
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
MINI REVIEW article
Front. Plant Sci. , 18 January 2023
Sec. Plant Symbiotic Interactions
Volume 13 - 2022 | https://doi.org/10.3389/fpls.2022.1092105
This article is part of the Research Topic The Role of Endophytes in Plant Health and Resistance Against Biotic and Abiotic Stresses View all 13 articles
 Sehrish Mushtaq1
Sehrish Mushtaq1 Muhammad Shafiq2*
Muhammad Shafiq2* Muhammad Rizwan Tariq3
Muhammad Rizwan Tariq3 Adnan Sami4
Adnan Sami4 Muhammad Shah Nawaz-ul-Rehman5
Muhammad Shah Nawaz-ul-Rehman5 Muhammad Hamza Tariq Bhatti1Muhammad Saleem Haider1
Muhammad Hamza Tariq Bhatti1Muhammad Saleem Haider1 Saleha Sadiq6Muhammad Taqqi Abbas7
Saleha Sadiq6Muhammad Taqqi Abbas7 Mujahid Hussain8
Mujahid Hussain8 Muhammad Adnan Shahid8*
Muhammad Adnan Shahid8*Endophytic bacteria are mainly present in the plant’s root systems. Endophytic bacteria improve plant health and are sometimes necessary to fight against adverse conditions. There is an increasing trend for the use of bacterial endophytes as bio-fertilizers. However, new challenges are also arising regarding the management of these newly discovered bacterial endophytes. Plant growth-promoting bacterial endophytes exist in a wide host range as part of their microbiome, and are proven to exhibit positive effects on plant growth. Endophytic bacterial communities within plant hosts are dynamic and affected by abiotic/biotic factors such as soil conditions, geographical distribution, climate, plant species, and plant-microbe interaction at a large scale. Therefore, there is a need to evaluate the mechanism of bacterial endophytes’ interaction with plants under field conditions before their application. Bacterial endophytes have both beneficial and harmful impacts on plants but the exact mechanism of interaction is poorly understood. A basic approach to exploit the potential genetic elements involved in an endophytic lifestyle is to compare the genomes of rhizospheric plant growth-promoting bacteria with endophytic bacteria. In this mini-review, we will be focused to characterize the genetic diversity and dynamics of endophyte interaction in different host plants.
Plants interact with diverse microbial populations in the ecosystem (Delaux et al., 2015). Microorganisms can colonize on plants’ surfaces or internal parts depending on the host genotype and the molecular signals released by plant roots. Microorganisms can colonize on plants’ surfaces or internal parts depending on the host genotype and the molecular signals released by plant roots. Endophytes are prokaryotic bacteria found within the healthy host tissue (Brader et al., 2014). Bacterial endophytes can benefit the host in several ways, such as biotic and abiotic stress resistance, increased availability of nutrients, degradation of toxic molecules, and production of phytohormones (Kandel et al., 2015).
Plant population dynamics have soil microbial intermediation. The plant has a microbial population in the phyllosphere, endophytes, or rhizospheric microbes. The ecology and phenotype of the plants can be affected by the influence of symbiotic microbes on the atmosphere and competition for soil resources.
The plant genotype affects the microbial make-up of the phyllosphere, rhizosphere, and endophytic microorganisms (Lynch et al., 2001). Although the precise method involves the plant-associated microorganisms and ecosystem function, the other specific mechanism is still unknown. Because they are co-evolved with bacteria, plants are immobile and need to control the results of their intricate interactions (Schnitzer and Klironomos, 2011). Different sorts of chemicals are continuously produced by plant roots, gathered, and secreted into the soil (Wood et al., 2012) known as the root exudates which contain enzymes, water, mucilage, H+ ions, and primary, secondary compounds made up of carbon (Singh, 2015). Every plant species’ rhizosphere is known to have a microorganism population that is 100 times higher than soil and is mostly controlled by compounds generated by roots (Jonkers et al., 2003; Bever, 2003). The favorable plant-soil microbial response enhances the microbial populations’ spatial spread (Schimel et al., 2007), while negative reaction results in plant replacement, which demands recolonization of locally specific roots (Bever et al., 2010; Pedrotti et al., 2013).
It has been proposed that endophytic bacteria vary from rhizobacteria in their genetic architecture, which may account for their capacity to colonise plant tissues internally. However, no specific gene or gene family has been found to explain the endophytic regime. In a 2014 study, the whole genomes of nine Proteobacteria were compared to identify a list of genes that may play a role in the endophytic activity. So yet, only a few of those genes have undergone experimental testing to determine whether they are involved in endophytic colonisation (Shen et al., 2013; Ouyabe et al., 2019). In this study, we have documented some mechanisms involved in plant endophyte interaction at the molecular level.
PGPEs enhance plant development through three interconnected mechanisms: phytostimulation, biofertilization, and biocontrol. Phytostimulation is the production of phytohormones for direct plant development (Vishwakarma et al., 2021). The amount of the plant hormone ethylene frequently declines as a result of the enzyme 1-aminocyclopropane-1-carboxylate (ACC) deaminase (Cruz Barrera et al., 2020). According to numerous studies, the pea plant and the pepper plant (Pseudomonas putida and Piper nigrum, respectively) both have bacterial endophytes that release ACC deaminase to aid plant growth (Ruduś et al, 2013). By controlling ethylene levels in plants, ACC deaminize production may minimize abiotic stress because an increase in ethylene can obstruct DNA synthesis, root and shoot growth, and cell division. However, the specific method for enhanced plant development is still unknown (González Candia, 2021). Bacterial strains also produced other hormones which include abscisic acid, indole-3-acetic acid, and jasmonic acid, to stimulate plant growth (Forchetti et al., 2007).The endophytes can enhance plant growth by increasing the availability of important nutrients known as bio-fertilization.
Nitrogen fixation is the most studied phenomenon of bio-fertilization which is the conversion of atmospheric nitrogen into ammonia (Mishra and Arora, 2016). Bacterial species like Azospirillum spp., Pantoea agglomerans, and Azoarcus spp. all are known to be involved in a substantial amount of nitrogen fixation in plant roots (Indiragandhi et al., 2008). Nonetheless, only 21 PGPEs can increase plant phosphorus availability by solubilizing phosphate. The metal cation linked to phosphorous is chelated as a result of the release of low molecular weight acids, making it more available to plants. The researchers have isolated, identified, and assessed the ability of Achromobacter xiloxidans and Bacillus pumilus to solubilize phosphate in sunflowers (Barrera et al., 2020). PGPEs were utilized to treat corn, lowering the quantity of artificial phosphorus fertilizer required while increasing yields by up to 50% (Cruz Barrera et al., 2019).
The protection of plants from phytopathogens and their growth promotion is known as biological control. Antibiotic and siderophores production are involved in biological control mechanisms. Siderophores like pyochelin and alicyclic acid and chelate iron are not directly involved in disease control due to their competition with pathogens for trace metals (Leopold, 1964). The disease can be suppressed in plants by antimicrobial metabolites secreted by bacterial endophytes such as 2,4-diacetylphloroglucinol (DAPG). Seed treatment of eggplant (Solanum melongena) with DAPG-producing bacterial endophytes reduced 70% of eggplant wilt caused by Ralstonia solanacearum (Rana et al., 2020a).
Burkholderia, Bacillus, Pseudomonas, Enterobacter, and Serratia are just a few of the bacterial endophyte strains that are successful at preventing the growth of pathogenic germs in both in vitro and in vivo settings (Khan and Doty, 2009). Aside from that, bacteria from the genera Bacillus, Enterobacter, Arthrobacter, Azotobacter, Isolptericola, Streptomyces, and Pseudomonas improved the crop’s stress resistance from heat, drought, and salt (Rana et al., 2020b; Khalil et al., 2021). The most important interaction between these endophytes and symbiotic plants allowed the plants to significantly increase their biomass and height while lowering stress. Although, it is not yet clear how bacterial endophytes lessen abiotic stress (Liu et al., 2014).
Rhizobium is a member of the family Rhizobiaceae and the class Alphaproteobacteria. Rhizobium, was the name given to this genus for the first time by Frank in 1889. There are 11 non-rhizobial species and 49 rhizobial species in the family Rhizobiaceae at the moment (Ledermann et al., 2021). The rhizobial species induce the nodules on the roots of plants (Fabaceae family) and are linked to symbiotic nitrogen-fixing bacteria. The nodule’s nitrogen fixation activity is extremely oxygen sensitive. The host plant receives continual supplies of reduced nitrogen from the bacterial enzyme system in this symbiotic connection, and the bacteria in exchange receive nutrients and energy from the plant (Van Rhijn and Vanderleyden, 1995). Nodules can occur in about 10% of legumes. The majority of the rhizobacteria in soil are oxygen sensitive and feed on the decomposing remains of other organisms.
In roots, nitrogen-fixing bacteria occur as irregular cells known as bacteroids, which are frequently Y, club-shaped and appear as straight rods with a regular structure (Figure 1). Bacteroidsencode genes that determine the rhizobium’s host specificity (Lodwig and Poole, 2003). Rhizobia that generate nodules but are unable to fix nitrogen are sometimes referred to as ineffective strains, whereas effective strains cause nitrogen fixation in nodules. Nodule development is controlled by certain genes known as nod genes i.e. nodF, nodE, nodL, nodP, nodQ, and nodH (Basile and Lepek, 2021). Some substances, such as flavonoids, are released by the root cells and trigger the production of nodules in bacteria by activating the nod gene. In essence, these chemicals are in charge of identifying the proper host and attaching to the root hairs.
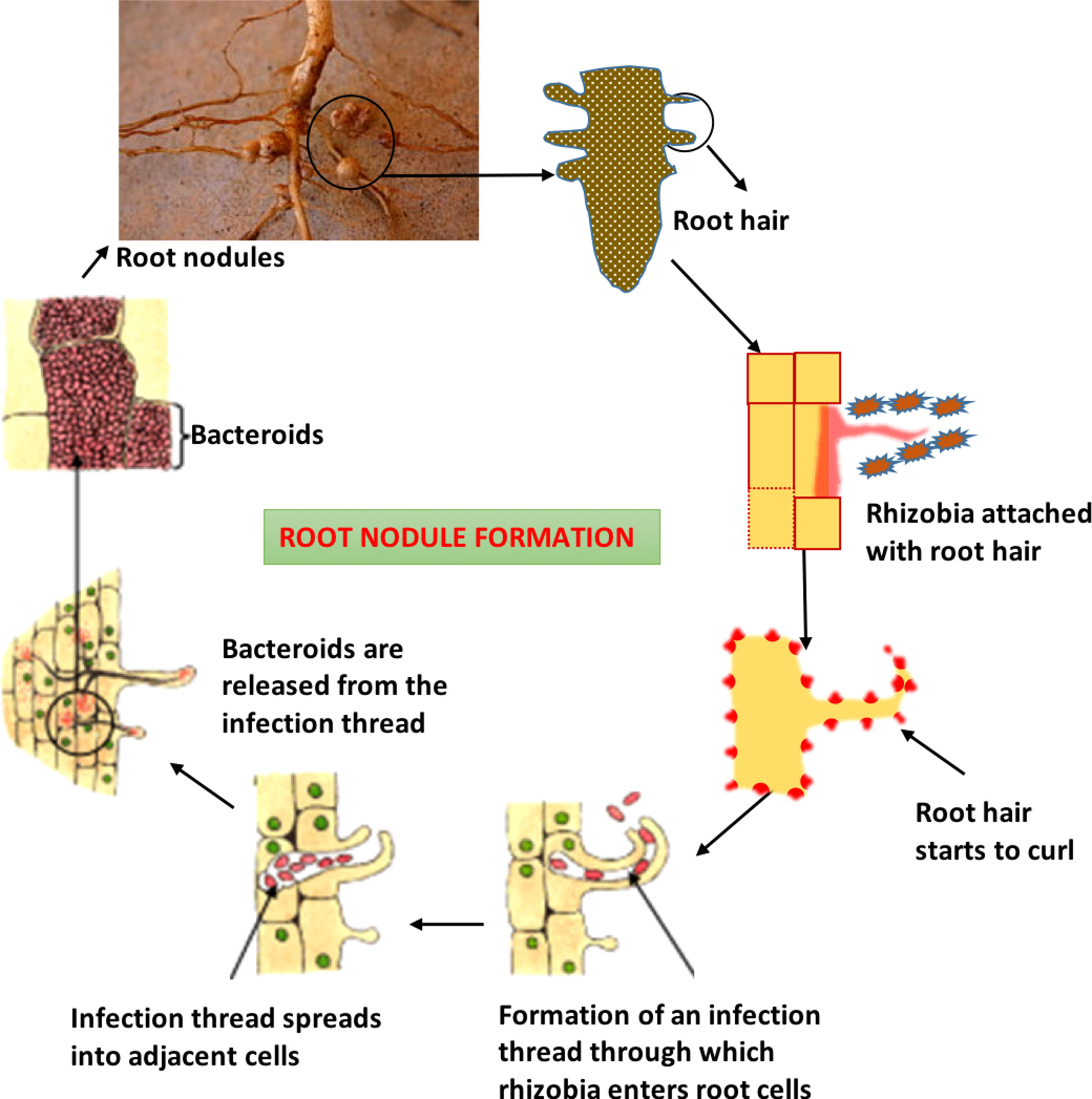
Figure 1 Diagrammatic representation of the whole process of nodule formation through rhizobia.
The nod factors, which are secreted by bacteria, cause the root hairs to curl (Moran, 1997). The root hair tip is damaged by rhizobia, which also causes the infection thread to arise. The thread then extends to neighboring cells by thread branching, and the bacteria continue to grow within the growing network of tubes, continuing to create nod factors that encourage the growth of the root cells and ultimately result in the formation of root nodules (Oldroyd et al., 2011). Following a week of infection, nodules are visible with the unaided eye and each nodule contains thousands of living rhizobium bacteria, the majority of which are malformed and are referred to as bacteroids. Small sections of the plant cell membranes called symbiosomes, which may or may not include multiple bacteroids, are located next to bacteroids and are active sites for nitrogen fixation (Ratu et al., 2021). Through the Nitrogenase enzyme, also known as Nitrogenize catalysis, nitrogen gas from the atmosphere is converted inside legume nodules into ammonia, which is then assimilated into amino acids, DNA, and RNA as well as significant energy molecules like ATP or other chemicals like vitamins, flavones, and hormones (Bergersen, 1961). The Nitrogenize complex is protected by a variety of mechanisms used by aerobic free-living bacteria, including physical barriers and fast metabolic rates. Azotobacter, for instance, circumvents this issue by maintaining the lowest oxygen concentration in its cells and the greatest rate of respiration of any organism. In the instance of Rhizobium, the nodule’s red iron-containing protein, similar to hemoglobin in function to bond with oxygen, maintains control over the oxygen level (Lindström and Mousavi, 2020). However, this avoids the accumulation of free oxygen to prevent the loss of Nitrogenize activity while still providing enough oxygen for the metabolic functioning of bacteriods. Rhizobia and plants work together to make leghemoglobin, something neither of them could ever do on their own. Even in poor soil with few nutrients and insufficient nitrogen to support the growth of other plants, these nodules increase crop output (Lodwig and Poole, 2003).
Plants and their microbial diversity vary throughout their life span of plants. These factors, prompt the structure and variety of the microbial community (Honma and Shimomura, 1978). Seed-born microbes gain entry into the germinating plant and take advantage of other colonizing microbes as well as opportunistic pathogens from the surrounding soil (Glick et al., 1999; Oteino et al., 2015). Hence the overall microbial biota and population changed dramatically throughout the life cycle of plants. The important ways of entry into host plants are through root hair cells, root cracks, and wounds whereas other sources include stomata particularly of young stems and leaves; lenticels, and germinating radicles (Figure 2). Vertical seed transmission is another possible way to receive endophytic bacteria through plant host generations (Bergersen, 1961).
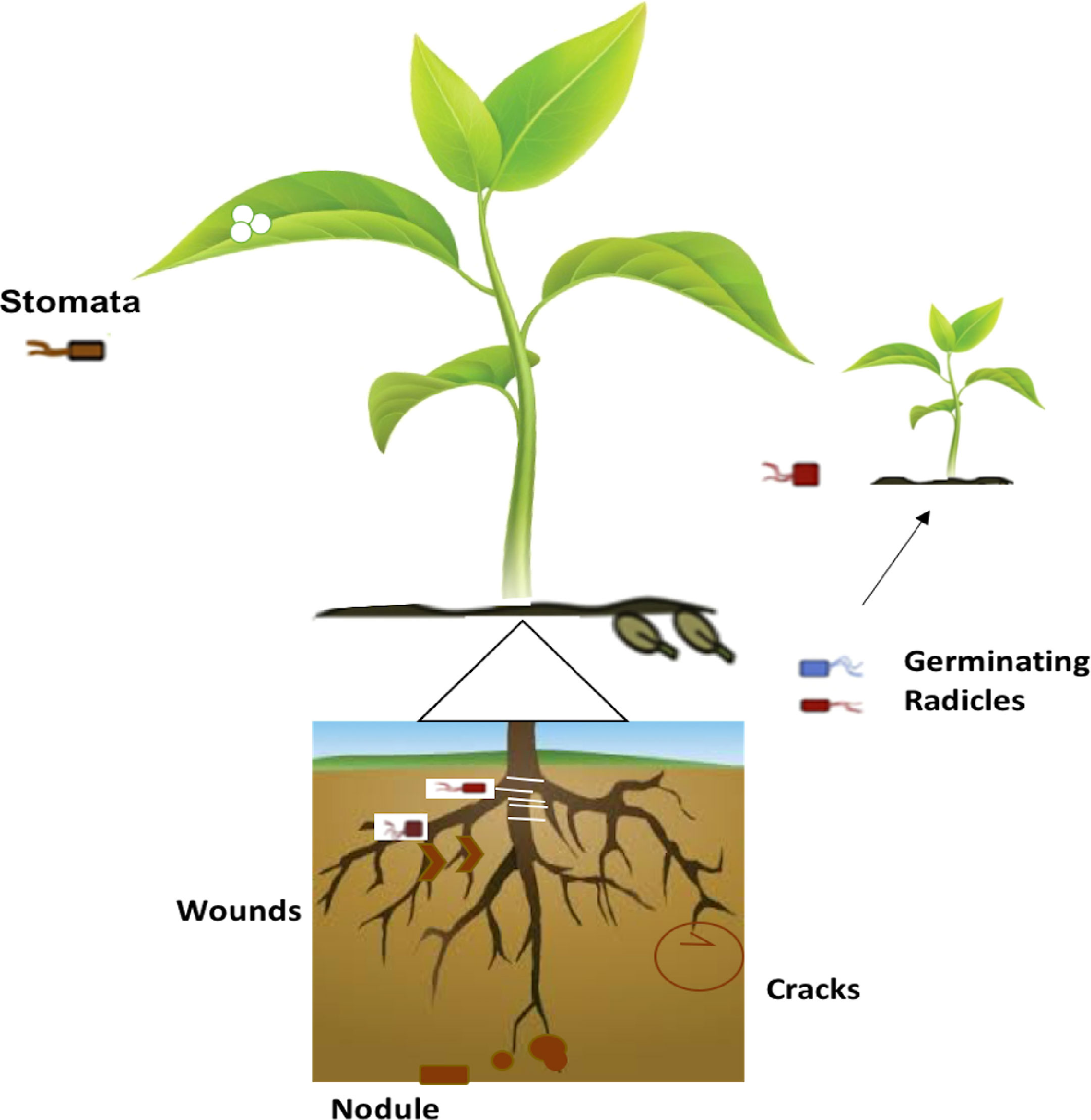
Figure 2 Overview of the endophytic bacterial mode of entry into different plant tissues.
Microorganisms associated with plants formed a complex network. Different studies suggested that plant-associated microbes live inside plant tissues or on the surface of plant parts such as leaves, stems, fruit, and roots (Clarholm, 1985). The microbiome studies of A. thaliana leaves showed that plant genotype, surrounding plants, and abiotic features affected the microbial population structure (Teixeira et al., 2013). These interactions are responsible for expediting the defense signals between plants and the efficacy of natural biological control agents (Morgan et al., 2005). Microbial populations might indirectly affect the other taxa of microbes by altering the host growth response or metabolites without direct interaction with microbes.
Plants usually take nutrients from the soil which constitutes a pool for microscopic life forms including bacteria, fungi, actinomycetes, algae, and protozoa. So, among them, the bacteria are the most common ones and have the maximum proportion in soil. The maximum number of bacteria present in the rhizosphere near the roots of plants is different from bulk soil (Luu et al., 2020). As these bacteria are present in more concentration in the soil so the bacteria may affect a plant through three different pathways (Edwards and Harding, 2004). PGPEs can promote plant growth directly by expediting the procurement of compounds or modifying levels of plant hormones and reducing the inhibitory effect of plant growth and pathogenicity by acting as biocontrol agents (Yan et al., 2019). The benefits provided by the endophytes to the host plants and their mechanisms are described in (Table 1).
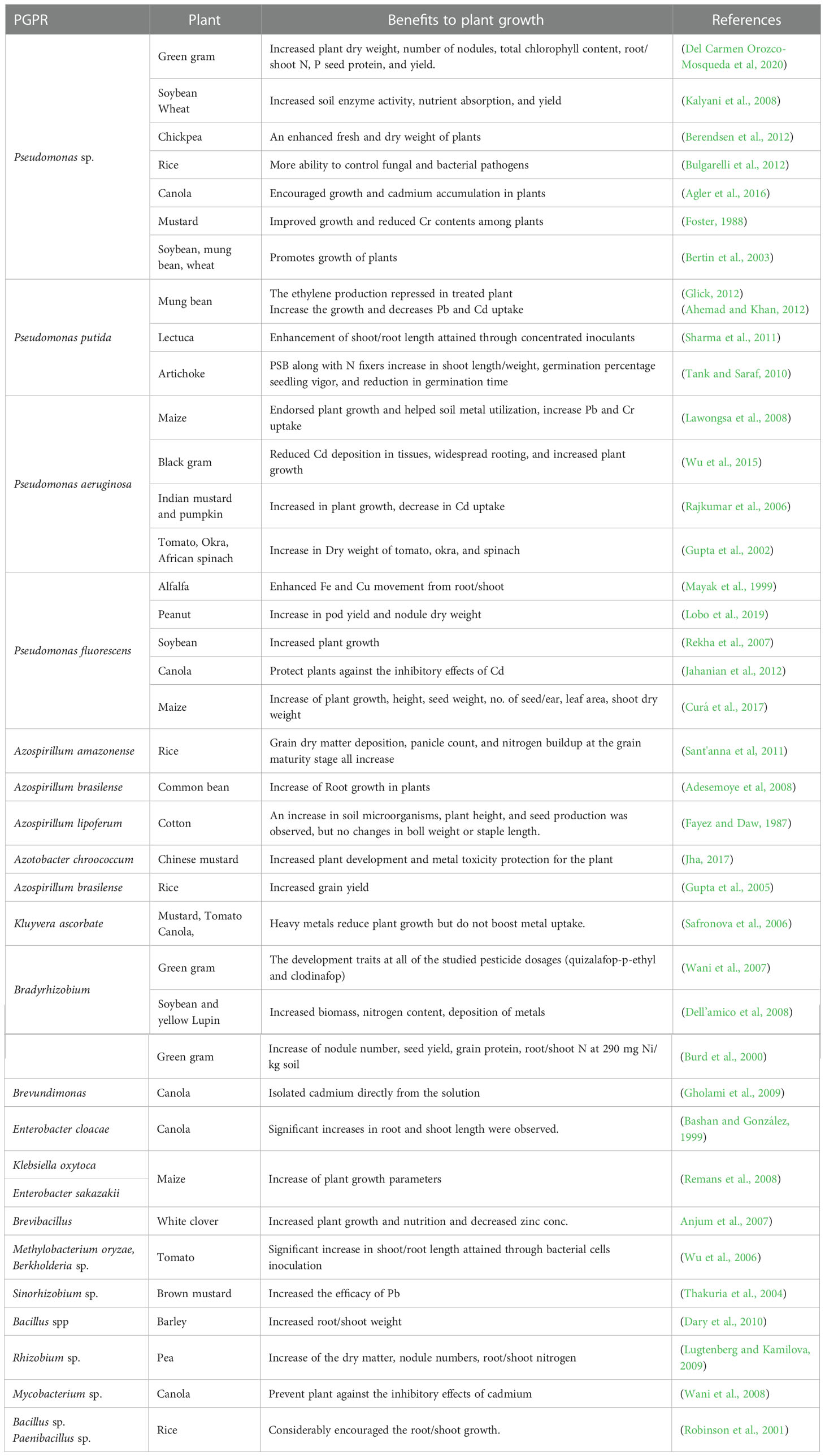
Table 1 Examples of plant growth-promoting rhizobacteria tested for various crop types.
Throughout their lives, plants are exposed to harmful abiotic and biotic stresses. The damage that bacteria, fungi, viruses, nematodes, viroids, and insects do to plants is referred to as “biotic stress.” Rhizobacteria that promote plant growth by generating phytohormones or facilitating the uptake of particular nutrients might affect plant growth through biotic stress (Tiwari et al., 2020). However, PGPR reduces or even eliminate the negative impacts of plant pathogens. For example, Pseudomonas fluorescens produces 2,4-Diacetyl Phloroglucinol, which inhibits the development of pathogenic fungi in plants (Suslow and Schroth, 1982). Chitinase and laminarinase, two extracellular enzymes generated by P. stutzeri, caused the lysis of Fusarium solani mycelia and root rot (Cano-Salazar et al., 2011). During a seven-month field trial, the endophytic B. cenocepacia reduced the prevalence of fusarium wilt disease in banana plants by 3.4%, compared to 24.5% in untreated infected plants (Sapak et al., 2008). The antibiotic Pyrrolnitrin, which helps to reduce cotton damping off losses brought on by Rhizoctonia solani, was developed by several endophytic Pseudomonas fluorescens strains (Timper et al., 2009). Fusarium oxysporum, which was used as a bio-agent to create resistance in tomato plants, was successfully protected against P. fluorescens in flowering plants (Dudai, 2011). A bacteria that inhabit plant roots called Bacillus amyloliquefaciens has the power to control plant diseases and promote plant growth (Vardi et al., 2021).
In a study, it was discovered that bacterial endophytes shield cucumber plants from the cucumber anthracnose produced by Pseudomonas fluorescents (Akköprü et al., 2021). It was once believed that Achromobacter sp., Streptomyces sp., and Bacillus licheniformis were responsible for the foliar disease known as downy mildew. The downy mildew disease infestation level was lowered by Pseudoperonospora cubensis (Basu et al., 2022), which ultimately resulted in an increased yield.
The management of pests, which has become a challenge for most crops since pests have evolved a tolerance to pesticides, is another use for these endophytic bacteria (Deng et al., 2014). Entomopathogenic bacteria have been used to combat pests that are immune to insecticides (Figure 3). A few fungi from the genera Podonectria, Verticillium, Hirsutella, Sphaerostilbe, Agerata, Metarhizium Aschersonia, and Myriangium are used for the biological management of pests. Brevibacillus laterosporus is effective against nematodes, Lepidoptera, Coleoptera, and toxic fungi in plants in addition to insects (Skinner et al., 2014).
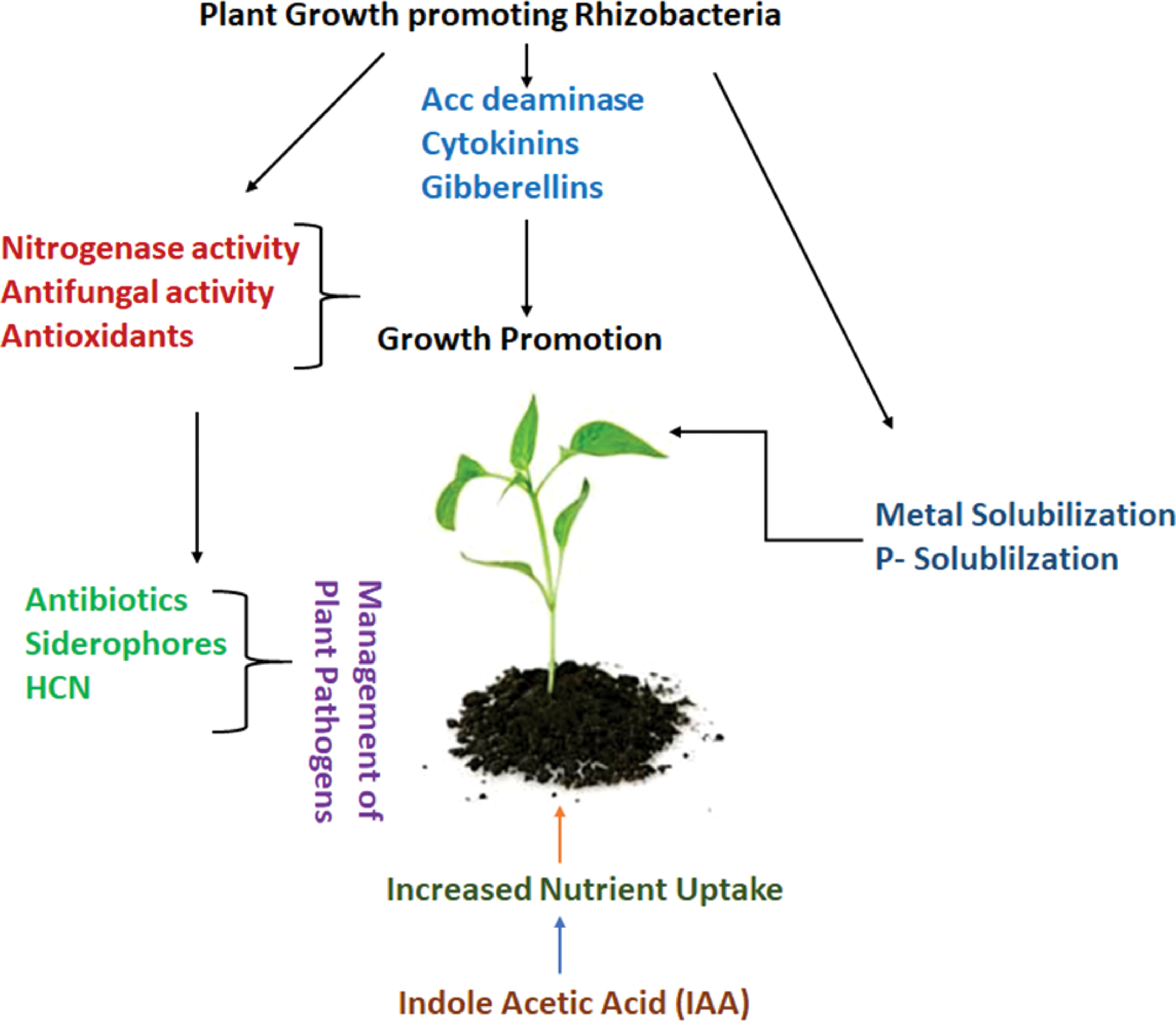
Figure 3 Mechanism of plant growth promotion by rhizobacteria (PGPR).
In recent years, next-generation sequencing (NGS) techniques have been utilized to study the whole population of cultivable or non-cultivable bacteria inside plants, as well as their genomes. The interaction of host and bacterial endophytes has insightful concerns for the biological functioning of plants. As a result of interactions, rapid changes in host phenotype occurs also it is assumed as a driving force for the speciation and co-evolution of both the genetic system of host and bacteria (Fawcett, 1944). Though old genetic techniques to study plant-microbe interaction are less efficient, time-consuming, costly, and labor-intensive required a wide range of experiments and are usually limited to certain known genes (De Oliveira et al., 2004) in comparison to investigating the host-microbe interactions in molecular levels, it is needed to understand the phenotypic phenomena and genomics in depth. So the development of NGS technologies or metagenomic studies has provided the best way to understand the host-pathogen system. Through this technology, we can construct genome models of different organisms, which includes strains, their natural populations over time and their evolutionary histories (Navas et al., 2017; Sharma et al., 2021).
These complicated interactions can be analyzed and integrated by viewing plant microbiomes as a system. To better understand endophytism, contemporary genomic investigations incorporating metaomics and comparative studies can be quite beneficial (Dubey et al., 2020). A better understanding of endophyte interactions could be used to improve agricultural management by increasing plant development, biocontrol, and bioremediation (Alaimo et al., 2018). Some of the tools being utilized or that could be used to understand the link between plants and endophytes include genome sequencing, comparative genomics, microarray, next-generation sequencing, metagenomics, and metatranscriptomics (Dixit et al., 2022). To study endophytes and their apparent function in host plant ecology, contemporary methods and approaches need to be investigated (Gaiero et al., 2013).
Another way to identify the endophytic bacteria interact with the plant is to isolate the endophytic bacteria culture and then classify based on its phenotypic traits, and a few isolates from each category are identified further through partial sequencing of the 16S rRNA gene (Khare et al., 2018). The results of partial sequencing show that the isolates belonged to the genera Pseudomonas, Stenotrophomonas, Bacillus, Pantoea, and Serratia of bacteria (Liu et al, 2017>2552). These isolates are tested for their ability to produce siderophores, phosphate solubilization, atmospheric nitrogen fixation, protease, and hydrogen cyanide, as well as phytohormones like auxin and gibberellin (Eid et al., 2019). Auxin and gibberellin, two plant growth hormones, can be produced by all strains, though to varying degrees. Almost all strains could solubilize phosphate (Lata et al., 2019). The outcomes of protease, siderophore, and atmospheric nitrogen-fixing ability vary between strains. These findings provide information on the relationship between endophytic bacteria and their host plant (Vandana et al., 2021).
Furthermost genomic methods require recognition of variations among sequences within species or populations, like point mutations, Addition/deletions, and structural variations in structures (Bulgarelli et al., 2013).
One of the great evolutionary changes in life is the development of advantageous symbioses between eukaryotic (plants) and prokaryotic creatures (Chebotar et al., 2015). According to certain theories, the relationship between endophytic bacteria and plants frequently depends on two fundamental elements: currency and a system for exchanging currency. The currency could be, for instance, a root exudate that bacteria can take up in the context of interactions between plants and endophytic bacteria (Mercado-Blanco and JJ Lugtenberg, 2014). Similarly, bacteria may release hormones that encourage plant growth, such as auxin and gibberellins, which may be favorable for plant growth (Maksimov et al., 2018). It is anticipated that selection will favor the evolution of mutualism when the exchange of currencies between the two parties is balanced. Therefore, it is hypothesized that increased mutualistic dependency develops through reciprocal co-evolution or adaptation by one of the partners through the selection of features directly related to the mutualistic interaction (Chen et al., 2021).
Competition for scarce shared resources like iron may also lead to asymmetrical currency exchange, which could help to explain why some plant-microbe interactions are hostile (Hong and Park, 2016). Furthermore, because the rhizosphere is open, the free diffusion of resources derived from plants may promote higher levels of cheating in which mutant bacterial genotypes take benefit of “public goods” without producing substances that aid plant growth (Pandey et al., 2017). Because of this, mutualistic plant-microbe interactions may need additional enforcement from the plant, such as penalizing dishonest bacterial genotypes or positively identifying genotypes that promote plant growth (Ryan et al., 2008).Intriguing research would also be done to see whether endophytic bacteria and plants may coevolve from first neutral interaction and whether plants can coevolve in response to rhizosphere bacteria (Santos et al., 2018). In conclusion, by showing that plant-associated bacteria can quickly evolve along the symbiotic connection within a few growth cycles, our results urge eco-evolutionary management of endophytic bacteria and plants interactions in agriculture (Aswani et al., 2020).
Crop productivity is impacted by a number of common plant diseases that are present worldwide. Some of the serious ones are wilt disease, root rot, powdery mildew, leaf spot, leaf curl, and blight. To counter these phytopathogens, endophytic bacteria are crucial (Latha et al., 2019).
By producing proteins associated with pathogenesis (PRPs) and defense enzymes that stop the growth of phytopathogens that cause disease, endophytic bacteria can produce siderophores, antimicrobial compounds, and systemic resistance (Pandey et al., 2019). Bacterial endophytes are also potentially useful biocontrol agents. Plant diseases degrade plant performance and crop quality, which reduces crop output (Muthukumar et al., 2017). It has been shown that the nitrogen-fixing bacteria Azotobacter chrococcum, the phosphate-solubilizing bacteria PSB (Pseudomonas cepacia), the endophytic bacterial strains Lysinibacillus sp. and Bacillus subtilis, and their combination as bio-fertilizers can reduce the incidence of bacterial wilt disease in chili plants by up to 80% (Tewari et al., 2019).
The endophytic bacterial strain B. subtilis showed the strongest (80%) illness suppression (Jacob et al., 2020). This endophyte could also considerably aid the growth of the chili. Chemical pesticides are typically used to manage such phytopathogens, but this tactic has raised concerns about environmental contamination and contributed to the emergence of resistance to specific chemicals over time (Prasad et al., 2020). New insecticides must always be developed to address this. Chemical pesticides are thought to be ineffective when compared to endophytic bacteria acting as biocontrol agents or bioinsecticides. A broad array of mechanisms, including direct antagonism via the generation of antibiotics, siderophores, hydrogen cyanide, hydrolytic enzymes (chitinases, proteases, and lipases), etc., are involved in the biocontrol of plant diseases (Puri et al., 2017).
Some of the bacterial endophytes or PGPR are commonly used to control different diseases and as biological control agents so nowadays most of the focus is the understanding of complex interactions and their mechanisms and outcome either beneficial or harmful. It is hard to find the exact mechanism of interaction among complex microbial populations residing in the soil and environment near to host. So that proper characterization and management strategies can be devised according to the current need of time. In recent time peoples are preferring organic food and disliked the use of fertilizers and chemicals in agriculture. As the world population is increasing and food shortage issues are raised, in the current situation food security is an important topic for debate. Hence bacterial endophytes can be used as an alternative to chemical fertilizers, nutrient sources, and biological control agents for various plant pathogens. Scientists are focusing on the use of these endophytes in the form of biopesticides, and biofertilizers with different trade names for the control of different diseases and sustainable agricultural systems. Although the application of these endophytes in combination may lead to the development of optimum PGPEs inoculants that robust, and slight variation of environmental factors will not affect the plant growth promotion.
SM, MN, MH, MS, and MA conceived and conceptualized the study. MAS, AS, MB provided materials and technical assistance. SM, MS wrote original draft. SS, MSH, MS and MT technically reviewed and finalized the draft. All authors contributed to the article and approved the submitted version.
The authors are thankful to the University of the Punjab Lahore, Pakistan for providing the facilities for study.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Adesemoye, A., Obini, M., Ugoji, E. (2008). Comparison of plant growth-promotion with pseudomonas aeruginosa and bacillus subtilis in three vegetables. Braz. J. Microbiol. 39, 423–426. doi: 10.1590/S1517-83822008000300003
Agler, M. T., Ruhe, J., Kroll, S., Morhenn, C., Kim, S.-T., Weigel, D., et al. (2016). Microbial hub taxa link host and abiotic factors to plant microbiome variation. PloS Biol. 14, e1002352. doi: 10.1371/journal.pbio.1002352
Ahemad, M., Khan, M. S. (2012). Effect of fungicides on plant growth promoting activities of phosphate solubilizing pseudomonas putida isolated from mustard (Brassica compestris) rhizosphere. Chemosphere 86, 945–950. doi: 10.1016/j.chemosphere.2011.11.013
Akköprü, A., Akat, Ş., Özaktan, H., Gül, A., Akbaba, M. (2021). The long-term colonization dynamics of endophytic bacteria in cucumber plants, and their effects on yield, fruit quality and angular leaf spot disease. Scientia Hortic. 282, 110005. doi: 10.1016/j.scienta.2021.110005
Alaimo, S., Marceca, G. P., Giugno, R., Ferro, A., Pulvirenti, A. (2018). Current knowledge and computational techniques for grapevine meta-omics analysis. Front. Plant Sci. 8, 2241. doi: 10.3389/fpls.2017.02241
Anjum, M., Sajjad, M., Akhtar, N., Qureshi, M., Iqbal, A., Rehman, A. (2007). Response of cotton to plant growth promoting rhizobacteria (PGPR) inoculation under different levels of nitrogen. J. Agric. Res. (Pakistan).
Aswani, R., Jishma, P., Radhakrishnan, E. (2020). “Endophytic bacteria from the medicinal plants and their potential applications,” in Microbial endophytes (Elsevier). Available at: https://www.sciencedirect.com/science/article/pii/B9780128187340000024.
Barrera, M. C., Jakobs-Schoenwandt, D., Gómez, M. I., Serrato, J., Ruppel, S., Patel, A. V. (2020). Formulating bacterial endophyte: Pre-conditioning of cells and the encapsulation in amidated pectin beads. Biotechnol. Rep. 26, e00463. doi: 10.1016/j.btre.2020.e00463
Bashan, Y., González, L. E. (1999). Long-term survival of the plant-growth-promoting bacteria azospirillum brasilense and pseudomonas fluorescens in dry alginate inoculant. Appl. Microbiol. Biotechnol. 51, 262–266. doi: 10.1007/s002530051391
Basile, L. A., Lepek, V. C. (2021). Legume–rhizobium dance: an agricultural tool that could be improved? Microbial Biotechnol. 14, 1897–1917.
Basu, S., Sarkar, S., Prasad, R., Kumar, G. (2022). Current approaches for alleviating abiotic stress tolerance in crops: Role of beneficial microorganisms. Beneficial Microorganisms Agric., 231–248. doi: 10.1007/978-981-19-0733-3_9
Berendsen, R. L., Pieterse, C. M., Bakker, P. A. (2012). The rhizosphere microbiome and plant health. Trends Plant Sci. 17, 478–486. doi: 10.1016/j.tplants.2012.04.001
Bergersen, F. (1961). The growth of rhizobium in synthetic media. Aust. J. Biol. Sci. 14, 349–360. doi: 10.1071/BI9610349
Bertin, C., Yang, X., Weston, L. A. (2003). The role of root exudates and allelochemicals in the rhizosphere. Plant Soil 256, 67–83. doi: 10.1023/A:1026290508166
Bever, J. D. (2003). Soil community feedback and the coexistence of competitors: conceptual frameworks and empirical tests. New Phytol. 157, 465–473. doi: 10.1046/j.1469-8137.2003.00714.x
Bever, J. D., Dickie, I. A., Facelli, E., Facelli, J. M., Klironomos, J., Moora, M., et al. (2010). Rooting theories of plant community ecology in microbial interactions. Trends Ecol. Evol. 25, 468–478. doi: 10.1016/j.tree.2010.05.004
Brader, G., Compant, S., Mitter, B., Trognitz, F., Sessitsch, A. (2014). Metabolic potential of endophytic bacteria. Curr. Opin. Biotechnol. 27, 30–37. doi: 10.1016/j.copbio.2013.09.012
Bulgarelli, D., Rott, M., Schlaeppi, K., Ver Loren Van Themaat, E., Ahmadinejad, N., Assenza, F., et al. (2012). Revealing structure and assembly cues for arabidopsis root-inhabiting bacterial microbiota. Nature 488, 91–95. doi: 10.1038/nature11336
Bulgarelli, D., Schlaeppi, K., Spaepen, S., Van Themaat, E. V. L., Schulze-Lefert, P. (2013). Structure and functions of the bacterial microbiota of plants. Annu. Rev. Plant Biol. 64, 807–838. doi: 10.1146/annurev-arplant-050312-120106
Burd, G. I., Dixon, D. G., Glick, B. R. (2000). Plant growth-promoting bacteria that decrease heavy metal toxicity in plants. Can. J. Microbiol. 46, 237–245. doi: 10.1139/w99-143
Cano-Salazar, L., Gregorio-Jáuregui, K. M., Juárez-Ordaz, A. J., Leon-Joublanc, E., Perez-Molina, A., Martínez-Hernández, J. L., et al. (2011). Thermodynamics of partitioning of chitinase and laminarinase in a soya lecithin liposome system and their antifungal effect against fusarium oxysporum. Biocatalysis Biotransformation 29, 60–70. doi: 10.3109/10242422.2011.563445
Chebotar, V., Malfanova, N., Shcherbakov, A., Ahtemova, G., Borisov, A. Y., Lugtenberg, B., et al. (2015). Endophytic bacteria in microbial preparations that improve plant development. Appl. Biochem. Microbiol. 51, 271–277. doi: 10.1134/S0003683815030059
Chen, W., Wang, E., Ji, Z., Zhang, J. (2021). Recent development and new insight of diversification and symbiosis specificity of legume rhizobia: mechanism and application. J. Appl. Microbiol. 131, 553–563. doi: 10.1111/jam.14960
Clarholm, M. (1985). Interactions of bacteria, protozoa and plants leading to mineralization of soil nitrogen. Soil Biol. Biochem. 17, 181–187. doi: 10.1016/0038-0717(85)90113-0
Cruz Barrera, F. M. (2019). Development of an integrated fermentation and formulation approach for the endophytic bacterium kosakonia radicincitans DSM16656T as a novel plant biostimulant.
Cruz Barrera, M., Jakobs-Schoenwandt, D., Persicke, M., Gómez, M. I., Ruppel, S., Patel, A. V. (2020). Anhydrobiotic engineering for the endophyte bacterium kosakonia radicincitans by osmoadaptation and providing exogenously hydroxyectoine. World J. Microbiol. Biotechnol. 36, 1–16.
Curá, J. A., Franz, D. R., Filosofía, J. E., Balestrasse, K. B., Burgueño, L. E. (2017). Inoculation with azospirillum sp. and herbaspirillum sp. bacteria increases the tolerance of maize to drought stress. Microorganisms 5, 41.
Dary, M., Chamber-Pérez, M., Palomares, A., Pajuelo, E. (2010). “In situ” phytostabilisation of heavy metal polluted soils using lupinus luteus inoculated with metal resistant plant-growth promoting rhizobacteria. J. Hazardous Materials 177, 323–330. doi: 10.1016/j.jhazmat.2009.12.035
Delaux, P.-M., Radhakrishnan, G. V., Jayaraman, D., Cheema, J., Malbreil, M., Volkening, J. D., et al. (2015). Algal ancestor of land plants was preadapted for symbiosis. Proc. Natl. Acad. Sci. 112, 13390–13395. doi: 10.1073/pnas.1515426112
Del Carmen Orozco-Mosqueda, M., Glick, B. R., Santoyo, G. (2020). ACC deaminase in plant growth-promoting bacteria (PGPB): an efficient mechanism to counter salt stress in crops. Microbiological Res. 235, 126439. doi: 10.1016/j.micres.2020.126439
Dell’amico, E., Cavalca, L., Andreoni, V. (2008). Improvement of brassica napus growth under cadmium stress by cadmium-resistant rhizobacteria. Soil Biol. Biochem. 40, 74–84. doi: 10.1016/j.soilbio.2007.06.024
Deng, M.-C., Li, J., Liang, F.-R., Yi, M., Xu, X.-M., Yuan, J.-P., et al. (2014). Isolation and characterization of a novel hydrocarbon-degrading bacterium achromobacter sp. HZ01 from the crude oil-contaminated seawater at the daya bay, southern China. Mar. pollut. Bull. 83, 79–86. doi: 10.1016/j.marpolbul.2014.04.018
De Oliveira, E. J., Rabinovitch, L., Monnerat, R. G., Passos, L. K. J., Zahner, V. (2004). Molecular characterization of brevibacillus laterosporus and its potential use in biological control. Appl. Environ. Microbiol. 70, 6657–6664. doi: 10.1128/AEM.70.11.6657-6664.2004
Dixit, R., Kamat, S., Srivastava, A., Kumari, M. (2022). “Molecular basis of plant-PGPM interactions during amelioration of biotic stress,” in Microbial biocontrol: Food security and post harvest management (Springer). Available at: https://link.springer.com/chapter/10.1007/978-3-030-87289-2_5.
Dubey, A., Malla, M. A., Kumar, A., Dayanandan, S., Khan, M. L. (2020). Plants endophytes: unveiling hidden agenda for bioprospecting toward sustainable agriculture. Crit. Rev. Biotechnol. 40, 1210–1231. doi: 10.1080/07388551.2020.1808584
Dudai, N. (2011). “Domestication and breeding of wild medicinal and aromatic plants-thirty years of experience in Israel,” in I International symposium on medicinal, aromatic and nutraceutical plants from mountainous areas (MAP-mountain 2011), vol. 955, 175–183. Available at: https://www.actahort.org/books/955/955_25.htm.
Edwards, R., Harding, K. G. (2004). Bacteria and wound healing. Curr. Opin. Infect. Dis. 17, 91–96. doi: 10.1097/00001432-200404000-00004
Eid, A. M., Salim, S. S., Hassan, S. E.-D., Ismail, M. A., Fouda, A. (2019). “Role of endophytes in plant health and abiotic stress management,” in Microbiome in plant health and disease (Springer). Available at: https://link.springer.com/chapter/10.1007/978-981-13-8495-0_6.
Fawcett, H. (1944). Fungus and bacterial diseases of insects as factors in biological control. Botanical Rev. 10, 327–348. doi: 10.1007/BF02861103
Fayez, M., Daw, Z. (1987). Effect of inoculation with different strains of azospirillum brasilense on cotton (Gossipium barbadense). Biol. fertility soils 4, 91–95. doi: 10.1007/BF00280357
Forchetti, G., Masciarelli, O., Alemano, S., Alvarez, D., Abdala, G. (2007). Endophytic bacteria in sunflower (Helianthus annuus l.): isolation, characterization, and production of jasmonates and abscisic acid in culture medium. Appl. Microbiol. Biotechnol. 76, 1145–1152. doi: 10.1007/s00253-007-1077-7
Foster, R. (1988). Microenvironments of soil microorganisms. Biol. fertility soils 6, 189–203. doi: 10.1007/BF00260816
Gaiero, J. R., Mccall, C. A., Thompson, K. A., Day, N. J., Best, A. S., Dunfield, K. E. (2013). Inside the root microbiome: bacterial root endophytes and plant growth promotion. Am. J. Bot. 100, 1738–1750. doi: 10.3732/ajb.1200572
Gholami, A., Shahsavani, S., Nezarat, S. (2009). The effect of plant growth promoting rhizobacteria (PGPR) on germination, seedling growth and yield of maize. Int. J. Agric. Biosyst. Eng. 3, 9–14.
Glick, B. R. (2012). Plant growth-promoting bacteria: mechanisms and applications. Scientifica 2012. doi: 10.6064/2012/963401
Glick, B. R., Holguin, G., Patten, C., Penrose, D. M. (1999). Biochemical and genetic mechanisms used by plant growth promoting bacteria (World Scientific). Available at: https://books.google.com.pk/books?hl=en&lr=&id=vPq3CgAAQBAJ&oi=fnd&pg=PP9&dq=Glick,+B.+R.,+Holguin,+G.,+Patten,+C.+%26+Penrose,+D.+M.+1999.+Biochemical+and+genetic+mechanisms+used+by+plant+growth+promoting+bacteria,+World+Scientific.&ots=7T3cBYAz5r&sig=5R7x1cvIBN5AVox2Xx_847srclY&redir_esc=y#v=onepage&q=Glick%2C%20B.%20R.%2C%20Holguin%2C%20G.%2C%20Patten%2C%20C.%20%26%20Penrose%2C%20D.%20M.%201999.%20Biochemical%20and%20genetic%20mechanisms%20used%20by%20plant%20growth%20promoting%20bacteria%2C%20World%20Scientific.&f=false.
González Candia, P. (2021). Rizobacterias que promueven el enraizamiento de miniestacas de híbridos de eucalyptus SP. y sus posibles mecanismos de acción. Available at: http://repositorio.udec.cl/handle/11594/9362.
Gupta, A., Meyer, J. M., Goel, R. (2002). Development of heavy metal-resistant mutants of phosphate solubilizing pseudomonas sp. NBRI 4014 and their characterization. Curr. Microbiol. 45, 323–327. doi: 10.1007/s00284-002-3762-1
Gupta, A., Rai, V., Bagdwal, N., Goel, R. (2005). In situ characterization of mercury-resistant growth-promoting fluorescent pseudomonads. Microbiological Res. 160, 385–388. doi: 10.1016/j.micres.2005.03.002
Hong, C. E., Park, J. M. (2016). Endophytic bacteria as biocontrol agents against plant pathogens: current state-of-the-art. Plant Biotechnol. Rep. 10, 353–357. doi: 10.1007/s11816-016-0423-6
Honma, M., Shimomura, T. (1978). Metabolism of 1-aminocyclopropane-1-carboxylic acid. Agric. Biol. Chem. 42, 1825–1831.
Indiragandhi, P., Anandham, R., Kim, K., Yim, W., Madhaiyan, M., Sa, T. (2008). Induction of defense responses in tomato against pseudomonas syringae pv. tomato by regulating the stress ethylene level with methylobacterium oryzae CBMB20 containing 1-aminocyclopropane-1-carboxylate deaminase. World J. Microbiol. Biotechnol. 24, 1037–1045. doi: 10.1007/s11274-007-9572-7
Jacob, J., Krishnan, G. V., Thankappan, D., Amma, D. K. B. N. S. (2020). “Endophytic bacterial strains induced systemic resistance in agriculturally important crop plants,” in Microbial endophytes (Elsevier). Available at: https://www.sciencedirect.com/science/article/pii/B9780128196540000041.
Jahanian, A., Chaichi, M., Rezaei, K., Rezayazdi, K., Khavazi, K. (2012). The effect of plant growth promoting rhizobacteria (PGPR) on germination and primary growth of artichoke (Cynara scolymus). Int. J. Agric. Crop Sci. (IJACS) 4, 923–929.
Jha, A. K. (2017). Effect of lime, compost and microbial inoculants on micronutrient removal by mustard and maize in trace metal contaminated soil of jharkhand. Curr. Agric. Res. J. 5, 196. doi: 10.12944/CARJ.5.2.07
Jonkers, H. M., Ludwig, R., De Wit, R., Pringault, O., Muyzer, G., Niemann, H., et al. (2003). Structural and functional analysis of a microbial mat ecosystem from a unique permanent hypersaline inland lake:’La salada de Chiprana’(NE Spain). FEMS Microbiol. Ecol. 44, 175–189. doi: 10.1016/S0168-6496(02)00464-6
Kalyani, D. C., Patil, P. S., Jadhav, J. P., Govindwar, S. P. (2008). Biodegradation of reactive textile dye red BLI by an isolated bacterium pseudomonas sp. SUK1. Bioresource Technol. 99, 4635–4641. doi: 10.1016/j.biortech.2007.06.058
Kandel, S., Herschberger, N., Kim, S., Doty, S. (2015). Diazotrophic endophytes of poplar and willow for growth promotion of rice plants in nitrogen-limited conditions. Crop Sci. 55, 1765–1772. doi: 10.2135/cropsci2014.08.0570
Khalil, A. M. A., Hassan, S. E.-D., Alsharif, S. M., Eid, A. M., Ewais, E. E.-D., Azab, E., et al. (2021). Isolation and characterization of fungal endophytes isolated from medicinal plant ephedra pachyclada as plant growth-promoting. Biomolecules 11, 140. doi: 10.3390/biom11020140
Khan, Z., Doty, S. L. (2009). Characterization of bacterial endophytes of sweet potato plants. Plant Soil 322, 197–207. doi: 10.1007/s11104-009-9908-1
Khare, E., Mishra, J., Arora, N. K. (2018). Multifaceted interactions between endophytes and plant: developments and prospects. Front. Microbiol. 9, 2732. doi: 10.3389/fmicb.2018.02732
Lata, R. K., Divjot, K., Nath, Y. A. (2019). Endophytic microbiomes: biodiversity, ecological significance and biotechnological applications. Res. J. Biotechnol. 14, 10.
Lawongsa, P., Boonkerd, N., Wongkaew, S., O’gara, F., Teaumroong, N. (2008). Molecular and phenotypic characterization of potential plant growth-promoting pseudomonas from rice and maize rhizospheres. World J. Microbiol. Biotechnol. 24, 1877–1884. doi: 10.1007/s11274-008-9685-7
Ledermann, R., Schulte, C. C., Poole, P. S. (2021). How rhizobia adapt to the nodule environment. J. bacteriology 203, e00539–e00520. doi: 10.1128/JB.00539-20
Leopold, A. C. (1964). “Plant growth and development,” in Plant growth and development. Available at: https://www.cabdirect.org/cabdirect/abstract/19670701019?freeview=true.
Lindström, K., Mousavi, S. A. (2020). Effectiveness of nitrogen fixation in rhizobia. Microbial Biotechnol. 13, 1314–1335. doi: 10.1111/1751-7915.13517
Liu, H., Carvalhais, L. C., Crawford, M., Singh, E., Dennis, P. G., Pieterse, C. M., et al. (2017>2552). Inner plant values: diversity, colonization and benefits from endophytic bacteria. Front. Microbiol. 8. doi: 10.3389/fmicb.2017.02552
Liu, W., Hou, J., Wang, Q., Ding, L., Luo, Y. (2014). Isolation and characterization of plant growth-promoting rhizobacteria and their effects on phytoremediation of petroleum-contaminated saline-alkali soil. Chemosphere 117, 303–308. doi: 10.1016/j.chemosphere.2014.07.026
Lobo, C. B., Tomás, M. S. J., Viruel, E., Ferrero, M. A., Lucca, M. E. (2019). Development of low-cost formulations of plant growth-promoting bacteria to be used as inoculants in beneficial agricultural technologies. Microbiological Res. 219, 12–25. doi: 10.1016/j.micres.2018.10.012
Lodwig, E., Poole, P. (2003). Metabolism of rhizobium bacteroids. Crit. Rev. Plant Sci. 22, 37–78. doi: 10.1080/713610850
Lugtenberg, B., Kamilova, F. (2009). Plant-growth-promoting rhizobacteria. Annu. Rev. Microbiol. 63, 541–556. doi: 10.1146/annurev.micro.62.081307.162918
Luu, M., Monning, H., Visekruna, A. (2020). Exploring the molecular mechanisms underlying the protective effects of microbial SCFAs on intestinal tolerance and food allergy. Front. Immunol. 11 doi: 10.3389/fimmu.2020.01225
Lynch, J. M., Brimecombe, M. J., De Leij, F. A. (2001). Rhizosphere. e LS. doi: 10.1038/npg.els.0000403
Maksimov, I., Maksimova, T., Sarvarova, E., Blagova, D., Popov, V. (2018). Endophytic bacteria as effective agents of new-generation biopesticides. Appl. Biochem. Microbiol. 54, 128–140. doi: 10.1134/S0003683818020072
Mayak, S., Tirosh, T., Glick, B. (1999). Effect of wild-type and mutant plant growth-promoting rhizobacteria on the rooting of mung bean cuttings. J. Plant Growth Regul. 18, 49–53. doi: 10.1007/PL00007047
Mercado-Blanco, J., Jj Lugtenberg, B. (2014). Biotechnological applications of bacterial endophytes. Curr. Biotechnol. 3, 60–75. doi: 10.2174/22115501113026660038
Morgan, J., Bending, G., White, P. (2005). Biological costs and benefits to plant–microbe interactions in the rhizosphere. J. Exp. Bot. 56, 1729–1739. doi: 10.1093/jxb/eri205
Muthukumar, A., Udhayakumar, R., Naveenkumar, R. (2017). “Role of bacterial endophytes in plant disease control,” in Endophytes: crop productivity and protection (Springer). Available at: https://link.springer.com/chapter/10.1007/978-3-319-66544-3_7.
Navas, L. E., Amadio, A. F., Ortiz, E. M., Sauka, D. H., Benintende, G. B., Berretta, M. F., et al. (2017). Complete sequence and organization of pFR260, the bacillus thuringiensis INTA Fr7-4 plasmid harboring insecticidal genes. Microbial Physiol. 27, 43–54. doi: 10.1159/000451056
Oldroyd, G. E., Murray, J. D., Poole, P. S., Downie, J. A. (2011). The rules of engagement in the legume-rhizobial symbiosis. Annu. Rev. Genet. 45, 119–144. doi: 10.1146/annurev-genet-110410-132549
Oteino, N., Lally, R. D., Kiwanuka, S., Lloyd, A., Ryan, D., Germaine, K. J., et al. (2015). Plant growth promotion induced by phosphate solubilizing endophytic pseudomonas isolates. Front. Microbiol. 6, 745. doi: 10.3389/fmicb.2015.00745
Ouyabe, M., Kikuno, H., Tanaka, N., Babil, P., Shiwachi, H. (2019). Endophytic nitrogen-fixing bacteria of water yam (Dioscorea alata l.) in relation with fertilization practices. Trop. Agric. Dev. 63, 122–130.
Pandey, P. K., Samanta, R., Yadav, R. N. S. (2019). Inside the plant: addressing bacterial endophytes in biotic stress alleviation. Arch. Microbiol. 201, 415–429. doi: 10.1007/s00203-019-01642-y
Pandey, P. K., Singh, M. C., Singh, S., Kumar, A., Pathak, M., Shakywar, R., et al. (2017). Inside the plants: endophytic bacteria and their functional attributes for plant growth promotion. Int. J. Curr. Microbiol. Appl. Sci. 6, 11–21. doi: 10.20546/ijcmas.2017.602.002
Pedrotti, L., Mueller, M. J., Waller, F. (2013). Piriformospora indica root colonization triggers local and systemic root responses and inhibits secondary colonization of distal roots. PloS One 8, e69352. doi: 10.1371/journal.pone.0069352
Prasad, M., Srinivasan, R., Chaudhary, M., Mahawer, S. K., Jat, L. K. (2020). “Endophytic bacteria: Role in sustainable agriculture,” in Microbial endophytes (Elsevier). Available at: https://www.sciencedirect.com/science/article/pii/B9780128187340000036.
Puri, A., Padda, K. P., Chanway, C. P. (2017). “Plant growth promotion by endophytic bacteria in nonnative crop hosts,” in Endophytes: crop productivity and protection (Springer). Available at: https://link.springer.com/chapter/10.1007/978-3-319-66544-3_2.
Rajkumar, M., Nagendran, R., Lee, K. J., Lee, W. H., Kim, S. Z. (2006). Influence of plant growth promoting bacteria and Cr6+ on the growth of Indian mustard. Chemosphere 62, 741–748. doi: 10.1016/j.chemosphere.2005.04.117
Rana, K. L., Kour, D., Kaur, T., Devi, R., Yadav, A. N., Yadav, N., et al. (2020a). Endophytic microbes: biodiversity, plant growth-promoting mechanisms and potential applications for agricultural sustainability. Antonie Van Leeuwenhoek 113, 1075–1107. doi: 10.1007/s10482-020-01429-y
Rana, K. L., Kour, D., Kaur, T., Sheikh, I., Yadav, A. N., Kumar, V., et al. (2020b). Endophytic microbes from diverse wheat genotypes and their potential biotechnological applications in plant growth promotion and nutrient uptake. Proc. Natl. Acad. Sciences India Section B: Biol. Sci. 90, 969–979. doi: 10.1007/s40011-020-01168-0
Ratu, S. T. N., Teulet, A., Miwa, H., Masuda, S., Nguyen, H. P., Yasuda, M., et al. (2021). Rhizobia use a pathogenic-like effector to hijack leguminous nodulation signalling. Sci. Rep. 11, 1–15. doi: 10.1038/s41598-021-81598-6
Rekha, P., Lai, W.-A., Arun, A., Young, C.-C. (2007). Effect of free and encapsulated pseudomonas putida CC-FR2-4 and bacillus subtilis CC-pg104 on plant growth under gnotobiotic conditions. Bioresource Technol. 98, 447–451. doi: 10.1016/j.biortech.2006.01.009
Remans, R., Beebe, S., Blair, M., Manrique, G., Tovar, E., Rao, I., et al. (2008). Physiological and genetic analysis of root responsiveness to auxin-producing plant growth-promoting bacteria in common bean (Phaseolus vulgaris l.). Plant Soil 302, 149–161. doi: 10.1007/s11104-007-9462-7
Robinson, B., Russell, C., Hedley, M., Clothier, B. (2001). Cadmium adsorption by rhizobacteria: implications for new Zealand pastureland. Agriculture Ecosyst. Environ. 87, 315–321. doi: 10.1016/S0167-8809(01)00146-3
Ruduś, I., Sasiak, M., Kępczyński, J. (2013). Regulation of ethylene biosynthesis at the level of 1-aminocyclopropane-1-carboxylate oxidase (ACO) gene. Acta Physiologiae Plantarum 35, 295–307. doi: 10.1007/s11738-012-1096-6
Ryan, R. P., Germaine, K., Franks, A., Ryan, D. J., Dowling, D. N. (2008). Bacterial endophytes: recent developments and applications. FEMS Microbiol. Lett. 278, 1–9. doi: 10.1111/j.1574-6968.2007.00918.x
Safronova, V. I., Stepanok, V. V., Engqvist, G. L., Alekseyev, Y. V., Belimov, A. A. (2006). Root-associated bacteria containing 1-aminocyclopropane-1-carboxylate deaminase improve growth and nutrient uptake by pea genotypes cultivated in cadmium supplemented soil. Biol. Fertility Soils 42, 267–272. doi: 10.1007/s00374-005-0024-y
Sant'anna, F. H., Almeida, L. G., Cecagno, R., Reolon, L. A., Siqueira, F. M., Machado, M. R., et al. (2011). Genomic insights into the versatility of the plant growth-promoting bacterium azospirillum amazonense. BMC Genomics 12, 1–14. doi: 10.1186/1471-2164-12-409
Santos, M. L. D., Berlitz, D. L., Wiest, S. L. F., Schünemann, R., Knaak, N., Fiuza, L. M. (2018). Benefits associated with the interaction of endophytic bacteria and plants. Braz. Arch. Biol. Technol. 61. doi: 10.1590/1678-4324-2018160431
Sapak, Z., Meon, S., Ahmad, Z. A. M. (2008). Effect of endophytic bacteria on growth and suppression of ganoderma infection in oil palm. Int. J. Agric. Biol. 10, 127–132.
Schimel, J., Balser, T. C., Wallenstein, M. (2007). Microbial stress-response physiology and its implications for ecosystem function. Ecology 88, 1386–1394. doi: 10.1890/06-0219
Schnitzer, S. A., Klironomos, J. (2011). Soil microbes regulate ecosystem productivity and maintain species diversity. Plant Signaling Behav. 6, 1240–1243. doi: 10.4161/psb.6.8.16455
Sharma, S., Chandra, D., Sharma, A. K. (2021). “Rhizosphere plant–microbe interactions under abiotic stress,” in Rhizosphere biology: Interactions between microbes and plants (Springer). Available at: https://link.springer.com/chapter/10.1007/978-981-15-6125-2_10.
Sharma, S. K., Johri, B. N., Ramesh, A., Joshi, O. P., Sai Prasad, S. (2011). Selection of plant growth-promoting pseudomonas spp. that enhanced productivity of soybean-wheat cropping system in central India. J. Microbiol. Biotechnol. 21, 1127–1142. doi: 10.4014/jmb.1012.12018
Shen, W., Cevallos-Cevallos, J. M., Nunes Da Rocha, U., Arevalo, H. A., Stansly, P. A., Roberts, P. D., et al. (2013). Relation between plant nutrition, hormones, insecticide applications, bacterial endophytes, and candidatus liberibacter ct values in citrus trees infected with huanglongbing. Eur. J. Plant Pathol. 137, 727–742. doi: 10.1007/s10658-013-0283-7
Singh, J. S. (2015). Microbes play major roles in the ecosystem services. Climate Change Environ. Sustainability 3, 163–167.
Skinner, M., Parker, B. L., Kim, J. S. (2014). Role of entomopathogenic fungi in integrated pest management. Integrated Pest Manage., 169–191. doi: 10.1016/B978-0-12-398529-3.00011-7
Suslow, T., Schroth, M. (1982). Role of deleterious rhizobacteria as minor pathogens in reducing crop growth. Phytopathology 72, 111–115. doi: 10.1094/Phyto-72-111
Tank, N., Saraf, M. (2010). Salinity-resistant plant growth promoting rhizobacteria ameliorates sodium chloride stress on tomato plants. J. Plant Interact. 5, 51–58. doi: 10.1080/17429140903125848
Teixeira, L. C., Yeargeau, E., Balieiro, F. C., Piccolo, M. C., Peixoto, R. S., Greer, C. W., et al. (2013). Plant and bird presence strongly influences the microbial communities in soils of admiralty bay, maritime Antarctica. PloS One 8, e66109. doi: 10.1371/journal.pone.0066109
Tewari, S., Shrivas, V. L., Hariprasad, P., Sharma, S. (2019). “Harnessing endophytes as biocontrol agents,” in Plant health under biotic stress (Springer). Available at: https://link.springer.com/chapter/10.1007/978-981-13-6040-4_10.
Thakuria, D., Talukdar, N., Goswami, C., Hazarika, S., Boro, R., Khan, M. (2004). Characterization and screening of bacteria from rhizosphere of rice grown in acidic soils of Assam. Curr. Sci., 978–985.
Timper, P., Kone, D., Yin, J., Ji, P., Gardener, B. B. M. (2009). Evaluation of an antibiotic-producing strain of pseudomonas fluorescens for suppression of plant-parasitic nematodes. J. Nematol. 41, 234.
Tiwari, R. K., Lal, M. K., Naga, K. C., Kumar, R., Chourasia, K. N., Subhash, S., et al. (2020). Emerging roles of melatonin in mitigating abiotic and biotic stresses of horticultural crops. Scientia Hortic. 272, 109592. doi: 10.1016/j.scienta.2020.109592
Vandana, U. K., Rajkumari, J., Singha, L. P., Satish, L., Alavilli, H., Sudheer, P. D., et al. (2021). The endophytic microbiome as a hotspot of synergistic interactions, with prospects of plant growth promotion. Biology 10, 101. doi: 10.3390/biology10020101
Van Rhijn, P., Vanderleyden, J. (1995). The rhizobium-plant symbiosis. Microbiological Rev. 59, 124–142. doi: 10.1128/mr.59.1.124-142.1995
Vardi, R., Mittermeier, J. C., Roll, U. (2021). Combining culturomic sources to uncover trends in popularity and seasonal interest in plants. Conserv. Biol. 35, 460–471. doi: 10.1111/cobi.13705
Vishwakarma, K., Kumar, N., Shandilya, C., Varma, A. (2021). “Unravelling the role of endophytes in micronutrient uptake and enhanced crop productivity,” in Symbiotic soil microorganisms (Springer). Available at: https://link.springer.com/chapter/10.1007/978-981-13-6040-4_10.
Wani, P. A., Khan, M. S., Zaidi, A. (2007). Effect of metal tolerant plant growth promoting bradyrhizobium sp.(vigna) on growth, symbiosis, seed yield and metal uptake by greengram plants. Chemosphere 70, 36–45. doi: 10.1016/j.chemosphere.2007.07.028
Wani, P. A., Khan, M., Zaidi, A. (2008). Effect of metal-tolerant plant growth-promoting rhizobium on the performance of pea grown in metal-amended soil. Arch. Environ. Contamination Toxicol. 55, 33–42. doi: 10.1007/s00244-007-9097-y
Wood, T. E., Cavaleri, M. A., Reed, S. C. (2012). Tropical forest carbon balance in a warmer world: A critical review spanning microbial-to ecosystem-scale processes. Biol. Rev. 87, 912–927. doi: 10.1111/j.1469-185X.2012.00232.x
Wu, W., Jin, Y., Bai, F., Jin, S. (2015). “Pseudomonas aeruginosa,” in Molecular medical microbiology (Elsevier). Available at: https://www.sciencedirect.com/science/article/pii/B978012397169200041X.
Wu, C. H., Wood, T. K., Mulchandani, A., Chen, W. (2006). Engineering plant-microbe symbiosis for rhizoremediation of heavy metals. Appl. Environ. Microbiol. 72, 1129–1134. doi: 10.1128/AEM.72.2.1129-1134.2006
Keywords: host endosymbiont interactions, mechanism of interaction, bacterial endophytes, plants, endophytic
Citation: Mushtaq S, Shafiq M, Tariq MR, Sami A, Nawaz-ul-Rehman MS, Bhatti MHT, Haider MS, Sadiq S, Abbas MT, Hussain M and Shahid MA (2023) Interaction between bacterial endophytes and host plants. Front. Plant Sci. 13:1092105. doi: 10.3389/fpls.2022.1092105
Received: 07 November 2022; Accepted: 22 December 2022;
Published: 18 January 2023.
Edited by:
Mamoona Rauf, Abdul Wali Khan University Mardan, PakistanReviewed by:
Julio Alves Cardoso Filho, Federal University of Alagoas, BrazilCopyright © 2023 Mushtaq, Shafiq, Tariq, Sami, Nawaz-ul-Rehman, Bhatti, Haider, Sadiq, Abbas, Hussain and Shahid. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Muhammad Adnan Shahid, bXNoYWhpZEB1ZmwuZWR1; Muhammad Shafiq, c2hhZmlxLmlhZ3NAcHUuZWR1LnBr
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.