2019, 2021). Wild germplasm is playing an important role in this. These examples stand out because phenology has played such a central role in the divergent evolution of crop and wild relative in both chickpea and lupin.
When the differences between crop and wild relative are more obscure, it becomes more difficult to generalize about the adaptive potential that may be exploited from CWR. Drought stress often occurs in marginal environments as described earlier. Among annual plants the principal adaptation strategy is stress avoidance through appropriate phenology, with some very limited capacity for drought postponement through water acquisition (e.g., deep roots) and conservative water use (e.g., stomatal closure), and very short-term tolerance through osmotic adjustment (Ludlow and Muchow, 1990; Berger et al., 2016). Given the widespread selection for earliness in domesticated crops compared to their wild relatives, it is difficult to argue that CWR will improve the capacity for drought avoidance in our crops. However, CWR may be a useful source of drought postponement or tolerance traits, given that these are likely to be under selection in wild populations where individuals compete amongst themselves and often also have long, indeterminate lifecycles which include periods of transient drought stress (Berger et al., 2016). Lysimetry has demonstrated that the wild Cicer species, C. echinospermum and C. reticulatum extract more water both under terminal drought and well-watered conditions than does domestic chickpea (Berger J. et al., 2020). At this stage the underlying mechanism is unknown. If the greater water use extraction of wild Cicer is explained by greater vegetative investment and associated lower water-use efficiency, it will be maladaptive in the chickpea crop; if it represents a capacity to tolerate lower water potential, however, it may be useful. Wild L. luteus has a greater capacity to maintain leaf water content at low water potential than the domesticated crop, probably through osmotic adjustment (Berger and Ludwig, 2014; Berger J. D. et al., 2020). Surprisingly this capacity was only found in high, rather than low rainfall ecotypes and was suggested as a bet-hedging strategy to postpone self-induced transient drought caused by very high water-use of high rainfall ecotypes. Currently, there are not enough ecophysiological studies of annual adaptive strategies over contrasting climates to generalize about the likelihood of finding these sorts of adaptive traits among CWR. There has been no evidence for similar drought stress tolerance on low and high rainfall ecotypes of wild L. angustifolius (Berger J. D. et al., 2020), Conversely Yousfi et al. (2010) detected greater osmotic adjustment capacity in low compared to high rainfall ecotypes of Medicago truncatula and M. laciniata. We hope that future germplasm screening efforts used in crop improvement will address this shortcoming by studying material from contrasting, well characterized collection sites.
Selection attributes according to the crop objective are related to classical domestication traits, such as reduced seed and fruit dispersal, changes in plant structure, plant phenology, seed dormancy, palatability, or acquisition of modified fruit size and shape in vegetable crops (Alonso-Blanco et al., 2009; Sakuma et al., 2011; Meyer et al., 2012; Zohary et al., 2012; Abbo et al., 2014; Fuller and Allaby, 2018; Ogutcen et al., 2018; Smýkal et al., 2018; Iqbal et al., 2020). Much progress has been made in the most widely grown and intensively selected crops (rice, wheat, soybean, sugarcane, tomato and potato), but improvement using CWR is more recently (Hajjar and Hodgkin, 2007; Shelef et al., 2017; Zhang et al., 2018; Zsögön et al., 2018; Fernie and Yan, 2019). CWRs provide unique reservoirs of genetic diversity for crop improvement (Cowling et al., 2009; Redden, 2013; Matesanz et al., 2020; Bohra et al., 2021). They have typically been used in breeding to introgress a small number of major genes (for insect/disease-resistant varieties), in contrast to adaptation that is controlled by many small-effect quantitative trait loci and complex interactions which are more difficult to harness (Hajjar and Hodgkin, 2007; Zhang et al., 2018; Zsögön et al., 2018). The introgression of adaptive diversity from CWR to domesticated crops provides breeders with new tools for crop improvement (Smýkal et al., 2018). As environmental changes associated with domestication should favor resource-acquisition strategies compared with the resource-conservation strategies of wild relatives (Redden, 2013; von Wettberg et al., 2014; Vilela and González-Paleo, 2015), breeders are faced with the challenge of a trade-off between productivity and adaptation. The development of a variety with high adaptation (survival) under a marginal environment, but very low productivity would not be desirable (Richards, 2006; Berger et al., 2016).
Cultivars with tolerance to drought, heat, salinity, soils with extreme pH (alkaline or acidic) and low fertility, as well as various biotic factors are known to be required in marginal environments (Redden, 2013; Quezada-Martinez et al., 2021). Irrespective of the pre-breeding technique it is important to understand the phenotypic traits that confer adaptation for identification in any given marginal environment and for agronomic management (Wilke and Snapp, 2008). Appropriate phenotypic traits (morphological, anatomical, and phenological) that are easily distinguishable and inexpensive to measure in the field, suitable for pre-breeding programs in developing countries, where marginal environments predominate, could be considered. However, the breeding efficiency might be low (Araus and Kefauver, 2018). On the contrary, adaptation to salinity through the evaluation of osmoprotectants, reactive oxygen scavengers (ROS), stress proteins and ion/proton transporters, or the resistance to diseases-pests not strongly associated with specific phenotypic traits, requires more complex screening and modern techniques (Araújo et al., 2015). Since many CWR can help stabilize and increase productivity in marginal agroecosystems, the phenotypic traits that provide these and other types of services of candidates for domestication should be described. Table 1 presents some phenotypic traits that are related to the plant responses to marginal environmental factors. These traits can guide breeders who are seeking new germplasm for domestication.
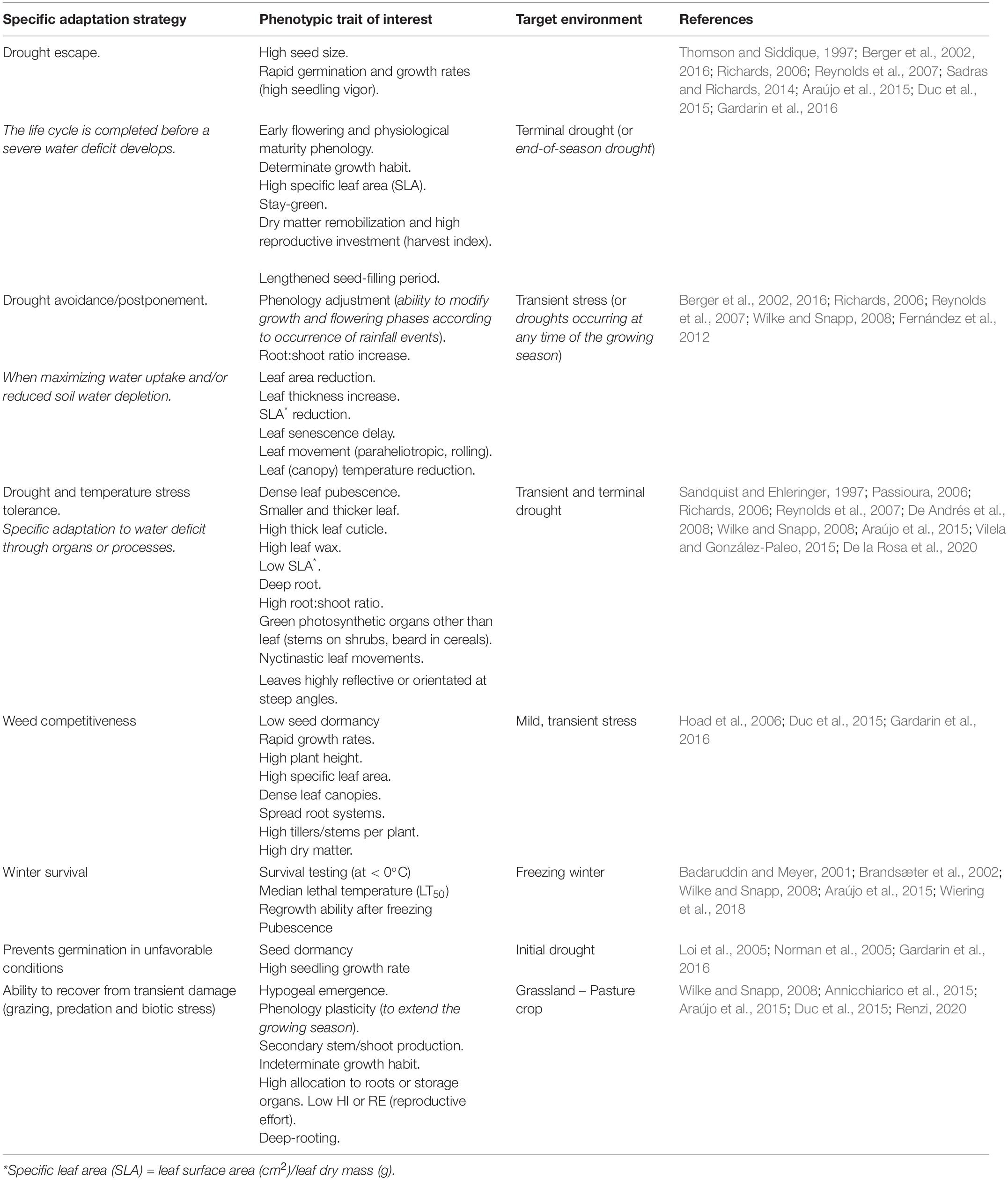
Table 1. Linking phenotypic traits and adaptation strategies that could be used as pre-breeding criteria in a population improvement program in grain and pasture crops.
Note that some traits are not directly related to yield, or even appear to be indirectly associated, but will favor adaptation in a marginal environment in the long term (Duc et al., 2015; Vilela and González-Paleo, 2015). For example, it is known that the use of high reproductive biomass (high harvest index) as a key selection criterion leads to increased wheat yield (Reynolds et al., 2007). However, in marginal environments in the northern Patagonian region of Argentina, it may be preferable to select high plant height and late-cycle wheat to cope with transient stress while maintaining the yield ceiling (∼ 1–1.5 t ha–1). Tall crops could favor competition with weeds, and the late-season genotype could prolong the duration of root growth, allowing the utilization of water and nutrients deeper in the soil profile (Richards, 2006). In addition, higher post-harvest residues with firmer stubble could increase soil cover, favoring soil moisture retention and avoid erosion processes that frequently occur in bare soils (Silenzi et al., 2012). Often remnant vegetative biomass is used as forage or for fuel. This is one of the reasons why traditional local varieties are still used instead of modern ones in this marginal environment (Cantamutto et al., 2016). In the case of forage legumes, the simple selection of a major trait (biomass) with four adaptive traits was useful for improving a semi-domesticated species such as hairy vetch (Vicia villosa) (Figure 2). The adaptive traits were late phenology, aboveground morphology (high plant branching), dense leaf pubescence and high seed dormancy. The long period of vegetative production was associated with the ability to modify the growth and flowering phases according to the occurrence of rainfall events. Successful adoption by farmers has occurred with the cultivar Patagonia INTA, with a high biomass and regrowth capacity after grazing, tolerance to transient drought, resistance to cold, and adequate natural reseeding (Renzi, 2020). The lesson is that adaptive traits have to be considered with the target environment and end user in mind; seed yield may not always be the dominant criterion.

Figure 2. Seed collection (A), plant breeding (B), multiplication (C), and agronomic use (D) of the improved cultivar of hairy vetch (Vicia villosa) cv. Patagonia INTA as forage and cover crop.
A less common but promising approach is the domestication of perennial grain crops. Traditional systems relying on annual crops have substantial negative impacts on ecosystem functions (i.e., nutrient cycling, water quality and carbon emissions, salinity, soil erosion and degradation) (Tilman et al., 2011). These problems can be reduced through the introduction of perennial crops in some environments (González-Paleo and Ravetta, 2015). This is illustrated by perennial Silphium oilseed (Heliantheae), with ideotypes with desirable traits for improving long-term yields and ecosystem services (Runck et al., 2014; Van Tassel et al., 2017). Kernza, a large-grained variety of intermediate wheatgrass (Thinopyrum intermedium) (DeHaan et al., 2020), is similar. The introduction of greater diversity in cropping systems through biotypes for mixed cultivation (“ideomixes”), with high resource-use complementarity, is also used as a novel approach by breeders (Litrico and Violle, 2015). For forage purposes, the domestication of shrub-tree legumes for livestock production has the potential not only to enhance productivity, but it could also provide multiple environmental benefits, such as providing shade for cattle, restoring degraded lands and mitigating greenhouse gas (GHG) emissions (Muir et al., 2018). The concept of the traditional ideotype in marginal environments needs to be discussed, and the ecosystem services of these should be considered (Duc et al., 2015). While introduction of perennial crops may not suit every marginal environment (such as those that experience extended hot and dry summer seasons), more research effort should be directed toward exploring the potential fit of perennial crops to different marginal environments.
Different drought scenarios (opportunity, extent, intensity and frequency of drought) and heat-stress are the most common and important stress factors in marginal lands (Chapman et al., 2000; Reynolds et al., 2007; Berger et al., 2016; Sehgal et al., 2018). This combination of changes in temperature and rainfall patterns is driving the development of drought-adapted crops. Table 1 details the different adaptive strategies to drought and the associated phenotypic traits; however, the integration of traits into an adaptive strategy that explains how species or genotypes address drought stress should be analyzed in a specific context that considers environmental selection pressures.
The phenotypic traits between drought postponement and tolerance may be overlapping in the same directions (co-gradient variation). Plant adjustments under stress conditions (plasticity) as opposed to tolerance as a constitutive trait of the species independent of water and/or heat stress are considered. Both the genetic differentiation in phenotypic traits and the plasticity of those traits are complementary mechanisms contributing to plant adaptation to environmental resource fluctuation (Matesanz et al., 2020). Uniform-rich habitats or consistently stressful environments (homogeneous and predictable low-resource environments) promote genetic differentiation rather than plasticity. On the contrary, unpredictable environments can lead to increased phenotypic plasticity (Vilela and González-Paleo, 2015; Espeland et al., 2018). Thus, the plasticity could represent most of the phenotype variation in some traits (phenology, leaf morphology) when compared with the genetic contribution; it has been suggested that it could be beneficial under climate change scenarios (Nicotra et al., 2010). In order to detect “plastic ideotypes” and exploit the plasticity in pre-breeding programs, a wide range of environments in order to fit models of phenotype versus environment should be included (de Felipe and Alvarez Prado, 2021).
In this context, CWR show high genetic variation for adaptation and may represent potential candidates for introduction into marginal areas. In addition to adaptation to different stress conditions, the ability to fix atmospheric nitrogen (N) is important in legumes and should be considered (Kebede, 2021). The percentage of nodulated plants and the number of nodules per plant are associated with N fixation in some species (Provorov and Tikhonovich, 2003). For pre-breeding purposes, the qualitative plant vigor rating, biomass, and the N measurements from non-destructive samples collected during flowering or maturity can be used (Muller et al., 2021). As the plant-associated microbes are closely linked, abiotic stresses can affect either the host plant, the Rhizobium, or the Rhizobium-legume symbioses, decreasing the N-fixing activity (Zahran, 1999). Marginal environments harbor and select diverse microbial communities, and the isolation of effective rhizobia from wild legumes for inoculating other legume crops could be an effective strategy (Ayangbenro and Babalola, 2020). Screening for tolerant N-fixing bacteria strains, as in other growth-promoting (PGP) organisms, should be considered in the selection of plant species/cultivar-specific microbial populations (Coba de la Peña and Pueyo, 2011).
Stress tolerant wild plants, such as halophytes, secretohalophytes, xerophytes and thermophytes, with a suite of desired features (food, economical or ecological) represent useful germplasm (Zhang et al., 2018), and the ecological niche models can be used to predict the adaptation in a target marginal environment (Phillips et al., 2006; Redden, 2013). Understanding the adaptive strategy simplifies crop improvement by allowing breeders to introgress new traits through traditional breeding methods or modern selection techniques (Berger et al., 2016). Likewise, knowledge of phenotypic traits in wild species would estimate the probability of adaptation a priori, and the global plant functional trait databases may be used as a screening tool (Kattge et al., 2011; Saatkamp et al., 2019).
Domestication could result in the loss of potentially useful “wild traits” that confer biotic and abiotic stress tolerance or specialized resource-use strategies (Van Tassel et al., 2017; Nagel et al., 2019). Pizza et al. (2021) showed that in Clarkia pulchella, after the eighth generation of farm cultivation, the relative fitness of the wild plants was significantly greater than the farmed plants, especially under drought stress. Genetic diversity lost over generations may be mitigated by growing out a large number of unrelated individuals (Basey et al., 2015). On the other hand, obtaining an ideal cultivar for a marginal environment can take many years of breeding resulting in abandoned programs due to low financial resources (Hajjar and Hodgkin, 2007; Muir et al., 2014; Fernie and Yan, 2019). Demographic modeling, especially in adapted pasture biotypes due to the predominance of livestock systems in marginal environments, can be used to advance their use in real production systems, until getting an advanced cultivar.
Adapted biotypes, which are not necessarily domesticated, can be very difficult to cultivate. Poor agronomic performance introduces numerous practical issues for breeders, such as difficulty in recovering a sufficient number of seeds and not being able to use standard equipment (Nutt and Loi, 1999; Nichols et al., 2012). For fodder production or restoration purposes, these traits can be used for the dispersal of desirable biotypes through demographic modeling. Dynamic population models have proven to be very useful for understanding survival and dispersal strategies of wild and semi-domesticated species. This approach is widely documented for reducing survival in weed species under different production systems (González-Andujar and Fernández-Quintanilla, 1993, 2004; Forcella et al., 2000; Colbach et al., 2005; Holst et al., 2007; Gardarin et al., 2012), as well as promoting it in legume and grass pastures (Taylor et al., 1991; Komatsuzaki, 2007; Thapa et al., 2011; Renzi et al., 2017). These models, in their simplest form, consist of five stages from seed production, seed dispersal and input into soil seed banks, through germination and seedling emergence to the reproductive phase (adult plants) (Figure 3). The study of these stages and processes can be the basis for identifying the causes of failure or success in the establishment and performance of the selected biotypes (Renzi et al., 2019).
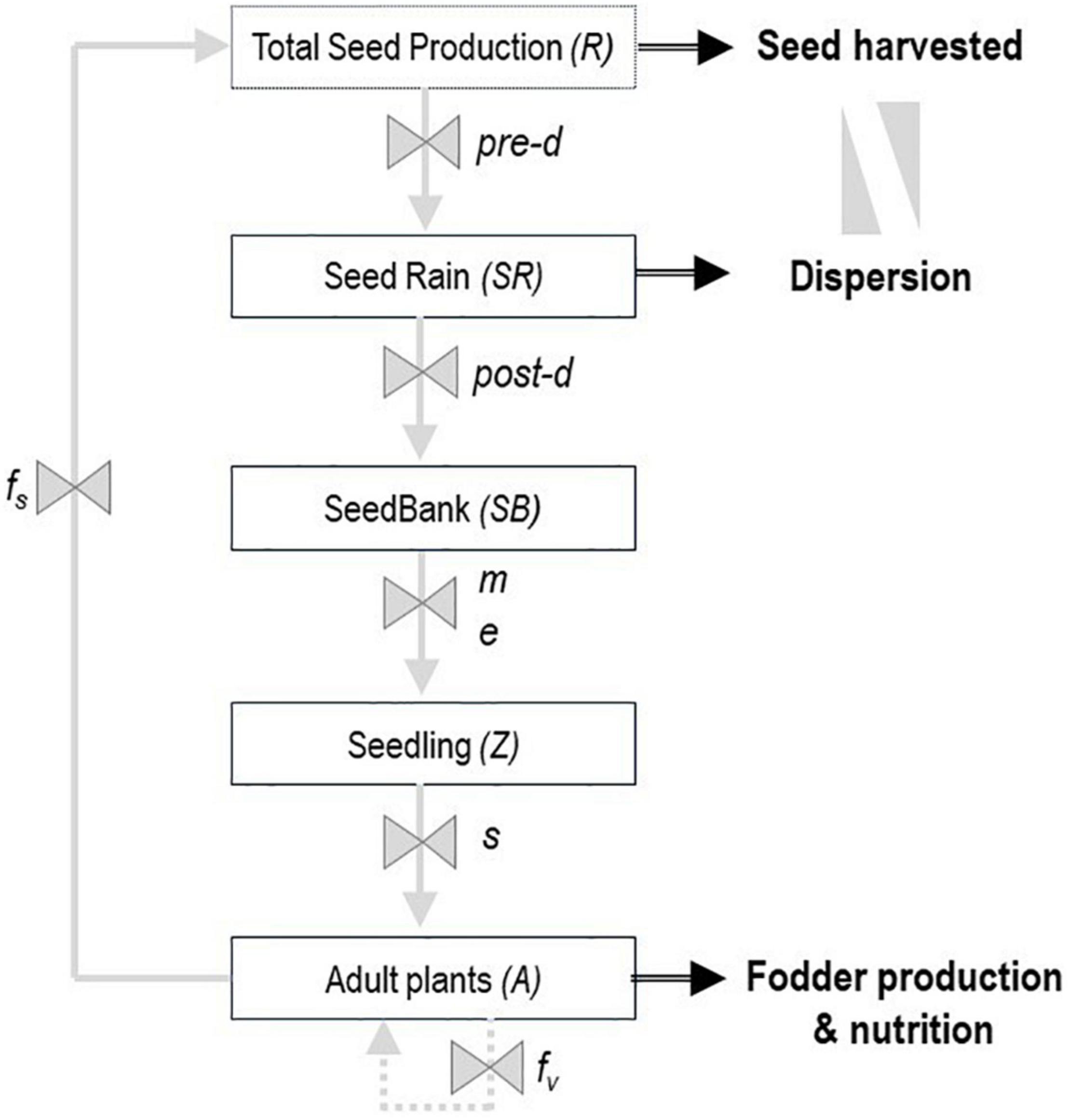
Figure 3. Conceptual model of the life cycle of an annual or perennial pasture biotype. Boxes indicate state in the plant life cycle. Gray arrows indicate the processes, and black arrows the agronomic traits of interest for pre-breeding. Process variables: pre-d, predation pre-dispersal; post-d, predation post-dispersal and seed rain losses; m, mortality of seeds in the soil; e, germination and seedling emergence; s, seedlings survivorship; fs, seed fecundity; and fv, vegetative reproduction.
The first requirement for this approach is to find the biotype(s) adapted to the target marginal environments, with acceptable forage yield and nutrition. Secondly, to reach a successful commercial scale, it is important to have seed available for farmers (Runck et al., 2020). To improve the harvestable seed yield, less seed shedding after physiological maturity, is a key objective for pre-breeding programs (Nichols et al., 2012). When the number of hectares that can be planted for cultivation I from a single hectare of seed production (S) is lower than 20 (C/S < 20), models that assist natural reseeding could be a good option. This is the case of Vicia villosa which is well-adapted to semi-arid temperate environments, but incompletely domesticated due to extreme indeterminate growth, non-uniform maturity, large seed losses due to uneven pod dehiscence, and seed dormancy (Kissing Kucek et al., 2020a,b; Renzi et al., 2020). The demographic model was useful for expanding their adoption by farmers in marginal lands in Argentina (Renzi et al., 2017, 2019). Several options of legumes with self-regeneration potential have been published (Carr et al., 2005; Loi et al., 2005, 2012; Norman et al., 2005; Ovalle et al., 2005; Driouech et al., 2008; Walsh et al., 2013). In a scenario of very low seed production, with a C/S ratio < 10, the natural seed dispersal should be considered in addition to the field emergence model. Under this premise, planting in patches or stripes (<<seed doses per ha) in function of the dispersal ability together with appropriate management will reduce the need for high initial seed availability. This management could be carried out in intercropping using biotypes with high resource-use complementarity (Litrico and Violle, 2015; Mazzafera et al., 2021). Thus, estimates of plant dispersal distances could be useful not only for ecological studies, but also for agronomic purposes. The dispersal traits of tall perennial pasture grasses suggest the potential applicability of this modeling approach (Thomson et al., 2011; Bullock et al., 2017). Nevertheless, complementary studies of temporal and spatial demographic dynamics are necessary.
Crop wild relatives are an untapped source of genetic diversity and a critical resource to meet food security needs and the challenges of new production systems, especially in response to climate change (Zhang et al., 2017; Bohra et al., 2021). However, three major factors discourage the use of CWRs in plant breeding (Zamir, 2001): (1) poor agronomic performance of CWRs, mostly due to the presence of undesirable traits such as shattering and seed dormancy; (2) crossing barriers between crops and their wild relatives, which limits the production of fertile offspring; and (3) the transfer of undesirable traits physically linked to the target trait, known as linkage drag (Zamir, 2001; Hajjar and Hodgkin, 2007; Shelef et al., 2017; Fernie and Yan, 2019). Therefore, to facilitate the use of CWRs in breeding programs, it is necessary to focus on the development of systematic strategies for the characterization and use of CWRs. Pre-breeding here refers to all activities designed to identify and transfer desirable traits or alleles from genetic materials that cannot be used directly in plant breeding (Prohens et al., 2017; Tefera, 2021). Pre-breeding produces intermediate genetic materials that can be used for breeders to produce elite cultivars. This section provides a review of distinct pre-breeding strategies to facilitate the use of CWRs in breeding programs, focused on the utilization of CWR for improving the tolerance to stressful conditions associated to marginal agricultural lands.
In the broad sense, strategies to incorporate CWR into breeding programs can be classified as “choose first” or “cross first” (Dempewolf et al., 2017). In “choose first” wild accessions are chosen from a larger number of accessions, based on phenotypic, genotypic, or eco-geographic merits. Chosen accessions are expected to carry a desirable trait, then they are used in a target crossing design. In “cross first” one or more wild accessions are crossed with elite cultivars, and then the progeny is evaluated, this approach is useful when the trait of interest cannot be directly measured in the wild donor.
Target hybridization is the classic example of the “choose first” strategy. Generally, a single wild accession carrying a single trait of interest (e.g., herbicide resistance, disease resistance) is used as donor and one elite cultivar, with good agronomic performance but lacking the trait of interest, is used as recurrent parent (Breseghello and Coelho, 2013). Crossing and backcrossing with positive selection for the target trait are used to incorporate the target trait and to recover most of the recurrent parent genome. This approach is routinely used in breeding programs to improve tolerance to pests and diseases (Hajjar and Hodgkin, 2007; Larkan et al., 2013; Seiler et al., 2017; Bohra et al., 2021). Using this approach, molecular markers can be used to map the region underlying the trait of interest, and to accelerate the recovery of the elite genome through assisted backcrossing (Breseghello and Coelho, 2013). The first step of target hybridization is to identify the suitable wild accession carrying the trait of interest (Zamir, 2001; Bahrami et al., 2021). As mentioned above, wild accessions can be selected based on phenotypic, genotypic, or eco-geographic merits. For phenotypic selection, several wild accessions are directly screened for the trait of interest along with elite cultivars; accessions outperforming elite cultivars for the target trait are selected for the next stage (Bahrami et al., 2021). Phenotypic screening has been used to identify wild accessions with increased tolerance to several stressful conditions associated to marginal agricultural lands, such as salinity in wild tomato (Pailles et al., 2020), freezing in wild sunflower (Hernández et al., 2020), drought in wheat and barley (Nevo and Chen, 2010), heat in wild rice (Scafaro et al., 2016; Bheemanahalli et al., 2017), wild sunflower (Hernández et al., 2018, 2020), and wild barley (Bahrami et al., 2021). Genotypic information can also be used to identify valuable wild accessions. For example, Buitrago-Bitar et al. (2021) identified wild accessions of tepary bean carrying novel alleles for previously identified drought-responsive genes. With the increasing amount of reported candidate genes for abiotic stress tolerance, genes-specific hybridization probes can be designed to target multiple candidate genes at once in many samples before next generation sequencing (Weitemier et al., 2014), thus accelerating the discovery of novel alleles of relevant genes. Lastly, eco-geographic tools can be used to identify possible sources of abiotic stress tolerance (e.g., Khazaei et al., 2013; Kantar et al., 2015; Stenberg and Ortiz, 2021). Eco-geographic approaches gather environmental variables from collection sites such as temperature, precipitation, and soil characteristics. This approach assumes that wild accessions from sites prone to a given abiotic stress (e.g., drought, cold or heat) likely carry adaptive alleles to cope with these environmental challenges (Kantar et al., 2015; Anderson et al., 2016). Phenotypic, genotypic, and eco-geographic approaches can be combined. Landscape genomics is a powerful approach that combines eco-geographic and genomic data to identify loci underlying environmental adaptation (Joost et al., 2007). This approach does not require phenotypic experiments, thus allowing the evaluation in silico of hundreds to thousands of accessions. Several climatic databased, including WorldClim (Fick and Hijmans, 2017), Envirem (Title and Bemmels, 2018), and SoilTemp (Lembrechts et al., 2021), along with genomic information can be used to link geographic coordinates of any accession with local environmental conditions and genomic data, increasing the value of ex situ collections. Recent examples focused on wild relatives of soybean (Anderson et al., 2016), wheat (Brunazzi et al., 2018), narrow-leafed lupin (Mousavi-Derazmahalleh et al., 2018), and chickpea (von Wettberg et al., 2018). Once the suitable accession is identified, a crossing and backcrossing design with an elite cultivar is initiated. As this approach uses only one wild accession as donor, genome-wide increases of genetic diversity are not expected. The advantage of this approach is the efficient transfer of traits regulated by a few genes, such as disease resistance (Seiler et al., 2017), and it is well-known by breeders. However, it is time-consuming and its success in traits with complex genetic architecture is not guaranteed.
The cross first strategy consists of developing populations of lines with introgressions from CWR into the genetic background of crops, aiming to generate elite materials carrying genome fragments from CWR (Tanksley and McCouch, 1997; Dempewolf et al., 2017; Prohens et al., 2017). Such populations can be used to identify genomic regions underlying adaptive traits, but they can also be directly incorporated into breeding programs (von Wettberg et al., 2018; El Haddad et al., 2021). There are many ways for developing populations with introgression from CWR; Chromosome Segment Substitution Lines (CSSL), Nested Association Mapping (NAM) populations, and Multiparent Advanced Generation Intercross (MAGIC) populations. These populations are often developed in an unfocused manner regarding target traits, i.e., the aim is to generate populations with as high genetic diversity from CWR as possible, thus populations are useful to map most agronomic traits in many environments. However, multi parent populations can be developed in a focused manner, selecting parents with particular features, such as tolerance to biotic or abiotic stress (reviewed in Prohens et al., 2017; Scott et al., 2020).
Chromosome Segment Substitution Lines were designed and used for mapping QTL in many crops (Balakrishnan et al., 2019). CSSL form a population of lines with chromosomal segments from one wild accession into an elite background (Balakrishnan et al., 2019). Ideally, each CSSL harbors a single segment of the donor and the whole donor genome is distributed segment-wise in the entire population of CSSL. In pre-breeding, CSSL has been used in several crops, such as rice, wheat, barley, maize, and soybean to identify and transfer alleles from wild species, including alleles associated to drought, heat, and freezing stress (reviewed in Balakrishnan et al., 2019). By definition, with this approach a single wild accession is used as donor and a single elite cultivar as recurrent parent, although CSSL populations using the same recurrent parent and distinct wild donors can be combined (Singh et al., 2020).
In the last decades, multiparent populations of crop species were developed and largely used to explore the genetic architecture of agronomic traits (Scott et al., 2020). Nested Association Mapping (NAM) and Multiparent Advanced Generation Intercross (MAGIC) populations are the most popular designs. They were designed to improve the resolution of QTL analysis with biparental populations and to reduce the confounding effects of population structure of association mapping (Gage et al., 2020). While NAM populations can be seen as a collection of biparental populations with a common parent (e.g., Yu et al., 2008; Hu et al., 2018), MAGIC populations have more complex crossing designs, commonly involving 4, 8, or 16 parents (Zamir, 2001; Gage et al., 2020; Scott et al., 2020). Such differences in the crossing designs determine that lines from NAM and MAGIC populations differ in their genetic composition, even when the same parents are used. In the NAM populations, lines are grouped into families, with as many families as donor parents, thus the genomes of NAM lines are a mosaic of only two parents, and haplotype diversity is constrained by the common parent (Zamir, 2001; Gage et al., 2020). On the contrary, the genomes of MAGIC lines are a mosaic of all founders, and haplotype diversity grows with the number of founders, although it is constrained by the number of lines in the population (Zamir, 2001; Scott et al., 2020). More than 50 multiparent populations were developed for several crop species (reviewed in Scott et al., 2020), however, only four of them included CWR as founders, three for barley and one for maize (Scott et al., 2020). In multiparent populations, the selection of founders is the most critical step, as it determines the pool of genetic variation segregating in the population. Choosing founders as genetically diverse as possible maximizes the population’s utility for mapping agronomic traits in multiple environments (Gage et al., 2020; Scott et al., 2020). However, for pre-breeding, a more conservative selection of founders will produce populations with higher overall agronomic performance, and therefore more attractive for breeders. In the case of NAM populations, after crossing the elite cultivar with CWR, one or more cycles of backcrosses can be implemented before selfing to improve the overall agronomic performance of the population (Chen et al., 2019). In the case of MAGIC populations, a trade-off between the overall agronomic performance and the novel genetic variation is expected, i.e., the larger the proportion of elite cultivars as founders the larger the overall agronomic performance and the lower the novel genetic variation, and vice versa.
The complex genetics controlling most abiotic stress tolerances presents a challenge for crop breeders to incorporate many CWR alleles into their breeding programs. Genomic selection (GS) is an efficient approach that was pioneered in animal breeding, where complex, polygenic traits are the norm (Goddard and Hayes, 2007). Genomic selection has been refined for plant breeding and adopted as a core methodology by many breeding programs (Crossa et al., 2017). This approach could be used to accumulate increasing numbers of alleles associated with stress tolerance and shift the breeding populations from stress-sensitive to stress-tolerant over many generations.
Genomic selection is increasingly common for a range of crops, e.g., barley (Gonzalez et al., 2021), pea (Al Bari et al., 2021), chickpea (Varshney et al., 2021), wheat (Muleta et al., 2017), and sorghum (Yu et al., 2016). However, GS to select within CWR collections has been proposed, but examples have yet to be published (Bohra et al., 2021). With the genotyping of larger CWR collections (e.g., Song et al., 2015; Sansaloni et al., 2020), it is expected this approach may be used in the future as it has been proven effective in plant breeding programs (Merrick and Carter, 2021). The importance of GS may increase for CWR pre-breeding programs as it allows for efficient introgression of beneficial alleles for polygenic traits with reduced linkage drag (Wang et al., 2017). Genetic selection has the advantage of selecting superior progeny based on genotypes alone using phenotyped training populations (Meuwissen et al., 2001). The first demonstration of GS in pre-breeding was in exotic maize populations successfully selecting for yield (Combs and Bernardo, 2013). Genomic selection in CWR pre-breeding was also validated recently in soybean (Beche et al., 2021). Very good prediction accuracies for several quantitative traits were reported using a NAM breeding population with three wild Plant Inventory parents (Beche et al., 2021). An excellent example for pre-breeding for marginal lands using focused identification of germplasm strategy (FIGS) is found in studies with barley, wheat and faba bean landraces (Endresen et al., 2011; Khazaei et al., 2013). FIGS is another machine learning application that takes multivariate data sets of a priori knowledge (phenotypes, environmental) to identify the potential most useful accessions (Bari et al., 2012). Stenberg and Ortiz (2021) suggest improving the FIGS model of adaptive processes (natural selection) by adding non-adaptive processes (gene flow and genetic drift) to hit the most intensive evolutionary hotspots to capture elusive traits followed by genomic selection.
In summary, the “choose first” strategy is more suitable for incorporating traits regulated by one or a few major loci (e.g., herbicide or disease resistance alleles) while the “cross first” is more suitable for polygenic traits, which is expected for most traits associated to adaptation to marginal agricultural lands. Multiple parent populations, especially MAGIC, are well-suited for the cross first strategy. Such populations are useful to discover agronomically relevant loci, to improve genetic diversity of pre-bred populations, and to directly incorporate them to breeding schemes. However, we identified a lack of multiparent populations (MPPs) that include CWR as founders, meaning that current MPPs are exploiting only a small fraction of species’ genetic diversity. Therefore, we call for a systematic development of MAGIC populations, which should include CWR, elite cultivars, and landraces as founders. The outlined advantages of MAGIC populations for both basic science and plant breeding should facilitate the creation of public-private pre-breeding partnership programs aiming to develop cultivars well-adapted to marginal agricultural lands. Although universal MAGIC populations can be used to discover agronomically relevant loci by multiple research groups, we encourage (when possible) the development of MAGIC populations with local founders to exploit local adaptation.
Since domestication began approximately 12,000 years ago, some significant events determined a deep change in the evolution of crops and their wild relatives. Fernie and Yan (2019) divided the history of crop improvement into four breeding generations, the first generation with breeding based on phenotype selection; the second generation with hybrid breeding and green revolution with dwarf plants and higher yields; a third generation called or “biotechnology-based breeding” with the arrival of genetically modified crops. Currently, the fourth generation of breeding is running, with genome editing and precision breeding. In fact, the fourth-generation breeding is determined by new plant breeding techniques like cis-genesis (Jacobsen and Schouten, 2007) and genome editing (Zsögön et al., 2017), mainly CRISPR/Cas (Sukegawa et al., 2021). Cis-genesis consists of the genetic transformation of a crop with CWR’s genes, without the introduction of reporters or selectable markers from other organisms, avoiding linkage drag. This is particularly interesting in the case of the secondary and tertiary genepool species (Harlan and de Wet, 1971) with strong hybridization barriers.
Genome editing techniques involve zinc-finger nucleases (ZFNs), transcription activator-like effector nucleases (TALENs), and clustered regularly interspaced short palindrome repeats associated protein 9 (Cas9), CRISPR/Cas systems (Manghwar et al., 2019; Sedeek et al., 2019; Van Tassel et al., 2020; Molla et al., 2021). ZFNs and TALENs are protein-based and require protein engineering for every user-defined sequence. Instead, CRISPR/Cas is a RNA-guided system that induces double-stranded DNA breaks by the action of Cas9 nuclease at a genome corresponding location (Belhaj et al., 2013; Altpeter et al., 2016; Rodríguez-Leal et al., 2017). Among genome editing techniques in plants, CRISPR/Cas9 has become the most popular (Altpeter et al., 2016; Pacher and Puchta, 2017; Das et al., 2019). The CRISPR/Cas9 system can generate stable and heritable mutations in genes that cannot be distinguished from a natural mutation at the same locus, without affecting the existing valuable traits (Belhaj et al., 2013; Arora and Narula, 2017). Since the main domestication genes in most of the major crops have been studied (see reviews by Kantar et al., 2017; Purugganan, 2019), the new gene editing tools would allow neo-domestication avoiding the linkage drag. De novo domestication or neo-domestication is defined as the targeted introduction of domestication genes into non-domesticated plants, this represents an important opportunity for fitting cultivated species to the climatic niche where they live (Fernie and Yan, 2019). In addition, genome editing allows to incorporate new adaptation attributes present in wild materials into cultivated ones, bringing resilience under the unfavorable conditions of climate change (Van Tassel et al., 2020). This is possible since the whole genome sequences for many crops and their wild species are now available (Bohra et al., 2021; Ramsay et al., 2021). However, it should be emphasized that this approach has yet to be fully tested and is unlikely to transfer all the beneficial traits accumulated in crops over centuries of generations, such as complex traits like grain yield, which is controlled by many genes.
Genome editing (GE) can be used for several purposes like to activate or suspend the function of any gene. Ghoshal et al. (2021) showed that with epigenome engineering it is possible to achieve heritable methylation, gene silencing, and delayed flowering time phenotypes. Hence, stable heritable epigenetic modifications in flowering time would allow researchers and breeders to overcome the crossing barriers of the secondary and tertiary genepool, facilitating crossings of CWRs with related crops (Cable et al., 2021). Genome editing can also be used to knock out genes. For example, increasing grain number and larger grain size in rice was possible by knocking out three mutations on negative regulators of yield (Mishra et al., 2018). In addition, knocking out the self-incompatibility gene S-Rnase allowed re-domestication of potato into an inbred-line based diploid crop for genetic improvement (Ye et al., 2018). This demonstrated the utility of this approach in four different Solanum tuberosum clones, opening possibilities in both future diploid breeding and basic research in self-incompatible crops.
Another application for GE is to generate novel alleles of any gene. For breeding drought tolerance in maize, novel allelic variation for ARGOS8 locus was generated by changing the DNA sequence at the native ARGOS8 (Shi et al., 2017). Genome editing could also add genes that do not exist in an original genome. There is also work on CRISPR/Cas9-mediated HDR (homology-directed repair) for editing genes that confer herbicide resistance, and to transfer an allele from a wild rice variant into a cultivated variant to increase yield potential. In addition, GE makes it possible to delete any sequence including large chromosomal fragments or even the entire chromosome (Xiao et al., 2013). This could be an advantage for the secondary and tertiary genepool species (Harlan and de Wet, 1971). Introgression of beneficial alleles from CWR into the cultivated gene pool has been a challenge when reproductive barriers are present. For instance, the genus Helianthus, native to North America, comprises 52 species including cultivated sunflower, H. annuus L. which has emerged as a model for genetic studies of adaptation, hybridization, and speciation. In particular, H. annuus and H. petiolaris are chromosomally divergent species, differing by a minimum of seven translocations and three inversions (Rieseberg et al., 1995). Notably, the first source of cytoplasmic male sterility for hybrid varieties’ development was discovered in H. petiolaris (Leclercq, 1969) and all currently grown sunflower hybrids are developed on cytoplasmic male sterility petiolaris type mothers. Chromosomal rearrangements are common in CWRs and drastically decrease recombination in crosses with crops by preventing the normal pairing of homologous chromosomes. The CRISPR/Cas system could be used to alter the frequency and guide homologous recombination to overcome these structural barriers between related species, thus opening new avenues to genomic restructuring and chromosomal engineering (Fauser et al., 2014).
The tertiary gene pool can also benefit from GE. Toda et al. (2019) demonstrated the possibility of genome editing in rice and maize zygotes, opening an avenue to gene manipulation in CWR wide crosses aiming to incorporate valuable traits to crops.
Utilizing CWR in breeding and crop improvement requires that researchers have access to germplasm resources. International agreements, particularly the Nagoya Protocol to the Convention on Biological Diversity and the International Treaty on Plant Genetic Resources for Food and Agriculture, the two international agreements that govern the collection and sharing of crop genetic resources, enshrine access and benefit sharing as core principles, so that those who give germplasm resources benefit from them too. Since much CWR diversity comes from less developed regions of the world, yet has historically primarily been harnessed in the less diverse global north, there are real historical inequalities that access and benefit sharing (ABS) frameworks should address. Negotiations around ABS in treaties have increasingly focused on Digital Sequence Information (DSI) associated with CWR (Rohden and Scholz, 2022; von Wettberg and Khoury, 2022). Digital Sequence Information may encompass not only DNA sequence data associated with germplasm accessions, but also CWR passport data, traditional ecological knowledge on the preparation or use of varieties (Brink and van Hintum, 2022). This information, as our discussion above shows, can be incredibly powerful as a breeding tool, yet is very unequally distributed globally.
For those seeking germplasm access, there are a number of data sources for CWR as well as other databases with additional data and access for ordering seed or propagule samples. The Global Biodiversity Information Facility (GBIF) repository remains an essential database and data repository for discovery of species locations that includes most herbaria globally1 (Lane and Edwards, 2007). Efforts by the Global Crop Diversity Trust have greatly improved knowledge of target CWR taxa through their Genesys database2 (Dempewolf et al., 2014) and one of their partners with GRIN-Global3 (Postman et al., 2010). A number of CWR characterization, evaluation and genotype data sets can be found here under the Genesys ‘Dataset’ repository. Genesys and GRIN-Global are data repositories only; no user analysis tools are present or planned in this resource.
As noted by Brink and van Hintum (2022), most genebanks do not plan to host genotype data resources. However, the GRIN-Global development team now has modules to improve the storage and downloading of crop genetic resource genotype data sets (see footnote 3). Although sequence resources for CWRs are increasing, currently most plant genetic resource sequencing datasets are on domesticated material, notable examples are found for several crops (Song et al., 2015; Sansaloni et al., 2020; Hellwig et al., 2021). A very promising development is the expansion of the Germinate database to handle pre-breeding populations from CWRs (Raubach et al., 2021). The Germinate software will provide the CWR pre-breeding community critical tools for working with and selection/advancement of introgression lines for growing in marginal production areas. Adoption of FAIR data standards will increase the availability of CWR evaluation data for pre-breeding efforts (Wilkinson et al., 2016).
For those harnessing CWRs, ex situ collections are critical as access points. But ex situ conservation alone is unable to fully conserve crop diversity, particularly for CWR that have historically been smaller parts of germplasm collections and harbor orders of magnitude more genetic diversity than domesticated accessions (Khoury et al., 2022). In situ conservation efforts for CWR have been growing over the last decade (Phillips et al., 2019) and can be essential to access traits of interest that are not covered by ex situ collections. In situ genetic reserve conservation may be defined as “the location, designation, management and monitoring of genetic diversity in natural wild populations within defined areas designated for active, long-term conservation” (Maxted et al., 1997). Some of the success in these efforts has been to harness existing conservation areas, often established for large animal preservation but also protecting CWR inadvertently. Dedicated efforts at in situ conservation have lagged, particularly in Vavilovian centers of origin where CWR taxa are most diverse and abundant. Protecting CWRs in their natural habitats is an essential strategy to prevent extinctions, especially where ex situ conservation of sufficient genetic diversity is practically or economically unfeasible (León-Lobos et al., 2012). There is a preference for in situ conservation, because it has the advantage of maintaining the dynamic evolution of the CWR diversity itself in relation to parallel biotic and abiotic changes. In the span of only three decades, for example, there was a shift in allelic diversity related to flowering time genes in wild barley (Qian et al., 2020) likely as response to global warming, and even an overwhelming local adaptation and ecological differentiation. Gap Analysis (e.g., Maxted et al., 2008) has been applied for hundreds of CWRs to find areas where ex situ collections and in situ conservation areas fail to protect the full range of diversity in CWR (Vincent et al., 2019; Khoury et al., 2020; Zair et al., 2021).
JR, JBr, CC, JBe, EW, MN, SU, FH, and PS contributed equally to writing and final editing. JR conceived the original idea. All authors contributed to the article and approved the submitted version.
PS was supported by the Grant Agency of the Czech Republic and Palacký University Grant Agency [IGA-2022_002]. JR work was supported by INTA PE-142 project. EW was supported by the USDA Hatch program through the Vermont State Agricultural Experimental Station. EW was also funded by the Ministry of Science and Higher Education of the Russian Federation as part of World-class Research Center program: Advanced Digital Technologies (contract No. 075-15-2022-311 dated 20.04.2022).
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
PS and JBr acknowledge the support from Palacký University for travel to INTA, Argentina during which initial idea on review topic was established.
Abbo, S., Berger, J., and Turner, N. C. (2003). Evolution of cultivated chickpea: four bottlenecks limit diversity and constrain adaptation. Funct. Plant Biol. 30, 1081–1087.
Abbo, S., Lev-Yadun, S., and Galwey, N. (2002). Vernalization response of wild chickpea. New Phytol. 154, 695–701.
Abbo, S., Pinhasi van-Oss, R., Gopher, A., Saranga, Y., Ofner, I., and Peleg, Z. (2014). Plant domestication versus crop evolution: a conceptual framework for cereals and grain legumes. Trends Plant Sci. 19, 351–360. doi: 10.1016/j.tplants.2013.12.002
Ahmadzai, H., Tutundjian, S., and Elouafi, I. (2021). Policies for sustainable agriculture and livelihood in marginal lands: a review. Sustainability 13:8692.
Al Bari, M. A. A., Worral, H., Szwiec, S., Ma, Y., Zheng, P., Main, D., et al. (2021). Harnessing genetic diversity in the USDA pea (Pisum sativum L.) germplasm collection through genomic prediction. Front. Genet. 12:707754. doi: 10.3389/fgene.2021.707754
Alonso-Blanco, C., Aarts, M. G. M., Bentsink, L., Keurentjes, J. J. B., Reymond, M., Vreugdenhil, D., et al. (2009). What has natural variation taught us about plant development, physiology, and adaptation? Plant Cell 21, 1877–1896. doi: 10.1105/tpc.109.068114
Altpeter, F., Springer, N. M., Bartley, L. E., Blechl, A. E., Brutnell, T. P., Citovsky, V., et al. (2016). Advancing crop transformation in the era of genome editing. Plant Cell 28, 1510–1520. doi: 10.1105/tpc.16.00196
Anderson, J. E., Kono, T. J. Y., Stupar, R. M., Kantar, M. B., and Morrell, P. L. (2016). Environmental association analyses identify candidates for abiotic stress tolerance in Glycine soja, the wild progenitor of cultivated soybeans. G3 Genes Genom. Genet. 6, 835–843. doi: 10.1534/g3.116.026914
Anderson, J. T., Willis, J. H., and Mitchell-Olds, T. (2011). Evolutionary genetics of plant adaptation. Trends Genet. 27, 258–266. doi: 10.1016/j.tig.2011.04.001
Annicchiarico, P., Barrett, B., Brummer, E. C., Julier, B., and Marshall, A. H. (2015). Achievements and challenges in improving temperate perennial forage legumes. Crit. Rev. Plant Sci. 34, 327–380. doi: 10.1080/07352689.2014.898462x
Araújo, S. S., Beebe, S., Crespi, M., Delbreil, B., González, E. M., Gruber, V., et al. (2015). Abiotic stress responses in legumes: strategies used to cope with environmental challenges. Crit. Rev. Plant Sci. 34, 237–280. doi: 10.1080/07352689.2014.898450
Araus, J. L., and Kefauver, S. C. (2018). Breeding to adapt agriculture to climate change: affordable phenotyping solutions. Curr. Opin. Plant Biol. 45(Pt. B), 237–247. doi: 10.1016/j.pbi.2018.05.003
Arora, L., and Narula, A. (2017). Gene editing and crop improvement using CRISPR-cas9 system. Front. Plant Sci. 8:1932. doi: 10.3389/fpls.2017.01932
Ayangbenro, A. S., and Babalola, O. O. (2020). Reclamation of arid and semi-arid soils: the role of plant growth-promoting archaea and bacteria. Curr. Plant Biol. 25, 100173. doi: 10.1016/j.cpb.2020.100173
Badaruddin, M. I., and Meyer, D. W. (2001). Factors modifying frost tolerance of legume species. Crop Sci. 41, 1911–1916.
Bahrami, F., Arzani, A., and Rahimmalek, M. (2021). Tolerance to high temperature at reproductive stage: trade-offs between phenology, grain yield and yield-related traits in wild and cultivated barleys. Plant Breed. 140, 812–826. doi: 10.1111/pbr.12953
Balakrishnan, D., Surapaneni, M., Mesapogu, S., and Neelamraju, S. (2019). Development and use of chromosome segment substitution lines as a genetic resource for crop improvement. Theor. Appl. Genet. 132, 1–25. doi: 10.1007/s00122-018-3219-y
Bari, A., Street, K., Mackay, M., Endresen, D. T. F., De Pauw, E., and Amri, A. (2012). Focused identification of germplasm strategy (FIGS) detects wheat stem rust resistance linked to environmental variables. Genet. Resour. Crop Evol. 59, 1465–1481.
Basey, A. C., Fant, J. B., and Kramer, A. T. (2015). Producing native plant materials for restoration: 10 rules to collect and maintain genetic diversity. Native Plants J. 16, 37–53. doi: 10.3368/npj.16.1.37
Beche, E., Gillman, J. D., Song, Q., Nelson, R., Beissinger, T., Decker, J., et al. (2021). Genomic prediction using training population design in interspecific soybean populations. Mol. Breed. 41:15. doi: 10.1007/s11032-021-01203-6
Belhaj, K., Chaparro-Garcia, A., Kamoun, S., and Nekrasov, V. (2013). Plant genome editing made easy: targeted mutagenesis in model and crop plants using the CRISPR/Cas system. Plant Methods 9, 1–10. doi: 10.1186/1746-4811-9-39
Berger, J. D., and Ludwig, C. (2014). Contrasting adaptive strategies to terminal drought stress gradients in Mediterranean legumes: phenology, productivity and water relations in wild and domesticated Lupinus luteus L. J. Exp. Bot. 65, 6219–6229. doi: 10.1093/jxb/eru006
Berger, J. D., Buck, R., Henzell, J. M., and Turner, N. C. (2005). Evolution in the genus Cicer – vernalisation response and low temperature pod set in chickpea (C. arietinum L.) and its annual wild relatives. Aust. J. Agric. Res. 56, 1191–1200.
Berger, J. D., Kumar, S., Nayyar, H., Street, K., Sandhu, J. S., Henzell, J. M., et al. (2012b). Temperature-stratified screening of chickpea (Cicer arietinum L.) genetic resource collections reveals very limited reproductive chilling tolerance compared to its annual wild relatives. Field Crops Res. 126, 119–129.
Berger, J. D., Buirchell, B., Luckett, D. J., and Nelson, M. N. (2012a). Domestication bottlenecks limit genetic diversity and constrain adaptation in narrow-leafed lupin (Lupinus angustifolius L.). Theor. Appl. Genet. 124, 637–652. doi: 10.1007/s00122-011-1736-z
Berger, J. D., Ludwig, C., and Whisson, K. (2020). Changing water use and adaptive strategies along rainfall gradients in Mediterranean lupins. Plant Biol. 22, 298–308. doi: 10.1111/plb.13076
Berger, J. D., Milroy, S. P., Turner, N. C., Siddique, K. H. M., Imtiaz, M., and Malhotra, R. (2011). Chickpea evolution has selected for contrasting phenological mechanisms among different habitats. Euphytica 180, 1–15.
Berger, J. D., Robertson, L. D., and Cocks, P. S. (2002). Agricultural potential of Mediterranean grain and forage legumes: key differences between and within Vicia species in terms of phenology, yield, and agronomy give insight into plant adaptation to semi-arid environments. Genet. Resour. Crop Evol. 49, 313–325.
Berger, J. D., Shrestha, D., and Ludwig, C. (2017). Reproductive strategies in mediterranean legumes: trade-offs between phenology, seed size and vigor within and between wild and domesticated Lupinus species collected along aridity gradients. Front. Plant Sci. 8:548. doi: 10.3389/fpls.2017.00548
Berger, J., Palta, J., and Vadez, V. (2016). Review: an integrated framework for crop adaptation to dry environments: responses to transient and terminal drought. Plant Sci. 253, 58–67. doi: 10.1016/j.plantsci.2016.09.007
Berger, J., Pushpavalli, R., Ludwig, C., Parsons, S., Basdemir, F., and Whisson, K. (2020). Wild and domestic differences in plant development and responses to water deficit in Cicer. Front. Genet. 11:607819. doi: 10.3389/fgene.2020.607819
Bheemanahalli, R., Sathishraj, R., Manoharan, M., Sumanth, H. N., Muthurajan, R., Ishimaru, T., et al. (2017). Is early morning flowering an effective trait to minimize heat stress damage during flowering in rice? Field Crops Res. 203, 238–242. doi: 10.1016/j.fcr.2016.11.011
Bohra, A., Kilian, B., Sivasankar, S., Caccamo, M., Mba, C., McCouch, S. R., et al. (2021). Reap the crop wild relatives for breeding future crops. Trends Biotechnol. 40, 412–431. doi: 10.1016/j.tibtech.2021.08.009
Brandsæter, L. O., Olsmo, A., Tronsmo, A. M., and Fykse, H. (2002). Freezing resistance of winter annual and biennial legumes at different developmental stages. Crop Sci. 42:437. doi: 10.2135/cropsci2002.4370
Breseghello, F., and Coelho, A. S. G. (2013). Traditional and modern plant breeding methods with examples in rice (Oryza sativa L.). J. Agric. Food Chem. 61, 8277–8286. doi: 10.1021/jf305531j
Brink, M., and van Hintum, T. (2022). Practical consequences of digital sequence information (DSI) definitions and access and benefit-sharing scenarios from a plant genebank’s perspective. Plants People Planet 4, 23–32. doi: 10.1002/ppp3.10201
Brozynska, M., Furtado, A., and Henry, R. J. (2016). Genomics of crop wild relatives: expanding the gene pool for crop improvement. Plant Biotechnol. J. 14, 1070–1085. doi: 10.1111/pbi.12454
Brunazzi, A., Scaglione, D., Talini, R. F., Miculan, M., Magni, F., Poland, J., et al. (2018). Molecular diversity and landscape genomics of the crop wild relative Triticum urartu across the Fertile Crescent. Plant J. 94, 670–684. doi: 10.1111/tpj.13888
Brus, J., Pechanec, V., and Machar, I. (2018). Depiction of uncertainty in the visually interpreted land cover data. Ecol. Inf. 47, 10–13. doi: 10.1016/j.ecoinf.2017.10.015
Buitrago-Bitar, M. A., Cortés, A. J., López-Hernández, F., Londoño-Caicedo, J. M., Muñoz-Florez, J. E., Carmenza Muñoz, L., et al. (2021). Allelic diversity at abiotic stress responsive genes in relationship to ecological drought indices for cultivated tepary bean, Phaseolus acutifolius A. Gray, and its wild relatives. Genes 12, 1–17. doi: 10.3390/genes12040556
Bullock, J. M., Mallada González, L., Tamme, R., Götzenberger, L., White, S. M., Pärtel, M., et al. (2017). A synthesis of empirical plant dispersal kernels. J. Ecol. 105, 6–19. doi: 10.1111/1365-2745.12666
Cable, J., Ronald, P. C., Voytas, D., Zhang, F., Levy, A. A., Takatsuka, A., et al. (2021). Plant genome engineering from lab to field—a Keystone Symposia report. Ann. N. Y. Acad. Sci. 1506, 35–54. doi: 10.1111/nyas.14675
Campbell, J. E., Lobell, D. B., Genova, R. C., and Field, C. B. (2008). The global potential of bioenergy on abandoned agriculture lands. Environ. Sci. Technol. 42, 5791–5794. doi: 10.1021/es800052w
Cantamutto, M. A., Bertucci, C. L., and Huarte, R. D. (2016). El trigo en el Sudoeste Bonaerense. Ed INTA. 76. Available online at: https://inta.gob.ar/sites/default/files/inta_trigo-sudoeste.bonaerense.2016.pdf (accessed February 1, 2022).
Carr, P. M., Poland, W. W., and Tisor, L. J. (2005). Forage legume regeneration from the soil seed bank in Western North Dakota. Agron. J. 97, 505–513.
Castro, H., and Castro, P. (2019). “Mediterranean marginal lands in face of climate change: biodiversity and ecosystem services,” in Climate Change-Resilient Agriculture and Agroforestry, eds P. Castro, A. Azul, W. Leal Filho, and U. Azeiteiro (Cham: Springer), 175–187.
Chapman, S. C., Cooper, M., Hammer, G. L., and Butler, D. G. (2000). Genotype by environment interactions affecting grain sorghum. II. Frequencies of different seasonal patterns of drought stress are related to location effects on hybrid yields. Aust. J. Agric. Res. 51:209. doi: 10.1071/AR99021
Chaturvedi, P., Wiese, A. J., Ghatak, A., Zaveska Drabkova, L., Weckwerth, W., and Honys, D. (2021). Heat stress response mechanisms in pollen development. New Phytol. 231, 571–585. doi: 10.1111/nph.17380
Chen, C., Fletcher, A., Lawes, R., Berger, J., and Robertson, M. (2017). Modelling phenological and agronomic adaptation options for narrow-leafed lupins in the southern grainbelt of Western Australia. Eur. J. Agron. 89, 140–147.
Chen, Q., Yang, C. J., York, A. M., Xue, W., Daskalska, L. L., DeValk, C. A., et al. (2019). TeoNAM: a nested association mapping population for domestication and agronomic trait analysis in maize. Genetics 213, 1065–1078. doi: 10.1534/genetics.119.302594
Chen, S., Stefanova, K., Siddique, K. H., and Cowling, W. A. (2021). Transient daily heat stress during the early reproductive phase disrupts pod and seed development in Brassica napus L. Food Energy Secur. 10:e262.
Clark, R. M., Linton, E., Messing, J., and Doebley, J. F. (2004). Pattern of diversity in the genomic region near the maize domestication gene tb1. Proc. Natl. Acad. Sci. U.S.A. 101, 700–707. doi: 10.1073/pnas.2237049100
Coba de la Peña, T., and Pueyo, J. J. (2011). Legumes in the reclamation of marginal soils, from cultivar and inoculant selection to transgenic approaches. Agron. Sustain. Dev. 32, 65–91. doi: 10.1007/s13593-011-0024-2
Colbach, N., Durr, C., Roger-Estrade, J., and Caneill, J. (2005). How to model the effects of farming practices on weed emergence. Weed Res. 45, 2–17.
Combs, E., and Bernardo, R. (2013). Genomewide selection to introgress semidwarf maize germplasm into U.S. Corn Belt inbreds. Crop Sci. 53, 1427–1436. doi: 10.2135/cropsci2012.11.0666
Cortes, A. J., and Lopez-Hernandez, F. (2021). Harnessing crop wild diversity for climate change adaptation. Genes 12:21. doi: 10.3390/genes12050783
Cowling, W. A., Buirchell, B. J., and Falk, D. E. (2009). A model for incorporating novel alleles from the primary gene pool into elite crop breeding programs while reselecting major genes for domestication or adaptation. Crop Pasture Sci. 60, 1009–1015.
Coyne, C. J., Shiv Kumar, S., von Wettberg, E. B., Marques, E., Berger, J. D., Redden, R. J., et al. (2020). Potential and limits of exploitation of crop wild relatives for pea, lentil and chickpea improvement. Legume Sci. 2:e36. doi: 10.1002/leg3.36
Crossa, J., Pérez-Rodríguez, P., Cuevas, J., Montesinos-López, O., Jarquín, D., de los Campos, G., et al. (2017). Genomic selection in plant breeding: methods, models, and perspectives. Trends Plant Sci. 22, 961–975. doi: 10.1016/j.tplants.2017.08.011
Das, A., Sharma, N., Prasad, M. (2019). CRISPR/Cas9: a novel weapon in the arsenal to combat plant diseases. Front. Plant Sci. 9:2008. doi: 10.3389/fpls.2018.02008
Dauber, J., Jones, M. B., and Stout, J. C. (2010). The impact of biomass crop cultivation on temperate biodiversity. GCB Bioenergy 2, 289–309.
De Andrés, E., Zambrana, E., Cadorniga, C., Martín, D., Marcos, T., De la Rosa, L., et al. (2008). Estudio de caracteres de resistencia a la sequía en la colección activa de Vicia sativa L del CRF-INIA. Actas Asoc. Esp. Leguminosas 3, 124–125.
de Felipe, M., and Alvarez Prado, S. (2021). Has yield plasticity already been exploited by soybean breeding programmes in Argentina? J. Exp. Bot. 72, 7264–7273. doi: 10.1093/jxb/erab347
De la Rosa, L., Zambrana, E., and Ramirez-Parra, E. (2020). Molecular bases for drought tolerance in common vetch: designing new molecular breeding tools. BMC Plant Biol. 20:71. doi: 10.1186/s12870-020-2267-z
DeHaan, L., Larson, S., López-Marqués, R. L., Wenkel, S., Gao, C., and Palmgren, M. (2020). Roadmap for accelerated domestication of an emerging perennial grain crop. Trends Plant Sci. 25, 525–537.
Dempewolf, H., Baute, G., Anderson, J., Kilian, B., Smith, C., and Guarino, L. (2017). Past and future use of wild relatives in crop breeding. Crop Sci. 57, 1070–1082. doi: 10.2135/cropsci2016.10.0885
Dempewolf, H., Eastwood, R. J., Guarino, L., Khoury, C. K., Müller, J. V., and Toll, J. (2014). Adapting agriculture to climate change: a global initiative to collect, conserve, and use crop wild relatives. Agroecol. Sustain. Food Syst. 38, 369–377.
Donald, C. M. (1981). “Competitive plants, communal plants, and yield in wheat crops,” in Wheat Science – Today and Tomorrow, eds L. T. Evans and W. J. Peackock (Cambridge: Cambridge University Press), 223–247. doi: 10.3389/fpls.2021.734056
Driouech, N., Abou Fayad, F., Ghanem, A., and Al-bitar, L. (2008). “Agronomic performance of annual self-reseeding legumes and their self-establishment potential in the Apulia region of Italy,” in Proceedings of the 16th IFOAM Organic World Congress, Modena.
Duc, G., Agrama, H., Bao, S., Berger, J., Bourion, V., De Ron, A. M., et al. (2015). Breeding annual grain legumes for sustainable agriculture: new methods to approach complex traits and target new cultivar ideotypes. Crit. Rev. Plant Sci. 34, 381–411. doi: 10.1080/07352689.2014.898469
Dürr, C., Dickie, J., Yang, X.-Y., and Pritchard, H. (2015). Ranges of critical temperature and water potential values for the germination of species worldwide: contribution to a seed trait database. Agric. For. Meteorol. 200, 222–232.
El Haddad, N., Sanchez-Garcia, M., Visioni, A., Jilal, A., El Amil, R., Sall, A. T., et al. (2021). Crop wild relatives crosses: multi-location assessment in durum wheat, barley, and lentil. Agronomy 11:2283. doi: 10.3390/agronomy11112283
El-Beltagy, A., and Madkour, M. (2012). Impact of climate change on arid lands agriculture. Agric. Food Secur. 1, 1–12. doi: 10.1186/2048-7010-1-3
Elbersen, B., Van Eupen, M., Mantel, S., Alexopoulou, E., Bai, Z., Boogard, H., et al. (2018). “Mapping marginal land potentially available for industrial crops in Europe,” in Paper presented at the European Biomass Conference and Exhibition Proceedings 26thEUBCE, Copenhagen.
Endresen, D. T. F., Street, K., Mackay, M., Bari, A., and De Pauw, E. (2011). Predictive association between biotic stress traits and eco-geographic data for wheat and barley landraces. Crop Sci. 5, 2036–2055.
Espeland, E. K., Johnson, R. C., and Horning, M. E. (2018). Plasticity in native perennial grass populations: implications for restoration. Evol. Appl. 11, 340–349.
Evenson, R. E., and Gollin, D. (2003). Assessing the impact of the Green Revolution, 1960 to 2000. Science 300, 758–762. doi: 10.1126/science.1078710
Fauser, F., Schiml, S., and Puchta, H. (2014). Both CRISPR/Cas-based nucleases and nickases can be used efficiently for genome engineering in Arabidopsis thaliana. Plant J. 79, 348–359. doi: 10.1111/tpj.12554
Fernández, I., Fraysse, M., Presotto, A., and Cantamutto, M. A. (2012). Evaluation of Argentine wild sunflower biotypes for drought stress during reproductive stage. Helia 35, 29–36. doi: 10.2298/hel1257029f
Fernie, A. R., and Yan, J. (2019). De novo domestication: an alternative route toward new crops for the future. Mol. Plant 12, 615–631.
Fick, S. E., and Hijmans, R. J. (2017). WorldClim 2: new 1-km spatial resolution climate surfaces for global land areas. Int. J. Climatol. 37, 4302–4315. doi: 10.1002/joc.5086
Forcella, F., Benech-Arnold, R. L., Sánchez, R., and Ghersa, C. M. (2000). Modeling seedling emergence. Field Crops Res. 67, 123–139.
Foyer, C., Lam, H. M., Nguyen, H., Siddique, K. H., Varshney, R. K., Colmer, T. D., et al. (2016). Neglecting legumes has compromised human health and sustainable food production. Nat. Plants 2:16112. doi: 10.1038/nplants.2016.112
Fuller, D. Q., and Allaby, R. (2009). “Seed dispersal and crop domestication: shattering, germination and seasonality in evolution under cultivation,” in Annual Plant Reviews: Fruit Development and Seed Dispersal, ed. L. Østergaard (Oxford: Wiley-Blackwell), 238–295.
Fuller, D. Q., and Allaby, R. (2018). “Seed dispersal and crop domestication: shattering, germination and seasonality in evolution under cultivation,” in Annual Plant Reviews online, ed. J. A. Roberts (Chichester: John Wiley & Sons, Ltd), 238–295. doi: 10.1002/9781119312994.apr0414
Fustier, M. A., Brandenburg, J. T., Boitard, S., Lapeyronnie, J., Eguiarte, L. E., Vigouroux, Y., et al. (2017). Signatures of local adaptation in lowland and highland teosintes from whole-genome sequencing of pooled samples. Mol. Ecol. 26, 2738–2756.
Gage, J. L., Monier, B., Giri, A., and Buckler, E. S. (2020). Ten years of the maize nested association mapping population: impact, limitations, and future directions. Plant Cell 32, 2083–2093. doi: 10.1105/tpc.19.00951
Gardarin, A., Coste, F., Wagner, M. H., and Dürr, C. (2016). How do seed and seedling traits influence germination and emergence parameters in crop species? A comparative analysis. Seed Sci. Res. 26, 317–331. doi: 10.1017/s0960258516000210
Gardarin, A., Durr, C., and Colbach, N. (2012). Modelling the dynamics and emergence of multispecies weed seed bank with species traits. Ecol. Modell. 240, 123–138.
Garibaldi, L. A., Aizen, M. A., Saez, A., Gleiser, G., Strelin, M. M., and Harder, L. D. (2021). The influences of progenitor filtering, domestication selection and the boundaries of nature on the domestication of grain crops. Funct. Ecol. 35, 1998–2011.
Ghoshal, B., Picard, C. L., Vong, B., Feng, S., and Jacobsen, S. E. (2021). CRISPR-based targeting of DNA methylation in Arabidopsis thaliana by a bacterial CG-specific DNA methyltransferase. Proc. Natl. Acad. Sci. U.S.A. 118:e2125016118. doi: 10.1073/pnas.2125016118
Gladstones, J. S. (1994). “An historical review of lupins in Australia,” in Proceedings of the 1st Lupin Technical Symposium, eds M. Dracup and J. A. Palta (Perth, WA: Department of Agriculture), 1–38. doi: 10.1002/ar.24517
Gladstones, J. S., and Hill, G. D. (1969). Selection for economic characters in Lupinus angustifulius and L. digitatus. 2. Time of flowering. Aust. J. Exp. Agric. Anim. Husbandry 9, 213–220.
Gonzalez, M. Y., Zhao, Y., Jiang, Y., Stein, N., Habekuss, A., Reif, J. C., et al. (2021). Genomic prediction models trained with historical records enable populating the German ex situ genebank bio-digital resource center of barley (Hordeum sp.) with information on resistances to soilborne barley mosaic viruses. Theor. Appl. Genet. 134, 2181–2196. doi: 10.1007/s00122-021-03815-0
González-Andujar, J. L., and Fernández-Quintanilla, C. (1993). Strategies for the control of Avena sterilis in winter wheat production systems in central Spain. Crop Protect. 12, 617–623.
González-Andujar, J. L., and Fernández-Quintanilla, C. (2004). Modelling the population dynamics of annual ryegrass (Lolium rigidum) under various weed management systems. Crop Protect. 23, 723–729.
González-Paleo, L., and Ravetta, D. A. (2015). Carbon acquisition strategies uncoupled from predictions derived from species life-cycle. Flora 212, 1–9. doi: 10.1016/j.flora.2015.02.004
Hajjar, R., and Hodgkin, T. (2007). The use of wild relatives in crop improvement: a survey of developments over the last 20 years. Euphytica 156, 1–13. doi: 10.1007/s10681-007-9363-0
Harlan, J. R., and de Wet, J. M. J. (1971). Toward a rational classification of cultivated plants. Taxon 20, 509–517. doi: 10.2307/1218252
Hellwig, T., Abbo, S., Sherman, A., and Ophir, R. (2021). Prospects for genetic diversity of crop wild relatives under climate change: the case of the wild pea Pisum fulvum. Plant Sci. 310:110957.
Hernández, F., Poverene, M., and Presotto, A. (2018). Heat stress effects on reproductive traits in cultivated and wild sunflower (Helianthus annuus L.): evidence for local adaptation within the wild germplasm. Euphytica 214:146. doi: 10.1007/s10681-018-2227-y
Hernández, F., Poverene, M., Mercer, K. L., and Presotto, A. (2020). Genetic variation for tolerance to extreme temperatures in wild and cultivated sunflower (Helianthus annuus) during early vegetative phases. Crop Pasture Sci. 71:578. doi: 10.1071/CP20005
Hoad, S. P., Davies, D. H. K., and Topp, C. E. F. (2006). How to select varieties for organic farming: science and practice. Aspects Appl. Biol. 79, 117–120.
Holst, N., Rasmussen, I. A., and Bastiaans, L. (2007). Field weed population dynamics: a review of model approaches and applications. Weed Res. 47, 1–14.
Hondelmann, W. (1984). The Lupin-ancient and modern crop plant. Theor. Appl. Genet. 68, 1–9. doi: 10.1007/BF00252301
Hradilová, I., Duchoslav, M., Brus, J., Pechanec, V., Hýbl, M., Kopecký, P., et al. (2019). Variation in wild pea (Pisum sativum subsp. elatius) seed dormancy and its relationship to the environment and seed coat traits. PeerJ 6:e6263. doi: 10.7717/peerj.6263
Hu, J., Guo, C., Wang, B., Ye, J., Liu, M., Wu, Z., et al. (2018). Genetic properties of a Nested Association Mapping population constructed with semi-winter and spring oilseed rapes. Front. Plant Sci. 9:1740. doi: 10.3389/fpls.2018.01740
Iqbal, M. M., Erskine, W., Berger, J. D., and Nelson, M. N. (2020). Phenotypic characterisation and linkage mapping of domestication syndrome traits in yellow lupin (Lupinus luteus L.). Theor. Appl. Genet. 133, 2975–2987. doi: 10.1007/s00122-020-03650-9
Jacobsen, E., and Schouten, H. J. (2007). Cisgenesis strongly improves introgression breeding and induced translocation breeding of plants. Trends Biotechnol. 25, 219–223. doi: 10.1016/j.tibtech.2007.03.008
Jones, H. P., Hole, D. G., and Zavaleta, E. S. (2012). Harnessing nature to help people adapt to climate change. Nat. Clim. Change 2, 504–509.
Joost, S., Bonin, A., Bruford, M. W., Després, L., Conord, C., Erhardt, G., et al. (2007). A spatial analysis method (SAM) to detect candidate loci for selection: towards a landscape genomics approach to adaptation. Mol. Ecol. 16, 3955–3969. doi: 10.1111/j.1365-294X.2007.03442.x
Kang, S., Post, W. M., Nichols, J. A., Wang, D., West, T. O., Bandaru, V., et al. (2013). Marginal lands: concept, assessment and management. J. Agric. Sci. 5:129.
Kantar, M. B., Nashoba, A. R., Anderson, J. E., Blackman, B. K., and Rieseberg, L. H. (2017). The genetics and genomics of plant domestication. BioScience 67, 971–982.
Kantar, M. B., Sosa, C. C., Khoury, C. K., Castañeda-Álvarez, N. P., Achicanoy, H. A., Bernau, V., et al. (2015). Ecogeography and utility to plant breeding of the crop wild relatives of sunflower (Helianthus annuus L.). Front. Plant Sci. 6:841. doi: 10.3389/fpls.2015.00841
Kattge, J., Díaz, S., Lavorel, S., Prentice, I. C., Leadley, P., Bönisch, G., et al. (2011). TRY—a global database of plant traits. Glob. Change Biol. 17, 2905–2935. doi: 10.1111/gcb.14869
Kebede, E. (2021). Contribution, utilization, and improvement of legumes-driven biological nitrogen fixation in agricultural systems. Front. Sustain. Food Syst. 5:767998. doi: 10.3389/fsufs.2021.767998
Khazaei, H., Street, K., Bari, A., Mackay, M., and Stoddard, F. L. (2013). The FIGS (Focused Identification of Germplasm Strategy) approach identifies traits related to drought adaptation in Vicia faba genetic resources. PLoS One 8:e63107. doi: 10.1371/journal.pone.0063107
Khoury, C. K., Brush, S., Costich, D. E., Curry, H. A., de Haan, S., Engels, J. M. M., et al. (2022). Crop genetic erosion: understanding and responding to loss of crop diversity. New Phytol. 233, 84–118. doi: 10.1111/nph.17733
Khoury, C. K., Carver, D., Greene, S. L., Williams, K. A., Achicanoy, H. A., Schori, M., et al. (2020). Crop wild relatives of the United States require urgent conservation action. Proc. Natl. Acad. Sci. U.S.A. 117, 33351–33357. doi: 10.1073/pnas.2007029117
Kissing Kucek, L., Riday, H., Rufener, B. P., Burke, A. N., Eagen, S. S., Ehlke, N., et al. (2020a). Pod dehiscence in hairy vetch (Vicia villosa Roth). Front. Plant Sci. 11:82. doi: 10.3389/fpls.2020.00082
Kissing Kucek, L., Azevedo, M. D., Eagen, S. S., Ehlke, N. J., Hayes, R. J., Mirsky, S. B., et al. (2020b). Seed dormancy in hairy vetch (Vicia villosa Roth) is influenced by genotype and environment. Agronomy 10:1804. doi: 10.3390/agronomy10111804
Komatsuzaki, M. (2007). New weed management strategy using subterranean clover reseeding under different tillage systems: numerical experiments with the subterranean clover-tillage dynamics model. Weed Biol. Manag. 7, 3–13.
Lane, M. A., and Edwards, J. L. (2007). “The global biodiversity information facility (GBIF),” in Biodiversity Databases, Vol. 73, eds G. B. Curry and C. J. Humphries (Boca Raton, FL: CRC Press).
Lark, T. J., Spawn, S. A., Bougie, M., and Gibbs, H. K. (2020). Cropland expansion in the United States produces marginal yields at high costs to wildlife. Nat. Commun. 11, 1–11. doi: 10.1038/s41467-020-18045-z
Larkan, N. J., Lydiate, D. J., Parkin, I. A. P., Nelson, M. N., Epp, D. J., Cowling, W. A., et al. (2013). The Brassica napus blackleg resistance gene LepR3 encodes a receptor-like protein triggered by the Leptosphaeria maculans effector AVRLM1. New Phytol. 197, 595–605. doi: 10.1111/nph.12043
Lembrechts, J. J., van den Hoogen, J., Aalto, J., Ashcroft, M. B., De Frenne, P., Kemppinen, J., et al. (2021). Global maps of soil temperature. Glob. Chang. Biol. 28, 3110–3144. doi: 10.1111/gcb.16060
León-Lobos, P., Way, M., Aranda, P. D., and Lima-Junior, M. (2012). The role of ex situ seed banks in the conservation of plant diversity and in ecological restoration in Latin America. Plant Ecol. Diver. 5, 245–258. doi: 10.1080/17550874.2012.713402
Lipper, L., Thornton, P., Campbell, B. M., Baedeker, T., Braimoh, A., Bwalya, M., et al. (2014). Climate-smart agriculture for food security. Nat. Clim. Change 4, 1068–1072. doi: 10.1016/j.agsy.2017.09.007
Litrico, I., and Violle, C. (2015). Diversity in plant breeding: a new conceptual framework. Trends Plant Sci. 2, 604–613. doi: 10.1016/j.tplants.2015.07.007
Liu, M., Li, Y., Ma, Y., Zhao, Q., Stiller, J., Qi, F., et al. (2020). The draft genome of a wild barley genotype reveals its enrichment in genes related to biotic and abiotic stresses compared to cultivated barley. Plant Biotechnol. J. 18, 443–456. doi: 10.1111/pbi.13210
Loi, A., Howieson, J. G., Nutt, B. J., and Carr, S. J. (2005). A second generation of annual pasture legumes and their potential for inclusion in Mediterranean-type farming systems. Aust. J. Agric. Res. 45, 289–299.
Loi, A., Nutt, B. J., Howieson, J. G., Yates, R. J., and Norman, H. C. (2012). Preliminary assessment of bladder clover (Trifolium spumosum L.) as an annual legume for ley farming systems in southern Australia. Crop Pasture Sci. 63, 582–591.
Ludlow, M. M., and Muchow, R. C. (1990). A critical evaluation of traits for improving crop yields in water-limited environments. Adv. Agron. 43, 107–153. doi: 10.1071/FP13149
Manghwar, H., Lindsey, K., Zhang, X., and Jin, S. (2019). CRISPR/Cas system: recent advances and future prospects for genome editing. Trends Plant Sci. 24, 1102–1125. doi: 10.1016/j.tplants.2019.09.006
Matesanz, S., Ramos-Muñoz, M., Moncalvillo, B., Rubio Teso, M. L., García de Dionisio, S. L., Romero, J., et al. (2020). Plasticity to drought and ecotypic differentiation in populations of a crop wild relative. AoB Plants 12:laa006. doi: 10.1093/aobpla/plaa006
Maxted, N., Dulloo, E., Ford-Lloyd, B. V., Iriondo, J. M., and Jarvis, A. (2008). Gap analysis: a tool for complementary genetic conservation assessment. Divers. Distrib. 14, 1018–1030. doi: 10.1111/j.1472-4642.2008.00512.x
Maxted, N., Hawkes, J., Guarino, L., and Sawkins, M. (1997). Towards the selection of taxa for plant genetic conservation. Genet. Resour. Crop Evol. 44, 337–348. doi: 10.1023/A:1008643206054
Mazzafera, P., Favarin, J. L., and Andrade, S. A. L. (2021). Intercropping Systems in Sustainable Agriculture. Front. Sustain. Food Syst. 5:634361. doi: 10.3389/fsufs.2021.634361
McCouch, S., Navabi, Z. K., Abberton, M., Anglin, N. L., Barbieri, R. L., Baum, M., et al. (2020). Mobilizing crop biodiversity. Mol. Plant 13, 1341–1344. doi: 10.1016/j.molp.2020.08.011
Merrick, L. F., and Carter, A. H. (2021). Comparison of genomic selection models for exploring predictive ability of complex traits in breeding programs. Plant Genome 14:e20158. doi: 10.1002/tpg2.20158
Meuwissen, T., Hayes, B., and Goddard, M. (2001). Prediction of total genetic value using genome-wide dense marker maps. Genetics 157, 1819–1829. doi: 10.1093/genetics/157.4.1819
Meyer, R. S., Duval, A. E., and Jensen, H. R. (2012). Patterns and processes in crop domestication: an analysis of 203 global food crops. New Phytol. 196, 29–48.
Mishra, R., Joshi, R. K., and Zhao, K. (2018). Genome editing in rice: recent advances, challenges, and future implications. Front. Plant Sci. 9:1361. doi: 10.3389/fpls.2018.01361
Molla, K. A., Sretenovic, S., Bansal, K. C., and Yiping, Q. (2021). Precise plant genome editing using base editors and prime editors. Nat. Plants 7, 1166–1187. doi: 10.1038/s41477-021-00991-1
Mousavi-Derazmahalleh, M., Bayer, P. E., Nevado, B., Hurgobin, B., Filatov, D., Kilian, A., et al. (2018). Exploring the genetic and adaptive diversity of a pan-Mediterranean crop wild relative: narrow-leafed lupin. Theor. Appl. Genet. 131, 887–901. doi: 10.1007/s00122-017-3045-7
Muir, J. P., Pitman, W. D., Dubeux, J. C., and Foster, J. L. (2014). The future of warm-season, tropical and subtropical forage legumes in sustainable pastures and rangelands. Afr. J. Range Forage Sci. 31, 187–198. doi: 10.2989/10220119.2014.884165
Muir, J. P., Pitman, W. D., Smith, F. S., Lloyd-Reilley, J., and Shadow, R. A. (2018). Challenges to developing native legume seed supplies: the Texas experience as a case study. Native Plants J. 19, 224–238. doi: 10.3368/npj.19.3.224
Muleta, K. T., Bulli, P., Zhang, Z., Chen, X., and Pumphrey, M. (2017). Unlocking diversity in germplasm collections via genomic selection: a case study based on quantitative adult plant resistance to stripe rust in spring wheat. Plant Genome 10:3. doi: 10.3835/plantgenome2016.12.0124
Muller, K., Guinness, J., Hecking, M., and Drinkwater, L. (2021). Estimating agronomically relevant symbiotic N fixation in green manure breeding programs. Crop Sci. 61, 3314–3330. doi: 10.7298/vrd4-4k23
Nagel, R., Durka, W., Bossdorf, O., and Bucharova, A. (2019). Rapid evolution in native plants cultivated for ecological restoration: not a general pattern. Plant Biol. 21, 551–558. doi: 10.1111/plb.12901
Nevo, E., and Chen, G. (2010). Drought and salt tolerances in wild relatives for wheat and barley improvement. Plant, Cell Environ. 33, 670–685. doi: 10.1111/j.1365-3040.2009.02107.x
Nichols, P. G. H., Revell, C. K., Humphries, A. W., Howie, J. H., Hall, E. J., Sandral, G. A., et al. (2012). Temperate pasture legumes in Australia – their history, current use, and future prospects. Crop Pasture Sci. 63, 691–725.
Nicotra, A. B., Atkin, O. K., Bonser, S. P., Davidson, A. M., Finnegan, J., Mathesius, U., et al. (2010). Plant phenotypic plasticity in a changing climate. Trends Plant Sci. 15, 684–692.
Norman, H. C., Cocks, P. S., and Galwey, N. W. (2002). Hardseededness in annual clovers: variation between populations from wet and dry environments. Aust. J. Agric. Res. 53, 821–829.
Norman, H. C., Cokcs, P. S., and Galwey, N. W. (2005). Annual clovers (Trifolium spp.) have different reproductive strategies to achieve persistence in Mediterranean-type climates. Aust. J. Agric. Res. 56, 33–43.
Nutt, B. J., and Loi, A. (1999). “Harvestability of annual Mediterranean pasture legumes using conventional crop machinery,” in “Proceedings of the 4th International Herbage Seed Conference”, eds M. Fancinelli and D. Rosellini (Perugia: University of Perugia), 78–82.
Ogutcen, E., Pandey, A., Khan, M. K., Marques, E., Penmetsa, R. V., Kahraman, A., et al. (2018). Pod shattering: a homologous series of variation underlying domestication and an avenue for crop improvement. Agronomy 8:137.
Ovalle, C. M., Del Pozo, A. L., Avendaño, J. R., Fernández, F. E., and Arredondo, S. S. (2005). Adaptation, growth and production of new annual forage legumes in the Mediterranean Zone of Chile. II. Species performance in granitic soils of the sub-humid interior dryland. Agric. Técnica 65, 265–277.
Pacher, M., and Puchta, H. (2017). From classical mutagenesis to nuclease-based breeding—directing natural DNA repair for a natural end-product. Plant J. 90, 819–833. doi: 10.1111/tpj.13469
Pailles, Y., Awlia, M., Julkowska, M., Passone, L., Zemmouri, K., Negrão, S., et al. (2020). Diverse traits contribute to salinity tolerance of wild tomato seedlings from the Galapagos Islands. Plant Physiol. 182, 534–546. doi: 10.1104/pp.19.00700
Passioura, J. (2006). Increasing crop productivity when water is scarce—from breeding to field management. Agric. Water Manage. 80, 176–196. doi: 10.1016/j.agwat.2005.07.012
Phillips, J., Whitehouse, K., and Maxted, N. (2019). An in situ approach to the conservation of temperate cereal crop wild relatives in the Mediterranean Basin and Asian centre of diversity. Plant Genet. Resour. 17, 185–195. doi: 10.1017/S1479262118000588
Phillips, S., Anderson, R., and Schapire, R. (2006). Maximum entropy modeling of species geographic distributions. Ecol. Modell. 190, 231–259.
Piano, P., Pecetti, L., and Carroni, A. M. (1996). Climatic adaptation in subterranean clover populations. Euphytica 92, 39–44.
Pingali, P. L. (2012). Green revolution: impacts, limits, and the path ahead. Proc. Natl. Acad. Sci. U.S.A. 109, 12302–12308. doi: 10.1073/pnas.0912953109
Pizza, R., Espeland, E., and Etterson, J. (2021). Eight generations of native seed cultivation reduces plant fitness relative to the wild progenitor population. Evol. Appl. 14, 1816–1829. doi: 10.1111/eva.13243
Polechová, J., and Barton, N. H. (2015). Limits to adaptation along environmental gradients. Proc. Natl. Acad. Sci. U.S.A. 112, 6401–6406. doi: 10.1073/pnas.1421515112
Postman, J., Hummer, K., Ayala-Silva, T., Bretting, P., Franko, T., Kinard, G., et al. (2010). GRIN-GLOBAL: an international project to develop a global plant Genebank information management system. Acta Hortic. 859, 49–55. doi: 10.17660/actahortic.2010.859.4
Prohens, J., Gramazio, P., Plazas, M., Dempewolf, H., Kilian, B., Diez, M. J., et al. (2017). Introgressiomics: a new approach for using crop wild relatives in breeding for adaptation to climate change. Euphytica 213:158. doi: 10.1007/s10681-017-1938-9
Provorov, N., and Tikhonovich, I. (2003). Genetic resources for improving nitrogen fixation in legume-rhizobia symbiosis. Genet. Resour. Crop Evol. 50, 89–99. doi: 10.1023/A:1022957429160
Purugganan, M. D. (2019). Evolutionary insights into the nature of plant domestication. Curr. Biol. 29, R705–R714. doi: 10.1016/j.cub.2019.05.053
Purugganan, M. D., and Fuller, D. Q. (2009). The nature of selection during plant domestication. Nature 457:843. doi: 10.1038/nature07895
Qian, C., Yan, X., Shi, Y., Yin, H., Chang, Y., Chen, J., et al. (2020). Adaptive signals of flowering time pathways in wild barley from Israel over 28 generations. Heredity 124, 62–76. doi: 10.1038/s41437-019-0264-5
Quezada-Martinez, D., Addo Nyarko, C. P., Schiessl, S. V., and Mason, A. S. (2021). Using wild relatives and related species to build climate resilience in Brassica crops. Theor. Appl. Genet. 134, 1711–1728. doi: 10.1007/s00122-021-03793-3
Ramsay, L., Koh, C. S., Kagale, S., Gao, D., Kaur, S., Haile, T., et al. (2021). Genomic rearrangements have consequences for introgression breeding as revealed by genome assemblies of wild and cultivated lentil species. bioRxiv [preprint]. doi: 10.1101/2021.07.23.453237
Raubach, S., Kilian, B., Dreher, K., Amri, A., Bassi, F. M., Boukar, O., et al. (2021). From bits to bites: advancement of the Germinate platform to support prebreeding informatics for crop wild relatives. Crop Sci. 61, 1538–1566.
Redden, R. (2013). New approaches for crop genetic adaptation to the abiotic stresses predicted with climate change. Agronomy 3, 419–432. doi: 10.3390/agronomy3020419
Redden, R., and Berger, J. D. (2007). “History and origin of chickpea,” in Chickpea Breeding and Management, eds S. S. Yadav, R. Redden, W. Chen, and B. Sharma (Wallingford: CABI), 1–13.
Renzi, J. P. (2020). Caracterización Agroecológica de Poblaciones Naturales de Vicia Villosa Roth (Fabaceae) de Argentina. Tesis de Doctorado en Agronomía. Bahía Blanca: Universidad Nacional del Sur, 152.
Renzi, J. P., Chantre, G. R., and Cantamutto, M. A. (2017). Self-regeneration of hairy vetch (Vicia villosa Roth) as affected by seedling density and soil tillage method in a semi-arid agroecosystem. Grass Forage Sci. 72, 535–544.
Renzi, J. P., Chantre, G. R., Smýkal, P., Presotto, A. D., Zubiaga, L., Garayalde, A. F., et al. (2020). Diversity of naturalized hairy vetch (Vicia villosa Roth) populations in central argentina as a source of potential adaptive traits for breeding. Front. Plant Sci. 11:189. doi: 10.3389/fpls.2020.00189
Renzi, J. P., Chantre, G., González-Andújar, J. L., and Cantamutto, M. A. (2019). Development and validation of a simulation model for hairy vetch (Vicia villosa Roth) self-regeneration under different crop rotations. Field Crops Res. 235, 79–86.
Reynolds, M. P., Saint Pierre, C., Saad, A. S. I., Vargas, M., and Condon, A. G. (2007). Evaluating potential genetic gains in wheat associated with stress adaptive trait expression in elite genetic resources under drought and heat stress. Crop Sci. 47, 172–189. doi: 10.2135/cropsci2007.10.0022ipbs
Ricardo, D. (2005). “From the principles of political economy and taxation,” in Readings in the Economics of the Division of Labor: The Classical Tradition, ed. S. Guang-Zhen (Hackensack, NJ: World Scientific), 127–130.
Richards, R. A. (2006). Physiological traits used in the breeding of new cultivars for water-scarce environments. Agric. Water Manag. 80, 197–211. doi: 10.1016/j.agwat.2005.07.013
Rieseberg, L. H., Van Fossen, C., and Desrochers, A. M. (1995). Hybrid speciation accompanied by genomic reorganization in wild sunflowers. Nature 375, 313–316.
Rodríguez-Leal, D., Lemmon, Z. H., Man, J., Bartlett, M. E., and Lippman, Z. B. (2017). Engineering quantitative trait variation for crop improvement by genome editing. Cell 171, 470–480. doi: 10.1016/j.cell.2017.08.030
Rohden, F., and Scholz, A. H. (2022). The international political process around Digital Sequence Information under the Convention on Biological Diversity and the 2018–2020 intersessional period. Plants People Planet 4, 51–60.
Runck, B. C., Kantar, M. B., Jordan, N. R., Anderson, J. A., Wyse, D. L., Eckberg, J. O., et al. (2014). The reflective plant breeding paradigm: a robust system of Germplasm development to support strategic diversification of Agroecosystems. Crop Sci. 54, 1939–1948. doi: 10.2135/cropsci2014.03.0195
Runck, B. C., Khoury, C. K., Ewing, P. M., and Kantar, M. (2020). The hidden land use cost of upscaling cover crops. Commun. Biol. 3:300. doi: 10.1038/s42003-020-1022-1
Russell, J., van Zonneveld, M., Dawson, I. K., Booth, A., Waugh, R., and Steffenson, B. (2014). Genetic diversity and ecological niche modeling of wild barley: refugia, large-scale post-LGM Range expansion and limited mid-future climate threats? PLoS One 9:e86021. doi: 10.1371/journal.pone.0086021
Saatkamp, A., Cochrane, A., Commander, L., Guja, L. K., Jimenez-Alfaro, B., Larson, J., et al. (2019). A research agenda for seed-trait functional ecology. New Phytol. 221, 1764–1775. doi: 10.1111/nph.15502
Sadras, V. O., and Richards, R. A. (2014). Improvement of crop yield in dry environments: benchmarks, levels of organization and the role of nitrogen. J. Exp. Bot. 65, 1981–1995. doi: 10.1093/jxb/eru061
Sakuma, S., Salomon, B., and Komatsuda, T. (2011). The domestication syndrome genes responsible for the major changes in plant form in the Triticeae crops. Plant Cell Physiol. 52, 738–749. doi: 10.1093/pcp/pcr025
Sandquist, D. R., and Ehleringer, J. R. (1997). Intraspecific variation of leaf pubescence and drought response in Encelia farinosa associated with contrasting desert environments. New Phytol. 135, 635–644. doi: 10.1046/j.1469-8137.1997.00697.x
Sansaloni, C., Franco, J., Santos, B., Percival-Alwyn, L., Singh, S., Petroli, C., et al. (2020). Diversity analysis of 80,000 wheat accessions reveals consequences and opportunities of selection footprints. Nat. Commun. 11:4572. doi: 10.1038/s41467-020-18404-w
Scafaro, A. P., Gallé, A., Van Rie, J., Carmo-Silva, E., Salvucci, M. E., and Atwell, B. J. (2016). Heat tolerance in a wild Oryza species is attributed to maintenance of Rubisco activation by a thermally stable Rubisco activase ortholog. New Phytol. 211, 899–911. doi: 10.1111/nph.13963
Scott, M. F., Ladejobi, O., Amer, S., Bentley, A. R., Biernaskie, J., Boden, S. A., et al. (2020). Multi-parent populations in crops: a toolbox integrating genomics and genetic mapping with breeding. Heredity 125, 396–416. doi: 10.1038/s41437-020-0336-6
Sebby, K. (2010). The Green Revolution of the 1960’s and its Impact on Small Farmers in India. Undergraduate thesis. Lincoln, NE: University of Nebraska–Lincoln.
Sedeek, K. E. M., Mahas, A., and Mahfouz, M. (2019). Plant genome engineering for targeted improvement of crop traits. Front. Plant Sci. 10:114. doi: 10.3389/fpls.2019.00114
Sehgal, A., Sita, K., Siddique, K. H. M., Kumar, R., Bhogireddy, S., Varshney, R. K., et al. (2018). Drought or/and heat-stress effects on seed filling in food crops: impacts on functional biochemistry, seed yields, and nutritional quality. Front. Plant Sci. 9:1705. doi: 10.3389/fpls.2018.01705
Seiler, G. J., Qi, L. L., and Marek, L. F. (2017). Utilization of sunflower crop wild relatives for cultivated sunflower improvement. Crop Sci. 57, 1083–1101. doi: 10.2135/cropsci2016.10.0856
Sharma, S., and Upadhyaya, H. D. (2019). Photoperiod response of annual wild cicer species and cultivated chickpea on phenology, growth, and yield traits. Crop Sci. 59, 632–639.
Shelef, O., Weisberg, P. J., and Provenza, F. D. (2017). The value of native plants and local production in an era of global agriculture. Front. Plant Sci. 8:2069. doi: 10.3389/fpls.2017.02069
Shi, J., Gao, H., Wang, H., Lafitte, H. R., Archibald, R. L., Yang, M., et al. (2017). ARGOS8 variants generated by CRISPR-Cas9 improve maize grain yield under field drought stress conditions. Plant Biotechnol. J. 15, 207–216. doi: 10.1111/pbi.12603
Silenzi, J. C., Echeverria, N. E., Vallejos, A. G., Bouza, M. E., and De Lucia, M. P. (2012). Wind erosion risk in the southwest of Buenos Aires province, Argentina and its relationship to the productivity index. Aeolian Res. 3, 419–425.
Singh, K. B., Malhotra, R. S., and Saxena, M. C. (1990). Sources for tolerance to cold in Cicer species. Crop Sci. 30, 1136–1138.
Singh, K. B., Malhotra, R. S., Halila, M. H., Knights, E. J., and Verma, M. M. (1994). Current status and future strategy in breeding chickpea for resistance to biotic and abiotic stresses. Euphytica 73, 137–149.
Singh, N., Wang, D. R., Ali, L., Kim, H., Akther, K. M., Harrington, S. E., et al. (2020). A coordinated suite of wild-introgression lines in Indica and Japonica elite backgrounds. Front. Plant Sci. 12:564824. doi: 10.3389/fpls.2020.564824
Skarbø, K., and VanderMolen, K. (2014). Irrigation access and vulnerability to climate-induced hydrological change in the Ecuadorian Andes. Cult. Agric. Food Environ. 36, 28–44.
Smýkal, P., Nelson, M., Berger, J., and von Wettberg, E. (2018). The impact of genetic changes during crop domestication. Agronomy 8:119. doi: 10.3390/agronomy8070119
Smýkal, P., Vernoud, V., Blair, M. W., Soukup, A., and Thompson, R. D. (2014). The role of the testa during development and in establishment of dormancy of the legume seed. Front. Plant Sci. 5:351. doi: 10.3389/fpls.2014.00351
Smýkal, P., Hradilová, I., Trněný, O., Brus, J., Rathore, A., Bariotakis, M., et al. (2017). Genomic diversity and macroecology of the crop wild relatives of domesticated pea. Sci. Rep. 7:17384.
Song, Q., Hyten, D. L., Jia, G., Quigley, C. V., Fickus, E. W., Nelson, R. L., et al. (2015). Fingerprinting soybean germplasm and its utility in genomic research. G3 Genes Genom. Genet. 5, 1999–2006. doi: 10.1534/g3.115.019000
Stenberg, J. A., and Ortiz, R. (2021). Focused identification of germplasm strategy (FIGS): polishing a rough diamond. Curr. Opin. Insect Sci. 45, 1–6. doi: 10.1016/j.cois.2020.11.001
Sukegawa, S., Saika, H., and Toki, S. (2021). Plant genome editing: ever more precise and wide reaching. Plant J. 106, 1208–1218. doi: 10.1111/tpj.15233
Tanksley, S. D., and McCouch, S. R. (1997). Seed banks and molecular maps: unlocking genetic potential from the wild. Science 277, 1063–1066. doi: 10.1126/science.277.5329.1063
Taylor, C. M., Garg, G., Berger, J. D., Ribalta, F. M., Croser, J. S., Singh, K. B., et al. (2021). A Trimethylguanosine Synthase1-like (TGS1) homologue is implicated in vernalisation and flowering time control. Theor. Appl. Genet. 134, 3411–3426. doi: 10.1007/s00122-021-03910-2
Taylor, C. M., Kamphuis, L. G., Zhang, W., Garg, G., Berger, J. D., Mousavi-Derazmahalleh, M., et al. (2019). INDEL variation in the regulatory region of the major flowering time gene LanFTc1 is associated with vernalization response and flowering time in narrow-leafed lupin (Lupinus angustifolius L.). Plant Cell Environ. 42, 174–187. doi: 10.1111/pce.13320
Taylor, G. B., Maller, L. A., and Rossiter, R. C. (1991). A model describing the influence of hard seededness on the persistence of an annual forage legume, in a ley farming system, in a mediterranean-type environment. Agric. Ecosyst. Environ. 37, 275–301.
Tefera, A. (2021). Pre-breeding concept and role in crop improvement. Int. J. Res. Appl. Sci. Biotechnol. 8, 2–7. doi: 10.31033/ijrasb.8.2.37
Thapa, R., Kemp, D. W., and Nichols, P. G. H. (2011). Low Cost Rehabilitation of Native Perennial Grass Pastures by Managing Seedling Recruitment. FFI CRC Technical Report 8-Farming systems. Crawley WA: The University of Western Australia.
Thomson, B., and Siddique, K. H. (1997). Grain legume species in low rainfall Mediterranean-type environments II. Canopy development, radiation interception, and dry-matter production. Field Crops Res. 54, 189–199.
Thomson, F. J., Moles, A. T., Auld, T. D., and Kingsford, R. T. (2011). Seed dispersal distance is more strongly correlated with plant height than with seed mass. J. Ecol. 99, 1299–1307. doi: 10.1111/j.1365-2745.2011.01867.x
Tilman, D., Balzer, C., Hill, J., and Befort, B. L. (2011). Global food demand and the sustainable intensification of agriculture. Proc. Natl. Acad. Sci. U.S.A. 108, 20260–20264. doi: 10.1073/pnas.1116437108
Title, P. O., and Bemmels, J. B. (2018). ENVIREM: an expanded set of bioclimatic and topographic variables increases flexibility and improves performance of ecological niche modeling. Ecography 41, 291–307. doi: 10.1111/ecog.02880
Toda, E., Koiso, N., Takebayashi, A., Ichikawa, M., Kiba, T., Osakabe, K., et al. (2019). An efficient DNA- and selectable-marker-free genome-editing system using zygotes in rice. Nat. Plants 5, 363–368. doi: 10.1038/s41477-019-0386-z
Václavík, T., Lautenbach, S., Kuemmerle, T., and Seppelt, R. (2013). Mapping global land system archetypes. Glob. Environ. Change 23, 1637–1647.
Van Tassel, D. L., Tesdell, O., Schlautman, B., Rubin, M. J., DeHaan, L. R., Crews, T. E., et al. (2020). New food crop domestication in the age of gene editing: genetic, agronomic and cultural change remain co-evolutionarily entangled. Front. Plant Sci. 11:789. doi: 10.3389/fpls.2020.00789
Van Tassel, D., Albrecht, K. A., Bever, J. D., Boe, A. A., Brandvain, Y., Crews, T. E., et al. (2017). Accelerating Silphium domestication: an opportunity to develop new crop ideotypes and breeding strategies informed by multiple disciplines. Crop Sci. 57, 1274–1284.
Varshney, R. K., Roorkiwal, M., Sun, S., Bajaj, P., Chitikineni, A., Thudi, M., et al. (2021). A chickpea genetic variation map based on the sequencing of 3,366 genomes. Nature 599, 622–627.
Vavilov, N. I. (1957). Agroecological Survey of the Main Field Crops. Moscow: Academy of Sciences of the USSR.
Vilela, A. E., and González-Paleo, L. (2015). Changes in resource-use strategy and phenotypic plasticity associated with selection for yield in wild species native to arid environments. J. Arid Environ. 113, 51–58. doi: 10.1016/j.jaridenv.2014.09.005
Vincent, H., Amri, A., Castañeda-Álvarez, N. P., Dempewolf, H., Dulloo, E., Guarino, L., et al. (2019). Modeling of crop wild relative species identifies areas globally for in situ conservation. Commun. Biol. 2:136. doi: 10.1038/s42003-019-0372-z
von Wettberg, E. J. B., Chang, P. L., Başdemir, F., Carrasquila-Garcia, N., Korbu, L. B., Moenga, S. M., et al. (2018). Ecology and genomics of an important crop wild relative as a prelude to agricultural innovation. Nat. Commun. 9:649. doi: 10.1038/s41467-018-02867-z
von Wettberg, E. J., Mukherjee, J. R., Adesky, N. D., Nesbeth, D., and Sistla, S. (2014). “The evolutionary ecology and genetics of stress resistance syndrome (SRS) traits: revisiting Chapin, Autumn and Pugnaire (1993),” in Plant Ecology and Evolution in Harsh Environments, eds N. Rajakaruna, R. S. Boyd, and T. B. Harris (New York, NY: Nova Science Publishers), 201–226.
von Wettberg, E., and Khoury, C. K. (2022). Biodiversity data: the importance of access and the challenges regarding benefit sharing. Plants People Planet 4, 2–4. doi: 10.1002/ppp3.10241
Walsh, M. J., Groose, R. W., Obour, A. K., Claypool, D. A., Delaney, R. H., and Krall, J. M. (2013). Seed persistence in soil of five medic cultivars in southeastern wyoming. Crop Sci. 53, 1–5.
Wang, C., Hu, S., Gardner, C., and Lübberstedt, T. (2017). Emerging avenues for utilization of exotic germplasm. Trends Plant Sci. 22, 624–637. doi: 10.1016/j.tplants.2017.04.002
Warschefsky, E., Varma Penmetsa, R., Cook, D. R., and von Wettberg, E. J. B. (2014). Back to the wilds: tapping evolutionary adaptations for resilient crops through systematic hybridization with crop wild relatives. Am. J. Bot. 101, 1791–1800. doi: 10.3732/ajb.1400116
Weiner, J., Du, Y.-L., Zhang, C., Qin, X.-L., and Li, F.-M. (2017). Evolutionary agroecology: individual fitness and population yield in wheat (Triticum aestivum). Ecology 98, 2261–2266. doi: 10.1002/ecy.1934
Weitemier, K., Straub, S. C. K., Cronn, R. C., Fishbein, M., Schmickl, R., McDonnell, A., et al. (2014). Hyb-Seq: combining target enrichment and genome skimming for plant phylogenomics. Appl. Plant Sci. 2:1400042. doi: 10.3732/apps.1400042
Wiering, N. P., Flavin, C., Sheaffer, C. C., Heineck, G. C., Sadok, W., and Ehlke, N. J. (2018). Winter hardiness and freezing tolerance in a hairy vetch collection. Crop Sci. 58:1594. doi: 10.2135/cropsci2017.12.0748
Wilke, B. J., and Snapp, S. S. (2008). Winter cover crops for local ecosystems: linking plant traits and ecosystem function. J. Sci. Food Agric. 88, 551–557. doi: 10.1002/jsfa.3149
Wilkinson, M. D., Dumontier, M., Aalbersberg, I. J., Appleton, G., Axton, M., Baak, A., et al. (2016). The FAIR Guiding Principles for scientific data management and stewardship. Sci. Data 3, 1–9. doi: 10.1038/sdata.2016.18
Xiao, A., Wang, Z., Hu, Y., Wu, Y., Luo, Z., Yang, Z., et al. (2013). Chromosomal deletions and inversions mediated by TALENs and CRISPR/Cas in zebrafish. Nucleic Acids Res. 41, 1–11. doi: 10.1093/nar/gkt464
Ye, M., Peng, Z., Tang, D., Yang, Z., Li, D., Xu, Y., et al. (2018). Generation of self-compatible diploid potato by knockout of S-RNase. Nat. Plants 4, 651–654. doi: 10.1038/s41477-018-0218-6
Yousfi, N., Slama, I., Ghnaya, T., Sayoure, A., and Abdelly, C. (2010). Effects of water deficit stress on growth, water relations and osmolyte accumulation in Medicago truncatula and M. laciniata populations. C. R. Biol. 333, 205–213. doi: 10.1016/j.crvi.2009.12.010
Yu, J., Holland, J. B., McMullen, M. D., and Buckler, E. S. (2008). Genetic design and statistical power of nested association mapping in maize. Genetics 178, 539–551. doi: 10.1534/genetics.107.074245
Yu, J., Jiang, M., and Guo, C. (2019). Crop pollen development under drought: from the phenotype to the mechanism. Inter. J. Mol. Sci. 20:1550. doi: 10.3390/ijms20071550
Yu, X., Li, X., Guo, T., Zhu, C., Wu, Y., Mitchell, S. E., et al. (2016). Genomic prediction contributing to a promising global strategy to turbocharge gene banks. Nat. Plants 2:16150. doi: 10.1038/nplants.2016.150
Zahran, H. H. (1999). Rhizobium-legume symbiosis and nitrogen fixation under severe conditions and in an arid climate. Microbiol. Mol. Biol. Rev. 63, 968–989. doi: 10.1128/mmbr.63.4.968-989.1999
Zair, W., Maxted, N., Brehm, J. M., and Amri, A. (2021). Ex situ and in situ conservation gap analysis of crop wild relative diversity in the Fertile Crescent of the Middle East. Genet. Resour. Crop Evol. 68, 693–709. doi: 10.1007/s10722-020-01017-z
Zamir, D. (2001). Improving plant breeding with exotic genetic libraries. Nat. Rev. Genet. 2, 983–989. doi: 10.1038/35103590
Zhang, H., Li, Y., and Zhu, J.-K. (2018). Developing naturally stress-resistant crops for a sustainable agriculture. Nat. Plants 4, 989–996. doi: 10.1038/s41477-018-0309-4
Zhang, H., Mittal, N., Leamy, L. J., Barazani, O., and Song, B. H. (2017). Back into the wild—Apply untapped genetic diversity of wild relatives for crop improvement. Evol. Appl. 10, 5–24. doi: 10.1111/eva.12434
Zohary, D., Hopf, M., and Weiss, E. (2012). Domestication of Plants in the Old World: The Origin and Spread of Domesticated Plants in Southwest Asia, Europe, and the Mediterranean Basin, 4th Edn. Oxford: Oxford University Press.
Zsögön, A., Cermak, T., Naves, E. R., Notini, M. M., Edel, K. H., Weinl, S., et al. (2018). De novo domestication of wild tomato using genome editing. Nat. Biotechnol. 36, 1211–1216. doi: 10.1038/nbt.4272
Keywords: abiotic stress, adaptation, breeding, crop wild relatives, legumes, marginal environment
Citation: Renzi JP, Coyne CJ, Berger J, von Wettberg E, Nelson M, Ureta S, Hernández F, Smýkal P and Brus J (2022) How Could the Use of Crop Wild Relatives in Breeding Increase the Adaptation of Crops to Marginal Environments? Front. Plant Sci. 13:886162. doi: 10.3389/fpls.2022.886162
Received: 28 February 2022; Accepted: 11 May 2022;
Published: 16 June 2022.
Edited by:
Diego Rubiales, Institute for Sustainable Agriculture (CSIC), SpainReviewed by:
Michael Benjamin Kantar, University of Hawai‘i, United StatesCopyright © 2022 Renzi, Coyne, Berger, von Wettberg, Nelson, Ureta, Hernández, Smýkal and Brus. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Juan Pablo Renzi, cmVuemlwdWduaS5qdWFuQGludGEuZ29iLmFy; Petr Smýkal, cGV0ci5zbXlrYWxAdXBvbC5jeg==; Jan Brus, amFuLmJydXNAdXBvbC5jeg==
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.