
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
REVIEW article
Front. Plant Sci., 17 May 2022
Sec. Plant Biotechnology
Volume 13 - 2022 | https://doi.org/10.3389/fpls.2022.782140
This article is part of the Research TopicOrphan Crops: Breeding and Biotechnology for Sustainable Agriculture, Food and NutritionView all 23 articles
 Jacob Olagbenro Popoola1*
Jacob Olagbenro Popoola1* Oluwadurotimi Samuel Aworunse1
Oluwadurotimi Samuel Aworunse1 Omena Bernard Ojuederie2,3
Omena Bernard Ojuederie2,3 Babasola Daniel Adewale4
Babasola Daniel Adewale4 Oluwapelumi Comfort Ajani1
Oluwapelumi Comfort Ajani1 Olaniyi Ajewole Oyatomi5
Olaniyi Ajewole Oyatomi5 Davelyne Ifechukwude Eruemulor1Taofeek Tope Adegboyega6
Davelyne Ifechukwude Eruemulor1Taofeek Tope Adegboyega6 Olawole Odun Obembe1,7*
Olawole Odun Obembe1,7*Poverty, food, and nutrition insecurity in sub-Saharan Africa (SSA) have become major concerns in recent times. The effects of climate change, drought, and unpredictable rainfall patterns threaten food production and sustainable agriculture. More so, insurgency, youth restiveness, and politico-economic instability amidst a burgeoning population requiring a sufficient and healthy diet remain front-burner issues in the region. Overdependence on only a few major staple crops is increasingly promoting the near extinction of many crops, especially orphan legumes, which possess immense potentials as protein and nutritional security crops. The major staple crops are declining in yield partly to their inability to adapt to the continuously changing climatic conditions. Remarkably, the orphan legumes are climate-smart crops with enormous agronomic features which foster sustainable livelihood. Research efforts on these crops have not attained a reasonable comparative status with most commercial crops. Though many research organizations and scientists have made efforts to promote the improvement and utilization of these orphan legumes, there is still more to be done. These legumes’ vast genetic resources and economic utility are grossly under-exploited, but their values and promising impacts are immeasurable. Given the United Nations sustainable development goals (SDGs) of zero hunger, improved nutrition, health, and sustainable agriculture, the need to introduce these crops into food systems in SSA and other poverty-prone regions of the world is now more compelling than ever. This review unveils inherent values in orphan legumes needing focus for exploitation viz-a-viz cultivation, commercialization, and social acceptance. More so, this article discusses some of the nutraceutical potentials of the orphan legumes, their global adaptability, and modern plant breeding strategies that could be deployed to develop superior phenotypes to enrich the landraces. Advanced omics technologies, speed breeding, as well as the application of genome editing techniques, could significantly enhance the genetic improvement of these useful but underutilized legumes. Efforts made in this regard and the challenges of these approaches were also discussed.
Africa’s population is currently estimated above 1.3 billion, and it is expected to hit 2.5 billion by the year 2050 (Fischer et al., 2009; Paliwal et al., 2021). This burgeoning figure is mounting pressure on food production. At the same time, factors such as poor soils, land degradation, climate change, lack of access to fertilizers, poor agricultural infrastructures, banditry, and insurgency impede sustainable agriculture. The continent is endowed with a rich agrobiodiversity and has excellent potential for self-sufficiency in food production. However, it is one of the food- and nutrition-deficient regions in the world. Africa’s agrobiodiversity is continuously under threat resulting in the erosion of valuable genetic resources (Popoola et al., 2020; Ikhajiagbe et al., 2021). The destruction of many agricultural fields and farms by overgrazing and unregulated nomadic pastoralism has fueled an upsurge in herder-farmer’s conflict in some regions, thereby subjecting many to poverty. Moreover, the internal displacement of persons is on the increase. Traditional agronomic practices for many indigenous species are on the verge of being lost, resulting in low yield and neglect. This could have far-reaching consequences on global food production and the supply chain. Whereas food insecurity and dietary deficiencies are global issues, their severities are more pronounced in Africa and some parts of Asia (Li and Siddique, 2020; Popoola et al., 2020). Nutritional transitions, overdependence on a few major staple foods, and improved socioeconomic status contribute to food and protein insecurity (Dixon, 2009; Chakona and Shackleton, 2017). Mitigating the impacts of these problems calls for global attention and concerted efforts by all stakeholders.
The promotion of indigenous biodiversity and incorporation of nutrient-dense crops into food value chains have been identified as measures that could ensure sustainable and resilient food systems, improve diet quality, and increase smallholder farmers’ incomes (Padulosi et al., 1999; Cheng et al., 2019; Hunter et al., 2019). The United Nations SDGs of zero hunger, achieving food security, improving nutrition, and promoting sustainable agriculture, are critical to alleviating poverty and malnutrition in Africa. To ensure sustainable food production and overcome dietary deficiencies, several strategies, including consistent cultivation, conservation, and genetic characterization of many unrecognized, abandoned, and under-exploited species alongside the reformation of traditional agronomic practices, are imperative (Popoola et al., 2020; Ikhajiagbe et al., 2021). Dietary diversification and incorporation of resilient crop species such as under-exploited leguminous species referred to as an orphan, minor, or underutilized into food systems will ensure food and protein security in many parts of Africa (Considine et al., 2017; Cullis and Kunert, 2017; Singh and Fernandes, 2018).
The term “Underutilized” or “orphan” alludes to the neglect of a species by international and indigenous research communities. Orphan species are generally classified as crops with little relevance at the global level (Muhammad et al., 2020). By way of definition, underutilized legumes are a diverse collection of domesticated pulses with beneficial properties, but with limited importance compared to major global crops like rice, maize, potato, and wheat due to utility and supply constraints. This set of crops regardless of their neglect is characteristically diverse with cultural values and inherent useful agronomic, genetic, and biochemical traits (Cullis and Kunert, 2017; Khan et al., 2021a). Orphan legumes play a vital role in several developing countries by generating revenue for smallholder farmers, as well as tackling micronutrient and protein deficiencies associated with the consumption of high calorie diet derived from the major crops researched by scientists and private corporations. Although these crops may be extensively distributed beyond their centers of origin, orphan legumes occupy special niches in traditional production and consumption systems. Whereas underutilized legumes are stapled food crops in many rural communities in sub-Saharan Africa (SSA), their economic potential for regional and international markets have not been fully tapped into, hence their neglected status and lack of genetic improvement, resulting in reduced quality and yield (Cullis and Kunert, 2017; Mabhaudhi et al., 2017). Although it is difficult to precisely define what attributes makes a crop underutilized, orphan legumes are marked by the following constraints: (i) often associated with the cultural heritage of their places of origin; (ii) poor documentation of their cultivation and use; (iii) adaptation to specific marginal land and agro-ecological niches; and (iv) no formal seed supply systems. In addition, the hard-to-cook (extended time required to achieve desired softening during cooking) characteristic, inadequate processing techniques, and seed coat hardness of many orphan legumes results in a lack of functional value chain that guarantees the delivery of processed and refined products from farmers to consumers (Khan et al., 2021c). Major legumes with a high quantity of oils particularly soybean and groundnut are preferred to orphan legumes in some regions owing to their numerous uses (Nedumaran et al., 2015; Muhammad et al., 2020). Consequently, orphan legumes which are subsistence food crops for the indigenous population are replaced with these profit crops given their export potentials (Muhammad et al., 2020). In Africa, many orphan legume crops abound which due to their dense nutritional profile, good adaptability to adverse climatic scenarios, and ability to grow in marginal soils hold potential for sustainable cultivation (Gulzar and Minnaar, 2017).
This review focuses on five orphan legume crops endemic to SSA. The crops include African yam bean (AYB; Sphenostylis stenocarpa), Bambara groundnut (BG; Vigna subterranean), Kersting’s groundnut (KG; Macrotyloma geocarpum), Lima bean (LB; Phaseolus lunatus), and Jack bean (JB; Canavalia ensiformis). In addition to the above-listed features, the criteria of low production rank and per capita consumption coupled with common trade in terms of export and import volumes (tons) at the international market compared to mainstream pulses like soybeans (Glycin max) and groundnut (Arachis hypogaea) were used in the selection of the five species. Although yield, production, and consumption of pulses had increased modestly in the last decades in SSA and on the global scale, orphan legume crops were not classed as commodities in the international trade framework. Statistics on their production, consumption, and trade remain scanty and, in some cases, nonexistent or aggregated with other pulses/dry beans. Therefore, this paper unveils inherent values in the selected orphan legumes needing focus for exploitation viz-a-viz cultivation/production, commercialization, and social acceptance. More so, this article discusses some of the nutraceutical potentials of the orphan legumes, their global adaptability, and modern plant breeding strategies that could be deployed to develop superior phenotypes to enrich the landraces. Also discussed are current biotechnology approaches and plant breeding strategies, including advanced omics technologies, speed breeding, genome editing tools, and the challenges of these approaches.
The production and yield quantities of pulses in Africa have surged in the last decade from 2010 to 2020, with a fall between 2015 and 2016 and a steady rise from 2017 to 2020 (Figure 1). Globally, in 2020, over 5.9 million hectares (Ha) of pulses were cultivated with a production of over 4.4 million tons (MT) and a yield of 7,503 kg/ha. Africa produced only about 1.35 MT of pulses on a harvested area of 2.12 million ha with a yield of 6,407 kg/ha. In the same year, the global production of soybean and groundnut was above 350 MT and 53 MT compared to 0.23 MT for BG (FAOSTAT, 2020). Currently, BG production is restricted to only seven countries: Burkina Faso, Cameroun, Democratic Republic of Congo (DRC), Mali, Togo, Niger, and Zimbabwe. Based on FAOSTAT, there are no production data on AYB, KG, LB, and JB (Table 1; Figure 2). Burkina Faso is the leading producer of BG with 57,429 tons, followed by Niger (55,570 tons) and Mali (26,996). Previously 100,000 tons were reported from Nigeria (Hillocks et al., 2012), but this is not captured on the current FAOSTAT data. The overview of the global area and production of pulses show that more tons of soybean and groundnut were produced globally and in SSA compared to BG and lack of production in others. The five orphan legume crops lag with little or no production data or aggregated under other pulses/beans compared to the major pulses (Ayenan and Ezin, 2016; FAOSTAT, 2020). Lima bean production is not reflecting on FAOSTAT, and just like BG, its production and that of others might not likely exceed 0.23 MT recorded for BG.
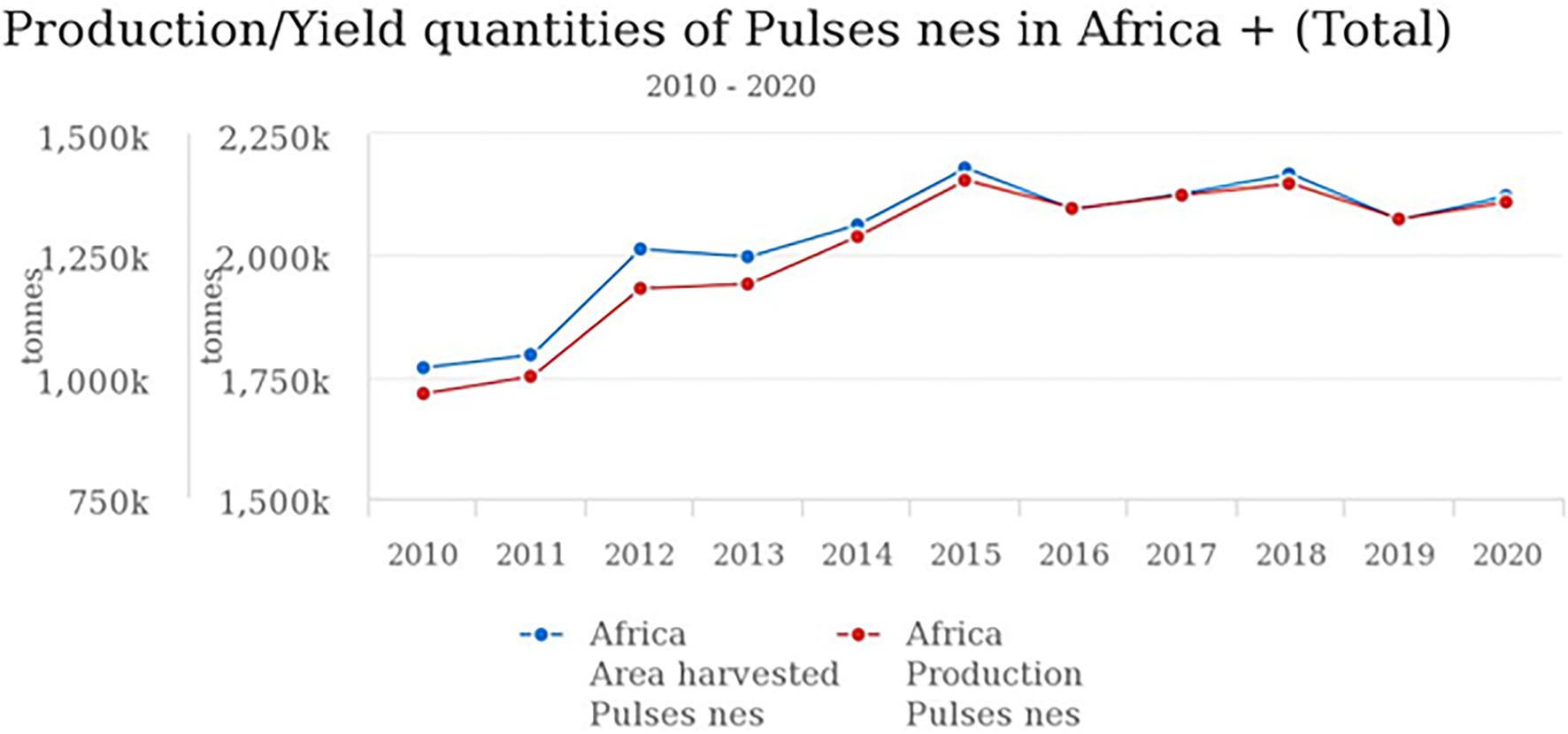
Figure 1. Pulses production/yield quantities in Africa.
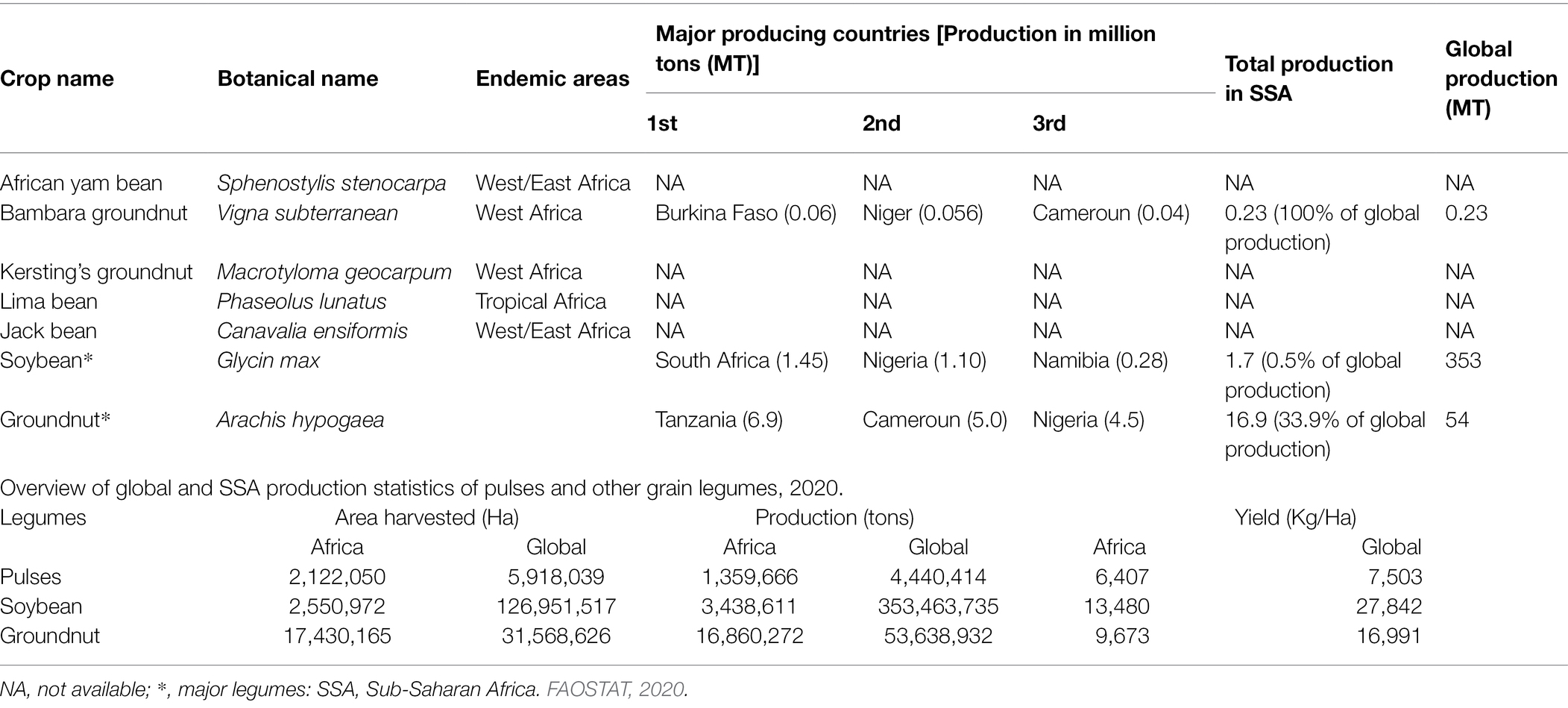
Table 1. Major producing countries of five selected orphan legumes of SSA and global production statistics FAOSTAT (2020).
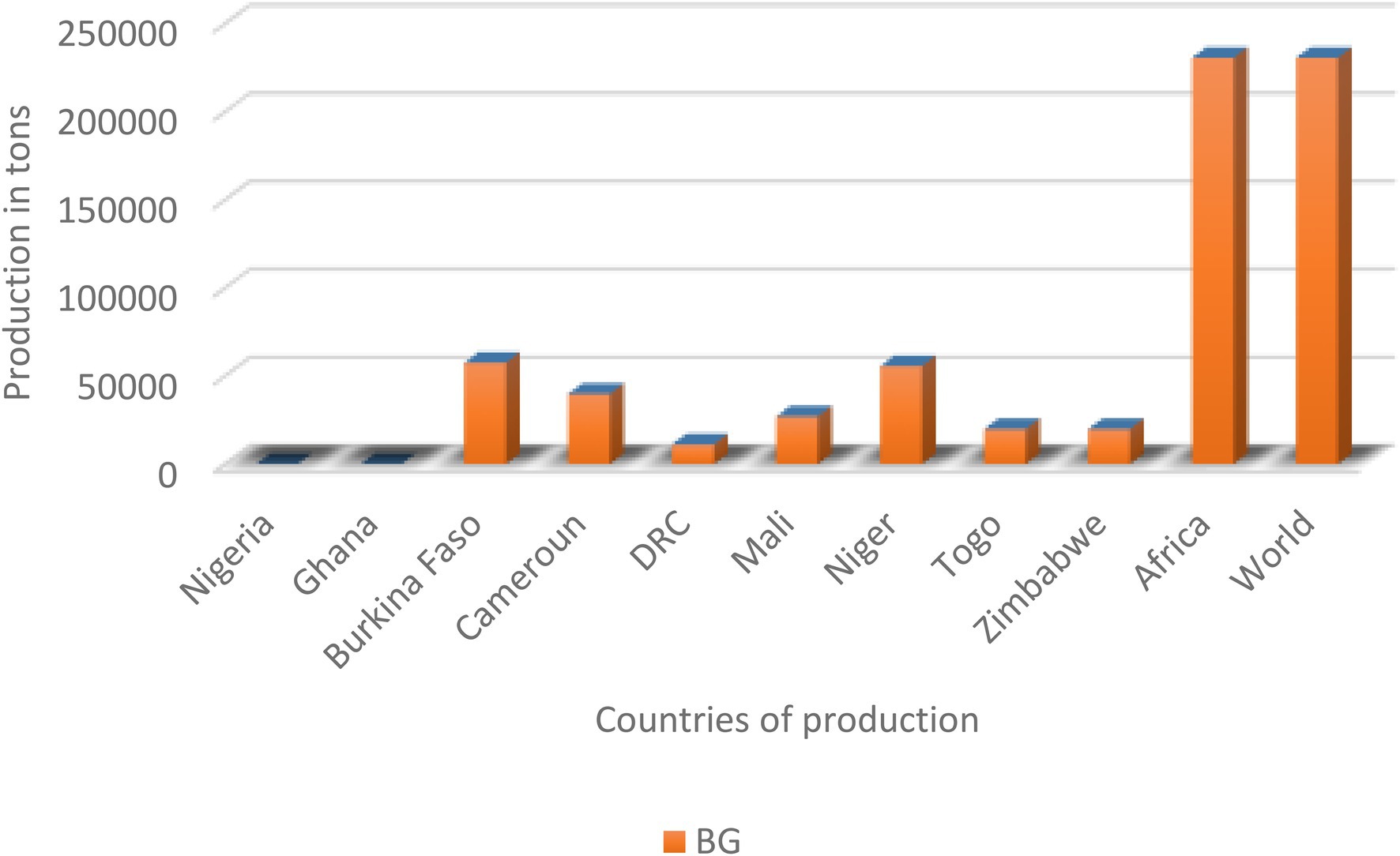
Figure 2. Production in tons of Bambara groundnut (BG). Only BG has data in FAOSTAT.
Consumption of pulses had been driven by factors like taste, accessibility, ease of cooking, population growth, dietary needs, income level, and other supply chain constraints (Nedumaran et al., 2015). Given SSA sizeable consumption requirements, the region consumed 37% of global roots and tubers and only 21% of global pulses (OECD/FAO, 2020). The per capita consumption of pulses recommended by the Food and Agriculture Organization of the United Nations (FAO) is 30 g/day/person. Remarkably, the average per capita daily consumption of pulses in SSA has significantly increased from about 21 g/day/per in 1985 to about 33 g/day/person in recent time (Rawal and Navarro, 2019; Akah et al., 2021). However, the most consumed pulses are the common bean, soybean, and groundnut. The per capita consumption of common bean is about 10 g/day/person. In contrast, soybean consumption has increased tremendously (about 32 g/day/person) and is driven by multiple utilizations in the poultry, fishery, and edible oil industries. Contrarily, the consumption of orphan legumes is meagre, and their per capita daily consumption level is unknown or nonexistent compared to the mainstream pulses. Most orphan legumes are fallback crops usually consumed in lean periods when popular pulses are unavailable and expensive. Several factors such as poor agronomic features, high cost of production, intensive labor, lack of improved varieties, changes in dietary patterns, and most critically, the hard-to-cook (HTC) phenomenon hinders their production and consumption (Alexandratos and Bruinsma, 2012; Tan et al., 2020; Khan et al., 2021c).
The orphan legumes are mainly cultivated by subsistence farmers, who lack the financial capability to adopt high-input farming practices needed to grow major staple crops (Conti et al., 2019). In some areas, these crops are referred to as “the poor man’s food.” In Niger and Mali, these crops were essential food sources for poor farmers and their livestock during a period of drought that lasted over 20 years (Chivenge et al., 2015). The orphan legume crops are well appreciated by the traditional farmers who have spent most of their years in their hometowns, where the origin and usefulness of these grains are well understood. Farmers are aware of these crops’ capacity to fix atmospheric nitrogen into the soil, tolerate drought, provide medicine, efficiently utilize soil moisture through their deep root system, and survive harsh environmental conditions. However, significant constraints such as long cooking time, poor shelf-life, low yield, sensitivity to daylight, presence of potent anti-nutritional factors (ANTs) have plunged their utilization as farmers are less interested to invest their time and energy in their cultivation and production (Nnamani et al., 2017; Popoola et al., 2020). More so, poor market price, poor demand, lack of buyers, lack of improved varieties, and inadequate capital have hampered their production (Khan et al., 2021c). Generally, the acceptance and utilization of these underutilized grain legumes are declining owing to urbanization, change in market needs, migration, land degradation, and other earlier mentioned factors. It is believed that these factors consequently reduce the transfer of traditional and agronomical knowledge on these crops from the older to younger generation. Thus, reliable production data are scanty, except for BG (Khan et al., 2021c) and as highlighted in Table 1.
Currently, several researchers such as those at the Genetic Resources Center, International Institute of Tropical Agriculture (IITA), Nigeria; Crops for the Future Malaysia; African Orphan Crops Consortium, among others, are reappraising the need to adopt the orphan legumes as a means of boosting child development, improving public health, and bridging the nutritional gaps in the yearly nutritional cycles (Cullis et al., 2018; Vidigal et al., 2019; Paliwal et al., 2021). Very recently, Ojuederie et al. (2020) reported that well-processed seeds and tubers of AYB (one of the orphan legumes) could be included in meals to reduce protein malnutrition while improving food and nutritional security in Africa. All these are geared toward enhancing the utilization of such crops to increase their production. Thus, to obtain significant benefits from these underutilized grain legumes, there is a need for systematic cultivation, wide acceptance, increased utilization, strategic crop improvement and funding, and the creation of global and local market spaces to enhance consumption across the SSA.
Diversity in seeds of the selected five underutilized legumes that can be cultivated to enhance Africa’s food and protein security is shown in Figure 3, while basic scientific information is listed in Table 2. In areas like East Africa, the Pacific, South Asia, and SSA, progress in the fight against poverty, food insecurity, and malnutrition is slow-paced compared to other regions of the world. Most importantly, the food system should be the first area of concern due to its capacity to provide nutrition to the people and stabilize the dilapidating economies of these countries. The need for the addition of food varieties like orphan legumes to the existing homogenous food-basket system in these regions can serve as safety nets to supply the nutritional needs of the people and generate support for local markets. Orphan legumes are resistant to biotic stress like pests and diseases, rich in quality protein and B vitamins, high in lysine content, and a good source of healthy oils. More so, orphan legumes are effective in fixing soil nitrogen, and capable of thriving in unfavorable environmental settings (Ojuederie et al., 2020; Popoola et al., 2020). A crop like Kersting’s groundnut is intolerant to soils with high moisture content. Cultivating this crop in drought-prone areas will have no adverse effect on the yield or nutritional content. While a few African countries such as Ethiopia, Kenya, and Malawi have emergency food reserve systems in place, some others are still unable to provide infrastructures critical to the conservation of these future food products. In the absence of appropriate storage infrastructure, embracing orphan legumes that require little or no special preservative techniques can be suitable in the short term.

Figure 3. Diversity in seeds of the selected orphan legume crops. (A) African yam bean (Sphenostylis stenocarpa). (B) Kersting’s groundnut (Macrotyloma geocarpum). (C) Jack bean (Canavalia ensiformis). (D) Lima bean (Phaseolus lunatus). (E) Bambara groundnut (Vigna subterranean). Source: seeds were collected from the International Institute of Tropical Agriculture (IITA, Ibadan, Nigeria).
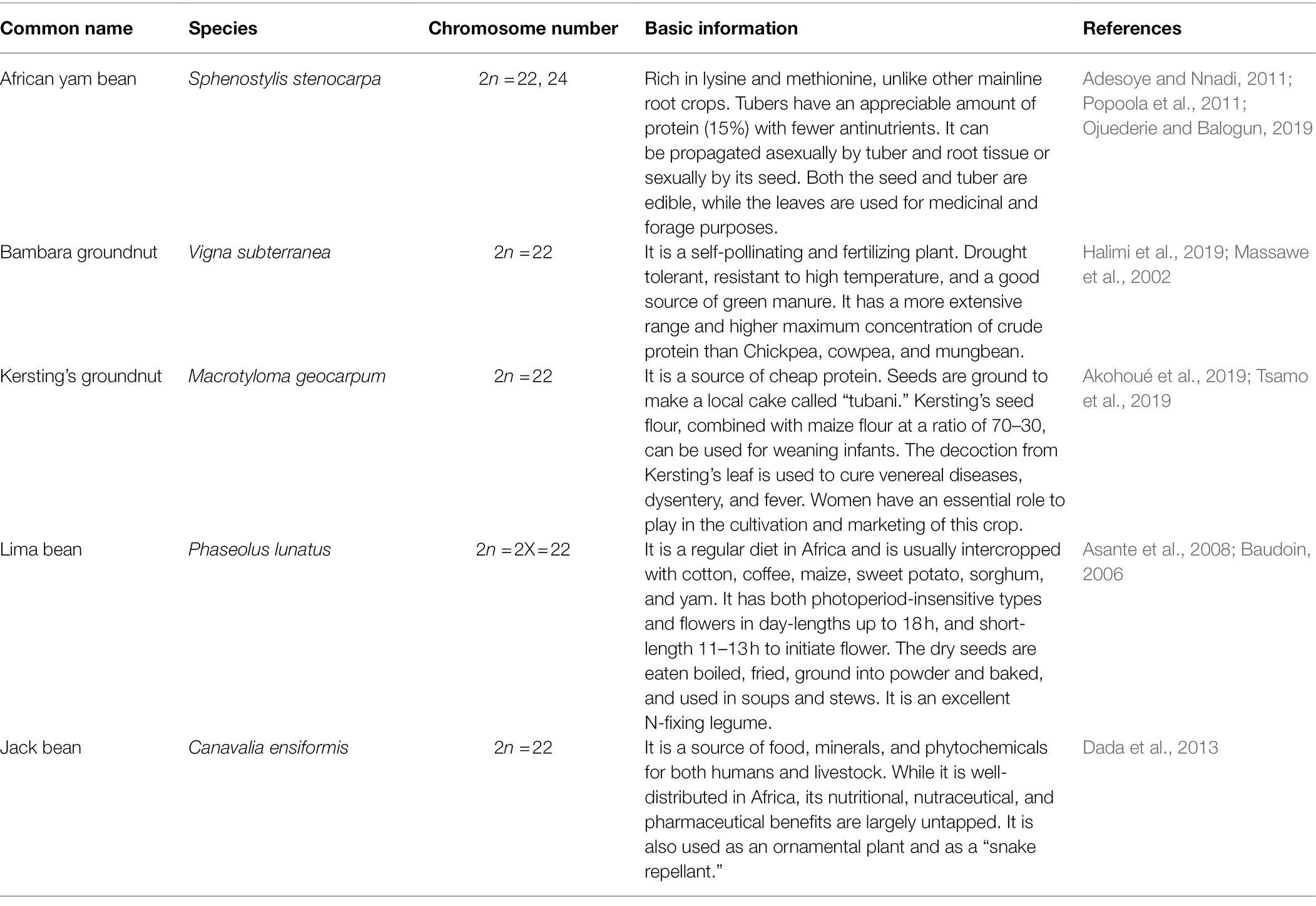
Table 2. Basic scientific information on selected orphan legumes in SSA.
The global pulse trade has grown with prices more volatile and increasing faster than traded volumes (Akibode and Maredia, 2011; Nedumaran et al., 2015). This trend situates the growing global demand relative to supply. Also, the export and import of pulses had greatly expanded across SSA (Fabinin, 2021). In 2014, the international export of pulses was worth over USD 10 billion, and Africa contributed only about 9.5%, while in 2020, it grew to over USD 12 billion (ITC, 2022). Major traded pulses include soybean, groundnut, lentils, peas, chickpeas, kidney beans, and black gram. Among the five orphan legumes considered in this review, only BG is on the list and traded but of low volume. In contrast, AYB, Kersting’s groundnut, Lima bean, and Jack bean are not traded or likely bundled under other pulses/beans. The market trend for many orphan legume crops is somehow sketchy. Nevertheless, local trading and markets exist, and we predict regional and international trade expansion in the coming years as awareness increases and the value chain expands.
This crop is grown locally in West/East/Southern African countries like Nigeria, Ghana, Benin, Cameroun, South Africa, Zambia, Uganda, and Angola. Its trade is restricted to local markets and not listed under crops and livestock products (production) of the Food and Agriculture Organization (FAO) nor listed under the Harmonized Tariff code System (HS) of the International Trade Centre (ITC). There are no data on its harvested area in hectares, production in tons, and market values. However, in Nigeria, an income of N74,000, equivalent to USD180, was generated monthly from the sales of prepared foods and derived products sold in open markets along the highways. This amount is more than twice the minimum wage (N30,000 or USUSD73) paid by the Federal Government of Nigeria (Nnamani et al., 2017). This portends a high potential to generate income for the rural population and their households.
It is a pulse with a subterranean fruit set, cultivated by subsistence farmers mostly in semi-arid regions of Africa (Olanrewaju et al., 2022). BG is emerging as an income source and gradually receiving more international research attention with export potential. Currently, it is not classed as a commodity in the global trade framework due to its low production rank (Halimi et al., 2019; Hasan et al., 2021), but it is classified as an edible leguminous vegetable and falls into the Harmonized Tariff code System (HS) as fresh (HS0708) or dried (HS0713). The trade outlook for BG is promising due to its high yielding potentials in varied agro-ecological areas and available intra-Africa trade outlets. The top exporter of BG in 2020 was Madagascar, with an export value of 213,000 USD, while South Africa was the leading importer with an import value of 282,000 USD. The awareness of the potential of BG as a climate-smart crop is expanding beyond its endemic areas (Jahanshiri et al., 2022). Recently, a total of USD 6 to 13 billion of yearly income to the global economy was predicted for BG using the mean potential areas with a modesty yield of 0.85 t/ha for one optimal season (June–September) with a modest price of 143 USD/T (Mayes et al., 2019; Hussin et al., 2020; Jahanshiri et al., 2022). Currently, in Nigeria, 50 kg of BG sells for N30,000 or USD 72.30, with a considerable profit from the sales of processed products (Onuche et al., 2020). Similar information has been reported from Mali (Majola et al., 2021).
It is a highly nutritious tropical crop adapted to drought-prone areas but neglected by researchers and policymakers. KG is declining in cultivation, and production statistics are unavailable due to intensive labor requirements, low yield, and improved varieties (Ayenan and Ezin, 2016). It is not yet traded globally, but it is well-known in Togo, Benin, Ghana, and Burkina Faso. It provides substantial incomes for the rural population, and its price can climb from CFA 1000 (USD 2) per kg in a period of plenty to CFA 4,000–5,000 (USD 8–10) per kg in a period of scarcity (Assogba et al., 2015; Akohoué et al., 2019). Farmers produce KG mainly for home consumption, and thus, market value is still low. Nonetheless, it is an essential source of income for the rural populace since gross revenue earned from growing the crop averages USD1200/ha (Assogba et al., 2015; Akohoué et al., 2019).
This crop is also known as butter beans. Though there is no matching HS code and trade statistics for LB, its trade is increasing globally and possibly lumped under the HS code for other pulses. Its utilization is expanding in the United States, Brazil, and India. Production and trade volume is low in SSA. In 2020, Morocco was the top exporter of LB with an export value of USD 271 M, while the United States was the leading importer valued at USD 198 M (Tridge Statistics, 2022)—data based on HS code 070820 (Phaseolus spp).1
This crop is grown in the traditional farming system in SSA, and there are no statistics on its trade. It has impressive nutritional contents, but it is largely untapped.
In summary, the prospects of these underutilized legumes look promising in SSA if stakeholders, including farmers, policymakers, government, and consumers, can embrace an integrated approach involving improved varieties, modern agronomic practices, increased funding, genetic manipulation using their rich genomic resources, and infrastructural development.
Several agronomic features confer on orphan legumes significant potential to enhance sustainable agriculture and human livelihood (Table 3). Poor research interest in these crops informs that there is no robust, comparable, and reliable empirical information which can be used to advocate for policy development (Mabhaudhi et al., 2019). This further establishes their poor competition with notable crops. However, the few reports considered in this review informed that orphan legumes are not useless but host huge possibilities for food and protein security. Critically, the continued existence of orphan crops within marginalized farming systems shows that they are adaptable to changing climatic scenarios. According to Vidigal et al. (2019), pulse crops offer a viable and sustainable strategy for upholding farming systems’ intercropping and production indices. The capacity to augment soil nitrogen makes orphan legumes an excellent partner in many farming systems, with additional contributions leading to soil fertility improvement, biodiversity conservation, ecosystem stabilization, farming risks reduction, and sustainable yield (Table 4).

Table 3. Exploitable agronomic wealth offered by underutilized legumes.

Table 4. Agronomic Utilities, Challenges, and plausible solutions of the five orphan legumes.
The survival of orphan legumes in a marginal environment may be due to their capacity to stimulate the colonization of a variety of nitrogen-fixing bacteria in different soil types (Naluwairo, 2011; Baldermann et al., 2016; Mabhaudhi et al., 2019).
As documented in Babalola et al. (2017), Allorhizobium, Azorhizobium, Bradyrhizobium, Mesorhizobium, Rhizobium, and Sinorhizobium are among the nitrogen-fixing bacteria which exist naturally in legumes. Moreover, the diversity of host legumes is accompanied by diversity in nodulating bacteria with significant differential capacity at taxa and cultivar levels (Sprent et al., 2010). This seems to inform that the use of legumes in soil amelioration would be a backbone for organic agriculture if their resources are well harnessed. There are very wide variations in nodule productivity and effectivity of both plant and rhizobial germplasms for optimizing nitrogen fixation (Sprent et al., 2010). Babalola et al. (2017) hinted that there are so many underutilized leguminous crops whose potentials have not been fully tapped to understand their functionalities within the realm of biological N fixation (BNF).
Agronomic features, specific challenges, and possible solutions of the considered orphan legumes are provided in Table 4. Notably, the contribution of the nitrogen-fixing bacteria which cooperatively function with orphan legumes is not well documented. However, the reasons for the protracted featuring of the most orphan leguminous crop in the cropping systems in SSA may be due to their contribution to soil fertility which farmers may have found to be highly sustainable. We speculate that the quantity of nitrogen they supply to the soil may be very significant, although this needs to be investigated.
Human dietary deficiencies are caused by a lack of essential nutrients in diets or the body’s inability to absorb and process these nutrients once ingested. Deficiency in nutritional requirements is the leading cause of various diseases today, like diabetes, protein-energy malnutrition (PEM), ‘beriberi’ (Thiamine deficiency), among others (Wokes et al., 1955; Flyman and Afolayan, 2006). Generally, legumes are a good source of quality proteins, dietary fiber, vital amino acids, and minerals, but scientific reports have shown that under-exploited legumes are nutritionally superior to the other commonly known legumes (Considine et al., 2017; Adegboyega et al., 2020). Many diseases are associated with a low diet, high in animal-based nutrients, overly processed foods, and low plant-based nutrients. Underutilized legumes are known to be a rich source of bioactive compounds, unsaturated fat, little or no cholesterol, and dietary fiber (Bhartiya et al., 2015; Li and Siddique, 2020). These dietary constituents promote health and sustainability by decreasing insulin production and preventing chronic diseases such as cancer, cardiovascular diseases, obesity, and the likes (Bhartiya et al., 2015; Salvi and Katewa, 2016; Hunter et al., 2019). A legume-based diet can guarantee a longer and healthier life. Without any doubt, the inclusion of under-exploited legumes into the dietary cycle of countries suffering from chronic deficiencies, malnutrition, hunger, and protein deficiency, will be a long-lasting solution due to its dense nutrient content, good quality protein, and micronutrients. It could also reduce the over-dependence on other major legumes like soybean, cowpea, chicken pea, groundnut, etc. In obesity, several studies have shown that consumption of processed underutilized legumes such as AYB and Lima bean could significantly aid weight loss (Onwuka et al., 2009; Crujeiras et al., 2010; Rebello et al., 2014). This could be attributed to the low-fat content and high dietary fiber. Many vegetarians include underutilized legumes as an alternative source of animal protein/meat without lagging in nutritional quality (Crujeiras et al., 2010; Vidigal et al., 2019). The low gastrointestinal tract nature of legumes’ carbohydrates aids in stabilizing the blood glucose level (Maphosa and Jideani, 2017). Generally, the B-Group vitamins such as folate, niacin, thiamin, riboflavin, pyridoxine, pantothenic acid among others are abundant in the orphan legumes (Table 5). These species are also a source of essential minerals like zinc, iron, calcium, potassium, copper, and selenium (Table 5). Calcium plays an important role in bone health and also eases the movement of blood within vessels. Zinc helps boost the immune system, iron synthesizes hemoglobin, and potassium helps prevent stroke (Barman et al., 2018). Orphan legumes with high zinc content would be very useful in preventing the symptoms associated with the Covid 19 virus if introduced into the human diet in the required amount. The high-quality protein (from 19.4 g per 100 g in KG to 22.5 g per 100 g in AYB) available in these orphan legumes makes them suitable and essential for every age group. However, some species like AYB, BG, KG, and JB are known to contain some toxic phytochemicals that can lead to bloating, flatulence, or activate allergic reactions in some people. Fortunately, most of these toxins can be easily neutralized by steaming, de-hulling, boiling, fermenting, roasting, and using advanced processing technology such as irradiation, infrared heating (micronization), and high-pressure cooking without jeopardizing their nutritional contents (Maphosa and Jideani, 2017; Adeleye et al., 2020; Tan et al., 2020; Khan et al., 2021c). Comparative nutritional profile of raw, mature seeds, and values per 100 g of the considered orphan legumes are shown in Table 5. The dense network of adequate quality protein, novel minerals, and vitamins all make up for their nutraceutical and pharmaceutical impact on human health, not exempting the animals’ health that feed on them.
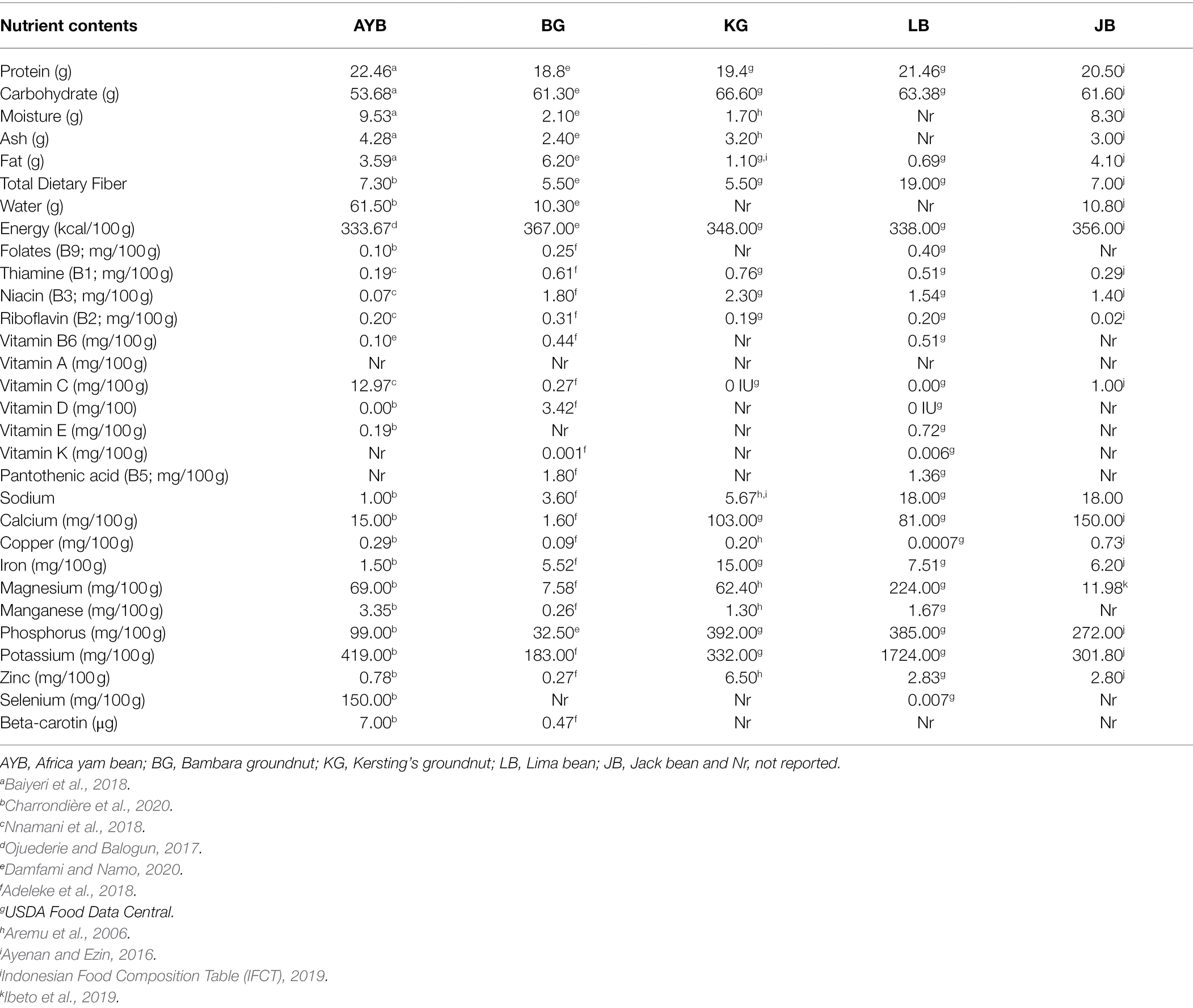
Table 5. Nutritional profile of raw, mature seeds, and values per 100 g of five orphan legume crops.
Regional differences exist in the utilization of these rich protein and nutrition security crops. For instance, for the AYB, the tubers are highly consumed in east and central Africa while the seeds are consumed in West Africa. Thus, there is the tendency of having more seed production of AYB in West Africa and the focus on tuber production in the east and central Africa. Studies by (Ojuederie and Balogun, 2017) and Ojuederie et al. (2020) showed that the tubers of AYB have a high protein content of up to 15% which is considerably higher than the protein content found in tuber crops such as cassava and sweet potato. Findings of their study on Wistar rats also found the tuber to possess fewer antinutrients than the seeds. This study has led to more research interest on the tuberization of AYB for food and nutrition security. There is therefore an urgent need to increase the production of these orphan legumes and others identified as being rich in protein and essential minerals to lessen malnutrition and enhance food and nutrition security in Africa.
A nutraceutical can be defined as a food or part of a food that provides medical or health benefits, including preventing or treating a disease. It can be an isolated nutrient, dietary products, processed foods, or beverages. Legume seeds can be considered a potent nutraceutical. It benefits human health and prevents or treats several diet-related diseases such as obesity, cardiovascular diseases, digestive tract diseases, diabetes, and others (Morris, 2003; Duranti, 2006). Regardless of the dense protein content in legume seeds, they are also known to contain an adequate amount of energy, carbohydrates, minerals, vitamins, dietary fiber, and low-fat content (Ojuederie and Balogun, 2017; Adegboyega et al., 2020; Ojuederie et al., 2020). Some bioactive compounds like glycosides, tannins, isoflavones, saponins, flavonoids, and so on contribute to its nutraceutical abilities (Carbonaro et al., 2015; Hoste et al., 2015). In most extensively pigmented legume seeds such as Lima bean, Jack, and Sword beans, there is a high phenolic acid and flavonoid level, contributing to the coloring alongside anti-oxidative functions (Morris, 2003; Carbonaro et al., 2015; Serventi and Dsouza, 2020). The seeds also contain enzyme inhibitors α-amylase, α-glucosidase, and γ-aminobutyric acid (GABA), for which it can be used as a nutraceutical molecule (Barman et al., 2018). Green legume seeds are also a good source of nutraceutical compounds. When legume seeds are processed, they enhance the present nutraceuticals’ bioavailability by inactivating ANTs like trypsin, growth inhibitors, and hemagglutinins (Suneja et al., 2011; Serventi and Dsouza, 2020). Some molecules in legume seeds that have been tagged toxic can be crucial to human health if consumed in the right proportion.
Underutilized legumes can be used as active components of drugs and other pharmaceutical products to promote human health status in terms of pharmaceuticals. Most under-exploited legumes are used in folk medicine in various parts of the world without knowing their active ingredients. For example, Kersting’s groundnut can be used to cure diarrhea, BG provides relief from menstrual cramps, cures insomnia, and promotes red blood cell production as well as prevent cancer (Ayenan and Ezin, 2016; Tsamo et al., 2019; Udeh et al., 2020). Similar properties abound in most of the underutilized legumes, including the AYB seeds where the paste made from the ground seeds have been used to treat ailments such as stomach aches and acute drunkenness when mixed with water in some West African countries; Ghana, Nigeria, and Togo (Adewale and Dumet, 2010). These pharmaceutical capabilities are attributed to bioactive agents like phenols capable of signaling structural polymers such as anthocyanin, flavonoids, tannins, and phenolic acids that chelate metal ions, inhibiting the peroxidation lipids and scavenge free radicals (Morris, 2003; Serventi and Dsouza, 2020). Phenolic compounds are responsible for the anti-oxidative nature of the seeds (Serventi and Dsouza, 2020). Also, phenolic compounds show anti-bacterial, anti-viral, anti-inflammatory, anti-mutagenic activities, and cancer prevention (Morris, 2003; Serventi and Dsouza, 2020; Udeh et al., 2020). Precisely, legumes exhibit pharmacological properties such as antioxidant, antimicrobial, anticancer, anti-inflammatory, anti-obesity, and heart protection (Serventi and Dsouza, 2020). The seeds have to be fermented to get the maximum anti-oxidative potentials of phenolic compounds (Rezende et al., 2018). Triterpenoids are commonly found saponins in legumes which play a major role in reducing carcinogenic substances in the colon. They can lower the risk of heart diseases and serve as immune stimulants by inducing cytokine production such as interferon (IFN) and interleukins (Singh et al., 2017; Serventi and Dsouza, 2020). Tannins are also abundant among the orphan legumes, which helps remove toxins from the intestinal tract because of their ability to bind proteins (Hoste et al., 2015; Barman et al., 2018). Anthocyanin, alkaloids, enzyme inhibitors such as α-Amylase and α-Glucosidase, phytic acid, and phytoestrogen have been isolated and reported among the orphan legumes (Barman et al., 2018; Serventi and Dsouza, 2020). The nutraceutical components of these grain legumes are mostly used as preventive measures of disease or to reduce some infections’ virulence (Duranti, 2006; Hoste et al., 2015; Barman et al., 2018). The introduction of these under-exploited legumes into our diet at a healthy amount will be of great health benefits to people in SSA.
Globally, tropical Africa and some regions in Asia are the most significant endemic areas and producers of orphan legumes (Cullis and Kunert, 2017; Popoola et al., 2020; Paliwal et al., 2021). These species’ ability to thrive in various environments, whether harsh or favorable, has enabled the species to survive in diverse areas. For instance, the Lima bean (Phaseolus lunatus) was initially endemic to West Africa, but it is now cultivated mainly in the temperate regions of the United States (Caicedo et al., 1999; Serrano-Serrano et al., 2010; Andueza-Noh et al., 2013). Furthermore, adaptability is the capacity to acclimatize to a wide range of environmental conditions effectively. Orphan legumes should be a reference masterpiece in terms of adaptability due to their ability to overcome neglect, near extinction, and under-exploitation before becoming the highlight of scientific research. These unpleasant conditions triggered various adaptation mechanisms (survival strategies) in their morphology, physiology, biochemical, and genetic constituents (Sita et al., 2017). Such survival mechanisms include; possession of a deep taproot system to enhance soil water capture during drought, regulation of stomata, reduction of canopy size and duration, increasing wax accumulation on the surface to prevent water loss, early maturation to allow reproduction before environmental conditions depreciate (mostly in annual crops), and nitrogen-fixing ability to self-enrich their soil (Sita et al., 2017; Kumari et al., 2020). The Canavalia species (C. ensiformis and C. gratilis) are characterized by an extensive rooting system that can be exploited for phytoremediation of polluted sites. Generally, grain legumes have various adaptation mechanisms for different threats and stresses; this ability makes them unique, and the need for their exploitation is crucial (Considine et al., 2017; Cullis and Kunert, 2017).
Plant genetic resources (PGR) are the plant’s vegetative and reproductive parts, from which the plants are propagated. In orphan legumes, every part is of great importance, ranging from the root, seeds, tubers, pods, flowers, and stems. Each part possesses heritable characteristics of potential value to plant breeders, varying from their dietary, medicinal, and social value. Due to the effortless adaptability of orphan legumes to almost all climatic conditions, each species has thousands of accessions, varying in phytochemical content, proximate, nutritional, and antimicrobial values. The availability of a diverse collection of PGR is the building block for any crop improvement program. Future development of improved varieties of orphan legumes depends largely on collections and preservation of their PGR. The Genetic Resources Center (GRC) of the International Institute of Tropical Agriculture (IITA) presently maintains a collection of about 6,747 accessions of various orphan legumes consisting of about 2000 accessions of BG (Atoyebi et al., 2017), with 19% of these collected from Nigeria, 456 accessions of AYB with 97% from Nigeria while 100% of the 22 accessions of the Kersting’s groundnut, were collected from Nigeria. Other legumes conserved at the Genetic Resources Center of IITA include winged bean (50 accessions) and other minor legumes, such as Jack bean, Mung bean, Canavalia species, and rice bean, made up of about 18% of the total 25,000 accessions of the seed crops conserved at the center.
The IITA Genebank has been developing a program of research on various aspects of these orphan legumes, particularly BG, AYB, and winged bean, and lately Kersting groundnut to understand their genetic diversity, nutrient composition and to evaluate them for various biotic and abiotic stresses (Paliwal et al., 2021). To identify valuable traits for improving and developing climate-smart varieties, it becomes necessary to conduct genetic diversity studies and quantitative trait loci (QTL) discovery for drought, yield-related traits, and climate-adaptive traits. Accessions of AYB have been evaluated for both nutritional and anti-nutritional properties, the result indicated a good nutritional profile of the seeds with high protein, carbohydrate, and other nutrients when compared with other important legumes (Adegboyega et al., 2020), while the assessment of the genetic variation of AYB accessions using the established crop descriptors and molecular markers has been carried out for some of the collections (Moyib et al., 2008; Adewale et al., 2014; Ojuederie et al., 2014; Shitta et al., 2016; Nnamani et al., 2019). Studies are ongoing on the seed processing procedure that will enhance the seed’s longevity in storage for both BG and AYB to minimize the high cost of conservation of their germplasm, thereby securing the availability of their PGR for future genetic improvement. The seeds of these crops were collected and stored in genetic resource centers, while the whole plant can be propagated in vitro using tissue culture techniques to ensure their sustainability (Ogunsola et al., 2016).
Major bottlenecks like the “hard-to-cook” factor, low yield, indeterminate growth habit, pest and disease attack, pod shattering, mandatory need for stakes or trellis, low seed supply, among others, cannot be solved at once. However, plant breeders and researchers working on the orphan legumes are to tackle these issues accordingly to improve the species for sustainable utilization. Plant breeders are currently studying how the entire genome sequence of some species such as AYB and BG can hasten their genetic improvement and, hence, better utilization. The genomes of some of these legumes are currently being sequenced by the African Orphan Crops Consortium (AOCC), which seeks to assemble and annotate the genomes of 101 traditional African food crops to improve their nutritional content (Ojuederie et al., 2021). Regardless of the ongoing efforts to see to the complete success of orphan legumes in the global food basket, the need for more crop improvement strategies is demanding, not just from conventional plant breeders alone, but biochemists, plant geneticists, botanists, technology providers, the government, and the traditional farmers. They have in-depth knowledge about these minor legumes. Presently, the impact of morphological markers in analyzing the diversity among germplasms of various under-exploited legumes is minimal since environmental conditions influence results. Future directions lie in the use of genome-editing tools, e.g., the Clustered Regularly Interspaced Short Palindromic Repeats (CRISPR), GBS (Genotyping by sequencing), High-throughput phenotyping, SNP (Single nucleotide polymorphism), CNV (Copy number variation), cloning, transformation, and core set development, to get improved traits. Exploring the genomics of these underutilized crops via conventional breeding, Genome-Wide Association Studies (GWAS), modern biotechnology, and computational technology, will hasten the species’ improvement for wide acceptability and utilization (Paliwal et al., 2021). In recent years, the development of large-scale genomic and genetic resources, including simple sequence repeat, expressed sequence tags, diversity array technology markers (DArT), and draft genomes, have enhanced genetic knowledge on these minor species (Varshney et al., 2009; Chang et al., 2019). These discoveries can accelerate gene discovery and pioneer breeding at the molecular level in these under-exploited crops.
The conventional method of producing plants with improved traits relied on time-consuming and laborious approaches. It could take up to 8–12 generations to obtain an improved crop with the desired trait, but other genes could also be transferred in the process. Thus, the use of an alternative method such as modern biotechnology has been considered by several researchers for the improvement of orphan legumes. However, most of the studies that have been conducted were based on the use of DNA or molecular markers for genetic diversity studies and marker-assisted breeding of these underutilized species. Molecular markers are segments of DNA revealing variations, which can be used to detect polymorphism between different genotypes or alleles of a gene for a particular sequence of DNA in a population or gene pool (Jiang, 2013). These molecular markers are short sequences of nucleotides positioned beside the DNA sequence of the desired gene, thus, they are genetically linked and can be transferred from one generation to the next (Abu et al., 2021). The polymorphisms present in molecular markers which arise from alteration of nucleotides or mutations within the genomic loci make them useful for the identification of genetic differences between individual organisms and assessment of the relationship between breeds (Igwe, 2021). Different molecular markers have been used to assess the genetic diversity within the germplasm of orphan legumes (Table 6). The initial molecular markers were PCR-based Restriction Fragment Length Polymorphism (RFLP), Random Amplified DNA (RAPD), Amplified, Fragment Length Polymorphism (AFLP) until the development of sequencing-based molecular markers such as single nucleotide polymorphism (SNP), diversity array technology (DArT) and expressed sequence tags (ESTs) which have provided much useful information for the improvement of orphan legumes. The relative ease and reduced cost of the next-generation sequencing platforms imply that the use of SSR and/or single nucleotide polymorphisms for genetic diversity analysis and association mapping could be supplanted by genotyping by sequencing (Cullis and Kunert, 2017).

Table 6. Molecular markers used for genetic diversity and population studies in orphan legumes.
The identification of quantitative trait loci (QTLs) for several traits in orphan legumes and the development of codominant molecular markers and linkage maps for some orphan crops have been made possible through marker-assisted selection with specific molecular markers. There is an urgent need to link phenotypic data with the genotypic data to appropriately select the landraces of these orphan legumes for genetic improvement.
Using a high-throughput DArTseq genotype-by-sequencing SNP approach, Oluwole et al. (2021b) generated a total of 3.6 k SNPs out of which 2.48 K quality SNPs were used for Genome-Wide Association Study (GWAS) in the AYB population. They identified quantitative trait loci (QTL) for genes that could be useful for the improvement of the protein, oil, and starch contents of AYB. Likewise, 493 SNPs were used for the genotyping of a population of 281 Kersting’s groundnut accessions from Benin using the DArTseqTM approach with about 10.6% of the SNPs found to be aligned to the reference genomes of adzuki bean and mung bean, an indication of an evolutionary relationship of Kersting’s groundnut with adzuki bean and mung bean (Akohoue et al., 2020; Paliwal et al., 2021). Uba et al. (2021) utilized DArTseq and SNP for genetic diversity and population structure studies on some genotypes of BG obtained from Nigeria, Cameroun, West Africa, East, and Southern Africa regions. The analyses of the results revealed that the mean gene diversity was highest 0.478) in Nigeria/Cameroon and West Africa region within the populations across the five regions and revealed the highest Shannon diversity index (0.787) in the West African region. Likewise, it also indicated that among the populations the genotypes in the unknown origin population from the United Kingdom was more closely related to the Western Africa population (0.018), and then the Nigeria/Cameroon populations (0.020) which were all grouped into a single subpopulation which was the largest based on the population structure generated by ADMIXTURE model among the 270 BG genotypes evaluated (Uba et al., 2021). Not much effort has been done in the use of gene technology or modern biotechnology for the genetic improvement of orphan legumes. Though there is public skepticism of the use of modern biotechnology for crop improvement adhering to biosafety measures and guidelines the technology could be very useful for the genetic enhancement of orphan legumes.
Advances in next-generation sequencing have provided a way for a new generation of different omics such as genomics, proteomics, metabolomics, and transcriptomics, which can proffer imperative solutions to these underutilized legumes, enhance crop improvement, and broaden the scope of the various germplasms (Popoola et al., 2020). These multi-omics approaches have been effectively utilized in elucidating growth, senescence, yield, and the responses to environmental stresses in several crops (Yang et al., 2021). Utilization of omics technologies coupled with high-throughput next-generation sequencing platforms, and bioinformatics, have allowed a greater comprehension of the plant system biology as well as the expression of genes and the various metabolic systems in plants (Ojuederie et al., 2022). The ever-increasing decline in the cost of sequencing has initiated the contribution of genomics to the improvement of orphan legumes. However, the use of omics in the characterization of orphan legumes is still in its infancy stage. This is because most of the genomes of these legumes are yet to be sequenced. Nevertheless, efforts are ongoing in the whole genome sequencing of some orphan legumes led by several organizations such as the African Orphan Crops Initiative2 to close the gap yet to be filled by the use of genomics transcriptomics and proteomics to extract useful information for the improvement of orphan legumes.
The sequenced genomes of legumes and the model species such as Glycine max, Vigna unguiculata, and Medicago trunculata, would speed up genomic advancements through comparative genomics in other important orphan legumes such as cluster bean, Dolichos bean AYB, as well as winged bean, which are still being sequenced (Dhaliwal et al., 2020).
Gene editing entails the use of engineered nucleases to instigate cellular DNA repair pathways to make precise, site-directed alterations to an organism’s genome (Bhowmik et al., 2021). Over the years, several genome editing techniques have been developed, including Zinc-finger nucleases (ZFNs), meganucleases, transcription activator-like effector nucleases (TALENs), clustered regularly interspaced short palindromic repeats (CRISPR)-CRISPR-associated protein 9 (Cas9) and homing endonucleases (Gaj et al., 2016). However, CRISPR/Cas9-mediated gene editing has emerged as the most straightforward, adaptable, and precise strategy for genetic manipulation in plants (Bhowmik et al., 2021; Nadakuduti and Enciso-Rodríguez, 2021). The mechanism of action that underpins the CRISPR technology has been extensively reviewed elsewhere (Gaj et al., 2016; Li et al., 2020; Wada et al., 2020; Bhowmik et al., 2021).
While the recalcitrant nature of legumes to in vitro gene transfer, low amenability to transformation, and regeneration efficiencies have posed a significant hurdle to the application of the CRISPR technology, some level of success has been recorded. CRISPR/Cas9-assisted gene editing has been achieved in some legume crops, including Medicago truncatula and Lotus japonicus, cowpea, and soybean (Wang et al., 2017; Zheng et al., 2020; Bhowmik et al., 2021). The success of the technique in these crops shows promise for its cross-application in underexploited legumes (Bhowmik et al., 2021). A critical requirement for improving crops by this technique is in-depth bioinformatic and genomic information, as the technology relies on functionally characterized target genes (Wolter et al., 2019). Primarily, the phytoene desaturase (PDS) gene which encodes an enzyme involved in carotenoid biosynthesis is the most commonly targeted gene in orphan legumes (Badejo, 2018; Venezia and Creasey Krainer, 2021). Furthermore, researchers at the international crops research institute for the semi-arid tropics (ICRISAT) are currently experimenting with the technique to knock down flowering time and photoreceptor genes correlated with photoperiod sensitivity in pigeons pea (ICRISAT, 2020).3 Other traits in orphan legumes that can be targeted for improvement using the CRISPR/Cas system include disease resistance, salinity tolerance, biomass yield, grain yield and quality, and nutrient use efficiency (Rasheed et al., 2021).
Albeit CRISPR provides an unparalleled opportunity to improve traits in the underutilized legumes, the first step remains to establish an efficient transformation protocol for guide RNA (gRNA) delivery and the availability of complete genome sequences in public databases. These are both lacking for the orphan legumes which this review focuses on Mousavi-Derazmahalleh et al. (2019). While the Bioscience eastern and central Africa International Livestock Research Institute (BecA-ILRI) has achieved significant progress in sequencing the entire genome of AYB, the availability of whole-genome sequences and annotations of the underexploited legumes could accelerate the deployment of CRISPR/Cas9 technology to enrich their commercial value, and improve traits associated with photosensitivity, prolonged maturity period, and hard seed coat responsible for long cooking times (ACACIA Africa, n.d.).4
Despite the potential benefits of CRISPR technology to underexploited legumes, regulatory bottlenecks remain a significant matter of concern (Bhowmik et al., 2021). Recently, a ruling by the European Court of Justice declared that targeted mutagenesis via genome editing should be subject to rigorous GMO regulations, even though the product is free of any foreign gene (Wolter et al., 2019; Zaidi et al., 2020). This calls for concerted efforts from policy makers and scientists to design a comprehensive framework for CRISPR-generated crops integration. In addition, large-scale field trials are also required to evaluate the performance of CRISPR-generated crops for traits that may be compromised owing to the disruption of specific genes (Zaidi et al., 2020).
Speed breeding is revolutionizing plant breeding, making it possible to have several generations of crops within a year. It utilizes enhanced light-emitting diode (LED) supplemental lighting and day-long regimes of up to 22 h at temperatures maintained between 17°C and 22°C to enhance photosynthesis and early flowering of the plants, which results in rapid growth and better yield. During the process, the LED lighting increases the breeding cycle of the exposed plants. Scientists at the University of Queensland, Australia, developed this technique. They successfully used it to increase the growth of spring wheat (Triticum aestivum), durum wheat (T. durum), barley (Hordeum vulgare), Chickpea (Cicer arietinum) per year in a temperature-controlled glasshouse fitted with high-pressure sodium lamps for up to six generations and four generations for canola (Brassica napus), in contrast to 2–3 generations under normal glasshouse conditions or field using conventional breeding methods (Watson et al., 2018). They achieved a significant reduction in the time taken for anthesis to occur for all crop species at 22 h photoperiod compared with the 12-h day-neutral photoperiod conditions, with a mean decrease of 22 ± 2 days (wheat), 64 ± 8 days (barley), 73 ± 9 days (canola) and for Chickpea, 33 ± 2 days (Watson et al., 2018). It, therefore, implies that plant breeders would be able to accelerate the rate of genetic improvement of several crops such as wheat, barley, rape, and pea for increased yield, disease resistance, and climate resilience in crops. The neglected and underutilized legumes could also benefit from this modern plant breeding technique, especially as most climate-resilient crops possess a rich nutritional profile.
Gaur et al. (2007) and O’Connor et al. (2013) successfully developed protocols for speed breeding of chickpea (Gaur et al., 2007; Chiurugwi et al., 2018) as well as Arachis hypogea, which resulted in a reduction in the generation time to 89 days from the usual 145 days in the second filial generation hybrids (O’Connor et al., 2013; Chiurugwi et al., 2018). They employed controlled temperature and constant light to fast-track plant development and accelerated a single seed descent (SSD) breeding program. Gaur et al. (2007) was able to achieve three seed-to-seed generations within a year by carrying out two generations under open field conditions and one under rainout shelters by exposing plants to lengthened (24-h) photoperiod (Saxena et al., 2019). Protocols have also been developed to shorten the generation time in some orphan legumes and oilseeds. These include lupin (Lupinus sp.; Croser et al., 2016), chickpea (Cicer arietinum; Sethi et al., 1981; Watson et al., 2018), subterranean clover (Trifolium subterraneum; Pazos-Navarro et al., 2017), lentil (Lens culinaris) as well as broad beans (Vicia faba; Mobini et al., 2015; Lulsdorf and Banniza, 2018). The application of cytokinins and auxins to induce early flowering enabled Mobini et al. (2015) to increase the number of generations to 7 generations for Faba bean (Vicia faba L.) and eight generations for lentil (Lens culinaris Medik), after which immature seeds were harvested for generation advancement (Saxena et al., 2019).
The AYB (Sphenostylis stenocarpa) has recently received more attention as a potential food and nutrition security crop. However, it has a long generation time between 6 and 7 months hence it is grown once a year. Could this dual-purpose crop that produces edible seeds and underground tubers also benefit from speed breeding? Some landraces of AYB are tuber producing while others are not. This characteristic has been linked to the photoperiod sensitivity of the crop, with short photoperiods being suggested as a probable stimulant for tuberization in the species. Using the speed breeding approach, which requires more extended photoperiod of 22 h to shorten the breeding cycle, may not initiate tubers in tuber-producing landraces but may shorten the generation time of the crop. However, a major challenge to the use of speed breeding techniques in Africa is the availability of a constant power supply. In some countries utilizing the technology, containers used for conveying manufactured goods across borders are being converted to a speed breeding facility with LED lights installed and powered by solar panels. Alternatively, a solar-powered glasshouse with LED fitted lighting could be used for speed breeding and adapted for orphan or underutilized legumes in Africa by the National Agricultural Research Centers in the various countries where these legumes are cultivated.
With the speed breeding technique, plant breeders and plant molecular geneticists would hasten the genetic variation inherent in wild relatives of these orphan legumes, thereby introducing elite varieties that will be widely accepted and grown by farmers. The ongoing discoveries of the vast genetic resources and nutraceutical components of these lesser-known legumes are increasingly attracting research and funding attention as potential future crops for sustainable agriculture by CGIAR, IITA, and other Genebank institutes around the world. International agricultural organizations such as the West Africa Centre for Crop Improvement (Ghana), the BeCa-ILRI Hub (Kenya), the World Vegetable Center (Taiwan), the African Orphan Crops Consortium (Kenya), Crops for the Future (Malaysia), the Global Pulse Confederation (UAE) and the CGIAR Centres and Research Programmes like the International Crops Research Institute for the Semi-Arid Tropics (ICRISAT), the International Institute of Tropical Agriculture (IITA), and the International Center for Agricultural Research in the Dry Areas are well-positioned to establish speed breeding facilities to give a boost to the breeding programs of these orphan legumes for enhanced utilization and food and nutrition security. Despite the potential of speed breeding in reducing the breeding cycle in crops, it has some limitations. The success of speed breeding depends on the crop and species and proper regulation of environmental factors such as the photoperiod, temperature, light intensity, and relative humidity. It may also be challenging to utilize speed breeding for short-day plants that require less light for floral induction. Simulating breeding set-ups that combine rapid generation advances with other technologies like genomic selection (GS) would be an excellent approach to optimize breeding methods cost-effectively (Hickey et al., 2019). Genomic selection (GS) accelerates the selection accuracy of superior genotypes, germplasm enhancement and aids in selecting targeted genes for particular traits from Genebank accession to elite lines. Thus, the genetic gain of key traits can be accelerated using integrative approaches of speed breeding and genomic selection, permitting more rapid production of improved cultivars.
The need to introduce these orphan legume crops into the food systems in Africa and other poverty-prone countries to attain zero hunger, improved nutrition, and sustainable agriculture goals of the SDG by 2030 is nonnegotiable. Their dense network of cheap, high, and quality protein, dietary fiber, vitamins, minerals, carbohydrates, low fat, the presence of diverse nutraceuticals and pharmaceutical components that can help prevent or alleviate diet-mediated disease cannot be overemphasized. Without any doubt, the inclusion of underutilized legumes into the dietary cycle could offer long-lasting solution to the high prevalence of malnutrition, hunger, and protein deficiencies in SSA malnutrition, hunger, and protein deficiency, will be a long-lasting solution due to its dense nutrient content, good quality protein, and micronutrients. Currently, the potentials of these minor legumes have begun to receive scientific attention. The target is to improve crop yield, eradicate toxic ANTs, reduce the cooking time of the seeds, speed up maturity rate, gain consumer acceptance, and solve food and protein insecurity. Efforts to elevate these nutrient-rich crops to the top of the food basket of any nation are demanding. However, modern technologies such as speed breeding and CRISPR/Cas9 in an integrative approach with genomic selection and other high-throughput methods could fast track the breeding cycle of orphan legumes to develop new varieties with traits that enable more than one cycle of cultivation per year. Still, with the various efforts being made worldwide, it is hoped that these legumes will contribute immensely to the food and nutritional security of most households and may likely become staple crops for Africans in the not-so-distant future.
JOP – conceived the idea, wrote the first draft, search the literature, and reviewed the manuscript. OSA and OBO – contributed to the writing and review of the manuscript. BDA – contributed to the writing and review. OCA – literature search and contributed to the writing. DIE – literature search. OAO – contributed to the writing of the manuscript. TTA – contributed to the writing. OOO – reviewed and approved the final manuscript. All authors contributed to the article and approved the submitted version.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
We acknowledge Covenant University Centre for Research, Innovation, and Discovery (CUCRID) for defraying the publication fees.
1. ^https://www.tridge.com/intelligences/butter-bean/export
2. ^http://africanorphancrops.org
3. ^https://www.icrisat.org/cgiar-focus-on-gene-editing-crops-for-a-food-and-income-secure-future/
4. ^https://acaciaafrica.org/bioinformatics-community-practice/full-genome-sequencing-and-annotation-of-the-african-yam-bean/
Abu, N. E., Ugwu, E. U., and Ojuederie, O. B. (2021). “The relevance of plant breeding to food security in Africa,” in Food Security and Safety African Perspective. (ed) O. O. Babalola (Switzerland: Springer), 495–527.
ACACIA Africa (n.d.). Project 1: Full-genome sequencing and annotation of the African yam bean (Sphenostylis stenocarpa). Available at: https://acaciaafrica.org/bioinformatics-community-practice/full-genome-sequencing-andannotation-of-the-african-yam-bean/ (Accessed February 5, 2021).
Adegboyega, T. T., Abberton, M. T., AbdelGadir, A. H., Dianda, M., Maziya-Dixon, B., Oyatomi, O. A., et al. (2020). Evaluation of nutritional and anti-nutritional properties of African yam bean (Sphenostylis stenocarpa (Hochst ex. A. Rich.) harms.) seeds. J. Food Qual. 2020:6569420. doi: 10.1155/2020/6569420
Adeleke, O. R., Adiamo, O. Q., and Fawale, O. S. (2018). Nutritional, physicochemical, and functional properties of protein concentrate and isolate of newly-developed Bambara groundnut (Vigna subterrenea L.) cultivars. Food Sci. Nutr. 6, 229–242. doi: 10.1002/fsn3.552
Adeleke, M. A., and Haruna, I. M. (2012). Residual nitrogen contributions from grain legumes to the growth and development of succeeding maize crop. ISRN Agron 2012:5. doi: 10.5402/2012/213729
Adeleye, O. O., Awodiran, S. T., Ajayi, A. O., and Ogunmoyela, T. F. (2020). Influence of extrusion cooking on physicochemical properties and starch digestion kinetics of Sphenostylis stenocarpa, Cajanus cajan, and Vigna subterranean grains. PLoS One 15:e0242697. doi: 10.1371/journal.pone.0242697
Adesoye, A. I., and Nnadi, N. C. (2011). Mitotic chromosome studies of some accessions of African yam bean Sphenostylis stenocarpa (Hochst. Ex. A. Rich.) harm. African J. Plant Sci. 5, 835–841. doi: 10.5897/AJPS11.199
Adewale, D. B., and Adegbite, A. E. (2018). Investigation of the breeding mechanism of African yam bean [Fabaceae](Sphenostylis stenocarpa Hochst. Ex. A. Rich) harms. Not. Sci. Biol. 10, 199–204. doi: 10.15835/nsb10210236
Adewale, D., and Dumet, D. (2010). AYB: A crop with food security potentials for Africa. Afr. Technol. Dev. Forum j. 6, 66–71.
Adewale, B., Vroh-Bi, I., Dumet, D., Nnadi, S., Kehinde, O., Ojo, D., et al. (2014). Genetic diversity in African yam bean accessions based on AFLP markers: towards a platform for germplasm improvement and utilization. Plant Genet. Resour. 13:111. doi: 10.1017/S1479262114000707
Adu-Gyamfi, R., Fearon, J., Bayorbor, T. B., Dzomeku, I. K., and Avornyo, V. K. (2011). The status of Kersting's groundnut (Macrotyloma geocarpum [harms] Marechal and Baudet) an underexploited legume in northern Ghana. Agriculture 40, 259–262. doi: 10.5367/2Foa.2011.0050
Akah, N. P., Kunyanga, C. N., Okoth, M. W., and Njue, L. G. (2021). Pulse production, consumption and utilization in Nigeria within regional and global context. Sustain. Agric. Res. 10:48. doi: 10.5539/sar.v10n2p48
Akibode, S., and Maredia, M. (2011). Global and Regional Trends in Production, Trade and Consumption of Food Legume Crops. Report Submitted to SPIA March 27, 2011.
Akohoue, F., Achigan-Dako, E. G., Sneller, C., Van Deynze, A., and Sibiya, J. (2020). Genetic diversity, SNP-trait associations, and genomic selection accuracy in a west African collection of Kersting’s groundnut [Macrotyloma geocarpum (harms) Maréchal & Baudet]. PLoS One 15:e0234769. doi: 10.1371/journal.pone.0234769
Akohoué, F., Sibiya, J., and Achigan-Dako, E. G. (2019). On-farm practices, mapping, and uses of genetic resources of Kersting’s groundnut [Macrotyloma geocarpum (harms) Maréchal et Baudet] across ecological zones in Benin and Togo. Genet. Resour. Crop. Evol. 66, 195–214. doi: 10.1007/s10722-018-0705-7
Akpapunam, M. A., and Sefa-Dedeh, S. (1997). Jack bean (Canavalia ensiformis): nutrition related aspects and needed nutrition research. Plant Foods Hum. Nutr. 50, 93–99. doi: 10.1007/BF02436029
Alexandratos, N., and Bruinsma, J. (2012). World agriculture towards 2030/2050: the 2012 revision. Food and Agriculture Organization of the United Nations, Rome.
Allito, B. B., Nana, E. M., and Alemneh, A. A. (2015). Rhizobia strain and legume genome interaction effects on nitrogen fixation and yield of grain legume: a review. Mol. Soil Biol. 6, 1–6. doi: 10.5376/msb.2015.06.0002
Amadou, H. I., Bebeli, P. J., and Kaltsikes, P. J. (2001). Genetic diversity in Bambara groundnut (Vigna subterranea L.) germplasm revealed by RAPD markers. Genome 44, 995–999. doi: 10.1139/g01-096
Andueza-Noh, R. H., Serrano-Serrano, M. L., Chacón Sánchez, M. I., Sanchéz del Pino, I., Camacho-Pérez, L., Coello-Coello, J., et al. (2013). Multiple domestications of the Mesoamerican gene pool of lima bean (Phaseolus lunatus L.): evidence from chloroplast DNA sequences. Genet. Resour. Crop. Evol. 60, 1069–1086. doi: 10.1007/s10722-012-9904-9
Aremu, M. O., Olaofe, O. A., and Emmanuel, T. (2006). A comparative study on the chemical and amino acid composition of some Nigerian under-utilized legume flours. Pak. J. Nutr. 5, 34–38. doi: 10.3923/pjn.2006.34.38
Asante, I. K., Offei, S. K., Addy, R., and Carson, A. G. (2008). Phenotypic and seed protein analysis in 31 Lima bean (Phaseolus lunatus) accessions in Ghana, west African. J. Appl. Ecol. 12. doi: 10.4314/wajae.v12i1.45775
Assefa, F. A., and Kleiner, D. (1997). Nodulation of African yam bean (Sphenostylis stenocarpa) by Bradyrhizobium sp. isolated from Erythrina brucei. Biol. Fertil. Soils 25, 209–210. doi: 10.1007/s003740050305
Assogba, P., Ewedje, E.-E. B. K., Dansi, A., Loko, Y. L. E., Adjatin, A., Dansi, M., et al. (2015). Indigenous knowledge and agro-morphological evaluation of the minor crop Kersting’s groundnut [Macrotyloma geocarpum (harms) Maréchal et Baudet] cultivars of Benin. Genet. Resour. Crop. Evol. 63, 513–529. doi: 10.1007/s10722-015-0268-9
Atoyebi, J. O., Osilesi, O., Adebawo, O., and Abberton, M. (2017). Evaluation of nutrient parameters of selected African accessions of Bambara groundnut (Vigna subterranea (L.) Verdc.). Am. J. Food Nutr. 5, 83–89. doi: 10.12691/ajfn-5-3-1
Ayenan, M. A. T., and Ezin, V. A. (2016). Potential of Kersting’s groundnut [Macrotyloma geocarpum (harms) Maréchal & Baudet] and prospects for its promotion. Agric. Food Secur. 5:10. doi: 10.1186/s40066-016-0058-4
Babalola, O. O., Olanrewaju, O. S., Dias, T., Ajilogba, C. F., Kutu, F. R., and Cruz, C. (2017). “Biological nitrogen fixation: the role of underutilized leguminous plants,” in Microorganisms for Green Revolution. eds. D. G. Panpatte, Y. K. Jhala, R. V. Vyas, and H. N. Shelat (Singapore: Springer), 431–443. doi: 10.1007/978-981-10-6241-4_20
Badejo, A. A. (2018). Elevated carotenoids in staple crops: the biosynthesis, challenges and measures for target delivery. J. Genet. Eng. Biotechnol. 16, 553–562. doi: 10.1016/j.jgeb.2018.02.010
Baiyeri, S. O., Uguru, M. I., Ogbonna, P. E., Samuel-Baiyeri, C. C. A., Okechukwu, R., Kumaga, F. K., et al. (2018). Evaluation of the nutritional composition of the seeds of some selected African yam bean (Sphenostylis stenocarpa Hochst ex. A. Rich (harms)) accessions. Agroscience 17, 37–44. doi: 10.4314/as.v17i2.5
Baldermann, S., Blagojević, L., Frede, K., Klopsch, R., Neugart, S., Neumann, A., et al. (2016). Are neglected plants the food for the future? CRC Crit. Rev. Plant Sci. 35, 106–119.
Barman, A., Marak, C. M., Barman, R. M., and Sangma, C. S. (2018). “Nutraceutical properties of legume seeds and their impact on human health,” in Legume Seed Nutraceutical Research. (UK: IntechOpen). doi: 10.5772/intechopen.78799
Baudoin, J. P. (2006). Phaseolus lunatus L. [Internet] Record from PROTA4U. Brink, M. & Belay, G. (Editors). PROTA (Plant Resources of Tropical Africa/Ressources végétales de l’Afrique tropicale), Wageningen, Netherlands. Available at: http://www.prota4u.org/search.asp (Accessed 27 March 2022).
Belel, M. D., Halim, R., Rafii, M., and Saud, H. (2014). Intercropping of corn with some selected legumes for improved forage production: a review. J. Agric. Sci. 6:48. doi: 10.5539/jas.v6n3p48
Bhartiya, A., Aditya, J. P., and Kant, L. (2015). Nutritional and remedial potential of an underutilized food legume horsegram (Macrotyloma uniflorum): a review. J. Anim. Plant Sci. 25, 908–920.
Bhowmik, P., Konkin, D., Polowick, P., Hodgins, C. L., Subedi, M., Xiang, D., et al. (2021). CRISPR/Cas9 gene editing in legume crops: opportunities and challenges. Legum. Sci. 3, 1–16. doi: 10.1002/leg3.96
Bria, E. J., and Bani, P. W. (2021). Genetic diversity of lima bean (Phaseolus lunatus L.) from Timor Island based on molecular marker inter-simple sequence repeats (ISSR). Floribunda 6, 257–263. doi: 10.32556/floribunda.v6i7.2021.354
Caicedo, A. L., Gaitán, E., Duque, M. C., Chica, O. T., Debouck, D. G., and Tohme, J. (1999). AFLP fingerprinting of Phaseolus lunatus L. and related wild species from South America. Crop Sci. 39, 1497–1507. doi: 10.2135/cropsci1999.3951497x
Carbonaro, M., Maselli, P., and Nucara, A. (2015). Structural aspects of legume proteins and nutraceutical properties. Food Res. Int. 76, 19–30. doi: 10.1016/j.foodres.2014.11.007
Chakona, G., and Shackleton, C. (2017). Minimum dietary diversity scores for women indicate micronutrient adequacy and food insecurity status in south African towns. Nutrients 9:812. doi: 10.3390/nu9080812
Chang, Y., Liu, H., Liu, M., Liao, X., Sahu, S. K., Fu, Y., et al. (2019). The draft genomes of five agriculturally important African orphan crops. GigaScience 8. doi: 10.1093/gigascience/giy152
Chapin, F., Zavaleta, E. S., Eviner, V. T., Chapin, F. S., Zavaleta, E. S., Eviner, V. T., et al. (2000). Consequences of changing biodiversity. Nature 405, 234–242. doi: 10.1038/35012241
Charrondière, U. R., Vincent, A., and Grande, F. (2020). “FAO/INFOODS Food Composition Table for Western Africa (2019): User Guide & Condensed Food Composition Table = Table de Composition Des Aliments FAO/INFOODS Pour l’Afrique de l’Ouest (2019): Guide d’utilisation & Table de Composition Des Aliments Condensée.” AGRIS: International Information System for the Agricultural Science and Technology, Available at: https://agris.fao.org/agrissearch/search.do?ecordID=XF2020000998 (Accessed May 2, 2022).
Cheng, A., Raai, M. N., Zain, N. A. M., Massawe, F., Singh, A., and Wan, W. A. A. Q. I. (2019). In search of alternative proteins: unlocking the potential of underutilized tropical legumes. Food Sec. 11, 1205–1215. doi: 10.1007/s12571-019-00977-0
Chiurugwi, T., Kemp, S., Powell, W., and Hickey, L. T. (2018). Speed breeding orphan crops. Theor. Appl. Genet. 132, 607–616. doi: 10.1007/s00122-018-3202-7
Chivenge, P., Mabhaudhi, T., Modi, A. T., and Mafongoya, P. (2015). The potential role of neglected and underutilized crop species as future crops under water scarce conditions in sub-Saharan Africa. Int. J. Environ. Res. Public Health 12, 5685–5711. doi: 10.3390/ijerph120605685
Considine, M. J., Siddique, K. H., and Foyer, C. H. (2017). Nature’s pulse power: legumes, food security and climate change. J. Exp. Bot. 68, 1815–1818. doi: 10.1093/jxb/erx099
Conti, M. V., Campanaro, A., Coccetti, P., De Giuseppe, R., Galimberti, A., Labra, M., et al. (2019). Potential role of neglected and underutilized plant species in improving women’s empowerment and nutrition in areas of sub-Saharan Africa. Nutr. Rev. 77, 817–828. doi: 10.1093/nutrit/nuz038
Coulibaly, M., Idohou, R., Akohoue, F., Peterson, A. T., Sawadogo, M., and Achigan-Dako, E. G. (2022). Coupling genetic structure analysis and ecological-niche modeling in Kersting’s groundnut in West Africa. Sci. Rep. 12:5590. doi: 10.1038/s41598-022-09153-5
Croser, J. S., Pazos-Navarro, M., Bennett, R. G., Tschirren, S., Edwards, K., Erskine, W., et al. (2016). Time to flowering of temperate pulses in vivo and generation turnover in vivo–in vitro of narrow-leaf lupin accelerated by low red to far-red ratio and high intensity in the far-red region. Plant Cell Tissue Org. Cult. 127, 591–599. doi: 10.1007/s11240-016-1092-4
Crujeiras, A. B., Goyenechea, E., and Martínez, J. A. (2010). “Fruit, vegetables, and legumes consumption: role in preventing and treating obesity,” in Bioactive Foods in Promoting Health. eds. R. R. Wason and V. R. Preedy (Elsevier), 359–380. doi: 10.1016/B978-0-12-374628-3.00024-4
Cullis, C., Chimwamurombe, P., Barker, N., Kunert, K., and Vorster, J. (2018). Orphan legumes growing in dry environments: Marama bean as a case study. Front. Plant Sci. 9:1199. doi: 10.3389/fpls.2018.01199
Cullis, C., and Kunert, K. J. (2017). Unlocking the potential of orphan legumes. J. Exp. Bot. 68, 1895–1903. doi: 10.1093/jxb/erw437
Dada, O. A., Faloye, B., and Dumet, J. D. (2013). Evaluation of variability in proximate compositions among accessions of sword bean (Canavalia gladiata Jacq. DC) and Jack bean (Canavalia ensiformis L. DC). Not. Sci. Biol. 5, 98–103. doi: 10.15835/nsb518274
Dahmardeh, M., Ghanbari, A., Syahsar, B., and Ramrodi, M. (2010). The role of intercropping maize (Zea mays L.) and cowpea (Vigna unguiculata L.) on yield and soil chemical properties. Afr. J. Agric. Res. 5, 631–636.
Damfami, A., and Namo, O. A. T. (2020). Bambara groundnut (Vigna subterranea (L.) Verd.): A review of its past, present and future role in human nutrition. J. Agric. Res. 3, 274–281.
Dhaliwal, S. K., Talukdar, A., Gautam, A., Sharma, P., Sharma, V., and Kaushik, P. (2020). Developments and prospects in imperative underexploited vegetable legumes breeding: a review. Int. J. Mol. Sci. 21:9615. doi: 10.3390/ijms21249615
Dixon, J. (2009). From the imperial to the empty calorie: how nutrition relations underpin food regime transitions. Agric. Hum. Values 26:321. doi: 10.1007/s10460-009-9217-6
Duranti, M. (2006). Grain legume proteins and nutraceutical properties. Fitoterapia 77, 67–82. doi: 10.1016/j.fitote.2005.11.008
Fabinin, A. N. (2021). Pulse, export and staple convenience food: market analysis in West Africa. Appl. Econ. 54, 764–773.
FAO (2010). Sustainable Diets and Biodiversity. Directions and Solutions for Policy, Research and Action. Proceedings of the International Scientific Symposium. Rome: FAO.
FAOSTAT (2020). FAO Statistical Databases (Food and Agriculture Organization of the United Nations) 2020 data. Available at: https://www.fao.org/faostat/en/#home (Accessed January 15, 2022).
Fischer, R. A., Byerlee, D. R., and Edmeades, G. O., (2009). Can technology deliver on the yield challenge to 2050? Conference paper by the FAO Expert Meeting on How to Feed the World in 2050 (Rome, 24–26 June 2009). doi: 10.22004/ag.econ.55481
Flyman, M. V., and Afolayan, A. J. (2006). The suitability of wild vegetables for alleviating human dietary deficiencies. S. Afr. J. Bot. 72, 492–497. doi: 10.1016/j.sajb.2006.02.003
Fofana, B., du Jardin, P., and Baudoin, J. P. (2001). Genetic diversity in the Lima bean (Phaseolus lunatus L.) as revealed by chloroplast DNA (cpDNA) variations. Genet. Resour. Crop. Evol. 48, 437–445. doi: 10.1023/A:1012016030318
Fofana, B., Vekemans, X., du Jardin, P., and Baudoin, J. P. (1997). Genetic diversity in Lima bean (Phaseolus lunatus L.) as revealed by RAPD markers. Euphytica 95, 157–165. doi: 10.1023/A:1002910114869
Gaj, T., Sirk, S. J., Shui, S. L., and Liu, J. (2016). Genome-editing technologies: principles and applications. Cold Spring Harb. Perspect. Biol. 8:a023754. doi: 10.1101/cshperspect.a023754
Gaur, P. M., Gowda, C. L. L., Knights, E. J., Warkentin, T. D., Acikgoz, N., Yadav, S. S., et al. (2007). “Breeding achievements,” in Chickpea Breeding and Management. eds. S. S. Yadav, R. J. Redden, W. Chen, and B. Sharma 391–416.
Gepts, P., Beavis, W. D., Brummer, E. C., Shoemaker, R. C., Stalker, H. T., Weeden, N. F., et al. (2005). Legumes as a model plant family. Genomics for food and feed report of the cross-legume advances through genomics conference. Plant Physiol. 137, 1228–1235. doi: 10.1104/pp.105.060871
Gomes, R. L. F., Costa, M. F., Alves-Pereira, A., Bajay, M. M., Viana, J. P. G., Valente, S. E. D. S., et al. (2019). A lima bean core collection based on molecular markers. Sci. Agric. 77. doi: 10.1590/1678-992X-2018-0140
Gulzar, M., and Minnaar, A. (2017). “Underutilized protein resources from African legumes,” in Sustainable Protein Sources. eds. S. R. Nadathur, J. P. D. Wanasundara, and L. Scanlin (Academic Press), 197–208.
Halimi, R. A., Barkla, B. J., Mayes, S., and King, G. J. (2019). The potential of the underutilized pulse bambara groundnut (Vigna subterranea (L.) Verdc.) for nutritional food security. J. Food Compos. Anal. 77, 47–59. doi: 10.1016/j.jfca.2018.12.008
Hasan, M., Uddin, M. K., Mohamed, M. T. M., Zuan, A. T. K., Motmainna, M., and Haque, A. N. A. (2021). Effect of nitrogen and phosphorus fertilizers on growth, yield, nodulation and nutritional composition of Bambara groundnut [Vigna subterranea (L.) Verdc.]. Legume research-An. Int. J. 1:6. doi: 10.18805/LR-617
Hickey, L. T., Amber, N. H., Robinson, H., Jackson, S. A., Leal-Bertioli, S. C. M., Tester, M., et al. (2019). Breeding crops to feed 10 billion. Nat. Biotechnol. 37, 744–754. doi: 10.1038/s41587-019-0152-9
Hillocks, R., Bennett, C., and Mponda, O. (2012). Bambara nut: a review of utilization, market potential and crop improvement. Afr. Crop. Sci. J. 20, 1–16.
Hoste, H., Torres-Acosta, J. F. J., Sandoval-Castro, C. A., Mueller-Harvey, I., Sotiraki, S., Louvandini, H., et al. (2015). Tannin containing legumes as a model for nutraceuticals against digestive parasites in livestock. Vet. Parasitol. 212, 5–17. doi: 10.1016/j.vetpar.2015.06.026
Hunter, D., Borelli, T., Beltrame, D. M., Oliveira, C. N., Coradin, L., Wasike, V. W., et al. (2019). The potential of neglected and underutilized species for improving diets and nutrition. Planta 250, 709–729. doi: 10.1007/s00425-019-03169-4
Hussin, H., Gregory, P. J., Julkifle, A. L., Sethuraman, G., Tan, X. L., Razi, F., et al. (2020). Enhancing the nutritional profile of noodles with Bambara groundnut (Vigna Subterranea) and Moringa (Moringa Oleifera): A food system approach. Front. Sustain. Food Syst. 4:59. doi: 10.3389/fsufs.2020.00059
Ibeto, A. U., Amoo, I. A., and Balogun, T. M. (2019). Comparative study of the chemical composition of Jack bean (Canavalia ensiformis) and sword bean (Canavalia gladiata) seeds. J. Chem. Res. 1, 246–255.
ICRISAT (2020). CGIAR Focus on Gene Editing Crops for a Food and Income Secure Future. Available at: https://www.icrisat.org/cgiar-focus-on-gene-editing-crops-for-a-food-and-income-secure-future/ (Accessed September 22, 2021).
Igwe, D. O. (2021). “Molecular markers: potential facilitators in plant breeding and Germplasm conservation” in Food Security and Safety: African Perspective (ed) O. O. Babalola (Switzerland: Springer), 611–646.
Ikhajiagbe, B., Ogwu, M. C., Ogochukwu, O. F., Odozi, E. B., Adekunle, I. J., and Omage, Z. E. (2021). The place of neglected and underutilized legumes in human nutrition and protein security in Nigeria. Crit. Rev. Food Sci. Nutr. 1–10. doi: 10.1080/10408398.2020.1871319
Indonesian Food Composition Table (IFCT) (2019). Nutrient contents in Jack Bean (Canavalia ensiformis). Data of Ministry of Health, TKPI, Indonesia.
ITC (2022). International Trade Statistics. Available at: https://www.intracen.org/itc/market-info-tools/trade-statistics/ (Accessed February 15, 2022).
Jahanshiri, E., Goh, E. V., Wimalasiri, E. M., Azam-Ali, S., Mayes, S., and Tengku Mohd Suhairi, T. A. S. (2022). The potential of Bambara groundnut: An analysis for the People’s republic of China. Food Energy Secur. e358. doi: 10.1002/fes3.358
Jiang, G. L. (2013). “Molecular markers and marker-assisted breeding in plants,” in Plant Breeding From Laboratories to Fields. (ed.) S. B. Andersen (UK: IntechOpen) 45–83.
Kafoutchoni, K. M., Agoyi, E. E., Agbahoungba, S., Assogbadjo, A. E., and Agbangla, C. (2021). Genetic diversity and population structure in a regional collection of Kersting’s groundnut (Macrotyloma geocarpum (harms) Maréchal & Baudet). Genet. Resour. Crop. Evol. 68, 3285–3300. doi: 10.1007/s10722-021-01187-4
Kambou, D. J., Nandkangre, H., Ouoba, A., Congo, A. K., Zida, W.-P. M. S. F., Sawadogo, N., et al. (2021). Molecular characterization using microsatellites of Bambara nut (Vigna subterranea [L] Verdcourt) landraces cultivated in Burkina Faso. Agric. Sci. 12, 1119–1128. doi: 10.4236/as.2021.1210072
Kanetro, B., Riyanto, M., Pujimulyani, D., and Huda, N. (2021). Improvement of functional properties of Jack bean (Canavalia ensiformis) flour by germination and its relation to amino acids profile. Curr. Res. Nutr. Food Sci. 9:812. doi: 10.12944/CRNFSJ.9.3.09
Khan, M. M. H., Rafii, M. Y., Ramlee, S. I., Jusoh, M., and Al Mamun, M. (2021a). Genetic analysis and selection of Bambara groundnut (Vigna subterranea [L.] Verdc.) landraces for high yield revealed by qualitative and quantitative traits. Sci. Rep. 11, 1–21. doi: 10.1038/s41598-021-87039-8
Khan, M. M. H., Rafii, M. Y., Ramlee, S. I., Jusoh, M., Al Mamun, M., and Halidu, J. (2021b). DNA fingerprinting, fixation-index (Fst), and admixture mapping of selected Bambara groundnut (Vigna subterranea [L.] Verdc.) accessions using ISSR markers system. Sci. Rep. 11:14527. doi: 10.1038/s41598-021-93867-5
Khan, M. M., Rafii, M. Y., Ramlee, I. S., Jusoh, M., and Al-Mamun, S. (2021c). Bambara groundnut (Vigna subterranea L. Verdc): A crop for the new millennium, its genetic diversity, and improvements to mitigate future food and nutritional challenges. Sustainability 15, 1–27. doi: 10.1038/s41598-021-93867-5
Khan, M. M. H., Rafii, M. Y., Ramlee, S. I., Jusoh, M., and Mamun, A. (2020). Genetic variability, heritability, and clustering pattern exploration of Bambara groundnut (Vigna subterranea L. Verdc) accessions for the perfection of yield and yield-related traits. Biomed. Res. Int. 2020:2195797. doi: 10.1155/2020/2195797
Konate, M. N., Nandkangré, H., Ouoba, A., Zida, S. F., Ouedraogo, M., Sawadogo, N., et al. (2019). Genetic diversity of Bambara groundnut accessions from Burkina Faso using random amplified polymorphic DNA markers. Agric. Food Sci. J. Ghana 12, 1050–1059.
Kumari, P., Rastogi, A., and Yadav, S. (2020). Effects of heat stress and molecular mitigation approaches in orphan legume, chickpea. Mol. Biol. Rep. 47, 4659–4670. doi: 10.1007/s11033-020-05358-x
Li, X., and Siddique, K. H. (2020). Future smart food: harnessing the potential of neglected and underutilized species for zero hunger. Matern. Child Nutr. 16:e13008. doi: 10.1111/mcn.13008
Li, H., Yang, Y., Hong, W., Huang, M., Wu, M., and Zhao, X. (2020). Applications of genome editing technology in the targeted therapy of human diseases: mechanisms, advances and prospects. Sig. Transduct. Target Ther. 5:1. doi: 10.1038/s41392-019-0089-y
Liu, M., Ding, M., Chen, L., Ouyang, K., Hui, W., Li, J., et al. (2014). Genetic diversity and relationships among Canavalia ensiformis (L.) DC. Accessions as revealed by sequence-related amplified polymorphism markers. Biochem. Syst. Ecol. 57, 242–249. doi: 10.1016/j.bse.2014.08.005
Lulsdorf, M. M., and Banniza, S. (2018). Rapid generation cycling of an F2 population derived from a cross between Lens culinaris Medik. and Lens ervoides (Brign.) Grande after aphanomyces root rot selection. Plant Breed. 137, 486–491. doi: 10.1111/pbr.12612
Mabhaudhi, T., Chimonyo, V. G., Chibarabada, T. P., and Modi, A. T. (2017). Developing a roadmap for improving neglected and underutilized crops: a case study of South Africa. Front. Plant Sci. 8:2143. doi: 10.3389/fpls.2017.02143
Mabhaudhi, T., Chimonyo, V. G. P., Hlahla, S., Massawe, F., Mayes, S., Nhamo, L., et al. (2019). Prospects of orphan crops in climate change. Planta 250, 695–708. doi: 10.1007/s00425-019-03129-y
Majola, N. G., Gerrano, A. S., and Shimelis, H. (2021). Bambara groundnut (Vigna subterranea [L.] Verdc.) production, utilization and genetic improvement in sub-Saharan Africa. Agronomy 11:1345. doi: 10.3390/agronomy11071345
Maphosa, Y., and Jideani, V. A., (2017). “The role of legumes in human nutrition,” in Functional Food-Improve Health through Adequate Food. (ed.) M. C. Hueda (UK: Intechopen) 1:13.
Martínez-Castillo, J., Zizumbo-Villarreal, D., Gepts, P., Delgado-Valerio, P., and Colunga-GarcíaMarín, P. (2006). Structure and genetic diversity of wild populations of Lima bean (Phaseolus lunatus L.) from the Yucatan peninsula, Mexico. Crop Sci. 46, 1071–1080. doi: 10.2135/cropsci2005.05-0081
Massawe, F. J., Dickinson, M., Roberts, J. A., and Azam-Ali, S. N. (2002). Genetic diversity in bambara groundnut (Vigna subterranea (L.) Verdc) landraces revealed by AFLP markers. Genome 45, 1175–1180. doi: 10.1139/g02-093
Mayes, S., Ho, W. K., Chai, H. H., Gao, X., Kundy, A. C., Mateva, K. I., et al. (2019). Bambara groundnut: an exemplar underutilized legume for resilience under climate change. Planta 250, 803–820. doi: 10.1007/s00425-019-03191-6
Minnaar-Ontong, A., Gerrano, A. S., and Labuschagne, M. T. (2021). Assessment of genetic diversity and structure of Bambara groundnut [Vigna subterranea (L.) verdc.] landraces in South Africa. Sci. Rep. 11, 1–9. doi: 10.1038/s41598-021-86977-7
Mobini, S. H., Lulsdorf, M., Warkentin, T. D., and Vandenberg, A. (2015). Plant growth regulators improve in vitro flowering and rapid generation advancement in lentil and faba bean. In Vitro Cell. Dev. Biol. Plant 51, 71–79. doi: 10.1007/s11627-014-9647-8
Mohale, K. C., Belane, A. K., and Dakora, F. D. (2014). Symbiotic N nutrition, C assimilation, and plant water use efficiency in Bambara groundnut (Vigna subterranea L. Verdc) grown in farmers’ fields in South Africa, measured using 15N and 13C natural abundance. Biol. Fertil. Soils 50, 307–319. doi: 10.1007/s00374-013-0841-3
Morris, B. (2003). Bio-functional legumes with nutraceutical, pharmaceutical, and industrial uses. Econ. Bot. 57:254. doi: 10.1663/0013-0001(2003)057[0254:BLWNPA]2.0.CO;2
Mousavi-Derazmahalleh, M., Bayer, P. E., Hane, J. K., Valliyodan, B., Nguyen, H. T., Nelson, M. N., et al. (2019). Adapting legume crops to climate change using genomic approaches. Plant Cell Environ. 42, 6–19. doi: 10.1111/pce.13203
Moyib, O., Gbadegesin, M., Aina, O., and Odunola, O. (2008). Genetic variation within a collection of Nigerian accessions of African yam bean (Sphenostylis stenocarpa) revealed by RAPD. Primers. Afr. J. Biotechnol. 7, 1839–1846. doi: 10.5897/AJB08.117
Mubaiwa, J., Fogliano, V., Chidewe, C., Bakker, E. J., and Linnemann, A. R. (2018). Utilization of bambara groundnut (Vigna subterranea (L.) Verdc.) for sustainable food and nutrition security in semi-arid regions of Zimbabwe. PLoS One 13:e0204817. doi: 10.1371/journal.pone.0204817
Mubaiwa, J., Fogliano, V., Chidewe, C., and Linnemann, A. R. (2017). Hard-to-cook phenomenon in bambara groundnut (Vigna subterranea (L.) Verdc.) processing: options to improve its role in providing food security. Food Rev. Int. 33, 167–194. doi: 10.1080/87559129.2016.1149864
Muhammad, I., Rafii, M. Y., Ramlee, S. I., Nazli, M. H., Harun, A. R., Oladosu, Y., et al. (2020). Exploration of bambara groundnut (Vigna subterranea (L.) Verdc.), an underutilized crop, to aid global food security: varietal improvement, genetic diversity and processing. Agronomy 10:766. doi: 10.3390/agronomy10060766
Mukakalisa, C., Kandawa-Schulz, M., and Mapaure, I. (2017). “Genetic diversity in landraces of bambara groundnut found in Namibia using RAPD markers.” in II International Symposium on Underutilized Plant Species: Crops for the Future-Beyond Food Security; June 27, 2017; (Malaysia: Kuala Lumpur) 979, 683–687.
Nadakuduti, S. S., and Enciso-Rodríguez, F. (2021). Advances in genome editing with CRISPR systems and transformation technologies for plant DNA manipulation. Front. Plant Sci. 11:2267. doi: 10.3389/fpls.2020.637159
Naluwairo, R. (2011). Promoting Agriculture Sector Growth and Development: A Comparative Analysis of Uganda’s Political Party Manifestos (2011–2016). Kampala, ACODE Policy Research Series, 41.
Nedumaran, S., Abinaya, P., Jyosthnaa, P., Shraavya, B., Rao, P., and Bantilan, C. (2015). “Grain legumes production, consumption and trade trends in developing countries. Working Paper Series, No 60.” in ICRISAT Research Program, Markets, Institutions and Policies. Patancheru 502 324, Telangana, India: International Crops Research Institute for the Semi-Arid Tropics. January 10, 2015; 64.
Nnamani, C., Afiukwa, C., Oselebe, H., Igwe, D., Uhuo, C., Idika, K., et al. (2019). Genetic diversity of some African yam bean accessions in Ebonyi state assessed using InterSimple sequence repeat (ISSR) markers. J. Und. Leg. 1, 20–33.
Nnamani, C. V., Ajayi, S. A., Oselebe, H. O., Atkinson, C. J., Adewale, D. B., Igwe, D. O., et al. (2018). Updates on nutritional diversity in Sphenostylis stenocarpa (Hoechst ex. A. Rich.) harms. For food security and conservation. Am. J. Agric. Biol. Sci. 13, 38–49. doi: 10.3844/ajabssp.2018.38.49
Nnamani, C. V., Ajayi, S. A., Oselebe, H. O., Atkinson, C. J., Igboabuchi, A. N., and Ezigbo, E. C. (2017). Sphenostylis stenocarpa (ex. A. Rich.) harms., a fading genetic resource in a changing climate: prerequisite for conservation and sustainability. Plan. Theory 6:30. doi: 10.3390/plants6030030
Ntundu, W. H., Bach, I. C., Christiansen, J. L., and Andersen, S. B. (2004). Analysis of genetic diversity in bambara groundnut [Vigna subterranea (L.) Verdc] landraces using amplified fragment length polymorphism (AFLP) markers. Afr. J. Biotechnol. 3, 220–225.
O’Connor, D. J., Wright, G. C., Dieters, M. J., George, D. L., Hunter, M. N., Tatnell, J. R., et al. (2013). Development and application of speed breeding technologies in a commercial peanut breeding program. Peanut Sci. 40, 107–114. doi: 10.3146/PS12-12.1
Ogunsola, K. E., Ojuederie, O. B., and Emmanuel, B. (2016). In vitro morphogenic responses of African yam bean (Sphenostylis stenocarpa (Hochst ex. A. Rich.) harms) accessions to plant growth regulators. Plant Cell Tissue Organ Cult. 127, 613–622. doi: 10.1007/s11240-016-1036-z
Ojuederie, O. B., Ajiboye, J. A., and Babalola, O. O. (2020). Biochemical and Histopathological studies of key tissues in healthy male Wistar rats fed on African yam bean seed and tuber meals. J. Food Qual. 2020:8892618. doi: 10.1155/2020/8892618
Ojuederie, O. B., and Balogun, M. O. (2017). Genetic variation in nutritional properties of African yam bean (Sphenostylis stenocarpa Hochst ex. A. Rich. Harms) accessions. NJAFE 13, 180–187.
Ojuederie, O. B., and Balogun, M. O. (2019). African yam bean (Sphenostylis stenocarpa) tubers for nutritional security. J. Und. Leg. 1, 56–68.
Ojuederie, O. B., Balogun, M. O., Fawole, I. I., Igwe, D. O., and Olowolafe, M. O. (2014). Assessment of the genetic diversity of African yam bean (Sphenostylis stenocarpa Hochst ex. A rich. Harms) accessions using amplified fragment length polymorphism (AFLP) markers. Afr. J. Biotechnol. 13, 1850–1858. doi: 10.5897/AJB2014.13734
Ojuederie, O. B., Igwe, D. O., and Popoola, J. O. (2022). “Transgenic plant-mediated phytoremediation: applications, challenges, and prospects,” in Assisted Phytoremediation. (ed) V. C. Pandey (USA: Elsevier), 179–202.
Ojuederie, O. B., Popoola, J. O., Aremu, C., and Babalola, O. O. (2021). “Harnessing the hidden treasures in African yam bean (Sphenostylis stenocarpa), an underutilized grain legume with food security potentials,” in Food Security and Safety. ed. O. O. Babalola (Cham: Springer).
Okpara, D. A., and Omaliko, C. P. E. (1997). Response of African yambean (Sphenostylis stenocarpa) to sowing date and plant density. Indian J. Agric. Sci. 67, 220–221.
Olanrewaju, O. S., Oyatomi, O., Babalola, O. O., and Abberton, M. (2022). Breeding potentials of Bambara groundnut for food and nutrition security in the face of climate change. Front. Plant Sci. 12:798993. doi: 10.3389/fpls.2021.798993
Oluwole, O. O., Aworunse, O. S., Aina, A. I., Oyesola, O. L., Popoola, J. O., Oyatomi, O. A., et al. (2021a). A review of biotechnological approaches towards crop improvement in African yam bean (Sphenostylis stenocarpa Hochst. Ex A. rich.). Heliyon 7:e08481. doi: 10.1016/j.heliyon.2021.e08481
Oluwole, O. O., Olomitutu, O. E., Paliwal, R., Oyatomi, O. A., Abberton, M. T., and Obembe, O. O. (2021b). Preliminary assessment of the association between DArT-SEQ SNP and some nutritional traits in African yam bean. Trop. J. Nat. Prod. Res. 4, 877–879.
Onuche, U., Ibitoye, S. J., and Anthony, T. (2020). Profitability and efficiency Of Bambara groundnut production In Nigeria: A case study. Review of Agricultural and Applied Economics 23, 92–101. doi: 10.15414/raae.2020.23.02.92-101
Onwuka, C. F., Ikewuchi, C. C., Ikewuchi, C. J., and Ayalogu, O. E. (2009). Effect of germination on the performance characteristics of African yam bean (Sphenostylis stenocarpa Hochst ex A rich) seed meal on albino rats. J. Appl. Sci. Environ. Manag. 13, 59–61.
Padulosi, S., Eyzaquirre, P., and Hodgkin, T. (1999). “Challenges and strategies in promoting conservation and use of neglected and underutilized crop species,” in Perspectives on New Crops and New Uses. (ed.) J. Janick (Alexandria, VA: ASHS Press) 140–145.
Paliwal, R., Adegboyega, T. T., Abberton, M., Faloye, B., and Oyatomi, O. (2021). Potential of genomics for the improvement of underutilized legumes in sub-Saharan Africa. Legum. Sci. 3, 1–16. doi: 10.1002/leg3.69
Pazos-Navarro, M., Castello, M., Bennett, R. G., Nichols, P., and Croser, J. (2017). In vitro assisted single-seed descent for breeding-cycle compression in subterranean clover (Trifolium subterraneum L.). Crop Pasture Sci. 68, 958–966. doi: 10.1071/CP170.67
Penha, J. S., Lopes, A. C. A., Gomes, R. L. F., Pinheiro, J. B., Assunção Filho, J. R., Silvestre, E. A., et al. (2017). Estimation of natural outcrossing rate and genetic diversity in Lima bean (Phaseolus lunatus L. var. lunatus) from Brazil using SSR markers: implications for conservation and breeding. Genet. Resour. Crop. Evol. 64, 1355–1364. doi: 10.1007/s10722-016-0441-9
Popoola, J. O., Adegbite, A. E., and Obembe, O. O. (2011). Cytological studies on Some accessions of African yam bean (AYB) (Sphenostylis stenocarpa Hochst. Ex. A. Rich. Harms). Int. Res. J. Plant Sci. 2, 249–253.
Popoola, J., Ojuederie, O., Omonhinmin, C., and Adegbite, A. (2020). “Neglected and underutilized legume crops: improvement and future prospects,” in Recent Advances in Grain Crops Research. eds. F. Shah, Z. Khan, A. Iqbal, M. Turan, and M. Olgun (UK: IntechOpen), 123–144. doi: 10.5772/intechopen.87069
Rasheed, A., Gill, R. A., Hassan, M. U., Mahmood, A., Qari, S., Zaman, Q. U., et al. (2021). A critical review: recent advancements in the use of CRISPR/Cas9 technology to enhance crops and alleviate global food crises. Curr. Issues Mol. Biol. 43, 1950–1976. doi: 10.3390/cimb43030135
Rebello, C. J., Greenway, F. L., and Finley, J. W. (2014). A review of the nutritional value of legumes and their effects on obesity and its related co-morbidities. Obes. Rev. 15, 392–407. doi: 10.1111/obr.12144
Rezende, A. A., Pacheco, M. T. B., da Silva, V. S. N., and de Castro Ferreira, T. A. P. (2018). Nutritional and protein quality of dry Brazilian beans (Phaseolus vulgaris L.). Food Sci. Technol. 38, 421–427. doi: 10.1590/1678-457X.05917
Rungnoi, O., Suwanprasert, J., Somta, P., and Srinives, P. (2012). Molecular genetic diversity of Bambara groundnut (Vigna subterranea L. Verdc.) revealed by RAPD and ISSR marker analysis. SABRAO J. Breed. Genet. 44, 87–101.
Salvi, J., and Katewa, S. S. (2016). A review: underutilized wild edible plants as a potential source of alternative nutrition. Int. J. Botany Stud. 1, 32–36.
Saxena, K. B., Saxena, R. K., Hickey, L. T., and Varshney, R. K. (2019). Can a speed breeding approach accelerate genetic gain in pigeonpea? Euphytica 215:202. doi: 10.1007/s10681-019-2520-4
Serrano-Serrano, M. L., Hernández-Torres, J., Castillo-Villamizar, G., Debouck, D. G., and Chacón Sánchez, M. I. (2010). Gene pools in wild Lima bean (Phaseolus lunatus L.) from the Americas: evidences for an Andean origin and past migrations. Mol. Phylogenet. Evol. 54, 76–87. doi: 10.1016/j.ympev.2009.08.028
Serventi, L., and Dsouza, L. V. (2020). “Bioactives in legumes,” in Upcycling Legume Water: From Wastewater to Food Ingredients. (ed). L. Serventi (Cham: Springer International Publishing), 139–153.
Sethi, S. C., Byth, D. E., Gowda, C. L. L., and Green, J. M. (1981). Photoperiodic response and accelerated generation turnover in chickpea. Field Crop Res. 4, 215–225. doi: 10.1016/0378-4290(81)90073-3
Shitta, N. S., Abberton, M. T., Adesoye, A. I., Adewale, D. B., and Oyatomi, O. (2016). Analysis of genetic diversity of African yam bean using SSR markers derived from cowpea. Plant Genet. Resour. 14, 50–56. doi: 10.1017/S1479262115000064
Singh, S., and Fernandes, M. (2018). Home-grown school feeding: promoting local production systems diversification through nutrition sensitive agriculture. Food Secur. 10, 111–119. doi: 10.1007/s12571-017-0760-5
Singh, B., Singh, J. P., Singh, N., and Kaur, A. (2017). Saponins in pulses and their health promoting activities: a review. Food Chem. 233, 540–549. doi: 10.1016/j.foodchem.2017.04.161
Sita, K., Sehgal, A., HanumanthaRao, B., Nair, R. M., Vara Prasad, P. V., Kumar, S., et al. (2017). Food legumes and rising temperatures: effects, adaptive functional mechanisms specific to reproductive growth stage and strategies to improve heat tolerance. Front. Plant Sci. 8:1658. doi: 10.3389/fpls.2017.01658
Sprent, J. I., Odee, D. W., and Dakora, F. D. (2010). African legumes: a vital but under-utilized resource. J. Exp. Bot. 61, 1257–1265. doi: 10.1093/jxb/erp342
Suneja, Y., Kaur, S., Gupta, A. K., and Kaur, N. (2011). Levels of nutritional constituents and anti-nutritional factors in black gram (Vigna mungo L. Hepper). Food Res. Int. 44, 621–628. doi: 10.1016/j.foodres.2010.12.020
Tan, X. L., Azam-Ali, S., Goh, E. V., Mustafa, M., Chai, H. H., Ho, W. K., et al. (2020). Bambara groundnut: An underutilized leguminous crop for global food security and nutrition. Front. Nutr. 7:601496. doi: 10.3389/fnut.2020.601496
Tridge Statistics (2022). Lima bean (Phaseolus spp). Available at: https://www.tridge.com/intelligences/butter-bean/export (Accessed February 15, 2022).
Tsamo, A. T., Mohammed, M., Ndibewu, P. P., and Dakora, F. D. (2019). Identification and quantification of anthocyanins in seeds of Kersting’s groundnut [Macrotyloma geocarpum (harms) Marechal & Baudet] landraces of varying seed coat pigmentation. J. Food Meas. Charact. 13, 2310–2317. doi: 10.1007/s11694-019-00150-3
Uba, C. U., Oselebe, H. O., Tesfaye, A. A., and Abtew, W. G. (2021). Genetic diversity and population structure analysis of bambara groundnut (Vigna subterrenea L) landraces using DArT SNP markers. PLoS One 16:e0253600. doi: 10.1371/journal.pone.0253600
Udeh, E. L., Nyila, M. A., and Kanu, S. A. (2020). Nutraceutical and antimicrobial potentials of Bambara groundnut (Vigna subterranean): A review. Heliyon 6:e05205. doi: 10.1016/j.heliyon.2020.e05205
Varshney, R. K., Close, T. J., Singh, N. K., Hoisington, D. A., and Cook, D. R. (2009). Orphan legume crops enter the genomics era! Curr. Opin. Plant Biol. 12, 202–210. doi: 10.1016/j.pbi.2008.12.004
Venezia, M., and Creasey Krainer, K. M. (2021). Current advancements and limitations of gene editing in orphan crops. Front. Plant Sci. 12:742932. doi: 10.3389/fpls.2021.742932
Vidigal, P., Romeiras, M. M., and Monteiro, F. (2019). “Crops diversification and the role of orphan legumes to improve the sub-Saharan Africa farming systems,” in Crop Production. (ed). M. Hasanuzzaman (UK: IntechOpen), 1–21.
Wada, N., Ueta, R., Osakabe, Y., and Osakabe, K. (2020). Precision genome editing in plants: state-of-the-art in CRISPR/Cas9-based genome engineering. BMC Plant Biol. 20:234. doi: 10.1186/s12870-020-02385-5
Wang, L., Wang, L., Zhou, Y., and Duanmu, D. (2017). Use of CRISPR/Cas9 for symbiotic nitrogen fixation research in legumes. Prog. Mol. Biol. Transl. Sci. 149, 187–213. doi: 10.1016/bs.pmbts.2017.03.010
Watson, A., Ghosh, S., Williams, M. J., Cuddy, W. S., Simmonds, J., Rey, M., et al. (2018). Speed breeding is a powerful tool to accelerate crop research and breeding. Nat. Plants 4, 23–29. doi: 10.1038/s41477-017-0083-8
Wokes, F., Badenoch, J., and Sinclair, H. M. (1955). Human dietary deficiency of vitamin B12. Am. J. Clin. Nutr. 3, 375–382. doi: 10.1093/ajcn/3.5.375
Wolter, F., Schindele, P., and Puchta, H. (2019). Plant breeding at the speed of light: the power of CRISPR/Cas to generate directed genetic diversity at multiple sites. BMC Plant Biol. 19:176. doi: 10.1186/s12870-019-1775-1
Yang, Y., Saand, M. A., Huang, L., Abdelaal, W. B., Zhang, J., Wu, Y., et al. (2021). Applications of multi-Omics Technologies for Crop Improvement. Front. Plant Sci. 12:563953. doi: 10.3389/fpls.2021.563953
Zaidi, S. S. A., Mahas, A., Vanderschuren, H., and Mahfouz, M. M. (2020). Engineering crops of the future: CRISPR approaches to develop climate-resilient and disease-resistant plants. Genome Biol. 21:289. doi: 10.1186/s13059-020-02204-y
Keywords: climate change, food security, malnutrition, orphan legumes, sustainable development goals
Citation: Popoola JO, Aworunse OS, Ojuederie OB, Adewale BD, Ajani OC, Oyatomi OA, Eruemulor DI, Adegboyega TT and Obembe OO (2022) The Exploitation of Orphan Legumes for Food, Income, and Nutrition Security in Sub-Saharan Africa. Front. Plant Sci. 13:782140. doi: 10.3389/fpls.2022.782140
Edited by:
Jill Margaret Farrant, University of Cape Town, South AfricaReviewed by:
Pranjal Yadava, Indian Agricultural Research Institute (ICAR), IndiaCopyright © 2022 Popoola, Aworunse, Ojuederie, Adewale, Ajani, Oyatomi, Eruemulor, Adegboyega and Obembe. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Jacob Olagbenro Popoola, amFjb2IucG9wb29sYUBjb3ZlbmFudHVuaXZlcnNpdHkuZWR1Lm5n, orcid.org/0000-0001-5302-4856; Olawole Odun Obembe, b2xhd29sZS5vYmVtYmVAY292ZW5hbnR1bml2ZXJzaXR5LmVkdS5uZw==, orcid.org/0000-0001-9050-8198
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.