
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
MINI REVIEW article
Front. Plant Sci. , 17 May 2022
Sec. Plant Systematics and Evolution
Volume 13 - 2022 | https://doi.org/10.3389/fpls.2022.854372
This article is part of the Research Topic Solanaceae VII: Biology, Genetics, and Evolution View all 15 articles
 Andrés Moreira-Muñoz1*†
Andrés Moreira-Muñoz1*† María Virginia Palchetti2,3
María Virginia Palchetti2,3 Vanezza Morales-Fierro4
Vanezza Morales-Fierro4 Valeria Soledad Duval5Rudy Allesch-Villalobos1Carlos E. González-Orozco6†
Valeria Soledad Duval5Rudy Allesch-Villalobos1Carlos E. González-Orozco6†There is a need to make substantial advances in the taxonomic, systematic, and distribution knowledge of plants, and find better ways of transmission of this information to society to surpass the general pattern described as “plant blindness.” The diversity of the plant family Solanaceae reaches its peak in South America; however, many of its species are threatened due to the expansion of the human footprint. Here, we examine the diversity patterns of the family in southern South America (Argentina and Chile) by means of species richness (SR), weighted endemism (WE), and corrected weighted endemism (CWE). We also evaluated conservation gaps in relation to protected areas and the human footprint as a proxy for potential impacts on this biodiversity. Results show two richness centers in NW and NE Argentina, with a high degree of overlap with protected areas, which, on the other side, show a relative high index of human footprint. Comparatively, coastal Atacama (Chile) shows lower richness values, but outstanding CWE and WE values. The coast of Atacama harbors high values due the presence of species of the genus Nolana with restricted distributions. Protected areas in this tight coastal strip are sparse, and the human footprint is also relatively high. The degree of protection based on these parameters is then unbalanced, highlighting the need for a geographically explicit strategy for the conservation of the family at subcontinental scale. In doing so, it is likely that other representatives of these unique centers of richness and endemism will benefit.
Plant conservation is limited by our knowledge of the diversity, distribution, and abundance of plant species (Gillson et al., 2020). This information is increasing but cannot keep pace with the threats plants suffer, leading to accelerated anthropogenic-caused extinctions and genetic erosion (Knapp, 2019; Wandersee and Schussler, 2001).
South America is one region where different approaches (e.g., taxonomy, phylogeny, biogeography, ethnobotany) are leading to the discovery of new species. Unfortunately, this diversity is dwindling across the continent (Ramirez-Villegas et al., 2012). Rapid land use changes, wildfires, and, in general, human footprint expansion (Zalles et al., 2021) are putting species and ecosystems increasingly under threat. Regional climate change amplifies these threats across biodiversity hotspots (Fuentes-Castillo et al., 2020).
Of special interest for plant conservation at a continental and subcontinental scale are several angiosperm groups that exhibit early diversification in South America, including the Bignoniaceae, Verbenaceae, Asteraceae, and Solanaceae (Olmstead, 2013; Dupin et al., 2017; Deanna et al., 2020). Obtaining a comprehensive understanding of the diversity and distribution of these taxa in South America is a challenge, but this effort is paramount to guide future conservation efforts.
The family Solanaceae encompasses approximately 2,800 species globally (98 genera). It is among the 10 families with the greatest diversity in countries considered to be megadiverse, such as Ecuador and Bolivia (Ulloa Ulloa et al., 2017). In addition, many solanaceous species are important food resources (Samuels, 2015).
Representatives of the Solanaceae family are distributed in the Americas from Alaska to Patagonia, from the sea level to the heights of the Andes (e.g., Solanum acaule, Jaborosa squarrosa, and Lycium humile (Barboza, 2013; Palchetti et al., 2021). Therefore, the family has inspired important biogeographic studies across the continent (Hijmans and Spooner, 2001; Anguiano-Constante et al., 2018).
The southern end of the continent is where the family reaches the greatest levels of diversity as the fifth largest family of the flora of the Southern Cone of South America after the Asteraceae, Poaceae, Fabaceae, and Orchidaceae (Zuloaga et al., 2019) (Figure 1).

Figure 1. Floral morphology and habitat diversity of southern Solanaceae. (A) Schizanthus porrigens and (B) Schizanthus parvula from Central Chile; (C) Fabiana denudata in Catamarca, Argentina; (D) Fabiana ramulosa in Chilean Altiplano; (E) Exodeconus flavus in Tarapacá Precordillera, north Chile; (F) Solanum trinominum in Chilean coastal sand dunes; (G) Salpichroa glandulosa and (H) Dunalia spinosa in the heights of Parinacota, Chile; (I,J) Nolana mollis and N. villosa on the lomas formation, Chile; (K) Lycium humile on salt-rich environments of the Altiplano highlands. Photographs by M. Virginia Palchetti, Rocío Deanna, and Andrés Moreira-Muñoz.
The type genus of the family is Solanum, a genus of nearly cosmopolitan distribution, published by Linnaeus (1753) in Species Plantarum. It is the second most diverse genus of the vascular plants in the Southern Cone (216 spp.) after Senecio (Asteraceae, 415 spp.) (Zuloaga et al., 2019).
Solanaceae is the fourth family in species richness in Argentina (Palchetti et al., 2020) as it is in Chile (Moreira-Muñoz, 2011). With 52% of endemic species, Chile stands out as the most relative endemism-rich country for the Solanaceae, followed by the megadiverse Peru and Brazil (47%) (Palchetti et al., 2020). More diverse genera in Argentina are Solanum and Lycium, while, in Chile, the highest diversity is represented by Solanum and Nolana. In Argentina, ecoregions with highest diversity are Chaco, Andes, and Pampa, and highest endemism occurs in Chaco, Andes, Yungas, and Monte ecoregions (Oyarzabal et al., 2018; Del Valle Elías and Aagesen, 2019; Palchetti et al., 2020; Arana et al., 2021). In Chile, the most outstanding ecoregion is the Desert Scrub, as defined by Luebert and Pliscoff (2017). The southern Andes has played a central role in the early diversification of the Solanaceae. The history of elevation change in the Andes occurred concurrently with plant evolution and influenced it, the mountains acting as a corridor, a barrier or providing a geodiversity framework for species diversification (Luebert and Weigend, 2014; Moreira-Muñoz et al., 2020).
Recent advances in the knowledge of the taxonomy of this family have challenged us to update the overall understanding of diversity and conservation priorities. Ca. 30 species are considered as threatened in Argentina (Palchetti et al., 2020) and 14 species in Chile, but most species have not been assessed yet.1
Our main goal in this study was to map and analyze the diversity of the Solanaceae in southern South America (Argentina and Chile), overlaying regional richness and endemism with protected areas to identify conservation gaps. Additionally, the human footprint in the existing protected areas is evaluated as a proxy for the degree of effective protection of the family. In this way, the areas of geographic concentration (richness micro-hotspots and centers of endemism) of the family can be identified, which can guide future floristic prospecting and identify areas under threat from land uses incompatible with conservation.
The distribution and richness analysis for the family was carried out through a compilation of a database, including different sources of information. Data for Argentina come mostly from the Documenta Florae Australis (2021), while the data for Chile collate specimen information from national (CONC and SGO) and international herbaria. The latter are available through the Global Biodiversity Information Facility (GBIF) platform. Specific status based on recent studies and reports published after April 2019 (not included in Palchetti et al., 2020) has been considered. This included studies, such as the revision of the genus Schizanthus (Morales-Fierro et al., 2020; Lavandero et al., 2021); updates in Nolana (Hepp and Dillon, 2018); the Morelloid clade of Solanum in Argentina (Knapp et al., 2020); cryptic species recently reported (Moreira-Muñoz and Muñoz-Schick, 2020); and new Petunia and Nicotiana species (Greppi et al., 2019; Santilli et al., 2021). After a first cleaning, data from ex situ living collections that introduced species, hybrid taxa, and records with doubtful or incomplete identification at the species level were excluded. Despite the fact that most of the records had georeferenced data in their originally source, errors were detected and corrected (19% of the total records). For this purpose, the Geonames2 and Mapcarta3 sites were used. Species nomenclature was based on the following sources: Flora del Cono Sur,4 POWO,5 and Solanaceae Source.6 After eliminating duplicate coordinates for each species, the final database consists of 15,510 records, which include 35 genera and 423 species (Supplementary Material).
Diversity maps were carried out by means of the Biodiverse 3.1 software. (Laffan et al., 2010).7 We used grid cells of 1 degree (latitude and longitude) (the most suited resolution at subcontinental scale), and three diversity indices were computed from the grid-cell data. The species richness (SR) of a cell is defined as the total number of species within that grid cell. Weighted endemism (WE) is the sum, over all species present in the window for that grid cell, of the number of grid cells in the window with that species divided by the range of that species. The range is defined as the total number of all grid cells in which that species is present. Crisp et al. (2001) defined corrected weighted endemism CWE as the weighted endemism (WE) divided by the total number of species in that window. This last division adjusts the index for the effect of SR. To assess the confidence in the identified centers of endemism, we conducted a randomization test (Laffan and Crisp, 2003). The test generated 999 random iterations, each of which preserves the observed SR of each geographically located cell, and the total number of cells, or ranges, for each species. CWE was then calculated for each random iteration, and the original ranked against the randomizations. Cells with CWE randomization ranks in the top 5% are significantly different from random at a threshold of alfa = 0.05 (González-Orozco et al., 2011) (Supplementary Material).
Gap analysis was carried out by means of the superposition of the richness units with available information on protected areas. Argentina-protected units were a downloaded official site.8 Chilean units are available on the SNIT Geoportal.9 Additionally, as a proxy for the state of protection of species within protected areas, we overlayed them with the “human footprint” index, as developed by Sanderson et al. (2002) by means of ArcGis 10.3 (ESRI, 2015). The human footprint is a quantitative analysis and representation of human influence across the planetary surface based upon four types of data: population density, land transformation, accessibility, and electrical power infrastructure. Human impact is represented on a scale of 0 (minimum) to 100 (maximum) on a resolution of one square kilometer (Sanderson et al., 2002). A score of 1 indicates the least human influence. The shape file for South America was obtained from10 and overlapped with the protected areas in Chile and Argentina. We calculated the mean of the human footprint in the set of pixels, encompassing a protected area (Table 1 and Supplementary Material).
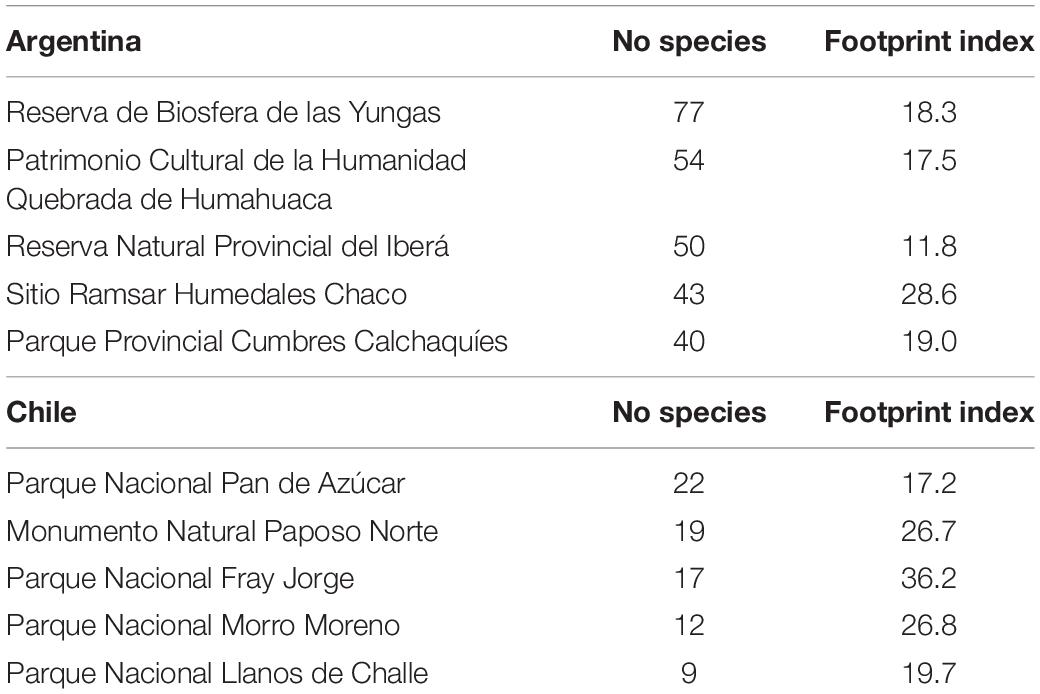
Table 1. Protected areas in Argentina and Chile with the higher number of Solanaceae species.
The number of native Solanaceae species in Argentina and Chile is 430 and belongs to 35 genera. Both countries share 18 genera and 55 species, of which 27 are endemic to Argentina and Chile. Argentina has 315 native species, distributed in 32 genera, of which 80 are endemic species (25%). Chile has 170 native species, distributed in 21 genera, of which 89 are endemic species (52%). Supplementary Material considering Argentina and Chile, a total of 8 genera are endemic (i.e., Benthamiella, Combera, Reyesia, Salpiglossis, and Schizanthus shared between countries; the monotypic genus Panthacantha only grows in Argentina, and the monotypic genera Latua and Vestia in Chile). The most speciose genus in Chile is Nolana, with 49 native species, while in Argentina is Solanum, with 126 native species.
According to Biodiverse 3.1 outputs, the primary centers of SR are in the northeast and northwestern regions of Argentina and coastal areas in the north of Chile (Figure 2). The SR scores ranged between 1 and 111 species, but SR maximum value of 47 at a threshold of 5–95% was found in a single-grid cell. We identified three main hotspots of WE: east and west northern corner of Argentina and the coastal areas in the north of Chile. WE scores ranged between 0.007 and 11.71, but a WE maximum value of 5.5 (5.5% of species are endemic to that grid cell) at a threshold of 5–95% was found in a single grid cell (Figure 2). Once species richness is being corrected, two of the main WE centers remained in the same location, and the one in the northwestern corner of Argentina tended to disappear. However, new areas of high CWE appeared in the south of Chile and Argentina as well as a few scattered grid cells in the central regions. Interestingly, the northern hotspot of endemism in Chile increases in size under the CWE. These changes are likely because of richness biases on specific grid cells. CWE scores ranged between 0 and 0.50, but a CWE maximum value of 0.15 (after a correction of richness, 15% of species are endemic to that grid cell) at a threshold of 5–95% was found in a single-grid cell. To test the validity of spatial CWE patterns, the randomization results show that all major identified centers of endemism were significantly different from random at a threshold of alfa = 0.05 (Figure 2).
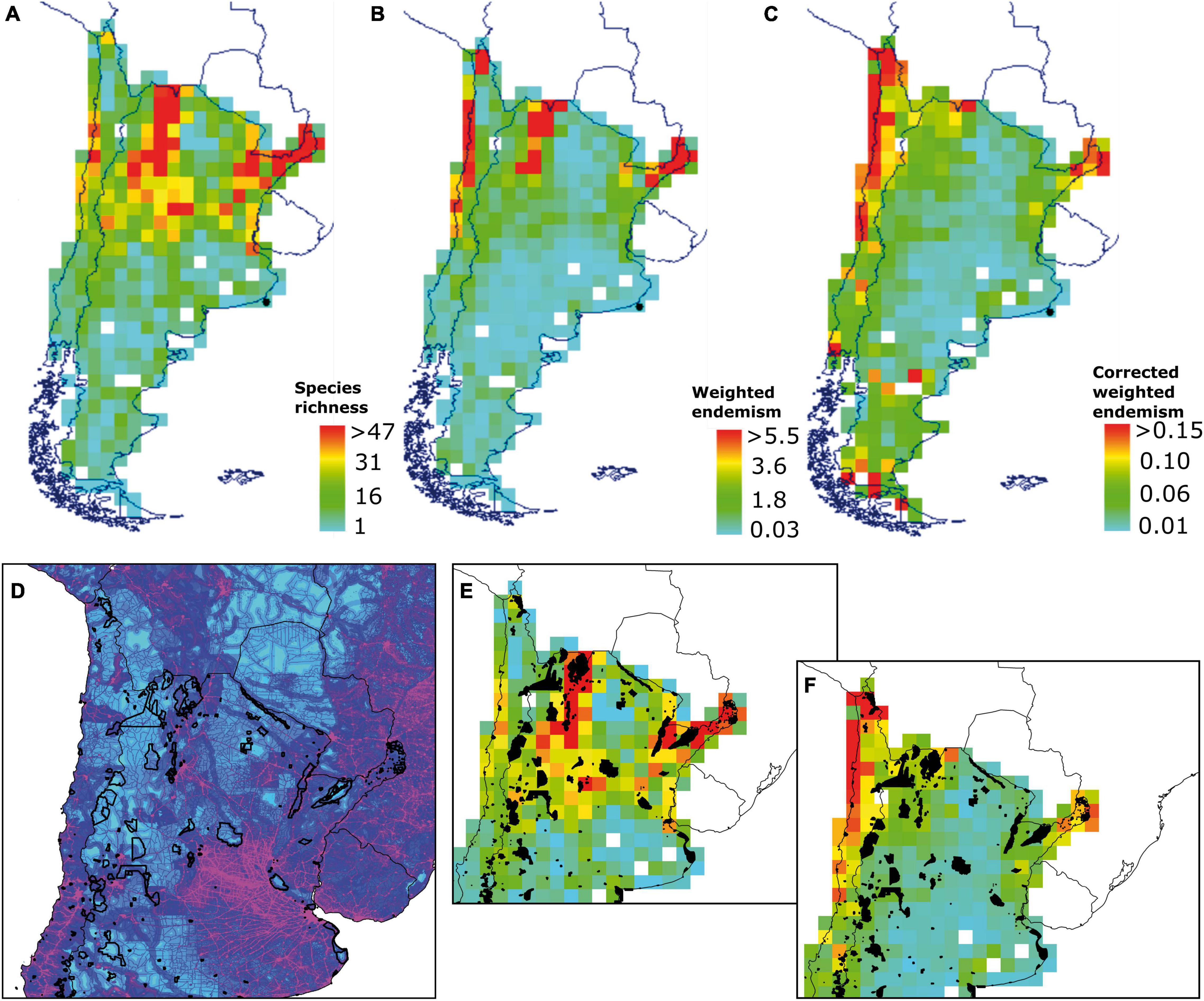
Figure 2. Diversity of Solanaceae species in southern South America, mapped using Biodiverse 3.1 on 1 × 1 degree matrix: (A) Species richness; (B) Weighted endemism; (C) Corrected weighted endemism; (D) A detailed area showing a footprint index and protected areas; (E) Protected areas overlapped with species richness; (F) Protected areas overlapped with corrected weighted endemism. Detailed maps and tables are available as (Supplementary Material).
In synthesis, NW and NE Argentina and the northern Atacama coast can be considered as richness/endemism centers, or micro-hotspots of biodiversity, at the margins of globally recognized biodiversity hotspots (Supplementary Material).
Gap analysis shows that, from 331 protected areas in Argentina, 129 have a degree of spatial coincidence with the distribution of Solanaceae. In Chile, 38 from 102 protected units superpose with cells with the distribution of Solanaceae. Protected areas with highest number of species in Chile and Argentina are those summarized in Table 1. Protected areas in the northwest of Argentina show a spatial match with richness centers and, to some lower degree, in NE Argentina (Figure 2E). The presence of protected areas in Chile is sparse at the coast of Atacama, where the highest endemism appears. The human footprint is higher in Central Chile and Central/northern Argentina, around metropolitan central areas (Supplementary Material). The mean value of the human print index in each protected area (PA) varies between 1.1 and 87 in Argentina (20.1 total mean). The index varies between 1.8 and 71 in the case of PA in Chile (19.7 total mean). PA areas encompassing high numbers of Solanaceae and a relatively high index are Sitio Ramsar Humedales Chaco in Argentina and Parque Nacional Fray Jorge in Chile (Table 1).
One of the greatest current challenges in conservation biogeography is identifying areas of high species richness and endemism, both to establish conservation priorities and to better understand the evolution of plant diversity. This is especially relevant in southern South America, a territory recognized as especially important in the evolution of diverse families, such as Solanaceae, Bignoniaceae, Verbenaceae, Asteraceae, Orchidaceae (Olmstead, 2013; Ulloa Ulloa et al., 2017). In southern South America, the wide geographic distribution of the family Solanaceae and its taxonomic richness is partly explained by a long evolutionary history since the early Eocene (Dupin et al., 2017; Deanna et al., 2020).
Diversity indexes applied to this wide study area show different regions as outstanding for Solanaceae species richness (SR) and endemism, respectively. Weighted endemism (WE) and corrected weighted endemism (CWE) are parameters that have shown great utility for analysis of restricted distributions (Sosa and de Nova, 2012; Rodríguez et al., 2018; Ruiz-Sánchez et al., 2020). SR shows two main areas in NW and NE Argentina. Richness areas show an important degree of protection mainly by “Reserva de Biosfera de las Yungas” and “Patrimonio Cultural de la Humanidad Quebrada de Humahuaca” in the NW, and “Reserva Natural Provincial del Iberá” and “Sitio Ramsar Humedales Chaco” in the NE. Both regions have been largely recognized as important sources of biodiversity and medicinal plants (Hilgert and Gil, 2006; Bernacki et al., 2015; Campanello et al., 2019). Especially Yungas has been recognized as an outstanding ecoregion for the conservation of different biotic groups (Grosso and Quintana, 2009; Arana et al., 2016; Torres and González-Reyes, 2017). When weighted endemism (WE) and corrected weighted endemism (CWE) are applied, another region clearly appears as outstanding at the regional scale: the coast of Atacama, mainly due the diversity and restricted distribution of species in genus Nolana. This genus is a main component of Lomas vegetation, a plant formation found along the coast from Peru to northern Chile, characterized by high endemism and richness maintained by the coastal fog reaching the coastal cliffs at an altitude around 1,000 m asl (Muñoz-Schick et al., 2001). The Solanaceae component of this remarkable environment has been emphasized by Dillon (2005, 2016), and other components of this unique biota have been recently highlighted (Moat et al., 2021; Pizarro-Araya et al., 2021).
The three outstanding centers of richness and endemism (micro-hotspots) of southern Solanaceae species are part of three different biodiversity hotspots at a continental scale: the northwest of Argentina is at the margin of the Tropical Andes hotspot; the northeast is at the margin of the Atlantic Forest hotspot, and the coastal Atacama is adjacent to the northern end of the Central Chile hotspot (Supplementary Material). Hotspots are defined as large areas of high species richness subjected to intense threats and landscape modification, as it gets clear by the expansion of the human footprint at a continental scale (Zalles et al., 2021). At least in Chile, protected areas have huge deficits in effective protection (Petit et al., 2018), and, certainly, much more efforts shall be done for effective protection at the landscape level, including target families, such as the Solanaceae. Results presented here remark the need for a geographically explicit strategy for the conservation of the family Solanaceae at subcontinental scale. In doing so, it is likely that other representatives of these centers of richness and endemism will benefit, promoting conservation and restoration at the landscape scale (Ianni and Geneletti, 2010; Malizia et al., 2012), and hopefully contributing also to reduce the generalized “plant blindness.”
AM-M and MP conceived the study and wrote the manuscript. VM-F compiled, cleaned, and updated the data base. VM-F and RA-V ran GIS analysis by means of ArcGis. VSD discussed the implications of biodiversity values in relation to protected areas. CG-O ran Biodiverse 3.1 analysis and analyzed results. All authors edited the manuscript and approved the submitted version.
AM-M has a grant from Agencia Nacional de Investigación y Desarrollo: Fondecyt-ANID 1180211. VSD was a CONICET Research Fellow.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Two reviewers provided very helpful comments on earlier drafts of the manuscript. We gratefully recognize the big efforts done by many botanists in the compilation of Documenta Florae Australis. RD invited us to this special issue and kindly provided photos. Mélica Muñoz-Schick and Alicia Marticorena shared information from SGO and CONC herbaria, respectively. Gloria Barboza, Juan José Cantero, and Miguel Dillon have been of constant help regarding taxonomic and distributional queries.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fpls.2022.854372/full#supplementary-material
Anguiano-Constante, M. A., Munguía-Lino, G., Ortiz, E., Villaseñor, J. L., and Rodríguez, A. (2018). Riqueza, distribución geográfica y conservación de Lycianthes serie Meizonodontae (Capsiceae, Solanaceae). Rev. Mex. Biodivers. 89, 516–529.
Arana, M. D., Larsen, C., and Ponce, M. M. (2016). Revisión y análisis panbiogeográfico de las Hymenophyllaceae de las Yungas meridionales de Argentina (Selva Tucumano-Boliviana). Rodriguésia 67, 55–75. doi: 10.1590/2175-7860201667105
Arana, M. D., Natale, E., Ferretti, N., Romano, G., Oggero, A., Martínez, G., et al. (2021). Esquema biogeográfico de la República Argentina. Opera Lilloa. 56:240.
Barboza, G. E. (Coord). (2013). “Solanaceae,” in Flora Argentina, 1st Edn, Vol. 13, eds F. O. Zuloaga, M. Belgrano, and A. M. Anton (San Isidro: IBODA-IMBIV, CONICET), 1–350. doi: 10.2307/j.ctt16vj2hs.4
Bernacki, F., Albornoz, P., Valoy, M., and Ordano, M. (2015). Anatomía de flor y fruto de Vassobia breviflora (Solanaceae) en el sur de las Yungas australes (Argentina). Phyton 84, 478–487.
Campanello, P. I., von Below, J., Hilgert, N. I., Cockle, K., Villagra, M., di Francescantonio, D., et al. (2019). ¿Es posible el uso sostenible del bosque en Misiones? Necesidades de manejo a diferentes escalas, investigación, intervenciones de alto impacto y más recursos económicos. Ecol. Aust. 29, 122–137. doi: 10.25260/EA.19.29.1.0.756
Crisp, M. D., Laffan, S. W., Linder, P., and Monro, A. (2001). Endemism in the Australian flora. J. Biogeogr. 28, 183–198. doi: 10.1046/j.1365-2699.2001.00524.x
Deanna, R., Wilf, P., and Gandolfo, M. A. (2020). New physaloid fruit-fossil species from early Eocene South America. Am. J. Bot. 107, 1–14. doi: 10.1002/ajb2.1565
Del Valle Elías, G., and Aagesen, L. (2019). Areas of endemism and recent speciation in the Southern Cone of South America, using Senecio (Asteraceae) as a proxy. Biol. J. Linn. Soc. 128, 70–82.
Dillon, M. O. (2005). “Solanaceae of the Lomas formations of Coastal Peru and Chile,” in A Festschrift for William G. D’Arcy: The Legacy of a Taxonomist, Monographs, eds V. Hollowell, T. Keating, W. Lewis, and T. Croa 131–155.
Dillon, M. O. (2016). “Nolana (Solanaceae),” in The Families and Genera of Vascular Plants. Asterales, et al Edn, Vol. 8, eds G. E. Barboza, A. T. Hunziker, and G. Bernardello (Berlin: Springer-Verlag), 343–344.
Documenta Florae Australis (2021). Available at http://www.darwin.edu.ar/iris/ (accessed July, 2021).
Dupin, J., Matzke, N. J., Särkinen, T., Knapp, S., Olmstead, R. G., Bohs, L., et al. (2017). Bayesian estimation of the global biogeographical history of the Solanaceae. J. Biogeogr. 44, 887–899. doi: 10.1111/jbi.12898
Fuentes-Castillo, T., Hernández, H. J., and Pliscoff, P. (2020). Hotspots and ecoregion vulnerability driven by climate change velocity in Southern South America. Reg. Environ. Chang. 20, 1–16. doi: 10.1007/s10113-020-01595-9
Gillson, L., Seymour, C. L., Slingsby, J. A., and Inouye, D. W. (2020). What are the grand challenges for plant conservation in the 21st Century? Font. Conserv. Sci. 1, 1–6. doi: 10.3389/fcosc.2020.600943
González-Orozco, A. C., Laffan, S. W., and Miller, J. T. (2011). Spatial distribution of species richness and endemism of the genus Acacia in Australia. Austr. J. Bot. 59, 600–608. doi: 10.1071/BT11112
Greppi, J. A., Hagiwara, J. C., and Stehmann, J. R. (2019). A new species of Petunia (Solanaceae) from Corrientes, Argentina. Phytotaxa 414, 289–295. doi: 10.11646/phytotaxa.414.6.3
Grosso, L. D. E., and Quintana, M. G. (2009). Can insect data be used to infer areas of endemism? An example from the Yungas of Argentina. Rev. Chil. Hist. Nat. 82, 507–522.
Hepp, J., and Dillon, M. O. (2018). A new endemic species of Nolana (Solanaceae-Nolaneae) from near Iquique, Chile. Arnaldoa 25, 323–338. doi: 10.22497/arnaldoa.252.25202
Hijmans, R. J., and Spooner, D. M. (2001). Geographic distribution of wild potato species. Am. J. Bot. 88, 2101–2112. doi: 10.2307/3558435
Hilgert, N. I., and Gil, G. E. (2006). Medicinal plants of the Argentine Yungas plants of the Las Yungas biosphere reserve, Northwest of Argentina, used in health care. Biodivers. Conserv. 15, 2565–2594. doi: 10.1007/s10531-005-3874-6
Ianni, E., and Geneletti, D. (2010). Applying the ecosystem approach to select priority areas for forest landscape restoration in the Yungas, Northwestern Argentina. Environ. Manage. 46, 748–760. doi: 10.1007/s00267-010-9553-8
Knapp, S. (2019). Are humans really blind to plants? Plants People Planet 1, 164–168. doi: 10.1002/ppp3.36
Knapp, S., Chiarini, F., Cantero, J. J., and Barboza, G. E. (2020). The Morelloid clade of Solanum L. (Solanaceae) in Argentina: nomenclatural changes, three new species and an updated key to all taxa. PhytoKeys 164, 33–66. doi: 10.3897/phytokeys.164.54504
Laffan, S. W., and Crisp, M. D. (2003). Assessing endemism at multiple spatial scales, with an example from the Australian vascular flora. J. Biogeogr. 30, 511–520. doi: 10.1046/j.1365-2699.2003.00875.x
Laffan, S. W., Lubarsky, E., and Rosauer, A. F. (2010). Biodiverse, a tool for the spatial analysis of biological and related diversity. Ecography 33, 643–647. doi: 10.1111/j.1600-0587.2010.06237.x
Lavandero, N., Chinga, J., Pinto, R., and Pérez, M. F. (2021). A new distinctive species of Schizanthus (Solanaceae) and the Reinstatement of Schizanthus fallax. Syst. Bot. 46, 456–469. doi: 10.1600/036364421X16231782047541
Luebert, F., and Pliscoff, P. (2017). Sinopsis Bioclimática y Vegetacional de Chile, 2nd Edn. Santiago: Editorial Universitaria.
Luebert, F., and Weigend, M. (2014). Phylogenetic insights into Andean plant diversification. Front. Ecol. Evol. 2:27. doi: 10.3389/fevo.2014.00027
Malizia, L., Pacheco, S., Blundo, C., and Brown, A. (2012). Caracterización altitudinal, uso y conservación de las Yungas Subtropicales de Argentina. Ecosistemas 1-2, 53–73.
Moat, J., Orellana-Garcia, A., Tovar, C., Arakaki, M., Arana, C., Cano, A., et al. (2021). Seeing through the clouds – mapping desert fog oasis ecosystems using 20 years of MODIS imagery over Peru and Chile. Int. J. Appl. Earth Obs. Geoinf. 103:102468. doi: 10.1016/j.jag.2021.102468
Morales-Fierro, V., Muñoz-Schick, M., and Moreira-Muñoz, A. (2020). Synopsis of Schizanthus Ruiz & Pav. (Solanaceae), a genus endemic to the southern Andes. PhytoKeys 154, 57–102. doi: 10.3897/phytokeys.154.49615
Moreira-Muñoz, A., and Muñoz-Schick, M. (2020). Rediscovery and taxonomic placement of Solanum polyphyllum Phil. (Solanaceae), a narrow endemic from the Chilean Atacama Desert. PhytoKeys 156, 47–54. doi: 10.3897/phytokeys.156.53703
Moreira-Muñoz, A., Scherson, R. A., Luebert, F., Román, M. J., Monge, M., Diazgranados, M., et al. (2020). Biogeography, phylogenetic relationships and morphological analyses of the South American genus Mutisia L.f. (Asteraceae) shows early connections of two disjunct biodiversity hotspots. Org. Divers. Evol. 20, 639–656. doi: 10.1007/s13127-020-00454-z
Muñoz-Schick, M., Pinto, R., Mesa, A., and Moreira-Muñoz, A. (2001). Fog oases during the El Niño Southern Oscillation 1997-1998, in the coastal hills south of Iquique, Tarapacá region, Chile. Rev. Chil. Hist. Nat. 74, 1–15.
Olmstead, R. G. (2013). Phylogeny and biogeography in Solanaceae, Verbenaceae and Bignoniaceae: a comparison of continental and intercontinental diversification patterns. Bot. J. Linn. Soc. 171, 80–102. doi: 10.1111/j.1095-8339.2012.01306.x
Oyarzabal, M., Clavijo, J., Oakley, L., Biganzoli, F., Tognetti, P., Barberis, I., et al. (2018). Unidades de vegetación de la Argentina. Ecol. Austr. 028, 40–63.
Palchetti, M. V., Cantero, J. J., and Barboza, G. E. (2020). Solanaceae diversity in South America and its distribution in Argentina. An. Acad. Bras. Cienc. 92, 1–17. doi: 10.1590/0001-3765202020190017
Palchetti, M. V., Cantero, J. J., Morales-Fierro, V., Barboza, G. E., and Moreira-Muñoz, A. (2021). Living in extreme environments: distribution of Lycium humile (Solanaceae), an endemic halophyte from the Altiplano-Puna region, South America. PhytoKeys 185, 1–15. doi: 10.3897/phytokeys.185.71377
Petit, I. J., Campoy, A. N., Hevia, M., Gaymer, C. F., and Squeo, F. A. (2018). Protected areas in Chile: are we managing them? Rev. Chil. Hist. Nat. 91, 1–8. doi: 10.1186/s40693-018-0071-z
Pizarro-Araya, J., Alfaro, F. M., Ojanguren-Affilastro, A., and Moreira-Muñoz, A. (2021). A Fine-Scale Hotspot at the Edge: epigean Arthropods from the Atacama Coast (Paposo-Taltal, Antofagasta Region, Chile). Insects 12, 1–15. doi: 10.3390/insects12100916
Ramirez-Villegas, J., Jarvis, A., and Touval, J. (2012). Analysis of threats to South American flora and its implications for conservation. J. Nat. Conserv. 20, 337–348. doi: 10.1016/j.jnc.2012.07.006
Rodríguez, A., Castro-Castro, A., Vargas-Amado, G., Vargas-Ponce, O., Zamora-Tavares, P., González-Gallegos, J., et al. (2018). Richness, geographic distribution patterns, and areas of endemism of selected angiosperm groups in Mexico. J. System. Evol. 56, 537–549. doi: 10.1111/jse.12457
Ruiz-Sánchez, E., Munguía-Lino, G., Vargas-Amado, G., and Rodríguez, A. (2020). Diversity, endemism and conservation status of native Mexican woody bamboos (Poaceae: Bambusoideae: Bambuseae). Bot. J. Linn. Soc. 192, 281–295.
Samuels, J. (2015). Biodiversity of food species of the solanaceae family: preliminary taxonomic inventory of subfamily solanoideae. Resources 4, 277–322. doi: 10.3390/resources4020277
Sanderson, E. W., Jaiteh, M., Levy, M. A., Redford, K. H., Wannebo, A. V., and Woolmer, G. (2002). The human footprint and the last of the wild. Bioscience. 52, 891–904. doi: 10.1641/0006-3568(2002)052[0891:thfatl]2.0.co;2
Santilli, L., Pérez, M. F., de Schrevel, C., Dandois, P., Mondaca, H., and Lavandero, N. (2021). Nicotiana rupicola sp. nov and Nicotiana knightiana (Paniculatae, Solanaceae), a new endemic and a new record for the flora of Chile. ARPHA Preprints 1:e73387. doi: 10.3897/arphapreprints.e73387
Sosa, V., and de Nova, J. A. (2012). Endemic angiosperm lineages in Mexico: hotspots for conservation. Acta Bot. Mex. 100, 293–315.
Torres, V. M., and González-Reyes, A. X. (2017). Diversidad taxonómica y funcional de arañas (Araneae) epigeas en bosques nativos de las Yungas (Salta. Argentina). Caldasia 39, 326–344.
Ulloa Ulloa, C., Acevedo-Rodríguez, P., Beck, S., Belgrano, M. J., Bernal, R., Berry, P. E., et al. (2017). An integrated assessment of the vascular plant species of the Americas. Science 358, 1614–1617. doi: 10.1126/science.aao0398
Wandersee, J., and Schussler, E. (2001). Toward a theory of plant blindness. Plant Sci. Bull. 47, 2–8.
Zalles, V., Hansen, M. C., Potapov, P. V., Parker, D., Stehman, S. V., Pickens, A. H., et al. (2021). Rapid expansion of human impact on natural land in South America since 1985. Sci. Adv. 7, 1–12. doi: 10.1126/sciadv.abg1620
Keywords: micro-hotspots, conservation biogeography, plant blindness, protected areas, biodiverse
Citation: Moreira-Muñoz A, Palchetti MV, Morales-Fierro V, Duval VS, Allesch-Villalobos R and González-Orozco CE (2022) Diversity and Conservation Gap Analysis of the Solanaceae of Southern South America. Front. Plant Sci. 13:854372. doi: 10.3389/fpls.2022.854372
Received: 13 January 2022; Accepted: 11 April 2022;
Published: 17 May 2022.
Edited by:
Rocio Deanna, University of Colorado, Boulder, United StatesReviewed by:
Eduardo Ruiz-Sanchez, University of Guadalajara, MexicoCopyright © 2022 Moreira-Muñoz, Palchetti, Morales-Fierro, Duval, Allesch-Villalobos and González-Orozco. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Andrés Moreira-Muñoz, YW5kcmVzLm1vcmVpcmFAcHVjdi5jbA==
†ORCID: Andrés Moreira-Muñoz, orcid.org/0000-0002-9136-1391; Carlos E. González-Orozco, orcid.org/0000-0002-9268-5224
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.