
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Plant Sci. , 15 June 2022
Sec. Plant Bioinformatics
Volume 13 - 2022 | https://doi.org/10.3389/fpls.2022.870876
This article is part of the Research Topic Digitizing Plant Breeding: Big Data Meets Genetic Mapping View all 6 articles
 Xuli Zhu1,2,3,4†
Xuli Zhu1,2,3,4† Fengshuo Sun1†
Fengshuo Sun1† Mengmeng Sang5
Mengmeng Sang5 Meixia Ye1,2,3,4
Meixia Ye1,2,3,4 Wenhao Bo1,2,3,4
Wenhao Bo1,2,3,4 Ang Dong1
Ang Dong1 Rongling Wu1,2,3,4
Rongling Wu1,2,3,4Heterophylly is an adaptive strategy used by some plants in response to environmental changes. Due to the lack of representative plants with typical heteromorphic leaves, little is known about the genetic architecture of heterophylly in plants and the genes underlying its control. Here, we investigated the genetic characteristics underlying changes in leaf shape based on the model species, Populus euphratica, which exhibits typical heterophylly. A set of 401,571 single-nucleotide polymorphisms (SNPs) derived from whole-genome sequencing of 860 genotypes were associated with nine leaf traits, which were related to descriptive and shape data using single- and multi-leaf genome-wide association studies (GWAS). Multi-leaf GWAS allows for a more comprehensive understanding of the genetic architecture of heterophylly by considering multiple leaves simultaneously. The single-leaf GWAS detected 140 significant SNPs, whereas the multi-leaf GWAS detected 200 SNP-trait associations. Markers were found across 19 chromosomes, and 21 unique genes were implicated in traits and serve as potential targets for selection. Our results provide novel insights into the genomic architecture of heterophylly, and provide candidate genes for breeding or engineering P. euphratica. Our observations also improve understanding of the intrinsic mechanisms of plant growth, evolution, and adaptation in response to climate change.
Heterophylly refers to the changes of leaf shape and size along the axis within a single plant, and is regarded as phenotypic plasticity occurring in response to environmental cues, such as temperature, light intensity, hypoxia, and water availability (Pigliucci and Marlow, 2001). Heterophylly occurs in various plants, including terrestrial and aquatic species (Jaya et al., 2010; Leigh et al., 2011). Morphological characters include leaf shape, leaf size, the relative positions of leaves on the stem, the rate of leaf initiation, etc. these reflect important biological phenomena, including heteroblasty, anisophylly, phase transitions, and heterophylly (Poethig, 1988; Dengler, 1999; Costa et al., 2012). The leaf form of heterophylly shows different morphological features according to genetic variation and environmental factors (Ernande and Dieckmann, 2004). For example, short and long leaves indicate the vital functional significance of heterophylly in Ginkgo biloba (Leigh et al., 2011). In contrast to heteroblasty and anisophylly, heterophylly is represented as the cue-induced switch between two or more leaf morphologies. In some situations, heterophylly is thought to be an adaptive mechanism, and is therefore of ecological and evolutionary significance (Zotz et al., 2011).
Previous studies mainly focused on the morphological, anatomical, and physiological aspects of heterophylly (Kaplan, 1980; Lee and Richards, 1991). For example, physiological indexes, including gas exchange and water use, showed significant differences between different leaf types on a single plant (Kitajima et al., 1997). Many studies have suggested that various hormones are involved in the regulation of heterophylly (Kuwabara et al., 2003; Iida et al., 2009). For example, abscisic acid (ABA) was shown to regulate leaf shape, and ethylene affected the cell elongation and shape of leaves in Ludwigia arcuata (Kuwabara et al., 2003; Sato et al., 2008). Recently, Nakayama et al. (2017) reviewed how phytohormones regulate heterophylly, and attempted to assess the biochemical mechanisms underlying heterophylly. The basic mechanisms of heterophylly at the molecular level have been elucidated in model plants, such as Arabidopsis thaliana (Yang et al., 2013; Rubio-Somoza et al., 2014; Xu et al., 2019). Over the past decade, a key regulatory module consisting of miR156 and its targets, the SQUAMOSA PROMOTER BINDING PROTEIN-LIKE (SPL) genes, was shown to play important roles in diverse aspects of plant development, including phase transition, heterophylly, and heteroblasty (Wu et al., 2009; Poethig, 2013; Manuela and Xu, 2020). Several studies have revealed the molecular mechanisms underlying heterophylly in non-model plants (Nakayama et al., 2014; Sicard et al., 2014; Kim et al., 2018). Studies in Rorippa aquatica suggested that the KNOTTED1-LIKE HOMEOBOX Class I (KNOX1) genes changed leaf primordia through the accumulation of gibberellin (GA) in response to environmental stimuli (Nakayama et al., 2014). Fu et al. (2012) reported that the miR156-SPL module is associated with the development of heterophylly in woody plants. However, few studies have investigated the genetic architecture of heterophyllous variation at the level of the whole genome.
The genome-wide association study (GWAS) has evolved as a powerful tool to understand the genetic control of complex traits, and has been successfully used to detect a substantial number of candidate genes for leaf traits in plants (Tian et al., 2011; Zhao et al., 2018; Mähler et al., 2020; Ye et al., 2020; Ahmed et al., 2021; Fu et al., 2021). However, these GWAS studies used only descriptive traits of leaf shape, such as width and length, and neglected information about the spatial distribution of shape changes. Therefore, the geometric morphometrics (GM) model has been used to detect leaf shape changes, and precisely describes leaf variation (Adams et al., 2013). To better understand the genetic mechanisms underlying leaf architecture, two GM approaches, namely landmark-based and outline analyses, have been used for genetic mapping. This new statistical framework, called shape mapping, can map specific shape quantitative trait loci (QTLs) that affect leaf shape variation (Langlade et al., 2005; Fu et al., 2010; Bo et al., 2014). However, the above models for shape mapping cannot be used to characterize heterophylly variation, and these methods neglect the differentiation of leaf shape at multiple positions within the same plant. To resolve this problem, the principle of GM-based shape mapping was expanded to map QTLs for the shapes of leaves in various positions, and this method can test specific QTLs potentially mediating heterophyllous variation throughout the genome (Sun et al., 2018). These shape mapping tools are valuable for exploring the genetic architecture of heterophylly.
Populus euphratica Oliv. is a vital woody plant of riparian ecosystems in arid and semi-arid regions, and exhibits extremely high tolerance to abiotic stresses, especially salt stress (Chen et al., 2004; Ling et al., 2015). Leaf morphology is linear, lanceolate, ovate, and broad–ovate from the bottom to the top of the canopy in adult P. euphratica (Figure 1). This transition of leaf shape shows that it has typical heterophylly (Zhao and Qin, 2017; Zhai et al., 2020). To date, most research regarding heterophylly of P. euphratica has focused mainly on leaf morphology, anatomical structure, leaf development, and physiological level (Zhai et al., 2020; Song et al., 2021). Some recent studies explored the molecular mechanisms underlying the genesis of P. euphratica heteromorphic leaves through high-throughput sequencing analyses (Iqbal et al., 2017; Qin et al., 2018; Zeng et al., 2019; Guo et al., 2020; Li et al., 2020; Fu et al., 2021). These studies suggested that specific genes, miRNAs, lncRNAs, and circRNAs may interact with each other in the regulation of leaf morphogenesis in P. euphratica (Song et al., 2021). To date, however, the genetic mechanisms underlying heterophylly of P. euphratica have rarely been studied.

Figure 1. Types and spatial distribution pattern of heterophylly in Populus euphratica. (A) An example of the four types of heterophylly in P. euphratica. (B) The spatial distribution of four types of heterophylly in the canopy of mature in P. euphratica.
In this study, heteromorphic leaves were collected from 860 diverse P. euphratica genotypes, photographed, and phenotyped through image analysis. Whole-genome sequencing produced a dense marker set of more than 400,000 single-nucleotide polymorphisms (SNPs). We further evaluated population stratification based on SNPs covering the P. euphratica genome. We performed single-leaf association tests between descriptive measures and leaf outline heterophylly using a mixed linear model of correlated traits. We also performed a multi-leaf GWAS, to increase the power and identify SNPs and candidate genes involved in controlling the formation of heterophylly in P. euphratica. This comprehensive GWAS provided a better understanding of the genetic architecture of heterophylly, and will facilitate molecular marker-assisted breeding in P. euphratica. It also provided new insights into how heterophylly has responded to the harsh environment during the evolution of P. euphratica.
The association natural population was composed of 860 P. euphratica genotypes, which were originally distributed in the Tarim River Basin of Yuli County, Xinjiang Uygur Autonomous Region, China (41°0′–41°2′N, 86°0′–86°2′E) (Supplementary Figure 1). The approximate age of the sampled trees varied from 50 to 100 years. Trees showing good growth and no disease or insect pests were randomly selected from within an area of 400 km2, and the distance between the sampled trees was at least 50 m to decrease kinship.
Four types of heterophylly were classified on the basis of the leaf index (LI, defined as leaf length [LL]/leaf width [LW]): linear (LI ≥ 4), lanceolate (4 > LI ≥ 2), ovoid (2 > LI ≥ 1), and broad–ovate (LI < 1) (Figure 1A). Heteromorphic leaves were picked from the bottom to the top position of the entire canopy for each mature P. euphratica in July 2017 (Figure 1B). A total of 3,644 leaves were collected for subsequent photographs.
Leaves were photographed against a red background with the same focal distance by a digital camera (DXM 5600; Nikon). Digital image analysis was performed as follows:
(1) Each leaf picture was read into R software (ver. 4.0; R Core Team, 2021) by package imager (Simon, 2021) and the RGB matrix was extracted to distinguish the leaf from the background.
(2) The RGB matrix of the leaf was converted into a binary matrix to capture the leaf outline.
(3) The effects of the scale, position, and orientation of each type of heteromorphic leaf were adjusted for according to the method described by Fu et al. (2013), and the effect of leaf size was adjusted for by an orthogonal Procrustes process (Viscosi and Cardini, 2011).
(4) A set of semi-landmarks describing the leaf boundary were generated at equal radial angles, and the Cartesian coordinates of each point on the leaf boundary were calculated.
The image analysis process was performed in R, Supplementary Figure 2 shows the pipeline used for obtaining the leaf boundary from a leaf image. Finally, the descriptive data, including LL, LW, LI, and leaf area (LA), and the coordinate matrix delineating the leaf outline as phenotypic data, were used in subsequent analyses.
Descriptive statistics, normality testing, coefficient of variation (CV), and Spearman’s rank correlation analysis of the descriptive data of leaf shape were performed. The statistical significance of differences among the descriptive values of four types of leaf was evaluated by one-way analysis of variance (ANOVA). Elliptic Fourier (EF) analysis was used to model the closed outline with two-dimensional (x, y) coordinate data for each leaf (Sun et al., 2018). Multivariate analysis of variance (MANOVA) was used to examine the significance of differences among the outlines of four types of leaf with the estimated Fourier coefficients. Multiple comparisons were performed using the Bonferroni method. To improve the performance and computational efficiency of GM-based GWAS, principal component analysis (PCA) was used to reduce the dimensions. Each principal component (PC) can reflect the specific morphology of heterophylly. In this study, PCs explaining >96% of the cumulative variance were chosen. To evaluate the variance and residual explained by each PC based on the CV, the maximum absolute value was calculated for each PC, and a new PC was generated to ensure that all genotypes were represented in each PC. Box–Cox transformation to a normal distribution was performed for PCs that were not normally distributed. PCA was calculated using the prcomp function in R. Additional details about the PCA of leaf shape were described previously by Fu et al. (2013). All statistical analyses were performed using R 4.0.
All P. euphratica genotypes were sequenced using the Illumina HiSeq 2000 platform (Illumina, San Diego, CA, United States). Clean data were obtained after removing low-quality bases and adapters from the raw data. Clean reads of the 860 genotypes were mapped to the P. euphratica reference genome (Ma et al., 2013) with BWA software (Li and Durbin, 2009) using the default parameters. The mapping results were generated in BAM format, and non-unique and unmapped reads were removed with SAMtools (Li et al., 2009). SNPs were called using GATK (McKenna et al., 2010) and were filtered by applying the following criteria in VCFtools (Danecek et al., 2011): mapping quality > 20; missing rate < 50%; minor allele frequency (MAF) < 5%; mean depth of coverage > 40; and p-value of Hardy–Weinberg equilibrium test > 0.05. Finally, 401,571 SNPs were screened for subsequent analysis. The genotype data has been deposited in the Genome Variation Map in National Genomics Data Center, under accession number GVM000321 that can be publicly accessible at https://bigd.big.ac.cn/gvm/getProjectFile?t=b9430bac.
A Bayesian clustering approach implemented in the software fastSTRUCTURE (Raj et al., 2014) was used to investigate the population structure of P. euphratica. K-values (i.e., number of individual ancestry proportions) ranging from 2 to 15 were tested by 10 replicates per K, using the default convergence criterion and priors. A reasonable range of K values was determined by maximizing marginal likelihood and model components, and using the chooseK.py function in fastSTRUCTURE. We also performed PCA to further investigate the structural components of the P. euphratica population, and the relationships among them, using the R package SNPRelate (Zheng et al., 2012). The King program was used to calculate the kinship matrix (Manichaikul et al., 2010) among the 860 genotypes used in subsequent analysis. Linkage disequilibrium (LD) was evaluated using PLINK (Purcell et al., 2007) with the following parameters: –ld-window-r2 0 –ld-window 99999 –ld-window-kb 1000.
Decay curves were fitted using the non-linear least-squares method based on an exponential equation. LD decay plots were drawn using R 4.0.
Let Zi represent the descriptive phenotypic or PC value for a particular type of leaf from tree i. We used a linear mixed effect model to detect genomic regions controlling the shape of the leaf; this model is expressed as:
where μ represents the intercept coefficient; X is a vector of SNP genotypes; α is a vector of the fixed effect of the SNP; P represents the population group; β is a vector of the fixed effect of the population structure; η is a random variable capturing the random polygenic effect, distributed as , where K is the realized kinship matrix among different genotypes and is the additive genetic variance; and ε is a vector of error, distributed as .
In this study, different methods were used to test the associations between genotypes and leaf phenotypes. Equation (1) is called QK model. If the fixed effect of population structure and the random effect of kinship are neglected in equation (1), which is simplified as linear regression (LM) model. If only the random polygenic effect η is moved in equation (1), which is used as a Q model to analyze continuous quantitative traits. The Q–Q plot and genomic inflation factors were used to evaluate the optimal GWAS model. Permutation tests were used to determine the critical thresholds by reshuffling real phenotypes 1,000 times to break the connections between these phenotypes and their corresponding genotypes (Churchill and Doerge, 1994), and the 5% of 1,000 LR values is used as a critical threshold. The QK model was implemented in the R package GMMAT (Chen et al., 2020).
In contrast to traditional leaf GWAS models, the method used in this study involved mapping of high-dimensional phenotypes for heterophylly. The outline of each leaf was described by selecting a set of L semi-landmarks in the clockwise direction, at equally spaced radial angles on the leaf boundary. Let Zi = (zi1, zi2, zi3, zi4) = ((xi1, yi1), (xi2, yi2), (xi3, yi3), (xi4, yi4)) = (((xi1(1), yi1(1)), … (xi1(L), yi1(L))), …, (((xi4(1), yi4(1)), … (xi4(L), yi4(L))))) represent the x- and y- coordinates of vectors of four types of heterophylly for an individual i. Let Ci = (ci1, ci2, ci3, ci4) denote the vectors of the phenotype and target PC of four heterophyllous leaves for an individual i. PCs are obtained by dimension reduction of the shape space coordinate Z with PCA. Assuming that there is a SNP that controls the formation of heterophylly, the likelihood of the phenotypes or PCs for a given SNP can be expressed as:
where J is the number of SNP genotypes observed for the marker; nj is the number of individuals of SNP genotype j; fj((ci1,ci2,ci3,ci4)|Θj,η) is a multivariate normal density distribution of phenotypes for individual i at SNP genotype j, where Θj is a set of expectation parameters that model the mean vectors for SNP genotype j, and η is the variance-covariance parameter common to all genotype groups that generated the covariance matrix via the SAD (1) model (Zhao et al., 2005).
Through likelihood (2), the existence of heterophylly QTLs can be tested using the following hypotheses:
H1: At least one of the equalities aforementioned does not hold,
where H0 corresponds to the reduced model and H1 corresponds to the full model, in which there exists a heterophylly QTL. The above hypotheses were tested by comparing the log-likelihood ratios of the reduced and full models. An empirical approach based on permutation tests can be used to determine the thresholds for the hypotheses (Churchill and Doerge, 1994). The above model can also be extended by fully considering the influence of population structure.
The SNP annotation and effect predictions were done using SnpEff software (Cingolani et al., 2012), based on the PopEup_1.0 genome sequences from the NCBI database (Ma et al., 2013). Genomic regions suitable for screening candidate genes were identified based on the gap between the left- and right-most significant SNPs in a Manhattan plot, and based on SNPs within ±100 kb of significant SNPs not inside the tower-like structure of the Manhattan plot. Candidate genes were annotated with the BLAST2GO tool (Conesa and Götz, 2008) using the non-redundant protein (Nr) database.
ANOVA indicated that the means of each descriptive measure (LL, LW, LI, and LA) were statistically significantly different among the four types of heteromorphic leaf classes (Figure 2A). The leaf outlines of a population of 860 P. euphratica accessions were reconstructed based on different types of leaves through EF analysis (Figure 2B). MANOVA demonstrated significant differences in outlines among leaves (Figure 2B).
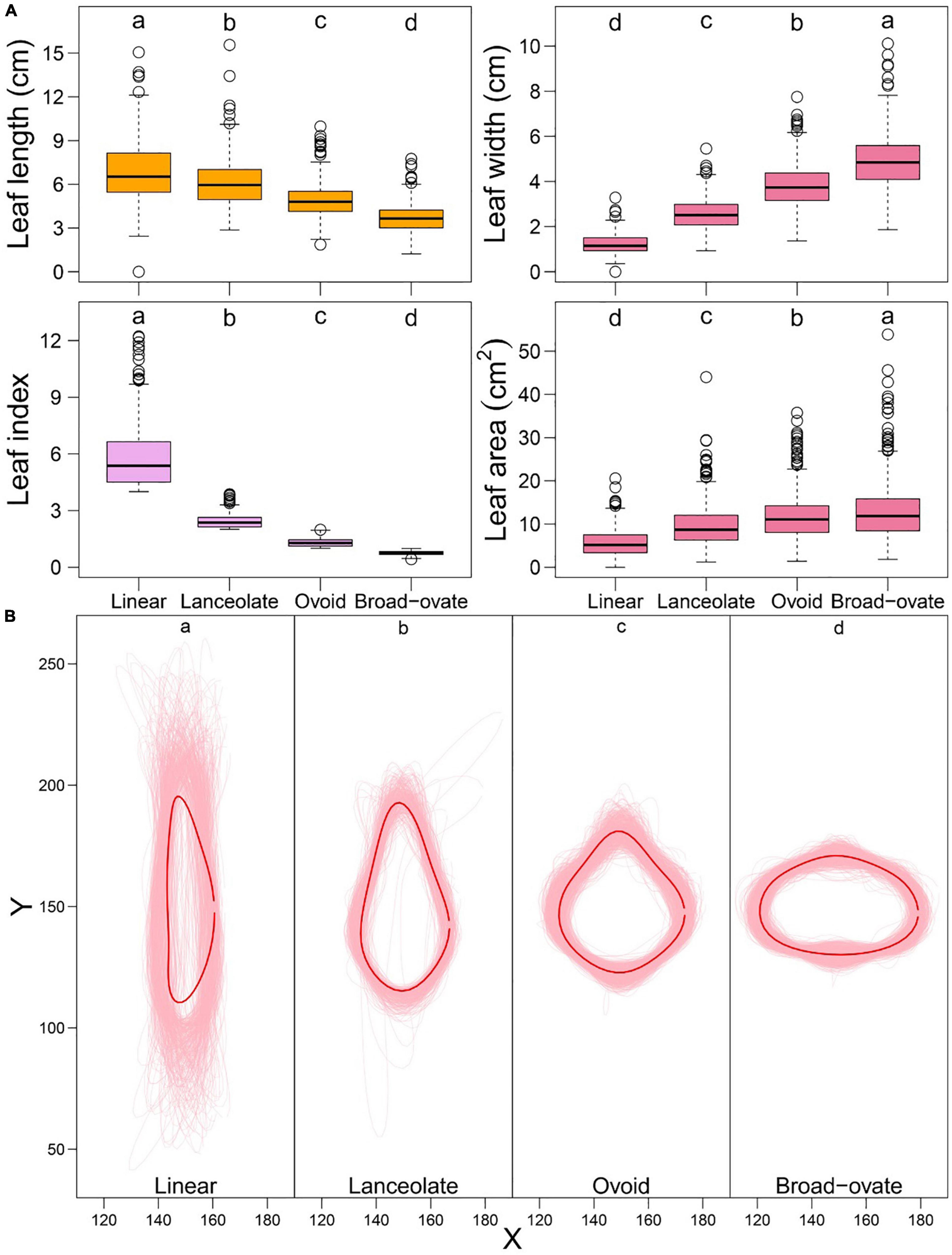
Figure 2. Descriptive data and shape distribution and MANOVA test for heterophylly, the letters (a, b, c, and d) indicate multiple comparison results at the significant level 0.05. (A) Descriptive data distribution and MANOVA for the descriptive data of heterophylly. (B) Shape distribution and MANOVA for the shape of heterophylly. The light pink thin line denotes the shape of heterophylly for each genotype, red thick line denotes the mean shape of all genotypes for four types of heterophylly.
To examine the variation of heterophylly in the whole sample, PCA was applied to the superimposed coordinates of the leaf boundaries. PCA showed that the first five orthogonal axes together explained 96.1% of the variance in accessions (57.7, 27.5, 6.7, 2.9, and 1.3%, respectively) (Figure 3A). Clear overlap was seen between the linear and lanceolate leaves, and the ovoid and broad–ovate leaves, but the linear, lanceolate and ovoid, broad–ovate leaves were separated along the PC2 axis (Figure 3B). As shown in Figure 3C, the outlines of linear, lanceolate, and broad–ovate leaves exhibited significant variation among the five PCs. Although the morphologies of ovoid leaves were similar among the five PCs, there were significant differences in leaf tip and leaf base (Figure 3C).
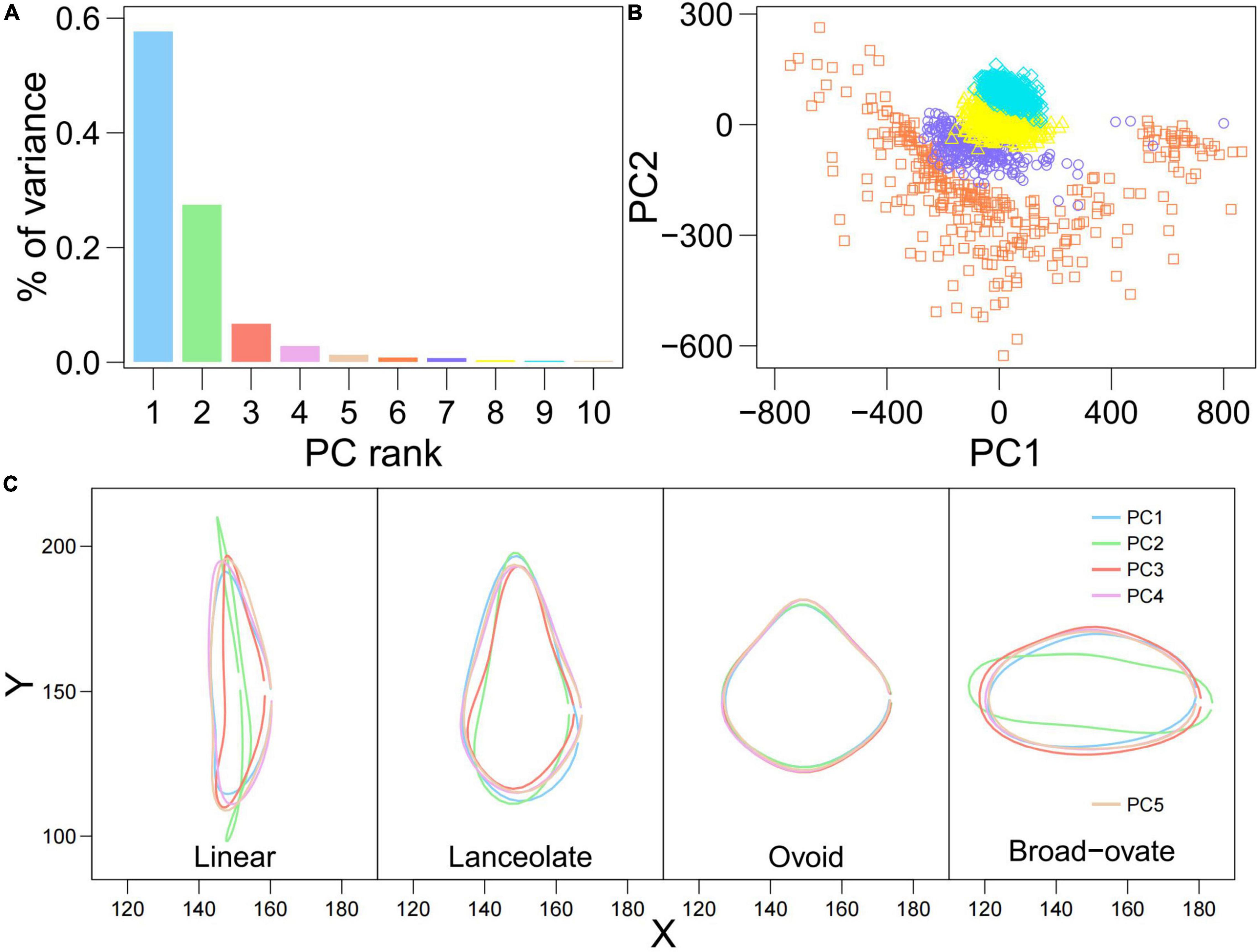
Figure 3. Principal component analysis (PCA) analysis calculated from the landmark data of 860 genotypes for heterophylly. (A) The barplot of PC contributions. (B) Biplot of the first and second principal components. The square, circle, triangle, and diamond symbols represent the linear, lanceolate, ovoid, and broad-ovate leaves, respectively. (C) The mean shape of four heterophyllous leaves on the first five PCs.
To further evaluate the variation and distribution of phenotypes among individuals, descriptive statistical analysis was performed using R. The CVs of all traits showed an extensive distribution range in the population, but the CVs of all traits showed no specific patterns within a given leaf type (Supplementary Table 1). Interestingly, the largest CVs for LA were seen in heteromorphic leaves, ranging from 56 to 58%. In general, the CVs of phenotypes LL, LW, LI, and LA were significantly greater than those of the PCs (Supplementary Table 1). Skewness and kurtosis coefficients were obtained to assess the data distribution using the Shapiro–Wilk test. Kurtosis of the traits varied from 2.5 to 13.67, and skewness from −0.08 to 2.47. These two parameters deviated markedly from normality for most traits, as verified by the Shapiro–Wilk test (P < 0.01). The results of descriptive statistical analysis suggested that the extensive variation in leaf outline was due not only to the use of traditional leaf measurement methods, but also to the PCs.
We assessed the relationships among all traits (LL, LW, LI, and LA), and the PCs, by correlation analysis (Supplementary Figure 3). Generally, the four traits showed strong correlations with each other within the same leaf type (Supplementary Figure 3). However, strong negative associations were found in the five PCs for leaf outline. The phenotypes exhibited moderate relationships for heteromorphic leaves. Interestingly, the five PCs showed weak or no correlations across different types of leaves, suggesting that each PC may reflect a unique morphological trait of heteromorphic leaves. We also found a high degree of phenotypic similarity among some descriptive traits, and some PCs, within the same type of leaf. For example, the positive correlations of PC5 with LW, LI, and LA for linear shape, and the negative correlation of PC2 and LI (Supplementary Figure 3) for ovoid shape, suggest that some PCs may contain information about LL, LW, LI, and LA.
Population structure was inferred by fastSTRUCTURE using all markers across the entire genome of 860 genotypes. The final number of subpopulations was two (K = 2), based on the marginal likelihood. There was a small degree of ancestral population stratification, although the majority of genotypes were classified into one “general” population (Supplementary Figure 4A). We performed a further PCA to investigate global genetic variation in the P. euphratica panel. The first two PCs accounted for 1.25 and 0.78% of the variation in the genotypic data, respectively (Supplementary Figure 4B). In particular, PC1 clearly distinguished the two subpopulations (Pop1 and Pop2). The fastSTRUCTURE and PCA results suggested that there was significant population structure difference among individuals of this natural population of P. euphratica. The LD decay with distance across the entire P. euphratica genome for all individuals was ∼61 Kb when the LD baseline was set to r = 0.1 (Supplementary Figure 4C). The LD decay observed in Pop1 was consistent with that of the whole population, but Pop2 showed a much slower rate of LD decay compared with both Pop1 and the entire population (Supplementary Figure 4C).
As the data of most traits showed a skewed distribution (Supplementary Table 1), Box–Cox transformations were required to normalize the data and reduce false-positive signals in the GWAS. The Shapiro–Wilk test showed that all phenotypes were approximately normally distributed at p < 0.05 (Supplementary Figure 5). Three different models (LM, Q, and Q + K) were used to test the associations between single-leaf traits and the 401,571 SNP genotypes evaluated in our 860 P. euphratica accessions. The Q–Q plot from the GWAS indicated that the three methods showed similar performance for all traits (Supplementary Figure 6), suggesting that population structure and cryptic relatedness had little influence on the precision of the GWAS analysis of this natural population. Therefore, the LM model (excluding both the population structure and kinship matrix) was used to detect significant associations between leaf traits and SNPs.
We examined a total of 140 significant SNP-trait associations using stringent significance thresholds based on permutation (Supplementary Table 2). However, no significant associations were found for LI or LA in lanceolate leaves, or for PC5 in linear leaves, or PC1 or PC4 in ovoid leaves (Supplementary Table 2 and Figure 4A). The number of markers associated with each trait/PC ranged from 1 (LA and PC3 of linear leaves, LW and PC1 of lanceolate leaves, PC3 of ovoid leaves, and PC5 of broad–ovate leaves) to 37 (LW of ovoid leaves). These markers were distributed on 19 chromosomes and several unanchored scaffolds (Figure 4A). Furthermore, no significantly overlapping SNPs were observed between the different leaf types (Supplementary Figure 7A), whereas two significantly overlapping SNPs were detected between the descriptive traits (Supplementary Figure 7C). One SNP was associated with both the descriptive traits and PCs (Supplementary Figure 7B). We also examined significantly overlapping SNPs for the different PCs (Supplementary Figure 7D). Three SNPs that affected both PC1 and PC4 were tested. Of the significant SNPs, 64 (45.7%) were located in intergenic regions, while 46 (32.9%) were in upstream, downstream, or intronic regions; 14 (10%) of the significant SNPs were in non-coding areas of genes (Supplementary Table 2). In addition, six missense and seven synonymous point mutations were detected (Supplementary Table 2), which most likely affected protein function and contributed to leaf variation. We also examined how much of the phenotypic variation was explained by individual SNPs. Notably, 12 (8.6%) significant individual SNPs explained >8% of the observed phenotypic variation.
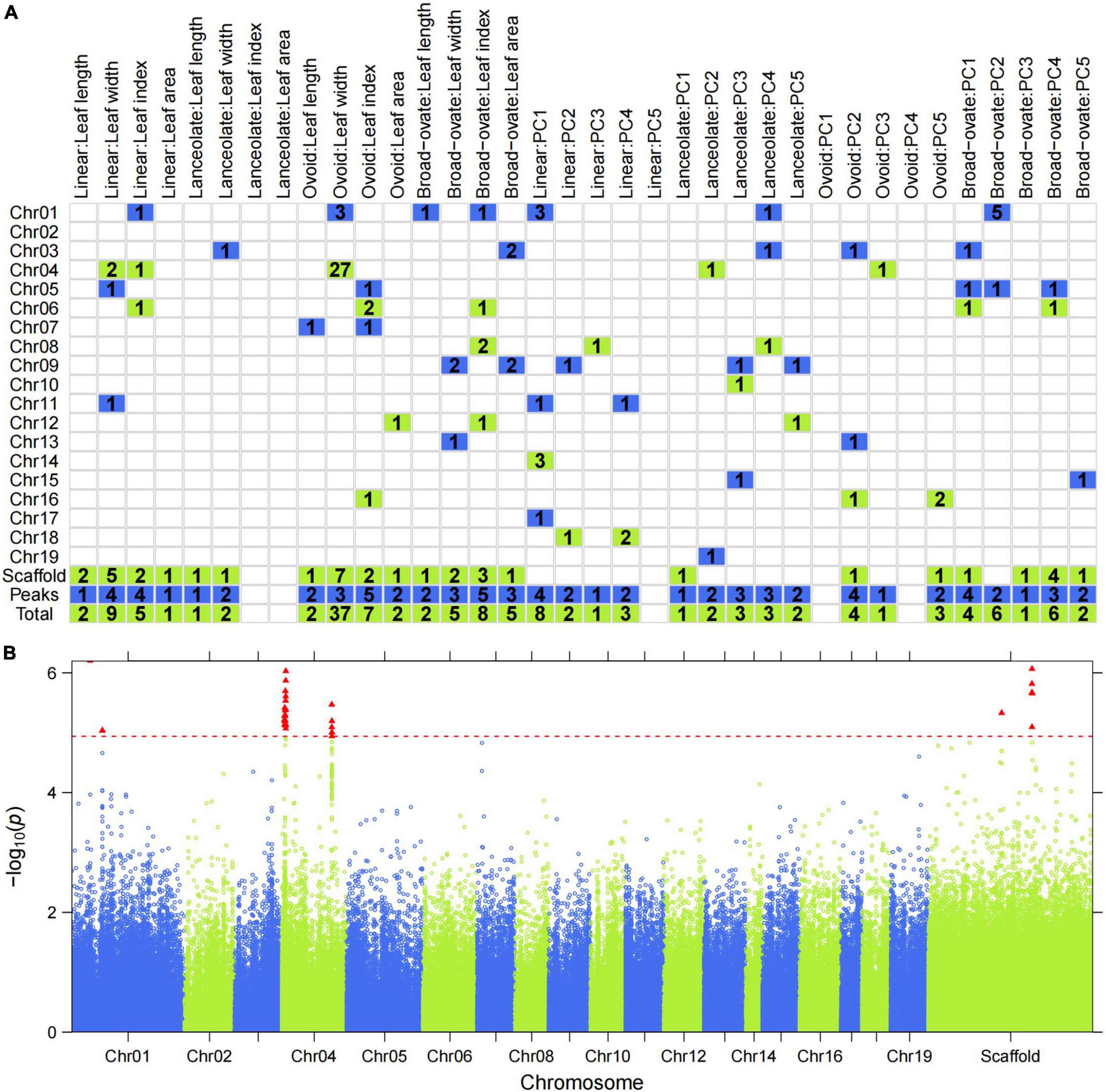
Figure 4. Genome-wide distribution of P. euphratica SNP loci detected based on single-leaf GWAS. (A) Distribution of the number of significant SNPs across each chromosome for all traits. (B) Manhattan plot displaying the GWAS result of the trait ‘Ovoid: Length width’ based on the LM model. The red triangle indicates the significantly associated SNPs. Redline indicates the threshold with permutation tests at 0.05.
As an example, Figure 4B shows the association signals of all SNPs along the chromosomes for LW in ovoid leaves. Most significant SNPs converged to form a tower-like structure in the Manhattan plot (Figure 4B), indicating the presence of only a few genomic regions controlling the LW of ovoid leaves. Two powerful association signals were detected for this phenotype, forming two QTL clusters on chromosome 4. The first cluster (first tower-like region include 21 SNPs) was located in four scaffolds in NW_011501471.1, NW_011500351.1, NW_011500738.1, and NW_011500414.1, with a peak SNP (missense mutation) at 0.104 Mbp in NW_011500414.1 (P = 5.67e-07, Supplementary Table 2). A total of six significant SNPs in the second cluster were mapped to NW_011500574.1 and NW_011500101.1, and two synonymous SNPs (SNP296047 and SNP296048) were in the same genomic region, which may play an important role in regulating LW in ovoid leaves.
Single-leaf GWAS were performed to characterize phenotypic variation in individual leaves, but failed to capture the differentiation of leaf shape among multiple classes, limiting their application for determination of the more complex genetic mechanisms of heterophylly. Here, multi-leaf GWAS were used to identify the genomic loci potentially controlling heterophylly in P. euphratica. The results of single-leaf GWAS showed that the inclusion of population stratification and the kinship matrix did not substantially increase confounding for the descriptive traits and PCs, so their effects were ignored in multi-leaf GWAS.
Multi-leaf GWAS for heterophylly detected 200 SNPs that passed permutation tests at the 5% level, among which 151 were associated with PCs (Supplementary Table 3, Figures 5, 6A, and Supplementary Figures 8–15A). Multi-leaf GWAS showed increased power but identified different genomic positions than single-leaf GWAS. Surprisingly, there was only one significantly overlapping SNP (SNP317395) between the single- and multi-leaf GWAS (Supplementary Tables 2, 3). We also found that only one missense SNP (SNP347849) was associated with both the descriptive traits and PCs describing leaf shape. A total of 101 significant SNPs were located in intergenic regions; 5 were missense mutations and 10 were synonymous changes (Supplementary Table 4). The phenotypic variance explained (PVE) by these SNPs ranged from 0.0008 to 9.21% for all traits.
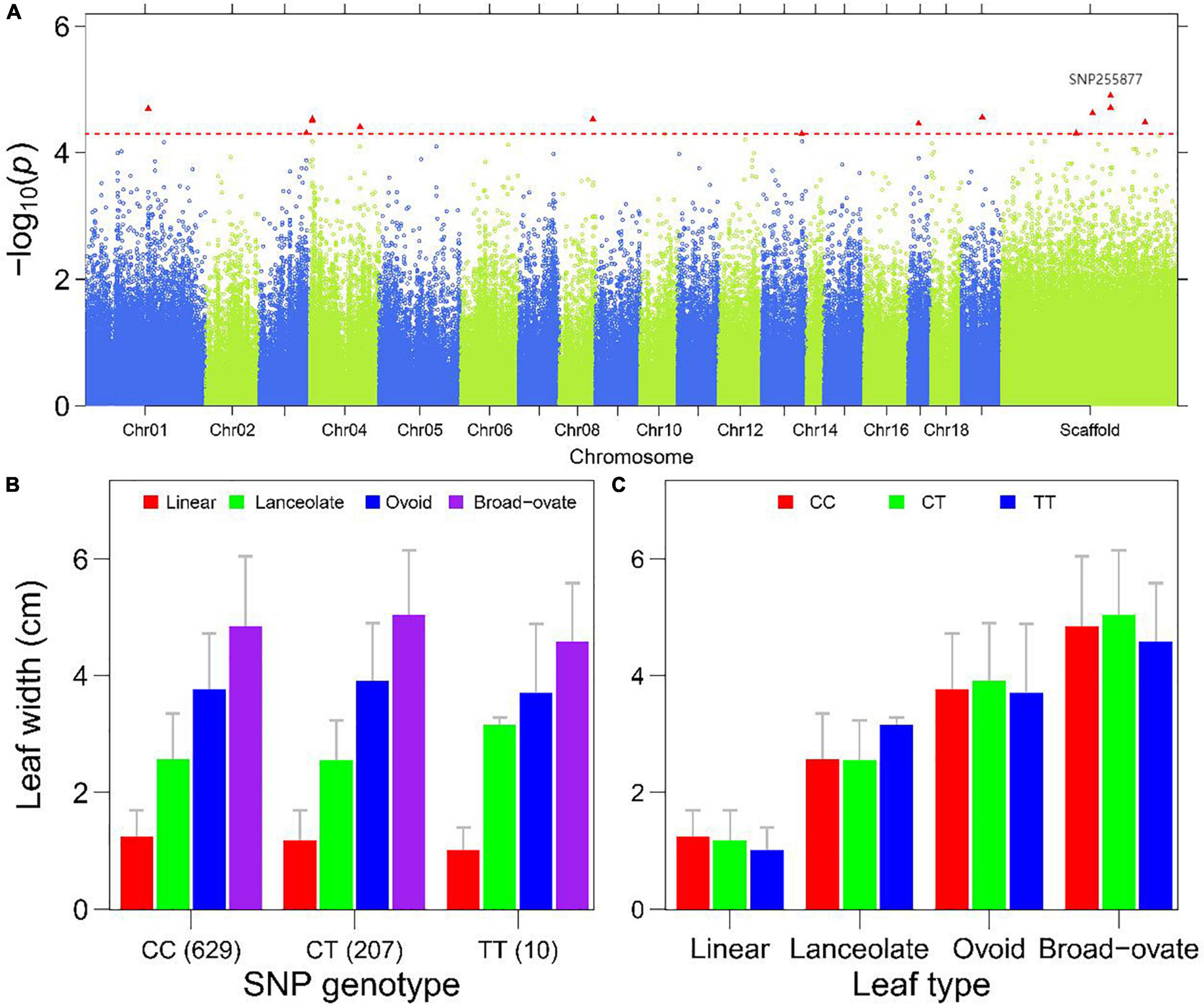
Figure 5. Genome-wide association mapping of the leaf width in P. euphratica. (A) Manhattan plot displaying the GWAS result of the trait ‘leaf width’ based on the single-leaf GWAS. The significantly associated SNP markers are labeled. (B) Histograms of leaf width of different leaf types plotted as a function of genotypes at SNP255877. (C) Histograms of allelic effects of SNP255877 for the heterophylly.
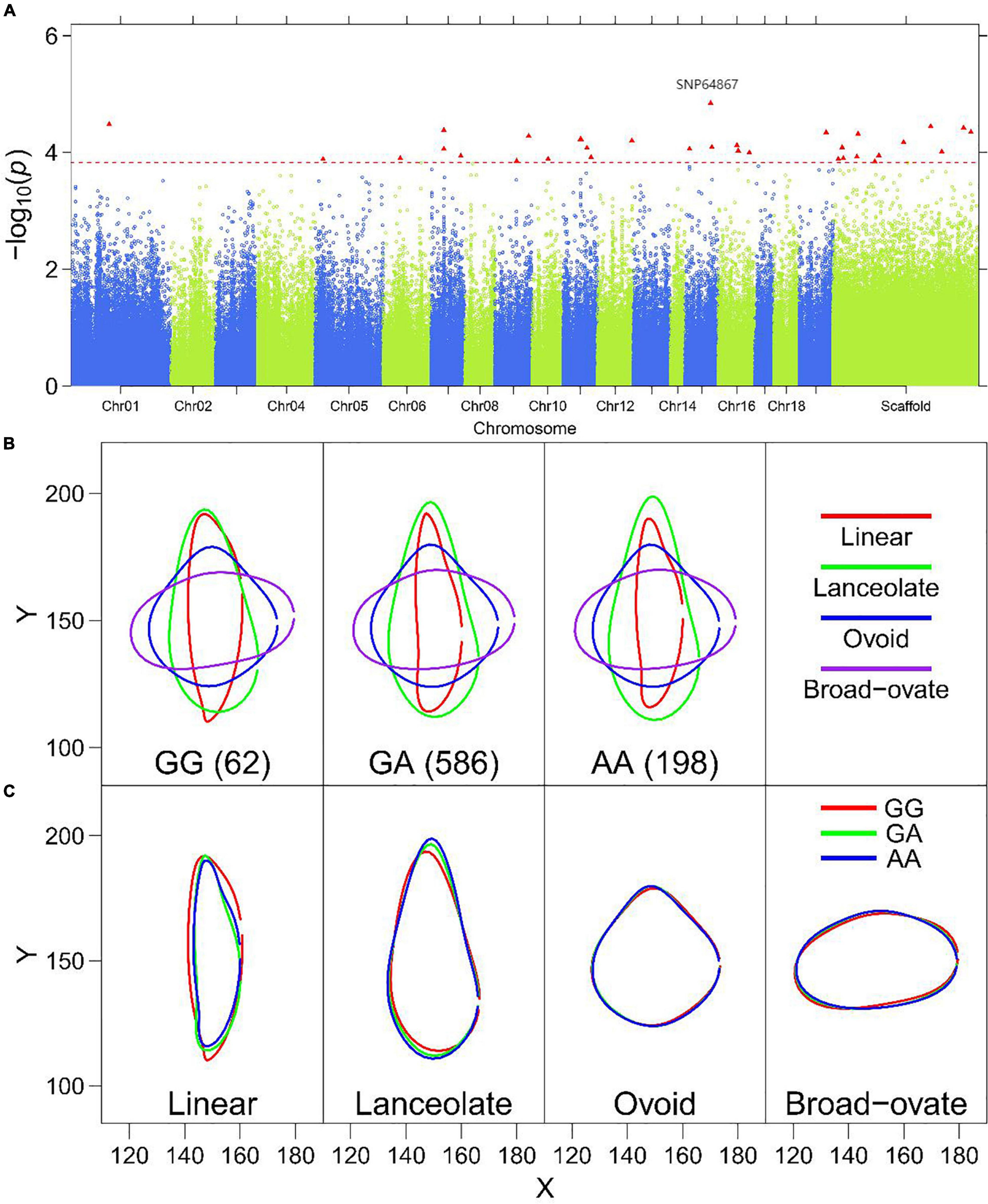
Figure 6. Multi-leaf GWAS for the PC1 of leaf structure in P. euphratica. (A) Manhattan plot displaying the GWAS result of the PC1 based on the multi-leaf GWAS. The significantly associated SNP markers are labeled. (B) Leaf outline of PC1 of heterophylly plotted as a function of genotypes at SNP64867. (C) Visualized variation in the shape of four heterophyllous leaves in P. euphratica at a significant SNP (SNP64867 on scaffold NW_011500277.1).
To show how a QTL controls heterophylly, we first compared the descriptive traits of heteromorphic leaves of P. euphratica among different genotypes for the significant SNPs, and then evaluated the variation of traits for the three genotypes within the same leaf type. The results presented in Figure 5B and Supplementary Figures 8–10B showed that there were obvious differences between different heteromorphic leaves of the same genotype, with significant QTLs for LL, LW, LI, and LA. SNP25589 was significantly associated with LL in the scaffold NW_011500852.1, and showed differences among the four leaf types (Figure 5C). Comparison among the genotypes of SNP62886 showed differences in LW between linear and lanceolate leaves (Supplementary Figure 8C). A similar result was found for the marker of LA, SNP57868 (Supplementary Figure 10C). For LI, we found that differences among the three genotypes occurred mainly in lanceolate, ovoid, and broad–ovate leaves (Supplementary Figure 9C).
To further show how a QTL controls leaf shape, we examined significant SNPs for each PC. In general, the significant SNPs for PC1 most affected leaf shape variation, as PC1 always explains the most variance. These SNPs were responsible for most of the variance in leaf shape in the ovoid and broad–ovate leaves, in particular. Figure 6A displaying the GWAS result of the PC1 based on the multi-leaf GWAS. Figure 6B shows PC1 for leaf shape for three genotypes, GG, GA, and AA; the QTL was detected by translating EF curves according to leaf shape. Three genotypes of SNP64867 explained the variation of linear and lanceolate leaves according to x- and y-coordinate values (Figure 6C). Regarding PC2, the shape of heteromorphic leaves associated with genotype GG was markedly different from those of genotypes GC and CC at SNP99561 (Supplementary Figure 11C). Similar results were obtained for the significant markers for PC3 (SNP232072), PC4 (SNP232072), and PC5 (SNP34584) (Supplementary Figures 12–14C). SNP232072 was associated with PC3 and PC4, suggesting that this QTL has a pleiotropic effect on multiple leaf shape features. These results indicate that these QTLs play vital roles in regulating the heterophylly of P. euphratica.
Functional annotation showed that 115 significant SNPs accorded with P. euphratica gene models (Supplementary Tables 1, 2). 20 candidate genes exhibited functional associations only with the trait of a single leaf (Table 1), and 29 genes were involved in the heterophylly of P. euphratica (Table 2). These genes acted on transcription factors and kinases, and in auxin signaling and cell wall modification. However, there were no overlapping genes between single-and multi-leaf GWAS. Therefore, heterophylly may involve highly specific metabolic pathways or regulatory mechanisms.
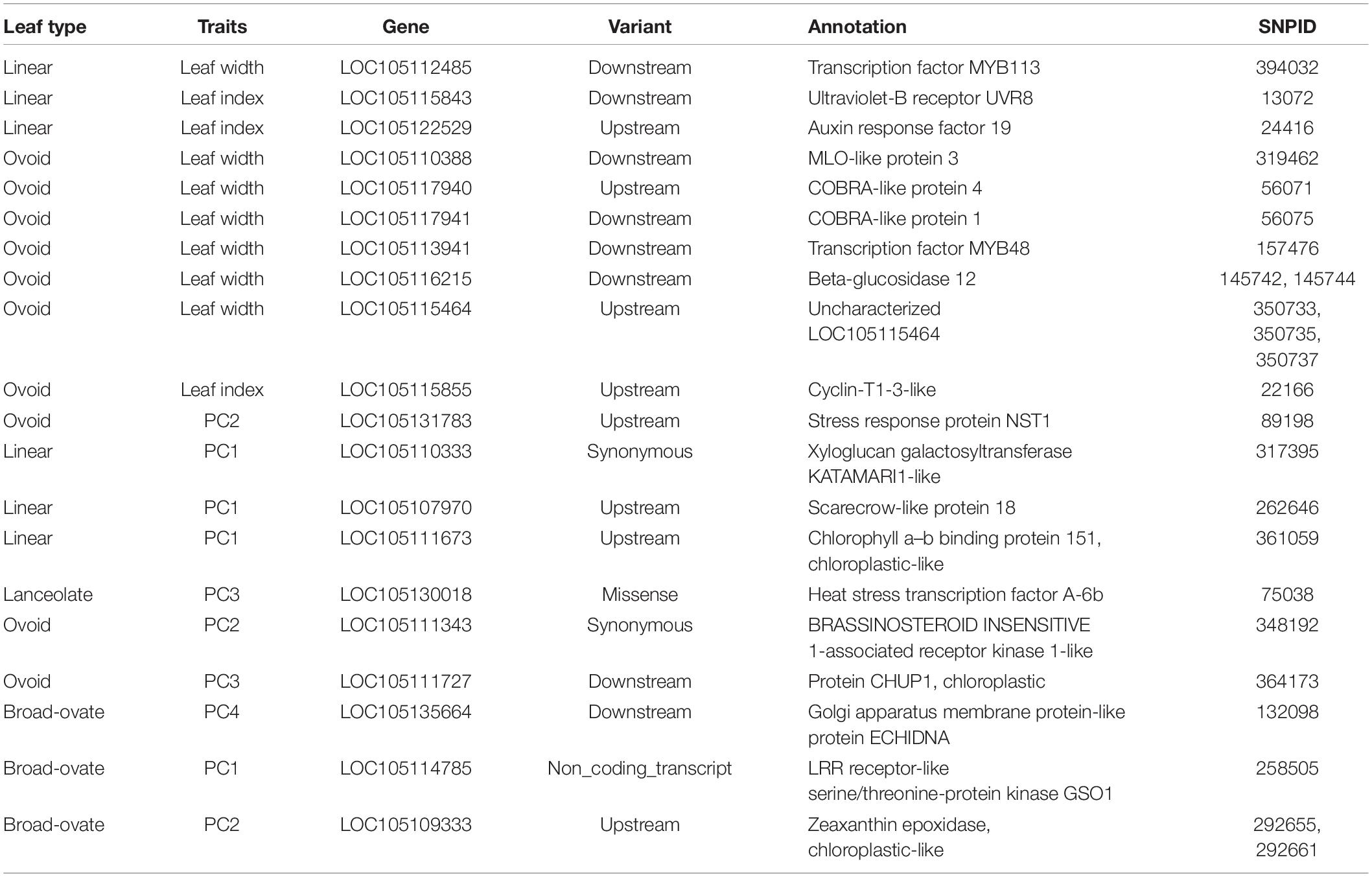
Table 1. Candidate genes identified according to significant SNPs for the different traits based on single-leaf.
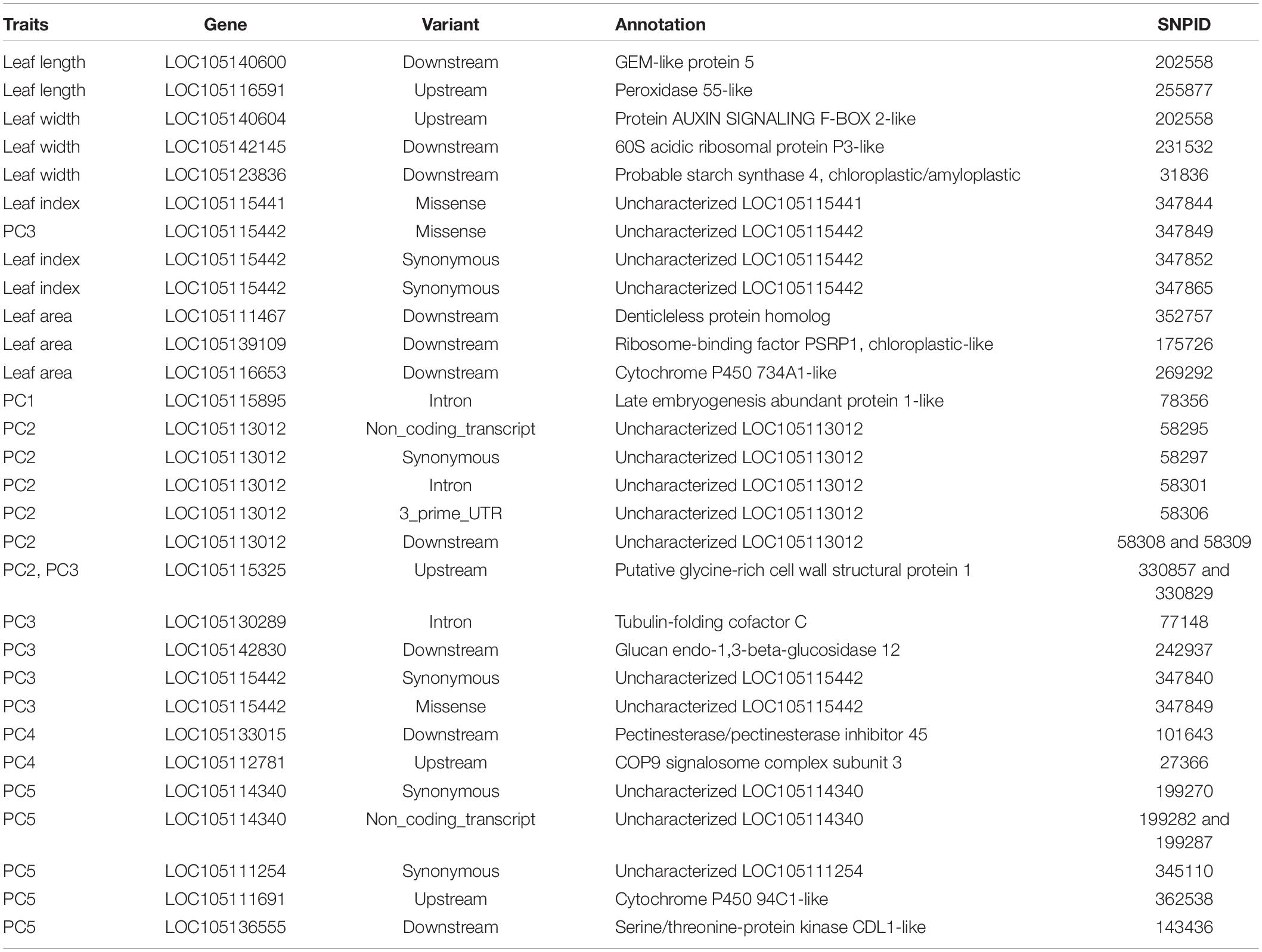
Table 2. Candidate genes identified according to significant SNPs for the different traits based on multi-leaf GWAS.
Among the candidate genes in the single-leaf GWAS, eight unique genes were closely related to the descriptive traits, while the remaining genes were associated with the PCs. LOC105112485 (transcription factors MYB113 and MYB113) and LOC105113941 (transcription factors MYB48 and MYB48) were associated with LW in linear and ovoid leaves, respectively (Table 1). LOC105115843 (ultraviolet-B receptor UVR8), which responds to ultraviolet and ultraviolet-B, was associated with LI in ovoid leaves (Table 1). Two genes (LOC105109333 and LOC105122529) were involved in the synthesis and regulation of auxin and ABA, and may play key roles in leaf development. For example, LOC105122529 (auxin response factor 19, ARF19), which binds specifically to the DNA sequence 5′-TGTCTC-3′ found in auxin-responsive promoter elements, and which could act as a transcriptional activator or repressor and be functionally redundant with ARF7, was associated with LI. Previously, it was reported that rice ARF19 controls leaf angle through positively regulating OsGH3-5 and OsBRI1 (Zhang et al., 2015). In addition, a recent study suggested that ARF6 and ARF7 control the flag leaf angle in rice by regulating secondary cell wall biosynthesis of lamina joints (Huang et al., 2021). Based on the function of ARF19 in rice, we hypothesized that the P. euphratica ARF19 protein may similarly regulate leaf development. Several genes, including LOC105135664, LOC105115855, LOC105117940, LOC105117941, and LOC105110388, may control leaf size by regulating cell division and expansion (Table 1).
Multi-leaf GWAS identified 21 genes with SNPs associated with variation in four types of leaves (Table 2). One QTL, associated with PC3 and LI, was located close to the P. euphratica LOC105115442 gene mapped on scaffold NW_011500570.1 (Supplementary Table 2), which contained three significant SNPs (SNP347849, missense; SNP347852, synonymous; and SNP347865, synonymous), but this gene was not functionally annotated (Table 2). SNP269292 (associated with LA) and SNP362538 (associated with PC5) were mapped downstream of LOC105116653 (cytochrome P450 734A1-like, CYP734A1) and upstream of LOC105111691 (cytochrome P450 94C1-like, CYP94C1), respectively (Table 2); these two genes belonged to the same CYP gene family. Chai et al. (2012) reported that the CYP78A5 gene, a member of the CYN gene family, is involved in regulating leaf development in Arabidopsis. This suggests that CYP734A1 and CYP94C1 may play important roles in heterophylly. In addition, we identified three genes involved in cell wall organization and modification (LOC105115325, LOC105142830, and LOC105133015) (Table 2), suggesting that cell wall synthesis is associated with photosynthetic performance, which affects leaf outline during the development of P. euphratica. Interestingly, we also found that the LOC105113012 gene included five significant SNPs, but the function of this gene remains unknown.
Many plant species exhibit high adaptability, altering leaf morphology in response to environmental perturbations. One of the most remarkable examples of this phenomenon is heterophylly, which is one way of dealing with the spatially and temporally varying environmental conditions presented to a plant throughout its life (Leigh et al., 2011). In some cases, heterophylly plays an important role in ecological and evolutionary processes (Nakayama et al., 2017); however, its genetic signature is still unknown (Nakayama and Kimura, 2015). Although heterophylly has been studied in several species, experimental resources for studying heterophylly in species with typical heteromorphic leaves are lacking (Nakayama et al., 2012). Therefore, a more versatile system to elucidate the mechanisms underlying heterophylly is needed. Existing studies on leaf development have focused on model species, such as A. thaliana, rice, and maize (Yang et al., 2011; Zhang et al., 2015; Du et al., 2020). However, these species do not exhibit significant heterophylly, and are therefore not ideal research targets. We used the desert tree P. euphratica as a new model species for studying heterophylly, given its marked variation in leaf shape.
P. euphratica grows linear, lanceolate, ovoid, and broad–ovate leaves between the juvenile and adult stages, and is therefore a good model species for studying leaf development (Zhao and Qin, 2017). The morphogenesis of P. euphratica heterophylly is closely related to its adaptation to high-temperature conditions, salinity, light stimuli, and other stresses (Song et al., 2021). For example, mature P. euphratica along river banks do not have more lanceolate leaves than those growing in water-scarce environments. In this study, we comprehensively evaluated the variation of heteromorphic leaves of P. euphratica at the population level, based on traditional morphological data and shape information from photographs. The morphologies of the four type leaves were obviously different, indicating heterophylly in this species. One of the highlights of this study was the quantitative description of heterophylly based on GM, which overcomes the inaccuracy of traditional shape analysis depending only on LL, LW, and LA. The shape data from the GM-based approach can precisely capture subtle changes in leaf outline, which is better for evaluating leaf variation among different genotypes of P. euphratica (Fu et al., 2013). Orthogonal PCA of “mathematical landmarks” of leaf outline was used to describe the shape changes of leaves. Five PCs explained the variance in heterophylly based on local and global facets of leaf outline.
It is important to obtain information regarding the population structure and genetic relatedness of P. euphratica accessions to identify regions of the genome associated with leaf traits. Our population structure analysis suggested that P. euphratica genotypes can be divided into two subpopulations accounting for the majority of genotypes. This was somewhat inconsistent with a previous study by Zhang et al. (2020), in which 297 genotypes sampled from the same P. euphratica population were classified into six subpopulations, with most individuals in fact originating from just one subpopulation. However, our study sample was relatively large, enabling population stratification with a high degree of confidence. Overall, the structure of our P. euphratica population did not exhibit a strong genetic signal, possibly because the genotypes were from a limited area. This may have reduced the false-positive rate in the GWAS.
To explore the genetic mechanism of heterophylly in P. euphratica more comprehensively, we used single- and multi-leaf GWAS to identify the genes controlling the formation of heteromorphic leaves. In P. euphratica, phenotypic variation may underlie the adaptability of different leaf types to environmental and climate changes. Therefore, the detailed characteristics of different leaf types were integrated into the GWAS framework to solve complex biological questions regarding heterophylly. Multi-leaf GWAS were designed to characterize leaf shape variation in multiple positions on the stem, considering descriptive traits and shape-related PCs through analysis of photographs depicting heterophylly. Linear mixed models have been widely used for association testing, to prevent the inflation associated with false-positives (Li and Zhu, 2013). However, population structure and kinship were not considered in our multi-leaf GWAS, mainly because the single-leaf GWAS demonstrated that including population structure and kinship, as in Q and Q+K models, cannot correct for inflation. Given the complex genetic correlations between multiple traits, mixed models of only one or two traits were used (Korte et al., 2012).
We performed GWAS to examine heterophylly variation based on SNPs obtained by whole-genome resequencing of the largest genome-wide dataset of genetic polymorphisms in P. euphratica reported to date. A total of 340 significant SNPs were involved in LL, LW, LI, and LA. There were 140 (41%) associations within 76 genes in the single-leaf GWAS, and 200 (59%) within 101 genes in the multi-leaf GWAS. Approximately half of the SNPs were located in intergenic regions, which may have been because the lack of high-quality complete genome sequences of P. euphratica affected the precision of SNP calling. Moreover, intergenic regions are widely distributed across the genome in plants (Yang et al., 2011), which may also be the case for P. euphratica. Intergenic areas were previously referred to as “junk DNA” because they were thought to be completely non-functional, lacking even the ability to encode RNA (Rapicavoli et al., 2013). However, some studies suggested that some intergenic regions contain potential open chromatin areas in Arabidopsis, rice, and maize, and may act as promoter elements regulating gene expression and quantitative traits (Yang et al., 2011; Du et al., 2020). These intergenic regions may play important roles in P. euphratica heterophylly. Despite the high degree of intraspecific variation in descriptive traits and PCs among accessions of P. euphratica, we found few variations in the phenotype of lanceolate leaves in the single-leaf GWAS (Supplementary Table 2). This may have been due to the low genomic coverage of the sequence data or to the effects of rare alleles, which are difficult to detect using GWAS.
The single-leaf GWAS identified two MYB transcription factors (MYB113 and MYB48) responsible for changes in leaf shape. Although a small number of MYB genes are involved in leaf development, previous studies focused on model plants and crops. For example, overexpression of OsMYB103L in rice was shown to affect the rolled leaf phenotype (Yang et al., 2014). In A. thaliana, MYB113 is associated with the gene EGL3, which affects the expression of enzymes involved in later stages of anthocyanin biosynthesis (Mondal and Roy, 2018), and MYB48, which regulates flavonol biosynthesis, primarily in cotyledons (Gaudinier et al., 2018). Therefore, MYB113 and MYB48 are closely related to the synthesis of flavonoids. Some studies suggested that flavonoid content changes during the development of leaves in various types of plants (Valkama et al., 2004), but the mechanism by which MYB regulates flavonoid synthesis in plants (and thus affects leaf development) is still unknown. Recently, Pan et al. (2020) reported that two types of MYB synergistically regulate transcription during leaf development in the hybrid poplar variety, 84K (Populus alba × Populus glandulosa). Thus, the P. euphratica genes MYB113 and MYB48 may play regulatory roles in heterophylly. A significant finding of this study was the identification of two candidate genes involved in hormone biosynthesis, ARF19 and ZEP (LOC105109333). ARF19 participates in auxin signaling in Arabidopsis (Li et al., 2006), and previous studies have shown that it also affects leaf angle in rice (Zhang et al., 2015). ZEP plays an important role in the xanthophyll cycle and ABA biosynthesis, and is highly upregulated in salt-stressed P. euphratica (Zhang et al., 2019). However, the function of the ZEP gene in leaf shape is still unknown.
Our multi-leaf GWAS showed that certain SNPs were associated with the traits of four leaf types. Importantly, the multi-leaf GWAS identified QTLs that control leaf morphology. The CYP734A1 and CYP94C1 genes identified in the GWAS have effects on LA. Cytochrome P450 is involved in brassinosteroid (BR) inactivation and regulation of BR homeostasis. CYP734A1 is associated with CYP72C1, inactivating BRs and modulating photomorphogenesis in Arabidopsis (Thornton et al., 2010). CYP94C1, encoding a bioactive phytohormone that participates in the jasmonate-mediated signaling pathway and plays a role in negative feedback control of jasmonoyl-l-isoleucine levels, also plays a role in the attenuation of jasmonate responses (Kandel et al., 2007). Noir et al. (2013) showed that methyl jasmonate inhibits leaf growth through the jasmonate receptor, COI1, by reducing cell number and size. Therefore, heterophylly appears to be inducible by CYP94C1, and its role in P. euphratica merits more in-depth analysis. We also identified a potentially pleiotropic QTL (LOC105115442) in P. euphratica that was associated with PC3 and LI. This QTL provided novel candidates underlying phenotypic variation, and suggested genomic regions important for heterophylly in P. euphratica. LOC105115441 is a missense gene neighboring LOC105115442. Notably, LOC105115442 was associated with five markers: two missense and three synonymous SNPs. Within LOC105113012, five SNPs significantly associated with PC2 were involved in multiple mutation types. This study identified novel candidate genes involved in heterophylly with potential as genetic markers for breeding P. euphratica.
The original contributions presented in the study are publicly available. This data can be found here: https://ngdc.cncb.ac.cn/, GVM000321.
XZ and FS performed the data analysis. MS, MY, WB, and AD performed the experiments and conducted fieldwork. RW and XZ conceived of the idea and designed the model. XZ and FS wrote the manuscript. All authors contributed to the article and approved the submitted version.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
This work was supported by Fundamental Research Funds for the Central Universities (No. BLX201912) and China Postdoctoral Science Foundation (2019M660496).
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fpls.2022.870876/full#supplementary-material
Adams, D. C., Rohlf, F. J., and Slice, D. E. (2013). A field comes of age: geometric morphometrics in the 21st century. Hystrix 24:7.
Ahmed, A. A., Mohamed, E. A., Hussein, M. Y., and Sallam, A. (2021). Genomic regions associated with leaf wilting traits under drought stress in spring wheat at the seedling stage revealed by GWAS. Environ. Exp. Bot. 184:104393.
Bo, W., Wang, Z., Xu, F., Fu, G., Sui, Y., Wu, W., et al. (2014). Shape mapping: genetic mapping meets geometric morphometrics. Brief. Bioinformatics 15, 571–581. doi: 10.1093/bib/bbt008
Chai, Y., Luo, D., and Yang, J. (2012). Cytochrome P450 CYP78A5/KLUH participates in regulating phase transition of leaf development in Arabidopsis thaliana. Plant Physiol. Commun. 48, 605–610.
Chen, H., Matthew, P., and Duy, T. (2020). GMMAT: Generalized Linear Mixed Model Association Tests Version 1.3. Houston, TX.
Chen, Y. N., Wang, Q., Ruan, X., Li, W. H., and Chen, Y. P. (2004). Physiological response of Populus euphratica to artificial water-recharge of the lower reaches of Tarim River. Acta Bot. Sin. 46, 1393–1401.
Churchill, G. A., and Doerge, R. W. (1994). Empirical threshold values for quantitative trait mapping. Genetics 138, 963–971. doi: 10.1093/genetics/138.3.963
Cingolani, P., Platts, A., Wang, L. L., Coon, M., Nguyen, T., Wang, L., et al. (2012). A program for annotating and predicting the effects of single nucleotide polymorphisms, SnpEff: SNPs in the genome of Drosophila melanogaster strain w1118; iso-2; iso-3. Fly 6, 80–92. doi: 10.4161/fly.19695
Conesa, A., and Götz, S. (2008). Blast2GO: a comprehensive suite for functional analysis in plant genomics. Int. J. Plant Genomics 2008, 619832. doi: 10.1155/2008/619832
Costa, M. M. R., Yang, S., Critchley, J., Feng, X., Wilson, Y., Langlade, N., et al. (2012). The genetic basis for natural variation in heteroblasty in Antirrhinum. New Phytol. 196, 1251–1259. doi: 10.1111/j.1469-8137.2012.04347.x
Danecek, P., Auton, A., Abecasis, G., Albers, C. A., Banks, E., DePristo, M. A., et al. (2011). The variant call format and VCFtools. Bioinformatics 27, 2156–2158. doi: 10.1093/bioinformatics/btr330
Dengler, N. G. (1999). Anisophylly and dorsiventral shoot symmetry. Int. J. Plant Sci. 160, S67–S80. doi: 10.1086/314218
Du, Y., Liu, L., Peng, Y., Li, M., Li, Y., Liu, D., et al. (2020). Unbranched3 expression and inflorescence development is mediated by unbranched2 and the distal enhancer, krn4, in maize. PLoS Genet. 16:e1008764. doi: 10.1371/journal.pgen.1008764
Ernande, B., and Dieckmann, U. (2004). The evolution of phenotypic plasticity in spatially structured environments: implications of intraspecific competition, plasticity costs and environmental characteristics. J. Evol. Biol. 17, 613–628. doi: 10.1111/j.1420-9101.2004.00691.x
Fu, C., Sunkar, R., Zhou, C., Shen, H., Zhang, J., Matts, J., et al. (2012). Overexpression of miR156 in switchgrass (Panicum virgatum L.) results in various morphological alterations and leads to improved biomass production. Plant Biotechnol. J. 10, 443–452.
Fu, G., Berg, A., Das, K., Jiahan, L. I., Runze, L. I., and Wu, R. (2010). A statistical model for mapping morphological shape. Theoretical Biol. Med. Modelling 7, 1–14. doi: 10.1186/1742-4682-7-28
Fu, G., Bo, W., Pang, X., Wang, Z., Chen, L., Song, Y., et al. (2013). Mapping shape quantitative trait loci using a radius-centroid-contour model. Heredity 110, 511–519. doi: 10.1038/hdy.2012.97
Fu, Y., Li, F., Mu, S., Jiang, L., Ye, M., and Wu, R. (2021). Heterophylly quantitative trait loci respond to salt stress in the desert tree Populus euphratica. Front. Plant Sci. 12:692494. doi: 10.3389/fpls.2021.692494
Gaudinier, A., Rodriguez-Medina, J., Zhang, L., Olson, A., Liseron-Monfils, C., Bågman, A. M., et al. (2018). Transcriptional regulation of nitrogen-associated metabolism and growth. Nature 563, 259–264. doi: 10.1038/s41586-018-0656-3
Guo, Z., Qin, S., Li, C., Bao, L., Yan, W., Wu, H., et al. (2020). Genome-wide identification and functional prediction of long non-coding RNAs in the genesis of Populus euphratica Oliv. heteromorphic leaves. J. Plant Biol. 63, 131–140.
Huang, G., Hu, H., van de Meene, A., Zhang, J., Dong, L., Zheng, S., et al. (2021). Auxin response factor 6 and 7 control the flag leaf angle in rice by regulating secondary cell wall biosynthesis of lamina joints. Plant Cell 33:koab175. doi: 10.1093/plcell/koab175
Iida, S., Miyagi, A., Aoki, S., Ito, M., Kadono, Y., and Kosuge, K. (2009). Molecular adaptation of rbcL in the heterophyllous aquatic plant Potamogeton. PLoS One 4:e4633. doi: 10.1371/journal.pone.0004633
Iqbal, A., Wang, T., Wu, G., Tang, W., Zhu, C., Wang, D., et al. (2017). Physiological and transcriptome analysis of heteromorphic leaves and hydrophilic roots in response to soil drying in desert Populus euphratica. Sci. Rep. 7, 1–15. doi: 10.1038/s41598-017-12091-2
Jaya, E., Kubien, D. S., Jameson, P. E., and Clemens, J. (2010). Vegetative phase change and photosynthesis in Eucalyptus occidentalis: architectural simplification prolongs juvenile traits. Tree Physiol. 30, 393–403. doi: 10.1093/treephys/tpp128
Kandel, S., Sauveplane, V., Compagnon, V., Franke, R., Millet, Y., Schreiber, L., et al. (2007). Characterization of a methyl jasmonate and wounding-responsive cytochrome P450 of Arabidopsis thaliana catalyzing dicarboxylic fatty acid formation in vitro. The FEBS journal 274, 5116–5127. doi: 10.1111/j.1742-4658.2007.06032.x
Kaplan, D. R. (1980). Heteroblastic leaf development in Acacia: morphological and Morphogenetic Implications. La Cellule 73, 137–203.
Kim, J., Joo, Y., Kyung, J., Jeon, M., Park, J. Y., Lee, H. G., et al. (2018). A molecular basis behind heterophylly in an amphibious plant, Ranunculus trichophyllus. PLos Genet. 14:e1007208. doi: 10.1371/journal.pgen.100720
Kitajima, K., Mulkey, S. S., and Wright, S. J. (1997). Decline of photosynthetic capacity with leaf age in relation to leaf longevities for five tropical canopy tree species. Am. J. Bot. 84, 702–708.
Korte, A., Vilhjálmsson, B. J., Segura, V., Platt, A., Long, Q., and Nordborg, M. (2012). A mixed-model approach for genome-wide association studies of correlated traits in structured populations. Nat. Genet. 44, 1066–1071. doi: 10.1038/ng.2376
Kuwabara, A., Ikegami, K., Koshiba, T., and Nagata, T. (2003). Effects of ethylene and abscisic acid upon heterophylly in Ludwigia arcuata (Onagraceae). Planta 217, 880–887. doi: 10.1007/s00425-003-1062-z
Langlade, N. B., Feng, X., Dransfield, T., Copsey, L., Hanna, A. I, Thébaud, C., et al. (2005). Evolution through genetically controlled allometry space. Proc. Natl. Acad. Sci. U.S.A. 102, 10221–10226. doi: 10.1073/pnas.0504210102
Lee, D. W., and Richards, J. H. (1991). “Heteroblastic development in vines,” in The Biology of Vines, eds F. E. Putz and H. A. Mooney (Cambridge: Cambridge University Press), 205–243. doi: 10.1002/pld3.28
Leigh, A., Zwieniecki, M. A., Rockwell, F. E., Boyce, C. K., Nicotra, A. B., and Holbrook, N. M. (2011). Structural and hydraulic correlates of heterophylly in Ginkgo biloba. New Phytol. 189, 459–470. doi: 10.1111/j.1469-8137.2010.03476.x
Li, C., Qin, S., Bao, L., Guo, Z., and Zhao, L. (2020). Identification and functional prediction of circRNAs in Populus Euphratica Oliv. heteromorphic leaves. Genomics 112, 92–98.
Li, G., and Zhu, H. (2013). Genetic studies: the linear mixed models in genome-wide association studies. Open Bioinformatics J. 7, 27–33.
Li, H., and Durbin, R. (2009). Fast and accurate short read alignment with Burrows-Wheeler transform. Bioinformatics 25, 1754–1760. doi: 10.1093/bioinformatics/btp324
Li, H., Handsaker, B., Wysoker, A., Fennell, T., Ruan, J., Homer, N., et al. (2009). The sequence alignment/map format and SAMtools. Bioinformatics 25, 2078–2079. doi: 10.1093/bioinformatics/btp352
Li, J., Dai, X., and Zhao, Y. (2006). A role for auxin response factor 19 in auxin and ethylene signaling in Arabidopsis. Plant Physiol. 140, 899–908. doi: 10.1104/pp.105.070987
Ling, H., Zhang, P., Xu, H., and Zhao, X. (2015). How to regenerate and protect desert riparian Populus euphratica forest in arid areas. Sci. Rep. 5, 1–12. doi: 10.1038/srep15418
Ma, T., Wang, J., Zhou, G., Yue, Z., Hu, Q., Chen, Y., et al. (2013). Genomic insights into salt adaptation in a desert poplar. Nat. Commun. 4, 1–9. doi: 10.1038/ncomms3797
Mähler, N., Schiffthaler, B., Robinson, K. M., Terebieniec, B. K., Vučak, M., Mannapperuma, C., et al. (2020). Leaf shape in Populus tremula is a complex, omnigenic trait. Ecol. Evol. 10, 11922–11940. doi: 10.1002/ece3.6691
Manichaikul, A., Mychaleckyj, J. C., Rich, S. S., Daly, K., Sale, M., and Chen, W. M. (2010). Robust relationship inference in genome-wide association studies. Bioinformatics 26, 2867–2873. doi: 10.1093/bioinformatics/btq559
Manuela, D., and Xu, M. (2020). Juvenile leaves or adult leaves: determinants for vegetative phase change in flowering plants. Int. J. Mol. Sci. 21:9753. doi: 10.3390/ijms21249753
McKenna, A., Hanna, M., Banks, E., Sivachenko, A., Cibulskis, K., Kernytsky, A., et al. (2010). The Genome Analysis Toolkit: a MapReduce framework for analyzing next-generation DNA sequencing data. Genome Res. 20, 1297–1303. doi: 10.1101/gr.107524.110
Mondal, S. K., and Roy, S. (2018). Genome-wide sequential, evolutionary, organizational and expression analyses of phenylpropanoid biosynthesis associated MYB domain transcription factors in Arabidopsis. J. Biomol. Struct. Dyn. 36, 1577–1601. doi: 10.1080/07391102.2017.1329099
Nakayama, H., and Kimura, S. (2015). Leaves may function as temperature sensors in the heterophylly of Rorippa aquatica (Brassicaceae). Plant Signal. Behav. 10:e1091909. doi: 10.1080/15592324.2015.1091909
Nakayama, H., Nakayama, N., Nakamasu, A., Sinha, N., and Kimura, S. (2012). Toward elucidating the mechanisms that regulate heterophylly. Plant Morphol. 24, 57–63.
Nakayama, H., Nakayama, N., Seiki, S., Kojima, M., Sakakibara, H., Sinha, N., et al. (2014). Regulation of the KNOX-GA gene module induces heterophyllic alteration in North American lake cress. Plant Cell 26, 4733–4748. doi: 10.1105/tpc.114.130229
Nakayama, H., Sinha, N. R., and Kimura, S. (2017). How do plants and phytohormones accomplish heterophylly, leaf phenotypic plasticity, in response to environmental cues. Front. Plant Sci. 8:1717. doi: 10.3389/fpls.2017.01717
Noir, S., Bömer, M., Takahashi, N., Ishida, T., Tsui, T. L., Balbi, V., et al. (2013). Jasmonate controls leaf growth by repressing cell proliferation and the onset of endoreduplication while maintaining a potential stand-by mode. Plant Physiol. 161, 1930–1951. doi: 10.1104/pp.113.214908
Pan, C., Bu, Y. F., and Jing, Y. P. (2020). Cloning and expression analysis of five MYB genes in poplar during leaf development. Genomics Appl. Biol. 39, 4090–4098.
Pigliucci, M., and Marlow, E. T. (2001). Differentiation for flowering time and phenotypic integration in Arabidopsis thaliana in response to season length and vernalization. Oecologia 127, 501–508. doi: 10.1007/s004420000613
Poethig, R. S. (1988). Heterochronic mutations affecting shoot development in maize. Genetics 119, 959–973. doi: 10.1093/genetics/119.4.959
Poethig, R. S. (2013). Vegetative phase change and shoot maturation in plants. Curr. Topics Dev. Biol. 105, 125–152. doi: 10.1016/B978-0-12-396968-2.00005-1
Purcell, S., Neale, B., Todd-Brown, K., Thomas, L., Ferreira, M. A., Bender, D., et al. (2007). PLINK: a tool set for whole-genome association and population-based linkage analyses. Am. J. Hum. Genet. 81, 559–575. doi: 10.1086/519795
Qin, S. W., Jiang, R. J., Zhang, N., Liu, Z. W., Li, C. L., Guo, Z. Z., et al. (2018). Genome-wide analysis of RNAs associated with Populus euphratica Oliv. heterophyll morphogenesis. Sci. Rep. 8, 1–9.
R Core Team (2021). R: A Language And Environment For Statistical Computing. Vienna: R Foundation for Statistical Computing.
Raj, A., Stephens, M., and Pritchard, J. K. (2014). fastSTRUCTURE: variational inference of population structure in large SNP data sets. Genetics 197, 573–589. doi: 10.1534/genetics.114.164350
Rapicavoli, N. A., Qu, K., Zhang, J., Mikhail, M., Laberge, R. M., and Chang, H. Y. (2013). A mammalian pseudogene lncRNA at the interface of inflammation and anti-inflammatory therapeutics. Elife 2:e00762. doi: 10.7554/eLife.00762
Rubio-Somoza, I., Zhou, C. M., Confraria, A., Martinho, C., von Born, P., Baena-Gonzalez, E., et al. (2014). Temporal control of leaf complexity by miRNA-regulated licensing of protein complexes. Curr. Biol. 24, 2714–2719. doi: 10.1016/j.cub.2014.09.058
Sato, M., Tsutsumi, M., Ohtsubo, A., Nishii, K., Kuwabara, A., and Nagata, T. (2008). Temperature-dependent changes of cell shape during heterophyllous leaf formation in Ludwigia arcuata (Onagraceae). Planta 228, 27–36. doi: 10.1007/s00425-008-0715-3
Sicard, A., Thamm, A., Marona, C., Lee, Y. W., Wahl, V., Stinchcombe, J. R., et al. (2014). Repeated evolutionary changes of leaf morphology caused by mutations to a homeobox gene. Curr. Biol. 24, 1880–1886. doi: 10.1016/j.cub.2014.06.061
Simon, B., David, T., Jan, W., Haz, E., and Shota, O. (2021). Image Processing Library Based on ‘CImg’. Grenoble.
Song, Z., Ni, X., Yao, J., and Wang, F. (2021). Progress in studying heteromorphic leaves in Populus euphratica: leaf morphology, anatomical structure, development regulation and their ecological adaptation to arid environments. Plant Signal. Behav. 16:1870842. doi: 10.1080/15592324.2020.1870842
Sun, L., Wang, J., Zhu, X., Jiang, L., Gosik, K., Sang, M., et al. (2018). Hp QTL: a geometric morphometric platform to compute the genetic architecture of heterophylly. Brief. Bioinformatics 19, 603–612. doi: 10.1093/bib/bbx011
Thornton, L. E., Rupasinghe, S. G., Peng, H., Schuler, M. A., and Neff, M. M. (2010). Arabidopsis CYP72C1 is an atypical cytochrome P450 that inactivates brassinosteroids. Plant Mol. Biol. 74, 167–181. doi: 10.1007/s11103-010-9663-y
Tian, F., Bradbury, P. J., Brown, P. J., Hung, H., Sun, Q., Flint-Garcia, S., et al. (2011). Genome-wide association study of leaf architecture in the maize nested association mapping population. Nat. Genet. 43, 159–162. doi: 10.1038/ng.746
Valkama, E., Salminen, J. P., Koricheva, J., and Pihlaja, K. (2004). Changes in leaf trichomes and epicuticular flavonoids during leaf development in three birch taxa. Ann. Bot. 94, 233–242. doi: 10.1093/aob/mch131
Viscosi, V., and Cardini, A. (2011). Leaf morphology, taxonomy and geometric morphometrics: a simplified protocol for beginners. PLoS One 6:e25630. doi: 10.1371/journal.pone.0025630
Wu, G., Park, M. Y., Conway, S. R., Wang, J. W., Weigel, D., and Poethig, R. S. (2009). The sequential action of miR156 and miR172 regulates developmental timing in Arabidopsis. Cell 138, 750–759. doi: 10.1016/j.cell.2009.06.031
Xu, Y., Qian, Z., Zhou, B., and Wu, G. (2019). Age-dependent heteroblastic development of leaf hairs in Arabidopsis. New Phytol. 224, 741–748. doi: 10.1111/nph.16054
Yang, C., Li, D., Liu, X., Ji, C., Hao, L., Zhao, X., et al. (2014). OsMYB103L, an R2R3-MYB transcription factor, influences leaf rolling and mechanical strength in rice (Oryza sativa L.). BMC Plant Biol. 14:158. doi: 10.1186/1471-2229-14-158
Yang, L., Xu, M., Koo, Y., He, J., and Poethig, R. S. (2013). Sugar promotes vegetative phase change in Arabidopsis thaliana by repressing the expression of MIR156A and MIR156C. Elife 2:e00260. doi: 10.7554/eLife.00260
Yang, X. H. W. C., Xia, X. Y., and Gang, S. H. (2011). Genome-wide analysis of intergenic regions in Arabidopsis thaliana suggests the existence of bidirectional promoters and genetic insulators. Curr.Topics Plant Biol. 12, 15–33.
Ye, M., Zhu, X., Gao, P., Jiang, L., and Wu, R. (2020). Identification of quantitative trait loci for altitude adaptation of tree leaf shape with Populus szechuanica in the qinghai-tibetan plateau. Front. Plant Sci. 11:632. doi: 10.3389/fpls.2020.00632
Zeng, M., He, S., Hao, L., Li, Y., Zheng, C., and Zhao, Y. (2019). Conjoint analysis of genome-wide lncRNA and mRNA expression of heteromorphic leavesin response to environmental heterogeneityin Populus euphratica. Int. J. Mol. Sci. 20:5148. doi: 10.3390/ijms20205148
Zhai, J. T., Li, Y. L., Han, Z. J., and Li, Z. J. (2020). Morphological, structural and physiological differences in heteromorphic leaves of Euphrates poplar during development stages and at crown scales. Plant Biol. 22, 366–375. doi: 10.1111/plb.13078
Zhang, C., Luo, W., Li, Y., Zhang, X., Bai, X., Niu, Z., et al. (2019). Transcriptomic analysis of seed germination under salt stress in two desert sister species (Populus euphratica and P. pruinosa). Front. Genet. 10:231. doi: 10.3389/fgene.2019.00231
Zhang, Q., Su, Z., Guo, Y., Zhang, S., Jiang, L., and Wu, R. (2020). Genome-wide association studies of callus differentiation for the desert tree, Populus euphratica. Tree Physiol. 40, 1762–1777. doi: 10.1093/treephys/tpaa098
Zhang, S., Wang, S., Xu, Y., Yu, C., Shen, C., Qian, Q., et al. (2015). The auxin response factor, OsARF 19, controls rice leaf angles through positively regulating OsGH 3-5 and OsBRI 1. Plant Cell Environ. 38, 638–654. doi: 10.1111/pce.12397
Zheng, X., Levine, D., Shen, J., Gogarten, S. M., Laurie, C., and Weir, B. S. (2012). A high-performance computing toolset for relatedness and principal component analysis of SNP data. Bioinformatics 28, 3326–3328.
Zhao, L., and Qin, S. (2017). Expression profiles of miRNAs in the genesis of Populus euphratica Oliv. heteromorphic leaves. Plant Growth Regul. 81, 231–242.
Zhao, W., Chen, Y. Q., Casella, G., Cheverud, J. M., and Wu, R. (2005). A non-stationary model for functional mapping of complex traits. Bioinformatics 21, 2469–2477. doi: 10.1093/bioinformatics/bti382
Zhao, X., Luo, L., Cao, Y., Liu, Y., Li, Y., Wu, W., et al. (2018). Genome-wide association analysis and QTL mapping reveal the genetic control of cadmium accumulation in maize leaf. BMC Genomics 19:91. doi: 10.1186/s12864-017-4395-x
Keywords: heterophylly, leaf shape, geometric morphometrics, genome-wide association study (GWAS), Populus euphratica
Citation: Zhu X, Sun F, Sang M, Ye M, Bo W, Dong A and Wu R (2022) Genetic Architecture of Heterophylly: Single and Multi-Leaf Genome-Wide Association Mapping in Populus euphratica. Front. Plant Sci. 13:870876. doi: 10.3389/fpls.2022.870876
Received: 07 February 2022; Accepted: 24 May 2022;
Published: 15 June 2022.
Edited by:
Josh Clevenger, HudsonAlpha Institute for Biotechnology, United StatesReviewed by:
Cheng-Ruei Lee, National Taiwan University, TaiwanCopyright © 2022 Zhu, Sun, Sang, Ye, Bo, Dong and Wu. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Rongling Wu, Uld1QHBocy5wc3UuZWR1
†These authors have contributed equally to this work
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.