 Arpit Gaur1,2Yogesh Jindal1Vikram Singh1Ratan Tiwari2Dinesh Kumar3Deepak Kaushik1Jogendra Singh4Sneh Narwal5Sarika Jaiswal3Mir Asif Iquebal3Ulavapp B. Angadi3Gyanendra Singh2Anil Rai3Gyanendra Pratap Singh2Sonia Sheoran2*
Arpit Gaur1,2Yogesh Jindal1Vikram Singh1Ratan Tiwari2Dinesh Kumar3Deepak Kaushik1Jogendra Singh4Sneh Narwal5Sarika Jaiswal3Mir Asif Iquebal3Ulavapp B. Angadi3Gyanendra Singh2Anil Rai3Gyanendra Pratap Singh2Sonia Sheoran2*
- 1Department of Genetics and Plant Breeding, CCS Haryana Agricultural University, Hisar, India
- 2ICAR-Indian Institute of Wheat and Barley Research, Karnal, India
- 3ICAR-Indian Agricultural Statistics Research Institute, New Delhi, India
- 4ICAR-Central Soil Salinity Research Institute, Karnal, India
- 5ICAR-Indian Agricultural Research Institute, New Delhi, India
Water-soluble carbohydrates (WSCs) play a vital role in water stress avoidance and buffering wheat grain yield. However, the genetic architecture of stem WSCs’ accumulation is partially understood, and few candidate genes are known. This study utilizes the compressed mixed linear model-based genome wide association study (GWAS) and heuristic post GWAS analyses to identify causative quantitative trait nucleotides (QTNs) and candidate genes for stem WSCs’ content at 15 days after anthesis under different water regimes (irrigated, rainfed, and drought). Glucose, fructose, sucrose, fructans, total non-structural carbohydrates (the sum of individual sugars), total WSCs (anthrone based) quantified in the peduncle of 301 bread wheat genotypes under multiple environments (E01-E08) pertaining different water regimes, and 14,571 SNPs from “35K Axiom Wheat Breeders” Array were used for analysis. As a result, 570 significant nucleotide trait associations were identified on all chromosomes except for 4D, of which 163 were considered stable. A total of 112 quantitative trait nucleotide regions (QNRs) were identified of which 47 were presumable novel. QNRs qWSC-3B.2 and qWSC-7A.2 were identified as the hotspots. Post GWAS integration of multiple data resources prioritized 208 putative candidate genes delimited into 64 QNRs, which can be critical in understanding the genetic architecture of stem WSCs accumulation in wheat under optimum and water-stressed environments. At least 19 stable QTNs were found associated with 24 prioritized candidate genes. Clusters of fructans metabolic genes reported in the QNRs qWSC-4A.2 and qWSC-7A.2. These genes can be utilized to bring an optimum combination of various fructans metabolic genes to improve the accumulation and remobilization of stem WSCs and water stress tolerance. These results will further strengthen wheat breeding programs targeting sustainable wheat production under limited water conditions.
Introduction
Current photosynthates and the photosynthates reserved in various vegetative tissues are two primary carbon sources for developing grain in wheat. Current photosynthates are directly transferred to developing grains during optimal conditions when photosynthesis is undergoing. When photosynthesis is absent during the dark period or depressed due to senescence and/or hostile growth conditions, developing grains become more dependent on the redistributed reserved photosynthates for their carbon requirement (Schnyder, 1993). So, it can be hypothesized that genotypes with more reserved photosynthates are likely to yield more under various growth conditions. In wheat and other temperate cereal crops, these reserved photosynthates are famously known as water-soluble carbohydrates (WSCs), which accumulate mainly in stem and leaf sheath from stem elongation to the early grain filling phase and remobilize to grains in the later grain filling stage, which further depends on the genotypic potential and environmental factors (Dreccer et al., 2009; Blum, 2011). The highest amount of WSCs is stored in the peduncle and penultimate of the wheat stem (Wardlaw and Willenbrink, 1994). Glucose, fructose, sucrose, and fructans are primary WSCs in wheat plants, which redistribute as a significant carbon source under depressed photosynthesis. Hence, it is evident that WSCs are involved in the complex plant growth and development system under optimal and stressed conditions. Many previous studies have advocated the significant role of WSCs in coping with various abiotic and biotic stresses. For instance, sucrose signaling pathways leading to fructans and anthocyanin accumulation play a significant role in abiotic and biotic stress (Van den Ende and El-Esawe, 2014). The role of WSCs in cold acclimatization and freezing has also been advocated by Livingston et al. (2006). During water stress, fructans protect the plants by maintaining membrane stability and reducing osmotic potential (Valluru and Van Den Ende, 2008; Livingston et al., 2009). The role of fructans in reactive oxygen species (ROS) scavenging mechanisms and phloem-mobile signaling has been proposed in plants under water-stressed conditions (Peshev et al., 2013; Van den Ende, 2013).
In wheat, remobilization of WSCs to developing grains may potentially contribute to 10-20% and 30-70% of total grain weight under well-watered and water-stressed conditions, respectively (Goggin and Setter, 2004; Rebetzke et al., 2008; Ovenden et al., 2017a). Previous studies have supported the increased amount of stem WSCs and their remobilization in drought-tolerant wheat genotypes under water-stressed conditions (Yáñez et al., 2017; Ram et al., 2018; Li et al., 2020). Indeed, under well-watered and water-stressed conditions, differentially expressed genes (DEGs) affecting the accumulation and remobilization of WSCs have been reported in drought-sensitive and tolerant wheat genotypes (Xue et al., 2008; Zhang et al., 2009; McIntyre et al., 2011; Yáñez et al., 2017). The 1-FEH (Zhang et al., 2009), 6-FEH (Van Riet et al., 2006), 1-SST and 6-SFT (Kawakami et al., 2005), and Ta-1FFT and Ta-6SFT genes (Huynh et al., 2008, 2012) have been cloned and identified as major responsible genes for WSCs metabolism in wheat; however, the important role of sugar transporter genes like TaSUT (Ahmed et al., 2018) and MYB gene TaMYb (Xue et al., 2011) has also been described earlier. Nevertheless, the correlation between wheat stem WSCs and final grain yield is still unclear as some studies advocate a positive correlation between these two (Xue et al., 2008; Gao et al., 2017), and some others demonstrate nonsignificant even negative correlations (del Pozo et al., 2016; Ovenden et al., 2017a). Studies have suggested that the recent progress in yield potential of wheat in Australia, the United Kingdom, and southern Yellow and Huai Valley of China is somehow due to intentional or unintentional selection for increased WSCs (van Herwaarden and Richards, 2002; Shearman et al., 2005; Gao et al., 2017). Therefore, improvement for WSCs in wheat can be valuable for improving grain yield under favorable and unfavorable growth conditions. And it can also help in enhancing water-stress tolerance in future wheat.
Variations among wheat genotypes for accumulation and remobilization of WSCs lay within the differential capacity of photosynthesis, respiration, and carbon use efficiency of individual genotypes (Tricker et al., 2018). Many studies have shown substantial genetic variations for stem WSCs under various growth conditions and environments in wheat (Ruuska et al., 2006; Rebetzke et al., 2007; Dong et al., 2016; Fu et al., 2020). These variations are very sensitive to the environment (Ruuska et al., 2006). Yang et al. (2007) observed that quantitative trait loci (QTLs) associated with the accumulation and redistribution of WSCs may have different expression patterns at various stages of growth and/or in different environments. Therefore, the need of identifying more stable markers for WSCs is suggested by Zhang et al. (2014). Several QTL-mapping studies have been conducted to dissect the complex genetic architecture of WSCs in wheat at different growth stages and environments (Snape et al., 2007; Yang et al., 2007; Rebetzke et al., 2008; McIntyre et al., 2012; Li et al., 2020). Nevertheless, these studies produced limited knowledge about the genetic architecture of WSCs in wheat as bi-parental mapping populations have limited genetic diversity and recombination events. However, the foundation laid by these QTL mapping studies cannot be denied. Genome-wide association studies (GWAS) help overcome the above-said drawbacks of bi-parental QTL mapping and improve the resolution to get precise and deeper insights into genetic aspects of any trait. Only seven GWAS for the accumulation of stem WSC at different growth stages and environments have been conducted to the best of our knowledge. The first GWAS for accumulation of WSC at flowering, mid-grain filling, and maturity in wheat was carried out by Zhang et al. (2014) and Li et al. (2015) using 209 simple sequence repeat (SSR) markers. Dong et al. (2016) carried out GWAS for WSC accumulation at 14DAA in wheat stems by using a 90K SNP array (Wang et al., 2014). Since then, only three GWAS for understanding the genetic makeup of WSC in the wheat stem have been conducted (Ovenden et al., 2017b; Fu et al., 2020; Guerra et al., 2021).
GWAS and QTL mapping studies on WSC in wheat have contributed to decipher the various genetic aspects of this trait. However, the number of these studies is few. Moreover, none covered the individual components of stem WSCs, viz. glucose, sucrose, fructose, and fructans. Based on the knowledge of stem WSC as a trait, this study covers the individual components of WSC as: (1) final content or concentration of total WSC depends on the initial content/concentration of each component, (2) fructans, which make nearly eighty per cent of the total stem WSC, are derived from the polymerization of sucrose, (3) the amount of glucose, fructose, and sucrose triggers the hydrolysis of fructans to hexose and monosaccharides, which later becomes the source of carbon to developing grains. Presumably, this is the first report on GWAS that aimed to unveil the genetic architecture of accumulation of WSC and its major components 15 days after anthesis (15DAA) under three water regimes (irrigated, rainfed, and drought).
Materials and Methods
Plant Material and Field Experiments
An IIWBR Wheat Association Mapping Panel (IWAMP) was developed with a total of 301 bread wheat (Triticum aestivum L.) genotypes selected based on the pedigree information and Shanon index from a hefty germplasm pool of seven thousand lines (Supplementary Table 1) (Sheoran et al., 2019). This IWAMP was evaluated for two consecutive cropping seasons, Rabi 2017-18 and Rabi 2018-19, under three water regimes [irrigated (IR), rainfed (RF), and complete drought (DT)] at two locations: Chaudhary Charan Singh Haryana Agricultural University (CCS HAU) Hisar (29°09′N 75°42′E) and ICAR-Indian Institute of Wheat and Barley (ICAR-IIWBR) Karnal (29.686°N 76.989°E). The Hisar location represents the hot typic arid subeco region 2.3 of India, and Karnal represents semiarid eco-subregion 4.1 (Mandal et al., 2014). Trials of IR were carried out at both locations for both cropping seasons, whereas trials of RF and DT were conducted at Hisar and Karnal, respectively. For IR, four pre-anthesis and two post-anthesis irrigations were given, whereas irrigations were entirely ceased for RF and DT. For DT, genotypes were raised under rainout shelters that only cover the whole cropping area on a rainy day. All the trails were established in an alpha lattice design consisting of two replications. Each genotype was hand sown using IIWBR-Dibbler to ensure maximum homogeneity among trials (Sharma et al., 2016). Henceforth, a total of 8 environments and pooled-over water-managements namely E1: Hisar-Irrigated-2017-18; E2: Hisar-Rainfed-2017-18; E3: Karnal-Irrigated-2017-18; E4: Karnal-Drought-2017-18; E5: Hisar-Irrigated-2018-19; E6: Hisar-Rainfed-2018-19; E7: Karnal-Irrigated-2018-19; E8: Karnal-Drought-2018-19; IR: BLUEs (pooling the environments of irrigated water management); RF: BLUEs (pooling the environments of rainfed management); DT: BLUEs (pooling the environments of complete drought management) were used for association analysis.
Phenotyping
Glucose, fructose, sucrose, fructans, and total WSC were assayed in each genotype’s peduncle (the first internode to spike base) under all the environments using Fourier-transformed near spectroscopy (FT-NIR). Briefly, from ten randomly chosen plants from each plot, peduncles of the main culm were sampled at 15DAA by cutting with a sharp knife just above the first internode and near the base of the spike, followed by immediate removal of leaf blades. Samples were dried for 60 min in a pre-heated (100°C) dry-air oven, followed by drying at 60°C for 48 h. Furthermore, each sample was chopped into small pieces of 1-2 mm. Finally, the whole sample was used for FT-NIR spectra acquisition. The in-house partial least square regression models (PLS-R), which are in line with Wang et al. (2011), were then used to quantify targeted traits (unpublished data). All PLS-R models were highly reliable with their high coefficient of determination (R2).
Statistical Analysis of Phenotypic Data
For the present study, location × year × water regime was considered as an environment. Following mixed linear models (MLM) fitted with restricted maximum likelihood (REML) methods were used to create a type III ANOVA table with Satterthwaite’s method for the eight individual environments and environments pooled across three water regimes in R-studio using the package “lmerTest” (Kuznetsova et al., 2017), and the best linear unbiased estimators (BLUE) for each line were subsequently calculated:
in the above models “(1| X)” indicates a random intercept of the source of variation with a fixed mean.
Broad-sense heritability (h2) was calculated from the obtained variances using the following equations and was expressed in percentage:
where the , , and , are genetic variance, residual variance, and variance due to interaction between genotype and environment, respectively. The “y” and “r” are the total number of environments and replications, respectively.
Genotyping
Genomic DNA was extracted from fresh leaves of 21-day-old seedlings using the CTAB method (Murray and Thompson, 1980). Genotyping was done using a 35K Axiom Wheat Breeders’ Array (Affymetrix UK Ltd., United Kingdom). SNP calling and filtering were done by PLINK (Purcell et al., 2007). Markers with missing values (GENO) more than 1%, minor allele frequency (MAF) less than 5%, and individuals with more than 10% missing SNP calls were rejected. The markers obtained were then ordered according to the genetic map available at CerealDb (Allen et al., 2017).
Genetic Diversity, Population Structure, and Linkage Disequilibrium
Genetic diversity (H) and the polymorphism information content (PIC) for extracted SNP markers were calculated with PowerMarker v3.2.5 (Liu and Muse, 2005). A population structure based on a Bayesian algorithm was inferred in STRUCTURE software v2.3.4 (Hubisz et al., 2009) using a K value between 1 and 10. Three independent iterations for each K value were carried out with 105 burn-in periods and Markov Chain Monte Carlo (MCMC) iterations. Optimal sub-population was identified using ad hoc statistics with ΔK (Evanno et al., 2005), calculated using the STRUCTURE HARVESTER tool (Earl and vonHoldt, 2012). Additionally, population stratification was validated using Principal Component Analysis (PCA) and p-value-based neighbor-joining (NJ) cluster analysis as performed with R-based GAPIT v2.0 (Tang et al., 2016) and MEGA-X (Kumar et al., 2018b), respectively. A marker-based VanRaden Kinship (VanRaden, 2008) (K-matrix) was also generated between all the 301 genotypes using GAPT-R v2.0 (Tang et al., 2016). Linkage disequilibrium (LD) was calculated separately for A, B, and D sub-genomes and the whole genome using PLINK software (Purcell et al., 2007). Later, the pattern of LD decay was estimated by plotting correlated pairwise r2 values against genetic distance with LOESS smoothening in R-studio using the “loess()” function. In the whole genome LD decay, a genetic distance at which the LOESS curve first touches the baseline r2 of 0.1 was considered the putative QTL-confidence interval (QTL-CI).
Nucleotide-Trait Association
A compressed mixed linear model (CMLM) was implicated in GAPIT-R v2.0 (Tang et al., 2016) to estimate the association between polymorphic loci and BLUE values of each trait observed under an individual environment and estimated across different water regimes. To avoid any spurious effect produced by population structure, the K-PC model (Zhao et al., 2007) was adopted in which the first three principal components (PC) were used as covariates together with kinship information (K-matrix). P-values thresholds were determined by adjusting the false discovery rate (FDR) to 10%. A nucleotide-trait association (NTA) with threshold -log10 p > 3.0 was declared significant. Quantile-Quantile and Manhattan plots were recreated with the CMplot-R package in R-studio.
Each significant SNP was called a quantitative trait nucleotide (QTN), and the region within the genome-wide attenuation distance was called a quantitative nucleotide region (QNR). Each QNR is called “q” followed by the abbreviation of the trait (in the capital), a hyphen, a chromosome, a period, and a sequence in Arabic numbers (e.g., qWSC-1A.1).
Quest for Putative Candidate Genes
Physical positions of significant SNPs in IWGSC RefSeq. v1.1 were retrieved from the CerealsDB (Wilkinson et al., 2020). Candidate genes within the 1-Mbp window of physically localized SNPs were retrieved from the EnsemblPlants database using the biomartr package of R, along with their genomic summary, InterPro, and GO term details. For in silico expression analysis, Affymetrix data of GSE9767 (Xue et al., 2008) and GSE87325 (Kumar et al., 2018a) were retrieved from NCBI’s Gene Expression Omnibus (GEO) database. The CEL files were processed with MAS5 algorithm with “affiyo” in R studio to extract the normalized transcript per million (TPM). The TPM was converted to log2 values in the MSExcel 2019. Additionally, the Genevestigator tool was used for the mRNA-based in silico expression analysis of overlapping genes under drought and related perturbation. The list of candidate genes was narrowed down to high-priority candidate genes by evaluating them on three candidate gene prioritization approaches, viz. (1) knowledge-based gene prioritization (KGP), (2) differential expression-based gene prioritization (DEGP), and (3) co-regulatory network-based gene prioritization (CNGP). The detailed method of gene prioritization can be found in Supplementary Method 1.
Results
Phenotypic Variability
The IWAMP carried continuous variation for all the studied traits under individual environments (E01-E08) and in the pooled data across three water regimes (Figure 1). Summary statistics on resulting BLUEs for individual environments and across three water regimes are detailed in Supplementary Table 2 and Table 1, respectively. Among all, fructans were predominating non-structural sugar, contributing 70 to 75 percent to total WSCs and TNSC. In contrast, the contribution of sucrose, glucose, and fructose was nearly 19-21, 7-8, and 6-7%, respectively. Values for all the soluble sugars were higher under stressed conditions than optimal conditions (Figure 1). An increment in all the studied traits showed a linear relationship with the degree of stress (Figure 1). Amount of glucose, fructose, sucrose, fructans, WSCs and TNSC under rainfed condition increased by 21, 17, 19, 9, 7, and 13%, respectively, and under drought stress 21, 21, 22, 61, 26, and 46%, respectively.
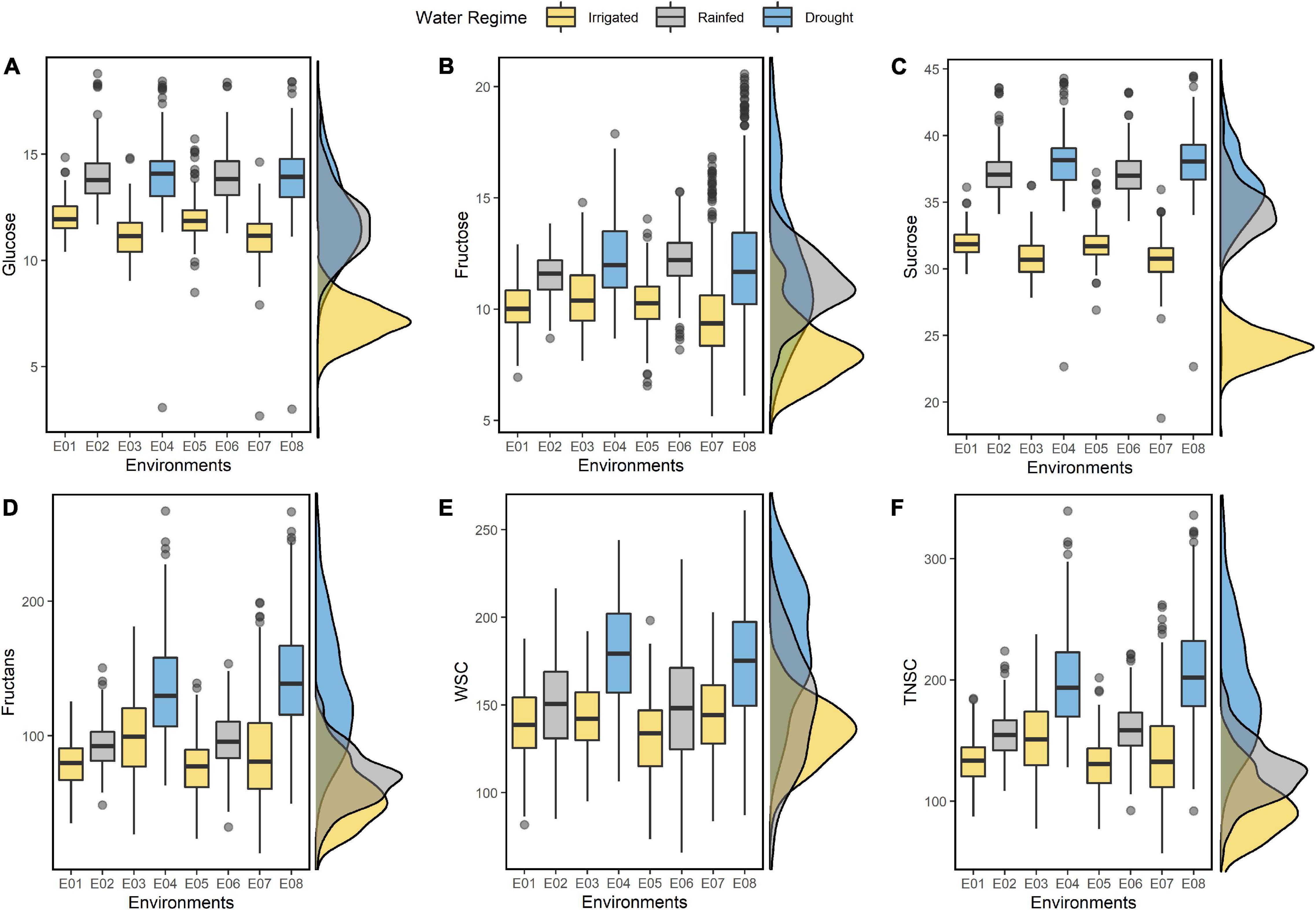
Figure 1. Boxplots summarizing the resulting BLUEs of glucose (A), fructose (B), sucrose (C), fructans (D), WSC (E), and TNSC (F) estimated under eight different environments (E01-E08). On the left of each boxplot, the corresponding density plots show the distribution pattern of the traits with resulting BLUEs estimated by pooling of environments across corresponding water regimes.
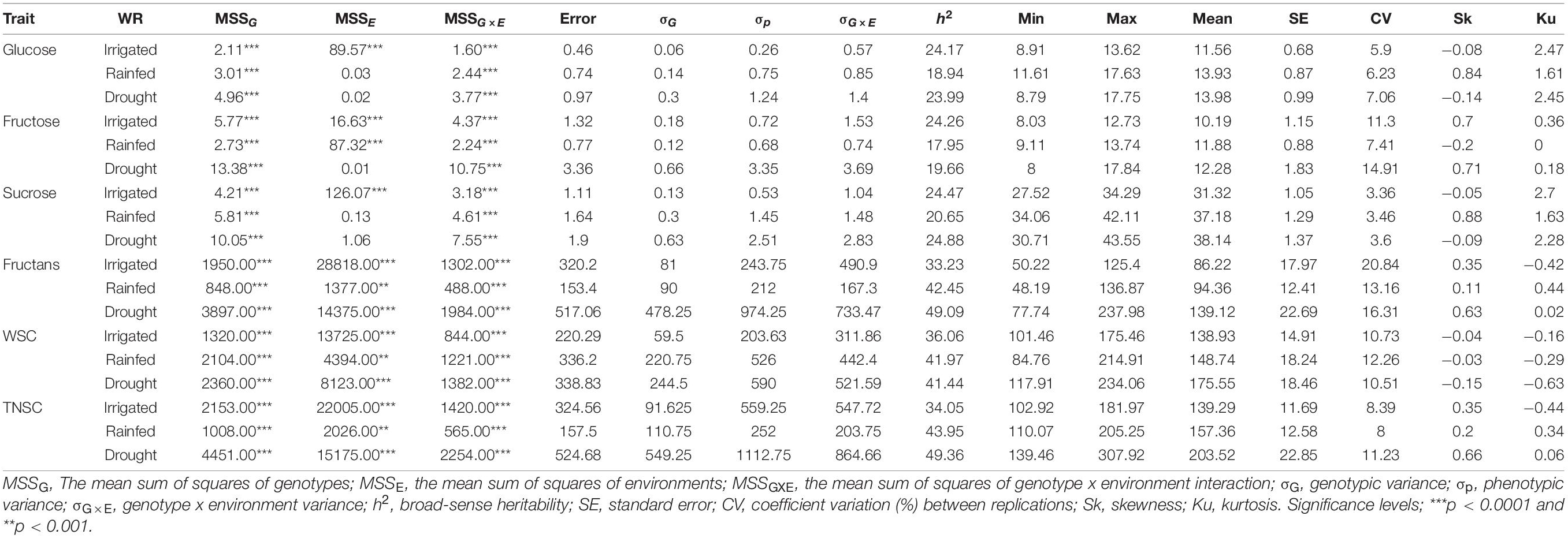
Table 1. A type-III ANOVA table with Satterthwaite’s method for a mixed linear model fitted by the restricted maximum likelihood method for the studied traits pooled across three different water regimes (irrigated, rainfed, and drought) and summary statistics.
The ANOVA for eight environments (Supplementary Table 2) and pooled across three water regimes (Table 1) indicated substantial phenotypic variability within IWAMP at p < 0.0001 for all the six traits. Pooled ANOVA further demonstrated a significant effect of genotype (G) x environment (E) interaction on all the studied traits at p < 0.0001. Each of the studied traits showed high environmental wise h2 (Supplementary Table 2), ranging between 63.18% (for sucrose under E05) and 86.78% (for TNSC under E04), whereas low-to-moderate pooled h2 (Table 1) varying from 17.95% (for fructose under RF) to 49.36% (for fructans and TNSC under DT) was estimated across three water regimes. Thus, the extent of genetic variability and differential response of accumulation of water-soluble sugars in the wheat peduncle of IWAMP to water availability was in favor to conduct the GWAS.
Marker Coverage, Population Structure, and Linkage Disequilibrium
A 35K SNP array-based genotyping of IWAMP produced 14,571 polymorphic SNPs after pre-processing of data. These markers covered a genome-wide genetic distance of 4,527 centimorgans (cM) with an average density of 0.31 cM (Figure 2A and Supplementary Table 3). About 5,728 SNPs were reported on sub-genome A, 7,357 on sub-genome B, and 1,486 on sub-genome D. The highest (1,360) and lowest (61) SNPs were reported on Chromosomes 2B and 4D, respectively. Furthermore, mean values for MAF, H, and PIC ranged from 0.16 (5D) to 0.32 (1D), 0.26 (5D) to 0.39 (1D) and 0.18 (5D) to 0.23 (5A), respectively. A summary of statistics generated from 14,751 polymorphic SNP markers is tabulated in Supplementary Table 3.
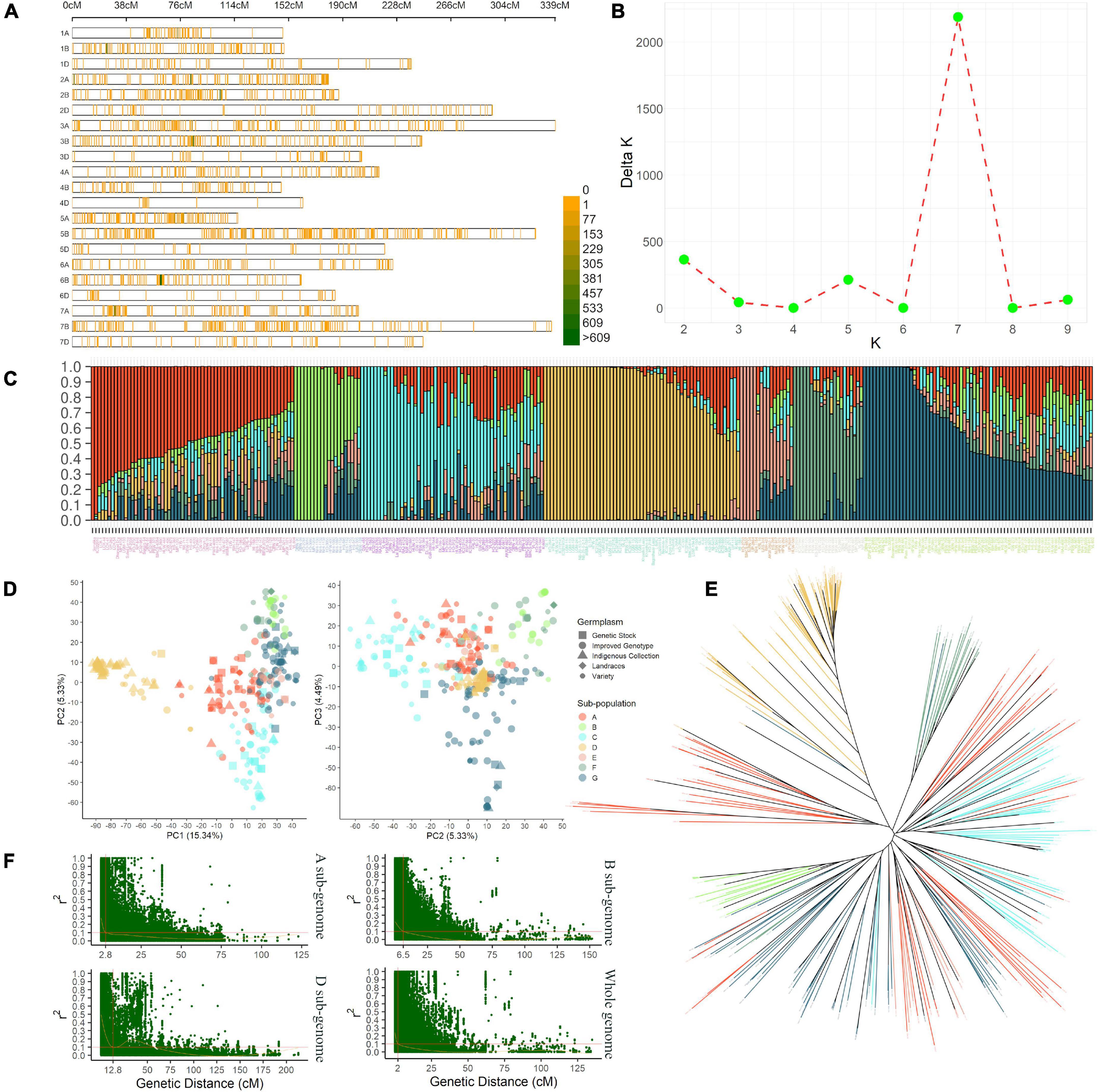
Figure 2. The density plot (A) shows the SNPs’ chromosome-wise distribution pattern with a 1 Mbp window in present IWAMP. The line graph (B) indicates the best K value estimated by the method described in Evanno et al. (2005) to identify the optimum number of sub-populations in the present AMP. The bar plot (C) summarizes the genetic composition of AMP; followed to this are PCA plots (D) and NJ cladogram (E), which show a nearly similar pattern for population stratification. Finally, the scatter plots (F) show the genetic distance at which LD decayed for sub-genomes (A,B,D) and the whole genome.
The stratification of IWAMP with STRUCTURE analysis defined seven subpopulations with substantial admixture (Figures 2B,C). The PCA explaining 25.16% with the first three PCs of the variation (Figure 2D) and an NJ-based cladogram (Figure 2E) showed fine consistency with population structure. The highest 69 genotypes clustered in sub-population “G” and the lowest 21 in sub-population “F.” The sub-population “G” was mainly composed of 32 improved genotypes (IG), which are ca. 36% of the total IG included in the present IWAMP. Contrastingly, sub-population “F” was a mixture of different germplasm types included in the present IWAMP. Interestingly, ca. 72% of the total indigenous collections (IC) clustered into sub-population “D,” whereas the rest were in sub-population “A,” “C,” and “G.” Based on decay in linkage disequilibrium (LD), the CI of a QNR was determined. At the cut-off value (r2 = 0.1), LD decayed at genetic distances 2.8 cM, 6.5 cM, and 12.8 cM for sub-genomes A, B, and D, respectively, and at 2 cM for the whole genome (Figure 2F).
QTNs for Stem Water-Soluble Carbohydrates
In the existing GWAS, eleven phenotypic datasets consisting of eight environments (E01-E08), which received three different water regimes, viz. irrigation, rainfed, and severe drought and pooled over different water regimes, were used along with 14,571 high-quality SNP markers. The quantile-quantile plots (Figure 3) indicated that the compressed maximum likelihood method (CMLM) was the best model to identify significant quantitative trait nucleotides (QTNs) with adequate control on the false-positive rate. The present GWAS yielded 382 significant QTNs at -log10 p values ≥ 3.0 for the targeted traits across eleven datasets (Figure 3) on all chromosomes except 4D. The number of QTNs detected for different traits in eleven datasets was 28 (E01), 67 (E02), 31 (E03), 66 (E04), 40 (E05), 78 (E06), 41 (E07), 56 (E08), 54 (IR), 30 (RF), and 79 (DT). Unlike previous reports, we called an association between QTN and a trait as nucleotide-trait association(s) (NTAs). Thus, there were a total of 570 NTAs in the present GWAS (Table 2). The maximum NTAs were identified on sub-genome B (191) and individual chromosome 5B (42). Dataset E08 and E01 produced the highest (33) and the lowest (14) NTAs in the present GWAS. In different datasets, there were a total of 86, 101, 88, 93, 113, and 89 NTAs for glucose, fructose, sucrose, fructans, TNSC, and WSC, respectively. Comparatively, there were more NTAs for fructans, TNSC, and WSC under water-stressed conditions and for glucose, fructose, and sucrose under irrigated conditions.
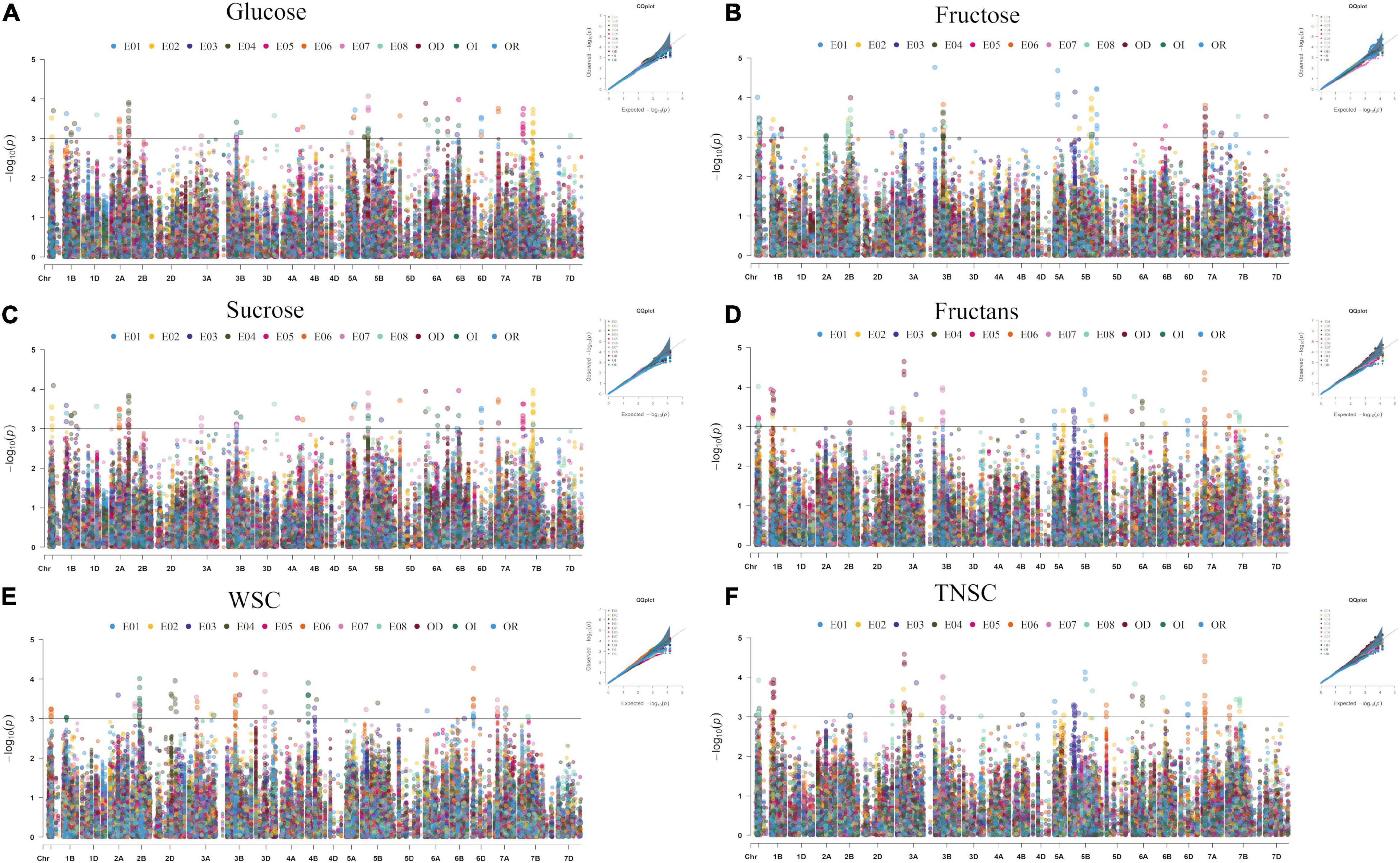
Figure 3. Manhattan and quantile-quantile plots summarizing the GWAS results for glucose (A), fructose (B), sucrose (C), fructans (D), TNSC (E), and WSC (F) in IWAMP of 301 bread wheat genotypes evaluated under eight distinct environments. The x-axis and y-axis refer to chromosomes and –log10(p) values for different traits, and the colors of dots refer to different datasets.
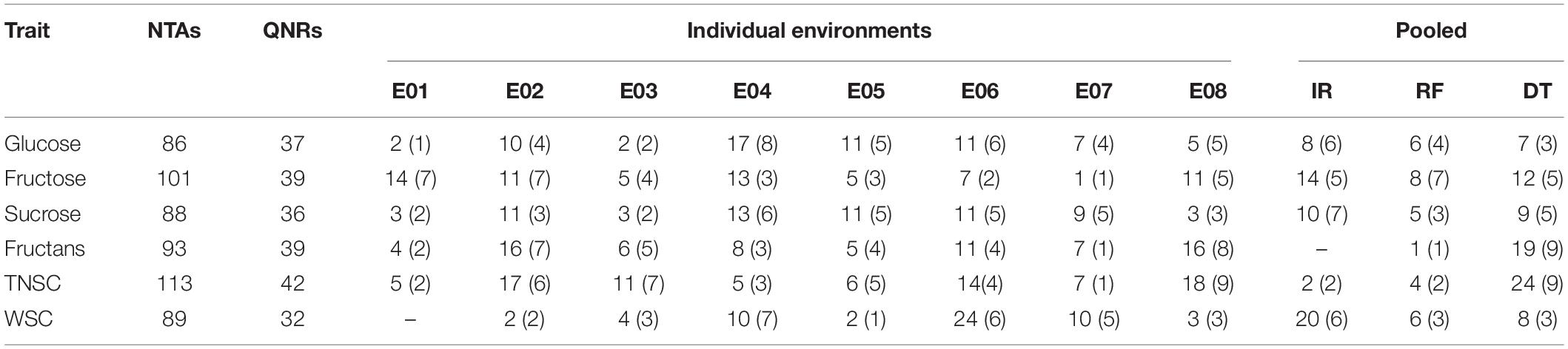
Table 2. Number of NTAs (QNRs) identified in the present GWAS for WSC and its major components with eleven datasets.
There was no significant QTN for WSC under E01 environment and fructans with pooled BLUEs corresponding to irrigated conditions. The effect of significant QTNs and explained phenotypic variance (PVE%) ranged from −0.61 to 0.72 and 3.47 to 6.69%, −1.10 to 1.47 and 3.55 to 7.76%, −0.87 to 1.03 and 3.47 to 7.01%, −23.19 to 15.09 and 3.16 to 10.31%, −24.51 to 16.34 and 3.1 to 9.7%, and −15.63 to 13.89 and 4.55 to 7.15% for individual traits glucose, fructose, sucrose, fructans TNSC, and WSC, respectively.
NTA common across two or more enviroments and/or with BLUEs was considered stable. Thus, we identified a total of 121 QTNs producing 163 stable NTAs (Supplementary Table 4), corresponding to different water regimes for six targeted traits, viz. glucose (21), fructose (34), sucrose (24), fructans (20), TNSC (30), and WSC (34), which were located on all chromosomes except on Chromosomes 1D, 4A, 4D, and 5D.
We used genome-wide LD attenuation distance (2 cM) to delimit detected QTNs into 112 genomic regions (Table 2 and Supplementary Table 4). These regions were designated as quantitative nucleotide regions (QNRs). The QNRs were unevenly dispersed across twenty wheat chromosomes (Figure 4). At most, fifteen QNRs were detected at Chromosome 5B and at least one at Chromosome 1D. Maximum (51) QNRs were dispersed within sub-genome B followed by A (44) and D (17). It indicated that sub-genome B is likely to be more critical for WSCs metabolism in bread wheat. The primitive sub-genome B is expected to inherit from Aegilops speltoides. The numbers of QNRs per trait were 37 (glucose), 39 (fructose), 36 (sucrose), 39 (fructans), 42 (TNSC), and 32 (WSC). We identified 25 QNRs that were consistent for two or more traits in more than one environment. A QNR, qWSC-3B.2, at physical position 3B:83.69-85.27cM, is likely to be a hotspot for WSCs metabolism since it carried 32 QTNs and was comprehensively associated with all the studied traits in different environments and water regimes. Another QNR, qWSC-7A.2 at 7A:29.86-30.66 cM harboring 22 NTAs for fructose, sucrose, fructans, and TNSC, was likely to be a hotspot for WSC metabolism under water-stressed environments; however, within this genomic region, there were two QTNs for WSC content under E05 (irrigated conditions).
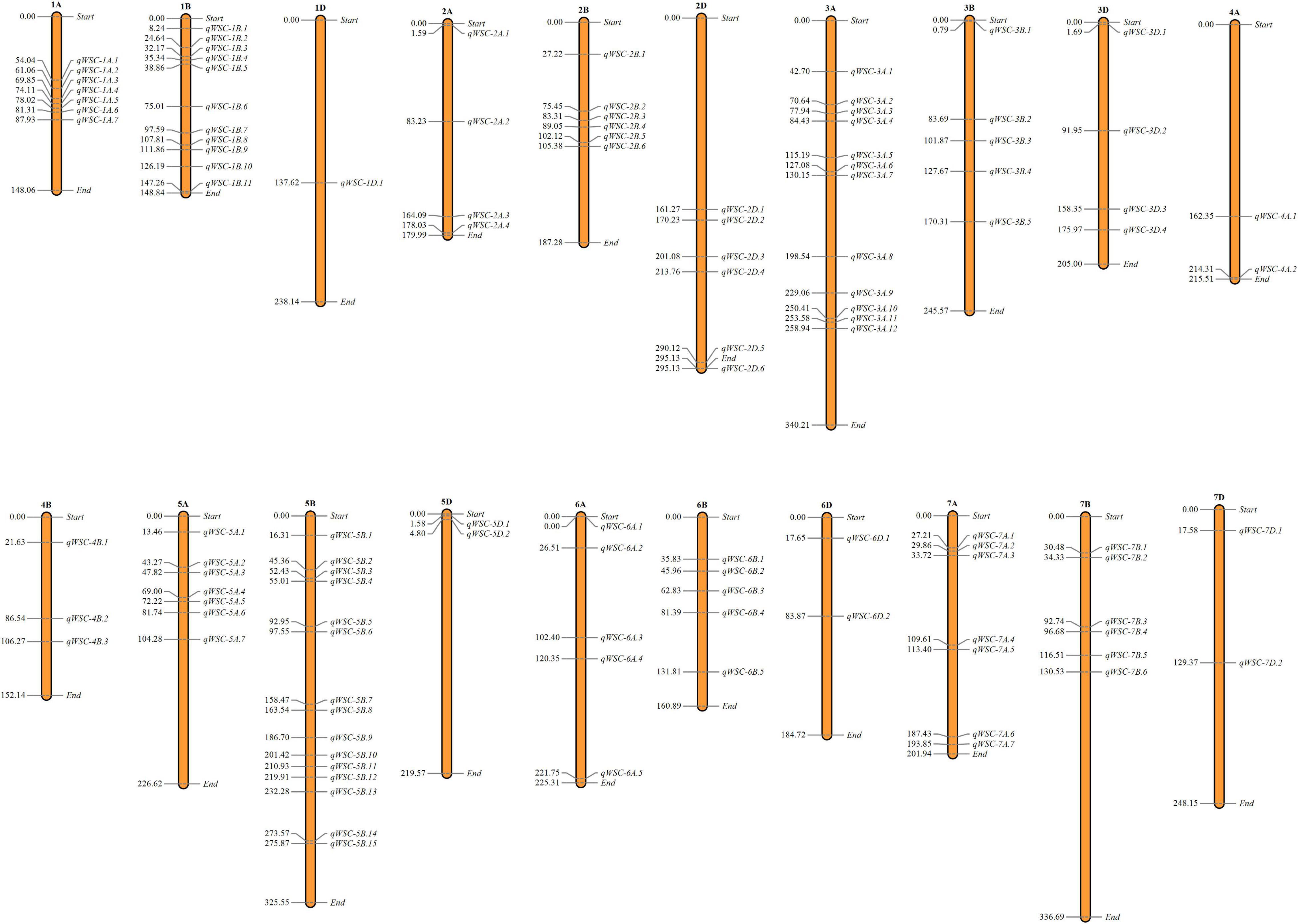
Figure 4. Dispersion of detected quantitative trait nucleotide regions (QNRs) across the bread wheat genome. The number on the left denotes the position of the first quantitative trait nucleotide (QTN) detected in the respective QNRs.
QTN Localization and Genes in QNRs
High confidence genomic locations were identified for 209 QTNs distributed under 81 QNRs. A total of 3,496 high-confidence IWGSC genes were pulled out within the 1 Mbp window of the localized QTNs (Supplementary Table 5). One hundred fifty genes overlapped 161 QTNs, indicating that 48 QTNs were intergenic (Supplementary Table 6). Maximum 3 QTNs were found in gene TraesCS7B02G446300 (7B: 709513978). Furthermore, nearly 1K genes were found in the 1-Mbp window of stable QTNs (Supplementary Table 7). All the genes retrieved were subjected to InterPro and gene ontology annotation using the EnsemblPlant and the AgriGO database in which InterPro domains and GO terms were retrieved for 2,939 and 2,501 genes, respectively. According to the ReviGO (Supek et al., 2011) summarization, these genes were engaged in 572 different biological processes in various cell components through their 480 molecular functions (Figure 5 and Supplementary Table 8). Maximum 203 genes were involved in the oxidation-reduction process followed by protein phosphorylation (173), regulation of transcription (127), transmembrane transport (79), proteolysis (70), and carbohydrate metabolism (60). Among overlapping genes, InterPro domains and GO terms were retrieved for 139 and 121 genes. Maximum 42 of overlapping genes were indulged in the oxidation-reduction process followed by protein phosphorylation (34), proteolysis (27), transmembrane transport (27), and translation (22) (Figure 5 and Supplementary Table 8).
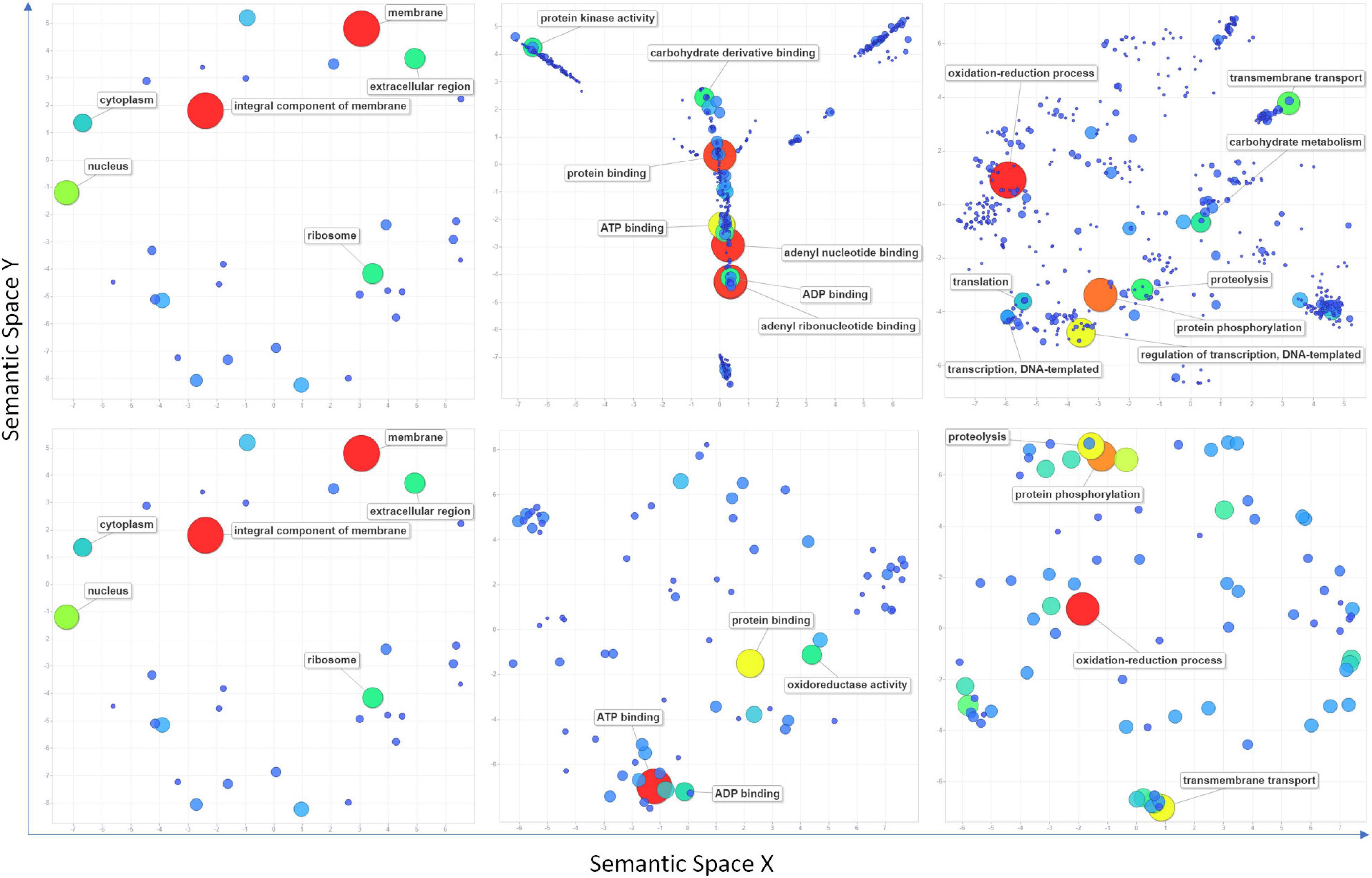
Figure 5. Scatter plots show cluster representatives of GO terms for cellular components, molecular functions, and biological processes (left to right) in semantic spaces for all the retrieved genes (above) within a 1 Mbp window from localized significant SNPs and overlapping genes (below). The size and colors of dots indicate the number of genes. Coordinates of each dot in semantic space are detailed in Supplementary Table 8. Terms with the highest values are mentioned in the figure.
We referred the public Affymetrix data of GSE9767 (Xue et al., 2008) and GSE87325 (Kumar et al., 2018a) for in silico expression analysis of genes retrieved within QNRs (Supplementary Table 5). There were 1,064 common genes with complete transcript information and valid results in both of the studies (Xue et al., 2008; Kumar et al., 2018a). In the peduncle of eight Seri/Bebax RILs contrasting to total stem WSCs at anthesis under rainfed environment, the average count of transcripts per million (tpm) ranged between 4.31 (log2 = 2.11; TraesCS6D02G392300) and 14151.17 (log2 = 13.79; TraesCS1B02G317700). In the latter experiment (Kumar et al., 2018a), the high-yielding drought-sensitive genotype WL-711 showed a range from 1.78 tpm (log2 = 0.83; TraesCS3A02G149500) to 161406.53 tpm (log2 = 17.30; TraesCS1B02G317700) in controlled environment and from 0.64 tpm (log2 = −0.64; TraesCS7A02G482400) to 153250.26 tpm (log2 = 17.23; TraesCS1B02G317700) in water stress environment. Likewise, the high-yielding drought-tolerant genotype C-306 carried a range between 1.30 tpm (log2 = 0.37; TraesCS1A02G317311) and 33444.93 tpm (log2 = 15.03; TraesCS5B02G496000) in controlled environment and 0.69 tpm (log2 = −0.54; TraesCS7A02G482400) to 32144.72 tpm (log2 = 14.97; TraesCS3A02G405700) in water stress environment. Additionally, a Genevestigator platform (Hruz et al., 2008)-based meta-transcriptomic analysis of 124 samples from various perturbation studies, including above two studies (control, water stress, heat, and heat + drought) suggested differential expression of all the overlapping genes (Supplementary Figure 1).
Gene Prioritization
To prioritize putative candidate genes, we implicated three heuristic approaches, viz. KGP, DEGP, and CNGP. The first two approaches prioritized 1,372 and 312 genes in this study. In the latter approach, 331 genes were prioritized from eight co-regulatory networks (Supplementary Figure 2). Genes equally prioritized by all three approaches were called high-priority candidate genes (HPCGs), whereas genes prioritized by DEGP and CNGP were called low-priority candidate genes (LPCGs). Thus, in this study, we prioritized 64 HPCGs and 109 LPCGs. Additionally, 51 overlapping genes were found in either coregulatory network and called specially prioritized genes (SPCG). Furthermore, seven and nine SCGPs were also identified as HPCGs and LPCGs. Finally, 208 putative candidate genes delimited into 64 QNRs are prioritized in this study, which can be critical to understanding the genetic architecture of stem WSCs accumulation in wheat under optimum and water-stressed environments (Supplementary Table 9). Furthermore, we identified nineteen causative stable QTNs for 24 prioritized genes (Supplementary Table 10).
Discussion
The Variabilities
Substantial phenotypic variability was reported for studied traits. The high-phenotypic and GxE variance suggested environmental influence on the traits studied; however, the values for h2 indicted the possibilities for selection for these traits in breeding programs. Careful early selection of fructans, TNSC, and WSC may be possible due to their mild heritability (h2 = 30-60%), but, due to low heritability (h2 < 30%), glucose, fructose, and sucrose will require multiple selection cycles. Additionally, the high GxE interaction suggests that varietal development programs must be environment specific for these traits. These findings agreed with previous studies (Ruuska et al., 2006; Zhang et al., 2014; del Pozo et al., 2016). As Yáñez et al. (2017) put it, WSCs showed a tradeoff with soil moisture. In our study, maximum 49, 40, 31, 74, 60, and 50% and 33, 37, 27, 54, 42, and 39% increases in glucose, fructose, sucrose, fructans, TNSC, and, WSC, respectively, were recorded under dry and rainfed conditions. The present study followed the general pattern of stem WSCs accumulation, i.e., drought-tolerant genotypes accumulate more stem WSCs in water stress. Indeed, genotypes that claim tolerance to heat stress, salinity stress, and a high degree of disease resistance also showed higher accumulation in water stress. However, drought-tolerant genotypes were more efficient in accumulating stem WSCs, which is likely due to the favorable conditions. Genotypes did not follow this pattern under irrigated trials. These results speculated the role of stem WSCs in drought stress and other stress conditions that must be further investigated.
Genotyping technique, marker coverage, and population structure are the determinating factors in GWAS, along with phenotypic variability. The present mapping panel was genotyped with Wheat Breeders’ Array (35K Axioms® Array), which is an improved version of Axiom® HD 820K wheat array and employed in previous studies (Alomari et al., 2017; Sheoran et al., 2019; Kumar et al., 2020). The number of polymorphic markers reported in the present study and their genome-wide coverage pattern agreed with the previous records (Berkman et al., 2013; Bajgain et al., 2015; Rahimi et al., 2019). As expected, roughly half of the total polymorphic SNPs mapped to B-genome followed by A (39%) and D (10%) genomes. For these polymorphic SNPs, genome-wide LD attenuated at 2 cM; however, for sub-genomes, the LD decayed at 2.8 cM (A-genome), 6.5 cM (B-genome), and 12.8 cM (D-genome). These values further presented the ample genetic diversity in our mapping panel. In addition, the LD decayed in the present study agreed with the previous studies (Ledesma-Ramírez et al., 2019; Hu et al., 2020).
The genetic population used in this study stratifies into seven distinct sub-populations (A to G). The genotypes clustered according to their pedigree or geographical origin. For instance, ten of the twenty members of the sub-population “B” had Pastor in their pedigree, and few others have “Bobwhite.” Interestingly, Pastor is the result of PFAU/SERI-82//(SIB)BOBWHITE (wheatpedigree.net). Likewise, in sub-population “D,” we can find a cluster of genotypes that are either collected from the Gujarat, Rajasthan, or Madhya Pradesh (MP) states of India or has been selected from the idigenous collection originated from these states. The substantial admixture further suggests that this population results from the extensive modern breeding program. On the other hand, most indigenous collections and some varieties (e.g., Sonalika and HUW-598) were admixture less. These results indicate the broad genetic base of our study material and suitability for a successful GWAS.
Comparison With Previous Studies
We have identified a substantial number of 382 QTNs delimited into 112 QNRs for the six targeted traits; many of which are endorsed by previous GWAS and bi-parental QTL mapping on stem WSCs. Forty-seven QNRs from the present GWAS overlapped with previously reported loci and genes for stem WSCs. These loci are likely to be the part of the same genomic region or loci (Supplementary Table 4). Presumably, our twenty novel QNRs carry 51 novel QTNs. Compared with previous studies, there were three intergenic QTNs (AX-94964616, AX-95114107, and AX-95169219), which Fu et al. (2020) also reported for stem WSC. In addition to this, five SNPs (AX-94428937, AX-94823847, AX-94474297, AX-110440726, and Ku_c56370_1155) from Fu et al. (2020) colocalized with our QTNs AX-94910470, AX-94708698, AX-94507107, and AX-94424668 on the same genes, viz. TraesCS2B02G556600, TraesCS6A02G407700, TraesCS2A02G562500, and TraesCS3A02G301500. Likewise, forty other QNRs overlapped the SNP markers identified by Fu et al. (2020). Two QTNs from qWSC-1A.1 (1A:54.04 cM), namely AX-94583145 and AX-95137931 were just 0.4 cM and 0.54 cM away from the SNPs previously reported for stem WSCs (Dong et al., 2016; Ovenden et al., 2017b).
Seven of the previously cloned genes of WSC importance were found within the vicinity of QTNs identified in this GWAS. A sugar transporter gene, TaSUT (2A:733.56 Mbp) (Ahmed et al., 2018), was identified ca. 15 Mbp downstream from the QTN AX-94416062 of qWSC-2A.3, which was associated with sucrose content in plants under optimal conditions. Transcriptional factor TaMYB13-1 (Xue et al., 2011) (1A:746.63 Mbp), which upregulates sucrose concentration and activates fructosyl transferase genes, was found within the vicinity of QTN AX-94838498 (3A: 743.75 Mbp) from qWSC-3A.12- controlling fructose content. QTN AX-94619716 (4A: 739.76 Mbp) from qWSC-4A.2 was associated with glucose and sucrose content under water-stressed condition and had 6-SFT (4A:739.06 Mbp), a gene responsible for β-2,6-linked fructans synthesis from sucrose (Kawakami and Yoshida, 2005), in its proximity. 1-FEH gene hydrolyses β-2,1-linkages of fructans to break it into simpler sugars. It has been found directly correlated with sucrose, fructans, and WSC content in parts of the wheat stem, especially under water-stressed conditions (Van Den Ende et al., 2003; Yang et al., 2004). In our study, this gene colocalized with QTNs AX-94556117 and AX-94543047 of region qWSC-6A.2 (6A:26.51 cM) controlling glucose, sucrose, fructans, TNSC, and WSC in water-stressed plants.
Furthermore, gene TraesCS4A02G485400 (6-SFT) found in the neighborhood (0.69 Mbp) of QTN AX-94619716 (4A:739763001 bp). Earlier, Huynh et al. (2008) identified a potential QTL, namely QGfc.aww-7A.1 (7A:2.2-8.2 cM), for grain fructans concentration in Berkut × Krichauff double-haploid lines, from which they further cloned a cluster of four fructans biosynthesis genes (Huynh et al., 2012). We BLAST the sequences of these genes against IWGSC RefSeq1.1 and retrieved two genes with 100% similarity, viz. TraesCS7A02G009200 (7A:4014510-4016977 bp) and TraesCS7A02G009800 (7A:4430034-4430058 bp), which they designated as Ta1-FFT and Ta6-SFT, respectively. These two genes were within the 1-Mbp window of QTN AX-95231996 (7A:3982822:4627547 bp) associated with fructose content under rainfed conditions. The QTN region, namely qWSC-7A.2 (7A:29.86-30.66 cM), to which these two genes assigned in this study, seems to be the same region. In the findings of Huynh et al. (2012), we found traces of inheritance of this QNR with favorable alleles from the CYMMIT line “Pastor.” Looking back into the pedigree information (Supplementary Table 1), at least 20 genotypes were directly developed by “Pastor” as one of the parents, and few others had it in their lineage. We closely correlated the pedigree information with the favorable allele analysis, which indicated that the genotypes with early CIMMYT breeding lines in their background accumulated more suitable combinations of favorable alleles. Thus, presumably, favorable alleles of stem WSCs are inherited by the CIMMYT breeding lines across the globe, for which inadvertent selections continued. Earlier, Rebetzke et al. (2009) also marked an increasing trend in the Western Australia wheat breeding program with the involvement of the CIMMYT lines. This finding needs a further detailed discussion with validatory reports that would be beyond the objectives of the present article.
Complex Genetic Architecture Regulates the Accumulation of Soluble Sugars
We observed that all the QTN regions were enriched with the genes, which may affect the carbohydrate metabolism and expression of studied traits in a C3 plant, depending upon the growth conditions. Furthermore, IPR and GO term enrichment analysis indicated that the genes with the molecular function of oxidation reduction, protein phosphorylation, and carrying genetic information overrepresented the defined QNRs (Figure 5). More specifically, the genes overlapping significant QTNs were located in the nucleus (100), chloroplast (30), mitochondria (12), cytoplasm (6), extracellular space (25), and various cellular membranes (Supplementary Table 8). Thus, they were likely to be responsible for the stem WSCs variation through other metabolic activities such as light-harvesting complex, electron transport system, sugar and ion transport, respiration, and many others. For instance, the QTN AX-94710575 associated with fructans and TNSC variation in water-stress condition tagged Os01g0227100 orthologous chlorophyll-b reductase gene (TraesCS3A02G151900). The chlorophyll-b reductase initiates the process of reduction of chlorophyll-b to 7-hydroxymethyl chlorophyll-a. The downregulation or absence of chlorophyll-b reductase genes in Arabidopsis and Oryza sativa has been associated with stay-green traits (Sato et al., 2009; Nakajima et al., 2012). Thus, this gene is reasonably associated with a more extended accumulation period for stem WSCs. A lower transcript level has been reported for TraesCS3A02G151900 in the drought-tolerant C-306 genotype in contrast to the drought-sensitive WL-711 genotype under water-stress conditions (Kumar et al., 2018a). Likewise, this gene showed higher expression in RILs with high WSC content in the peduncle under rainfed conditions than RILs with low WSC content (Xue et al., 2008). A PsbQ gene (TraesCS2B02G395700) overlapped the QTN AX-94491160 associated with fructose content under severe drought-stress environment E04. The PsbQ protein is a part of the photosystem-II oxygen-evolving complex and is required for the calcium ion retention that is crucial for the stability of photosystem-II (Sasi et al., 2018). The previous study on Arabidopsis has suggested that the long-term water stress markedly decreases the PsbQ protein, thereby affecting the functionality of photosystem and photosynthesis. In our research, the E04 environment represents the long-term drought condition. The TraesCS3A02G425500, a sucrose synthase/phosphatase-encoding gene, was tagged by a stable QTN AX-94501648 responsible for TNSC variation in our GWAS panel under severe drought stress. Increased expression of sucrose synthase genes is negatively correlated with total WSC content in the bread wheat peduncle under water-stressed conditions (Xue et al., 2008). A sugar transporter gene (TraesCS4B02G371700) overlapped the QTN AX-94619716 associated with glucose and sucrose content under a rainfed environment (E06). As revealed by Knetminer, the TraesCS4B02G371700 has a direct physical relationship with the sucrose transporter gene (SUT4) that cooccurs with carbohydrate and sucrose content in bread wheat. Likewise, there were a total of 72 overlapping genes that had a direct or indirect connection with either or all the studied traits in Knetminers. About 26 stable QTNs were likely to be associated with 24 genes prioritized for stem WSCs metabolism in the present study (Supplementary Table 10). These genes were involved in a wide array of molecular functions and biological processes in wheat. For instance, AX-94865722 (2A:765292309 bp), a stable QTN associated with glucose and sucrose content under severe water-stress environment (E04 and DT), tagged two NAC-like genes (TraesCS2A02G566000 and TraesCS2A02G566100). Two stable QTNs (AX-94995102 and AX-94495814) for fructans and TNSC content under severe water stress were found causative from a MYB-domain protein TraesCS1B02G316200. Earlier, Kooiker et al. (2013) demonstrated the role of MYB-domain protein in enhanced accumulation of fructans in bread wheat. The NAC genes affect carbohydrate metabolism, phytohormones, and transmembrane transporters-related genes, hence crucial for drought tolerance and reducing grain yield losses (Chung et al., 2018). Likewise, a sugar/inositol transporter gene (TraesCS4B02G371700) was tagged under optimum water conditions for total stem WSC content with a stable QTN AX-94546364 (4B:657172059 bp). The stable and novel QTN from novel QNR qWSC-3A.7, AX-94501648 (3A:667793607 bp), was significant for TNSC in severe drought, tagged Os01g0919400-like sucrose phosphate synthase gene TraesCS3A02G425500. Furthermore, the co-regulatory network (CRNs) study based on the publicly available Affymetrix data (GSE9767 and GSE87325) indicated that a complex network of many genes works behind the veil for regulating the accumulation of stem WSCs. We reported that the number and type of participatory genes significantly change with the genetic background and growth conditions at an individual level. The twelve CRNs allowed us to better narrow down to a few hub genes (301). No hub gene was reported common across the twelve CRNs. However, there were hub genes common between different genetic backgrounds. The functional enrichment analysis followed by functional group-based networking (Figures 6A,B) of hub genes left us with the information on the interaction of various functional groups interacting with major fructans metabolic genes, which we reported in QNRs qWSC-4A.2 and qWSC-7A.2, and other sugar metabolic genes, in a complex and coordinated network. Figure 6A indicates various biological functions, such as chlorophyll biosynthesis, photosynthesis, and respiration, correlate with the metabolic process of WSCs.
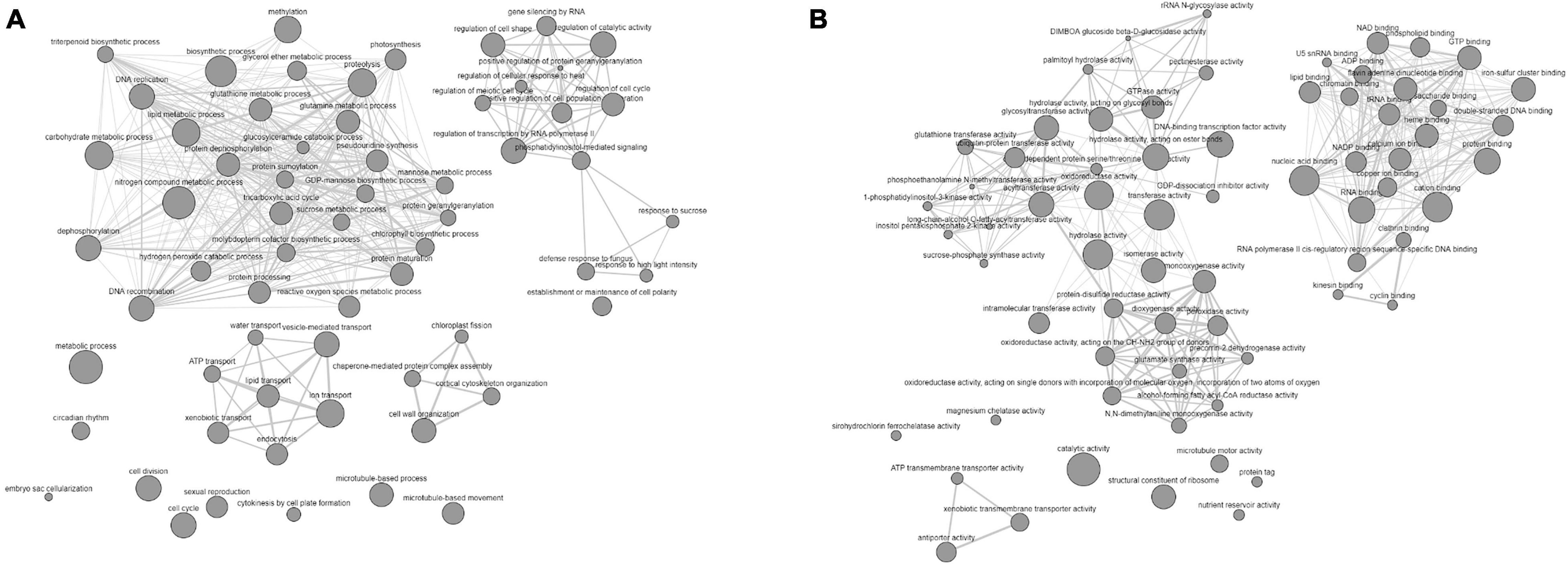
Figure 6. GO enrichment-based biological (A) and molecular (B) networks of hub genes indicating the involvement of various biological and functional groups in the accumulation of WSCs in the wheat stem under differential water conditions.
Prospects for Stem Water-Soluble Carbohydrates Improvement in the Present Study
Consistent with many previous studies, our results demonstrate several advantages, such as a more significant number of genotypes with substantial genetic variability at phenotypic and genetic levels, consideration of multiple environments pertaining to different water regimes, and consideration of individual components of water-soluble carbohydrates as a determining factor in final stem WSCs content. However, the present investigation carried the well-defined limitation of GWAS, which includes the inability to detect rare alleles and epistasis. In the current research, we have identified twenty novel QNRs responsible for the expression of studied traits.
With the keen observation of the results, we realized that the accumulation of stem WSCs is a low to moderately heritable and environmentally influenced trait. The complexity in improving this trait lies with the higher degree of genotype x environment (GxE) and QTN x environment (QnxE) interactions. Stability of QNRs over the QTNs across the environments indicated that the phenotypic expression of the studied traits is limited to the fewer genomic regions with a substantial number of genes bound within a single and complex regulatory network with a fewer number of hub genes that further modulate with the genetic background of an individual and external stimuli. Within 1.5Kbp upstream from the starting coordinates (bp) of the possible candidate and overlapping genes, we identified 130 different cis-regulatory elements (CREs) with the help of PlantCARE (Lescot, 2002). These CREs were responsive to various hormones (ABA, gibberellin, auxins), lux, temperature, water, and circadian rhythm, including those having tissue-specific expression (Supplementary Figure 3). Probably, this is why we detected a lesser number of stable QTNs and traces of environmental pleiotropy. Furthermore, the strong correlation among the studied traits is likely due to the stacking of genes rather than being controlled by a single gene.
Furthermore, the present study identified two genomic regions, qWSC-4A.2 and qWSC-7A.2, with the clusters of fructans metabolic genes, two of which were the center of gene networks; however, their expression was subjected to genotype and water conditions. The genevestigator based in silico expression analysis of these genes showed differential expression, depending upon the pertubation (Supplementary Figure 4). Furthermore, these genes were subjected to phylogeny, evolutionary, and protein-modeling studies (Supplementary Method 2) for mining further opportunities. We found that these clusters carried multiple orthologs of fructosyl transferase genes translating into theoretically stable (except TraesCS4A02G485000 and TraesCS7A02G009100) but structurally different proteins. The multiple sequence alignment showed many conserved amino acids across the fructans metabolic genes (Figure 7A). As put forward by Huynh et al. (2012), the phylogenetic analysis (Figure 7B) in this study indicated that fructosyl transferase genes (SST/SFT/FFT) have evolved from exohydrolase/invertase genes. Of the three different fructosyl genes, SFT type of genes is likely to evolve first, followed by SST and FFT. With MEME suite analysis, we identified three significant motifs of the glycosyl hydrolase family (INV_N, Glyco hydro 32N, and Glyco Hydro 32C) conserved across the fructans metabolic genes (Figure 7B). However, multiple sequence alignment of these motifs indicated that the substitutions of amino acids at few positions are likely to bring structural and functional changes. The Glyco hydro 32N and INV motifs are likely to carry a more significant number of conserved sites than the Glyco hydro 32C motif (Supplementary Figure 5). These substitutions are presumably located at active sites, bringing the difference in the functionality of FEH-, SFT-, SST-, and FFT-type genes despite the similar protein structure. Previously, Lammens et al. (2012) demonstrated that the major differences between functionality of various fructans metabolic genes lay within the few amino acid substitutions at active sites. Therefore, one type of the fructans metabolic gene can be converted to another type with the help of advanced gene-editing techniques like CRISPR to bring different fructans genes to an optimum combination that improves the accumulation and remobilization of stem WSCs. Earlier, in tobacco, Bie et al. (2012) demonstrated improved fructans biosynthesis and abiotic stress tolerance with the combinational transformation of wheat’s 1-FFT, 6-SFT, and 1-SST genes. Additionally, 3D protein models (Supplementary Figure 6) were predicted with the deep learning methods (Wang and Xu, 2013; Ma et al., 2015) as applied in the RaptorX (Källberg et al., 2012). The validatory reports on the 3D models generated with Structure Validation Server (SAVESv6.0) indicated the high reliability of the predicated models (Supplementary Table 11).
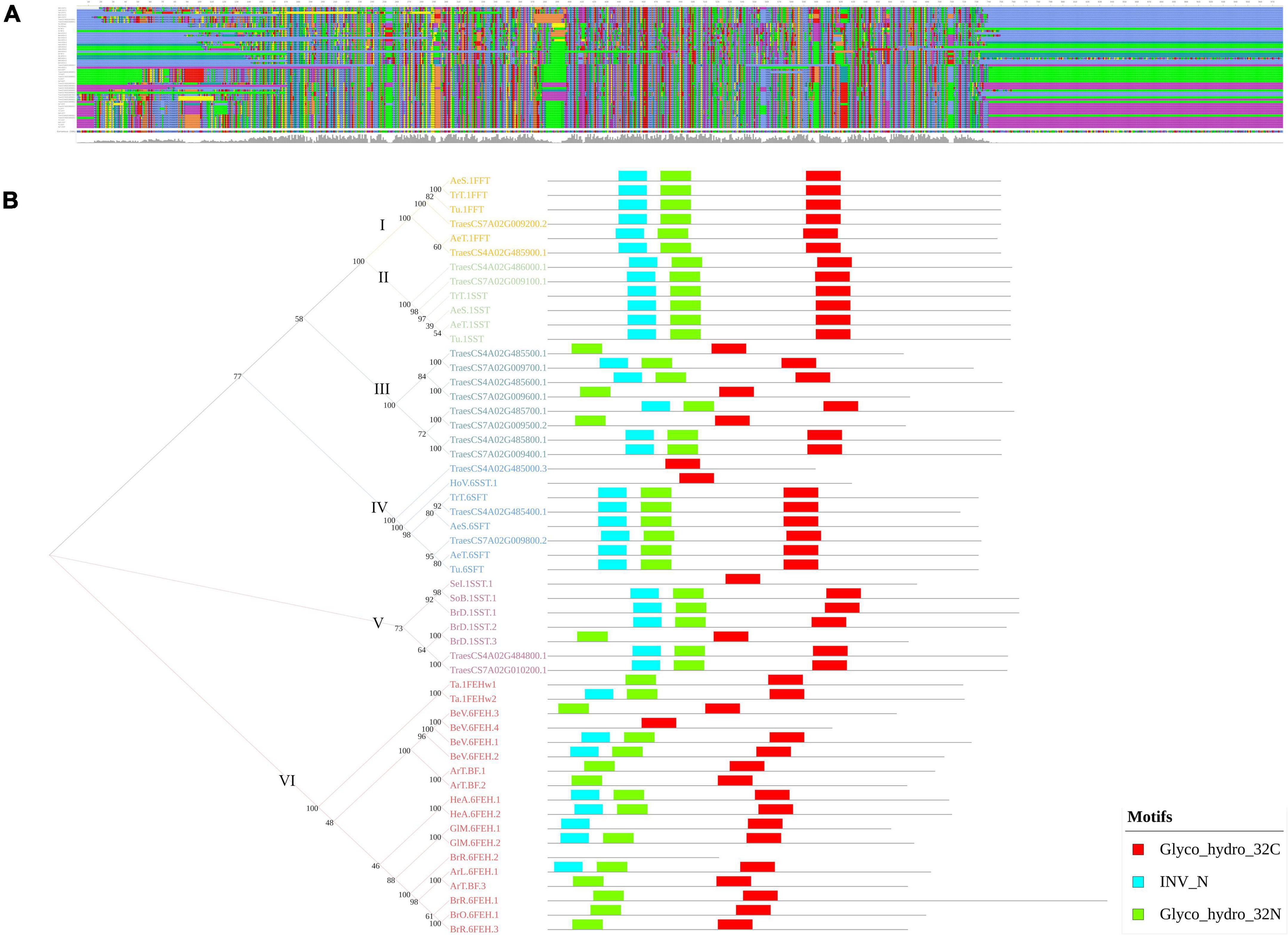
Figure 7. The multiple sequence alignment (A) and phylogram with a comparison between motif structures of the fructans metabolic genes of Triticum aestivum and model plant species (B). In (B), the Arabic numbers indicate the bootstrap values, whereas the Roman numbers indicate the cluster number.
Conclusion
Finally, based on our results, we conclude that the final content of total WSC in stem depends on the initial partitioning of the individual sugars, viz. glucose, fructose, and sucrose and their conversion to fructans. Therefore, the accumulation of stem WSCs in wheat shall depend on the machinery affecting the metabolism of these sugars, which further strengthens with the identification of candidate genes involved in electron transport systems, photosynthesis, respiration, and signaling. Unfortunately, due to the presence of CREs, these genes are quite sensitive to external stimuli and, accordingly, changes their expression pattern. In the present study, we identified novel QTNs and QNRs, showing the positive pyramiding effect of a few representative QTNs on total stem WSCs. But, bringing improvement in total stem WSC content with the pyramiding of this large number of QTN and candidate genes further subjected to large GxE and QnxE interaction remains a big challenge, even with the advanced genomic selection algorithms. Fructan metabolic genes identified on chromosomes 4A and 7A can be used to enhance the fructans biosynthesis and remobilization for improving the drought tolerance. Thus, alongside identifying novel QTNs and QNRs, and narrowing the knowledge of the molecular and genetic basis of stem WSCs accumulation in different water conditions, the present investigation indicates vast opportunities to study CREs as a breeding target for environmentally influenced traits and gene editing in improving fructans metabolism.
Data Availability Statement
The datasets presented in this study can be found in online repositories. The names of the repository/repositories and accession number(s) can be found in the article/Supplementary Material.
Author Contributions
SS, AG, and YJ designed the study. SS, RT, and AG managed the trials at ICAR-IIWBR, Karnal. AG, YJ, VS, and DeK managed the trials at CCS HAU, Hisar. AG carried out phenotyping at Karnal and Hisar. SN guided in wet lab analysis. JS provided the FTNIR spectroscope facility. AG, SS, SJ, MI, and UA conducted GWAS. AG and SS conducted post GWAS analysis. AG and SS drafted the manuscript. YJ, SS, VS, GS, DK, GPS, and AR provided overall guidance and finalized the edited manuscript. All authors read and approved the final version of the manuscript.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s Note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Acknowledgments
The authors acknowledge ICAR (Indian Council of Agricultural Research)-sponsored project “Incentivizing Research in Agriculture” for funding and ICAR-Indian Institute of Wheat & Barley Research Institute, Karnal for providing infrastructure facilities. The authors are thankful to the Indian Council of Agricultural Research, Ministry of Agriculture and Farmers’ Welfare, Govt. of India for Advanced Super Computing Hub for Omics Knowledge in Agriculture (ASHOKA) facility and CABin Project at ICAR-IASRI, New Delhi, India.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fpls.2022.825687/full#supplementary-material
References
Ahmed, S. A. S., Zhang, J., Ma, W., and Dell, B. (2018). Contributions of TaSUTs to grain weight in wheat under drought. Plant Mol. Biol. 98, 333–347. doi: 10.1007/s11103-018-0782-1
Allen, A. M., Winfield, M. O., Burridge, A. J., Downie, R. C., Benbow, H. R., Barker, G. L. A., et al. (2017). Characterization of a Wheat Breeders’ Array suitable for high-throughput SNP genotyping of global accessions of hexaploid bread wheat (Triticum aestivum). Plant Biotechnol. J. 15, 390–401. doi: 10.1111/pbi.12635
Alomari, D. Z., Eggert, K., Von Wirén, N., Pillen, K., and Röder, M. S. (2017). Genome-wide association study of calcium accumulation in grains of European wheat cultivars. Front. Plant Sci. 8:1797. doi: 10.3389/fpls.2017.01797
Bajgain, P., Rouse, M. N., Bulli, P., Bhavani, S., Gordon, T., Wanyera, R., et al. (2015). Association mapping of North American spring wheat breeding germplasm reveals loci conferring resistance to Ug99 and other African stem rust races. BMC Plant Biol. 15:249. doi: 10.1186/s12870-015-0628-9
Berkman, P. J., Visendi, P., Lee, H. C., Stiller, J., Manoli, S., Lorenc, M. T., et al. (2013). Dispersion and domestication shaped the genome of bread wheat. Plant Biotechnol. J. 11, 564–571. doi: 10.1111/pbi.12044
Bie, X., Wang, K., She, M., Du, L., Zhang, S., Li, J., et al. (2012). Combinational transformation of three wheat genes encoding fructan biosynthesis enzymes confers increased fructan content and tolerance to abiotic stresses in tobacco. Plant Cell Rep. 31, 2229–2238. doi: 10.1007/S00299-012-1332-Y
Blum, A. (2011). Plant Breeding for Water-Limited Environments. New York, NY: Springer doi: 10.1007/978-1-4419-7491-4
Chung, P. J., Jung, H., Choi, Y., and Do, Kim, J. K. (2018). Genome-wide analyses of direct target genes of four rice NAC-domain transcription factors involved in drought tolerance. BMC Genomics 19:40. doi: 10.1186/s12864-017-4367-1
del Pozo, A., Yáñez, A., Matus, I. A., Tapia, G., Castillo, D., Sanchez-Jardón, L., et al. (2016). Physiological Traits Associated with Wheat Yield Potential and Performance under Water-Stress in a Mediterranean Environment. Front. Plant Sci. 7:987. doi: 10.3389/fpls.2016.00987
Dong, Y., Liu, J., Zhang, Y. Y., Geng, H., Rasheed, A., Xiao, Y., et al. (2016). Genome-wide association of stem water soluble carbohydrates in bread wheat. PLoS One 11:e0164293. doi: 10.1371/journal.pone.0164293
Dreccer, M. F., van Herwaarden, A. F., and Chapman, S. C. (2009). Grain number and grain weight in wheat lines contrasting for stem water soluble carbohydrate concentration. F. Crop. Res. 112, 43–54. doi: 10.1016/j.fcr.2009.02.006
Earl, D. A., and vonHoldt, B. M. (2012). STRUCTURE HARVESTER: a website and program for visualizing STRUCTURE output and implementing the Evanno method. Conserv. Genet. Res. 4, 359–361. doi: 10.1007/s12686-011-9548-7
Evanno, G., Regnaut, S., and Goudet, J. (2005). Detecting the number of clusters of individuals using the software STRUCTURE: a simulation study. Mol. Ecol. 14, 2611–2620. doi: 10.1111/j.1365-294X.2005.02553.x
Fu, L., Wu, J., Yang, S., Jin, Y., Liu, J., Yang, M., et al. (2020). Genome-wide association analysis of stem water-soluble carbohydrate content in bread wheat. Theor. Appl. Genet. 133, 2897-2914. doi: 10.1007/s00122-020-03640-x
Gao, F., Ma, D., Yin, G., Rasheed, A., Dong, Y., Xiao, Y., et al. (2017). Genetic progress in grain yield and physiological traits in Chinese wheat cultivars of southern Yellow and Huai Valley since 1950. Crop Sci. 57, 760–773. doi: 10.2135/cropsci2016.05.0362
Goggin, D. E., and Setter, T. L. (2004). Fructosyltransferase activity and fructan accumulation during development in wheat exposed to terminal drought. Funct. Plant Biol. 31, 11–21. doi: 10.1071/FP03123
Guerra, F. P., Yáñez, A., Matus, I., and Del Pozo, A. (2021). Genome-wide association of stem carbohydrate accumulation and remobilization during grain growth in bread wheat (Triticum aestivum l.) in mediterranean environments. Plants 10, 539. doi: 10.3390/plants10030539
Hruz, T., Laule, O., Szabo, G., Wessendorp, F., Bleuler, S., Oertle, L., et al. (2008). Genevestigator V3: a Reference Expression Database for the Meta-Analysis of Transcriptomes. Adv. Bioinform. 2008:420747. doi: 10.1155/2008/420747
Hu, W., Gao, D., Wu, H., Liu, J., Zhang, C., Wang, J., et al. (2020). Genome-wide association mapping revealed syntenic loci QFhb-4AL and QFhb-5DL for Fusarium head blight resistance in common wheat (Triticum aestivum L.). BMC Plant Biol. 20:29. doi: 10.1186/s12870-019-2177-0
Hubisz, M. J., Falush, D., Stephens, M., and Pritchard, J. K. (2009). Inferring weak population structure with the assistance of sample group information. Mol. Ecol. Resour. 9, 1322–1332. doi: 10.1111/j.1755-0998.2009.02591.x
Huynh, B. L., Mather, D. E., Schreiber, A. W., Toubia, J., Baumann, U., Shoaei, Z., et al. (2012). Clusters of genes encoding fructan biosynthesizing enzymes in wheat and barley. Plant Mol. Biol. 80, 299–314. doi: 10.1007/s11103-012-9949-3
Huynh, B. L., Wallwork, H., Stangoulis, J. C. R., Graham, R. D., Willsmore, K. L., Olson, S., et al. (2008). Quantitative trait loci for grain fructan concentration in wheat (Triticum aestivum L.). Theor. Appl. Genet. 117, 701–709. doi: 10.1007/s00122-008-0811-6
Källberg, M., Wang, H., Wang, S., Peng, J., Wang, Z., Lu, H., et al. (2012). Template-based protein structure modeling using the RaptorX web server. Nat. Protoc. 7, 1511–1522.
Kawakami, A., and Yoshida, M. (2005). Fructan:fructan 1-fructosyltransferase, a key enzyme for biosynthesis of graminan oligomers in hardened wheat. Planta 223, 90–104. doi: 10.1007/s00425-005-0054-6
Kawakami, A., Yoshida, M., and Van Den Ende, W. (2005). Molecular cloning and functional analysis of a novel 6&1-FEH from wheat (Triticum aestivum L.) preferentially degrading small graminans like bifurcose. Gene 358, 93–101. doi: 10.1016/j.gene.2005.05.029
Kooiker, M., Drenth, J., Glassop, D., McIntyre, C. L., and Xue, G. P. (2013). TaMYB13-1, a R2R3 MYB transcription factor, regulates the fructan synthetic pathway and contributes to enhanced fructan accumulation in bread wheat. J. Exp. Bot. 64, 3681–3696.
Kumar, D., Chhokar, V., Sheoran, S., Singh, R., Sharma, P., Jaiswal, S., et al. (2020). Characterization of genetic diversity and population structure in wheat using array based SNP markers. Mol. Biol. Rep. 47, 293–306. doi: 10.1007/s11033-019-05132-8
Kumar, S., Stecher, G., Li, M., Knyaz, C., and Tamura, K. (2018b). MEGA X: molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 35, 1547–1549. doi: 10.1093/molbev/msy096
Kumar, J., Gunapati, S., Kianian, S. F., and Singh, S. P. (2018a). Comparative analysis of transcriptome in two wheat genotypes with contrasting levels of drought tolerance. Protoplasma 255, 1487–1504. doi: 10.1007/s00709-018-1237-x
Kuznetsova, A., Brockhoff, P. B., and Christensen, R. H. B. (2017). lmerTest Package: tests in Linear Mixed Effects Models. J. Stat. Softw. 82, 1–26. doi: 10.18637/jss.v082.i13
Lammens, W., Le Roy, K., Yuan, S., Vergauwen, R., Rabijns, A., Van Laere, A., et al. (2012). Crystal structure of 6-SST/6-SFT from Pachysandra terminalis, a plant fructan biosynthesizing enzyme in complex with its acceptor substrate 6-kestose. Plant J. 70, 205–219. doi: 10.1111/j.1365-313X.2011.04858.x
Ledesma-Ramírez, L., Solís-Moya, E., Iturriaga, G., Sehgal, D., Reyes-Valdes, M. H., Montero-Tavera, V., et al. (2019). GWAS to Identify Genetic Loci for Resistance to Yellow Rust in Wheat Pre-Breeding Lines Derived From Diverse Exotic Crosses. Front. Plant Sci. 10:1390. doi: 10.3389/fpls.2019.01390
Lescot, M. (2002). PlantCARE, a database of plant cis-acting regulatory elements and a portal to tools for in silico analysis of promoter sequences. Nucl. Acids Res. 30, 325–327. doi: 10.1093/nar/30.1.325
Li, M., Liu, Y., Ma, J., Zhang, P., Wang, C., Su, J., et al. (2020). Genetic dissection of stem WSC accumulation and remobilization in wheat (Triticum aestivum L.) under terminal drought stress. BMC Genet. 21:50. doi: 10.1186/s12863-020-00855-1
Li, W., Zhang, B., Li, R., Chang, X., and Jing, R. (2015). Favorable alleles for Stem water-soluble carbohydrates identified by association analysis contribute to grain weight under drought stress conditions in wheat. PLoS One 10:e0119438. doi: 10.1371/journal.pone.0119438
Liu, K., and Muse, S. V. (2005). PowerMaker: an integrated analysis environment for genetic maker analysis. Bioinformatics 21, 2128–2129. doi: 10.1093/bioinformatics/bti282
Livingston, D. P., Hincha, D. K., and Heyer, A. G. (2009). Fructan and its relationship to abiotic stress tolerance in plants. Cell. Mol. Life Sci. 66, 2007–2023. doi: 10.1007/s00018-009-0002-x
Livingston, D. P., Premakumar, R., and Tallury, S. P. (2006). Carbohydrate partitioning between upper and lower regions of the crown in oat and rye during cold acclimation and freezing. Cryobiology 52, 200–208. doi: 10.1016/j.cryobiol.2005.11.001
Ma, J., Wang, S., Wang, Z., and Xu, J. (2015). Protein contact prediction by integrating joint evolutionary coupling analysis and supervised learning. Bioinformatics 31, 3506–3513.
Mandal, C., Mandal, D. K., Bhattacharyya, T., Sarkar, D., Pal, D. K., Prasad, J., et al. (2014). Revisiting agro-ecological sub-regions of India–a case study of two major food production zones. Curr. Sci. 107, 1519–1536.
McIntyre, C. L., Casu, R. E., Rattey, A., Dreccer, M. F., Kam, J. W., Van Herwaarden, A. F., et al. (2011). Linked gene networks involved in nitrogen and carbon metabolism and levels of water-soluble carbohydrate accumulation in wheat stems. Funct. Integr. Genomics 11, 585–597. doi: 10.1007/s10142-011-0232-5
McIntyre, C. L., Seung, D., Casu, R. E., Rebetzke, G. J., Shorter, R., and Xue, G. P. (2012). Genotypic variation in the accumulation of water soluble carbohydrates in wheat. Funct. Plant Biol. 39, 560–568. doi: 10.1071/FP12077
Murray, M. G., and Thompson, W. F. (1980). Rapid isolation of high molecular weight plant DNA. Nucl. Acids Res. 8, 4321–4326. doi: 10.1093/nar/8.19.4321
Nakajima, S., Ito, H., Tanaka, R., and Tanaka, A. (2012). Chlorophyll b reductase plays an essential role in maturation and storability of Arabidopsis seeds. Plant Physiol. 160, 261–273. doi: 10.1104/pp.112.196881
Ovenden, B., Milgate, A., Lisle, C., Wade, L. J., Rebetzke, G. J., and Holland, J. B. (2017a). Selection for water-soluble carbohydrate accumulation and investigation of genetic × environment interactions in an elite wheat breeding population. Theor. Appl. Genet. 130, 2445–2461. doi: 10.1007/s00122-017-2969-2
Ovenden, B., Milgate, A., Wade, L. J., Rebetzke, G. J., and Holland, J. B. (2017b). Genome-Wide Associations for Water-Soluble Carbohydrate Concentration and Relative Maturity in Wheat Using SNP and DArT Marker Arrays. G3. 7, 2821–2830. doi: 10.1534/g3.117.039842
Peshev, D., Vergauwen, R., Moglia, A., Hideg, É, and Van Den Ende, W. (2013). Towards understanding vacuolar antioxidant mechanisms: a role for fructans? J. Exp. Bot. 64, 1025–1038. doi: 10.1093/jxb/ers377
Purcell, S., Neale, B., Todd-Brown, K., Thomas, L., Ferreira, M. A. R., Bender, D., et al. (2007). PLINK: a tool set for whole-genome association and population-based linkage analyses. Am. J. Hum. Genet. 81, 559–575. doi: 10.1086/519795
Rahimi, Y., Bihamta, M. R., Taleei, A., Alipour, H., and Ingvarsson, P. K. (2019). Genome-wide association study of agronomic traits in bread wheat reveals novel putative alleles for future breeding programs. BMC Plant Biol. 19:541. doi: 10.1186/s12870-019-2165-4
Ram, K., Kumar, R., Ta, S., and Munjal, R. (2018). Stem Reserve Mobilization in Relation to Yield under Different Drought and High Temperature Stress Conditions in Wheat (Triticum aestivum L.) Genotypes. Int. J. Curr. Microbiol. Appl. Sci 7, 3695–3704. doi: 10.20546/ijcmas.2018.704.415
Rebetzke, G. J., Chapman, S. C., Lynne Mcintyre, C., Richards, R. A., Condon, A. G., Watt, M., et al. (2009). “Grain Yield Improvement in Water-Limited Environments,” in Wheat Science and Trade, ed B. F. Carver (New Jersey: Wiley-Blackwell), 215–249. doi: 10.1002/9780813818832.ch11
Rebetzke, G. J., Herwaarden, A. F., van, Jenkins, C., Ruuska, S., Tabe, L., et al. (2007). “Genetic Control of Water-Soluble Carbohydrate Reserves in Bread Wheat,” in Wheat Production in Stressed Environments, eds H. T. Buck, J. E. Nisi, and N. Salomón (Netherlands: Springer), 349–356. doi: 10.1007/1-4020-5497-1-43
Rebetzke, G. J., Van Herwaarden, A. F., Jenkins, C., Weiss, M., Lewis, D., Ruuska, S., et al. (2008). Quantitative trait loci for water-soluble carbohydrates and associations with agronomic traits in wheat. Aust. J. Agric. Res. 59, 891–905. doi: 10.1071/AR08067
Ruuska, S. A., Rebetzke, G. J., Van Herwaarden, A. F., Richards, R. A., Fettell, N. A., Tabe, L., et al. (2006). Genotypic variation in water-soluble carbohydrate accumulation in wheat. Funct. Plant Biol. 33, 799-809. doi: 10.1071/fp06062
Sasi, S., Venkatesh, J., Daneshi, R. F., and Gururani, M. A. (2018). Photosystem ii extrinsic proteins and their putative role in abiotic stress tolerance in higher plants. Plants 7:100. doi: 10.3390/plants7040100
Sato, Y., Morita, R., Katsuma, S., Nishimura, M., Tanaka, A., and Kusaba, M. (2009). Two short-chain dehydrogenase/reductases, NON-YELLOW COLORING 1 and NYC1-LIKE, are required for chlorophyll b and light-harvesting complex II degradation during senescence in rice. Plant J. 57, 120–131. doi: 10.1111/j.1365-313X.2008.03670.x
Schnyder, H. (1993). The role of carbohydrate storage and redistribution in the source-sink relations of wheat and barley during grain filling — a review. New Phytol. 123, 233–245. doi: 10.1111/j.1469-8137.1993.tb03731.x
Sharma, D., Singh, R., Rane, J., Gupta, V. K., Mamrutha, H. M., and Tiwari, R. (2016). Mapping quantitative trait loci associated with grain filling duration and grain number under terminal heat stress in bread wheat (Triticum aestivum L.). Plant Breed. 135, 538–545. doi: 10.1111/pbr.12405
Shearman, V. J., Sylvester-Bradley, R., Scott, R. K., and Foulkes, M. J. (2005). Physiological processes associated with wheat yield progress in the UK. Crop Sci. 45, 175–185. doi: 10.2135/cropsci2005.0175
Sheoran, S., Jaiswal, S., Kumar, D. D. D., Raghav, N., Sharma, R., Pawar, S., et al. (2019). Uncovering Genomic Regions Associated With 36 Agro-Morphological Traits in Indian Spring Wheat Using GWAS. Front. Plant Sci. 10:527. doi: 10.3389/fpls.2019.00527
Snape, J. W., Foulkes, M. J., Simmonds, J., Leverington, M., Fish, L. J., Wang, Y., et al. (2007). Dissecting gene x environmental effects on wheat yields via QTL and physiological analysis. Euphytica 154, 401–408. doi: 10.1007/s10681-006-9208-2
Supek, F., Bošnjak, M., Škunca, N., and Šmuc, T. (2011). Revigo summarizes and visualizes long lists of gene ontology terms. PLoS One 6:e21800. doi: 10.1371/journal.pone.0021800
Tang, Y., Liu, X., Wang, J., Li, M., Wang, Q., Tian, F., et al. (2016). GAPIT version 2: an enhanced integrated tool for genomic association and prediction. Plant Genome 9:plantgenome2015-11. doi: 10.3835/plantgenome2015.11.0120
Tricker, P. J., Elhabti, A., Schmidt, J., and Fleury, D. (2018). The physiological and genetic basis of combined drought and heat tolerance in wheat. J. Exp. Bot. 69, 3195–3210. doi: 10.1093/jxb/ery081
Valluru, R., and Van Den Ende, W. (2008). Plant fructans in stress environments: emerging concepts and future prospects. J. Exp. Bot. 59, 2905–2916. doi: 10.1093/jxb/ern164
Van den Ende, W. (2013). Multifunctional fructans and raffinose family oligosaccharides. Front. Plant Sci. 4:247. doi: 10.3389/fpls.2013.00247
Van Den Ende, W., Clerens, S., Vergauwen, R., Van Riet, L., Van Laere, A., Yoshida, M., et al. (2003). Fructan 1-exohydrolases. β-(2,1)-Trimmers during graminan biosynthesis in stems of wheat? Purification, characterization, mass mapping, and cloning of two fructan 1-exohydrolase isoforms. Plant Physiol. 131, 621–631. doi: 10.1104/pp.015305
Van den Ende, W., and El-Esawe, S. K. (2014). Sucrose signaling pathways leading to fructan and anthocyanin accumulation: a dual function in abiotic and biotic stress responses? Environ. Exp. Bot. 108, 4–13. doi: 10.1016/j.envexpbot.2013.09.017
van Herwaarden, A., and Richards, R. (2002). “Water-soluble carbohydrate accumulation in stems is related to breeding progress in Australian wheats,” in Proceedings of the 12th Australasian Plant Breeding Conference, ed. J. A. McComb (Perth: Plant Breeding for the 11th Millenium), 878–882. Available online at: http://hdl.handle.net/102.100.100/198752?index=1
Van Riet, L., Nagaraj, V., Van Den Ende, W., Clerens, S., Wiemken, A., and Van Laere, A. (2006). Purification, cloning and functional characterization of a fructan 6-exohydrolase from wheat (Triticum aestivum L.). J. Exp. Bot. 57, 213–223. doi: 10.1093/jxb/erj031
VanRaden, P. M. (2008). Efficient methods to compute genomic predictions. J. Dairy Sci. 91, 4414–4423. doi: 10.3168/jds.2007-0980
Wang, S., Wong, D., Forrest, K., Allen, A., Chao, S., Huang, B. E., et al. (2014). Characterization of polyploid wheat genomic diversity using a high-density 90 000 single nucleotide polymorphism array. Plant Biotechnol. J. 12, 787–796. doi: 10.1111/pbi.12183
Wang, Z., Liu, X., Li, R., Chang, X., and Jing, R. (2011). Development of near-infrared reflectance spectroscopy models for quantitative determination of water-soluble carbohydrate content in wheat stem and glume. Anal. Lett. 44, 2478–2490. doi: 10.1080/00032719.2011.551859
Wang, Z., and Xu, J. (2013). Predicting protein contact map using evolutionary and physical constraints by integer programming. Bioinformatics 29, i266–i273.
Wardlaw, I. F., and Willenbrink, J. (1994). Carbohydrate storage and mobilisation by the culm of wheat between heading and grain maturity: the relation to sucrose synthase and sucrose-phosphate synthase. Aust. J. Plant Physiol. 21, 255–271. doi: 10.1071/PP9940255
Wilkinson, P. A., Allen, A. M., Tyrrell, S., Wingen, L. U., Bian, X., Winfield, M. O., et al. (2020). CerealsDB-new tools for the analysis of the wheat genome: update 2020. Database 2020:baaa060. doi: 10.1093/database/baaa060
Xue, G. P., Kooiker, M., Drenth, J., and McIntyre, C. L. (2011). TaMYB13 is a transcriptional activator of fructosyltransferase genes involved in β-2,6-linked fructan synthesis in wheat. Plant J. 68, 857–870. doi: 10.1111/j.1365-313X.2011.04737.x
Xue, G. P., McIntyre, C. L., Jenkins, C. L. D., Glassop, D., Van Herwaarden, A. F., and Shorter, R. (2008). Molecular dissection of variation in carbohydrate metabolism related to water-soluble carbohydrate accumulation in stems of wheat. Plant Physiol. 146, 441–454. doi: 10.1104/pp.107.113076
Yáñez, A., Tapia, G., Guerra, F., and del Pozo, A. (2017). Stem carbohydrate dynamics and expression of genes involved in fructan accumulation and remobilization during grain growth in wheat (Triticum aestivum L.) genotypes with contrasting tolerance to water stress. PLoS One 12:e0177667. doi: 10.1371/journal.pone.0177667
Yang, D.-L. L., Jing, R.-L. L., Chang, X.-P. P., and Li, W. (2007). Identification of quantitative trait loci and environmental interactions for accumulation and remobilization of water-soluble carbohydrates in wheat (Triticum aestivum L.) stems. Genetics 176, 571–584. doi: 10.1534/genetics.106.068361
Yang, J., Zhang, J., Wang, Z., Zhu, Q., and Liu, L. (2004). Activities of fructan- and sucrose-metabolizing enzymes in wheat stems subjected to water stress during grain filling. Planta 220, 331–343. doi: 10.1007/s00425-004-1338-y
Zhang, B., Li, W., Chang, X., Li, R., and Jing, R. (2014). Effects of favorable alleles for water-soluble carbohydrates at grain filling on grain weight under drought and heat stresses in wheat. PLoS One 9:e102917. doi: 10.1371/journal.pone.0102917
Zhang, J., Dell, B., Conocono, E., Waters, I., Setter, T., and Appels, R. (2009). Water deficits in wheat: Fructan exohydrolase (1-FEH) mRNA expression and relationship to soluble carbohydrate concentrations in two varieties. New Phytol. 181, 843–850. doi: 10.1111/j.1469-8137.2008.02713.x
Keywords: water soluble carbohydrates, bread wheat, GWAS, QTN, gene prioritization, water stress
Citation: Gaur A, Jindal Y, Singh V, Tiwari R, Kumar D, Kaushik D, Singh J, Narwal S, Jaiswal S, Iquebal MA, Angadi UB, Singh G, Rai A, Singh GP and Sheoran S (2022) GWAS to Identify Novel QTNs for WSCs Accumulation in Wheat Peduncle Under Different Water Regimes. Front. Plant Sci. 13:825687. doi: 10.3389/fpls.2022.825687
Received: 30 November 2021; Accepted: 27 January 2022;
Published: 03 March 2022.
Edited by:
Yuri Shavrukov, Flinders University, AustraliaReviewed by:
Deepmala Sehgal, International Maize and Wheat Improvement Center (Mexico), MexicoXiongming Du, State Key Laboratory of Cotton Biology, Cotton Institute of the Chinese Academy of Agricultural Sciences (CAAS), China
Copyright © 2022 Gaur, Jindal, Singh, Tiwari, Kumar, Kaushik, Singh, Narwal, Jaiswal, Iquebal, Angadi, Singh, Rai, Singh and Sheoran. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Sonia Sheoran, Sonia.Sheoran@icar.gov.in