 Mariam Kourani
Mariam Kourani Fady Mohareb
Fady Mohareb Faisal I. Rezwan1†
Faisal I. Rezwan1† Maria Anastasiadi
Maria Anastasiadi John P. Hammond
John P. Hammond
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
REVIEW article
Front. Plant Sci. , 05 April 2022
Sec. Plant Abiotic Stress
Volume 13 - 2022 | https://doi.org/10.3389/fpls.2022.832147
This article is part of the Research Topic Insights in Plant Abiotic Stress: 2021 View all 14 articles
Given the current rise in global temperatures, heat stress has become a major abiotic challenge affecting the growth and development of various crops and reducing their productivity. Brassica napus, the second largest source of vegetable oil worldwide, experiences a drastic reduction in seed yield and quality in response to heat. This review outlines the latest research that explores the genetic and physiological impact of heat stress on different developmental stages of B. napus with a special attention to the reproductive stages of floral progression, organogenesis, and post flowering. Several studies have shown that extreme temperature fluctuations during these crucial periods have detrimental effects on the plant and often leading to impaired growth and reduced seed production. The underlying mechanisms of heat stress adaptations and associated key regulatory genes are discussed. Furthermore, an overview and the implications of the polyploidy nature of B. napus and the regulatory role of alternative splicing in forming a priming-induced heat-stress memory are presented. New insights into the dynamics of epigenetic modifications during heat stress are discussed. Interestingly, while such studies are scarce in B. napus, opposite trends in expression of key genetic and epigenetic components have been identified in different species and in cultivars within the same species under various abiotic stresses, suggesting a complex role of these genes and their regulation in heat stress tolerance mechanisms. Additionally, omics-based studies are discussed with emphasis on the transcriptome, proteome and metabolome of B. napus, to gain a systems level understanding of how heat stress alters its yield and quality traits. The combination of omics approaches has revealed crucial interactions and regulatory networks taking part in the complex machinery of heat stress tolerance. We identify key knowledge gaps regarding the impact of heat stress on B. napus during its yield determining reproductive stages, where in-depth analysis of this subject is still needed. A deeper knowledge of heat stress response components and mechanisms in tissue specific models would serve as a stepping-stone to gaining insights into the regulation of thermotolerance that takes place in this important crop species and support future breeding of heat tolerant crops.
Over the last century, global warming has become a major environmental challenge triggered anthropogenic emissions of greenhouse gases driving the continuous rise in ambient temperature (Pachauri et al., 2014; Zandalinas et al., 2021). Furthermore, the climate projections for 2100 suggest an increase in the number of drought affected areas by 50% (Battisti and Naylor, 2009). The combination of these environmental changes has the potential to destabilize plant growth and development and could result in drastic yield reductions (Lamaoui et al., 2018). Although the impact of elevated CO2 on crops is still under debate with potential benefits on photosynthesis, Frenck et al. (2011) showed that rising CO2 could not compensate for the negative impact on yield caused by high temperature stress.
On a global scale, environmental climate change is aggravating different abiotic stresses, including heat and water deficit across many biomes. Plants, being sessile organisms, cannot escape their environment and must endure a wide variety of stresses (Saraswat et al., 2017). Although plant stress responses are dynamic and encompass a complex cross-talk between different molecular and cellular interactions to establish physiological and morphological adaptations (Krasensky and Jonak, 2012), temperature exceeding the threshold of adaptation will substantially influence plant survivability (Hasanuzzaman et al., 2014). In particular, this affects cellular functioning, which in turn, hinders plant growth and development and weakens resilience to biotic stresses (Ahmed et al., 2020).
Several studies have demonstrated the effect of heat stress, often in combination with drought or other stresses, on crop production and its impact on plant metabolism, reproduction, and physiology. These studies have reported an extensive reduction in yield of various crops (Frenck et al., 2011; Hasanuzzaman et al., 2014). Lesk et al. (2016) studied the influence of extreme weather disasters on global crop production and found that, between 1964 and 2007, 1.82 and 1.19 billion metric tons of cereals were lost due to droughts and extreme heat respectively. Using four different process-based crop models, Zhao et al. (2017) demonstrated that for each 1°C rise in global mean temperature would cause a reduction in the annual production of wheat, rice, maize and soybean. In addition to its impact on yield, the increase in climate temperature has also been found to negatively impact the quality of oil in oilseed crops such as soybean, oilseed rape and sunflower, where a negative correlation between temperature and the proportion of essential fatty acids such as C18:2 and C18:3 was reported (Werteker et al., 2010; Namazkar et al., 2016). Thus, the effect of climate change is inevitably impeding vital components of human diet and nutrition.
Oilseed rape (Brassica napus L.) is the second-largest source of vegetable oil after soybean worldwide (United States Department of Agriculture [USDA], 2021). Given its eco-physiological adaptations to temperate climates, it is currently considered the predominant oil crop in Europe (Namazkar et al., 2016; Sun et al., 2017). B. napus belongs to the genus Brassica from the mustard Family Brassicaceae (Liu G. et al., 2018), where selection has given rise to several agronomically important lines such as oilseed rape, rutabaga, fodder rape, and kale morphotypes (Chalhoub et al., 2014). These are cultivated globally for vegetables, good quality oil, fodder, and bio-diesel industries (Chalhoub et al., 2014; Pokharel et al., 2020). As a cool-season crop, Brassica crops can be extremely sensitive to increasing temperatures, which significantly affect its production (Yu et al., 2014; Ahmed et al., 2020). Though studies in different Brassica species have found negative relationships between heat stress and seed yield and quality, the underlying mechanisms are still not well understood and require further investigation (Yu et al., 2014). Thus, to increase resilience against adverse environmental growth conditions and to maintain a stable oil supply to the global population, understanding and improving heat tolerance in B. napus is critical.
The acclimation mechanisms of plants to heat stress are complex and mediated by the activation of different physiological, cellular, and metabolic processes depending on their developmental stage (Zandalinas et al., 2018). These mechanisms normally encompass a combination of stress avoidance and tolerance (thermotolerance) approaches that vary with genotypes (Chaves et al., 2002). At a molecular level, the abundance in transcriptomic data facilitates the analysis of the expression of genes that play regulatory roles in different stages and tissues of plant growth and development under various stress conditions. However, the challenge lies in linking this information with the relevant biological processes taking place during stress (Chaves et al., 2009). So, uncovering the controlling mechanisms for heat tolerance is therefore imperative for successful breeding programs and future production sustainability.
In this review, we outline the latest research exploring heat stress responses in B. napus, focusing on the physiological impact at different developmental stages along with their underlying mechanisms. We further explore new insights into the dynamics of epigenetic changes and systems biology approaches of this plant species under heat stress. However, due to the scarcity of information specific to B. napus, many of the reviewed findings stem from research on the model organism Arabidopsis thaliana and other Brassica species.
As thermotolerance is regulated both developmentally and tissue-specifically, addressing the impact of high temperature on different developmental stages is imperative to produce heat-tolerant cultivars capable of coping with heat stress (Figure 1). Different optimum temperatures also differ considerably among species or genotypes (Angadi et al., 2000; Baux et al., 2013). In general, heat stress takes place when soil and ambient temperature is beyond a plants’ growth and photosynthetic temperature range such that, permanent harm occurs. A plants performance under heat stress is also influenced by the concomitant presence of other types of stress, such as drought (Frenck et al., 2011).
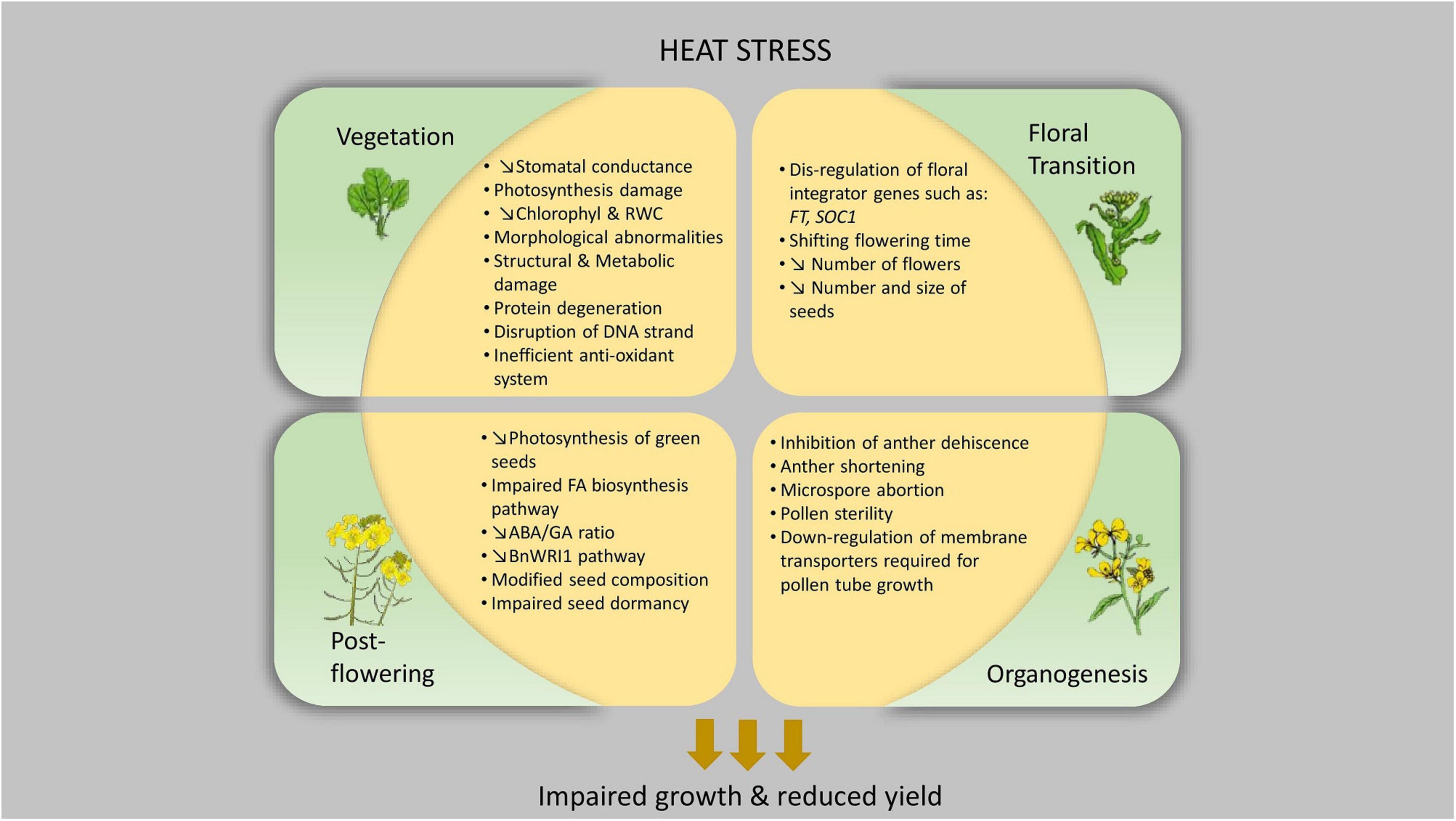
Figure 1. Physiological impact of heat stress at different developmental stages of Brassica napus.
Under heatwave conditions during early vegetative growth, well-watered B. napus experiences an increase in saturating light (Asat), transpiration (E) and in the ratio of intercellular to ambient CO2 concentration (Ci/Ca) (Dikšaitytė et al., 2019). The increase in ambient temperature enhances photosynthesis performance and increases above-ground growth. However, in the presence of drought, the combined stress conditions lead to severely impaired photosynthesis, which is accompanied by a profound decrease in stomatal conductance (Dikšaitytė et al., 2019). To avoid overheating, plants open their stomata and increase transpiration, thus preventing leaf damage. In contrast, drought triggers stomatal closure as a conservative mechanism to protect plants from excessive water loss (Elferjani and Soolanayakanahally, 2018). During the flowering stage, B. napus responds to combined heat and drought stress by a rapid closure of their stomata. This avoids further loss of water through transpiration at the expense of evaporative cooling. Consequently, this results in an excessive increase in leaf temperature and more photosynthetic damage (Elferjani and Soolanayakanahally, 2018). This s paradox requires plants to maintain a balance between avoiding overheating and minimizing water loss (Zandalinas et al., 2018).
Additionally, impairment of chlorophyll biosynthesis and disruption of the biochemical reactions of photosystems were exhibited in 10-day-old B. napus seedlings subjected to 38°C (Hasanuzzaman et al., 2014). This was manifested through a significant reduction in chlorophyll and leaf relative water content as well as through an inefficient antioxidant defense system (Hasanuzzaman et al., 2014). During heat stress, the physiological changes that occur at the level of photosynthesis and transpiration alter the synthesis and usage of assimilates, which in turn translate into morphological abnormalities in the floral organs (Echer et al., 2014).
At the molecular level, heat stress can either induce structural damage through altering membrane proteins and stability or metabolic damage through disrupting enzymatic activities and production of toxic metabolites (Mittler et al., 2012). In A. thaliana, heat stress induced Reactive Oxygen Species (ROS) production and subsequently a higher lipid peroxidation and membrane injury (Rocco et al., 2013). Moreover, protein degradation, pigment bleaching and disruption of DNA strands are also consequences of heat stress (Mathur and Jajoo, 2014). Therefore, heat stress during vegetative and seedling growth works as an induction point for a cascade of events that can impact on reproductive organs later in development and consequently lead to reduced productivity.
Since the developmental progression of plants from seed to seed is bound to the success of sexual reproduction, susceptibility to heat stress at this stage can have deleterious implications on yield potential. Plants exhibit higher sensitivity to extreme temperature fluctuations during the reproductive stage of development than during their vegetative growth, often leading to impaired growth and reduced seed production (Zhang et al., 2017; Lamaoui et al., 2018), especially during meiosis and fertilization (Zhang et al., 2017), with the male reproductive organs being the most vulnerable (Giorno et al., 2013). This sensitivity fluctuates within different reproductive processes, such as: pollen development, pollen tube growth, pollen/pistil interactions, fertilization and embryo development (Erickson and Markhart, 2002; Young et al., 2004; Baron et al., 2012; Giorno et al., 2013; Zhang et al., 2017). The after-effects of heat stress during reproduction influence plant physiology in three main areas: reduction in the number of flowers at pre-anthesis, reduction in flower fertility, and poor pods and seeds development (Morrison and Stewart, 2002). Nevertheless, this has been poorly studied and specific responses at each of these processes remain elusive. Similarly, in A. thaliana only a limited number of reports addressed the effect of heat stress on its reproductive development (Antoun and Ouellet, 2013; Guan et al., 2014; Bac-Molenaar et al., 2015; Chao et al., 2017). Under short-term heat exposure, a genome-wide association (GWA) mapping approach in A. thaliana revealed different QTLs associated with heat response at pre- and post- anthesis, suggesting that these genetic regulators are developmental stage-specific (Bac-Molenaar et al., 2015). Moreover, when A. thaliana was exposed to 37°C for 24 h, it exhibited a severe growth impairment of stamen filaments which prevented pollination (Baron et al., 2012). At the structural level, heat stress caused abnormalities in the reproductive organs including vacuolation, collapse within microspores and abnormally shaped pollen sacs, incapable of accumulating carbohydrates (Baron et al., 2012). In spring wheat, pre-anthesis associated events such as gametogenesis and gamete development appeared highly sensitive to high-temperature treatment as heat stress resulted in reduced floret fertility and consequently a significant reduction in seed number and weight (Bheemanahalli et al., 2019).
In agriculture, flowering is a key prerequisite stage for the transition from the vegetative phase to reproduction. It is regulated by the expression of a set of floral integrator genes such as FLOWERING LOCUS T (FT), to activate the meristem identity genes LEAFY (LFY), APETALA1 (AP1), SEPALLATA3 (SEP3) and FRUITFULL (FUL), causing the transition from vegetative to a floral meristem (Blümel et al., 2015). Under different stress conditions, plants utilize different mechanisms to alter flowering time (FTi) as a strategy to ensure the success of reproduction (Figure 2; Simpson, 2004; Ko et al., 2010; Blümel et al., 2015; Kazan and Lyons, 2016). However, these mechanisms are known to impact crop productivity (Kazan and Lyons, 2016).
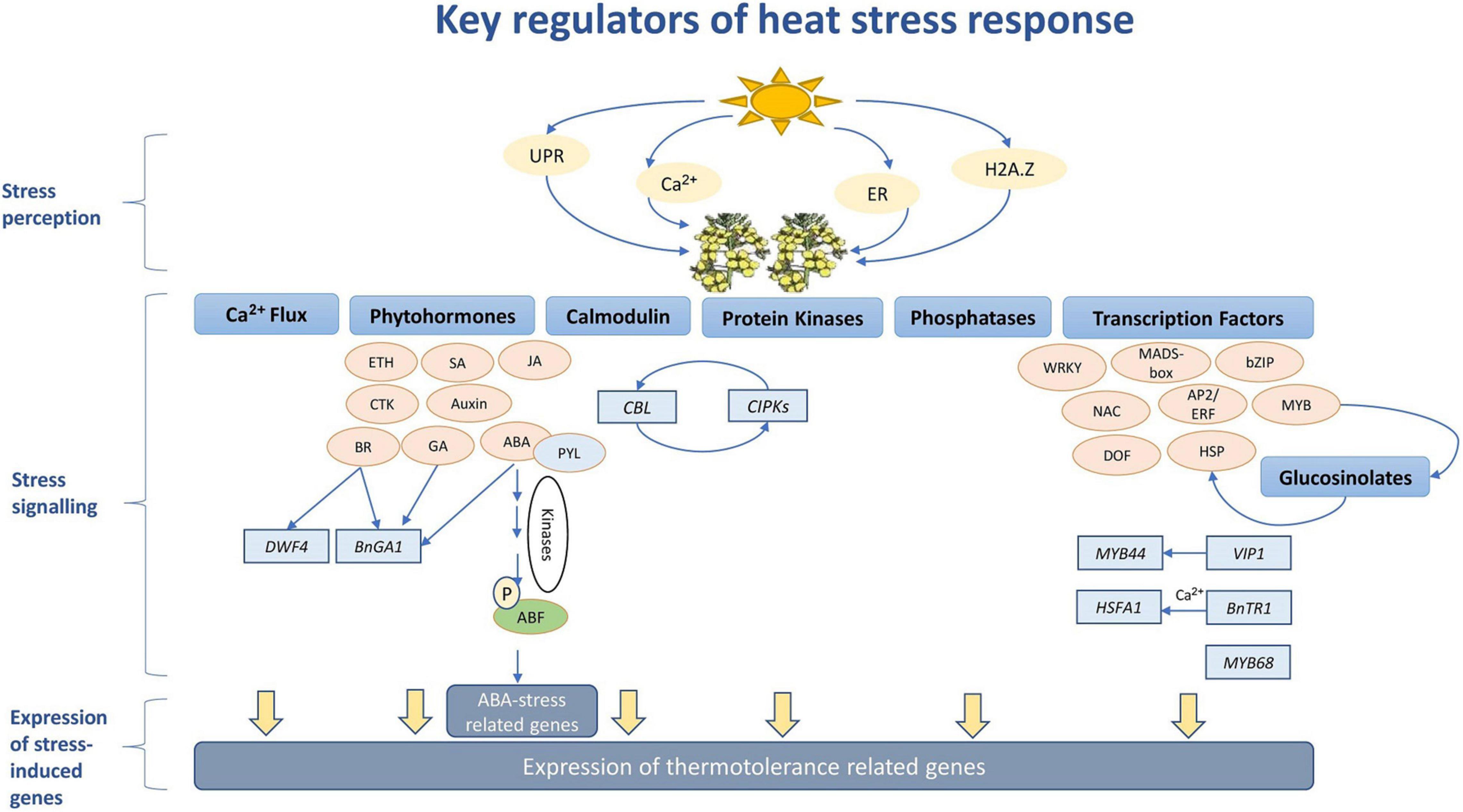
Figure 2. Major flowering time regulators in Brassica napus.
During flowering, energy demand spikes due to different processes, such as: formation of flowers, nitrogen use efficiency and reduced photosynthesis (Schiessl, 2020). Such crucial physiological period has been found sensitive to various biotic and abiotic stress and particularly to heat, which has a pronounced effect on the timing of flowering, depending on its duration, frequency, and intensity (Lohani et al., 2020b). In a split-plot experiment, different genotypes of B. napus were exposed to high-temperature stress (31°C/14°C - day/night) during flowering across two sowing periods (Koscielny et al., 2018). Results showed a reduction in yield by 55% in the heat treatment during winter and by 41% during the subsequent autumn experimental run (Koscielny et al., 2018). At different developmental stages, severe damage to reproductive organs of three Brassica species (B. juncea L., B. napus L., B. rapa L.) occurred when exposed to 35°C/15°C for 7 days resulting in yield reductions (Angadi et al., 2000). It was noted that yield was affected by heat stress at flowering more than at pod development, indicating a vital developmental threshold passed by pods rendering them more heat tolerant. Interestingly, among the three studied Brassica species, only B. napus could not recover completely after heat stress was removed, producing abnormal, plump, and short pods (Angadi et al., 2000). A reduction in seed yield was observed following 29.5°C temperature treatment during flowering, primarily due to the reduction in the number of flowers and in the number and size of seeds produced per flower (Morrison and Stewart, 2002).
During flower opening in rice air temperature is crucial in determining the fate of reproduction (Bheemanahalli et al., 2017) and the resulting earlier or delayed flowering can negatively impact crop growth and quality. In a recent study carried out by Pokharel et al. (2020), high night temperature stress-induced a shift in B. napus peak flower opening time into earlier and cooler morning hours, suggesting an adaptation toward the heat escape response, yet this was accompanied by a significant reduction in yield. Whilst B. napus is not strictly dependent on pollinators, it has been shown that pollinators enhance seed yield and quality (Zou et al., 2017). So, shifting flowering time drives the flower opening out of sync with pollinator activity, and could potentially lead to reductions in yield.
Although FTi regulator genes and their associated networks have been well characterized in A. thaliana, the conservation of these genes in other Brassica species indicates that their regulation takes place in the same way as in this model system. However, environmental adaptation has led to the evolution of new FTi regulator genes with no functional equivalence in A. thaliana (Blümel et al., 2015). Moreover, some FTi gene copy numbers were found to either change or lose functionality over evolutionary periods resulting in sub-functionalization (Schiessl, 2020). As a result, it is difficult to functionally characterize crop FTi regulators and pathways based on inferences from this model species. This was manifested in a study by Del Olmo et al. (2019) when B. rapa delayed flowering was associated with reduced BraA.FT.a mRNA levels upon high-temperature treatment. High levels of H2A.Z were also found to occupy the BraA.FT.a locus, which in turn affected chromatin conformation and hindered its accessibility. The same conserved chromatin regulatory mechanism in A. thaliana showed an opposite trend and resulted in accelerated flowering (Del Olmo et al., 2019). This brings attention to the fact that thermosensory pathways perform differently in different crops to alter FTi regulators.
Considering the scarcity of studies addressing the impact of increased ambient temperature on FTi-related signaling pathways in B. napus, the continuous rise in global temperature will need to drive future research to explore these pathways and the underlying regulators under heat stress. Such studies will help identify key regulators, direct genetic modifications to candidate genes involved in these pathways and to provide more reliable data for breeding programmes to deliver heat stress tolerant cultivars.
The next stage after floral transition is the differentiation of floral organs or organogenesis. In B. napus as in other flowering plants, gametogenesis and reproduction are highly vulnerable to high-temperature stress (Angadi et al., 2000; Young et al., 2004). From inhibition of anther dehiscence and anther shortening to reducing pollen germinability and viability (Rahaman et al., 2018), the impact of heat stress on different reproductive organs has been reported in various species, such as: wheat, soybean, Brassicas and tomato (Giorno et al., 2013; Draeger and Moore, 2017; Djanaguiraman et al., 2019). Moreover, heat stress during microsporogenesis leads either to microspore abortion or pollen sterility (Lohani et al., 2020b). The variety of affected species suggests the existence of common processes involved in heat stress-related infertility (Kim et al., 2001; Rang et al., 2011; Sage et al., 2015) and yield loss (Morrison and Stewart, 2002; Frenck et al., 2011; Zhao et al., 2017). However, in B. napus, such processes are still not very well studied.
In Arabidopsis thaliana, during anther development, heat stress causes male sterility in a stage-specific manner (Kim et al., 2001). Disruption of male meiotic processes was associated with abnormal pollen development and morphological analysis with a failure in the separation of pollen mother cells and a subsequent microspore differentiation (Kim et al., 2001). Heat stress also caused a dramatic down-regulation in membrane transporters required for pollen tube growth such as K+ and carbohydrate co-transporters, which would adversely impact pollination (Poidevin et al., 2020). To discover the effect of heat stress after flowering initiation in B. napus, Young et al. (2004) reported that following heat exposure at 35°C, both micro and megagametophytes were found thermosensitive and resulted in a significant reduction in pollen viability and germinability as well as in seed development. An increase in temperature was also reported to induce sterility and lead to a decrease in fertility rate, pod-setting ratio and seed number per pod (Zhang et al., 2012). Similarly, in a comparative transcriptomic analysis, Tang et al. (2019) found that genes encoding heat shock proteins, skeleton proteins, GTPase and calmodulin were potentially involved in the mechanism of thermosensitive genic male sterility (TGMS) under high temperature inducement. Moreover, genes encoding plant hormone signaling pathways (auxin, gibberellins, jasmonic acid, abscisic acid, brassinosteroid signalings), along with some well-known transcription factors (MADS, NFY, HSF, MYB/C, and WRKY) were also found involved in the regulation of TGMS in the flowers grown under 25°C (Tang et al., 2019).
Although many studies have reported that male reproductive organs are more sensitive to heat stress than female reproductive organs, changes in the female reproductive organs following heat stress have also been documented (Hedhly, 2011). Additionally, according to the identity of the cell, the female gametophyte possesses a unique and differentially mediated response to heat stress (Ambastha and Leshem, 2020). These findings suggest that temperature plays a vital role in affecting the fertility of pollen and the morphology of the flower reproductive organs.
The thermotolerance of seed setting rate and filling, are also crucial in determining grain yield and composition. During later reproductive growth, the biosynthesis and deposition of oils and proteins in seeds take place at the seed filling stage. Hence, it is imperative to strictly regulate this stage for the generation of high-quality and balanced oil (Brunel-Muguet et al., 2015).
Photosynthesis in B. napus green seeds and pods induces the formation of the necessary components of the fatty acid biosynthesis pathway (Huang et al., 2019). This pathway was found to be regulated by the transcription factor BnWRI1 (Wu et al., 2014). In a recent study, heat stress suppressed oil deposition in developing seeds and impaired fatty acid biosynthesis where the inhibition of the de novo fatty acid biosynthesis pathway was associated with the inhibition of photosynthesis and downregulation of many genes in the BnWRI1 pathway (Huang et al., 2019). These findings have highlighted the crucial function of BnWRI1 as a key regulator within the intricate regulatory systems of heat stress and biosynthesis of storage compounds. Brunel-Muguet et al. (2015) also showed that high temperature during seed development modified seed composition and impaired the acquisition of seed dormancy. This was accompanied by a decrease in abscisic acid/gibberellic acid (ABA/GA) ratio, which plays a role in controlling seed composition and storage capacity (Brunel-Muguet et al., 2015). Similarly, the total lipids and all the fatty acids studied were significantly lower in B. napus seed oil when temperature was raised from 19°C to 24°C, resulting in imbalanced proportions of essential fatty acids (Namazkar et al., 2016).
Interestingly, not only high day temperature was found to impact seed quality and yield, but also high night temperature was reported to have detrimental effect on yield and seed fatty acid composition in B. napus. During flowering and silique-filling stages, Pokharel et al. (2020) reported that high night temperature caused significant alterations in the reproductive organs and led to a substantial reduction in seed yield and number of siliques, which was further explained by the thermal impairment of photosynthetic assimilation enzymes. Likewise, exposure of susceptible B. napus cultivars to high night temperature stress between flowering and seed filling stages also induced a profound reduction in total fatty acids and in fatty acid composition in seeds (Zhou et al., 2018). It is worth noting that the magnitude of heat stress and the cardinal temperature unequivocally contribute to the degree of impact on each crop species (Mittler et al., 2012).
In summary, high-temperature stress induces adverse effects on different processes and organs during B. napus reproduction. From floral transition to seed filling, such environmental threats will eventually translate into unfavorable agronomic and quality traits and ultimately challenge the production of seed oil with strong nutritional values. Knowledge about heat stress during these stages is limited and more insight on this topic will help scientists and breeders to design strategies to protect this important crop species from the impact of the stress potentially caused by climate change presently and in the future.
During postharvest storage, high-temperature stress has also been found to impact the safety and nutritional qualities of stored seeds (Krasucki et al., 2002). Brassica seeds are rich in bioactive compounds that exhibit antioxidant action and can play a vital role in reducing blood cholesterol levels (Ratnayake and Daun, 2004). In B. napus seeds, the concentrations of phytosterols can be twice that of sunflower and soybean oils (Vlahakis and Hazebroek, 2000), and the content of phenolic compounds can be 10-fold greater than in seeds of other crops (Siger et al., 2018). The abundance of such important biologically active compounds is sensitive to storage conditions. High temperature and moisture content were associated with the deterioration of seeds and degradation of its biochemical contents (Gawrysiak-Witulska et al., 2012). After 18 days of storage at 25°C and 30°C, the amount of phytosterols decreased significantly, reaching 24 and 58% less in B. napus seeds with a moisture content of 15.5% (Gawrysiak-Witulska et al., 2012). Under similar storage conditions, tocopherols dropped by 14.4% while plastochromanol-8 loss ranged from 4 to 24% (Gawrysiak-Witulska et al., 2011). These natural antioxidants play a crucial role in determining lipid stability and inhibition of auto-oxidation in stored seeds (Olejnik et al., 1997). As a result, the degradation of such compounds not only alters the quality of oil but can also lead to the formation of oxidized derivatives with cytotoxic properties (Rudzińska et al., 2009). To investigate the effect of inappropriate storage conditions (13.5% moisture content and 25°C temperature) on seed quality in B. napus, Siger et al. (2018) reported an intense growth of fungi along with a decrease in sinapic acid derivatives, the main phenolic compounds in B. napus (Siger et al., 2018). The development of fungal growth not only reduces the time of safe storage but also causes a risk of seed deterioration and mycotoxin contamination posing a serious threat to human and animal health (Siger et al., 2018). Furthermore, fungal growth and respiration could also result in the production of additional heat and moisture in the seeds (Mills, 1989), which further exacerbates the problem of temperature stress and microbial infestation.
Plants respond to environmental adversities, such as changes in ambient temperature through a cascade of signaling pathways, resulting in cellular readjustments at the transcriptome, epigenome, proteome and metabolome levels. Eventually, these adjustments provide the required balance through multiple adaptive strategies to ensure survival under the new conditions. Such intricate machinery involves an interconnected crosstalk between different components of the cell, initiated by stress perception, carried out through stress signaling and ultimately end with the expression of a specific set of downstream stress-induced genes that result in a phenotypic response to the stress.
Different sensors are involved in the initial stress perception (Lamaoui et al., 2018; Baillo et al., 2019), and these include plasma membrane mediated calcium flux, endoplasmic reticulum (ER) and cytosolic unfolded protein response (UPR) sensors, and decreased histone occupancy in the nuclei (Mittler et al., 2012). The generated signals are then integrated through a network of multiple events that includes calcium fluxes, calmodulin, phosphatases, transcriptional regulators, hormones, and protein kinases such as CDPKs, MAPKs (Figure 3; Verma et al., 2016; Lohani et al., 2020a). Hence, the complexity of heat response lies in the broad range of pathways involved, which appear to be tissue and developmental stage specific (Zhang et al., 2017).
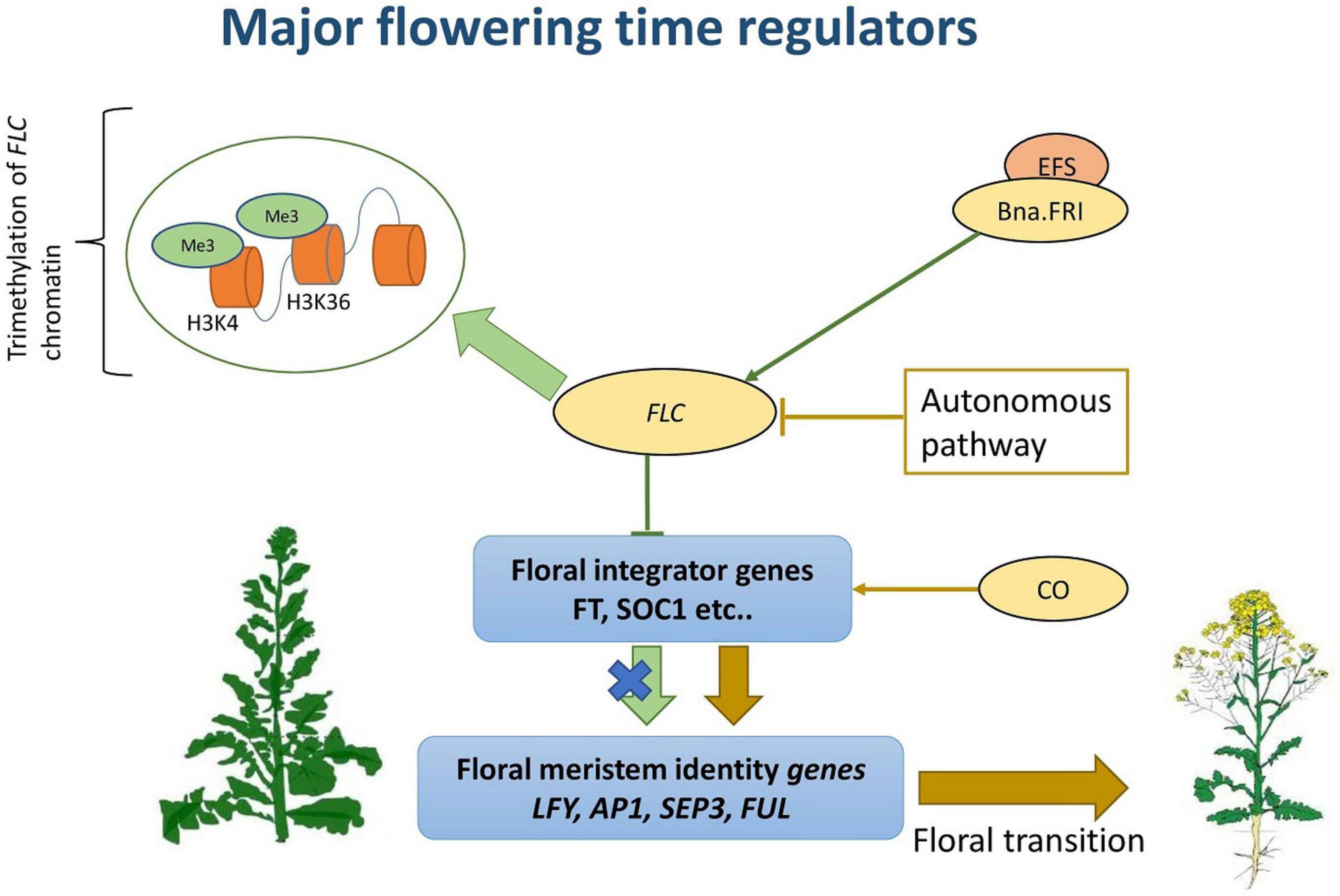
Figure 3. Regulatory and signaling events involved in plant heat stress response.
One of the principal regulators of abiotic stress responses are phytohormones, which encompass several compounds, such as: abscisic acid (ABA), brassinosteroids (BR), salicylic acid, jasmonates (JA), ethylene, auxins, cytokinins, and gibberellins (Verma et al., 2016). These phytohormones have essential roles in protecting plants against environmental stress (Lohani et al., 2020a). The intricate signaling role of these compounds is carried out either directly or through crosstalk between interacting arrays of various secondary messengers (Verma et al., 2016). During biotic and abiotic stress responses, ABA plays a powerful role in mediating plant adaptation to stress (Baron et al., 2012). In A. thaliana, temperature stress induced the differential expression of several ABA biosynthesis genes (ABA1, ABA2, ABA4, AAO3, NCED3) in an organ-specific manner (Baron et al., 2012; Table 1). Moreover, in B. napus, Di et al. (2018) identified 14 differentially expressed pyrabactin resistance 1-like (PYL) genes, which play core roles in ABA signaling networks, at different time points of heat treatment. BnPYR1-3, BnPYL1-2, BnPYL3-1, BnPYL4-2, BnPYL5-4, BnPYL6-1, BnPYL7-2, and BnPYL 9-2 were up-regulated while BnPYR1-4, BnPYL2-2, BnPYL4-6, BnPYL8-5, BnPYL8-6, BnPYL9-1 were down regulated (Table 1). Also, some BnPYL genes such as BnPYR1-3, BnPYL1-2, and BnPYL7-2 had similar expression patterns under drought, salinity, and heat treatments suggesting that these genes might be important candidates for improving tolerance to abiotic stress (Di et al., 2018). In addition to its role during abiotic stress response, tissue ABA concentrations were also found to influence seed components during grain filling (Chandrasekaran et al., 2014). Under high-temperature conditions, changes in ABA content in seeds were involved in the modification of downstream expression of genes associated with the biosynthesis of seed storage compounds in castor beans (Chandrasekaran et al., 2014). Thus, the quality determining characteristics of seeds such as seed germination, vigor and storability can be negatively affected by the impact of heat stress through inducing changes in the concentration of important hormonal signaling molecules (Gao et al., 2010).
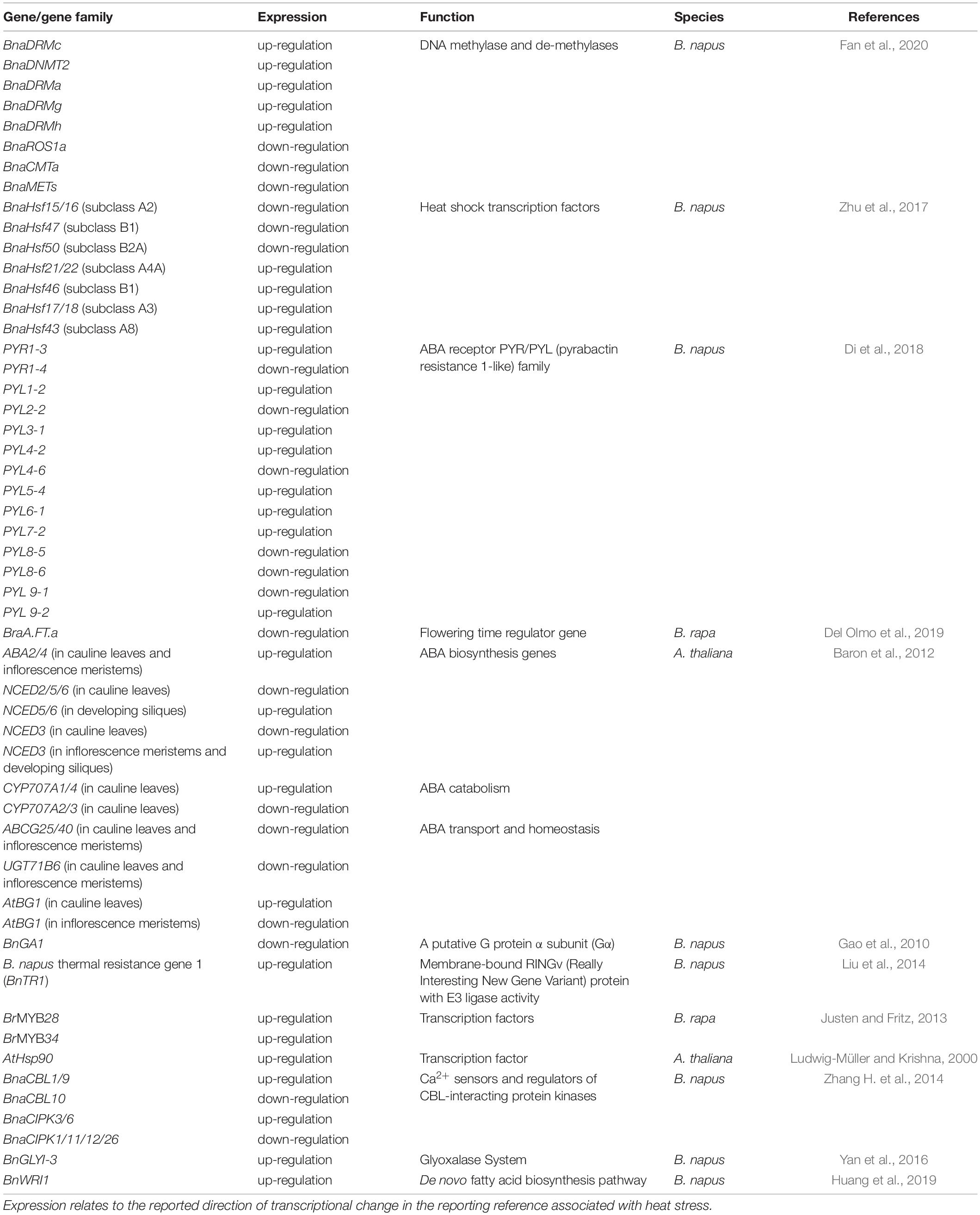
Table 1. Heat stress responsive genes identified in Brassica napus and related species.
Although many studies addressed the role of ABA as a major stress-responsive hormone, the role of other stress hormones has also been studied in response to various abiotic stress factors. For example, high concentrations of ABA and BR and low concentrations of gibberellic acid 3 (GA3) significantly induced BnGA1 expression, a gene that encodes a putative G protein α subunit (Gα) in B. napus (Gao et al., 2010). BnGA1 was initially found strongly expressed at the bolting and fruiting stages. However, different abiotic stresses prompted different expression patterns with an apparent low expression in response to heat. This reduction in transcription could be attributed to one of the physiological events in decelerating cellular processes under temperature stress (Gao et al., 2010). BR were also found to enhance seedling tolerance to drought and cold stress and counterbalance the inhibitory impact of salt stress on seed germination in A. thaliana and B. napus (Kagale et al., 2007). Transgenic B. napus overexpressing the Arabidopsis BR biosynthetic gene DWF4 exhibited increased seed yield and enhanced stress response to heat and drought (Sahni et al., 2016). At higher concentrations, stress hormones such as JA inhibit growth as a strategy to mobilize the metabolic reservoir between growth and stress response processes (Heinrich et al., 2013). The overexpression of BR biosynthetic gene DWF4 induced a downregulation of several JA biosynthesis and signaling genes, suggesting a mechanism played by BR to balance the inhibitory effect of JA on growth (Sahni et al., 2016). Furthermore, analysis of global transcription profiles after heat stress showed diverse expression patterns between different hormone signaling pathways. While genes encoding proteins involved in the biosynthesis of ethylene and GAs were all reduced, genes encoding auxin synthesis, binding and transport were all induced in seeds (Yu et al., 2014). These studies suggest a complex interaction among different phytohormones, through switching the activities of certain genes as needed depending on the stress situation. Thereby, functioning as key regulators to achieve a state of homeostasis and maintain a balance between stress tolerance and plant productivity.
Different TF families have been identified in plants such as AP2/ERF, MYB, HSP, bZIP, NAC, DOF, MADS-box, etc. (Ghorbani et al., 2020; Table 1). In a recent study, Wang et al. (2018) identified 2,167 TFs belonging to five different families in B. napus with 7.6% identified as novel and unique to B. napus. Interestingly, among the DEG identified in response to different abiotic stresses, 70% were responsive to heat after 12 h of treatment and these include: 62 BnAP2/EREBPs, 32 BnbZIPs, 61 BnMYBs, 40 BnNACs, and 27 BnWRKYs (Wang et al., 2018).
During plant heat stress, the heat shock response is a conserved response during which heat shock transcription factors (HSF) regulate heat shock proteins (HSP) through recognizing and binding to conserved palindromic motifs in the promoter region of heat-stress responsive genes (Scharf et al., 2012). As a result, HSP bind to denatured proteins and inhibit their aggregation, thus maintaining protein homeostasis and eventually thermotolerance (Su and Li, 2008). Unlike the limited number of HSF encoding genes in yeast and animals, plants possess large and diverse HSF gene families with complex species-specific functions (Scharf et al., 2012). There are 21 HSF genes in the A. thaliana genome (Nover et al., 2001), 25 HSF genes in the rice genome (Chauhan et al., 2011), and 56 HSF genes in the wheat genome (Xue et al., 2014). Interestingly, 64 HSF encoding genes were discovered in B. napus, rendering it the largest HSF family in plants identified so far (Zhu et al., 2017). The diversity of HSF genes in plants has likely arisen from whole-genome duplications events that took place during evolution (Chalhoub et al., 2014). In the case of B. napus, it is believed that the allopolyploidy process contributes significantly to the expansion of its HSF gene families, a key element for its adaptation and survivability under different environmental conditions (Zhu et al., 2017).
An example of a novel yet-conserved gene that plays a key role in the thermal resistance in B. napus is Brassica napus thermal resistance gene 1 (BnTR1). BnTR1 belongs to a new type of membrane-bound RINGv (Really Interesting New Gene Variant) protein with E3 ligase activity and was found to control the expression of HSFs related genes such as HSFA1a through the regulation of cytosolic Ca2+ concentration under heat stress, thus enhancing thermotolerance (Liu et al., 2014). It was also noticed that BnTR1 was able to increase heat tolerance in O. sativa without producing any defective growth phenotypes as seen with other thermal-resistant genes, such as: DREB1A, AtHSFA3 and BhHSF1 (Yoshida et al., 2008; Hong et al., 2009; Zhu et al., 2009). Liu et al. (2014) also found that the expression of BnTR1 increased the yield of rice suggesting its role in alleviating adverse environmental impacts on the crop through functioning as a master regulator of heat stress-responsive genes.
Other heat-responsive markers have also been identified in B. napus heat stress tolerance mechanisms. MYB genes were concomitantly reprogrammed and induced in the silique wall and seeds of B. napus following exposure to heat stress (Shamloo-Dashtpagerdi et al., 2018). After constructing a gene regulatory network of key stress regulatory genes that respond to drought and salt tolerance in B. napus, comparative genomic analysis from A. thaliana and Eutrema salsugineum and their B. napus homologs showed that transcription factor MYB44 and its direct regulator VIP1 had the largest number of connectivity and achieved the highest centrality values among the studied genes, implying its pivotal role in the regulatory network and ultimately in the plant stress tolerance system (Shamloo-Dashtpagerdi et al., 2018). At the physiological level, TFs also play important roles in plant growth and development. In A. thaliana, the ectopic expression of transcription factor AtMYB68 in B. napus following severe heat stress at flowering enhanced pollen viability and led to significant improvement in yield (Deng et al., 2020). These findings show the key role of TFs in the reproductive heat tolerance system pertained by A. thaliana and other related species (Yu et al., 2014) and altogether suggest a conserved stress response with key resistance factors in different tissues and plants (Yu et al., 2014; Huang et al., 2019).
The involvement of TFs in modulating glucosinolate concentrations under heat stress has also been reported. Glucosinolates are secondary metabolites found almost exclusively in the Brassicaceae (Fahey et al., 2001). Although they are known to be present in all parts of the plant, their concentration varies with different tissues and developmental stages (Fieldsend and Milford, 1994). While seeds are considered the major compartment for their storage, their synthesis takes place after flowering in the leaves and the reproductive tissues such as the silique wall (Nour-Eldin et al., 2012). Some glucosinolates play protective roles in the plant stress defense mechanisms against biotic and abiotic stress (Ludwig-Müller and Krishna, 2000; Martínez-Ballesta et al., 2013). Elevated temperature was found to increase glucosinolate concentrations in B. rapa, with a noticeable involvement of BrMYB28 and BrMYB34 transcription factors in this process (Justen and Fritz, 2013). In A. thaliana, mutants deficient in glucosinolate metabolism experienced reduced expression of cytoplasmic Hsp90 and less thermostability under elevated temperature (Ludwig-Müller and Krishna, 2000), whilst the supply of two exogenous glucosinolate hydrolysis products increased the expression of HSPs and improved thermotolerance (Hara et al., 2013). Conversely, after heat treatment of 20d-old siliques of B. napus, Yu et al. (2014) reported a dramatic decrease in the expression of several transcripts involved in glucosinolate metabolism such as those involved in the indolic and benzenic pathways, suggesting the suppression of glucosinolate biosynthesis in the silique wall due to heat stress. This shows that down-regulation of such pathways under stressful conditions could act as a protective measure to conserve energy (Yu et al., 2014), but also to optimize redistribution of glucosinolates (Martínez-Ballesta et al., 2013), ensuring survival under adverse environmental conditions. Such opposite trends in glucosinolate concentrations in response to heat treatment and the involvement of distinct signaling molecules, such as TFs, reveal a complex stress response powered by various factors such as the intensity and duration of stress as well as the plant developmental stage. As little is known about the biological functions of glucosinolate metabolism under heat stress, there is a need for more research to further understand the auxiliary roles of individual compounds in modulating the plant physiological status. Thus, revealing the interconnectivity between different components and pathways involved in an effective heat tolerance system in B. napus.
Polyploidy or whole-genome duplication (WGD) is one of the major forces in plant evolution (Soltis et al., 2014). The resulting speciation and biodiversity have influenced the structure and evolution of plant genomics, giving rise to highly dynamic genomes with broad gene expression changes (Wang R. et al., 2019). Most of the world crops have undergone polyploidy including wheat, cotton, tobacco, oats, potato and coffee. A recent allopolyploidy event took place around 7,500 years ago between ancestors of B. oleracea and B. rapa resulting in the formation of the highly adaptive species B. napus. This in addition to earlier polyploidization events, has caused an aggregate of 72x genome multiplication since the origin of angiosperms (Chalhoub et al., 2014). These evolutionary events have led to the acquisition of strong and valuable agronomic traits, giving these species the ability to adapt and survive different climatic conditions. For example, one copy of FLOWERING LOCUS C (FLC), a key adaptive gene controlling vernalization and photoperiod responses, exists in A. thaliana, while four copies exist in B. rapa and B. oleracea and nine or more in B. napus (Chalhoub et al., 2014).
There has been an increasing interest in studying the expression patterns of such duplicated genes. In polyploidy, duplicated genes may also be differentially spliced by the action of alternative splicing (AS) (Wang R. et al., 2019). This mechanism is characterized by the excision of introns and the production of differential exon combinations. The resulting genetic modifications alter transcript abundance and gene expression and eventually produce variant proteins (Liu Z. et al., 2018). During plant growth and development, AS plays key roles in different biological processes and most importantly, in response to biotic and abiotic stress (Guo et al., 2020). It is becoming increasingly evident that AS is not a by-product of stress but a regulatory part of the larger stress response (Kannan et al., 2018). Heat stress was found to induce an increase in AS events in genes that are associated with abiotic stress response such as those relating to protein folding (Chang et al., 2014; Kannan et al., 2018). In a recent study, analysis of differential AS landscape among A, B, and D wheat subgenomes under salt treatment found that homologous genes displayed differential AS responses under salt stress and more AS events were identified in the salt-sensitive cultivar than in the salt-tolerant one. Interestingly, specific AS events were found induced at different time points of salt stress (Guo et al., 2020). When multiple stresses were assessed, combined heat and drought stresses (HD) induced specific AS events compared with those induced by individual stresses. Upon comparison of differentially spliced genes (DSGs) and differentially expressed genes (DEGs), 40% of DSGs were overlapping with DEGs under heat stress and HD conditions (Liu Z. et al., 2018). These findings demonstrate the coordination between AS regulation and transcriptional regulation to orchestrate an effective stress response and ultimately contribute to abiotic stress tolerance. Similarly, in B. napus, RNA-Seq of plants subjected to cold, heat, and drought stress treatments also showed variation in expression and AS levels within the duplicated genes, with an overall AT subgenome biases in gene expression and CT subgenome biases in the extent of AS (Lee and Adams, 2020). Saminathan et al. (2015) hypothesized that polyploidy can influence AS, resulting in transcriptome modifications. In two independently synthesized lines of B. napus, alterations in AS events after polyploidy were reported (Zhou et al., 2011). Also, in response to heat, the number and type of AS events in the two related species A. thaliana and the highly thermotolerant polyploid Boechera depauperate varied significantly, suggesting a role for polyploidy in this variance. Moreover, this also demonstrates that AS responses to heat stress are species-specific (Kannan et al., 2018). In summary, the divergence in gene expression and AS patterns within the homologous genes may contribute to gene functional evolution which explains the flexibility of polyploids when responding to different abiotic stress (Zhang et al., 2010; Lee and Adams, 2020).
To further explore the role of AS in heat stress, Ling et al. (2018) investigated the existence of a “splicing memory” which is thought to be a key factor for achieving thermotolerance in Arabidopsis. As intron retention (IR) is the main AS form present in plants, the study aimed to investigate whether primed plants would preserve the same levels of these events during the recovery and memory-establishment phases and whether their response would differ from the response of non-primed plants after a recurrent exposure to heat. It was observed that heat-stressed plants accumulated unprocessed transcripts through splicing repression with marked levels of IR events, which eventually decreased to normal levels during recovery. In the second heat exposure, primed plants exhibited a very distinct response from non–primed plants. Primed plants remembered to undergo splicing and correctly processed transcripts similar to control plants under normal conditions. As a result, primed plants maintained a “splicing memory” that can carry out correct splicing and produce the necessary transcripts and proteins needed for the growth and development of plants after the cessation of stress, and subsequently ensuring the survivability of plants following another stress event (Ling et al., 2018). This further supports the role of AS an integral component in forming a priming-induced heat-stress memory in Arabidopsis (Sanyal et al., 2018). Further elucidation of this mechanism is needed to explore whether the discovered splicing-linked stress memory can be inherited across generations, or its function is limited somatically within an individual generation. Taken together, the polyploidy nature of B. napus and the regulatory role of AS in response to abiotic stress entail a further exploration of these evolutionary mechanisms under heat stress aiming to unveil novel species-specific interactions and to provide more understanding on its impact on thermotolerance, and more broadly, how polyploidy can impact the organismal response to heat stress.
In plants, high throughput technologies such as whole-genome bisulfite sequencing (WGBS) and chromatin immunoprecipitation (ChIP), and more recently nanopore technology, provide the opportunity to study epigenetic modifications in species and genotypes at genome-wide levels (Laird, 2010). Different types of epigenetic mechanisms exist and the most common are DNA methylation, histone modifications and small RNAs. Along with its vital role in maintaining genome integrity, increasing evidence indicates that epigenetic mechanisms in plants play a substantial part in modulating gene expression under different environmental conditions, including salinity, drought, extreme temperatures and exposure to heavy metals (Gao et al., 2014; Fan et al., 2020; Ueda and Seki, 2020).
In higher plants, 30-50% of all cytosine residues of the nuclear DNA are methylated (Gao et al., 2014), and DNA methylation occurs in all three sequence contexts: the symmetric CG and CHG, and the asymmetric CHH, where H stands for A, C, or T (Li et al., 2016; Klungland and Robertson, 2017). Despite the availability of epigenetic research in A. thaliana, these studies are still very limited in Brassica crops. Different methylation levels can produce different profiles of gene expression and transcription, giving rise to different phenotypic responses. In plants, genome methylation status is maintained by the dynamic function of DNA methyltransferase and demethylase enzymes. These enzymes are classified into different categories depending on their protein structure and function (Liu G. et al., 2018). Fan et al. (2020) identified 22 methylase genes and six demethylase genes through studying two varieties of B. napus (YN171 and 93275; Table 1). In addition to the variations in methylation patterns detected at different sequence locations (gene regions and up/downstream regions) and contexts (CHG, CHH, and CG), analysis of gene expression in 22 tissues across different developmental stages showed that these genes have different regulatory roles in plant growth and development.
Several reports have demonstrated that plant adaptations to different abiotic stresses are associated with differences in methylation levels (Marconi et al., 2013; Dhar et al., 2014; Liu et al., 2017). In B. napus, a large set of genes were associated with changes in cytosine methylation in response to heat treatment. More specifically, this variation was prominent at the genotype level where more DNA methylation occurred in the heat-sensitive cultivar and more DNA demethylation occurred in the heat-tolerant one (Gao et al., 2014). The expression level of DNA methyltransferase and demethylase enzymes changed significantly in response to salt and heat stress, indicating that methylation of some genes is essential for the response to abiotic stress in plants (Fan et al., 2020; Table 1). Similarly, position- and context-dependent methylation variations were also seen in B. rapa under heat stress. Analysis of differential methylation and differential gene expression found that low-expressed genes were enriched in genes with high methylation levels and vice versa, except for mCG methylation in exon regions. Interestingly, different sets of differentially methylated DEGs were involved at the early and late stages of heat treatment (Liu G. et al., 2018).
During the reproductive stages, more complex relationships between DNA methylation and gene expression have also been elucidated. During the two pollen pathways (gametophytic development and pollen embryogenesis) and in response to heat treatment, in vitro-cultured microspores of B. napus changed their gametophytic developmental pathway toward embryogenesis through an epigenetic reprogramming control. This was associated with a decrease in global DNA methylation and activation of cell proliferation, while DNA methylation increased with pollen and embryo differentiation in a cell-type-specific manner (Solís et al., 2012). On the contrary, Li et al. (2016) reported that short term heat shock treatment induced a decrease in global DNA methylation in cultured microspores of B. napus, suggesting a lack of trend in the changes of DNA methylation under heat stress in different species or cell types.
Histone modifications, including methylation, demethylation, and acetylation, are recognized as ubiquitous epigenetic marks that play a vital role in regulating chromatin structure and modulating gene expression (Xu et al., 2017). Other posttranslational modifications (PTMs), such as phosphorylation and ubiquitination have also been identified in transcriptional regulation in Arabidopsis (Füßl et al., 2018). The presence or absence of methylation on the lysine and arginine residues in histones affects their interaction with reader proteins and results in chromatin conformational changes. Eventually, these changes lead to either transcriptional activation or repression (Teperino et al., 2010). In plants, two Arg methylation sites (H3R17 and H4R3) and five Lys methylation sites (H3K4, H3K9, H3K27, H3K36, and H4K20) have been identified (Liu et al., 2010). In Arabidopsis, histone methylation involves both repressive (H4R3me2, H3K9me2/3, and H3K27me3) and active modifications (H4R3me2, H3K4me3, and H3K36me2/3) (Wang et al., 2016). In contrast to methylation, histone acetylation marks neutralize basic charges in histones, thus weakening histone/DNA interactions and making the DNA more accessible for the transcriptional machinery (Allis and Jenuwein, 2016). Histone modifications involve a complex network of epigenetic modifiers that serve to recognize, add or remove chemical moieties on histone tails or core domains (Xu et al., 2017). In response to stress, the combined activity of writers, readers and erasers regulate the level and type of histone modifications. Because tissue-specific expression is essential for stress response, these modifiers need to be recruited to specific loci for their participation. Various elements, such as: transcription factors and long non-coding RNAs have been proposed to act as recruiters of these regulators into specific chromatin sites (Deng et al., 2018). However, their role is still poorly explored in B. napus.
Recent studies have highlighted the role of histone modifications and their modifiers in response to abiotic stress. Alam et al. (2019) reported 18 B. rapa PHD (Plant homeodomain) finger genes as highly responsive to drought and salt treatments. As “epigenome readers,” PHD finger proteins can bind specifically to several histone modifications and confer activation or repression of the corresponding gene. Interestingly, some of these genes were found to be induced by more than 18-fold and others displayed different expression patterns at different time points of the treatments (Alam et al., 2019). In another study, 15 paralogous pairs of histone methyltransferase and demethylase genes also showed high variation in their expression profile in response to heat and cold stress where dynamic variations also took place in specific tissues and treatments, suggesting a complex role of these genes as robust candidates in the stress tolerance mechanisms of B. rapa (Liu G. et al., 2019).
Adding to this complexity, epigenetic regulators were found to interact with other stress-responsive elements to promote tolerance (Liu C. et al., 2019). To explore this relationship, Liu and colleagues investigated the role of polycomb repressive complex 2 (PRC2) in regulating ABA stress response at a genome level (Liu C. et al., 2019). PRC2 is one of the major families of polycomb group (PcG) factors that functions to induce a state of chromatin inactivation. Its core enzymes are responsible for histone H3 lysine-27 tri-methylation (H3K27me3), which is one of the major epigenetic marks that causes gene silencing (Liu C. et al., 2019). In A. thaliana more than 20% of genes are covered by H3K27me3 in one given organ (Lafos et al., 2011) and around 60% of protein-coding genes are silenced by H3K27me3 in different cell types (You et al., 2017). Therefore, H3K27me3 is considered a hallmark of gene repression, but its exact mechanism of action is not fully understood. H3K27me3 was found to preferentially target ABA-induced senescence-associated genes (SAGs) (Liu C. et al., 2019). ABA-induced senescence allows for the recycling and redistributing of nutrients to new growing organs and seeds, thus boosting the plant’s fitness and tolerance against abiotic stress (Zhao et al., 2016). In their study, Liu and colleagues found that mutants of PRC2 enzymes were hypersensitive to ABA. Accordingly, SAGs were derepressed and appeared highly induced in comparison to the wild type. This indicates that PRC2-mediated H3K27me3 plays a key role in attenuating the ABA response through buffering the expression of ABA-induced genes. Hence, fine tuning of ABA-induced senescence to protect the plant from oversensitive response (Liu C. et al., 2019). The role of PRC2 in regulating stress responses through modification of histones were also previously explored in different abiotic conditions such as drought, cold and salt where H3K27me3 modified the promotors of major stress response genes (Kwon et al., 2009).
Another component of the epigenetic machinery thought to play a role in the response to abiotic stress are non-coding RNAs, which are involved in the regulation of gene expression, along with other roles in different biological processes (Zhang Y. C. et al., 2014). The short (20-24 nucleotides) small RNAs (sRNAs) and the long (more than 200 nucleotides) non–coding RNAs (lncRNAs) have no or limited discernible protein-coding functionalities.
Small RNAs include micro-RNAs (miRNAs) which are the most thoroughly characterized, and small interfering RNAs (siRNAs) (Bartel, 2009). sRNAs are known to affect gene expression via two main mechanisms: transcriptional (TGS) and posttranscriptional gene silencing (PTGS). To date, several studies have identified both conserved and novel miRNAs from various model plants such as Arabidopsis and rice in response to different abiotic stress, yet there is limited knowledge about the small RNA population in B. napus where only a small number of miRNAs have been identified and their role in response to heat stress is largely unknown (Jian et al., 2016). In a recent study, miRNAs mediated thermotolerance through the action of miR160 which altered the expression of heat shock proteins and improved seed germination and seedling survival under heat stress in A. thaliana (Lin et al., 2018). Additionally, five conserved miRNAs families along with four novel ones were found responsive to heat stress and resulted in either positive or negative regulation of their putative genes in B. rapa (Yu et al., 2012). Using computational prediction methods, Xie et al. (2007) were the first to study miRNAs in B. napus where they detected five miRNAs in response to auxin, cadmium stress and phosphate starvation. However, this study was carried out prior to the availability of a reference sequence which explains the small number of miRNAs detected by their approach. Subsequently, a set of differentially expressed miRNAs in response to sulfate deficiency and cadmium stress have been identified (Huang et al., 2010). High-throughput sequencing revealed a diverse and complex set of core miRNAs in B. napus that were involved in seed imbibition response to salt and drought stresses, suggesting the role of miRNAs in response to stressful conditions during seed germination (Jian et al., 2016). In B. napus, the regulatory role of miRNAs has been explored and detected in response to different abiotic stresses, yet little or no information is available in response to heat stress. Hence, more studies are warranted on this aspect to gain insight about their role in thermotolerance.
Non–coding RNAs were found to function as precursors of small RNAs (Joshi et al., 2016). They are grouped according to their genomic positions and orientations as follows: long intergenic non-coding RNAs (lincRNAs), long non-coding natural antisense transcripts (lncNATs), long intronic non-coding RNAs, and sense lncRNAs. In plants, lncRNAs research is still in its infancy with only a few studies showing their role in plant development and adaptation to abiotic stress (Jha et al., 2020). Nonetheless, lncRNAs have been found highly expressed in a tissue-specific or stage-specific manner, with some having preferential expression in the reproductive stages as well (Zhang Y. C. et al., 2014). There is increasing evidence about the role of lncRNAs as “biological regulators” for various processes including developmental, biotic and abiotic stress in animals and plants (Chekanova, 2015). To explore the current understanding of the participatory role of lncRNAs in the tolerance response to different abiotic stress conditions in plants, Jha et al. (2020) reviewed several studies that identified lncRNAs under drought, heat, cold, nutrient deficiency and metal toxicity. As a result, a plethora of differentially expressed heat stress lncRNAs and their corresponding heat stress-responsive target genes were reported from different crops. Song et al. (2016) identified 9,687 novel lncRNAs in B. rapa in response to heat treatment using RNA-seq. Likewise, two up-regulated lncRNAs (TalnRNA27 and TalnRNA5), which are miRNA precursors, were up-regulated in wheat in response to heat stress (Xin et al., 2010). During pollen development and fertilization, a systematic analysis in B. rapa revealed that 47 cis-acting lncRNAs and 451 trans-acting lncRNAs were highly co-expressed with their target genes (Huang et al., 2018). Furthermore, a co-expression network of B. rapa depicted 210 DEGs, 4 miRNAs and 33 lncRNAs under heat stress, where the biological functions identified through GO enrichment analysis revealed terms such as “brassinosteroid mediated signaling pathway”, “response to stimulus” and “response to heat”, suggesting their contribution in the heat response through these signaling pathways (Wang A. et al., 2019). At the seedling stage, two genotypes of B. napus exhibiting different sensitivity to drought were exposed to drought stress and rehydration treatment. Genome-wide analysis showed that the up-and down-regulated mRNAs co-expressed with lncRNAs participated in different metabolic pathways and were involved in different regulatory mechanisms, which suggested the existence of divergent mechanisms to modulate the response to drought stress and re-watering treatment (Tan et al., 2020). These findings show that lncRNAs in plants could form an additional layer of complexity to other stress response mechanisms during abiotic stress events. In B. napus, only a few studies were found addressing the roles of lncRNAs in response to certain stress conditions such as pathogens (Joshi et al., 2016), cadmium (Feng et al., 2016) and drought stress (Tan et al., 2020), while to the best of our knowledge, no studies have investigated the role of lncRNAs in responses to heat stress in this species. There is a significant knowledge gap concerning the roles of different epigenetic mechanisms in response to heat stress in B. napus. Owing to its complexity and poorly understood mechanisms, a deeper knowledge of these intricate epigenetic components would serve as a stepping-stone to gain insights into the genomic regulation behind heat stress-mediated responses in this important crop species.
Omics approaches are now routinely used in helping to understand plant responses and adaptations to different biotic and abiotic conditions. However, their exploitation in B. napus is limited and the knowledge of the impact of heat stress on its yield determining reproductive stages is still fragmentary. Nevertheless, the availability of reference genomes in Brassica species has contributed to major improvements in understanding the genetics of these crops. This has created a foundation for identifying possible interactions that link genotypes and phenotypes. Subsequently, emerging genomics and transcriptomics studies have resulted in the development of transgenic cultivars with potentially stronger agronomic properties than their non-transgenic cultivars (Raza et al., 2021). In B. napus and other related species, several genomic projects have been conducted to study the effect of different abiotic stresses on different genetic and physiological properties. Using GWAS, numerous studies examined genomic regions and quantitative trait loci (QTL) controlling various traits that are affected by heat stress (Bac-Molenaar et al., 2015; Rahaman et al., 2018). Others identified transcription factors that play regulatory roles in abiotic stress responses including heat (Yoshida et al., 2008; He et al., 2016; Wang et al., 2018; Liang et al., 2019; Ghorbani et al., 2020) and some looked at gene families that are involved in controlling different hormonal signaling networks during heat stress (Justen and Fritz, 2013; Di et al., 2018).
At the transcript level, global transcriptome profiling allows for dynamic quantification of gene expression and their isoforms at different developmental stages in different tissues (Wang et al., 2009). For instance, combined RNA-seq and qRT-PCR identified and analyzed the expression patterns of 64 genes encoding BnaHSF proteins in the B. napus genome across different tissues and under heat, drought and high CO2 stresses (Zhu et al., 2017; Table 1). Similarly, organ-specific expression was detected using transcription profiling in B. napus siliques at the seed-filling stage, where some genes were preferentially expressed in heat-stressed silique walls or seeds and others were involved in specific biological functions of each tissue, such as glucosinolate metabolism in silique wall and flavonoid synthesis in seeds (Yu et al., 2014).
Examining the impact of high night temperature on the fatty acid composition of B. napus seeds, in-depth RNA-seq analysis revealed up-regulated gibberellin signaling which was also associated with active expression of genes involved in fatty acid catabolism and glyoxylate metabolism pathways (Zhou et al., 2018). Furthermore, a genome-wide analysis allowed the identification and functional analysis of 321 putative AP2/ERF TFs belonging to five major subfamilies, including DREB, ERF, AP2, RAV, and BnSoloist in B. napus (Owji et al., 2017). Gene expression analysis using RNA-seq revealed a high expression of AP2/ERF genes in the roots as compared to other organs. Gene ontology annotation, regulatory elements identification, and functional association network showed that most of these TFs are responsive to different abiotic stresses such as drought, salinity, and cold and play significant roles in the developmental processes, particularly organ speciation and embryogenesis (Owji et al., 2017).
Plant responses to heat are depicted by many quantitative factors such as time, intensity and duration of stress. At the molecular level, these responses result in profound structural and metabolic modifications, ranging from disruption of membrane and cytoskeleton stability to metabolic alterations of enzyme activity and accumulation of toxic metabolites such as reactive oxygen species (ROS). Although genomic and transcriptomic studies using RNA-seq have offered revolutionary approaches to interpret genomic data, these data do not always reflect the exact proteomic and metabolic state as gene products are also influenced by other processes such as post-translational and co-translational modifications. Hence, integration of metabolic and proteomic approaches will further help to better understand the events taking place at the molecular level (Rocco et al., 2013). Recently, omics approaches have been fine-tuned to elucidate the abiotic stress-related genes and develop tolerant cultivars that are capable to withstand current climatic and environmental changes (Raza et al., 2021). For example, studying the proteome response of a crop species during high-temperature stress will help to reveal the proteins and their corresponding biological mechanisms responding to these stresses. Moreover, as proteins act as direct effectors of phenotype, this proximity allows the prediction of affected phenotypes and hence, improving the breeding outcomes (Kosová et al., 2015). Although the proteome comprises of all protein components in a living organism, this set of proteins varies considerably depending on the developmental stage, growth conditions, tissue and cell type, giving rise to infinite proteomes within one organism (Kosová et al., 2015). Therefore, to gain a comprehensive insight into the protein profiles and pathways that are associated with heat, extensive research needs to be carried out not just at the organismal level but also looking at specific developmental stages and growth conditions.
Although numerous proteomic studies addressing the impact of various abiotic stresses in model species and various crops have been reported and reviewed (Benešová et al., 2012; Hossain et al., 2012; Kosová et al., 2013; Li et al., 2013), there is very limited knowledge to date on the proteome of B. napus under heat stress. The existing studies have identified proteins that are thought to be involved in thermotolerance, redox homeostasis, carbohydrate metabolism and protein synthesis and degradation (Lee et al., 2007; Neilson et al., 2010). In A. thaliana, analysis of the protein repertoire in the rosette leaves after exposing seedlings to short-term heat stress revealed differentially represented proteins that grouped into two main broad classes including energy and metabolism and stress response proteins (Rocco et al., 2013). In the first proteomic report in B. napus in response to heat stress, Ismaili et al. (2015) observed an increase in the abundance of ascorbate peroxidase protein and its corresponding mRNA transcript, suggesting its role in short-term high-temperature stress response. Also, the up regulation of proteins involved in energy and metabolism is thought to have utilized most of the nutritive reserves in the seedlings, resulting in a noticeable reduction in growth as a response to heat (Ismaili et al., 2015). Similarly, the first proteome analysis of B. napus response to drought stress also reported a reduction in growth that was associated with decreased abundance of HSP 70 and tubulin beta-2 proteins in the drought-sensitive and hybrid F1 lines (Mohammadi et al., 2012). Using yeast two-hybrid assay, the interactions among various Calcineurin B-like proteins (CBLs) and CBL-interacting protein kinases (CIPKs) were demonstrated in B. napus under several abiotic stresses (Zhang H. et al., 2014). Furthermore, expression levels of six selected BnaCBLs and 12 BnaCIPKs genes in response to salt, drought, cold, heat, and ABA showed that the two gene families were responsive to multiple stimuli, suggesting that the CBL-CIPK network in B. napus may be a common link for a variety of signaling pathways (Zhang H. et al., 2014). Yan et al. (2016) used a comparative proteomic approach to screen for thermotolerance genes in seeds of B. napus. As a result, a B. napus GLYI gene was identified. GLYI is a member of glyoxalase system that plays a role in methylglyoxal detoxification during carbohydrate and lipid metabolism. Two-dimensional gel analyses showed that the abundance of BnGLYI-3 protein in thermotolerant seeds significantly increased in response to heat stress, suggesting its role in inducing thermotolerance to high temperature stress in B. napus seeds (Yan et al., 2016).
Along with proteomics and other omics approaches, metabolomics has emerged as an exciting scientific discipline where cellular metabolites are profiled to reveal connections among genes, phenotypes and other cellular biomarkers in different biological species (Raza, 2020). The metabolome is the total pool of metabolites present in an organism (Razzaq et al., 2019). Through metabolomics studies, it is now possible to identify and measure primary and secondary metabolites to characterize genetic or environmental variations in plants (Razzaq et al., 2019). Under heat stress conditions, metabolomics provides a systems biology approach to understand complex metabolic networks and enhance thermotolerance and can be a useful tool in distinguishing between genotypes with different heat sensitivity and understanding how changes in the transcriptome and proteome manifest themselves at the cellular and tissue levels. Koscielny et al. (2018) identified 25 metabolic markers that discriminated between the heat-tolerant and -susceptible genotypes of B. napus during the reproduction stage under heat stress, through an untargeted metabolic assessment using gas chromatography-mass spectrometry. The latter finding demonstrates how genetic variations exhibited by different genotypes are translated into distinct agronomic traits that eventually allow its survivability under suboptimal conditions. In addition, metabolome information collected from stressed plants has also helped define the severity and timing of heat stress (Koscielny et al., 2018). In another metabolomic study, numerous unknown potential regulatory relationships involved in glucosinolate metabolism were identified through a doubled-haploid mapping population of B. napus. Feng et al. (2012) demonstrated the impact of genetic variation on the biochemical pathways through combining genetic and metabolic analysis of different glucosinolate components in the leaves and seeds. Subsequently, this has resulted in the identification of 105 metabolite QTLs and the construction of an advanced metabolic network for the glucosinolate composition in both leaves and seeds (Feng et al., 2012). Integration of metabolomic approaches with other omics tools has therefore the potential to reveal crucial interactions and regulatory networks between genes, proteins, and many essential regulatory components, where different responses meet to form a complex machinery of plant stress tolerance.
Despite current advances in exploring the impact of different biotic and abiotic stress factors within different Brassica species, this review has identified several knowledge gaps regarding the impact of heat stress on B. napus during its yield determining reproductive stages, where in-depth analysis of this subject is still needed. The opposite trends in expression of key genetic and epigenetic components identified in different species and in cultivars within the same species under various abiotic stresses call for further research in this field. Moreover, variations in intensity, time and duration of heat stress have been found to generate modified responses in plants. Therefore, to understand its impact on B. napus crops in specific geographical areas, regional studies on local cultivars should take place to simulate field conditions specific to these areas. To date, only a few studies have addressed the impact of heat stress on the metabolome and proteome of B. napus, and the underline cellular mechanisms are still poorly understood. On this account, combining genetic studies with different omics approaches will provide the means to explore the intricate responses arising following exposure to heat stress and accomplish a system-level understanding of how heat stress alters the yield and quality of the B. napus crop. It is expected that the findings of these studies will help breeders to produce cultivars with strong thermotolerance traits and to mitigate the detrimental effect of global warming on agriculture through contributing to sustainable food production and security.
FM, FR, MA, and JH conceived the research. MK undertook the review of the literature, prepared the figures, and drafted the manuscript. All authors contributed to and revised the manuscript, given the opportunity to comment, and approved the final version of the manuscript.
MK was supported by the BBSRC FoodBioSystems DTP (BB/T008776/1). JH was supported by the UK Department for the Environment, Food and Agriculture through project CH0110 Oilseed Rape Genetic Improvement Network (OREGIN).
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Ahmed, W., Xia, Y., Li, R., Bai, G., Siddique, K. H. M., and Guo, P. (2020). Non-coding RNAs: functional roles in the regulation of stress response in Brassica crops. Genomics 112, 1419–1424. doi: 10.1016/j.ygeno.2019.08.011
Alam, I., Liu, C. C., Ge, H. L., Batool, K., Yang, Y. Q., and Lu, Y. H. (2019). Genome wide survey, evolution and expression analysis of PHD finger genes reveal their diverse roles during the development and abiotic stress responses in Brassica rapa L. BMC Genomics 20:773. doi: 10.1186/s12864-019-6080-8
Allis, C. D., and Jenuwein, T. (2016). The molecular hallmarks of epigenetic control. Nat. Rev. Genet. 17, 487–500. doi: 10.1038/nrg.2016.59
Ambastha, V., and Leshem, Y. (2020). Differential cell persistence is observed in the Arabidopsis female gametophyte during heat stress. Plant Reprod. 33, 111–116. doi: 10.1007/s00497-020-00390-0
Angadi, S. V., Cutforth, H. W., Miller, P. R., Mcconkey, B. G., Entz, M. H., Brandt, S. A., et al. (2000). Response of three Brassica species to high temperature stress during reproductive growth. Can. J. Plant Sci. 80, 693–701.
Antoun, M., and Ouellet, F. (2013). Growth temperature affects inflorescence architecture in Arabidopsis thaliana. Botany 91, 642–651. doi: 10.1139/CJB-2013-0011
Bac-Molenaar, J. A., Fradin, E. F., Becker, F. F. M., Rienstra, J. A., van der Schoot, J., Vreugdenhil, D., et al. (2015). Genome-wide association mapping of fertility reduction upon heat stress reveals developmental stage-specific QTLs in Arabidopsis thaliana. Plant Cell 27, 1857–1874. doi: 10.1105/tpc.15.00248
Baillo, E. H., Kimotho, R. N., Zhang, Z., and Xu, P. (2019). Transcription factors associated with abiotic and biotic stress tolerance and their potential for crops improvement. Genes 10:771. doi: 10.3390/genes10100771
Baron, K. N., Schroeder, D. F., and Stasolla, C. (2012). Transcriptional response of abscisic acid (ABA) metabolism and transport to cold and heat stress applied at the reproductive stage of development in Arabidopsis thaliana. Plant Sci. 188–189, 48–59. doi: 10.1016/j.plantsci.2012.03.001
Bartel, D. P. (2009). MicroRNAs: target recognition and regulatory functions. Cell 136, 215–233. doi: 10.1016/j.cell.2009.01.002
Battisti, D. S., and Naylor, R. L. (2009). Historical warnings of future food insecurity with unprecedented seasonal heat. Science 323, 240–244. doi: 10.1126/science.1164363
Baux, A., Colbach, N., Allirand, J. M., Jullien, A., Ney, B., and Pellet, D. (2013). Insights into temperature effects on the fatty acid composition of oilseed rape varieties. Eur. J. Agron. 49, 12–19. doi: 10.1016/j.eja.2013.03.001
Benešová, M., Holá, D., Fischer, L., Jedelský, P. L., Hnilička, F., Wilhelmová, N., et al. (2012). The physiology and proteomics of drought tolerance in Maize: early stomatal closure as a cause of lower tolerance to short-term dehydration? PLoS One 7:e3801. doi: 10.1371/journal.pone.0038017
Bheemanahalli, R., Sathishraj, R., Manoharan, M., Sumanth, H. N., Raveendran, M., Ishimaru, T., et al. (2017). Is early morning flowering an effective trait to minimize heat stress damage during flowering in rice? Field Crops Res. 203, 238–242. doi: 10.1016/j.fcr.2016.11.011
Bheemanahalli, R., Sunoj, V. S. J., Saripalli, G., Prasad, P. V. V., Balyan, H. S., Gupta, P. K., et al. (2019). Quantifying the impact of heat stress on pollen germination, seed set, and grain filling in spring wheat. Crop Sci. 59, 684–696. doi: 10.2135/cropsci2018.05.0292
Blümel, M., Dally, N., and Jung, C. (2015). Flowering time regulation in crops-what did we learn from Arabidopsis? Curr. Opin. Biotechnol. 32, 121–129. doi: 10.1016/j.copbio.2014.11.023
Brunel-Muguet, S., D’Hooghe, P., Bataillé, M. P., Larré, C., Kim, T. H., Trouverie, J., et al. (2015). Heat stress during seed filling interferes with sulfur restriction on grain composition and seed germination in oilseed rape (Brassica napus L.). Front. Plant Sci. 6:213. doi: 10.3389/fpls.2015.00213
Chalhoub, B., Denoeud, F., Liu, S., Parkin, I. A. P., Tang, H., Wang, X., et al. (2014). Early allopolyploid evolution in the post-Neolithic Brassica napus oilseed genome. Science 345, 950–953. doi: 10.1126/science.1253435
Chandrasekaran, U., Xu, W., and Liu, A. (2014). Transcriptome profiling identifies ABA mediated regulatory changes towards storage filling in developing seeds of castor bean (Ricinus communis L.). Cell Biosci. 4:33. doi: 10.1186/2045-3701-4-33
Chang, C.-Y., Lin, W.-D., and Tu, S.-L. (2014). Genome-wide analysis of heat-sensitive alternative splicing in Physcomitrella patens. Plant Physiol. 165, 826–840. doi: 10.1104/pp.113.230540
Chao, L. M., Liu, Y. Q., Chen, D. Y., Xue, X. Y., Mao, Y. B., and Chen, X. Y. (2017). Arabidopsis transcription factors SPL1 and SPL12 confer plant thermotolerance at reproductive stage. Mol. Plant 10, 735–748. doi: 10.1016/j.molp.2017.03.010
Chauhan, H., Khurana, N., Agarwal, P., and Khurana, P. (2011). Heat shock factors in rice (Oryza sativa L.): genome-wide expression analysis during reproductive development and abiotic stress. Mol. Genet. Genom. 286, 171–187. doi: 10.1007/s00438-011-0638-8
Chaves, M. M., Flexas, J., and Pinheiro, C. (2009). Photosynthesis under drought and salt stress: regulation mechanisms from whole plant to cell. Ann. Bot. 103, 551–560. doi: 10.1093/aob/mcn125
Chaves, M. M., Pereira, J. S., Maroco, J., Rodrigues, M. L., Ricardo, C. P. P., Osório, M. L., et al. (2002). How plants cope with water stress in the field. Photosynthesis and growth. Ann. Bot. 89, 907–916. doi: 10.1093/aob/mcf105
Chekanova, J. A. (2015). Long non-coding RNAs and their functions in plants. Curr. Opin. Plant Biol. 27, 207–216. doi: 10.1016/j.pbi.2015.08.003
Del Olmo, I., Poza-Viejo, L., Piñeiro, M., Jarillo, J. A., and Crevillén, P. (2019). High ambient temperature leads to reduced FT expression and delayed flowering in Brassica rapa via a mechanism associated with H2A.Z dynamics. Plant J. 100:343356. doi: 10.1111/tpj.14446
Deng, M., Wang, Y., Kuzma, M., Chalifoux, M., Tremblay, L., Yang, S., et al. (2020). Activation tagging identifies Arabidopsis transcription factor AtMYB68 for heat and drought tolerance at yield determining reproductive stages. Plant J. 104, 1535–1550. doi: 10.1111/tpj.15019
Deng, X., Qiu, Q., He, K., and Cao, X. (2018). The seekers: how epigenetic modifying enzymes find their hidden genomic targets in Arabidopsis. Curr. Opin. Plant Biol. 45, 75–81. doi: 10.1016/j.pbi.2018.05.006
Dhar, M. K., Vishal, P., Sharma, R., and Kaul, S. (2014). Epigenetic dynamics: role of epimarks and underlying machinery in plants exposed to abiotic stress. Int. J. Genom. 2014, 187146. doi: 10.1155/2014/187146
Di, F., Jian, H., Wang, T., Chen, X., Ding, Y., Du, H., et al. (2018). Genome-wide analysis of the PYL gene family and identification of PYL genes that respond to abiotic stress in Brassica napus. Genes 9:156. doi: 10.3390/genes9030156
Dikšaitytė, A., Viršilė, A., Žaltauskaitė, J., Januškaitienė, I., and Juozapaitienė, G. (2019). Growth and photosynthetic responses in Brassica napus differ during stress and recovery periods when exposed to combined heat, drought and elevated CO2. Plant Physiol. Biochem. 142, 59–72. doi: 10.1016/j.plaphy.2019.06.026
Djanaguiraman, M., Schapaugh, W., Fritschi, F., Nguyen, H., and Prasad, P. V. V. (2019). Reproductive success of soybean (Glycine max L. Merril) cultivars and exotic lines under high daytime temperature. Plant Cell Environ. 42, 321–336. doi: 10.1111/pce.13421
Draeger, T., and Moore, G. (2017). Short periods of high temperature during meiosis prevent normal meiotic progression and reduce grain number in hexaploid wheat (Triticum aestivum L.). Theor. Appl. Genet. 130, 1785–1800. doi: 10.1007/s00122-017-2925-1
Echer, F. R., Oosterhuis, D. M., Loka, D. A., and Rosolem, C. A. (2014). High night temperatures during the floral bud stage increase the abscission of reproductive structures in cotton. J. Agron. Crop Sci. 200, 191–198. doi: 10.1111/jac.12056
Elferjani, R., and Soolanayakanahally, R. (2018). Canola responses to drought, heat, and combined stress: shared and specific effects on carbon assimilation, seed yield, and oil composition. Front. Plant Sci. 9:1224. doi: 10.3389/fpls.2018.01224
Erickson, A. N., and Markhart, A. H. (2002). Flower developmental stage and organ sensitivity of bell pepper (Capsicum annuum L.) to elevated temperature. Plant Cell Environ. 25, 123–130. doi: 10.1046/j.0016-8025.2001.00807.x
Fahey, J. W., Zalcmann, A. T., and Talalay, P. (2001). The chemical diversity and distribution of glucosinolates and isothiocyanates among plants. Phytochemistry 56, 5–51. doi: 10.1016/s0031-9422(00)00316-2
Fan, S., Liu, H., Liu, J., Hua, W., Xu, S., and Li, J. (2020). Systematic analysis of the DNA methylase and demethylase gene families in rapeseed (Brassica napus l.) and their expression variations after salt and heat stresses. Int. J. Mol. Sci. 21:253. doi: 10.3390/ijms21030953
Feng, J., Long, Y., Shi, L., Shi, J., Barker, G., and Meng, J. (2012). Characterization of metabolite quantitative trait loci and metabolic networks that control glucosinolate concentration in the seeds and leaves of Brassica napus. New Phytol. 193, 96–108. doi: 10.1111/j.1469-8137.2011.03890.x
Feng, S. J., Zhang, X. D., Liu, X. S., Tan, S. K., Chu, S. S., Meng, J. G., et al. (2016). Characterization of long non-coding RNAs involved in cadmium toxic response in: Brassica napus. RSC Adv. 6, 82157–82173. doi: 10.1039/c6ra05459e
Fieldsend, J., and Milford, G. F. J. (1994). Changes in glucosinolates during crop development in single- and double-low genotypes of winter oilseed rape (Brassica napus): production and distribution in vegetative tissues and developing pods during development and potential role in the recycling of sulphur within the crop. Ann. Appl. Biol. 124, 531–542. doi: 10.1111/j.1744-7348.1994.tb04157.x
Frenck, G., van der Linden, L., Mikkelsen, T. N., Brix, H., and Jørgensen, R. B. (2011). Increased [CO2] does not compensate for negative effects on yield caused by higher temperature and [O3] in Brassica napus L. Eur. J. Agron. 35, 127–134. doi: 10.1016/j.eja.2011.05.004
Füßl, M., Lassowskat, I., Née, G., Koskela, M. M., Brünje, A., Tilak, P., et al. (2018). Beyond histones: new substrate proteins of lysine deacetylases in Arabidopsis nuclei. Front. Plant Sci. 9:461. doi: 10.3389/fpls.2018.00461
Gao, G., Li, J., Li, H., Li, F., Xu, K., Yan, G., et al. (2014). Comparison of the heat stress induced variations in DNA methylation between heat-tolerant and heat-sensitive rapeseed seedlings. Breed. Sci. 64:125133. doi: 10.1270/jsbbs.64.125
Gao, Y., Li, T., Liu, Y., Ren, C., Zhao, Y., and Wang, M. (2010). Isolation and characterization of gene encoding G protein α subunit protein responsive to plant hormones and abiotic stresses in Brassica napus. Mol. Biol. Rep. 37, 3957–3965. doi: 10.1007/s11033-010-0054-x
Gawrysiak-Witulska, M., Rudzińska, M., Wawrzyniak, J., and Siger, A. (2012). The effect of temperature and moisture content of stored rapeseed on the phytosterol degradation rate. J. Am. Oil Chem. Soc. 89, 1673–1679. doi: 10.1007/s11746-012-2064-4
Gawrysiak-Witulska, M., Siger, A., Wawrzyniak, J., and Nogala-Kalucka, M. (2011). Changes in tocochromanol content in seeds of Brassica napus L. during adverse conditions of storage. J. Am. Oil Chem. Soc. 88, 1379–1385. doi: 10.1007/s11746-011-1793-0
Ghorbani, R., Zakipour, Z., Alemzadeh, A., and Razi, H. (2020). Genome-wide analysis of AP2/ERF transcription factors family in Brassica napus. Physiol. Mol. Biol. Plants 26, 1463–1476. doi: 10.1007/s12298-020-00832-z
Giorno, F., Wolters-Arts, M., Mariani, C., and Rieu, I. (2013). Ensuring reproduction at high temperatures: the heat stress response during anther and pollen development. Plants 2, 489–506. doi: 10.3390/plants2030489
Guan, Q., Yue, X., Zeng, H., and Zhu, J. (2014). The protein phosphatase RCF2 and its interacting partner NAC019 are critical for heat stress-responsive gene regulation and thermotolerance in Arabidopsis. Plant Cell 26, 438–453. doi: 10.1105/tpc.113.118927
Guo, W., Yu, K., Han, L., Li, X., Wang, H., Liu, Y., et al. (2020). Global profiling of alternative splicing landscape responsive to salt stress in wheat (Triticum aestivum L.). Plant Growth Regul. 92, 107–116. doi: 10.1007/s10725-020-00623-2
Hara, M., Harazaki, A., and Tabata, K. (2013). Administration of isothiocyanates enhances heat tolerance in Arabidopsis thaliana. Plant Growth Regul. 69, 71–77. doi: 10.1007/s10725-012-9748-5
Hasanuzzaman, M., Nahar, K., Alam, M. M., and Fujita, M. (2014). Modulation of antioxidant machinery and the methylglyoxal detoxification system in selenium-supplemented Brassica napus seedlings confers tolerance to high temperature stress. Biol. Trace Element Res. 161, 297–307. doi: 10.1007/s12011-014-0120-7
He, Y., Mao, S., Gao, Y., Zhu, L., Wu, D., Cui, Y., et al. (2016). Genome-wide identification and expression analysis of WRKY transcription factors under multiple stresses in Brassica napus. PLoS One 11:e0157558. doi: 10.1371/journal.pone.0157558
Hedhly, A. (2011). Sensitivity of flowering plant gametophytes to temperature fluctuations. Environ. Exp. Bot. 74, 9–16. doi: 10.1016/j.envexpbot.2011.03.016
Heinrich, M., Hettenhausen, C., Lange, T., Wünsche, H., Fang, J., Baldwin, I. T., et al. (2013). High levels of jasmonic acid antagonize the biosynthesis of gibberellins and inhibit the growth of Nicotiana attenuatastems. Plant J. 73, 591–606. doi: 10.1111/tpj.12058
Hong, B., Ma, C., Yang, Y., Wang, T., Yamaguchi-Shinozaki, K., and Gao, J. (2009). Over-expression of AtDREB1A in chrysanthemum enhances tolerance to heat stress. Plant Mol. Biol. 70, 231–240. doi: 10.1007/s11103-009-9468-z
Hossain, Z., Nouri, M. Z., and Komatsu, S. (2012). Plant cell organelle proteomics in response to abiotic stress. J. Proteome Res. 11, 37–48. doi: 10.1021/pr200863r
Huang, L., Dong, H., Zhou, D., Li, M., Liu, Y., Zhang, F., et al. (2018). Systematic identification of long non-coding RNAs during pollen development and fertilization in Brassica rapa. Plant J. 96, 203–222. doi: 10.1111/tpj.14016
Huang, R., Liu, Z., Xing, M., Yang, Y., Wu, X., Liu, H., et al. (2019). Heat stress suppresses Brassica napus seed oil accumulation by inhibition of photosynthesis and BnWRI1 pathway. Plant Cell Physiol. 60, 1457–1470. doi: 10.1093/pcp/pcz052
Huang, S. Q., Xiang, A. L., Che, L. L., Chen, S., Li, H., Song, J. B., et al. (2010). A set of miRNAs from Brassica napus in response to sulphate deficiency and cadmium stress. Plant Biotechnol. J. 8, 887–899. doi: 10.1111/j.1467-7652.2010.00517.x
Ismaili, A., Salavati, A., and Mohammadi, P. P. (2015). A comparative proteomic analysis of responses to high-temperature stress in hypocotyl of canola (Brassica napus L.). Protein Pept. Lett. 2, 285–299. doi: 10.2174/0929866521666141124102755
Jha, U. C., Nayyar, H., Jha, R., Khurshid, M., Zhou, M., Mantri, N., et al. (2020). Long non-coding RNAs: emerging players regulating plant abiotic stress response and adaptation. BMC Plant Biol. 20:466. doi: 10.1186/s12870-020-02595-x
Jian, H., Wang, J., Wang, T., Wei, L., Li, J., and Liu, L. (2016). Identification of rapeseed MicroRNAs involved in early-stage seed germination under salt and drought stresses. Front. Plant Sci. 7:658. doi: 10.3389/fpls.2016.00658
Joshi, R. K., Megha, S., Basu, U., Rahman, M. H., and Kav, N. N. V. (2016). Genome wide identification and functional prediction of long non-coding RNAs responsive to Sclerotinia sclerotiorum infection in Brassica napus. PLoS One 11:e0158784. doi: 10.1371/journal.pone.0158784
Justen, V. L., and Fritz, V. A. (2013). Temperature-induced glucosinolate accumulation is associated with expression of BrMYB transcription factors. HortScience 48, 47–52. doi: 10.21273/HORTSCI.48.1.47
Kagale, S., Divi, U. K., Krochko, J. E., Keller, W. A., and Krishna, P. (2007). Brassinosteroid confers tolerance in Arabidopsis thaliana and Brassica napus to a range of abiotic stresses. Planta 225, 353–364. doi: 10.1007/s00425-006-0361-6
Kannan, S., Halter, G., Renner, T., and Waters, E. R. (2018). Patterns of alternative splicing vary between species during heat stress. AoB Plants 10:ly013. doi: 10.1093/aobpla/ply013
Kazan, K., and Lyons, R. (2016). The link between flowering time and stress tolerance. J. Exp. Bot. 67, 47–60. doi: 10.1093/jxb/erv441
Kim, S. Y., Hong, C. B., and Lee, L. (2001). Heat shock stress causes stage-specific male sterility in Arabidopsis thaliana. J. Plant Res. 114, 301–307. doi: 10.1007/PL00013991
Klungland, A., and Robertson, A. B. (2017). Oxidized C5-methyl cytosine bases in DNA: 5-hydroxymethylcytosine; 5-formylcytosine; and 5-carboxycytosine. Free Rad. Biol. Med. 107, 62–68. doi: 10.1016/j.freeradbiomed.2016.11.038
Ko, J. H., Mitina, I., Tamada, Y., Hyun, Y., Choi, Y., Amasino, R. M., et al. (2010). Growth habit determination by the balance of histone methylation activities in Arabidopsis. EMBO J. 29, 3208–3215. doi: 10.1038/emboj.2010.198
Koscielny, C. B., Hazebroek, J., and Duncan, R. W. (2018). Phenotypic and metabolic variation among spring Brassica napus genotypes during heat stress. Crop Pasture Sci. 69, 284–295. doi: 10.1071/CP17259
Kosová, K., Prášil, I. T., and Vítámvás, P. (2013). Protein contribution to plant salinity response and tolerance acquisition. Int. J. Mol. Sci. 14, 6757–6789. doi: 10.3390/ijms14046757
Kosová, K., Vítámvás, P., Urban, M. O., Klíma, M., Roy, A., and Tom Prášil, I. (2015). Biological networks underlying abiotic stress tolerance in temperate crops-a proteomic perspective. Int. J. Mol. Sci. 16, 20913–20942. doi: 10.3390/ijms160920913
Krasensky, J., and Jonak, C. (2012). Drought, salt, and temperature stress-induced metabolic rearrangements and regulatory networks. J. Exp. Bot. 63, 1593–1608. doi: 10.1093/jxb/err460
Krasucki, W., Tys, J., Szafran, K., Rybacki, R., and Orlicki, Ł (2002). Influence of drying temperature on chemical composition of seeds of oilseed rape. Oilseed Crop 23, 427–438.
Kwon, C. S., Lee, D., Choi, G., and Chung, W.-I. I. (2009). Histone occupancy-dependent and -independent removal of H3K27 trimethylation at cold-responsive genes in Arabidopsis. Plant J. 60, 112–121. doi: 10.1111/j.1365-313x.2009.03938.x
Lafos, M., Kroll, P., Hohenstatt, M. L., Thorpe, F. L., Clarenz, O., and Schubert, D. (2011). Dynamic regulation of H3K27 trimethylation during Arabidopsis differentiation. PLoS Genet. 7:e1002040. doi: 10.1371/journal.pgen.1002040
Laird, P. (2010). Principles and challenges of genome-wide DNA methylation analysis. Nat. Rev. Genet. 11, 191–203. doi: 10.1038/nrg2732
Lamaoui, M., Jemo, M., Datla, R., and Bekkaoui, F. (2018). Heat and drought stresses in crops and approaches for their mitigation. Front. Chem. 6:26. doi: 10.3389/fchem.2018.00026
Lee, D. G., Ahsan, N., Lee, S. H., Kyu, Y. K., Jeong, D. B., Lee, I. J., et al. (2007). A proteomic approach in analyzing heat-responsive proteins in rice leaves. Proteomics 7, 3369–3383. doi: 10.1002/pmic.200700266
Lee, J. S., and Adams, K. L. (2020). Global insights into duplicated gene expression and alternative splicing in polyploid Brassica napus under heat, cold, and drought stress. Plant Genome 13:e20057. doi: 10.1002/tpg2.20057
Lesk, C., Rowhani, P., and Ramankutty, N. (2016). Influence of extreme weather disasters on global crop production. Nature 529, 84–87. doi: 10.1038/nature16467
Li, J., Huang, Q., Sun, M., Zhang, T., Li, H., Chen, B., et al. (2016). Global DNA methylation variations after short-term heat shock treatment in cultured microspores of Brassica napus cv. Topas. Sci. Rep. 6:38401. doi: 10.1038/srep38401
Li, W., Wei, Z., Qiao, Z., Wu, Z., Cheng, L., and Wang, Y. (2013). Proteomics analysis of alfalfa response to heat stress. PLoS One 8:e82725. doi: 10.1371/journal.pone.0082725
Liang, Z., Li, M., Liu, Z., and Wang, J. (2019). Genome-wide identification and characterization of the Hsp70 gene family in allopolyploid rapeseed (Brassica napus L.) compared with its diploid progenitors. PeerJ 7:e7511. doi: 10.7717/peerj.7511
Lin, J. S., Kuo, C. C., Yang, I. C., Tsai, W. A., Shen, Y. H., Lin, C. C., et al. (2018). MicroRNA160 modulates plant development and heat shock protein gene expression to mediate heat tolerance in Arabidopsis. Front. Plant Sci. 9:68. doi: 10.3389/fpls.2018.00068
Ling, Y., Serrano, N., Gao, G., Atia, M., Mokhtar, M., Woo, Y. H., et al. (2018). Thermopriming triggers splicing memory in Arabidopsis. J. Exp. Bot. 69, 2659–2675. doi: 10.1093/jxb/ery062
Liu, C., Cheng, J., Zhuang, Y., Ye, L., Li, Z., Wang, Y., et al. (2019). Polycomb repressive complex 2 attenuates ABA-induced senescence in Arabidopsis. Plant J. 97, 368–377. doi: 10.1111/tpj.14125
Liu, C., Lu, F., Cui, X., and Cao, X. (2010). Histone methylation in higher plants. Annu. Rev. Plant Biol. 61, 395–420. doi: 10.1146/annurev.arplant.043008.091939
Liu, G., Khan, N., Ma, X., and Hou, X. (2019). Identification, evolution, and expression profiling of histone lysine methylation moderators in Brassica rapa. Plants 8:526. doi: 10.3390/plants8120526
Liu, G., Xia, Y., Liu, T., Dai, S., and Hou, X. (2018). The DNA methylome and association of differentially methylated regions with differential gene expression during heat stress in Brassica rapa. Int. J. Mol. Sci. 19:1414. doi: 10.3390/ijms19051414
Liu, T., Li, Y., Duan, W., Huang, F., and Hou, X. (2017). Cold acclimation alters DNA methylation patterns and confers tolerance to heat and increases growth rate in Brassica rapa. J. Exp. Bot. 68, 1213–1224. doi: 10.1093/jxb/erw496
Liu, Z. B., Wang, J. M., Yang, F. X., Yang, L., Yue, Y. F., Xiang, J. B., et al. (2014). A novel membrane-bound E3 ubiquitin ligase enhances the thermal resistance in plants. Plant Biotechnol. J. 12, 93–104. doi: 10.1111/pbi.12120
Liu, Z., Qin, J., Tian, X., Xu, S., Wang, Y., Li, H., et al. (2018). Global profiling of alternative splicing landscape responsive to drought, heat and their combination in wheat (Triticum aestivum L.). Plant Biotechnol. J. 16, 714–726. doi: 10.1111/pbi.12822
Lohani, N., Jain, D., Singh, M. B., and Bhalla, P. L. (2020a). Engineering multiple abiotic stress tolerance in canola, Brassica napus. Front. Plant Sci. 11:3. doi: 10.3389/fpls.2020.00003
Lohani, N., Singh, M. B., and Bhalla, P. L. (2020b). High temperature susceptibility of sexual reproduction in crop plants. J. Exp. Bot. 71, 555–568. doi: 10.1093/jxb/erz426
Ludwig-Müller, J., and Krishna, P. (2000). A glucosinolate mutant of Arabidopsis is thermosensitive and defective in cytosolic Hsp90 expression after heat stress. Plant Physiol. 123, 949–958. doi: 10.1104/pp.123.3.949
Marconi, G., Pace, R., Traini, A., Raggi, L., Lutts, S., Chiusano, M., et al. (2013). Use of MSAP markers to analyse the effects of salt stress on DNA methylation in rapeseed (Brassica napus var. oleifera). PLoS One 8:e75597. doi: 10.1371/journal.pone.0075597
Martínez-Ballesta, M., del, C., Moreno, D. A., and Carvajal, M. (2013). The physiological importance of glucosinolates on plant response to abiotic stress in Brassica. Int. J. Mol. Sci. 14, 11607–11625. doi: 10.3390/ijms140611607
Mathur, S., and Jajoo, A. (2014). “Effects of heat stress on growth and crop yield of wheat (Triticum aestivum),” in Physiological Mechanisms and Adaptation Strategies in Plants Under Changing Environment, eds P. Ahmad and M. Wani (New York, NY: Springer), doi: 10.1007/978-1-4614-8591-9_8
Mills, J. T. (1989). Spoilage and Heating of Stored Agricultural Products. Prevention, Detection, and Control. Publ. 1823E. Ottawa, Ont: Agriculture Canada.
Mittler, R., Finka, A., and Goloubinoff, P. (2012). How do plants feel the heat? Trends Biochem. Sci. 37, 118–125. doi: 10.1016/j.tibs.2011.11.007
Mohammadi, P. P., Moieni, A., and Komatsu, S. (2012). Comparative proteome analysis of drought-sensitive and drought-tolerant rapeseed roots and their hybrid F1 line under drought stress. Amino Acids 43, 2137–2152. doi: 10.1007/s00726-012-1299-6
Morrison, M. J., and Stewart, D. W. (2002). Heat stress during flowering in summer Brassica. Crop Sci. 42, 797–803. doi: 10.2135/cropsci2002.7970
Namazkar, S., Stockmarr, A., Frenck, G., Egsgaard, H., Terkelsen, T., Mikkelsen, T., et al. (2016). Concurrent elevation of CO2, O3 and temperature severely affects oil quality and quantity in rapeseed. J. Exp. Bot. 67, 4117–4125. doi: 10.1093/jxb/erw180
Neilson, K. A., Gammulla, C. G., Mirzaei, M., Imin, N., and Haynes, P. A. (2010). Proteomic analysis of temperature stress in plants. Proteomics 10, 828–845. doi: 10.1002/pmic.200900538
Nour-Eldin, H. H., Andersen, T. G., Burow, M., Madsen, S. R., Jørgensen, M. E., Olsen, C. E., et al. (2012). NRT/PTR transporters are essential for translocation of glucosinolate defence compounds to seeds. Nature 488, 531–534. doi: 10.1038/nature11285
Nover, L., Bharti, K., Dö, P., Mishra, S. K., Ganguli, A., and Scharf, K.-D. (2001). Arabidopsis and the heat stress transcription factor world: how many heat stress transcription factors do we need? Cell Stress Chaperones 6, 177–189. doi: 10.1379/1466-12682001006<0177:aathst<2.0.co;2
Olejnik, D., Gogolewski, M., and Nogala-Kałucka, M. (1997). Isolation and some properties of plastochromanol-8. Nahrung 41, 101–104. doi: 10.1002/food.19970410209
Owji, H., Hajiebrahimi, A., Seradj, H., and Hemmati, S. (2017). Identification and functional prediction of stress responsive AP2/ERF transcription factors in Brassica napus by genome-wide analysis. Comput. Biol. Chem. 71, 32–56. doi: 10.1016/j.compbiolchem.2017.09.004
Pachauri, R. K., Allen, M. R., Barros, V. R., Broome, J., Cramer, W., Christ, R., et al. (2014). IPCC, 2014: Contribution of Working Groups, I, II and III to the fifth assessment report of the Intergovernmental Panel on Climate Change. Climate Change 2014: Synthesis Report. Geneva: IPCC.
Poidevin, L., Forment, J., Unal, D., and Ferrando, A. (2020). Transcriptome and translatome changes in germinated pollen under heat stress uncover roles of transporter genes involved in pollen tube growth. Plant Cell Environ. 44, 2167–2184. doi: 10.1111/pce.13972
Pokharel, M., Chiluwal, A., Stamm, M., Min, D., Rhodes, D., and Jagadish, S. V. K. (2020). High night-time temperature during flowering and pod filling affects flower opening, yield and seed fatty acid composition in canola. J. Agron. Crop Sci. 206, 579–596. doi: 10.1111/jac.12408
Rahaman, M., Mamidi, S., and Rahman, M. (2018). Genome-wide association study of heat stress-tolerance traits in spring-type Brassica napus L. under controlled conditions. Crop J. 6, 115–125. doi: 10.1016/j.cj.2017.08.003
Rang, Z. W., Jagadish, S. V. K., Zhou, Q. M., Craufurd, P. Q., and Heuer, S. (2011). Effect of high temperature and water stress on pollen germination and spikelet fertility in rice. Environ. Exp. Bot. 70, 58–65. doi: 10.1016/j.envexpbot.2010.08.009
Ratnayake, W. M. N., and Daun, J. K. (2004). “Chemical composition of canola and rapeseed oils,” in Rapeseed and Canola Oil. Production, Processing, Properties and Uses, ed. F. D. Gunstone (Boca Raton, FL: CRC Press), 37–78.
Raza, A. (2020). Metabolomics: a systems biology approach for enhancing heat stress tolerance in plants. Plant Cell Rep. doi: 10.1007/s00299-020-02635-8
Raza, A., Razzaq, A., Mehmood, S. S., Hussain, M. A., Wei, S., He, H., et al. (2021). Omics: the way forward to enhance abiotic stress tolerance in Brassica napus L. GM. Crops Food 12, 251–281. doi: 10.1080/21645698.2020.1859898
Razzaq, A., Sadia, B., Raza, A., Hameed, M. K., and Saleem, F. (2019). Metabolomics: a way forward for crop improvement. Metabolites 9:303. doi: 10.3390/metabo9120303
Rocco, M., Arena, S., Renzone, G., Scippa, G. S., Lomaglio, T., Verrillo, F., et al. (2013). Proteomic analysis of temperature stress-responsive proteins in Arabidopsis thaliana rosette leaves. Mol. BioSyst. 9, 1257–1267. doi: 10.1039/c3mb70137a
Rudzińska, M., Przybylski, R., and Wąsowicz, E. (2009). Products formed during thermo-oxidative degradation of phytosterols. J. Am. Oil Chem. Soc. 86, 651–662. doi: 10.1007/s11746-009-1397-0
Sage, T. L., Bagha, S., Lundsgaard-Nielsen, V., Branch, H. A., Sultmanis, S., and Sage, R. F. (2015). The effect of high temperature stress on male and female reproduction in plants. Field Crops Res. 182, 30–42. doi: 10.1016/j.fcr.2015.06.011
Sahni, S., Prasad, B. D., Liu, Q., Grbic, V., Sharpe, A., Singh, S. P., et al. (2016). Overexpression of the brassinosteroid biosynthetic gene DWF4 in Brassica napus simultaneously increases seed yield and stress tolerance. Sci. Rep. 6:28298. doi: 10.1038/srep28298
Saminathan, T., Nimmakayala, P., Manohar, S., Malkaram, S., Almeida, A., Cantrell, R., et al. (2015). Differential gene expression and alternative splicing between diploid and tetraploid watermelon. J. Exp. Bot. 66, 1369–1385. doi: 10.1093/jxb/eru486
Sanyal, R. P., Misra, H. S., and Saini, A. (2018). Heat-stress priming and alternative splicing-linked memory. J. Exp. Bot. 69, 2431–2434. doi: 10.1093/jxb/ery111
Saraswat, S., Yadav, A. K., Sirohi, P., and Singh, N. K. (2017). Role of epigenetics in crop improvement: water and heat stress. J. Plant Biol. 60, 231–240. doi: 10.1007/s12374-017-0053-8
Scharf, K. D., Berberich, T., Ebersberger, I., and Nover, L. (2012). The plant heat stress transcription factor (Hsf) family: structure, function and evolution. Biochim. Biophys. Acta 1819, 104–119. doi: 10.1016/j.bbagrm.2011.10.002
Schiessl, S. (2020). Regulation and subfunctionalization of flowering time genes in the allotetraploid oil crop Brassica napus. Front. Plant Sci. 11:605155. doi: 10.3389/fpls.2020.605155
Shamloo-Dashtpagerdi, R., Razi, H., Ebrahimie, E., and Niazi, A. (2018). Molecular characterization of Brassica napus stress related transcription factors, BnMYB44 and BnVIP1, selected based on comparative analysis of Arabidopsis thaliana and Eutrema salsugineum transcriptomes. Mol. Biol. Rep. 45, 1111–1124. doi: 10.1007/s11033-018-4262-0
Siger, A., Gawrysiak-Witulska, M., and Wawrzyniak, J. (2018). Changes in contents of phenolic compounds (sinapic acid derivatives) in seeds of Brassica napus L.under adverse storage conditions. Acta Sci. Pol. Technol. Aliment. 17, 367–375. doi: 10.17306/j.afs.2018.0596
Simpson, G. G. (2004). The autonomous pathway: epigenetic and post-transcriptional gene regulation in the control of Arabidopsis flowering time. Curr. Opin. Plant Biol. 7, 570–574. doi: 10.1016/j.pbi.2004.07.002
Solís, M. T., Rodríguez-Serrano, M., Meijón, M., Cañal, M. J., Cifuentes, A., Risueño, M. C., et al. (2012). DNA methylation dynamics and MET1a-like gene expression changes during stress-induced pollen reprogramming to embryogenesis. J. Exp. Bot. 63, 6431–6444. doi: 10.1093/jxb/ers298
Soltis, D. E., Visger, C. J., and Soltis, P. S. (2014). The polyploidy revolution then.and now: stebbins revisited. Am. J. Bot. 101, 1057–1078. doi: 10.3732/ajb.1400178
Song, X., Liu, G., Huang, Z., Duan, W., Tan, H., Li, Y., et al. (2016). Temperature expression patterns of genes and their coexpression with LncRNAs revealed by RNA-Seq in non-heading chinese cabbage. BMC Genomics 17:297. doi: 10.1186/s12864-016-2625-2
Su, P.-H., and Li, H. (2008). Arabidopsis stromal 70-kd heat shock proteins are essential for plant development and important for thermotolerance of germinating seeds. Plant Physiol. 146, 1231–1241. doi: 10.1104/pp.107.114496
Sun, F., Fan, G., Hu, Q., Zhou, Y., Guan, M., Tong, C., et al. (2017). The high-quality genome of Brassica napus cultivar ‘ZS11’ reveals the introgression history in semi-winter morphotype. Plant J. 92, 452–468. doi: 10.1111/tpj.13669
Tan, X., Li, S., Hu, L., and Zhang, C. (2020). Genome-wide analysis of long non-coding RNAs (lncRNAs) in two contrasting rapeseed (Brassica napus L.) genotypes subjected to drought stress and re-watering. BMC Plant Biol. 20:81. doi: 10.1186/s12870-020-2286-9
Tang, X., Hao, Y.-J., Lu, J.-X., Lu, G., and Zhang, T. (2019). Transcriptomic analysis reveals the mechanism of thermosensitive genic male sterility (TGMS) of Brassica napus under the high temperature inducement. BMC Genom. 20:644. doi: 10.1186/s12864-019-6008-3
Teperino, R., Schoonjans, K., and Auwerx, J. (2010). Histone methyl transferases and demethylases; Can they link metabolism and transcription? Cell Metab. 12, 321–327. doi: 10.1016/j.cmet.2010.09.004
Ueda, M., and Seki, M. (2020). Histone modifications form epigenetic regulatory networks to regulate abiotic stress response1. Plant Physiol. 182, 15–26. doi: 10.1104/pp.19.00988
United States Department of Agriculture [USDA] (2021). Oilseeds. Washington, DC: United States Department of Agriculture, Foreign Agricultural Service.
Verma, V., Ravindran, P., and Kumar, P. P. (2016). Plant hormone-mediated regulation of stress responses. BMC Plant Biol. 16:86. doi: 10.1186/s12870-016-0771-y
Vlahakis, C., and Hazebroek, J. (2000). Phytosterol accumulation in canola, sunflower, and soybean oils: effects of genetics, planting location, and temperature. J. Am. Oil Chem. Soc. 77, 49–53.
Wang, A., Hu, J., Gao, C., Chen, G., Wang, B., Lin, C., et al. (2019). Genome-wide analysis of long non-coding RNAs unveils the regulatory roles in the heat tolerance of Chinese cabbage (Brassica rapa ssp.chinensis). Sci. Rep. 9:5002. doi: 10.1038/s41598-019-41428-2
Wang, J., Meng, X., Yuan, C., Harrison, A. P., and Chen, M. (2016). The roles of crosstalk epigenetic patterns in Arabidopsis thaliana. Brief Funct. Genomics 15, 278–287.
Wang, P., Yang, C., Chen, H., Luo, L., Leng, Q., Li, S., et al. (2018). Exploring transcription factors reveals crucial members and regulatory networks involved in different abiotic stresses in Brassica napus L. BMC Plant Biol. 18:202. doi: 10.1186/s12870-018-1417-z
Wang, R., Liu, H., Liu, Z., Zou, J., Meng, J., and Wang, J. (2019). Genome-wide analysis of alternative splicing divergences between Brassica hexaploid and its parents. Planta 250, 603–628. doi: 10.1007/s00425-019-03198-z
Wang, Z., Gerstein, M., and Snyder, M. (2009). RNA-Seq: a revolutionary tool for transcriptomics. Nat. Rev. Genet. 10, 57–63. doi: 10.1038/nrg2484
Werteker, M., Lorenz, A., Johannes, H., Berghofer, E., and Findlay, C. S. (2010). Environmental and varietal influences on the fatty acid composition of rapeseed, soybeans, and sunflowers. J. Agron. Crop Sci. 196, 20–27. doi: 10.1111/j.1439-037X.2009.00393.x
Wu, X. L., Liu, Z. H., Hu, Z. H., and Huang, R. Z. (2014). BnWRI1 coordinates fatty acid biosynthesis and photosynthesis pathways during oil accumulation in rapeseed. J. Integr. Plant Biol. 56, 582–593. doi: 10.1111/jipb.12158
Xie, F. L., Huang, S. Q., Guo, K., Xiang, A. L., Zhu, Y. Y., Nie, L., et al. (2007). Computational identification of novel microRNAs and targets in Brassica napus. FEBS Lett. 581, 1464–1474. doi: 10.1016/j.febslet.2007.02.074
Xin, M., Wang, Y., Yao, Y., Xie, C., Peng, H., Ni, Z., et al. (2010). Diverse set of microRNAs are responsive to powdery mildew infection and heat stress in wheat (Triticum aestivum L.). BMC Plant Biol. 10:123. doi: 10.1186/1471-2229-10-123
Xu, Y., Zhang, S., Lin, S., Guo, Y., Deng, W., Zhang, Y., et al. (2017). WERAM: a database of writers, erasers and readers of histone acetylation and methylation in eukaryotes. Nucleic Acids Res. 45, D264–D270. doi: 10.1093/nar/gkw1011
Xue, G. P., Sadat, S., Drenth, J., and McIntyre, C. L. (2014). The heat shock factor family from Triticum aestivum in response to heat and other major abiotic stresses and their role in regulation of heat shock protein genes. J. Exp. Bot. 65, 539–557. doi: 10.1093/jxb/ert399
Yan, G., Lv, X., Gao, G., Li, F., Li, J., Qiao, J., et al. (2016). Identification and characterization of a glyoxalase I gene in a rapeseed cultivar with seed thermotolerance. Front. Plant Sci. 7:150. doi: 10.3389/fpls.2016.00150
Yoshida, T., Sakuma, Y., Todaka, D., Maruyama, K., Qin, F., Mizoi, J., et al. (2008). Functional analysis of an Arabidopsis heat-shock transcription factor HsfA3 in the transcriptional cascade downstream of the DREB2A stress-regulatory system. Biochem. Biophys. Res. Commun. 368, 515–521. doi: 10.1016/j.bbrc.2008.01.134
You, Y., Sawikowska, A., Neumann, M., Posé, D., Capovilla, G., Langenecker, T., et al. (2017). Temporal dynamics of gene expression and histone marks at the Arabidopsis shoot meristem during flowering. Nat. Commun. 8:15120. doi: 10.1038/ncomms15120
Young, L. W., Wilen, R. W., and Bonham-Smith, P. C. (2004). High temperature stress of Brassica napus during flowering reduces micro- and megagametophyte fertility, induces fruit abortion, and disrupts seed production. J. Exp. Bot. 55, 485–495. doi: 10.1093/jxb/erh038
Yu, E., Fan, C., Yang, Q., Li, X., Wan, B., Dong, Y., et al. (2014). Identification of heat responsive genes in Brassica napus siliques at the seed-filling stage through transcriptional profiling. PLoS One 9:e101914. doi: 10.1371/journal.pone.0101914
Yu, X., Wang, H., Lu, Y., de Ruiter, M., Cariaso, M., Prins, M., et al. (2012). Identification of conserved and novel microRNAs that are responsive to heat stress in Brassica rapa. J. Exp. Bot. 63, 1025–1038. doi: 10.1093/jxb/err337
Zandalinas, S. I., Fritschi, F. B., and Mittler, R. (2021). Global warming, climate change, and environmental pollution: recipe for a multifactorial stress combination disaster. Trends Plant Sci. 26, 588–599. doi: 10.1016/j.tplants.2021.02.011
Zandalinas, S. I., Mittler, R., Balfagón, D., Arbona, V., and Gómez-Cadenas, A. (2018). Plant adaptations to the combination of drought and high temperatures. Physiol. Plant. 162, 2–12. doi: 10.1111/ppl.12540
Zhang, H., Yang, B., Liu, W.-Z., Li, H., Wang, L., Wang, B., et al. (2014). Identification and characterization of CBL and CIPK gene families in canola (Brassica napus L.). BMC Plant Biol. 14:8. doi: 10.1186/1471-2229-14-8
Zhang, P. G., Huang, S. Z., Pin, A. L., and Adams, K. L. (2010). Extensive divergence in alternative splicing patterns after gene and genome duplication during the evolutionary history of Arabidopsis. Mol. Biol. Evol. 27, 1686–1697. doi: 10.1093/molbev/msq054
Zhang, S. S., Yang, H., Ding, L., Song, Z. T., Ma, H., Chang, F., et al. (2017). Tissue-specific transcriptomics reveals an important role of the unfolded protein response in maintaining fertility upon heat stress in Arabidopsis. Plant Cell 29, 1007–1023. doi: 10.1105/tpc.16.00916
Zhang, T., Shen, L. Y., Wang, R. X., Zou, Y., Zhao, J. H., Li, R. C., et al. (2012). Fertility alteration and utilization of male-sterile line 160S in Brassica napus. Acta Bot. Bor. Occid. Sin. 32, 35–41.
Zhang, Y. C., Liao, J. Y., Li, Z. Y., Yu, Y., Zhang, J. P., Li, Q. F., et al. (2014). Genome-wide screening and functional analysis identify a large number of long noncoding RNAs involved in the sexual reproduction of rice. Genome Biol. 15:512. doi: 10.1186/s13059-014-0512-1
Zhao, C., Liu, B., Piao, S., Wang, X., Lobell, D. B., Huang, Y., et al. (2017). Temperature increase reduces global yields of major crops in four independent estimates. Proc. Natl. Acad. Sci. U.S.A. 114, 9326–9331. doi: 10.1073/pnas.1701762114
Zhao, Y., Chan, Z., Gao, J., Xing, L., Cao, M., Yu, C., et al. (2016). ABA receptor PYL9 promotes drought resistance and leaf senescence. Proc. Natl. Acad. Sci. U.S.A. 113, 1949–1954. doi: 10.1073/pnas.1522840113
Zhou, L., Yan, T., Chen, X., Li, Z., Wu, D., Hua, S., et al. (2018). Effect of high night temperature on storage lipids and transcriptome changes in developing seeds of oilseed rape. J. Exp. Bot. 69, 1721–1733. doi: 10.1093/jxb/ery004
Zhou, R., Moshgabadi, N., and Adams, K. L. (2011). Extensive changes to alternative splicing patterns following allopolyploidy in natural and resynthesized polyploids. Proc. Natl. Acad. Sci. U.S.A. 108, 16122–16127. doi: 10.1073/pnas.1109551108
Zhu, X., Huang, C., Zhang, L., Liu, H., Yu, J., Hu, Z., et al. (2017). Systematic analysis of HsF family genes in the Brassica napus genome reveals novel responses to heat, drought and high CO2 stresses. Front. Plant Sci. 8:1174. doi: 10.3389/fpls.2017.01174
Zhu, Y., Wang, Z., Jing, Y., Wang, L., Liu, X., Liu, Y., et al. (2009). Ectopic over-expression of BhHsf1, a heat shock factor from the resurrection plant Boea hygrometrica, leads to increased thermotolerance and retarded growth in transgenic Arabidopsis and tobacco. Plant Mol. Biol. 71, 451–467. doi: 10.1007/s11103-009-9538-2
Keywords: Brassica napus, heat stress, flowering, alternative splicing, epigenetic modifications
Citation: Kourani M, Mohareb F, Rezwan FI, Anastasiadi M and Hammond JP (2022) Genetic and Physiological Responses to Heat Stress in Brassica napus. Front. Plant Sci. 13:832147. doi: 10.3389/fpls.2022.832147
Received: 09 December 2021; Accepted: 04 March 2022;
Published: 05 April 2022.
Edited by:
Luisa M. Sandalio, Spanish National Research Council (CSIC), SpainReviewed by:
Fugui Zhang, Anhui Agricultural University, ChinaCopyright © 2022 Kourani, Mohareb, Rezwan, Anastasiadi and Hammond. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Fady Mohareb, Zi5tb2hhcmViQGNyYW5maWVsZC5hYy51aw==; John P. Hammond, ai5wLmhhbW1vbmRAcmVhZGluZy5hYy51aw==
†Present address: Faisal I. Rezwan, Department of Computer Science, Aberystwyth University, Aberystwyth, United Kingdom
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.