
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Plant Sci. , 28 July 2022
Sec. Plant Symbiotic Interactions
Volume 13 - 2022 | https://doi.org/10.3389/fpls.2022.884716
This article is part of the Research Topic Bio-based Solutions for Sustainable Development of Agriculture View all 17 articles
 Ailton Ferreira de Paula1,2
Ailton Ferreira de Paula1,2 Felipe de Paula Nogueira Cruz1,3Naiana Barbosa Dinato2,4
Felipe de Paula Nogueira Cruz1,3Naiana Barbosa Dinato2,4 Paulo Henrique Marques de Andrade1,2Amanda Carolina Prado de Moraes1,3Waldomiro Barioni Junior4
Paulo Henrique Marques de Andrade1,2Amanda Carolina Prado de Moraes1,3Waldomiro Barioni Junior4 Alberto Carlos de Campos Bernardi4
Alberto Carlos de Campos Bernardi4 Bianca Baccili Zanotto Vigna4Alessandra Pereira Fávero2,4
Bianca Baccili Zanotto Vigna4Alessandra Pereira Fávero2,4 Paulo Teixeira Lacava1,2,3*
Paulo Teixeira Lacava1,2,3*The genus Paspalum belongs to the family Poaceae and has several species that are native to Brazil. The Paspalum Germplasm Bank (GB) of the Brazilian Agricultural Research Corporation comprises approximately 450 accessions from 50 species. Among these accessions, Paspalum atratum (BGP 308) has economic potential for forage purposes. However, the endophytic and rhizospheric microbial communities within this accession and their ability to promote plant growth remain unknown. The present study aimed to isolate the endophytic and rhizospheric bacteria associated with P. atratum and to assess their potential for plant growth improvement, so-called plant growth-promoting bacteria (PGPB). For the in vitro tests, the ability of nitrogen-fixing bacteria (NFB), phosphate solubilization (PS) and indoleacetic acid (IAA) production were evaluated. A total of 116 endophytic and rhizosphere bacteria were obtained from the isolation. In the in vitro tests, 43 (37.00%) of these isolates showed positive NFB, PS, and IAA results. These isolates were identified by 16S rDNA sequencing. The phosphate solubilization index (PSI) ranged from 2 to 3.61, all 43 strains performed biological nitrogen fixation and the IAA production ranged from 12.85 to 431.41 μg ml−1. Eight of these 43 isolates were evaluated in vivo in a greenhouse using P. atratum caryopsis. The pots were filled with soil prepared with three different phosphate sources and one control without phosphate. After growth, the plants were submitted to morphological, bromatological and chemical determination. Data were analyzed using analysis of variance (ANOVA) and principal component analysis (PCA). In the in vivo test, treatments 105 (Pseudomonas sp.) and 458 (Pseudomonas sp.) were the most significant for the crystalline phosphate source, 109 (Bacillus sp.) for the sedimentary phosphate source and, as for the soluble phosphate source most treatments that received bacterial isolates had higher phosphorus content in the dry matter than the uninoculated soluble phosphate control. The 105FCR (crystalline phosphate + Pseudomonas sp.), 109FSE (sedimentary phosphate + Bacillus sp.), and 110 FSE (sedimentary phosphate + Enterobacter sp.) treatments showed the best results for plant growth promotion. This work made it possible to determine the bacterial community associated with P. atratum (BGP308) and to obtain new potential plant growth-promoting strains.
Pasture is the main form of animal nutrition used for herds. Due to the potential for growth in different soils and climate conditions and requiring little management (Rodrigues et al., 2014), the genus Urochloa currently constitutes the majority of Brazilian pastures used for animal feed. The main problem in regard to Urochloa spp. is exposure to ecological imbalances owing to its low genetic variability (Compant et al., 2010). However, Paspalum is an important and highly diverse genus in the Poaceae family in the Americas (Novo et al., 2016), occurring throughout Brazil, Bolivia, Paraguay, Argentina, Chile and Uruguay (Zuloaga and Morrone 2005; Novo et al., 2019). Therefore, Paspalum spp. could potentially replace pastures composed of Urochloa spp. or could occupy areas where these grasses do not grow. Brazil is one of the largest meat producers and exporters globally; therefore, there is a need to research new sources and ways to carry out animal nutrition. According to the Brazilian Association of Meat Exporting Industries (ABIEC), Brazil exported approximately 8.50 million tons of beef in 2020 and 4.38 million tons in the first half of 2021, significantly contributing to Brazil’s economy (ABIEC, 2021).
Another critical point is that with the increase in cultivated areas, agrochemical use and environmental impacts also increase. Thus, alternative nonpolluting and more economical methods of promoting plant growth have gained greater attention (Coelho et al., 2019; Xiang et al., 2020).
Phosphate fertilizers are among the most commonly used agrochemicals in agriculture, and the lack of adequate levels of these fertilizers is a limiting factor for plant growth (Crous et al., 2019). Therefore, plants usually receive soluble phosphorus through industrialized fertilizers. In addition, some phosphate rocks can be applied for direct soil fertilization (Ahemad and Kibret, 2014). First, however, it is necessary to transform this phosphorus into a soluble form for plants. Phosphate-solubilizing bacteria (PSB) transform this phosphorus by chelation, ion exchange, and organic acid production (Khan et al., 2009; Afzal et al., 2019). Among the types of insoluble phosphate rocks are the sedimentary rocks of Arad (33.0% P2O5) and the crystalline rocks of Cajati/SP (5.0% P2O5) (Alves and Hagni, 2008; Ramos et al., 2009).
Paspalum accessions from the germplasm bank of Embrapa Pecuaria Sudeste have been evaluated for different uses, such as forage (Marcón et al., 2018) and turf (Souza et al., 2020). The studies developed with Paspalum plants from this GB have included characterization regarding shade stress (Barro et al., 2012), water stress (De Pezzopane et al., 2017), insect tolerance (Gusmão et al., 2016), and cryopreservation for new hybrid production (Dinato et al., 2018). However, studies related to the endophytic and rhizospheric bacterial microbiota diversity associated with this germplasm collection are still scarce.
Among the Paspalum accessions evaluated in the Brazilian Agricultural Research Corporation breeding program, BGP 308 from P. atratum Swallen is a promising accession for becoming a forage cultivar and for composing the preliminary studies of the endophytic and rhizospheric microbiota.
The analysis of the bacterial diversity associated with this species can indicate new microorganisms to be used for plant growth promotion in the forage plant P. atratum. Thus, to collaborate with works that seek alternative and sustainable ways to use phosphorus to avoid the environmental and economic impacts caused by the industrial process of obtaining industrial phosphorus.
The objective of this study was to search for phosphate-solubilizing bacteria, evaluate their ability to solubilize phosphate rocks, and promote plant growth.
The present study is the first report to examine phosphate rock in the nutrition of P. atratum plants, intermediated by cultivable endophytic and rhizospheric bacteria.
The work was divided into steps in vitro and in vivo. In the in vitro stage, there was isolation, identified and evaluated of the functional capacity of the plant growth-promoting bacterial strains. The three main characterizations performed were for phosphate solubilization (PS), nitrogen-fixing bacteria (NFB) and indoleacetic acid (IAA) production. In the in vivo stage, P. atratum plants were characterized in a morphological, nutritional and mineral manner (emphasizing phosphate solubilization), when inoculated or not with plant growth-promoting bacterial strains.
Samples (rhizospheric soils, roots, and leaves) from an adult plant of P. atratum BGP 308 (BRA 030078/VRcMmSv 14,525) were collected in August 2016 (dry season) and January 2017 (rainy season). Access BGP 308 belongs to the Paspalum germplasm bank, located at Brazilian Agricultural Research Corporation1 (21°57′42″S, 47°50′28″W, 860 m), São Carlos, SP, Brazil.
The endophytic bacterial community was isolated according to Araújo et al. (2014) and Bogas et al. (2015). Plant tissues were superficially disinfected by serial washes in 70% ethanol (EtOH) for 2 minutes, followed by 3% sodium hypochlorite for 3 minutes, 1 minute in 70% EtOH, and two rinses with sterile distilled water. Plant tissues were incubated in phosphate buffered saline (PBS) for 2 h at 28°C/200 rpm. Aliquots of 100 μl of decimal dilutions were inoculated in duplicate in plates containing tryptone soya agar (TSA) supplemented with Benlate (50 μg ml−1) to prevent fungal growth.
The isolation of rhizospheric bacteria was performed according to Mohite (2013) with modifications, in which the temperature used was 28°C, and aliquots of 100 μl of decimal dilutions were inoculated in duplicate in plates containing tryptic soy agar (TSA) supplemented with Benlate (50 μg ml−1). Bacterial cultures were preserved in tryptone soya broth (TSB) supplemented with glycerol (1:1) at −80°C until further study.
Total DNA was extracted according to Aljanabi and Martinez (1997). The 16S gene was amplified using the primers V3F (5′-ACTCCTACGGGAGGCAGCAG-3′) and V6R (5′ ACAGCCATGCANCACCT 3′; Yang et al., 2016). Polymerase chain reaction (PCR) containing 60 ng of genomic DNA, 25 μl of Thermo Scientific PCR Master Mix (1.25 U of Taq polymerase enzyme, 1 × PCR buffer (200 mM Tris pH 8.4, 500 mM KCl), 50 mM MgCl2, and 1.25 mM dNTP) and 3 pmol of each primer was performed for the selected isolates. The reaction conditions consisted of an initial 95°C step for 3 min, followed by 31 cycles of 95°C for 30 s., 60°C for 30 s., 72°C for 1 min. and a final extension of 10 min at 72°C in a BioRad T100 thermocycler. The amplicons were examined by 0.7% agarose gel electrophoresis and purified by using a QIAquick (Qiagen) kit. The sequencing reactions were performed using the BigDye® Terminator v3.1 Cycle Sequencing Kit (Thermo Fisher Scientific) and sequenced using an ABI 3730 DNA Analyzer. Sequences were analyzed by Sequencing Analysis 5.3.1 software using the Base Caller KB, and the low-quality sequences were visualized and edited using BioEdit software (Hall, 1999). The final sequence was subjected to BLASTn bacterial identification (Altschul et al., 1997; Maidak et al., 2000; Garrity et al., 2004).
The phylogenetic tree was obtained by multiple alignments of P. atratum endophytes and 16S rRNA sequences were retrieved from NCBI using the algorithm ClustalW and generated using MEGA X software (Kumar et al., 2018) using the maximum likelihood method in combination with a general time reversible model with 1,000 bootstrap replications as branch support (Saitou and Nei, 1987; Tamura et al., 2011). Halobacterium sp. (NR_113428.1) sequence was used as an outgroup.
Bacteria isolated from P. atratum were qualitatively/quantitatively screened in triplicate for their ability to solubilize inorganic calcium phosphate. Strains were incubated in nutrient agar supplemented with Ca3(PO4)2 for 96 h at 28°C (Rodríguez and Fraga, 1999; Verma et al., 2001). As a result, the phosphate solubilization index (PSI) was calculated as the ratio of the total diameter (colony + halo zone) to the colony diameter and was classified as low (PSI < 2), medium (2 < PSI < 3), and high (PSI > 3; Silva Filho and Vidor, 2000; Pande et al., 2017).
The nitrogen-fixing bacteria (NFB) assay was carried out by growing the strains in a semisolid nitrogen-free medium twice for 72 h/28°C (Döbereiner et al., 1995). For the quantification of auxin production, the strains were grown in the broth tryptone de soy 10% medium supplemented with L-tryptophan for 72 h/28°Ca. This method was initially proposed by Bric et al. (1991) and adapted as a quantitative method (Husen, 2016).
Caryopsis of P. atratum were germinated and inoculated with eight different strains that presented PSI ≥ 2.0. The experiment was conducted in a greenhouse at the Brazilian Agricultural Research Corporation (see footnote 1), São Carlos, SP, Brazil. A randomized block design with three replications was used in a factorial scheme of 8 × 3 + 1 with pots containing 0.5 l and 4.5 l of soil.
The soil was prepared with three different sources of phosphate plus the control without phosphate, totaling 36 treatments and 108 pots. The three sources of P used were (1) soluble phosphate—triple granulated superphosphate (46.0% P2O5 soluble in neutral ammonium citrate + water), (2) sedimentary phosphate—Arad phosphate rock concentrate (33.0% P2O5 total), and (3) crystalline phosphate—Cajati phosphate rock concentrate (5.0% P2O5 total) with a dose of 200 mg kg−1 P or 458 mg kg−1 P2O5.
The chemical characteristics of the soil were determined according to Van Raij et al. (2001). Based on the soil analysis results, dolomitic limestone (total neutralizing power ratio, TNPR = 70%) was added to achieve a base saturation of 60% prior to transplanting. Then, the three phosphorus sources were applied at the transplanting of the seedlings, and the pots were fertilized with K2SO4 (60% K2O) until K reached 3% of the cation exchange capacity (CEC).
The caryopses of the spikelets were removed and subjected to the disinfection process in a closed desiccator using the protocol described by Quesenberry et al. (2010). The caryopses were inserted into 16 × 100 mm test tubes containing Murashige and Skoog (MS) medium according to the Orbovic and Grosser (2006) seed germination protocol. After 14 days, the seedlings were removed from the test tube and inserted into a Falcon tube with 15 ml of bacterial suspension for 30 min at 28°C. The liquid MS medium was standardized at 109 CFU/ml and used in this step. Thus, seedlings were transplanted into 500 ml pots containing limed soil without correction of nutrients. Five more inoculations were performed: in 500 ml pots, they received two more inoculations (15 and 30 days after transplant), and in 4.5 kg pots, they received three more inoculations (45, 60, and 75 days after transplant). Finally, the 6 ml volume of the standardized bacterial suspension was inoculated into the soil close to each plant’s root. The control was inoculated with phosphate buffered saline (PBS) solution without the bacterial isolate.
The seedlings remained in the 500 ml pots for 43 days and were transplanted into the 4.5 kg pots for the four treatments. At 50 days after transplanting, the aerial parts of the plants were cut 15 cm from ground level and measured.
The aerial part samples were placed in an oven with forced circulation at 60°C for 72 h. After drying the samples and determining the dry leaf weight, the material was crushed in a Wiley mill with 1 mm sieves. The collected material was stored in a plastic bottle.
Thirty-two descriptors were evaluated: EP (phosphorus extract), EN (nitrogen extract), ECa (calcium extract), CP (crude protein), MM (mineral matter), LIG (lignin), Ca (calcium), Mg (magnesium), P (phosphorus), K (potassium), S (sulfur), Mn (manganese), Zn (zinc), N (nitrogen), SPAD (SPAD index), AFW (aerial fresh weight), ADW (aerial dry weight), LW (leaf width), NL (number of leaves), NT (number of tillers), ANT (presence of anthocyanin), LA (leaf area), EZn (zinc extract), DIV (in vitro digestibility), FDN (neutral fiber detergent), Ll (leaf length), PHe (plant height), Fe (iron), FDA (acid detergent fiber), MS (dry matter), EE (ether extract) and Cu (copper). Dry matter, crude protein, neutral detergent fiber, acid detergent fiber, lignin, and in vitro digestibility were determined using a near-infrared spectrometer (NIRS; Büchi Labortechnik, 2007) with a calibrated curve for Paspalum.
Based on Nogueira et al. (1998), the total nutrient content was determined. Nitrogen was determined in the extract of sulfuric digestion by the semimicro Khjeldhal method. The determination of K was made in the extract of nitro-perchloric digestion and determined by flame photometry. The other macronutrients (P, Ca, Mg, and S) and micronutrients (Cu, Fe, Mn, and Zn) were determined in the same nitro-perchloric extract and determined by induced plasma spectrometry (ICP–OES).
Thirty-two traits (morphological, nutritional, and mineral) were analyzed. Data were analyzed using analysis of variance (ANOVA) and principal component analysis (PCA) with SAS® 9.3 software (SAS Institute Inc, 2011).
In the rainy season isolation, the bacterial population ranged from 02 × 10−1 cfu gm−1 (leaf) to 43 × 10−3 cfu gm−1 (root), whereas in the dry season isolation, the bacterial population ranged from 05 × 10−1 cfu gm−1 (leaf) to 35 × 10−2 cfu gm−1 (rhizosphere; Table 1). In the rainy season, the soil had a pH of 5.4 (water) and 4.8 (CaCl2), and in the dry season, it was 5.6 (water) and 5.0 (CaCl2).
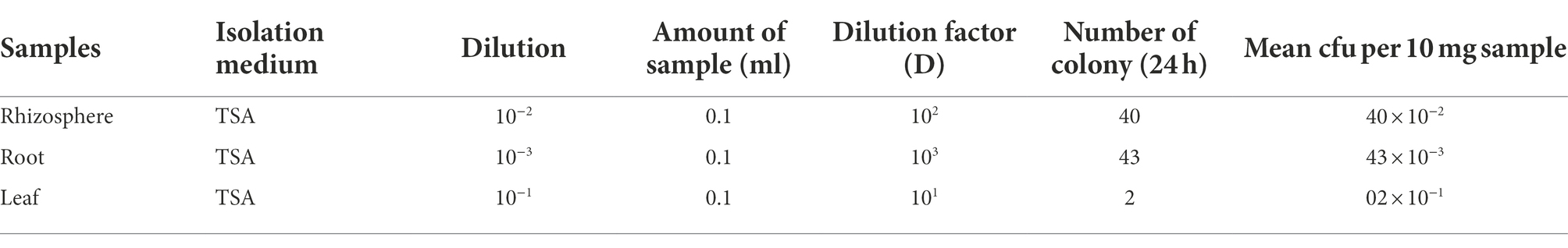
Table 1. Isolation of endophytic and rhizospheric bacteria from soil samples.
A total of 116 bacterial isolates were collected from P. atratum BGP 308, where 43 (37.00%) were obtained from rhizospheric soils, 42 (36.20%) from roots, and 31 (26.70%) from leaves. A total of 74 (63.70%) strains were isolated in the rainy season and 42 (36.20%) in the dry season.
Among the 116 strains obtained, 43 (37.00%) showed positive NFB, SF, and IAA results. From these, the strains that belonged to the genera Enterobacter (46.50%), Pseudomonas (32.50%), and Pantoea (13.90%) were the most abundant (Figure 1). On the other hand, Bacillus, Microbacterium, and Micrococcus strains represented only 6.90%. The phylogenetic analysis showed highly significant support (>98%) for the groups formed of samples from each genus and its correspondent reference sequences for the genus. Moreover, nodes forming a group with all the samples from genera Enterobacter, Pantoea and Pseudomonas and another group with all the samples from genera Bacillus, Microbacterium and Micrococcus also showed significant support (>90%), showing the relation among genera identified in this study (Figure 1).
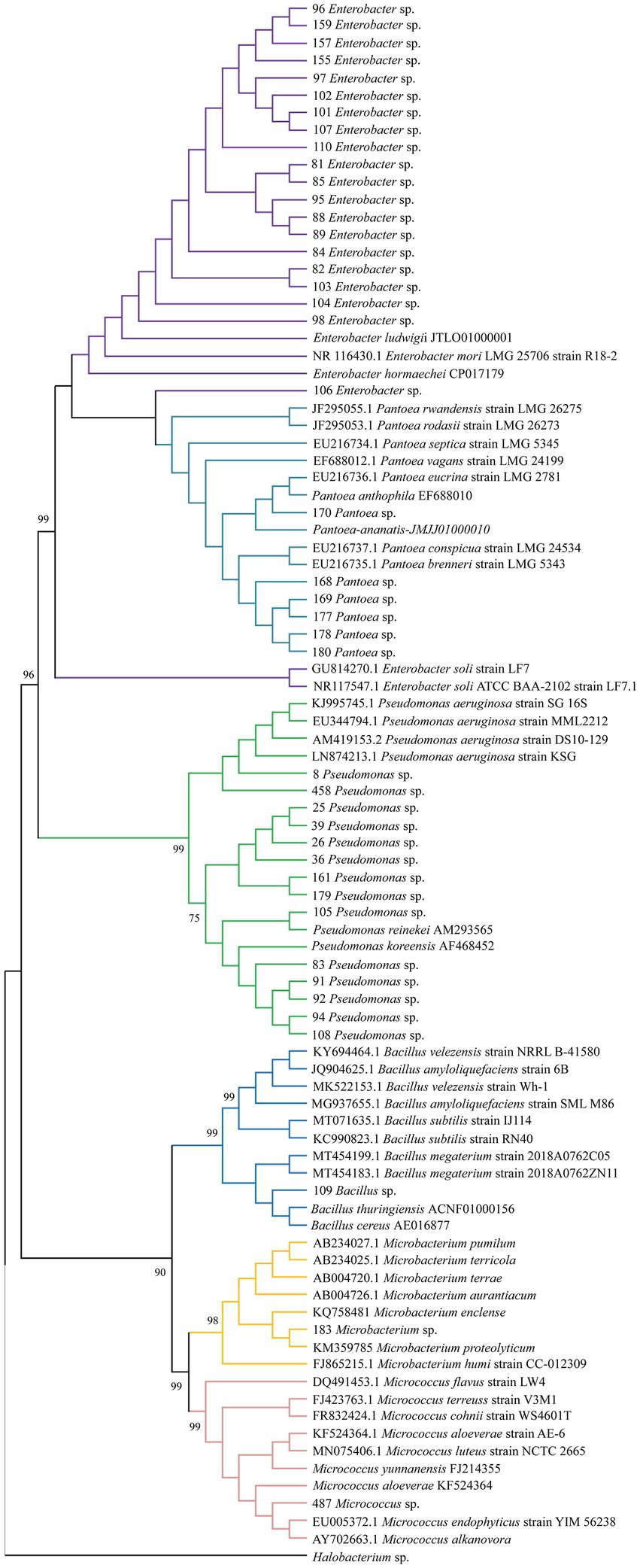
Figure 1. Consensus tree obtained from a maximum likelihood phylogenetic analysis using the general time reversible model (bootstrap with 1,000 replicates) based on 670 bp of the partial 16S rDNA gene.
The phosphate solubilization index (PSI) ranged from 2 to 3.61 (Table 2). Strains 103, 89, and 102, which are root endophytes and belong to the Enterobacter genus, showed the best results, with PSI values of 3.61, 3.58, and 3.56, respectively. All 43 strains in Table 2 performed biological nitrogen fixation, characterized by a semisolid nitrogen-free medium. The IAA production ranged from 12.85 to 431.41 μg ml−1. The endophytic leaf strain 170 (Pantoea sp.) showed the highest IAA production (431.41 μg ml−1), followed by 18 Enterobacter spp.; endophytes from roots presented 105.05–263.74 μg ml−1 of IAA production.
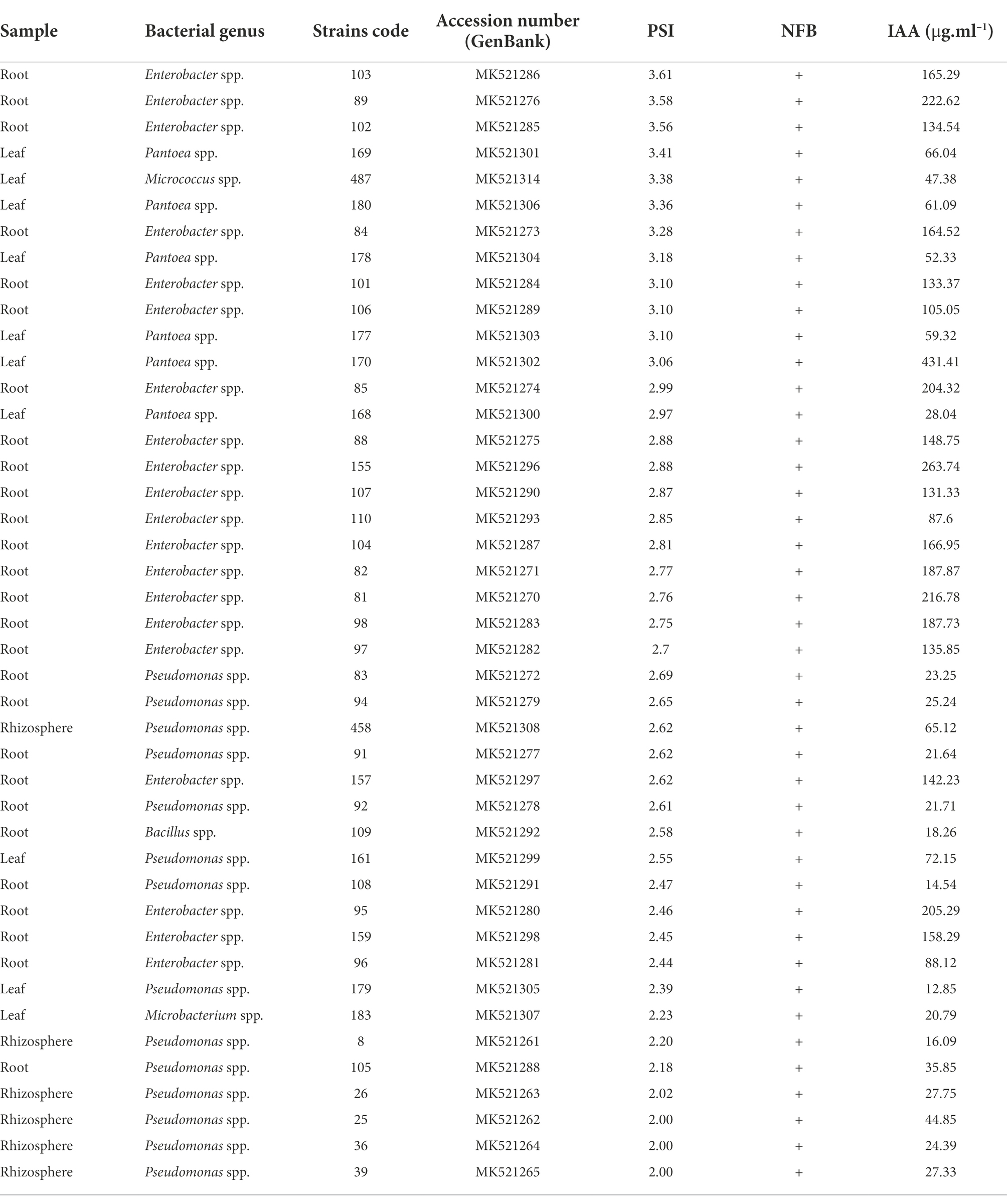
Table 2. Molecular identification of the genera of the 43 bacterial strains that showed positive results for the phosphate solubilization index (PSI), nitrogen-fixing bacteria (NFB), and indole acetic acid (IAA) production.
All strains belonging to Microbacterium, Micrococcus, Pantoea, Bacillus, and Enterobacter were endophytes, while the Pseudomonas strains were either endophytic or rhizospheric. Among the 43 strains selected for the in vitro tests, six were isolated from the rhizosphere, 27 from the root, and 10 from the leaf (Table 3).
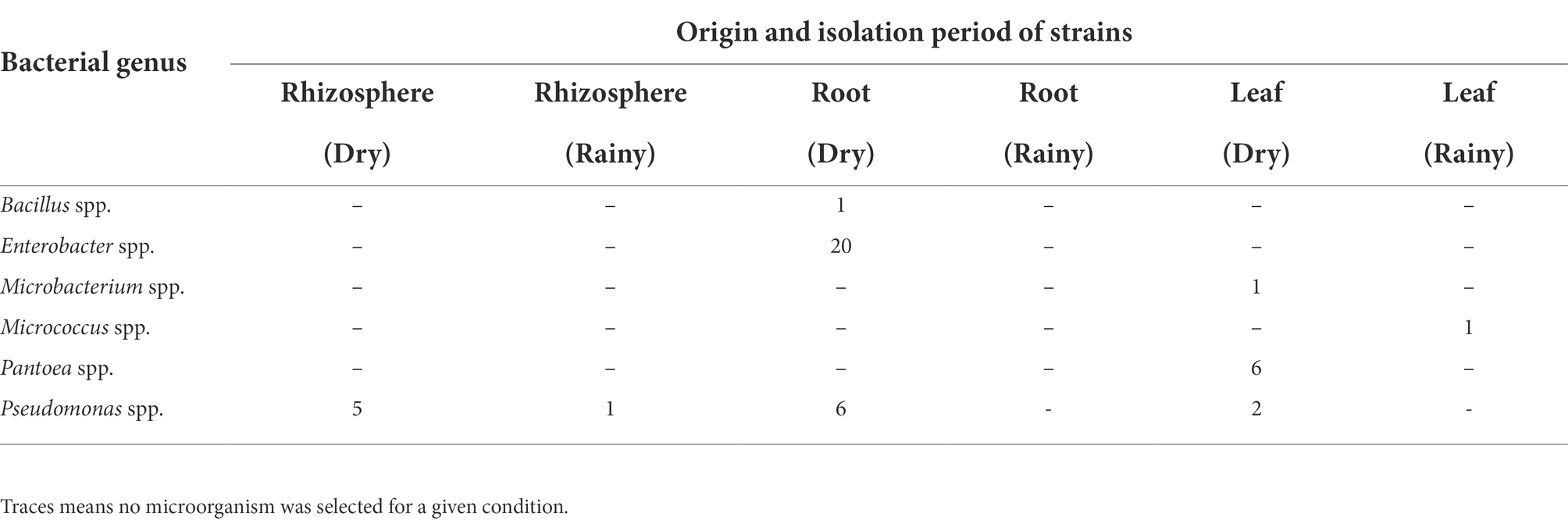
Table 3. Bacterial genus, number of strains, origin and isolation period of strains of BGP 308 from P. atratum that were positive for phosphate solubilization, biological nitrogen fixation, and indole acetic acid production.
In the plant growth promotion assay, the selected strains belonged to the rhizosphere, root and leaf. The PSI ranged from 2 to 3.61, and all were positive for NFB and IAA. The rhizospheric bacteria selected for the in vivo test were Isolates 25 (Pseudomonas sp.) and 458 (Pseudomonas sp.), and the endophytic bacteria were Isolates 103 (Enterobacter sp.), 105 (Pseudomonas sp.), 109 (Bacillus sp.), 110 (Enterobacter sp.), 161 (Pseudomonas sp.), and 170 (Pantoea sp.).
The triple interaction [Source of Phosphorus (3) vs. Isolate (9) vs. Cut (3)] for Phosphorus extract (P) was significant with p-value = 0.0023.
The P extract showed that in treatments with crystalline phosphate, the mean phosphorus content in dry matter ranged from 0.90 to 3.82 kg ha–1, considering treatments and cuts (Figure 2). The treatments that received strains 105 (Pseudomonas sp.) and 458 (Pseudomonas sp.) had a higher phosphorus content than the control crystalline phosphate in the first cut and showed a drop in the second third cut. The treatment containing Strain 110 (Enterobacter sp.) presented a high level of phosphorus in the second cut compared to the control. Within this treatment, P extract increased in the second cut, and decreased it in the first and third cut. Despite the numerical variations, in the phosphorus content, there was no statistical difference (p > 0.05). In treatments with sedimentary phosphate, the mean phosphorus content in dry matter ranged from 0.89 to 11.82 kg ha–1, considering treatments and cuts. The treatment that received the bacterial isolate 109 (Bacillus sp.) had a higher phosphorus content than the sedimentary phosphate control in the first cut (p > 0.05). Within this treatment, the phosphorus content was higher in the first cut, followed by the second and third cut (p ≤ 0.05). The treatment that received strain 103 (Enterobacter sp.) also showed an increase in phosphorus content in the first cut. In the other treatments there were no (p > 0.05) significant changes in the phosphorus content in the cuts.
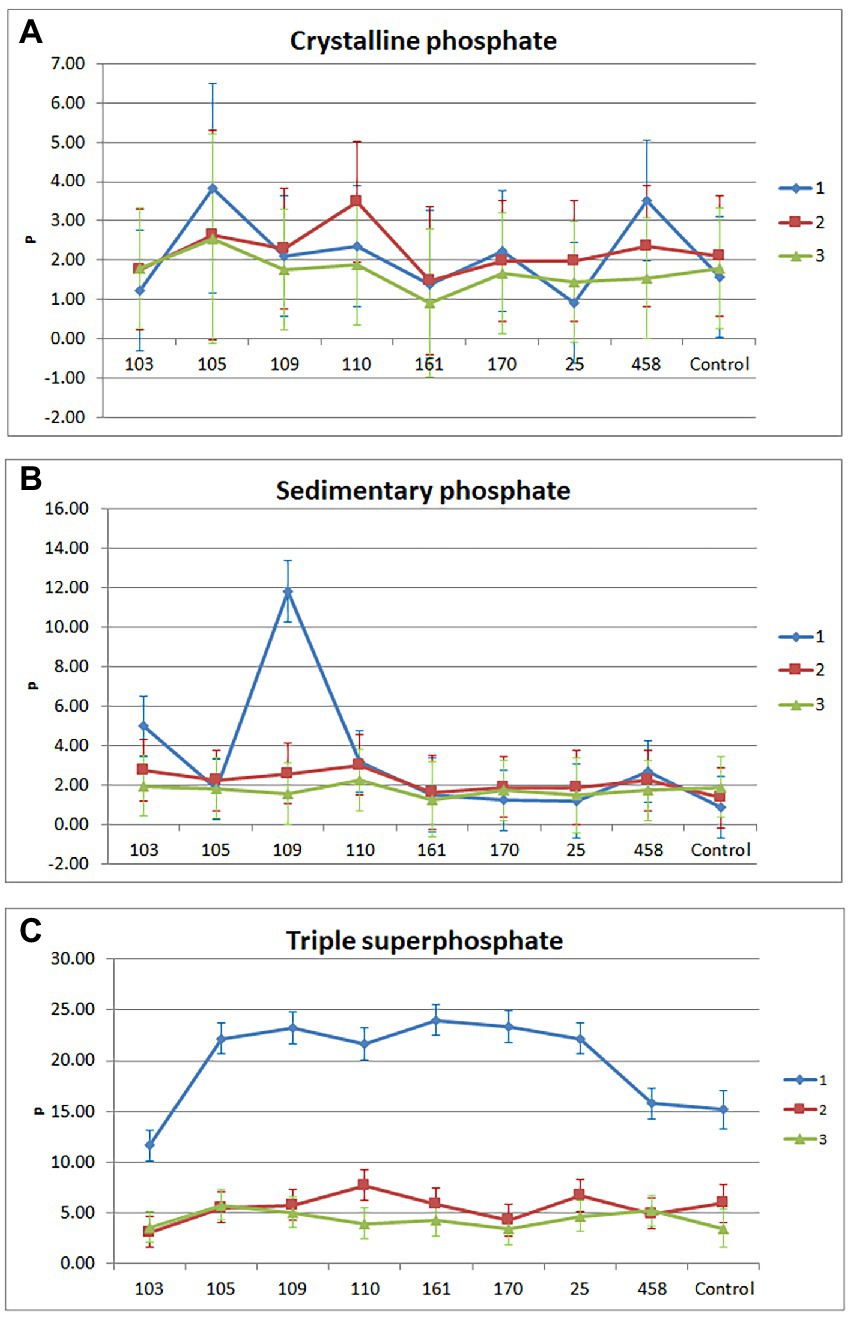
Figure 2. Triple interaction (source of phosphorus vs. isolate vs. cut). Treatments that received crystalline (A), sedimentary (B), and soluble (C) phosphate as a source of phosphorus.
In treatments with soluble phosphate, the mean phosphorus content in dry matter ranged from 3.14 to 24.00 kg ha–1, considering treatments and cuts. Regardless of treatment, the phosphorus content was higher in the first cut (p ≤ 0.05). In this cut, the lowest phosphorus content in the dry matter was in the treatment that received the strain 103 (Enterobacter sp.) and the highest was in the treatment that received the strain 161 (Pseudomonas sp.) with 11.65 and 24.00 kg ha–1, respectively. The treatment that received the bacterial isolate 110 (Enterobacter sp.) showed a significant difference (p ≤ 0.05) between the three cuts, for the phosphorus content, with 21.66, 7.73 and 3.98 kg ha–1, respectively. The phosphorus content, in cut1, for the control of uninoculated soluble phosphate was 15.21 kg ha–1. In cuts 2 and 3, the maximum phosphorus content reached was 7.73 kg ha–1, regardless of the treatments.
The results of the principal component analysis (PCA) showed that Component 1 (PRIN1 = 65.31%) and Component 2 (PRIN2 = 8.52%) explained 73.83% of the variance observed. A total of 22 variables were significant (correlation > 50%) in the discrimination of treatments (Table 4) among the 32 variables used.
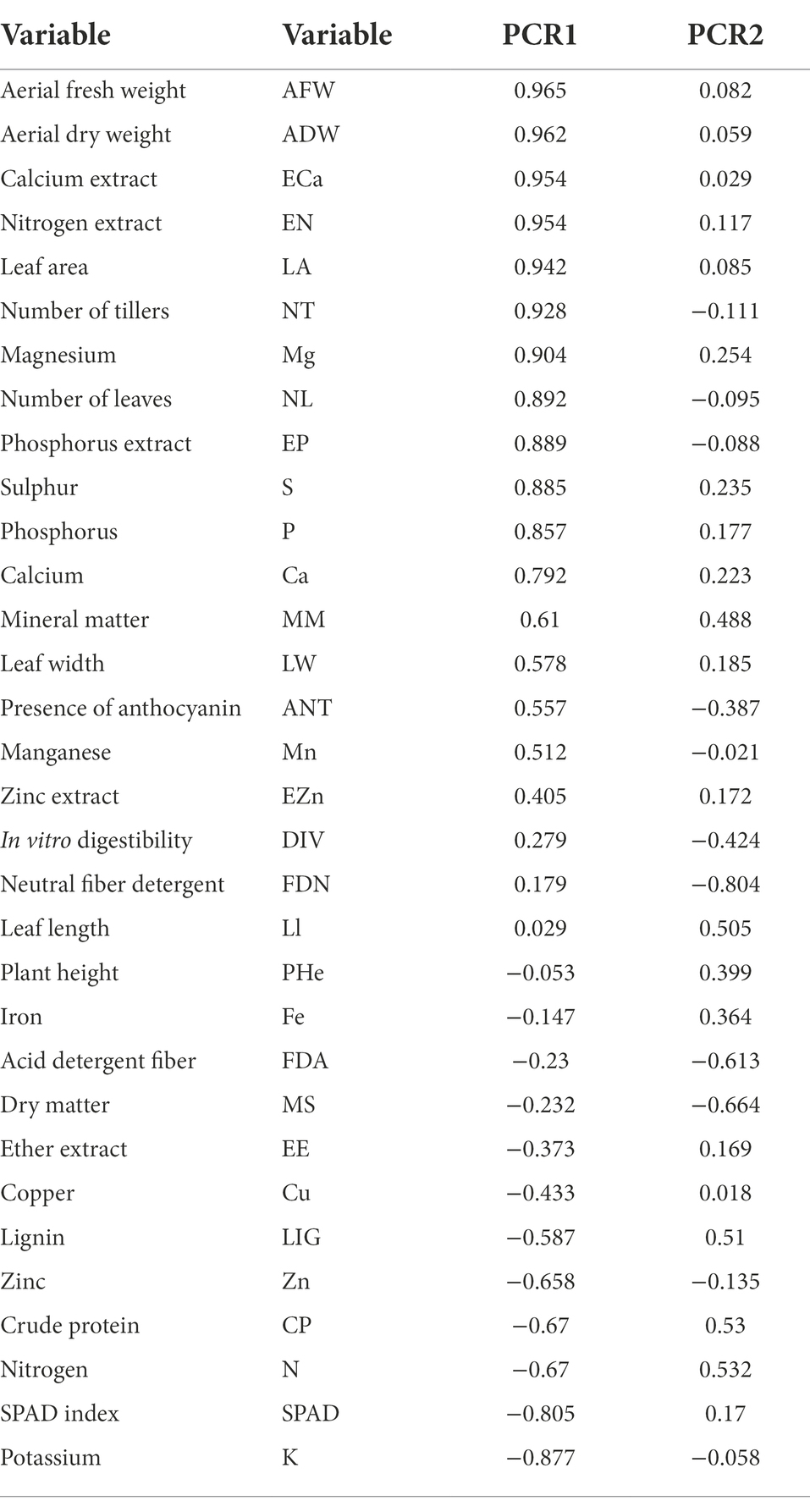
Table 4. Contribution of morphological descriptors for principal component analysis.
When evaluating the phosphate source used in the experiment, the analyzed variables divided the treatments into two groups (Figure 3), directed by Principal Components 1 (PRIN1) and 2 (PRIN2). The treatments that received soluble phosphate are distributed on the left side of the graph. In contrast, the treatments without phosphate (control) and those receiving sedimentary and crystalline phosphate are distributed on the right side. For the most significant morphological, mineral and bromatological variables, the principal component analysis showed that Principal Component 1 had significant associations with the SPAD index (SPAD), potassium (K), zinc (Zn), crude protein (CP), lignin (LIG) and nitrogen (N) (Figure 2). Furthermore, these variables were responsible for grouping the nine treatments (103 FSO, 105 FSO, 109 FSO, 110 FSO, 161 FSO, 170 FSO, 25 FSO, 458 FSO, and CAFSO) that received the soluble phosphate source in Group 1.
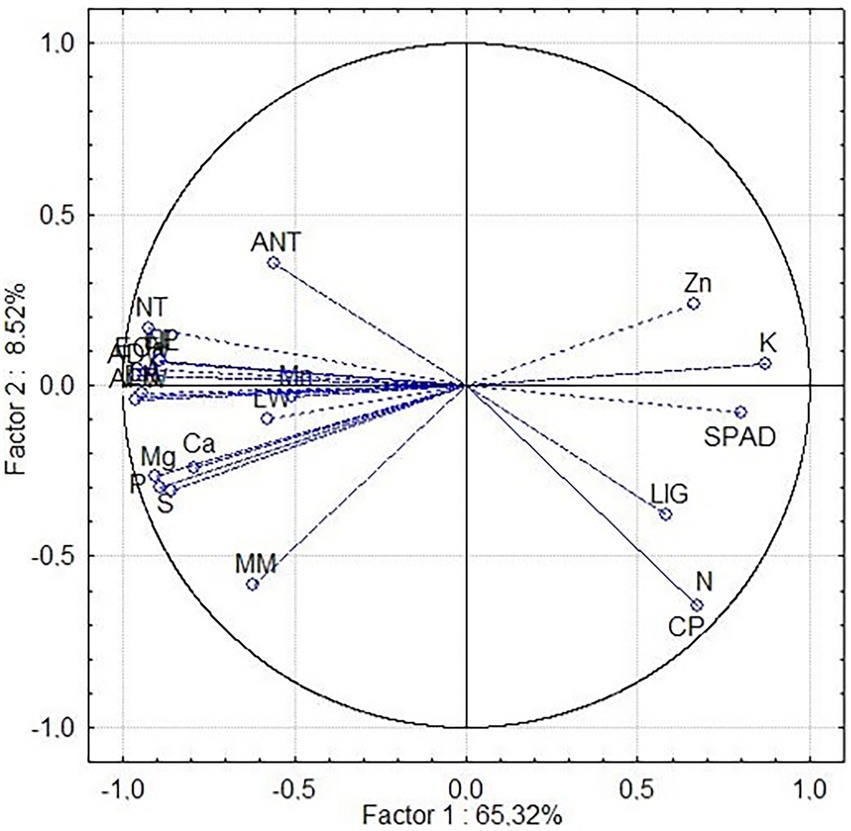
Figure 3. Principal component analysis of the 22 descriptors evaluated. Correlation between the evaluated descriptors. Phosphorus extract (EP), nitrogen extract (EN), calcium extract (ECa), crude protein (CP), mineral matter (MM), lignin (LIG), calcium (Ca), magnesium (Mg), phosphorus (P), potassium (K), sulfur (S), manganese (Mn), zinc (Zn), nitrogen (N), SPAD index (SPAD), aerial fresh weight (AFW), aerial dry weight (ADW), leaf width (LW), number of leaves (NL), number of tillers (NT), presence of anthocyanin (ANT), and leaf area (LA).
Principal Component 2 showed strong associations with aerial fresh weight (AFW), aerial dry weight (ADW), leaf width (LW), number of leaves (NL), number of tillers (NT), anthocyanin (ANT), area leaf (LA), phosphorus extract (EP), calcium extract (ECa), calcium (Ca), magnesium (Mg), phosphorus (P), sulfur (S), manganese (Mn), nitrogen (EN) and mineral matter (MM) (Figure 2). These variables grouped the 27 treatments (103FCR, 103FSE, 103SF, 105FCR, 105FSE, 105SF, 109FCR, 109FSE, 109SF, 110FCR, 110FSE, 110SF, 161FCR, 161FSE, 161SF, 170FCR, 170FSE, 170SF, 25FCR, 25FSE, 25SF, 458FCR, 458FSE, 458SF, CAFCR, CAFSE and CASF) that did not receive the soluble phosphate source in Group 2.
Tables 5, 6 describe the order of descriptors that most contributed to the morphological variation observed in Principal Component 1 (PRIN 1) of the principal component analysis (PCA) of all treatments evaluated in this work. Analyzing the most important descriptors (SPAD, K, Zn, CP, LIG and N), it can be observed that the treatments that received soluble phosphate presented lower values (Tables 5, 6) for these descriptors, and this located these treatments in group 1 (Figure 3). The inverse of values for these descriptors are represented in the procedures that they located in group 2.
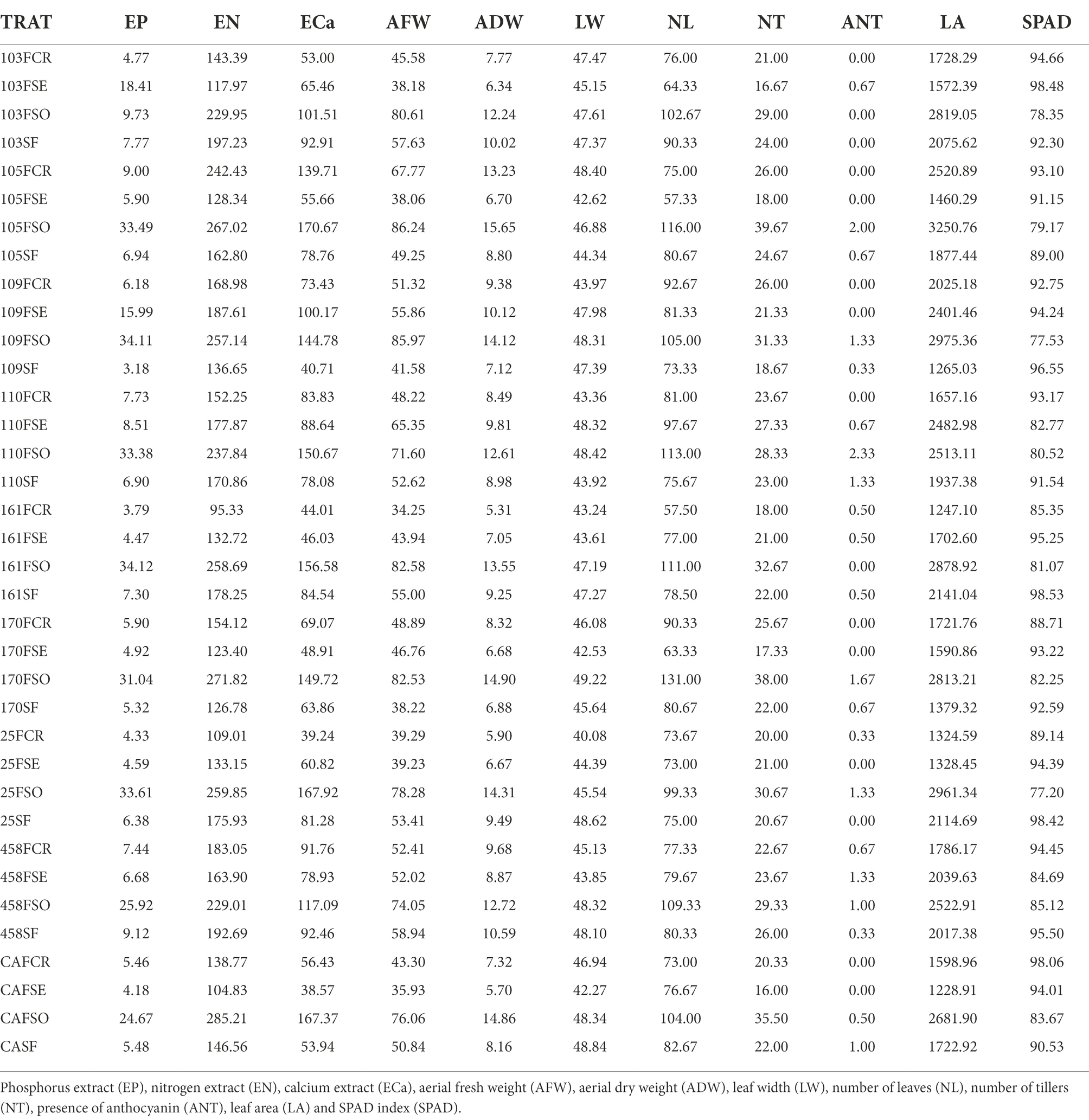
Table 5. Values of extracts and morphological descriptors evaluated on average.

Table 6. Values of mineral and bromatological descriptors evaluated on average.
In Figure 4, the principal component graph shows the two groups and treatments closest to those receiving phosphorus from the soluble phosphate source. The group on the right side of the graph belongs to treatments with soluble phosphate. In contrast, the group to the left and central part of the graph belongs to treatments containing sedimentary phosphate, crystalline phosphate, and no phosphate (control). Any treatment that received sedimentary and crystalline phosphate was not observed, composing the group of those that received the soluble phosphate. Nevertheless, the ones that came closest were the 105FCR (crystalline phosphate + Pseudomonas sp.), 109FSE (sedimentary phosphate + Bacillus sp.), and 110FSE (sedimentary phosphate + Enterobacter sp.) treatments.
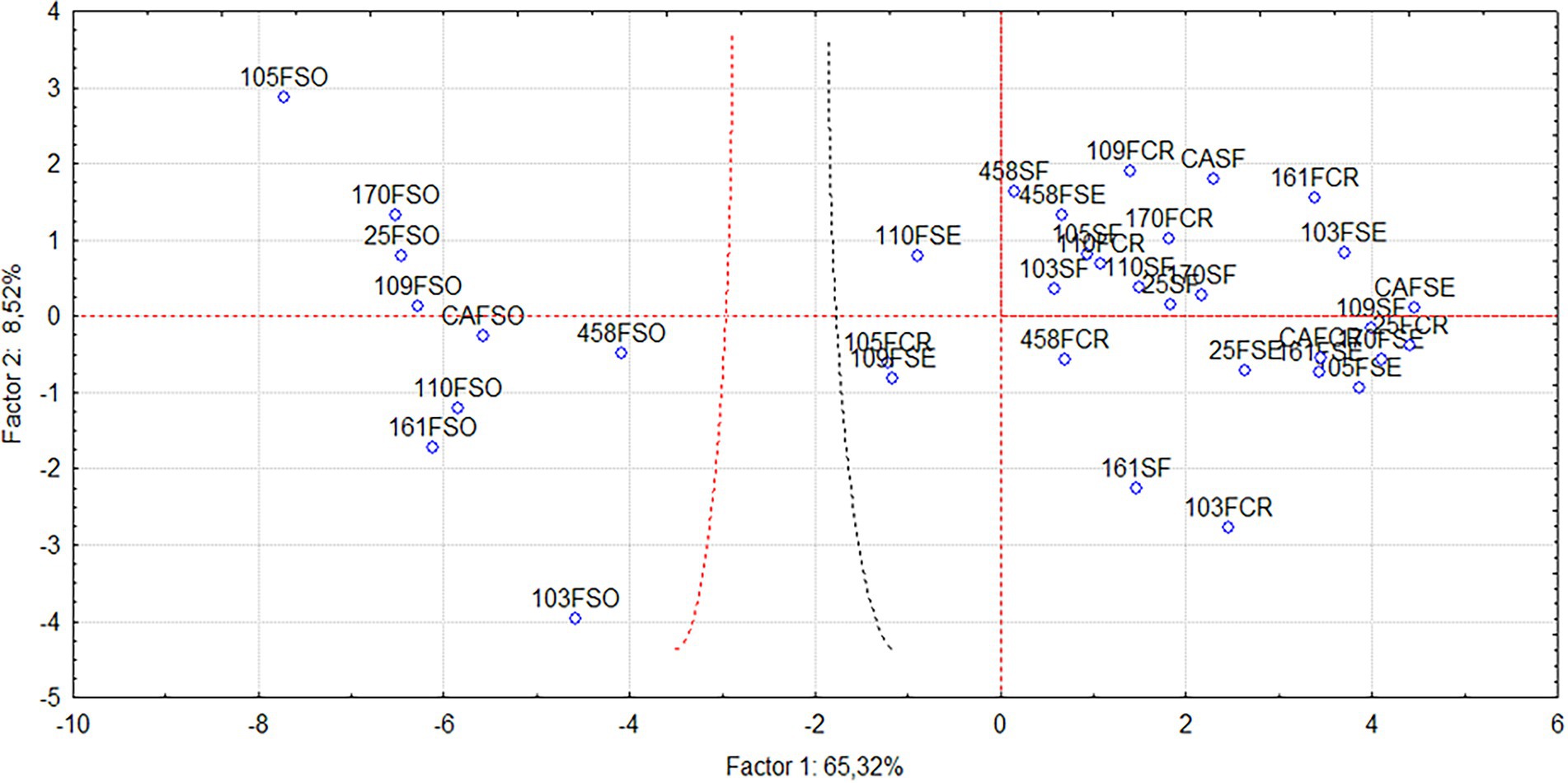
Figure 4. Biplot graph resulting from the 36 treatments obtained by principal component analysis considering the 22 descriptors for Principal Components 1 and 2. Control (CA), crystalline phosphate (FCR), sedimentary phosphate (FSE), phosphate-free (SFO), and soluble phosphate (FSO). The number that precedes the treatment refers to the strain code.
In the production of pastures for animal nutrition, there is great concern regarding the cultivation of plants. For cultivation, plants obtain most of their nutrients from some industrialized source. In 2018, Brazil consumed 852.4 thousand tons of triple superphosphate (largest consumer on a global scale) and 2,437.7 thousand tons of ammonium phosphate (third consumer on a global scale). For nitrogen, phosphorus, and potassium (NPK) fertilizer, Brazil consumed 15507.1 thousand tons and imported 13338.7 thousand tons, the largest importer of NPK on a worldwide scale (International Fertilizer Industry Association, 2021). Microorganisms can provide nutrients from alternative and nonindustrialized sources to plants. The main focus of this work was to address phosphate solubilization by plant growth-promoting bacteria (PGPB).
Studies involving the endophytic and rhizospheric microbiota of Paspalum accessions are scarce. Zhao et al. (2021) isolated endogenous Enterobacter strains from Paspalum vaginatum and evaluated their influence on promoting salt tolerance in the plant. Pérez and Martínez (2016) isolated and identified endophytic bacteria that is resistant to mercury associated with P. arundinaceum in Mina Santa Cruz, Bolivar., Colombia, aiming to obtain plant growth-promoting strains with potential for phytoremediation of mercury-contaminated soil. Amaral et al. (2021) isolated and characterized plant growth-promoting rhizobacteria (PGPR) from 10 genotypes of Paspalum and evaluated the effect of inoculation on P. regnellii, P. atratum, and P. malacophyllum. The genotypes were also collected from the Paspalum germplasm bank at the Brazilian Agricultural Research Corporation; however, the genotype of P. atratum studied was cv. Pojuca BGP 098. For studies involving phosphate solubilization in Paspalum with potential for forage, this work is the first to examine phosphate rock in the nutrition of P. atratum plants, intermediated by cultivable endophytic and rhizospheric microorganisms.
Based on the present study results, the bacterial population was higher in the rhizosphere and the root. The rhizosphere exhibits different physical and chemical characteristics than nonrhizospheric soil, as plants can produce root exudates, which provide bacterial nutrition and make this environment favorable for bacterial colonization (Wu et al., 2018). In the case of endophytic bacteria, the root is the main entrance way for microorganisms in plants (White et al., 2014). Thus, it is also expected that the roots present a greater population of microorganisms than other plant segments, such as stems and leaves. Abedinzadeh et al. (2019) found similar data, in which the population size was 3.4 ± 2.12 × 106 and 6.8 ± 1.20 × 103 for rhizospheric and endophytic bacteria in maize plants.
Among the 116 strains isolated, 43 showed positive results for NFB, SF, and IAA. The 43 strains belonged to Proteobacteria, Firmicutes, and Actinobacteria. Genetic diversity studies have reported that these phyla are both endophytic and rhizospheric (Rosenblueth and Martínez-Romero, 2006; Pisa et al., 2011; Prabha et al., 2018) and generally present strains with potential for plant growth promotion.
In this work, through in vitro tests, the main strains that showed the potential to promote plant growth belonged to Bacillus, Enterobacter, Microbacterium, Micrococcus, Pantoea, and Pseudomonas (Amaral et al., 2021). The main PGPB isolated from Paspalum genotypes were Acinetobacter, Bacillus, Cupriavidus, Dyadobacter, Enterobacter, Paraburkholderia, Pseudomonas, and Rhizobium.
In the in vivo test, treatments were separated into two groups. Group 1 received alternative phosphate sources, and Group 2 received a soluble phosphate source. The most significant morphological descriptor for Group 1 was SPAD (SPAD index). The mineral descriptor was potassium (K), and as for the results of the bromatological analysis, the crude protein (CP) content was the most significant. These descriptors showed lower values in treatments that received the soluble phosphate source. For Group 2, the most significant morphological descriptors were aerial fresh weight (AFW) and aerial dry weight (ADW), the mineral was calcium extract (ECa), and the bromatological descriptor was nitrogen extract (EN). Phosphorus was the 11th most significant descriptor in Principal Component 1 and the 12th most significant in Principal Component 2; thus, it had equal importance for both groups.
The in vitro tests showed that the PSI ranged from 2 to 3.61, and the phosphate-solubilizing bacteria belonged to Bacillus, Enterobacter, Microbacterium, Micrococcus, Pantoea, and Pseudomonas. These genera are already described in the literature as phosphate-solubilizing bacteria. De Assumpção et al. (2009) found Pseudomonas sp. with 5.3 and 8.3 of PSI and Pantoea sp. with PSI = 6.0. Suleman et al. (2018), when prospecting bacteria with the potential for phosphate solubilization for wheat plants, found that the best results were with Enterobacter, presenting 2.2–5.8 of PSI. Similarly, the strain with the highest PSI also belonged to the Enterobacter genus in the present study.
Regarding the in vivo test in a greenhouse, the treatments with crystalline phosphate plus Isolates 105 (Pseudomonas sp.) and 458 (Pseudomonas sp.) showed available phosphorus in the initial period of plant development. However, the treatment that received the 110 (Enterobacter sp.) strain showed higher phosphorus content in the second cut, suggesting that the 110 strain needs a more extended period to make the phosphorus available to the plant. In the literature, there is a search for an alternative source of phosphorus for plant nutrition, with Arad rock phosphate being one of those rocks with potential for this purpose. Gatiboni et al. (2003) used natural phosphates from Arad as a source of phosphorus for white clover (Trifolium repens) and ryegrass (Lolium multiflorum) pastures. Gatiboni et al. (2003) observed that the use of natural phosphate from Arad was effective in moderate to high soil and that liming increased the efficiency of superphosphate and decreased the efficiency of rock phosphate as a source of phosphorus. Guedes et al. (2012), evaluating the use of natural Arad phosphate and liming in two tropical grass species in degraded Amazon soil, observed better results in Megathyrsus maximus than in Urochloa brizantha, noting that the success in fertilization was dependent on the cultivated species and soil acidity. These studies sought to explore the gradual capacity of natural rocks to release phosphorus. However, they only used Arad’s natural phosphate in crops without studying the endophytic and rhizospheric microbiota of the host plant. In the present study, the primary method was the optimization of the phosphorus contained in the phosphate rock through selected microorganisms isolated from soil and plant tissue of P. atratum. Another characteristic observed in P. atratum was the acidic soil; both in the rainy and dry seasons, the soil pH of the soil was not higher than 5.6. As mentioned by Guedes et al. (2012), this feature facilitates the solubilization of phosphate from natural rocks.
Principal component analysis showed that the 110FCR (Enterobacter sp.) and 458FCR (Pseudomonas sp.) treatments were very close. The 105FCR (Pseudomonas sp.) treatment stood out, being the treatment with crystalline phosphate that came closest to the treatments that received soluble phosphate. The literature shows that the main phosphate solubilizers are Arthrobacter, Azospirillum, Azotobacter, Bacillus, Beijerinckia, Burkholderia, Enterobacter, Erwinia, Flavobacterium, Mesorhizobium, Microbacterium, Pseudomonas, Rhizobium, Rhodococcus, and Serratia (Bhattacharyya and Jha, 2012; Gouda et al., 2018). There is a search to optimize phosphate rock from Cajati as an alternative source of phosphorus. Lemos et al. (2013) sought to use the rock in the diet of Nellore cattle, as Bernardi and Oliveira (2021) sought to associate the phosphate rock of Cajati with zeolite minerals to nourish the alfalfa crop.
In treatments with sedimentary phosphate, the 109FSE (Bacillus sp.) treatment was the closest to the treatments with soluble phosphate and presented the highest phosphorus content in the first cut. One hypothesis to explain the higher content of phosphorus in the first cut is that there was great solubilization of sedimentary phosphate during the initial periods of the plant, thus depleting almost all available phosphorus sources in the first cut and, consequently, reducing these in dry matter in the second and third cut. This fact shows the importance of topdressing after grazing.
The results observed in treatments with soluble phosphate that included the bacterial isolates also suggest that the source of phosphorus was depleted during the initial stages of plant development. Of the eight treatments that received the strains, seven showed higher phosphorus content than the soluble phosphate control dry matter. The exception was treatment 103FSO (Enterobacter sp.). Therefore, future studies should explore the possibility of using less soluble phosphorus by inoculating phosphate-solubilizing bacteria, seeking to optimize the use of soluble phosphorus in agriculture. When looking for bacteria with the potential for phosphate solubilization in peas, Oteino et al. (2015) conducted an experiment involving Bacillus sp. and Pseudomonas sp. They used soluble phosphate as a control and tricalcium phosphate (Ca3(PO4)2) insoluble in the treatments. As a result, it was observed that the strains increased the phosphorus content in the plant compared to the insoluble control. Nevertheless, no treatment equaled or surpassed the phosphorus content present in the dry matter of the plants treated with soluble phosphate. Similar to the results presented by Oteino et al. (2015), no alternative treatments had equal or higher phosphorus levels as seen in treatments that received the soluble phosphate source. However, a significant difference was that treatments with soluble phosphate also received the bacteria, indicating the potential of these microorganisms to optimize the soluble phosphate in the plant.
During the in vitro tests, using the NFB medium, there was an expectation of finding Azorhizophilus paspali (Azotobacter paspali), a nitrogen-fixing bacteria, found by Döbereiner et al. (1995) when developing the culture medium (Baldani et al., 2014). Batista et al. (2018), when using NFB medium, observed that the main strains with potential for biological nitrogen fixation belonged to Bacillus and Burkholderia. As in these studies, no strains of Azotobacter were obtained in the present study. However, the growth of other microorganisms in the NFB medium was justified because the medium was not highly selective; therefore, strains with the potential to use malic acid as a carbon source and with a pH of 6.8 can grow (Baldani et al., 2014). This method characterized the strains with potential for nitrogen fixation in this work. The cultivable bacterial community with the potential for nitrogen fixation will undoubtedly increase by using another culture medium for isolation or biochemical characterization.
The bromatological descriptors crude protein (CP) and nitrogen (N) content correlated with the morphological descriptor index SPAD in the plant. The CP descriptor was calculated by multiplying the nitrogen content by 6.25 (Druzian et al., 2012; De Medeiros et al., 2015). The SPAD index was generated by the SPAD-502 chlorophyll meter (Soil Plant Analysis Development), which indirectly measures the leaf chlorophyll content without destroying the leaf (D’Oliveira et al., 2020), and the chlorophyll concentration positively correlated with the nitrogen content (Benati et al., 2021).
Among the most significant descriptors to assess the variation between treatments and plant growth promotion, crude protein content was significantly crucial in Group 1 treatments. The 161SF (Pseudomonas sp.), 103FSE (Enterobacter sp.), 25SF (Pseudomonas sp.), CAFCR, and 109SF (Bacillus sp.) were the ones with the highest SPAD index. On the other hand, the 103FCR (Enterobacter sp.), 25FCR (Pseudomonas sp.), 109SF (Bacillus sp.), 161SF (Pseudomonas sp.), and 105 FSE (Pseudomonas sp.) treatments presented the highest levels of CP and N.
Leite et al. (2001) studied the growth and chemical composition of P. atratum cv. Pojuca grass in soil with satisfactory nutrients and nitrogen fertilization during the rainy season. The researchers found CP contents between 6.90 and 12.11%. By sampling the nutritional contents of three cultivars, Porto et al. (2009) found CP contents of 11.1, 11.9, and 9.4% for Tanzania grass (M. maximus), Stargrass (Cynodon), and marandu grass (U. brizantha cv. Marandu), respectively.
Crude protein contents lower than 7% in the dry matter limit animal nutrition (De Abreu et al., 2006). All treatments studied in the present work had a protein content greater than 7%, ranging from 10.12 to 12.46%, values similar to those found by Leite et al. (2001) for the Pojuca cultivar (P. atratum) and by Porto et al. (2009) for the cultivars Capim-tanzânia, Grama-estrela, and Capim-marandu. The leaf protein content in the Pojuca cultivar ranges from 8 to 10% (Karia and de Andrade, 2001). The values found in this work were also superior to the results obtained by Lopes et al. (2010) for U. brizantha, U. decumbens, U. humidicola, and U. Ruziziensis, which ranged from 6.4 to 7.5% CP in dry matter.
In the present study, none of the treatments received nitrogen fertilization. Nevertheless, many of them had similar or superior CP results compared to other studies that evaluated cultivars already on the market. Even the phosphate-free control (CSF), which did not receive any phosphorus source or bacterial inoculum, showed CP results superior to those found by Leite et al. (2001). This fact shows the potential of this genotype as a forage plant.
The main potential NFB were 25 (Pseudomonas sp.), 103 (Enterobacter sp.), 105 (Pseudomonas sp.), 109 (Bacillus sp. and 161 (Pseudomonas sp.). The 103FSO (Enterobacter sp.) treatment stood out due to high levels of CP and N in the dry matter. The Isolate 103 (Enterobacter sp.) also showed good levels of CP and N in treatments with a sedimentary and crystalline phosphate source, making it a good candidate for biological nitrogen fixation investigation. Potassium was another significantly important mineral descriptor observed in Group 1 treatments. In the K+ format, potassium regulates the osmotic potential and activates enzymes involved in respiration and photosynthesis in the plant (Taiz et al., 2016). Therefore, potassium was among the most significant descriptors in the principal component analysis. The treatments that presented the lowest K content received the soluble phosphate source. The treatments that showed the highest K content were 105FSE (Pseudomonas sp.), 103FSE (Enterobacter sp.), 170FSE (Pantoea sp.), 25FCR (Pseudomonas sp.), and 161FCR (Pseudomonas sp.), followed by the control treatments CASF, CAFCR, and CAFSE. The microbiological modification technique enables the direct application of rocks in agriculture. Citric and oxalic acids produced by microorganisms release potassium from biotite, a common mineral in the silicate class (Van Straaten, 2010).
The plants inoculated with the bacterial Isolates 105FCR (Pseudomonas sp.), 109FSE (Bacillus sp.), 110FSE (Enterobacter sp.), 103SF (Enterobacter sp.), 458SF (Pseudomonas sp.), and 458FCR (Pseudomonas sp.) showed growth similar to those treated with a soluble phosphate source. Therefore, they can be selected for future plant growth-promotion experiments.
A total of 116 cultivable endophytic and rhizospheric strains were isolated from rhizospheric soil samples, roots, and leaves of P. atratum.
As for the capacity of the plant growth-promoting bacterial strains, 43 (37.00%) strains showed positive NFB, SF, and IAA results and belonged to Enterobacter (46.50%), Pseudomonas (32.50%), and Pantoea (13.90%), and Bacillus, Microbacterium, and Micrococcus strains represented 6.90%.
The phosphate solubilization index (PSI) ranged from 2 (Pseudomonas spp.) to 3.61 (Enterobacter spp.) and the IAA production ranged from 12.85 (Pseudomonas spp.) to 431.41 (Pantoea spp.) μg ml−1.
In the in vivo test, treatments 105 (Pseudomonas sp.) and 458 (Pseudomonas sp.) were the most significant for the crystalline phosphate source, 109 (Bacillus sp.) for the sedimentary phosphate source and, as for the soluble phosphate source most treatments that received bacterial isolates had higher phosphorus content in the dry matter than the uninoculated soluble phosphate control.
For the morphological, mineral and bromatological variables, the principal component analysis showed that Principal Component 1 had significant associations with the SPAD index (SPAD), potassium (K), zinc (Zn), crude protein (CP), lignin (LIG) and nitrogen (N). While, principal Component 2 showed strong associations with the other 16 descriptors.
These diverse cultivable bacterial genera have the potential to promote plant growth, and the 105FCR (crystalline phosphate + Pseudomonas sp.), 109FSE (sedimentary phosphate + Bacillus sp.), and 110 FSE (sedimentary phosphate + Enterobacter sp.) treatments showed the best results in the plant growth promotion assay.
Other treatments showed isolated characteristics of interest for one or another descriptor analyzed, such as dry weight, potassium, and nitrogen content in the leaves.
The datasets presented in this study can be found in online repositories. The names of the repository/repositories and accession number(s) can be found at: https://www.ncbi.nlm.nih.gov/genbank/, MK521286, MK521276, MK521285, MK521301, MK521314, MK521306, MK521273, MK521304, MK521284, MK521289, MK521303, MK521302, MK521274, MK521300, MK521275, MK521296, MK521290, MK521293, MK521287, MK521271, MK521270, MK521283, MK521282, MK521272, MK521279, MK521308, MK521277, MK521297, MK521278, MK521292, MK521299, MK521291, MK521280, MK521298, MK521281, MK521305, MK521307, MK521261, MK521288, MK521263, MK521262, MK521264, and MK521265.
All authors listed have made a substantial, direct, and intellectual contribution to the work and approved it for publication.
This work was supported by grants from the São Paulo Research Foundation, FAPESP (Proc. No. 2020/11315-6).
ND, WJ, AB, BV, and AF were employed by Embrapa Pecuária Sudeste.
The remaining authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Abedinzadeh, M., Etesami, H., and Alikhani, H. A. (2019). Characterization of rhizosphere and endophytic bacteria from roots of maize (Zea mays L.) plant irrigated with wastewater with biotechnological potential in agriculture. Biotechnol. Rep. 21:e00305. doi: 10.1016/j.btre.2019.e00305
ABIEC (2021). Associação Brasileira das Indústrias Exportadoras de Carnes. Available at: http://abiec.com.br/exportacoes/# (Accessed July, 2022).
Abreu, E. M. A.De, Fernandes, A. R., Martins, A. R. A., and Rodrigues, T. E. (2006). Produção de forragem e valor nutritivo de espécies forrageiras sob condições de pastejo, em solo de várzea baixa do Rio Guamá. Acta Amaz. 36, 11–18. doi: 10.1590/S0044-59672006000100003
Afzal, I., Shinwari, Z. K., Sikandar, S., and Shahzad, S. (2019). Plant beneficial endophytic bacteria: mechanisms, diversity, host range and genetic determinants. Microbiol. Res. 221, 36–49. doi: 10.1016/j.micres.2019.02.001
Ahemad, M., and Kibret, M. (2014). Mechanisms and applications of plant growth promoting rhizobacteria: current perspective. J. King Saud Univ. Sci. 26, 1–20. doi: 10.1016/j.jksus.2013.05.001
Aljanabi, S. M., and Martinez, I. (1997). Universal and rapid salt-extraction of high quality genomic DNA for PCR-based techniques. Nucleic Acids Res. 25, 4692–4693. doi: 10.1093/nar/25.22.4692
Altschul, S. F., Madden, T. L., Schäffer, A. A., Zhang, J., Zhang, Z., Miller, W., et al. (1997). Gapped BLAST and PSI-BLAST: a new generation of protein database search programs. Nucleic Acids Res. 25, 3389–3402. doi: 10.1093/nar/25.17.3389
Alves, P. R., and Hagni, R. D. (2008). The carbonatite-hosted apatite deposit of Jacupiranga, Southeast Brazil - styles of mineralisation, ore characterisation and association with mineral processing. Australas. Inst. Min. Metall. Publ. Ser. 381–389.
Amaral, M. B., Ribeiro, T. G., Alves, G. C., Coelho, M. R. R., Matta, F. D. P., Baldani, J. I., et al. (2021). The occurrence of rhizobacteria from paspalum genotypes and their effects on plant growth. Sci. Agric. 79, 1–10. doi: 10.1590/1678-992x-2020-0240
Araújo, W. L., Quecine, M. C., Lacava, P. T., Aguilar-Vildoso, C. I., Marcon, J., Lima, A. O. S., et al. (2014). Micro-organismos Endofíticos: Aspectos Teóricos e Práticos de Isolamento e Caracterização. Santarém: UFOPA.
Baldani, J. I., Reis, V. M., Videira, S. S., Boddey, L. H., and Baldani, V. L. D. (2014). The art of isolating nitrogen-fixing bacteria from non-leguminous plants using N-free semi-solid media: a practical guide for microbiologists. Plant Soil 384, 413–431. doi: 10.1007/s11104-014-2186-6
Barro, R. S., Varella, A. C., Lemaire, G., De Medeiros, R. B., De Saibro, J. C., Nabinger, C., et al. (2012). Forage yield and nitrogen nutrition dynamics of warm-season native forage genotypes under two shading levels and in full sunlight. Rev. Bras. Zootec. 41, 1589–1597. doi: 10.1590/S1516-35982012000700006
Batista, B. D., Lacava, P. T., Ferrari, A., Teixeira-Silva, N. S., Bonatelli, M. L., Tsui, S., et al. (2018). Screening of tropically derived, multi-trait plant growth-promoting rhizobacteria and evaluation of corn and soybean colonization ability. Microbiol. Res. 206, 33–42. doi: 10.1016/j.micres.2017.09.007
Benati, J. A., Nava, G., and Mayer, N. A. (2021). Spad index for diagnosis of nitrogen status in ‘esmeralda’ peach. Rev. Bras. Frutic. 43, 1–6. doi: 10.1590/0100-29452021093
Bernardi, C. D., and Oliveira, C. D. (2021). Improved alfalfa phosphate utilization using zeolite amendments in low pH soil. J. Soil Sci. Plant Nutr. 21, 1307–1317. doi: 10.1007/s42729-021-00441-z
Bhattacharyya, P. N., and Jha, D. K. (2012). Plant growth-promoting rhizobacteria (PGPR): emergence in agriculture. World J. Microbiol. Biotechnol. 28, 1327–1350. doi: 10.1007/s11274-011-0979-9
Bogas, A. C., Ferreira, A. J., Araújo, W. L., Astolfi-Filho, S., Kitajima, E. W., Lacava, P. T., et al. (2015). Endophytic bacterial diversity in the phyllosphere of Amazon Paullinia cupana associated with asymptomatic and symptomatic anthracnose. Springerplus 4:258. doi: 10.1186/s40064-015-1037-0
Bric, J. M., Bostock, R. M., and Silverstone, S. E. (1991). Rapid in situ assay for indoleacetic acid production by bacteria immobilized on a nitrocellulose membrane. Appl. Environ. Microbiol. 57, 535–538. doi: 10.1128/aem.57.2.535-538.1991
Büchi Labortechnik, A. (2007). NIRCal, Quick Guide number 93119-93123. NIRWare Software Suite Quick Guide. Version B, 1–3.
Coelho, F. E., Lopes, L. C., Cavalcante, R. M., Corrêa, G. C., and Leduc, A. O. (2019). Brazil unwisely gives pesticides a free pass. Science 365, 552–553. doi: 10.1126/science.aay3150
Compant, S., Van Der Heijden, M. G. A., and Sessitsch, A. (2010). Climate change effects on beneficial plant-microorganism interactions. FEMS Microbiol. Ecol. 73, 197–214. doi: 10.1111/j.1574-6941.2010.00900.x
Crous, K. Y., Wujeska-Klause, A., Jiang, M., Medlyn, B. E., and Ellsworth, D. S. (2019). Nitrogen and phosphorus retranslocation of leaves and stemwood in a mature Eucalyptus forest exposed to 5 years of elevated CO2. Front. Plant Sci. 10, 1–13. doi: 10.3389/fpls.2019.00664
D’Oliveira, P. S., Suzuki, L. S., Da Costa Carneiro, J., Machado, J. C., Prates, J. F., Rocha, D. A. S. D. C., et al. (2020). Use of SPAD index in elephant grass pre-breeding. Semin. Agrar. 41, 1047–1052. doi: 10.5433/1679-0359.2020v41n3p1047
De Medeiros, S. R., Gomes, R. C., and Bungenstab, D. J. (2015). Nutrição de bovinos de corte Fundamentos e aplicações. Campo Grande: Embrapa.
De Assumpção, L. C., Lacava, P. T., Dias, A. C. F., De Azevedo, J. L., and Menten, J. O. M. (2009). Diversidade e potencial biotecnológico da comunidade bacteriana endofítica de sementes de soja. Pesqui. Agropecuária Bras. 44, 503–510. doi: 10.1590/S0100-204X2009000500010
Dinato, N. B., Santos, I. R. I., Leonardecz, E., Burson, B. L., Quarín, C. L., de Paula, A. F., et al. (2018). Storage of bahiagrass pollen at different temperatures. Crop Sci. 58, 2391–2398. doi: 10.2135/cropsci2018.03.0164
Döbereiner, J., Baldani, V. L. D., and Baldini, J. I. (1995). Como Isolar e identificar bactérias diazotróficas de plantas não-leguminosas. Brasília: Embrapa-SPI.
Druzian, J. I., Machado, B. A. S., De Souza, C., Fraga, L. M., De Duran, V. A., De Burghgrave, U. S., et al. (2012). Influência da dieta suplementada com óleo de soja na composição centesimal e perfil lipídico de tilápias do Nilo (Oreochromis niloticus). Rev. Inst. Adolfo Lutz 71, 85–92.
Garrity, G. M., Bell, J. A., Lilburn, T. G., and Lansing, E. (2004). Taxonomic Outline of the Prokaryotes. Bergey’s Manual of Systematic Bacteriology. New York, Berlin, Heidelberg: Springer.
Gatiboni, L. C., Kaminski, J., Rheinheimer, D. S., and Brunetto, G. (2003). Superphosphate and rock phosphates as phosphorus sources for grass-clover pasture on a limed acid soil in southern Brazil. Commun. Soil Sci. Plant Anal. 34, 2503–2514. doi: 10.1081/CSS-120024782
Gouda, S., Kerry, R. G., Das, G., Paramithiotis, S., Shin, H.-S., and Patra, J. K. (2018). Revitalization of plant growth promoting rhizobacteria for sustainable development in agriculture. Microbiol. Res. 206, 131–140. doi: 10.1016/j.micres.2017.08.016
Guedes, E. M. S., Fernandes, A. R., Lobato, A. K.Da, Guedes, R. S., de Ávila, W. S., and da Silva, S. P. (2012). Natural phosphate and liming improves phosphorus extraction in two tropical grasses grown in degraded Amazon soil. J. Food Agric. Environ. 10, 1165–1168.
Gusmão, M. R., Valério, J. R., Matta, F. P., Souza, F. H. D., Vigna, B. B. Z., Fávero, A. P., et al. (2016). Plant resistance warm-season (C4) Turfgrass genotypes resistant to spittlebugs (Hemiptera: Cercopidae). J. Econ. Entomol. Adv. 109, 1914–1921. doi: 10.1093/jee/tow135
Hall, T. (1999). BioEdit: a user-friendly biological sequence alignment editor and analysis program for windows 95 / 98 / NT. Nucleic Acids Symp. Ser. 41, 95–98.
Husen, E. (2016). Screening of soil bacteria for plant growth promotion activities in vitro. Indones. J. Agric. Sci. 4, 27–31. doi: 10.21082/ijas.v4n1.2003.27-31
International Fertilizer Industry Association (2021). Available at https://www.ifastat.org/databases/plant-nutrition (Accessed February 23, 2021).
Khan, A. A., Jilani, G., Saleem Akhtar, M., Naqvi, M. S., and Rasheed, M. (2009). A phosphorus solubilizing bacteria: occurrence, mechanisms and their role in crop production. J. Agric. Biol. Sci. 1, 48–58. doi: 10.5923/j.re.20120201.10
Kumar, S., Stecher, G., Li, M., Knyaz, C., and Tamura, K. (2018). MEGA X: molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 35, 1547–1549. doi: 10.1093/molbev/msy096
Leite, G. G., Silveira, L. F.Da, Fernandes, F. D., and Gomes, A. C. (2001). Crescimento e Composição Química do Capim P. atratum cv. Pojuca. Planaltina.
Lemos, G. C., Costa, R. M., Neto, M. G., and Malafaia, P. (2013). Desempenho ponderal de bovinos Nelore suplementados com fontes alternativas de fósforo. Pesqui. Vet. Bras. 33, 188–192. doi: 10.1590/S0100-736X2013000200009
Lopes, F. C. F., Paciullo, D. S. C., Mota, E. F., Pereira, J. C., Azambuja, A. A., Motta, A. C. S., et al. (2010). Composição química e digestibilidade ruminal in situ da forragem de quatro espécies do gênero Brachiaria. Arq. Bras. Med. Veterinária e Zootec. 62, 883–888. doi: 10.1590/s0102-09352010000400018
Maidak, B. L., Cole, J. R., Lilburn, T. G., Parker, C. T., Saxman, P. R., Stredwick, J. M., et al. (2000). The RDP (Ribosomal Database Project) continues. Nucleic Acids Res. 28, 173–174. doi: 10.1093/nar/28.1.173
Marcón, F., Urbani, M. H., Quarin, C. L., and Acuña, C. A. (2018). Agronomic characterization of P. atratum Swallen and P. lenticulare Kunth. Trop. Grasslands-Forrajes Trop. 6, 70. doi: 10.17138/tgft(6)70-81
Mohite, B. (2013). Isolation and characterization of indole acetic acid (IAA) producing bacteria from rhizospheric soil and its effect on plant growth. J. Soil Sci. Plant Nutr. 13, 638–649. doi: 10.4067/s0718-95162013005000051
Nogueira, A. R. A., Machado, P. L. O. A., Carmo, C. A. F. S., and Ferreira, J. R. (1998). Manual de Laboratório: Solo, água, nutrição vegetal, nutrição animal e alimentos. São Carlos: EMBRAPA - CPPSE, 72p.: il. Available at: https://www.infoteca.cnptia.embrapa.br/infoteca/bitstream/doc/44394/1/ManualdeLaboratorios.pdf
Novo, P. E., Francisco, J., Valls, M., Galdeano, F., Honfi, A. I., Espinoza, F., et al. (2016). Interspecific hybrids between Paspalum plicatulum and P. oteroi: a key tool for forage breeding. Sci. Agric. 73, 356–362. doi: 10.1590/0103-9016-2015-0218
Novo, P. E., Galdeano, F., Espinoza, F., and Quarin, C. L. (2019). Cytogenetic relationships, polyploid origin, and taxonomic issues in Paspalum species: inter-and intraspecific hybrids between a sexual synthetic autotetraploid and five wild apomictic tetraploid species. Plant Biol. 21, 267–277. doi: 10.1111/plb.12931
Orbovic, V., and Grosser, J. W. (2006). “Citrus” in Agrobacterium Protocols. ed. K. Wang (Nova Iorque: Humana Press Inc), 177–189.
Oteino, N., Lally, R. D., Kiwanuka, S., Lloyd, A., Ryan, D., Germaine, K. J., et al. (2015). Plant growth promotion induced by phosphate solubilizing endophytic Pseudomonas isolates. Front. Microbiol. 6, 1–9. doi: 10.3389/fmicb.2015.00745
Pande, A., Pandey, P., Mehra, S., Singh, M., and Kaushik, S. (2017). Phenotypic and genotypic characterization of phosphate solubilizing bacteria and their efficiency on the growth of maize. J. Genet. Eng. Biotechnol. 15, 379–391. doi: 10.1016/j.jgeb.2017.06.005
Pérez, A., and Martínez, D. B. Z. (2016). Bacterias endófitas asociadas a los géneros Cyperus y Paspalum en suelos contaminados con mercurio. Rev. U.D.C.A Actual. & Divulg. Científica 19, 67–76.
Pezzopane, C.De, Lima, A. G., Cruz, P. G., Beloni, T., Fávero, A. P., and Santos, P. M. (2017). Evaluation and strategies of tolerance to water stress in Paspalum germplasm. Trop. Grasslands-Forrajes Trop. 5, 153–162. doi: 10.17138/tgft(5)153-162
Pisa, G., Magnani, G. S., Weber, H., Souza, E. M., Faoro, H., Monteiro, R. A., et al. (2011). Diversity of 16S rRNA genes from bacteria of sugarcane rhizosphere soil. Brazilian J. Med. Biol. Res. 44, 1215–1221. doi: 10.1590/S0100-879X2011007500148
Porto, P. P., Deresz, F., Tadeu, G., César, F., Lopes, F., Cecato, U., et al. (2009). Produção e composição química do leite, consumo e digestibilidade de forragens tropicais manejadas em sistema de lotação intermitente. Rev. Bras. Zootec. 38, 1422–1431. doi: 10.1590/S1516-35982009000800005
Prabha, R., Singh, D. P., Verma, M. K., Sahu, P., and Kumar, P. (2018). Bacterial diversity in rhizosphere of Paspalum scrobiculatum L. (kodo millet) is revealed with shotgun metagenome sequencing and data analysis. Data Br. 20, 1653–1657. doi: 10.1016/j.dib.2018.09.006
Quesenberry, K. H., Dampier, J. M., Lee, Y. Y., Smith, R. L., and Acuña, C. A. (2010). Doubling the chromosome number of bahiagrass via tissue culture. Euphytica 175, 43–50. doi: 10.1007/s10681-010-0165-4
Ramos, S. J., Faquin, V., Rodrigues, C. R., Silva, C. A., and Boldrin, P. F. (2009). Biomass production and phosphorus use of forage grasses fertilized with two phosphorus sources. Rev. Bras. Cienc. do Solo 33, 335–343. doi: 10.1590/S0100-06832009000200011
Rodrigues, R. C., Sousa, T. V. R., Melo, M. A. A., Araújo, J. S., Lana, R. P., Costa, C. S., et al. (2014). Agronomic, morphogenic and structural characteristics of tropical forage grasses in Northeast Brazil. Trop. Grasslands-Forrajes Trop. 2, 214–222. doi: 10.17138/TGFT(2)214-222
Rodríguez, H., and Fraga, R. (1999). Phosphate solubilizing bacteria and their role in plant growth promotion. Biotechnol. Adv. 17, 319–339. doi: 10.1016/S0734-9750(99)00014-2
Rosenblueth, M., and Martínez-Romero, E. (2006). Bacterial Endophytes and their interactions with hosts. Mol. Plant-Microbe Interact. 19, 827–837. doi: 10.1094/mpmi-19-0827
Saitou, N., and Nei, M. (1987). The neighbor-joining method: a new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 4, 406–425. doi: 10.1093/oxfordjournals.molbev.a040454
SAS Institute Inc. (2011). Base SAS® 9.3 Procedures Guide: Statistical Procedures. SAS Institute Inc: Cary, NC.
Silva Filho, G. N., and Vidor, C. (2000). Solubilização de fostatos por microrganismos na presença de fontes de carbono. Rev. Bras. Ciência do Solo 24, 311–319. doi: 10.1590/s0100-06832000000200008
Souza, F. H. D., Gusmão, M. R., Cavallari, M. M., and Barioni, W. (2020). Characterization of the potential of native grasses for use as lawns. Ornam. Hortic. 26, 109–120. doi: 10.1590/2447-536X.v26i1.2067
Suleman, M., Yasmin, S., Rasul, M., Yahya, M., Atta, B. M., and Mirza, M. S. (2018). Phosphate solubilizing bacteria with glucose dehydrogenase gene for phosphorus uptake and beneficial effects on wheat. PLoS One 13, e0204408–e0204428. doi: 10.1371/journal.pone.0204408
Taiz, L., Zeiger, E., Moller, I. M., and Murphy, A. (2016). Fisiologia e desenvolvimento vegetal. 6th Edn. ed. P. L. de Oliveira (São Paulo).
Tamura, K., Peterson, D., Peterson, N., Stecher, G., Nei, M., and Kumar, S. (2011). MEGA5: molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol. Biol. Evol. 28, 2731–2739. doi: 10.1093/molbev/msr121
Van Raij, B., Andrade, J. C., Cantarella, H., and Quaggio, J. A. (2001). Análise química para avaliação da fertilidade de solos tropicais. Campinas Inst. Agronômico 775, 26–32. doi: 10.1016/j.mrfmmm.2015.03.010
Van Straaten, P. (2010). “Rochas e minerais como fertilizantes alternativos na agricultura: uma experiência internacional” in Agrominerais para o Brasil (Rio de Janeiro: CETEM/MCT), 235–264.
Verma, S. C., Ladha, J. K., and Tripathi, A. K. (2001). Evaluation of plant growth promoting and colonization ability of endophytic diazotrophs from deep water rice. J. Biotechnol. 91, 127–141. doi: 10.1016/S0168-1656(01)00333-9
White, J. F., Torres, M. S., Somu, M. P., Johnson, H., Irizarry, I., Chen, Q., et al. (2014). Hydrogen peroxide staining to visualize intracellular bacterial infections of seedling root cells. Microsc. Res. Tech. 77, 566–573. doi: 10.1002/jemt.22375
Wu, Z., Liu, Q., Li, Z., Cheng, W., Sun, J., Guo, Z., et al. (2018). Environmental factors shaping the diversity of bacterial communities that promote rice production. BMC Microbiol. 18, 51–11. doi: 10.1186/s12866-018-1174-z
Xiang, T., Malik, T. H., and Nielsen, K. (2020). The impact of population pressure on global fertiliser use intensity, 1970–2011: an analysis of policy-induced mediation. Technol. Forecast. Soc. Change 152, 119895. doi: 10.1016/j.techfore.2019.119895
Yang, B., Wang, Y., and Qian, P. Y. (2016). Sensitivity and correlation of hypervariable regions in 16S rRNA genes in phylogenetic analysis. BMC Bioinform. 17, 135. doi: 10.1186/s12859-016-0992-y
Zhao, X., Chen, X., Li, Z., Zhang, J., and Liu, T. (2021). An evaluation of the effects of the plant endophyte Enterobacter on the salt tolerance of bermudagrass. Acta Prataculturae Sin. 30, 127–136. doi: 10.11686/cyxb2020317
Keywords: bacterial community, genetic resources, biological nitrogen fixation, indoleacetic acid, phosphate solubilization
Citation: de Paula AF, Cruz FPN, Dinato NB, de Andrade PHM, de Moraes ACP, Junior WB, Bernardi ACC, Vigna BBZ, Fávero AP and Lacava PT (2022) Endophytic and rhizospheric bacteria associated with Paspalum atratum and its potential for plant growth promotion with different phosphate sources. Front. Plant Sci. 13:884716. doi: 10.3389/fpls.2022.884716
Edited by:
Helena M. V. M. Soares, University of Porto, PortugalReviewed by:
Roxana Vidican, University of Agricultural Sciences and Veterinary Medicine of Cluj-Napoca, RomaniaCopyright © 2022 de Paula, Cruz, Dinato, de Andrade, de Moraes, Junior, Bernardi, Vigna, Fávero and Lacava. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Paulo Teixeira Lacava, cHRsYWNhdmFAdWZzY2FyLmJy
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.