
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Plant Sci. , 05 January 2023
Sec. Sustainable and Intelligent Phytoprotection
Volume 13 - 2022 | https://doi.org/10.3389/fpls.2022.1098048
This article is part of the Research Topic New Insights in Small Fruit Diseases View all 5 articles
 Lina Dėnė1*†
Lina Dėnė1*† Kristina Laužikė2†
Kristina Laužikė2† Neringa Rasiukevičiūtė1
Neringa Rasiukevičiūtė1 Simona Chrapačienė1
Simona Chrapačienė1 Aušra Brazaitytė2
Aušra Brazaitytė2 Akvilė Viršilė2
Akvilė Viršilė2 Viktorija Vaštakaitė-Kairienė2
Viktorija Vaštakaitė-Kairienė2 Jurga Miliauskienė2
Jurga Miliauskienė2 Rūta Sutulienė2
Rūta Sutulienė2 Giedrė Samuolienė2
Giedrė Samuolienė2 Alma Valiuškaitė1
Alma Valiuškaitė1Essential oils and extracts are investigated in sustainable plant protection area lately. Alternative antifungal substances are especially relevant for major economic-relevance pathogens, like Botrytis cinerea (causal agent of strawberry grey mold), control. However, the reaction of plants to alternative protection with plant-origin products is currently unknown. Induced stress in plants causes changes in antioxidant and photosynthetic systems. The aim of the research was to determine the defense response of strawberry plants under application of coriander seed products. In the first step of the research, we determined coriander seed (Coriandrum sativum), black seed (Nigella sativa) and peppermint leaf (Menta × piperita) products’ antifungal activity against B. cinerea in vitro. Secondly, we continued evaluation of antifungal activity under controlled environment on strawberry plants of the most effective coriander seed products. Additionally, we evaluated the antioxidant and photosynthetic parameters in strawberries, to examine the response of plants. Antifungal activity on strawberries was determined based on grey mold incidence and severity after application of coriander products. Impact on photosynthetic system was examined measuring photosynthetic rate, transpiration rate, stomatal conductance, and intercellular to ambient CO2 concentration. Strawberry leaves were collected at the end of the experiment to analyze the antioxidant response. The highest antifungal activity both in vitro and on strawberries had coriander seed essential oil, which decreased grey mold severity. Coriander extract increased the photosynthetic capacity and antioxidant response of strawberry plants, however had negative effect on suppression of grey mold. In most cases, the essential oil activated antioxidant response of strawberry plants lower than extract. Our study results provide no direct impact of increased photosynthetic capacity values and antifungal effect after treatment with natural oils. The highest concentrations of coriander essential oil and extract potentially demonstrated a phytotoxic effect.
Healthy fruit-bearing plants and high-quality yield are prioritized in the horticultural market. Currently, in agriculture, a lower usage of chemical pesticides for disease management should be achieved to reduce existing and occurring consequences to the environment and food safety (Nxumalo et al., 2021). Therefore, attention is paid to the investigation of the effectiveness of naturally occurring pesticides from plants against pathogens (Raveau et al., 2020; Soleha et al., 2022). The plant’s defense response is a complex system depending on various elements. Despite decreasing damage to pathogens, natural compounds can also have a growth-enhancing effect (Andresen and Cedergreen, 2010), affect photosynthetic capacity (Yao et al., 2020), and phenolic compounds (Karlund et al., 2014; Wei et al., 2018). An additional value would be provided to the natural compounds by registering not only the antifungal effect but also the impact on other plant factors.
Previous research on antifungal activity shows that there is a demand for non-chemical measures for control of plant pathogens (Zaker and Mosallanejad, 2010; Mohammadi et al., 2014; Zabka et al., 2014; Ikeura and Koabayashi, 2015; Zatla et al., 2017; Moghadam et al., 2019). Coriander leaves and stem extracts showed high antifungal activity against Botryotinia fuckeliana, Glomerella cingulata, Fusarium oxysporum, and Pectobacterium carotovorum subsp. carotovorum (Ikeura and Koabayashi, 2015). The essential oil of coriander had an inhibitory effect on Aspergillus flavus (Lasram et al., 2019), and Leucoagaricus gongylophorus (Morais et al., 2015). Besides, it demonstrated antimicrobial properties for food preservation (Zamindar et al., 2016; El-Sayed et al., 2022), while the anti-yeast effect of the black seed extract was observed (Nadaf et al., 2015). Although there are studies on the effectiveness of coriander, peppermint, and black seed extracts in vitro against fungal pathogens, only a few studies on Botrytis cinerea under in vitro conditions could be found (Mohammadi et al., 2014). Additionally, the comparison of different forms of natural compounds from plants is not widely investigated as well as the effectiveness of the extracts in vivo.
Based on the literature mentioned above, the oils and extracts of coriander, peppermint, and black seed can potentially have antifungal activity against B. cinerea and become a biopesticide for strawberry grey mold. Also, these products demonstrated antioxidant activity (Wangensteen et al., 2004; Zheljazkov et al., 2014). Phenolic compounds and antioxidant activity were mainly investigated in berries but not in the plant itself (Wei et al., 2018; Abd-Elkader et al., 2021). The content of phenolic compounds is one of the main indicators of the response of plants to stress (Elstner et al., 1993; Rivero et al., 2001) and indicate plants natural defense against pathogen (Karlund et al., 2014). Studies show fruits’ total phenolics and antioxidant activity are highly responsive to B. cinerea (Bui et al., 2019; Tzortzakis, 2019). Nevertheless, a preharvest antioxidant response in strawberry plants after treatment with extracts and essential oils was not investigated.
An important primary metabolic process for plants is photosynthesis. It provides energy and material for plant growth and is responsible for plant defense mechanisms. One of the stomata’s vital roles in plants is to regulate gas exchange. However, its channels may also be favorable for pathogen entry (Francesconi and Balestra, 2020). Reducing stomata size (to defend from pathogens) affects gas exchange that leads to inhibition in CO2 assimilation and photosynthetic rate (Ahammed et al., 2021; Geng et al., 2022). Pathogens induce stress in strawberry plants, leading to elevated antioxidant activities (Petrasch et al., 2019; Sekmen Cetinel et al., 2021). In addition, with the help of biological control agents, secondary metabolites in their synthesis and metabolic pathways enhance resistance to stress, biotic and abiotic (Ferreira and Musumeci, 2021). Although there are reports on the antifungal activity of extracts and essential oils and their antioxidant activity itself, so far there is no complex research conducting antifungal activity evaluation together with the impact on the antioxidant system of plants.
Therefore, the aim of the study was to determine the defense response of strawberry plants under application of coriander seed products. To obtain this, firstly, we selected the extract from coriander seed (Coriandrum sativum), black seed (Nigella sativa) and peppermint leaf (Menta × piperita) extracts based on the highest antifungal activity against B. cinerea in vitro. Secondly, we examined the possibilities of strawberry B. cinerea control under controlled environment on strawberries using three different forms (extract, essential oil, and hydrosol) of selected extract (C. sativum) and determined strawberry antioxidant and physiological response.
In this part of study, natural products were prepared and investigated for antifungal activity against B. cinerea in vitro. Further, the most effective extract was prepared in three different forms and investigated for minimal inhibitory concentrations (MIC), which were used in the experiments with strawberry plants.
Coriander seed (Coriandrum sativum), peppermint leaf (Menta × piperita), and black seed (Nigella sativa) CO2 extracts were used for the first part of the evaluation of the antifungal effect against Botrytis cinerea in vitro at different concentrations. Coriander (Coriandrum sativum) essential oil, extract, and hydrosol were used for the second part of the evaluation of the antifungal effect in vitro and on strawberry plants under controlled environment.
Dried plant material: coriander seeds, peppermint leaves, and black seeds were obtained from Origanum (Lithuania). Extracts were produced by the previously published method and parameters of subcritical CO2 extraction (Šernaitė et al., 2020). Meanwhile, essential oil and hydrosol were obtained by hydrodistillation with Clevenger apparatus.
The first part of the experiment in vitro on the antifungal effect of the extracts was carried out by mixing coriander, peppermint, and black seed extracts with potato dextrose agar (PDA) at 0.02-0.20% concentrations and pouring them into Petri dishes. Each Petri dish was inoculated with a 7-day old 7 mm diameter mycelium disc of B. cinerea. The experiment was kept at 22 ± 2°C in the dark for 7 days. The antifungal effect was evaluated by measuring the radial colony growth of the pathogen at 2, 4, and 7 days after inoculation (DAI). The extract, causing the lowest radial colony growth of B. cinerea at investigated concentrations, was considered as the most effective and was used for the determination of MIC.
In the second part of the experiment in vitro, three different forms of the most effective coriander extract were used. Extract, essential oil, and hydrosol of coriander seed were separately mixed with PDA at 0.06-0.18% concentrations. Inoculation and evaluation were made as described in the first part of in vitro experiments. We considered MIC as the lowest concentration with the highest antifungal activity, and MIC of the essential oil as the lowest concentration with less than 1 cm radial colony growth of B. cinerea at 5 DAI. These concentrations were used as a base for the applications on strawberry plants.
Strawberries (cv. Deluxe) were planted, in the 15x15 cm plastic pots with 1:1 black soil and peat soil for vegetable mix (Durpeta, Lithuania) in greenhouse, under supplemental high-pressure sodium lamps lighting (200 μmol m-2 s-1), under 16 h photoperiod. Plants were watered as needed and fertilized every week from 1.5 months after planting, with N 34 (10 g/L) and NPK 14-10-25 (10 g/L) (Baltic Agro). Strawberries were transferred into a controlled environment at 21°C temperature, and 70% humidity 2 months after planting. The acclimatization lasted for 2 days.
24h before the inoculation, the humidity was increased to 100% and plants were covered under agro-film to make suitable conditions for the infection of grey mold. After 24h, spraying mixtures with three coriander seed products were prepared by mixing them with sterilized distilled water at 0.12-1.6% concentrations and 1% Tween 80 as a surfactant (Table 1). Hydrosol and sterilized distilled water mixture (1:1) was used instead of only water in two spraying mixtures (Ext 0.16+h, Eo 0.12+h). Inoculated control (Control inoc) was sprayed only with sterile distilled water. Additionally, non-inoculated and not sprayed control treatment was used in the experiment for comparison (Control 0).

Table 1 Spraying mixtures for strawberry application under controlled environment.
The experiment was arranged in randomized blocks, 3 replicates (five plants) per treatment. 5 ml of prepared mixtures were sprayed on each treatment. Three wounds were made on three different middle leaves of each strawberry trifoliate. 9 mm mycelium plug of B. cinerea was put on each wound. Strawberries were covered under agro-film again and kept at 21°C and 100% humidity for 48h. After 48h, agro-film was removed.
The experiment was evaluated daily, and the first grey mold incidence and severity registered when the first infection symptoms were observed. For each replicate, disease severity in inoculated leaves area firstly was evaluated with scores from 0 to 5, where: 0 – no infection; 1 -<5%; 2 -<10%; 3 -<15%; 4< 20% and 5 – >21% infected (Figure 1), and then scores were used to calculate percentage value using formula (Dabkevičius and Brazauskienė, 2007):

Figure 1 Grey mold severity scale on strawberry leaves suggested by the authors.
where: A – number of inspected leaves, B – number of leaves damaged by grey mold, a – number of leaves with the same damage score, K – the highest score of the scale, Σ – the sum of the products of the number of leaves with the same damage score and the score values.
Disease incidence was calculated as the proportion of infected leaves. All evaluations were made 3, 6, and 8 days after inoculation.
Extracts were prepared by grinding 0,5 g of strawberry leaves with liquid nitrogen and diluting with 5 mL of 80% methanol. Each of three biological replicates consisted of at least five conjugated plants leaves and were repeated in three analytical replicates. Further, biochemical analyses were performed from the prepared extract. The antioxidant assay was carried out as in Perez-Lopez et al. (2014), using the radical cation ABTS (2,2’-azino-di[3-ethylbenzthiazoline sulphonate]) generated following Re et al. (1999). The activity was assayed after the addition of 10 μl of the extract to the radical solution, previously diluted to reach an absorbance value of 0.70 ± 0.05 at 734 nm. After 10 min of reaction, the decrease in absorbance was compared to that caused by Trolox standard solutions in the range 0.2–1.5 mM. The activity was expressed as μmol Trolox equivalent antioxidant capacity (TEAC) g−1 dry weight (DW) of plant material.
The antioxidant activity (by DPPH) of methanol extracts of the investigated plants was evaluated spectrophotometrically relating to the 2,2–diphenyl–1–picrylhydrazyl (DPPH•) free radical scavenging capacity (Ragaee et al., 2006). The absorbance scanned after 16 minutes from the beginning of the reaction at 515 nm was used for the calculation of the ability of seed material to scavenge DPPH• free radicals (μmol g-1).
Total phenols in methanolic extracts were determined as previously reported by Mazzoncini et al. (2015) following the reaction at 735 nm between phenols and the Folin-Ciocalteu reagent. Gallic acid was used as a standard to prepare a calibration curve to quantify phenolics amounts. Total phenols were expressed as gallic acid equivalents (GAE) g−1 DW.
Photosynthetic rate (Pr, μmol CO2 m−2 s−1), transpiration rate (Tr, mmol H2O m−2 s−1), stomatal conductance (gs, mol H2O m−2 s−1), intercellular to ambient CO2 concentration (Ci/Ca) was determined 9:00-12:00 am by using an LI-6400XT portable open flow gas exchange system (Li-COR 6400XT Biosciences, Lincoln, USA). Fully developed healthy leaves near the inoculated leaf from the plant, were chosen for measurements, five plants were measured for one minute. Reference air (CO2 400 μmol mol-1), light intensity (1000 μmol m-2 s-1), and the flow rate of gas pump (500 mmol s-1) were set.
Statistical analysis of antifungal activity data was made using SAS Enterprise Guide 7.1 program (SAS Inc., USA). One-way analysis of variance (ANOVA) procedure was performed. Means of three replicates (n = 3) were compared by Duncan’s multiple range test (p< 0.05). Statistical analysis for plant physiology (photosynthetic and antioxidant systems response) data was performed using Microsoft Excel 2016 and Addinsoft XLSTAT 2022.1 XLSTAT statistical and data analysis (Long Island, NY, USA). The data are presented as means of three replicates (n = 3) linked to the sampling points. One-way (ANOVA) followed by Tukey’s significant difference test (p< 0.05) for multiple comparisons was used to evaluate differences between means of measurements. Multivariate principal component analysis (PCA) was performed to determine the statistical relations between natural product applications, used in the experiment. The results are presented in a three PCA scatterplot that indicates distinct levels of: 1. combined extracts antifungal activity and treated strawberry plants physiological data; 2. only treated strawberry plants physiological data; 3. only extracts antifungal activity data.
Three plant extracts were evaluated for antifungal activity against B. cinerea in vitro. The radial colony growth of B. cinerea under the action of black seed, coriander, and peppermint extracts is presented in Figure 2. The use of 0.16% of coriander and peppermint extracts and 0.20% of coriander extract resulted in the lowest growth of the pathogen at 2 DAI. Coriander extract remained with the highest antifungal activity at 0.16-0.20% concentration range at 4 DAI. The most effective concentration was 0.16%. Peppermint extract lost efficiency at 4 DAI. The lowest antifungal activity was observed with blackseed extract at both DAI.
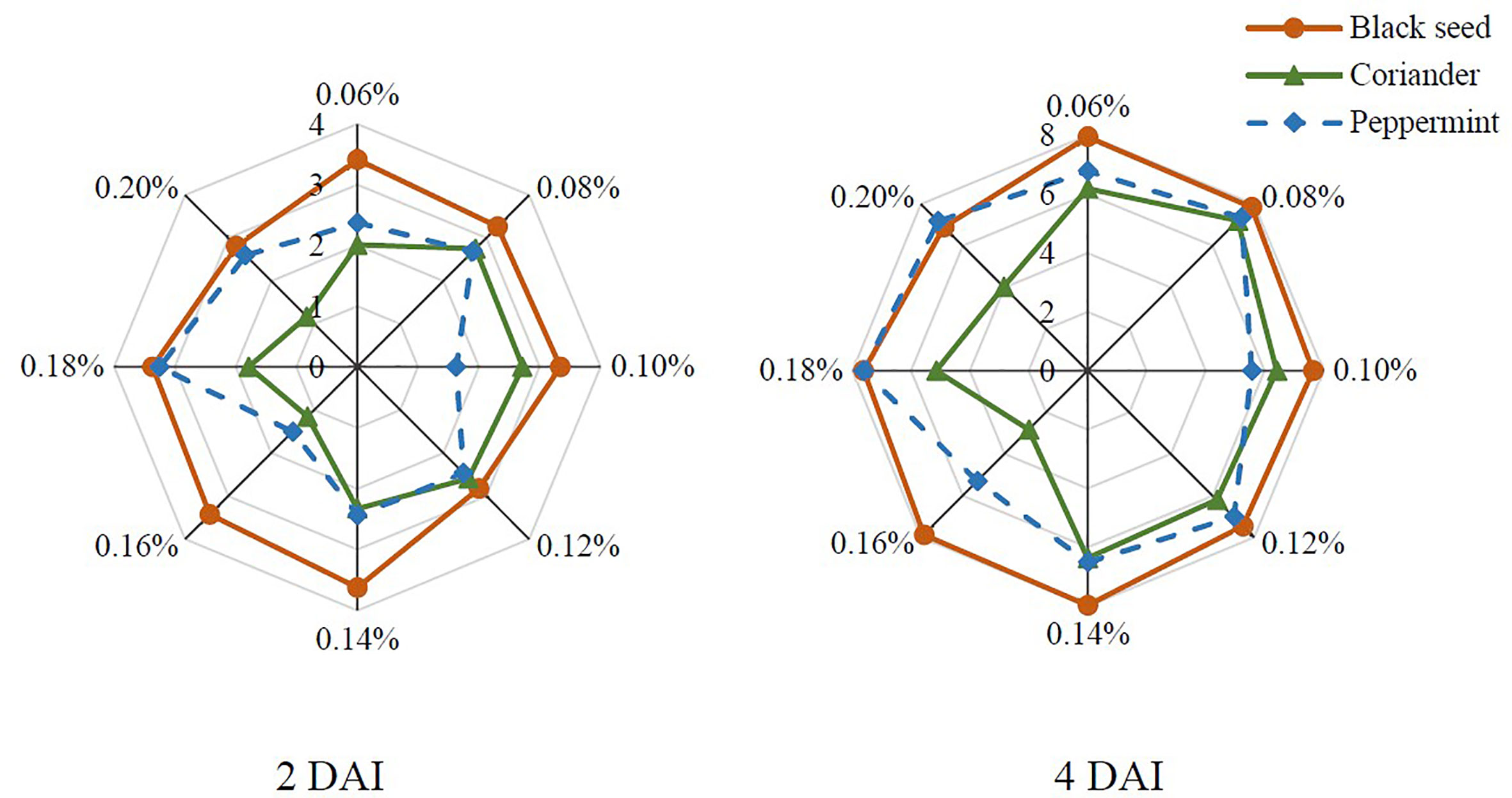
Figure 2 The radial colony growth (cm) of B. cinerea on the PDA with black seed (Nigella sativa), coriander (Coriandrum sativum), and peppermint (Menta × piperita) extracts at different concentrations (%); DAI – days after inoculation.
As coriander extract was observed as the most effective against B. cinerea, in vitro study was continued with three different products from coriander seed. Determined MIC (Table 2) were used for strawberry treatment under controlled environment. Coriander essential oil showed the lowest MIC. Meanwhile, B. cinerea was not inhibited by any investigated concentration of hydrosol.
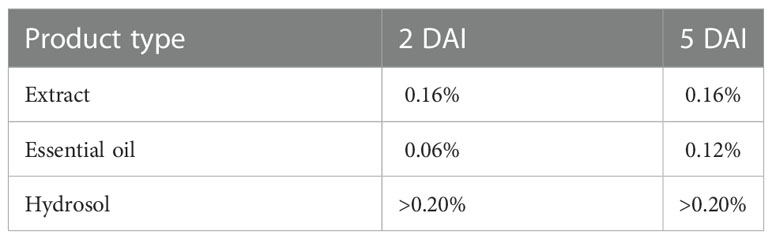
Table 2 Minimal inhibitory concentrations (MIC) of products from coriander seed.
Disease incidence and severity were evaluated to investigate the antifungal activity of natural products from coriander seed against strawberry grey mold induced by B. cinerea. The lowest disease incidence and severity were observed in essential oil treatments Eo 0.12 and Eo 0.12+h at 3 and 6 DAI (Figure 3). Eo 0.12+h and Eo 1.2 treated strawberries had the lowest grey mold parameters at 8 DAI. Treatments with extract (Extr1, Extr1+h, Extr2) were not effective in reducing the infection and had higher disease incidence and severity than control treatment at all DAI. Ext 0.16+h demonstrated the best results compared to the two other extract applications.
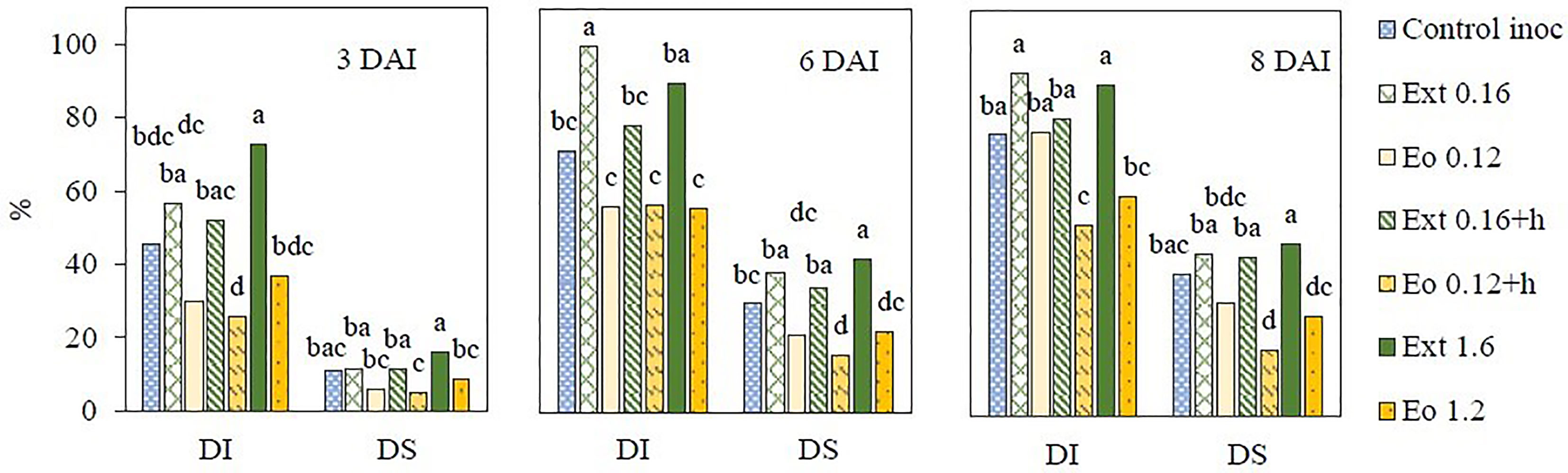
Figure 3 Grey mould incidence (DI, %) and severity (DS, %) on strawberry plants after application of coriander seed products; DAI – days after inoculation. Different letters indicate significant differences between the means of the treatments (Duncan’s multiple range test, p < 0.05).
The radical scavenging activity and total phenolic compounds (TPC) in strawberry plants at 8 DAI are shown in Table 3. Inoculation increased strawberry’s antioxidant system activity up to two times, as well as total phenol content. The highest DPPH, ABTS and TPC were determined in Ext 1.6 and Eo 1.2 treatments. Lower than inoculated control DPPH and ABTS scavenging activity was observed in other treatments except for Ext 0.16+h, which was almost equal to control at ABTS. The highest total phenolic compounds were in Ext 1.6 and Eo 1.2 treatments. Meanwhile, Eo 0.12, Ext 0.16+h and Eo 0.12+h treatments resulted in lower than control treatment TPC.
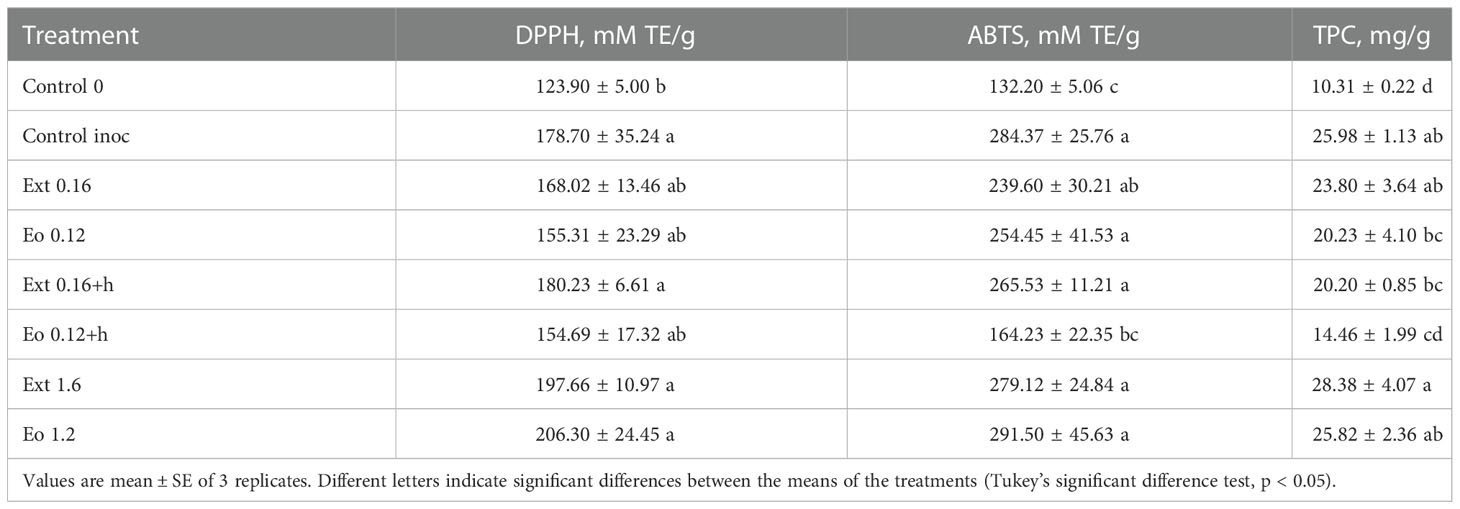
Table 3 Effect of coriander seed products applications on antioxidant system response of strawberry leaves.
Inoculation decreased the photosynthetic rate of strawberries, but increased transpiration compared to not inoculated strawberries in the majority of applications (Table 4). All the applications significantly increased photosynthetic rate except Eo 0.12 and Eo 1.2. Eo 1.2 significantly decreased the photosynthetic rate of strawberry leaves. Meanwhile, Eo 0.12 effect was not significant compared to the control. The Eo 1.2 effect was manifested by the closing of stomata, which caused a drop in transpiration and the photosynthetic rate (Table 4). Ext 0.16 and Ext 0.16+h opened the stomata and increased stomatal conductance up to two times compared to both control treatments. Transpiration increased significantly 26.6 and 48.5% in treatments, respectively, compared to the not inoculated plants. Ext 1.6 and Eo 1.2 decreased intercellular CO2 mostly compared to inoculated plants.
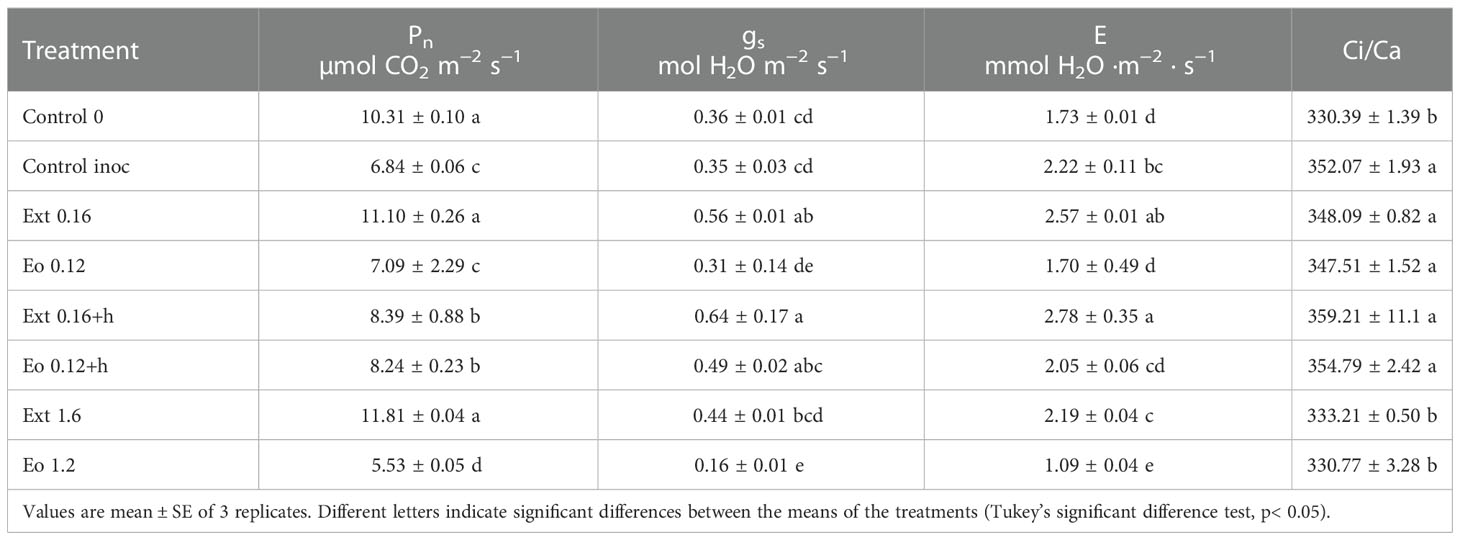
Table 4 Photosynthetic capacity of strawberry leaves: Photosynthetic rate (Pn), Stomatal conductance gs, transpiration rate (E), intercellular CO2 (Ci/Ca).
After evaluating the results of the experiment, the principal component analysis (PCA) covered 79.09% of all data and divided the used applications into separate groups (Figure 4A). The F1 axis covered 59.02% of the data and the applications according to the effect on strawberries were divided into two groups, with essential oils and control in one, and extracts in the other. The F2 axis (covering 20.07% of the data) separated the treatments by concentrations. Analyzing data only from treated strawberry plants’ physiological response, PCA covered 79.65% of all data. Strawberries’ physiological response is divided into similar groups: one extracts and inoculated control and the second one essential oil and not inoculated control (Figure 4B). Meanwhile, PCA data from antifungal activity aspects covered 93.13% data. According to antifungal activity data, essential oils and extracts were also divided into groups by effect on disease severity (Figure 4C).
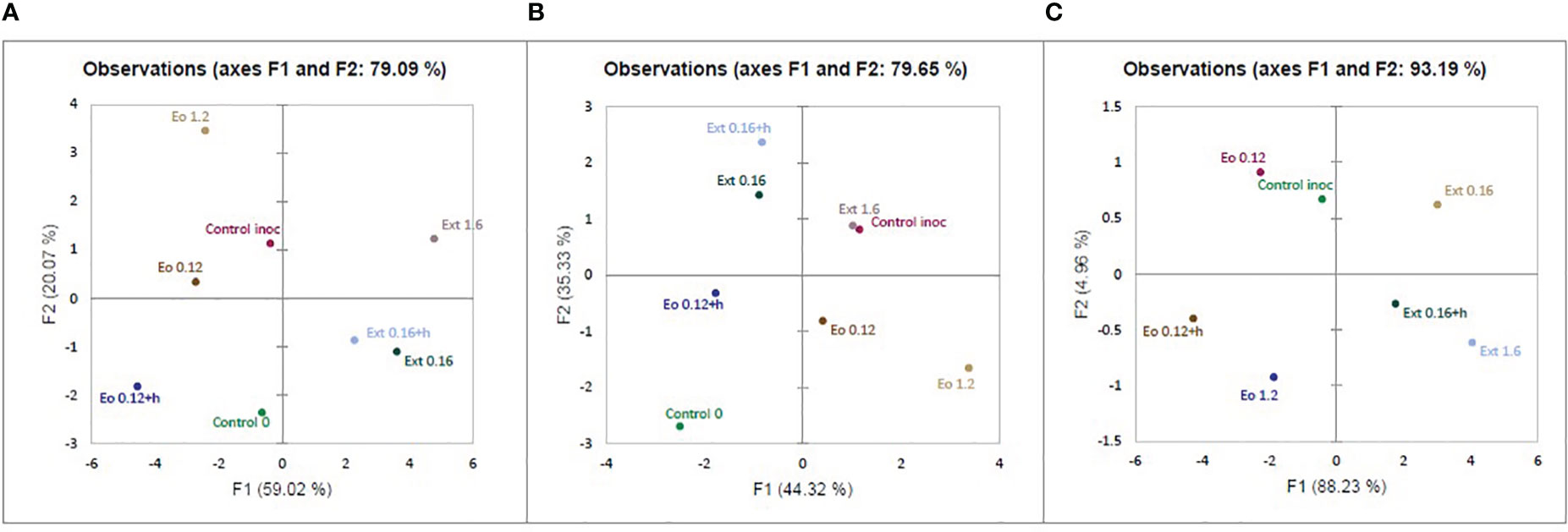
Figure 4 Principal component analysis (PCA) of: combined extracts antifungal activity and treated strawberry plants physiological data (A); only treated strawberry plants physiological data (B), and only extracts antifungal activity data (C).
The results of our study provide relevant information on the further step of natural oils application in vivo for grey mold prevention. In the first part of our study, we determined coriander CO2 extract as the most effective amongst other investigated extracts. Until now, coriander essential oil or extract were mostly investigated for antifungal activity in vitro (Zaker and Mosallanejad, 2010; Mohammadi et al., 2014; Ikeura and Koabayashi, 2015; Zatla et al., 2017; Moghadam et al., 2019). The high antifungal activity of coriander essential oil against Alternaria alternata, Stachybotrys chartarum, Cladosporium cladosporioides, Aspergillus niger fungi was stated in other study (Zabka et al., 2014), which agrees with our in vitro results. It revealed the differences in coriander oil antifungal activity, depending on the active components of the investigated distilled oil fraction (Delaquis et al., 2002). Through the time of our experiments with strawberries, coriander essential oil lowered grey mold severity in vivo. However, the opposite effect was observed after treatment with coriander extract. Such data emphasize the differences between the products obtained using different techniques. However, the lower concentration of coriander essential oil (0.12%) was grouped statistically with inoculated control according to their similar effect on disease severity. Moreover, we did not observe the positive effect of increased product concentration, as higher antifungal activity was registered after adding hydrosol than with a higher concentration. There is no data of application of plant extracts on strawberries under controlled climate conditions so far, thus our results provide valuable information for the further practical use of coriander extracts on horticultural crops.
Antioxidant capacity, both DPPH and ABTS, increased together with increased concentration. Active antioxidant system response after treatment with coriander extract was not efficient in decreasing disease severity and incidence. Evaluating disease incidence, a negative effect of coriander extract was observed, as more plants were found to be infected compared to an inoculated plant. Coriander essential oil activated antioxidant response of strawberry plant lower than extract (except with Eo 1.2 treatment). Reduced stress for the plant after inoculation may have resulted in decreased disease parameters. On the contrary, more observations should be performed with the highest used concentrations (Eo 1.2 and Ext 1.6 treatments) as both extract and essential oil increased strawberries’ total phenolic compounds and antioxidant capacity. Inoculation increases plants’ antioxidant system response. Speaking about the increase level, we could conclude that a slight increase of the antioxidant system was more useful in preventing grey mold infection than a higher increase. A similar effect was found with birch wood distillate, which increased the phenolics in strawberry leaves (Karlund et al., 2014). Phenolic compounds are described as responsible for antifungal activity in tomatoes (Rodrigues and Furlong, 2022). Most of the studies evaluated postharvest application on strawberry fruits against fungal infections (Zatla et al., 2017; Wei et al., 2018; Abd-Elkader et al., 2021; de Oliveira Filho et al., 2021; Hassan et al., 2021). Preharvest application of tea tree oil gradually increased total phenolic content in strawberry fruits in the first days of storage (Wei et al., 2018). We registered gradually increasing antioxidant activity after preharvest treatment with coriander extract, however treatment with essential oil resulted in varied antioxidant capacity. There is a lack of information on how the preharvest application of plant products affects plants during growth. Our observations suggest an interesting hypothesis that secondary products like hydrosol could be useful additives to increase antifungal activity in vivo.
Stomatal conductance, transpiration rate and other photosynthetic capacity indices of plants may be affected by many environmental factors, like heavy metal pollution (Stancheva et al., 2014) or salt stress (Faghih et al., 2017). Coriander extract showed the potential to increase the photosynthetic capacity of infected strawberries. Similar data were obtained by (Yao et al., 2020), where seaweed extracts increased photosynthetic rate, chlorophyll content, and intercellular CO2 in tomatoes. The exception in our study was treatment with Ext 0.16+h, which did not increase photosynthetic rate that much. However, mostly increased stomatal conductance, transpiration rate and the ratio of intercellular CO2 (Ci/Ca) values. Essential oil, which in our case had the highest antifungal activity against grey mold, did not enhance the photosynthetic capacity of strawberries. Our study results provide no direct impact of increased photosynthetic capacity and antifungal effect after treatment with natural oils. It suggests that treatment with coriander extract may not be favorable for the plant, and its reaction was directed not to disease fighting but more to internal processes.
Clear differences after treatment with highest tested concentrations of essential oil and extract were indicated in our study. According to statistical analysis of strawberry’s physiological response to application of coriander products, inoculated plants and higher concentration (1.6%) coriander extract was grouped by similar plant response. Coriander extract in higher concentration caused plant stress similar to infection. Essential oils in high concentrations can have herbicidal activity, and completely kill plants (Tworkoski, 2002). Coriander essential oil increased photosynthetic capacity in low concentrations (0.12%) but with increased concentration (1.2%) photosynthetic rate and other photosynthetic parameters decreased lower compared to control plants. The herbicidal effect on the plant was possibly manifested under 1.2% of coriander essential oil and extract. High total phenolic compound contents and antioxidant scavenging activity can contribute to the statement.
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Conceptualization, LD, KL, AB, NR, and AVa; methodology, LD, KL, NR, and AVa; software, LD and KL; validation, AB and GS; performing the experiment, collecting the samples for analysis, RS, JM, KL, LD, and SC; formal analysis, LD and KL; investigation, AB, VV-K, NR, AVi, AVa, and GS; writing-original draft preparation, LD and KL; writing-review and editing, AB, NR, AVi, AVa, VV-K, and GS; visualization, LD and KL. All authors have read and agreed to the published version of the manuscript.
This project has received funding from European Regional Development Fund (project No 01.2.2-LMT-K-718-03-0035) under grant agreement with the Research Council of Lithuania (LMTLT).
The research team would like to express their gratitude to the laboratory assistants Marytė Smilgaitienė and Galina Starkuvienė for their help during the project implementation.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Abd-Elkader, D. Y., Salem, M. Z. M., Komeil, D. A., Al-Huqail, A. A., Ali, H. M., Salah, A. H., et al. (2021). Post-harvest enhancing and botrytis cinerea control of strawberry fruits using low cost and eco-friendly natural oils. Agronomy 11,1246. doi: 10.3390/agronomy11061246
Ahammed, G. J., Li, X., Mao, Q., Wan, H., Zhou, G., Cheng, Y. (2021). The SlWRKY81 transcription factor inhibits stomatal closure by attenuating nitric oxide accumulation in the guard cells of tomato under drought. Physiol. Plant 172, 885–895. doi: 10.1111/ppl.13243
Andresen, M., Cedergreen, N. (2010). Plant growth is stimulated by tea-seed extract: A new natural growth regulator? HortScience 45, 1848–1853. doi: 10.21273/hortsci.45.12.1848
Bui, T. T. A., Al Wright, S. I., Falk, A. B., Vanwalleghem, T., Van Hemelrijck, W., Hertog, M. L. A. T. M., et al. (2019). Botrytis cinerea differentially induces postharvest antioxidant responses in ‘Braeburn’ and ‘Golden delicious’ apple fruit. J. Sci. Food Agric. 99, 5662–5670. doi: 10.1002/jsfa.9827
Dabkevičius, Z., Brazauskienė, I. (2007). Augalų patologija (Plant pathology). 1st ed. Eds. Švedienė, R., Akademija, D.P. (IDP Solutions: Akademija, Lithuania).
Delaquis, P. J., Stanich, K., Girard, B., Mazza, G. (2002). Antimicrobial activity of individual and mixed fractions of dill, cilantro, coriander and eucalyptus essential oils. Int. J. Food Microbiol. 74, 101–109. doi: 10.1016/S0168-1605(01)00734-6
de Oliveira Filho, J. G., da Cruz Silva, G., de Aguiar, A. C., Cipriano, L., de Azeredo, H. M. C., Bogusz Junior, S., et al. (2021). Chemical composition and antifungal activity of essential oils and their combinations against botrytis cinerea in strawberries. J. Food Meas. Charact. 15, 1815–1825. doi: 10.1007/s11694-020-00765-x
El-Sayed, H. S., Fouad, M. T., El-Sayed, S. M. (2022). Enhanced microbial, functional and sensory properties of herbal soft cheese with coriander seeds extract nanoemulsion. Biocatal. Agric. Biotechnol. 45, 102495. doi: 10.1016/j.bcab.2022.102495
Elstner, N., Singh, R. R. P., Young, A. P. (1993). Finite temperature properties of the spin-1/2 Heisenberg antiferromagnet on the triangular lattice. Phys. Rev. Lett. 71, 1629. doi: 10.1103/PhysRevLett.71.1629
Faghih, S., Ghobadi, C., Zarei, A. (2017). Response of strawberry plant cv. ‘Camarosa’ to salicylic acid and methyl jasmonate application under salt stress condition. J. Plant Growth Regul. 36, 651–659. doi: 10.1007/s00344-017-9666-x
Ferreira, F. V., Musumeci, M. A. (2021). Trichoderma as biological control agent: scope and prospects to improve efficacy. World J. Microbiol. Biotechnol. 37, 90. doi: 10.1007/s11274-021-03058-7
Francesconi, S., Balestra, G. M. (2020). The modulation of stomatal conductance and photosynthetic parameters is involved in fusarium head blight resistance in wheat. PloS One 15, e0235482. doi: 10.1371/journal.pone.0235482
Geng, L., Fu, Y., Peng, X., Yang, Z., Zhang, M., Song, Z., et al. (2022). Biocontrol potential of trichoderma harzianum against botrytis cinerea in tomato plants. Biol. Control. 174, 105019. doi: 10.1016/j.biocontrol.2022.105019
Hassan, E. A., Mostafa, Y. S., Alamri, S., Hashem, M., Nafady, N. A. (2021). Biosafe management of botrytis grey mold of strawberry fruit by novel bioagents. Plants 10, 2737. doi: 10.3390/plants10122737
Ikeura, H., Koabayashi, F. (2015). Antimicrobial and antifungal activity of volatile extracts of 10 herb species against glomerella cingulata. J. Agric. Sci. 7, 77–84. doi: 10.5539/jas.v7n9p77
Karlund, A., Salminen, J. P., Koskinen, P., Ahern, J. R., Karonen, M., Tiilikkala, K., et al. (2014). Polyphenols in strawberry (Fragaria × ananassa) leaves induced by plant activators. J. Agric. Food Chem. 62, 4592–4600. doi: 10.1021/jf405589f
Lasram, S., Zemni, H., Hamdi, Z., Chenenaoui, S., Houissa, H., Saidani Tounsi, M., et al. (2019). Antifungal and antiaflatoxinogenic activities of carum carvi l., coriandrum sativum l. seed essential oils and their major terpene component against aspergillus flavus. Ind. Crops Prod. 134, 11–18. doi: 10.1016/j.indcrop.2019.03.037
Mazzoncini, M., Antichi, D., Silvestri, N., Ciantelli, G., Sgherri, C. (2015). Organically vs conventionally grown winter wheat: Effects on grain yield, technological quality, and on phenolic composition and antioxidant properties of bran and refined flour. Food Chem. 175, 445–451. doi: 10.1016/j.foodchem.2014.11.138
Moghadam, Z. A., Hosseini, H., Hadian, Z., Asgari, B., Mirmoghtadaie, L., Mohammadi, A., et al. (2019). Evaluation of the antifungal activity of cinnamon, clove, thymes, zataria multiflora, cumin and caraway essential oils against ochratoxigenic aspergillus ochraceus. J. Pharm. Res. Int. 26, 1–16. doi: 10.9734/jpri/2019/v26i130126
Mohammadi, S., Aroiee, H., Aminifard, M. H., Tehranifar, A., Jahanbakhsh, V. (2014). Effect of fungicidal essential oils against botrytis cinerea and rhizopus stolonifer rot fungus in vitro conditions. Arch. Phytopathol. Plant Prot. 47, 1603–1610. doi: 10.1080/03235408.2013.853456
Morais, W. C. C., Lima, M. A. P., Zanuncio, J. C., Oliveira, M. A., Bragança, M. A. L., Serrão, J. E., et al. (2015). Extracts of ageratum conyzoides, coriandrum sativum and mentha piperita inhibit the growth of the symbiotic fungus of leaf-cutting ants. Ind. Crops Prod. 65, 463–466. doi: 10.1016/j.indcrop.2014.10.054
Nadaf, N. H., Gawade, S. S., Muniv, A. S., Waghmare, S. R., Jadhav, D. B., Sonawane, K. D. (2015). Exploring anti-yeast activity of nigella sativa seed extracts. Ind. Crops Prod. 77, 624–630. doi: 10.1016/j.indcrop.2015.09.038
Nxumalo, K. A., Aremu, A. O., Fawole, O. A. (2021). Potentials of medicinal plant extracts as an alternative to synthetic chemicals in postharvest protection and preservation of horticultural crops: A review. Sustain 13,5897. doi: 10.3390/su13115897
Perez-Lopez, U., Pinzino, C., Quartacci, M. F., Ranieri, A., Sgherri, C. (2014). Phenolic omposition and related antioxidant properties in differently colored lettuces: A study by electron paramagnetic resonance (EPR) kinetics. J. Agric. Food Chem. 62, 12001–12007. doi: 10.1021/jf503260v
Petrasch, S., Knapp, S. J., van Kan, J. A. L., Blanco-Ulate, B. (2019). Grey mould of strawberry, a devastating disease caused by the ubiquitous necrotrophic fungal pathogen botrytis cinerea. Mol. Plant Pathol. 20, 877–892. doi: 10.1111/mpp.12794
Ragaee, S., Abdel-Aal, E. S. M., Noaman, M. (2006). Antioxidant activity and nutrient composition of selected cereals for food use. Food Chem. 98, 32–38. doi: 10.1016/j.foodchem.2005.04.039
Raveau, R., Fontaine, J., Lounès-Hadj Sahraoui, A. (2020). Essential oils as potential alternative biocontrol products against plant pathogens and weeds: A review. Foods 9, 365. doi: 10.3390/foods9030365
Re, R., Pellegrini, N., Proteggente, A., Pannala, A., Yang, M., Rice-Evans, C. (1999). Development and characterisation of carbon nanotube-reinforced polyurethane foams. Free Radic. Biol. Med. 26, 1231–1237. doi: 10.1016/S0891-5849(98)00315-3
Rivero, R. M., Ruiz, J. M., Garcia, P. C., Lopez-Lefebre, L. R., Sanchez, E., Romero, L. (2001). Resistance to cold and heat stress: accumulation of phenolic compounds in tomato and watermelon plants. Plant Sci. 160, 315–321. doi: 10.1016/S0168-9452(00)00395-2
Rodrigues, M. H. P., Furlong, E. B. (2022). Fungal diseases and natural defense mechanisms of tomatoes (Solanum lycopersicum): A review. Physiol. Mol. Plant Pathol. 122, 101906. doi: 10.1016/j.pmpp.2022.101906
Sekmen Cetinel, A. H., Gokce, A., Erdik, E., Cetinel, B., Cetinkaya, N. (2021). The effect of trichoderma citrinoviride treatment under salinity combined to rhizoctonia solani infection in strawberry (Fragaria x ananassa duch.). Agronomy 11,1589. doi: 10.3390/agronomy11081589
Šernaitė, L., Rasiukevičiūtė, N., Dambrauskienė, E., Viškelis, P., Valiuškaitė, A. (2020). Biocontrol of strawberry pathogen botrytis cinerea using plant extracts and essential oils. Zemdirbyste-Agriculture 107, 147–152. doi: 10.13080/z-a.2020.107.019
Soleha, S., Muslim, A., Suwandi, S., Kadir, S., Pratama, R. (2022). The identification and pathogenicity of fusarium oxysporum causing acacia seedling wilt disease. J. For. Res. 33, 711–719. doi: 10.1007/s11676-021-01355-3
Stancheva, I., Geneva, M., Markovska, Y., Tzvetkova, N., Mitova, I., Todorova, M., et al. (2014). A comparative study on plant morphology, gas exchange parameters, and antioxidant response of ocimum basilicum l. and origanum vulgare l. grown on industrially polluted soil. Turkish. J. Biol. 38, 89–102. doi: 10.3906/biy-1304-94
Tworkoski, T. (2002). Herbicide effects of essential oils. Weed. Sci. 50, 425–431. doi: 10.1614/0043-1745(2002)050[0425:HEOEO]2.0.CO;2
Tzortzakis, N. (2019). Physiological and proteomic approaches to address the active role of botrytis cinerea inoculation in tomato postharvest ripening. Microorganisms 7, 681. doi: 10.3390/microorganisms7120681
Wangensteen, H., Samuelsen, A. B., Malterud, K. E. (2004). Antioxidant activity in extracts from coriander. Food Chem. 88, 293–297. doi: 10.1016/j.foodchem.2004.01.047
Wei, Y., Shao, X., Wei, Y., Xu, F., Wang, H. (2018). Effect of preharvest application of tea tree oil on strawberry fruit quality parameters and possible disease resistance mechanisms. Sci. Hortic. (Amsterdam). 241, 18–28. doi: 10.1016/j.scienta.2018.06.077
Yao, Y., Wang, X., Chen, B., Zhang, M., Ma, J. (2020). Seaweed extract improved yields, leaf photosynthesis, ripening time, and net returns of tomato (Solanum lycopersicum mill.). ACS Omega. 5, 4242–4249. doi: 10.1021/acsomega.9b04155
Zabka, M., Pavela, R., Prokinova, E. (2014). Antifungal activity and chemical composition of twenty essential oils against significant indoor and outdoor toxigenic and aeroallergenic fungi. Chemosphere 112, 443–448. doi: 10.1016/j.chemosphere.2014.05.014
Zaker, M., Mosallanejad, H. (2010). Antifugal activity of some plant extracts on alternaria alternata, the causal agent of alternaria leaf spot of potato. Pakistan J. Biol. Sci. 13, 1023–1029. doi: 10.3923/pjbs.2010.1023.1029
Zamindar, N., Sadrarhami, M., Doudi, M. (2016). Antifungal activity of coriander (Coriandrum sativum l.) essential oil in tomato sauce. J. Food Meas. Charact. 10, 589–594. doi: 10.1007/s11694-016-9341-0
Zatla, A. T., El Amine Dib, M., Djabou, N., Ilias, F., Costa, J., Muselli, A. (2017). Antifungal activities of essential oils and hydrosol extracts of daucus carota subsp. sativus for the control of fungal pathogens, in particular gray rot of strawberry during storage. J. Essent. Oil Res. 29, 391–399. doi: 10.1080/10412905.2017.1322008
Keywords: biocontrol, coriandrum sativum, grey mold, disease severity, antioxidant activity, photosynthesis, sustainable plant protection
Citation: Dėnė L, Laužikė K, Rasiukevičiūtė N, Chrapačienė S, Brazaitytė A, Viršilė A, Vaštakaitė-Kairienė V, Miliauskienė J, Sutulienė R, Samuolienė G and Valiuškaitė A (2023) Defense response of strawberry plants against Botrytis cinerea influenced by coriander extract and essential oil. Front. Plant Sci. 13:1098048. doi: 10.3389/fpls.2022.1098048
Received: 14 November 2022; Accepted: 14 December 2022;
Published: 05 January 2023.
Edited by:
Daolong Dou, Nanjing Agricultural University, ChinaReviewed by:
Alessandra Carrubba, Università di Palermo, ItalyCopyright © 2023 Dėnė, Laužikė, Rasiukevičiūtė, Chrapačienė, Brazaitytė, Viršilė, Vaštakaitė-Kairienė, Miliauskienė, Sutulienė, Samuolienė and Valiuškaitė. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Lina Dėnė, bGluYS5kZW5lQGxhbW1jLmx0
†These authors have contributed equally to this work and share first authorship
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.