 Sung-Je Yoo1†
Sung-Je Yoo1† Nicolás M. Cecchini
Nicolás M. Cecchini Jean T. Greenberg
Jean T. Greenberg Ho Won Jung
Ho Won Jung- 1Department of Molecular Genetics, Dong-A University, Busan, South Korea
- 2Department of Applied Bioscience, Dong-A University, Busan, South Korea
- 3Departamento de Química Biológica Ranwel Caputto, Centro de Investigaciones en Química Biológica de Córdoba (CIQUIBIC), Consejo Nacional de Investigaciones Científicas y Técnicas (CONICET), Universidad Nacional de Córdoba, Córdoba, Argentina
- 4Department of Molecular Genetics and Cell Biology, The University of Chicago, Chicago, IL, United States
Immunity in plants arises from defense regulatory circuits that can be conceptualized as modules. Both the types (and isolates) of pathogen and the repertoire of plant receptors may cause different modules to be activated and affect the magnitude of activation. Two major defense enzymes of Arabidopsis are ALD1 and ICS1/SID2. ALD1 is an aminotransferase needed for producing the metabolites pipecolic acid, hydroxy-pipecolic acid, and possibly other defense signals. ICS1/SID2 produces isochorismate, an intermediate in the synthesis of salicylic acid (SA) and SA-derivatives. Metabolites resulting from the activation of these enzymes are found in petiole exudates and may serve as priming signals for systemic disease resistance in Arabidopsis. Mutants lacking ALD1 are known to have reduced SA accumulation. To further investigate the role of ALD1 in relation to the SA-related module, immunity phenotypes of double mutants that disrupt ALD1 and ICS1/SID2 or SA perception by NPR1 were compared with each single mutant after infection by different Pseudomonas strains. Exudates collected from these mutants after infection were also evaluated for their ability to confer disease resistance when applied to wild-type plants. During infection with virulent or attenuated strains, the loss of ALD1 does not increase the susceptibility of npr1 or sid2 mutants, suggesting the main role of ALD1 in this context is in amplifying the SA-related module. In contrast, after an infection that leads to strong pathogen recognition via the cytoplasmic immune receptor RPS2, ALD1 acts additively with both NPR1 and ICS1/SID2 to suppress pathogen growth. The additive effects are observed in early basal defense responses as well as SA-related events. Thus, there are specific conditions that dictate whether the modules independently contribute to immunity to provide additive protection during infection. In the exudate experiments, intact NPR1 and ICS1/SID2, but not ALD1 in the donor plants were needed for conferring immunity. Mixing exudates showed that loss of SID2 yields exudates that suppress active exudates from wild-type or ald1 plants. This indicates that ICS1/SID2 may not only lead to positive defense signals, but also prevent a suppressive signal(s).
Introduction
Plants have multilayered barriers to defend themselves against pathogen attacks. In the last few decades, plant immune receptors that initiate disease resistance against different infectious agents have been identified in model and crop plants (Han and Jung, 2013; Michelmore et al., 2013; Macho and Zipfel, 2014). Receptors that control resistance to bacterial pathogens mostly fall into two classes: cell surface-localized pattern recognition receptor (PRR) and cytoplasmic disease resistance (R) protein (also known as NOD-LIKE RECEPTOR [NLR] and nucleotide-binding site [NBS]-Leucine-rich repeat [LRR] protein). PRRs recognize microbe/pathogen-associated molecular patterns (MAMPs/PAMPs) and initiate pattern-triggered immunity (PTI), whereas R proteins are responsible for effector-triggered immunity (ETI) through direct or indirect recognition of the pathogen-derived effectors (Chisholm et al., 2006; Jones and Dangl, 2006; Jones et al., 2016). In general, the execution of PTI is characterized by an early Ca2+ and oxidative burst, transient dual phosphorylation of MITOGEN-ACTIVATED PROTEIN KINASE3 (MPK3) and MPK6, and induction of MAMP-responsive genes such as FLG22-INDUCED RECEPTOR-LIKE KINASE1 (FRK1) and NDR1/HIN1-LIKE10 (NHL10) (Nicaise et al., 2009; Serrano et al., 2007; Tsuda et al., 2013; Bigeard et al., 2015). In addition, salicylic acid (SA), a phytohormone critical for plant immunity to (hemi)biotrophic microbes, controls PRRs’ levels and intensifies the responsiveness of plants to MAMPs/PAMPs (Tateda et al., 2014). On the other hand, the most well-studied attribute of ETI is a hypersensitive response (HR), a type of programmed cell death occurring at infection sites (Mur et al., 2008). HR is accompanied by diverse cellular responses such as rapid ion leakage, chloroplast disruption, and DNA laddering (Coll et al., 2011). Various types of infections and/or MAMP treatments can have non-autonomous signaling effects that cause distal tissues to become more resistant to subsequent infections, a process called systemic acquired resistance (SAR) (Ryals et al., 1996; Mishina and Zeier, 2006; 2007; Zeidler et al., 2010; Jelenska et al., 2017; Cecchini et al., 2019).
Although PTI and ETI are initiated by different receptor systems, they share key defense components (Katagiri and Tsuda, 2010; Tsuda et al., 2013). Moreover, recent studies demonstrate that ETI enhances PTI and requires PTI for full ETI against avirulent Pseudomonas infection (Ngou et al., 2021; Yuan et al., 2021). However, there are still many questions about the relationship between PTI and ETI. One of the shared defense signals is SA, which is implicated in both PTI and ETI pathways (Glazebrook, 2005; Spoel and Dong, 2008; Tsuda et al., 2013; Tateda et al., 2014). In addition, SA is needed for the establishment of SAR (Métraux et al., 1990; Durrant and Dong, 2004). Dozens of proteins bring SA-dependent defenses to completion. Among them, the crucial defense factors, ICS1/SID2, NPR1, and ALD1, are important for the interconnectivity between PTI, ETI and a SA-dependent defense response. ISOCHORISMATE SYNTHASE1/SALICYLIC ACID INDUCTION DEFICIENT2 (ICS1/SID2) is directly engaged in SA biosynthesis (Nawrath and Métraux, 1999; Wildermuth et al., 2001), whereas NONEXPRESSOR OF PATHOGENESIS-RELATED PROTEINS1 (NPR1), a SA receptor, is necessary for the expression of SA-responsive genes (Cao et al., 1997; Fu et al., 2012; Wu et al., 2012). AGD2-LIKE DEFENSE PROTEIN1 (ALD1), a diaminopimelate aminotransferase, is responsible for early SA accumulation during ETI as well as for the biosynthesis of pipecolic acid (Pip)/N-hydroxy-Pip (NHP) during infections that lead to SAR (Song et al., 2004a; 2004b; Návarová et al., 2012; Sobolev et al., 2013; Hartmann et al., 2017; 2018; Chen et al., 2018; Jiang et al., 2021). ALD1 is also involved in PTI and the basal production of unidentified non-Pip molecule(s) that induces/maintains local disease resistance, but not systemic resistance (Cecchini et al., 2015). Pip, NHP, and other non-Pip molecules are proposed to be present in the petiole exudates of leaves infected with SAR-inducing Pseudomonas (Návarová et al., 2012; Chen et al., 2018; Hartmann et al., 2018; Jiang et al., 2021). Moreover, both ALD1 and ICS1/SID2 are required for the accumulation of the receptor for the MAMP flagellin, FLAGELLIN-SENSING2 (FLS2), in Arabidopsis, and NPR1 participates in the flg22 (a flagellin-derived peptide that triggers FLS2 signaling)-induced early immune response (Cecchini et al., 2015; Yi et al., 2014; 2015).
Two well-known R proteins activating strong ETI in Arabidopsisare the cytoplasmic receptors RESISTANCE TO PSEUDOMONAS SYRINGAE2 (RPS2) and RESISTANCE TO P. SYRINGAE PV MACULICOLA1 (RPM1). RPS2 and RPM1 confer resistance against infection by avirulent Pseudomonas syringae carrying AvrRpt2 and AvrRpm1, respectively (Kunkel et al., 1993; Bisgrove et al., 1994). Although RPS2- and RPM1-mediated immunity require common genes (e.g., NON-RACE-SPECIFIC DISEASE RESISTANCE1 [NDR1]), RPS2 signaling results in stronger SA-related defenses upon infection (Century et al., 1995; Belkhadir et al., 2004; Day et al., 2006; Lee et al., 2007). Avirulent P. syringae derivatives grow better, but cause weak or no detectable visible symptoms in leaves of ald1, npr1, and sid2 mutants, compared with wild type (WT), when the inoculum concentrations are low (e.g., OD600 = 0.0001) (Nawrath and Métraux, 1999; Clarke et al., 2000; Song et al., 2004b; Raffaele et al., 2006; Venugopal et al., 2009). Additionally, macroscopic HR-associated leaf collapse still occurs in these mutants, although NPR1 and ICS1/SID2 somewhat affect the development of HR symptoms after infection with avirulent strains of P. syringae (> OD600 = 0.01). (Nawrath and Métraux, 1999; Dewdney et al., 2000; Rate and Greenberg, 2001; Song et al., 2004b; Zhang et al., 2004; Munch et al., 2014). Microscopic early cell death response after similar avirulent P. syringae infections is highly reduced in sid2 plants (Seguel et al., 2018). These previous observations show that ALD1, NPR1, and ICS1/SID2 are engaged in R-mediated immunity as well as SA-dependent defense when avirulent pathogens infect Arabidopsis plants.
Although several genetic studies exist (Lu et al., 2009; Ng et al., 2011), the importance of different defense modules mediated by ALD1 and NPR1 or ICS1/SID2 during PTI and ETI have yet to be fully elucidated. In particular, because of ALD1’s role in positively affecting SA levels, we wished to test whether ALD1 acts primarily by amplifying SA levels or if it has an additive function with SA synthesis and/or transduction. We addressed this gap in knowledge and found that there are different requirements for ALD1, NPR1 and ICS1/SID2, depending on the P. syringae strain used for infection. In many infections, ALD1’s role is to amplify SA, but one avirulent infection uncovered an additive function for ALD1 with SA. We infer that the receptors used for detecting different strains influence how the defense network is modulated to achieve pathogen suppression. Finally, we also investigated the roles of ALD1, NPR1 and ICS1/SID2 in the production of active exudates that may contain mobile defense signals. We find different roles for these components in the production of active exudates and discuss the implications of these findings for understanding systemic signaling.
Materials and methods
Biological materials
A. thaliana seeds treated with 95% ethanol for 2 ~ 3 min were soaked in 50% bleach supplemented with 0.01% Triton X-100 for 15 min. The seeds were washed with deionized water several times to remove residual bleach. For most experiments (except those involving exudates-see section below), the resulting sterilized seeds were sown in potting mixture consisting of 59.26% peat moss, 20% cocopeat, and 20% perlite (Nongwoo Bio, Korea). Plants grew from 24 to 28 days in an environmentally controlled walk-in growth chamber (21 ± 1°C, 12 h day and 12 h night, 50 to 60% humidity). All seeds were in the Columbia (Col-0) background. To generate double mutants, ald1-T2 (ald1) was crossed with npr1-1 (npr1) or sid2-1 (sid2), and the resulting mutants were screened with T-DNA verification primers (ald1) and dCAPS markers (npr1 and sid2) (Table 1) (Cao et al., 1997; Nawrath and Métraux, 1999; Song et al., 2004a).

Table 1 Nucleotide sequences of primers used in this study.
Two virulent strains, P. syringae pv. maculicola ES4326 (re-classified as P. cannabina pv. alisalensis [Bull et al., 2010]) (PsmES4326) and P. syringae pv. tomato DC3000 (PstDC3000), and an attenuated strain PstDC3000 ΔAvrPto/ΔAvrPtoB (Lin and Martin, 2005) were employed in this study. Four different previously described avirulent derivatives of PsmES4326 and PstDC3000 carrying AvrRpt2 or AvrRpm1 were used (Mudgett and Staskawicz, 1999; Guttman and Greenberg, 2001).
Pathogen infection
Pseudomonas strains were cultured on King’s B (KB) media (10 g proteose peptone, 1.5 g K2HPO4, 15 g glycerol, and 5 mM MgSO4 per liter) supplemented with an appropriate antibiotic(s) at 28°C. Freshly cultured Pseudomonas strains were diluted to an optical density at 600 nm (OD600) = 0.0001, 0.001 or 0.1 in 10 mM MgSO4. Bacterial suspensions diluted to OD600 = 0.001 or 0.0001 were infiltrated into the leaves of WT and mutants with a needleless syringe. Infected plants were covered with a transparent plastic dome to keep high relative humidity for successful disease development. For spray-inoculation, 0.05% Silwet® L-77 was added into the bacterial suspension (OD600 = 0.1) before application.
The number of bacteria was determined using serial-dilutions by plating on KB media on day 3 (syringe-inoculation) or 5 (spray-inoculation) after infection. To monitor visible HR formation in the WT and mutants induced by either PsmES4326/AvrRpt2 or PsmES4326/AvrRpm1, 3 leaves per plant were infected by a high dose of these avirulent strains (OD600 = 0.01) and the number of leaves showing visible or no HR was scored 10 h (for RPM1-AvrRpm1 interaction) and 20 h (for RPS2-AvrRpt2 interaction) after inoculation.
Free SA measurement and electrolyte leakage analysis
Free SA levels were quantified as described with minor modifications (Seskar et al., 1998; Jung et al., 2009). An avirulent derivative of Pseudomonas, PsmES4326/AvrRpt2 (OD600 = 0.01), was infiltrated into the leaves of Arabidopsis plants. The infected leaves were harvested at given time points to check the level of free SA induced by pathogen infection. o-anisic acid (W394300, Sigma-Aldrich) was spiked into the samples before extraction to calibrate the recovery rate after extraction and HPLC analysis. Extracts were dried, dissolved in 55% methanol and applied in 10 μl aliquots to an HPLC coupled with a fluorescence detector (Agilent 1100). The experiments were repeated twice with similar results.
The degree of electrolyte leakage was measured in the leaves of WT and mutants infected with either PsmES4326/AvrRpt2 or PsmES4326/AvrRpm1 (OD600 = 0.05) until 25 h after inoculation. Ten leaf discs per biological replicate (4 replicates) were taken from the infected leaves, rinsed with distilled water for 30 min, and submerged in distilled water (Sohn et al., 2012). The conductivity was measured with a LAQUAtwin compact water quality metre (Horiba Scientific).
Real-time quantitative reverse transcription PCR analysis
Bacterial strains employed in this study were infiltrated into the leaves of Arabidopsis (OD600 = 0.01). Plants were kept in a walk-in growth chamber without a plastic dome during infection. Total RNAs from the infected Arabidopsis leaves were isolated with the TRIzol™ reagent according to the manufacturer’s instruction (Thermo Fisher Scientific). In order to minimize genomic DNA contamination in total RNAs, TURBO™ DNase (Thermo Fisher Scientific) was added to the total RNAs at 37°C for 30 min. About 1 μg of total RNA was used to synthesize first-strand cDNAs with Superscript™ II reverse transcriptase (Thermo Fisher Scientific). qRT-PCR was performed with the SYBR™ Green PCR mixture (Takara Bio) using the cycling program as follows: 95°C for 5 min followed by 40 cycles at 95°C for 15 s, 60°C for 15 s, and 72°C for 35 s (CFX384™/C1000™, Bio-Rad). ACTIN2 (At3g18780) was used as an internal control. Oligonucleotide sequences used in this study were listed in Table 1. Relative expression levels were calculated by a relative standard curve method (Larionov et al., 2005). The mRNA analyses were repeated at least three times with similar expression patterns.
Western blot analysis
Leaves of Arabidopsis plants inoculated with 10 mM MgSO4, PstDC3000, PstDC3000 ΔAvrPto/ΔAvrPtoB, and PsmES4326/AvrRpt2 (OD600 = 0.01) were collected at indicated time points. The finely ground leaf powder was homogenized in an equal volume of protein extraction buffer (20 mM Tris-HCl [pH 7.5], 150 mM NaCl, 1 mM EDTA, 1% Triton X-100, 0.1% sodium dodecyl sulfate [SDS], 5 mM dithiothreitol, and 1 X proteinase inhibitor [Thermo Fisher Scientific]) and centrifuged at 16,200 x g for 30 min in order to eliminate certain debris (Lee et al., 2007). SDS-polyacrylamide gel electrophoresis (SDS-PAGE) using Tris-glycine electrophoresis buffer was carried out as described by Sambrook et al. (1989). Target proteins and concentration of primary antibodies were used in this study as follows: Phospho-P44/P42 MAPK (Erk1/2) (#4370L, Cell Signaling Technology), 1:1000; MPK3 (M8318, Sigma-Aldrich), 1:800; MPK6 (A7104, Sigma-Aldrich), 1:5000; and BAK1 (AS12 1858, Agrisera), 1:5000. A goat anti-rabbit immunoglobulin G (H+L) conjugated with horseradish peroxidase (Thermo Fisher Scientific) was employed as secondary antibodies (1:5000). The signal was visualized with SuperSignal™ Chemiluminescent Substrate (Thermo Fisher Scientific). All immunoblot analyses were repeated at least twice, and the relative band intensities against a Coomassie Brilliant Blue (CBB) staining were quantified using the Image J program.
Petiole exudate analysis
WT and mutant plants were grown on Pro-Mix BX (Premier Tech) soil for 24 ~ 28 days under 21 ± 1°C and 12 h day/12 h night conditions (Greenberg et al., 2000). Leaves of A. thaliana were inoculated with either 10 mM MgSO4 or PsmES4326/AvrRpt2 (OD600 = 0.01) and detached 12 to 15 h after inoculation. To collect exudates, leaves were surface sterilized with 70% ethanol, and petioles were submerged into 1 mM EDTA supplemented with 50 μg/ml carbenicillin and streptomycin (Duchefa) at room temperature under continuous light as described (Jung et al., 2009). Exudates were diluted five times in 10 mM MgSO4 and stored at -80°C. For treatments with mixed exudates, we added an equal volume of 10 mM MgSO4 or sid2-exudates into WT-exudates or ald1-exudates. To test whether or not petiole exudates could confer resistance to bacterial infection in Arabidopsis, diluted petiole exudates were syringe-infused into recipient WT leaves 2 days prior to challenge-inoculation with virulent PsmES4326 (OD600 = 0.0001) (Jung et al., 2009). Bacterial growth was enumerated on day 3 after inoculation.
Results
Loss of ALD1 does not make npr1 and sid2 mutants more susceptible to infection with virulent Pseudomonas
Mutation of NPR1, but not ICS1/SID2, was reported to adversely affect dual phosphorylation of MPK3 and MPK6 in Arabidopsis after infection with P. syringae or flg22 treatment (Tsuda et al., 2013; Yi et al., 2015). To test if ALD1 cooperates with NPR1 and ICS1/SID2 for establishing early basal defense, we first monitored levels of MPK3 and MPK6 and the amount of activated MPK3 and MPK6 (phosphorylated forms) in leaves of WT, ald1, npr1, sid2, ald1npr1, and ald1sid2 plants before and after infection with a virulent or an attenuated strain of P. syringae. We used the attenuated strain PstDC3000 ΔAvrPto/ΔAvrPtoB, because the AvrPto and AvrPtoB secreted virulence effectors inhibit early basal defense responses initiated by MAMP-perception as well as the level of PRR complexes in Arabidopsis (Block and Alfano, 2011). Untreated plants showed no changes in the total MPK3 and MPK6 levels (upper panel in Figure 1A), indicating that basal accumulation of MPK3 and MPK6 occurs independently of ALD1, NPR1, and SID2. Phosphorylated MPK3 and MPK6 levels were very low in untreated WT and all of the mutants (upper panel in Figure 1A). Infection with PstDC3000 or PstDC3000 ΔAvrPto/ΔAvrPtoB led to robust phosphorylation of MPK3 and MPK6 in WT 30 min after infection (lower panel in Figure 1A). The extent of phosphorylated MPK3 and MPK6 was reduced in the ald1 mutant as compared with that in the WT. In npr1, sid2, ald1npr1, and ald1sid2 plants, no phosphorylated MPK3 or MPK6 was detected (lower panel in Figure 1A). Mock inoculation with 10 mM MgSO4 only slightly activated MPK3 and MPK6 in WT 30 min after infiltration. These results suggest that ALD1 is only partially needed, whereas NPR1 and SID2 are strictly required for the phosphorylated-forms to accumulate at 30 min after PstDC3000 ΔAvrPto/ΔAvrPtoB or PstDC3000 inoculation. ALD1 may influence the transient phosphorylation of MPK3 and MPK6 in NPR1- and ICS1-dependent manners. The levels of phosphorylated MPK3 and MPK6 observed in npr1 and sid2 mutants in this study differ from those previously reported (Tsuda et al., 2013; Yi et al., 2015). This might be due to the non-sterile growth conditions used here and differences in the time points used among experiments done in different labs.
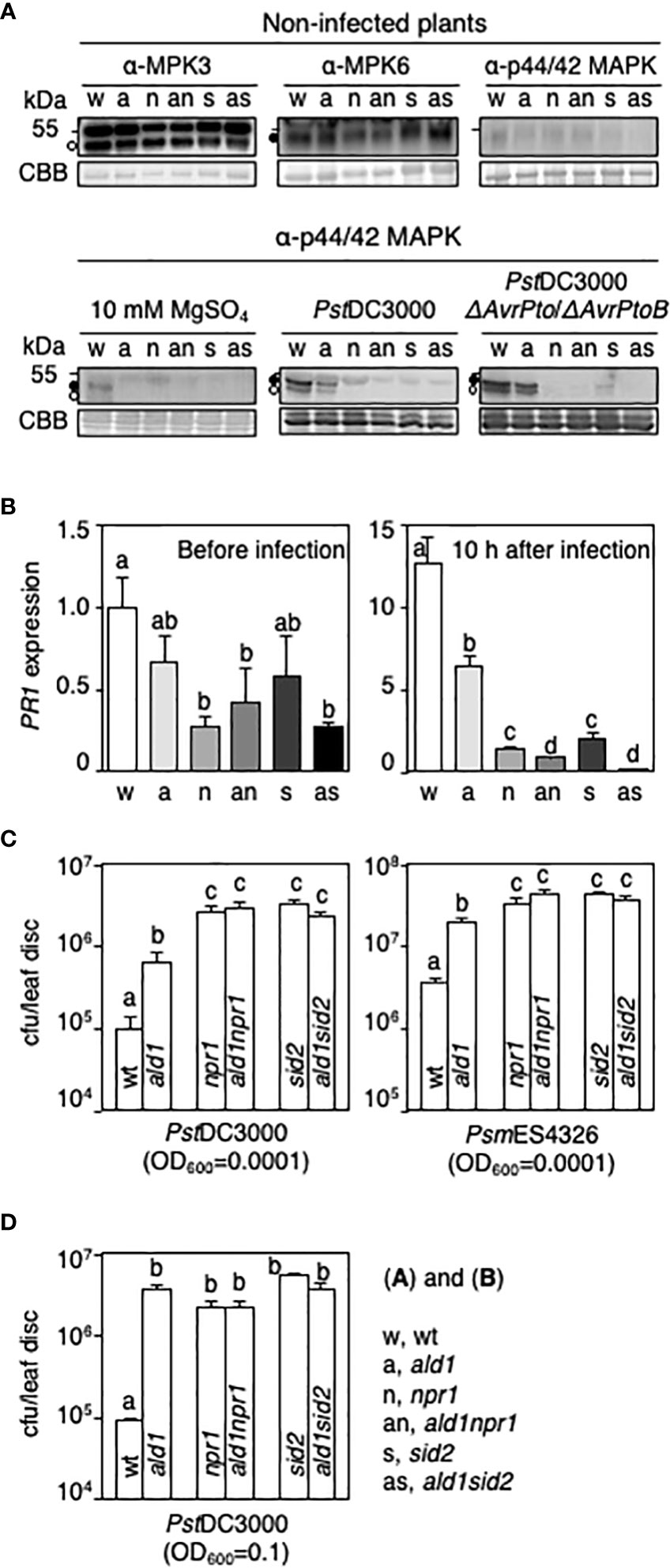
Figure 1 Loss of ALD1 in npr1 or sid2 mutants does not alter the susceptibility of Arabidopsis against infection by virulent strains of Pseudomonas. (A) Levels of MPK3, MPK6, and phosphorylated MPK3 and MPK6 in leaves of WT (w), ald1 (a), npr1 (n), ald1npr1 (an), sid2 (s), and ald1sid2 (as) without (upper panel) and with pathogen infection (lower panel). Open circles and filled circles point out MPK3 and MPK6, respectively. Membranes were stained with Coomassie Brilliant Blue R-250 (CBB) to verify equal loading. Infected leaves with 10 mM MgSO4, PstDC3000, or PstDC3000 AvrPto/AvrPtoB (OD600 = 0.01) were taken 30 min after infiltration. (B) Relative PR1 mRNA level in leaves of WT and mutants during PsmES4326 infection (OD600 = 0.01). mRNA level in the non-infected WT plant (PR1/ACTIN2) was set to 1, and then mRNA levels of each condition were normalized by the value of the non-infected WT plant. Error bars represent the standard deviation from three biological replicates (number of technical replicates = 3). (C) Growth of virulent strains of Pseudomonas in Arabidopsis plants 3 days after syringe-inoculation. Leaves of 24- to 28-day-old plants were inoculated with either a virulent strain of PstDC3000 (left panel) or PsmES4326 (right panel) (OD600 = 0.0001) by using a needleless syringe. (D) Growth of PstDC3000 in leaves of Arabidopsis plants 5 days after spray-inoculation (OD600 = 0.1). Error bars indicate standard error (n = 8 for syringe-inoculation; n = 12 for spray-inoculation) (C, D). The experiments were repeated more than three times with the same results. Different letters above bars indicate statistically significant differences (p < 0.05, one-way ANOVA) (B-D).
To test whether SA-responsive defenses are reduced in the double mutants compared to each single mutant, we examined the expression of PATHOGENESIS-RELATED PROTEIN1 (PR1) in leaves of Arabidopsis with or without pathogen infection. Under uninfected conditions, the level of PR1 transcript was diminished in npr1, ald1npr1, and ald1sid2, as compared with those in WT (left panel in Figure 1B). Mutation of NPR1 or ICS1/SID2 strongly decreased PR1 transcription more than that of ALD1 during PsmES4326 infection (OD600 = 0.01) (right panel in Figure 1B). Interestingly, PR1 levels in ald1npr1 and ald1sid2 were lower than those in npr1 and sid2 mutants 10 h after inoculation (right panel in Figure 1B). Considering the additive effect between ALD1 and NPR1 or ICS1/SID2, the results strongly support that ALD1 can impact PR1 expression during infection in a SA- and NPR1-independent manner as described previously using a constitutive defense mutant (Song et al., 2004b; Ng et al., 2011).
To analyze if the changes in MPK3 and MPK6 activation and PR1 transcripts by the simultaneous mutation of ALD1, NPR1 and/or ICS1 are related to resistance against pathogens, we syringe-inoculated Arabidopsis plants with virulent strains of Pseudomonas and measured bacterial growth in infected leaves 3 days post inoculation. The ald1 mutant was less susceptible than npr1 and sid2 mutants to PstDC3000 and PsmES4326 (Figure 1C). The mutation of ALD1 in the npr1 or sid2 background did not affect the susceptibility to either strain (Figure 1C). These results suggest that the residual resistance in ald1 is due to the SA pathway regulated by NPR1 and ICS1/SID2, at least when bacteria is infiltrated.
Epidermal cells provide a substantial barrier to infection, possibly due to the cuticle and the dynamics of natural openings such as stomata. Moreover, ALD1 restricted to the epidermal plastids rescues several immune responses in Arabidopsis (Jiang et al., 2021). One way to assess the role of epidermal defenses is to spray-inoculate bacteria onto plant leaves and measure bacterial growth (Melotto et al., 2006; Panchal et al., 2016). Therefore, we spray-inoculated WT and our suite of Arabidopsis mutants with PstDC3000. The ald1 plants showed more growth of bacteria relative to that seen in WT. Additionally, ald1 susceptibility was indistinguishable from that of npr1, sid2, ald1npr1, and ald1sid2 (Figure 1D). Thus, ALD1 and the SA biosynthesis/action components (ICS1/SID2 and NPR1) are needed together to suppress strong colonization after spray-inoculation, as suggested by a previous study (Jiang et al., 2021). This may be due to ALD1’s role in amplifying SA accumulation.
Loss of ALD1 in npr1 and sid2 mutants does not additively increase susceptibility to infection with attenuated Pseudomonas
The reduced dual phosphorylation of MPK3 and MPK6 in single and double mutants 30 min after inoculation (Figure 1A) prompted us to test whether ALD1, NPR1, and ICS1/SID2 control basal defense responses to PstDC3000 ΔAvrPto/ΔAvrPtoB infection. We analyzed transcript levels of FRK1, NHL10, and PR1 in WT and mutants during infection with the attenuated Pseudomonas strain (Lin and Martin, 2005; Rosebrock et al., 2007; Boudsocq et al., 2010; Mersmann et al., 2010). Although differences in FRK1 transcript levels were not found among WT and mutant plants 10 h after PstDC3000 ΔAvrPto/ΔAvrPtoB inoculation, the level of FRK1 mRNA significantly decreased in the mutants, especially npr1, sid2, ald1npr1, and ald1sid2, 15 h after inoculation (left panel in Figure 2A). Transcript levels of NHL10, another MAMP-responsive gene, decreased in all mutants tested in this study after PstDC3000 ΔAvrPto/ΔAvrPtoB infection compared with that in WT. NHL10 mRNA expression was also significantly reduced in double mutants compared with those in each single mutant 10 h after inoculation (right panel in Figure 2A). In the case of PR1, there was no induction of the gene in any of the mutants at 10 h and only the ald1mutant displayed an increase (although less than in WT) at 15 h after PstDC3000 ΔAvrPto/ΔAvrPtoB infection (Figure 2B). Expression levels of PR1 in the ald1npr1 and ald1sid2 mutants did not differ from those in npr1 and sid2 mutants after PstDC3000 ΔAvrPto/ΔAvrPtoB infection (Figure 2B). This supports the view that NPR1 and ICS1/SID2 are the leading players in regulating PR1 expression in Arabidopsis (Cao et al., 1997; Wildermuth et al., 2001). Taken together, these mRNA analyses reveal that NPR1 and ICS1/SID2 have a stronger influence on FRK1, NHL10, and PR1 mRNA expression than ALD1 in PstDC3000 ΔAvrPto/ΔAvrPtoB-infected leaves.
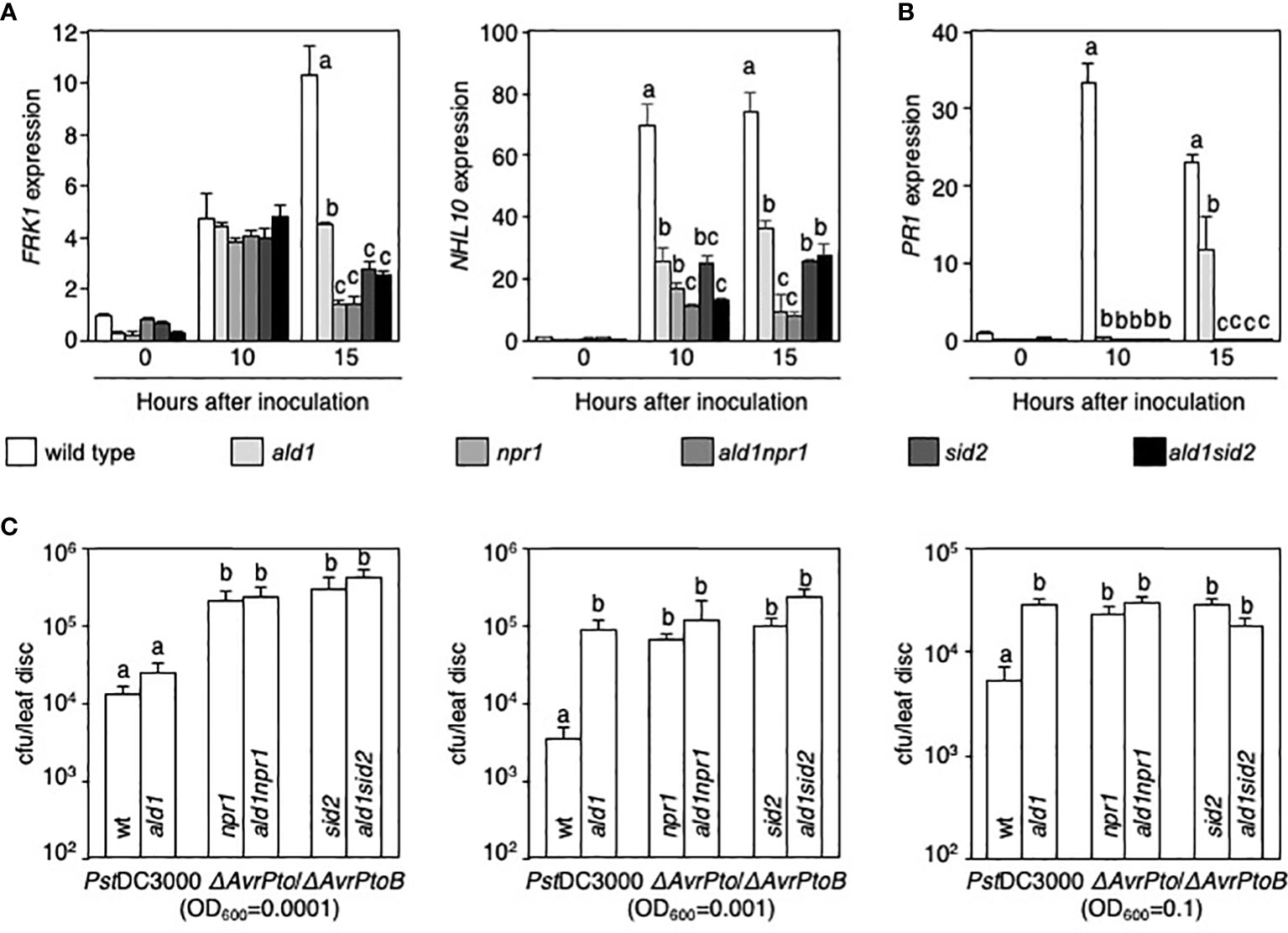
Figure 2 Mutations of ALD1, NPR1, and ICS1/SID2 cause hypo-resistance against an attenuated strain of Pseudomonas syringae infection. (A, B) Relative expression levels of FRK1 and NHL10 (A) and PR1 (B) in leaves of WT and mutants infected with PstDC3000 ΔAvrPto/ΔAvrPtoB (OD600 = 0.01). The columns and bars show the average with standard deviation obtained from three biological replications (number of technical replicates = 3). (C) Growth of PstDC3000 ΔAvrPto/ΔAvrPtoB strain in WT and mutants. An attenuated strain was syringe-infiltrated into the leaves of WT, single, and double mutants (OD600 = 0.0001 or 0.001) (left and middle panels), or the strain was also applied to Arabidopsis plants with a spray (OD600 = 0.1) (right panel). The number of bacteria was counted on day 3 or 5 after inoculation. Error bars indicate standard error (n = 10 or 12). The experiment was repeated twice with the same results. Statistically significant differences are shown using different letters (p < 0.05 [A, B], p < 0.01 [C], one-way ANOVA).
We analyzed the growth of the attenuated strain in WT and mutant plants. The bacterial titer was slightly higher in the ald1 mutant than in WT when PstDC3000 ΔAvrPto/ΔAvrPtoB (OD600 = 0.0001) were infiltrated into Arabidopsis leaves; however, the difference was not statistically significant (left panel in Figure 2C). Interestingly, relative to WT, ald1 showed increased susceptibility to PstDC3000 ΔAvrPto/ΔAvrPtoB infection with a higher concentration (OD600 = 0.001) (middle panel in Figure 2C) as well as after spray-inoculation of the strain (OD600 = 0.1) (right panel in Figure 2C). Populations of the attenuated strain were higher in npr1 and sid2 mutants than in the WT plant, and a further mutation of ALD1 in npr1 and sid2 mutants did not render plants more susceptible to PstDC3000 ΔAvrPto/ΔAvrPtoB infection (Figure 2C). This finding, together with the intermediate effect of mutation of ALD1 on MPK3/MPK6 activation and defense genes expression supports the view that ALD1 amplifies the ICS1/SID2 and NPR1 module in this infection condition.
Both ald1npr1 and ald1sid2 mutants are more susceptible than each single mutant to Pseudomonas carrying AvrRpt2, but not AvrRpm1
To examine if disrupting ALD1 and NPR1 or ICS1/SID2 might have an additive effect on ETI in Arabidopsis, we inoculated avirulent derivatives of PsmES4326 and PstDC3000 carrying AvrRpt2 or AvrRpm1 into leaves of WT and mutant plants. Although visible HR was not affected by any mutations tested in this study after avirulent Pseudomonas infection (OD600 = 0.01), the rate of electrolyte leakage was slightly delayed by these mutations (Figures 3A, B). Previously, it was reported that the npr1 mutant showed stronger HR than the WT after infection with P. syringae carrying AvrRpm1 (OD600 = 0.03 or 0.1), but exhibited a WT-like response to P. syringae carrying AvrRpt2 (OD600 = 0.03) (Rate and Greenberg, 2001). These current and previous studies suggest that the cell death response of npr1 partly relies on the concentration of inoculum and the genetic composition of P. syringae.
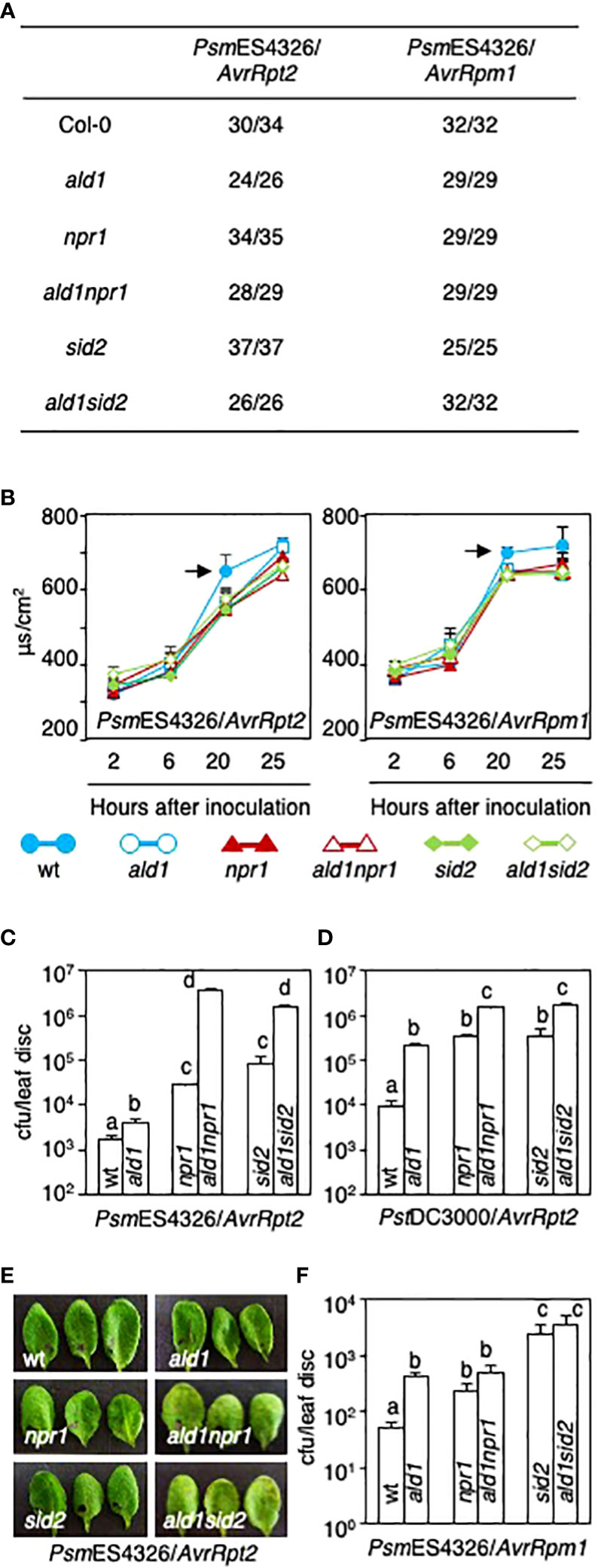
Figure 3 Both ald1npr1 and ald1sid2 mutants show susceptibility in response to avirulent derivatives of Pseudomonas carrying AvrRpt2, but not AvrRpm1. (A) The number of leaves exhibiting HR out of the number of leaves infected with two different avirulent derivatives, PsmES4326/AvrRpt2 or PsmES4326/AvrRpm1 (OD600 = 0.01). (B) Electrolyte leakage from leaves of Arabidopsis after infection with PsmES4326/AvrRpt2 (left panel) or PsmES4326/AvrRpm1 (right panel) (OD600 = 0.05). Data represent the averages with standard deviations (n = 4). Arrows indicate statistically significant differences between WT and mutants (p < 0.01, two-tailed Student’s t-test). The experiments were repeated twice with the same results. (C, D) Pathogen growth in leaves of ald1npr1 and ald1sid2 mutants after PsmES4326/AvrRpt2 infection (C) and PstDC3000/AvrRpt2 infection (OD600 = 0.0001) (D). (E) Symptom development in the leaves of ald1npr1 and ald1sid2 mutants after PsmES4326/AvrRpt2 infection (OD600 = 0.0001). The photos were taken on day 3 after inoculation. (F) Growth of an avirulent derivative, PsmES4326/AvrRpm1, in leaves of Arabidopsis 3 days after inoculation (OD600 = 0.0001). Data (C, D, F) represent the averages with standard errors (n = 8, or 12). Different letters indicate statistically significant differences (p < 0.01, one-way ANOVA). The experiments were repeated more than three times with the same results.
Interestingly, the growth of PsmES4326/AvrRpt2 and PstDC3000/AvrRpt2 dramatically increased more than 10-fold in the ald1npr1 and ald1sid2 mutants compared with those in each single mutant (Figures 3C, D). Additionally, the double mutants exhibited visible disease symptoms after PsmES4326/AvrRpt2 infection, while no evident disease lesions in WT or single mutants were observed (Figure 3E). In contrast to the effects of strains carrying AvrRpt2, growth of PsmES4326/AvrRpm1 did not increase in the ald1npr1 and ald1sid2 mutants compared to that in each single mutant (Figure 3F). These results show that effective signaling via RPS2, the AvrRpt2 receptor, requires more than just an amplifying effect of ALD1 on SA accumulation previously described (Song et al., 2004b). Thus, we suggest that ALD1 provides additional SA-independent defenses for suppressing the growth of strains that carry AvrRpt2.
ALD1 and NPR1 have an additive effect on SA and PR1 accumulation against an avirulent derivative of Pseudomonas carrying AvrRpt2
To understand the roles of ALD1, NPR1, and ICS1/SID2 in the immune response during infection with PsmES4326/AvrRpt2 in detail, we examined the strength of SA-related defenses in WT and mutants after infection. Free SA and PR1 mRNA levels in the ald1 mutant were lower than those in WT plant after PsmES4326/AvrRpt2 infection (Figures 4A, B), as previously reported (Song et al., 2004b; Cecchini et al., 2015). NPR1 mutation resulted in reduced SA accumulation, and PR1 transcription after PsmES4326/AvrRpt2 infection at 10 h and 20 h after inoculation compared to WT plants (Figures 4A, B). We note that the lower SA level in npr1 versus WT differed from previous studies (using strain PstDC3000/AvrRpt2) in which the opposite findings were reported (Delaney et al., 1995; Zhang et al., 2010; Ding et al., 2015). These studies used longer infection times and in one case a lower infection dose for the experiments, parameters that affect free SA accumulation. The SA and PR1 mRNA levels significantly decreased in the infected leaves of the ald1npr1 mutant after PsmES4326/AvrRpt2 infection compared with those in ald1 and npr1 mutants (Figures 4A, B). On the other hand, both SA and PR1 mRNA did not accumulate to significant levels in sid2 and ald1sid2 mutants after infection (Figures 4A, B), which is consistent with ICS1/SID2 being directly involved in SA biosynthesis (Wildermuth et al., 2001; Garcion et al., 2008). These results suggest that ALD1 acts additively with NPR1 to execute requisite defenses in RPS2-mediated ETI.
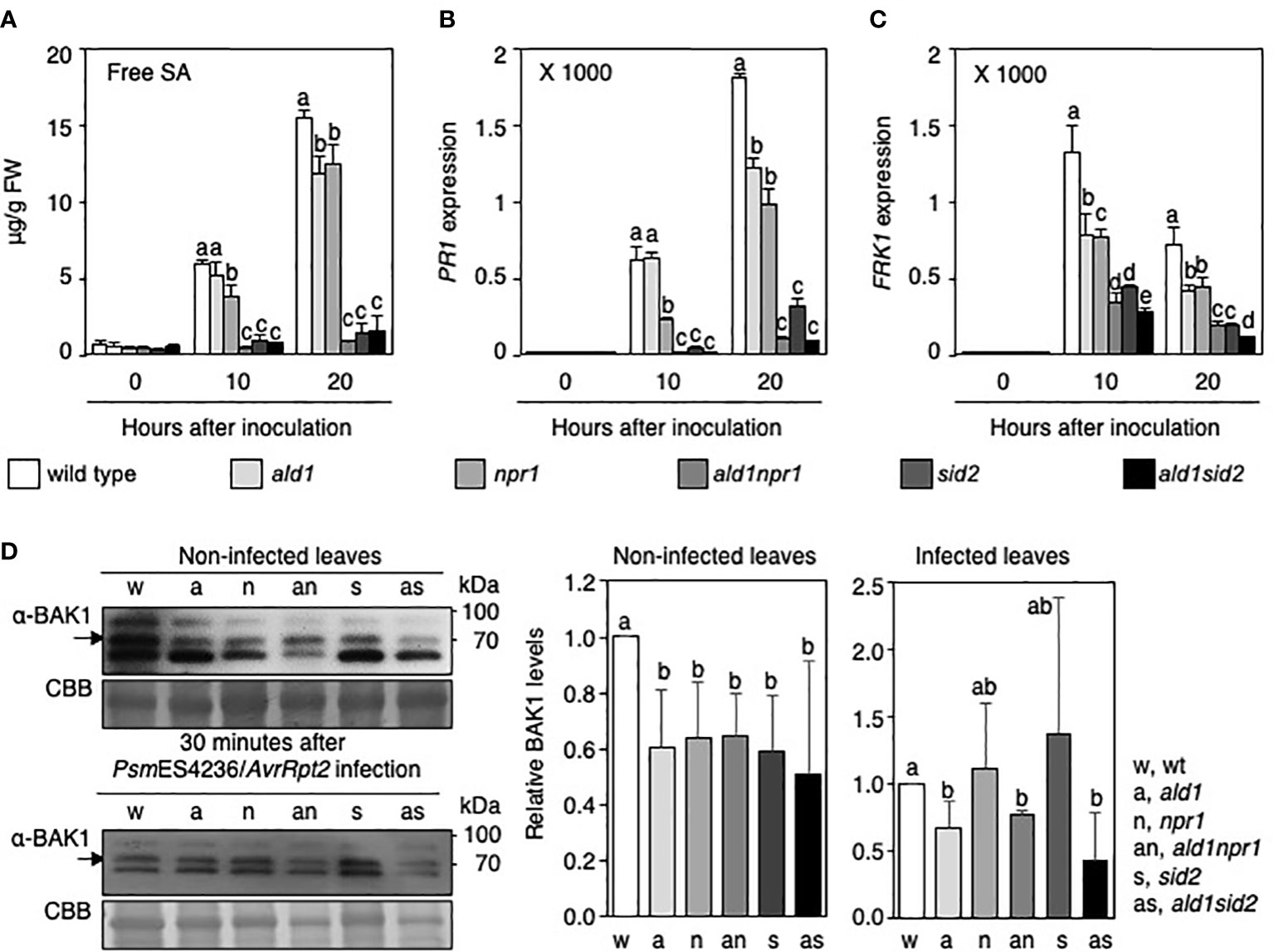
Figure 4 ALD1, NPR1, and SID2 are necessary to fully express RPS2-mediated immunity against an avirulent Pseudomonas infection. (A) Free SA levels in the leaves of WT, ald1, npr1, ald1npr1, sid2, and ald1sid2 plants during avirulent PsmES4326/AvrRpt2 infection. Data show averages with standard deviations from five biological replications (n = 5). (B) Deregulation of PR1 transcription in ald1npr1 and ald1sid2 mutants after infection. PR1 expression levels relative to an ACTIN2 gene in non-inoculated leaves of WT plant was set to 1. The columns and bars show the average with standard deviation obtained from three biological replications (number of technical replicates = 3). (C) Relative level of FRK1 mRNA in leaves of Arabidopsis plants infected with PsmES4326/AvrRpt2. We normalized expression levels and presented the results mentioned above (B). (D) Level of BAK1 in leaves of WT and mutants in the absence or presence of avirulent pathogen infection. Upper, BAK1 protein levels in leaves of non-infected Arabidopsis plants; lower, BAK1 levels after PsmES4326/AvrRpt2 infection. Bands not indicated are non-specific. Graphs on the right side of D show the normalized BAK1 levels in Arabidopsis at the indicated time points (average ± standard deviation, n = 3, or 4). Different letters indicate statistically significant differences (p < 0.05 [D], p < 0.01 [A-C], one-way ANOVA).
Accumulation of BAK1 and FRK1 expression requires ALD1, NPR1, and ICS1/SID2 in avirulent PsmES4326/AvrRpt2-infected Arabidopsis
An interconnection between MAPK signaling and SA-related defense was well-described in Arabidopsis infected with various derivatives of P. syringae (Tsuda et al., 2013), and ALD1, NPR1 and ICS1 are also known to participate in basal defense responses (Laluk et al., 2011; Yi et al., 2014; 2015; Cecchini et al., 2015). Based on this knowledge, it seemed possible that additional mutation of ALD1 in npr1 or sid2 mutant background could weaken early defense against PsmES4326/AvrRpt2 infection. To address this, we examined representative basal defense responses in leaves of WT, single, and double mutants infected with PsmES4326/AvrRpt2. FRK1 mRNA levels were severely reduced in ald1npr1 and ald1sid2 at 10 and 20 h after PsmES4326/AvrRpt2 infection compared with ald1, npr1 and sid2, respectively (Figure 4C). Compared with FRK1 transcript levels in WT and mutants under infection of an attenuated P. syringae strain (scale of Y-axis was up to 12) (Figure 2A), infection of avirulent strain PsmES4326/AvrRpt2 could actively trigger the transcription of FRK1 (scale of Y-axis was up to 2,000). The results support prior observations that the expression of MAMP-responsive defense responses is intensified during ETI in Arabidopsis (Tsuda et al., 2013; Ngou et al., 2021; Yuan et al., 2021). Furthermore, BRI1-ASSOCIATED RECEPTOR KINASE1 (BAK1) protein levels decreased in the mutants compared with that in WT plant in the absence of Pseudomonas infection (Figure 4D). After infection, however, BAK1 proteins accumulated to the WT level in npr1 and sid2 mutants, and BAK1 levels in the double mutants could not be distinguished from npr1 and sid2 mutants (Figure 4D). Only the ALD1 mutation adversely affected the level of BAK1 protein during PsmES4326/AvrRpt2 infection (Figure 4D). This indicates that ALD1 is involved in controlling MAMP-receptor levels before infection as described previously (Cecchini et al., 2015) as well as during infection. Taken together, these results suggest that the reduction of SA-related defense and early defense responses might be the cause of the high susceptibility to Pseudomonas carrying AvrRpt2 of the ald1sid2 and ald1npr1 double mutants.
Both NPR1 and ICS1/SID2 are required for producing active petiole exudates capable of inhibiting bacterial growth in Arabidopsis
PTI and ETI that occur in local tissues can initiate SAR by inducing the accumulation of mobile signals. Application of PsmES4326/AvrRpt2-induced petiole exudate (PEX) from WT and exudate from ALD1-overexpressing plants to recipient WT plants confers disease resistance against virulent P. syringae infection (Jung et al., 2009; Cecchini et al., 2015). Although it is well-known that npr1 and sid2 mutants have a defect in systemic resistance, the role of NPR1 and ICS1/SID2 in the biosynthesis of mobile SAR signal compounds in Arabidopsis is still not known. Therefore, we introduced PEXs collected from WT, ald1, npr1, and sid2 infected with PsmES4326/AvrRpt2 into leaves of recipient WT plants 2 days prior to challenge-inoculation with a virulent P. syringae strain (PsmES4326). PEXs from WT and ald1 mutant successfully inhibit PsmES4326 growth in the leaves of recipient WT plants (Figure 5A). However, neither mock-induced exudates (MEX) nor PEXs from npr1 and sid2 mutants could reduce bacterial growth in the recipient WT leaves after subsequent PsmES4326 infection (Figure 5A). ald1npr1 and ald1sid2 mutants displaying increased susceptibility to Pseudomonas carrying AvrRpt2 also failed to produce active PEXs (Figure 5B). These results suggest that NPR1 and ICS1/SID2 are required for producing active PEX in Arabidopsis. We note that SA production/transduction in Arabidopsis could be needed to enable tissue to produce enough mobile signals to confer disease resistance or SA might be needed as a component of the exudate, even if it is not the only mobile signal in the exudate. Our results also show a requirement for the basal level of SA before pathogen infection and accumulation of SA in local infected leaves.
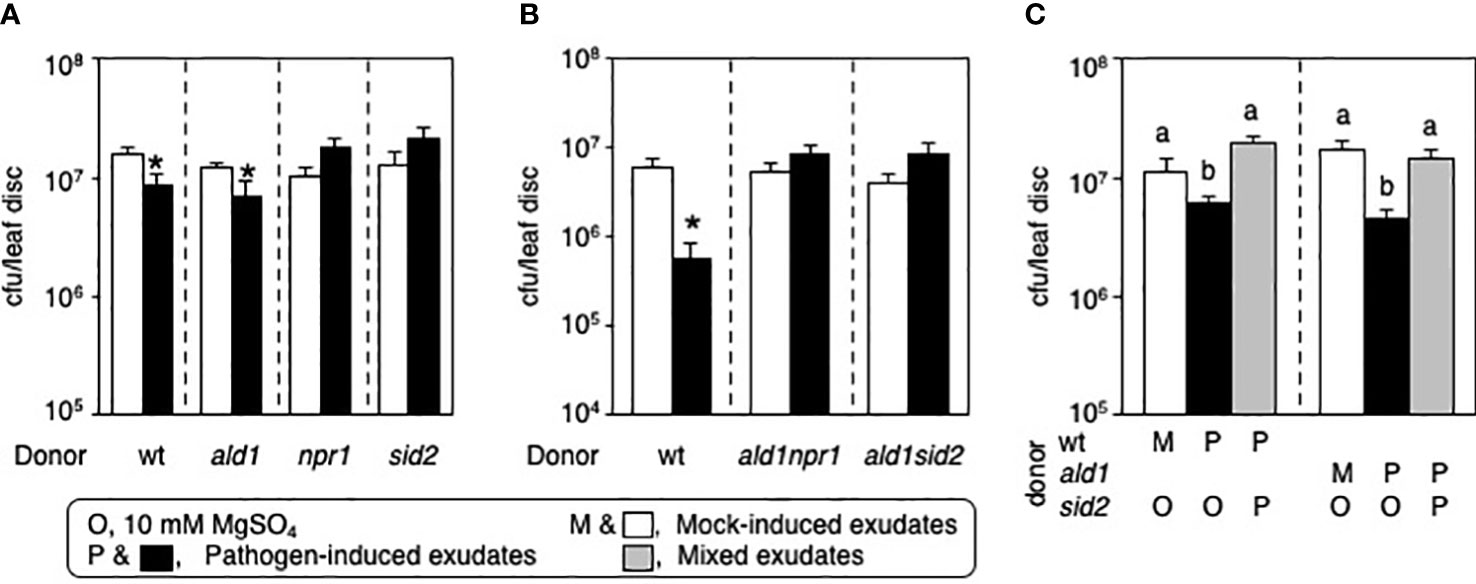
Figure 5 NPR1 and SID2 are needed to make active exudates capable of inducing disease resistance in plants. (A) Bacterial growth in the recipient WT plants pre-immunized with mock-induced exudates (MEX, white boxes) and pathogen-induced exudates (PEX, black boxes) from WT, ald1, npr1, and sid2 plants. (B) Biological activity of PEXs collected from ald1npr1 and ald1sid2 double mutants. (C) Inhibitory effect of PEX from sid2 mutant on active PEXs collected from WT and ald1 infected with SAR-inducing PsmES4326/AvrRpt2. Mixed exudates were pre-applied into the leaves of recipient WT plants prior to challenge-inoculation. m, mock-induced exudates; p, pathogen-induced exudates. Data represent the averages with standard errors. The asterisks (p < 0.05, two-tailed Student’s t-test, n = 8) and different letters (p < 0.05, one-way ANOVA, n = 8) indicate statistically significant differences. The experiments were repeated at least twice with similar results.
In a different scenario, it could also be possible that an inactive PEX can inhibit the activity of an active PEXs. To examine if inactive sid2-PEX can suppress active PEX’s activity, we produced the mixed PEXs collected from different plants and introduced them into recipient WT plants before subsequent pathogen inoculation. Remarkably, the WT-PEX and ald1-PEX lost their immune-inducing activity in the presence of PEX from the sid2 mutant (Figure 5C). This result indicates that an unidentified metabolite(s) increased in the infected sid2 plant, or a Pseudomonas-derived compound(s) that accumulates more in the sid2 mutant, may override the positive effects of WT-PEX and ald1-PEX.
Discussion
The activities of ALD1, NPR1 and ICS1/SID2 can influence each other at the transcriptional level (Song et al., 2004b; Cecchini et al., 2015). This study showed that their functional relationship with respect to signaling output and pathogen suppression is complex. The loss of ALD1-dependent signaling does not make npr1 and sid2 mutants more susceptible to infection with an attenuated strain, virulent strains, and an avirulent strain of P. syringae carrying AvrRpm1. However, to fully activate the immune response to an avirulent P. syringae strain carrying AvrRpt2 requires ALD1 in addition to NPR1 and ICS1/SID2. This highlights the differences in signaling requirements even with closely related P. syringae strains. Our working model is that under many conditions, the major role of ALD1 is to amplify the accumulation of SA. However, under very defense-demanding conditions ALD1 provides an additional defense function to provide robust disease resistance.
In most infections that we studied, each defense module relying on ALD1, NPR1 or ICS1/SID2 can be sufficient to inhibit attenuated or virulent P. syringae growth; an additive effect between ALD1 and NPR1 or ICS1/SID2 is not detectable during these Arabidopsis-P. syringae interactions. Interestingly, the ald1 mutant shows disease susceptibility that differs from WT in a manner that depends on the infection dose. When the dose is low, ald1 shows a WT-like disease response to the attenuated strain, but higher doses elicit stronger phenotypes. A plausible explanation is that basal immunity, although reduced, is strong enough to confer disease resistance against the low dose of attenuated P. syringae. Possibly different proteins, such as EDS1 and PAD4, that act upstream of ICS1/SID2 and NPR1, are still active in the ald1 mutant (Falk et al., 1999; Jirage et al., 1999) and consequently they can regulate the functions of ICS1/SID2 and NPR1. An alternative scenario is that ALD1 may be necessary for executing a vigorous immune response when Arabidopsis plants are exposed to a high inoculum of virulent pathogen. For another clue, the ald1sid2 double mutant was less resistant than each single mutant when plants were inoculated with a higher concentration of a virulent Pseudomonas strain (OD600 = 0.001) (Bernsdorff et al., 2016) than was used in this study (OD600 = 0.0001). The previous report supports the possibility that the strength of the immune response correlates with the intensity of pathogen threat and is determined by the relationship between individual defense modules controlled by key defense proteins. This scenario can also support ALD1’s role as an amplifier of defenses (Song et al., 2004b).
ALD1 is more important in the plant defense response during pathogenesis after spray-inoculation (i.e. indistinguishable from that of npr1, sid2, ald1npr1, and ald1sid2) than after syringe-inoculation. This is consistent with ALD1 playing a key role in the epidermis (Jiang et al., 2021), the cell layer that first comes in contact with sprayed bacteria. Moreover, it is known that the PTI response in plant epidermal cells is critical for sensing invading pathogens and preventing pathogens’ colonization (Melotto et al., 2006; Zeng and He, 2010; Henty-Ridilla et al., 2013; Kang et al., 2014) and that ALD1 regulates levels and responsiveness of PRRs (Cecchini et al., 2015). Thus, we suggest that ALD1 can boost PTI responses in the epidermis.
Additional mutation of ALD1 in the sid2 mutant does not affect SA biosynthesis and PR1 mRNA expression. However, transcription of FRK1 during infection significantly decreases in the ald1sid2 double mutant compared to the single mutants. These results suggest that SA- and ALD1-mediated defense molecules have an additive effect on the amplification of FRK1 expression in plants during ETI (Tsuda et al., 2013; Bigeard et al., 2015). The reduced immunity in ald1npr1 and ald1sid2 mutants after avirulent P. syringae carrying AvrRpt2 infection could be due to the weakened SA-related defense as well as basal defense response resulting from the simultaneous mutation of ALD1 and NPR1 or ICS1/SID2. Therefore, we infer that AvrRpt2-triggered immunity may depend on PTI more than AvrRpm1-dependent immunity, where there is no additive effect of such simultaneous mutations. Taken together, we propose that these three defense proteins modulate the strength of the immune response in an infection-specific manner.
Petiole exudates from infected plants contain signal molecules, some of which may be mobile and involved in systemic immunity. It may be surprising that ald1 exudates still have biological activity, given the proposed function of ALD1-dependent metabolites Pip and NHP as systemic signals (Návarová et al., 2012; Chen et al., 2018; Hartmann et al., 2018). However, previous experiments using chimeric plants that only produced ALD1 in epidermal cells of leaves used for generating SAR signals showed that exudates from such plants lacked Pip and NHP (Jiang et al., 2021). These chimeric plants still showed robust SAR. The present results showing that ald1 plants can produce active exudates supports the view that SAR can occur without appreciable accumulation of vascular-mobilized Pip or NHP. This is consistent with the idea that there may be multiple priming signals that together form a priming threshold that enables strong systemic resistance.
Unlike exudates from WT and ald1, the exudates from npr1 and sid2 mutants infected with PsmES4326/AvrRpt2 are biologically nonfunctional. We do not have any direct evidence to explain the inability of npr1- and sid2-exudates collected after infection to confer immunity, but one possibility is that the npr1 and sid2 mutants can not produce enough signal molecule(s). It is possible that, unlike in tobacco (Vernooij et al., 1994), SA-related immunity may be needed in local tissues. This could be addressed in the future using chimeric Arabidopsis plants. The Pip level in petiole exudates from a sid2 mutant was reported to be similar to that from WT plant (Návarová et al., 2012), but other proposed mobile signal molecules, such as MeSA, azelaic acid, glycerol-3-phosphate, and dehydroabietinal, have not yet been quantified in PEXs from npr1 and sid2 mutants (Jung et al., 2009; Chanda et al., 2011; Chaturvedi et al., 2012; Dempsey and Klessig, 2012; Návarová et al., 2012). Thus, we infer that all or some of these proposed signals may not accumulate in npr1- and sid2-PEX. Alternatively, it is also conceivable that both mutants impair the ability to mobilize signals out of the leaves into the exudate.
Another plausible explanation for the non-functionality of the npr1 and sid2 exudates may be related to systemic induced susceptibility (SIS, Cui et al., 2005; Haney et al., 2018). This is a phenomenon wherein plants inoculated on their lower leaves with a strain that produces the secreted toxin coronatine (a mimic of jasmonic acid; Mittal and Davis, 1995; Bender et al., 1999; Katsir et al., 2008) become more susceptible to infection of the upper leaves (Cui et al., 2005). PEXs from the hypersusceptible npr1 and sid2 mutants may contain a high level of coronatine and/or plant metabolite(s) that suppress systemic resistance in plants. This may occur because PsmES4326/AvrRpt2 is more virulent in these mutants and more secreted coronatine may end up in the npr1 and sid2 exudates. The fact that mixed exudates between active exudates from WT and ald1 and PEX from the sid2 mutant can not suppress bacterial growth in the recipient WT plants strongly supports this last explanation. We note that coronatine inhibits SA-dependent defenses by disturbing SA accumulation, resulting in local and systemic susceptibility (Geng et al., 2012; Zheng et al., 2012). Therefore, coronatine is a strong bacterial-derived candidate that accumulates in sid2-PEX. It is also possible that the jasmonic acid accumulated in the leaves of sid2 donor plants inoculated with P. syringae carrying AvrRpt2 (Liu et al., 2016) inhibits the PEX-induced resistance in recipient WT plants by blocking the activation of SA-related defenses (Koornneef and Pieterse, 2008).
In conclusion, to control bacterial growth, ALD1 acts independently or together with NPR1 or ICS1/SID2 as a defense signal amplifier, depending on the type of Arabidopsis-Pseudomonas interaction. Both defense modules are required to confer maximal immunity to infection by an avirulent pathogen that triggers resistance via perception of the AvrRpt2 bacterial effector.
Data availability statement
The original contributions presented in the study are included in the article/supplementary material. Further inquiries can be directed to the corresponding author.
Author contributions
HWJ and JG conceived and designed the experiments. HWJ, S-JY, HJC, and SWN performed the experiments. HWJ, JG, and NC analyzed and interpreted data and wrote the paper. All authors contributed to the article and approved the submitted version.
Funding
This work was supported by Basic Science Research Program through the National Research Foundation of Korea funded by the Ministry of Education (2020R1A6A1A03047729) and the Green Fusion Technology Graduate School Program of the Ministry of Environment to HJ, a grant from NSF (NSFIOS1456904) to JG, and grants from ANPCYT and CONICET (PICT-2017-0589, PICT-2020-0483, and PIP_2021-2023) to NC.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Belkhadir, Y., Nimchuk, Z., Hubert, D. A., Mackey, D., Dangl, J. L. (2004). Arabidopsis RIN4 negatively regulates disease resistance mediated by RPS2 and RPM1 downstream or independent of the NDR1 signal modulator and is not required for the virulence functions of bacterial type III effectors AvrRpt2 or AvrRpm1. Plant Cell 16 (10), 2822–2835. doi: 10.1105/tpc.104.024117
Bender, C. L., Alarcón-Chaidez, F., Gross, D. C. (1999). Pseudomonas syringae phytotoxins: mode of action, regulation, and biosynthesis by peptide and polyketide synthetases. Microbiol. Mol. Biol. Rev. 63 (2), 266–292. doi: 10.1128/MMBR.63.2.266-292.1999
Bernsdorff, F., Döring, A. C., Gruner, K., Schuck, S., Bräutigam, A., Zeier, J. (2016). Pipecolic acid orchestrates plant systemic acquired resistance and defense priming via salicylic acid-dependent and -independent pathways. Plant Cell 28 (1), 102–129. doi: 10.1105/tpc.15.00496
Bigeard, J., Colcombet, J., Hirt, H. (2015). Signaling mechanisms in pattern-triggered immunity (PTI). Mol. Plant 8 (4), 521–539. doi: 10.1016/j.molp.2014.12.022
Bisgrove, S. R., Simonich, M. T., Smith, N. M., Sattler, A., Innes, R. W. (1994). A disease resistance gene in arabidopsis with specificity for two different pathogen avirulence genes. Plant Cell 6 (7), 927–933. doi: 10.1105/tpc.6.7.927
Block, A., Alfano, J. R. (2011). Plant targets for pseudomonas syringae type III effectors: Virulence targets or guarded decoys? Curr. Opin. Microbiol. 14 (1), 39–46. doi: 10.1016/j.mib.2010.12.011
Boudsocq, M., Willmann, M. R., McCormack, M., Lee, H., Shan, L., He, P., et al. (2010). Differential innate immune signalling via Ca(2+) sensor protein kinases. Nature 464 (7287), 418–422. doi: 10.1038/nature08794
Bull, C. T., Manceau, C., Lydon, J., Kong, H., Vinatzer, B. A., Fischer-Le Saux, M. (2010). Pseudomonas cannabina pv. cannabina pv. nov., and pseudomonas cannabina pv. alisalensis (Cintas koike and bull 2000) comb. nov., are members of the emended species pseudomonas cannabina (ex sutic and dowson 1959) gardan, shafik, belouin, brosch, grimont and grimont 1999. Syst. Appl. Microbiol. 33 (3), 105–115. doi: 10.1016/j.syapm.2010.02.001
Cao, H., Glazebrook, J., Clarke, J. D., Volko, S., Dong, X. (1997). The arabidopsis NPR1 gene that controls systemic acquired resistance encodes a novel protein containing ankyrin repeats. Cell 88 (1), 57–63. doi: 10.1016/s0092-8674(00)81858-9
Cecchini, N. M., Jung, H. W., Engle, N. L., Tschaplinski, T. J., Greenberg, J. T. (2015). ALD1 regulates basal immune components and early inducible defense responses in arabidopsis. Mol. Plant-Microbe Interact. 28 (4), 455–466. doi: 10.1094/MPMI-06-14-0187-R
Cecchini, N. M., Roychoudhry, S., Speed, D. J., Steffes, K., Tambe, A., Zodrow, K., et al. (2019). Underground azelaic acid-conferred resistance to pseudomonas syringae in arabidopsis. Mol. Plant-Microbe Interact. 32 (1), 86–94. doi: 10.1094/MPMI-07-18-0185-R
Century, K. S., Holub, E. B., Staskawicz, B. J. (1995). NDR1, a locus of arabidopsis thaliana that is required for disease resistance to both a bacterial and a fungal pathogen. Proc. Natl. Acad. Sci. United States America 92 (14), 6597–6601. doi: 10.1073/pnas.92.14.6597
Chanda, B., Xia, Y., Mandal, M. K., Yu, K., Sekine, K. T., Gao, Q. M., et al. (2011). Glycerol-3-phosphate is a critical mobile inducer of systemic immunity in plants. Nat. Genet. 43 (5), 421–427. doi: 10.1038/ng.798
Chaturvedi, R., Venables, B., Petros, R. A., Nalam, V., Li, M., Wang, X., et al. (2012). An abietane diterpenoid is a potent activator of systemic acquired resistance. Plant J. 71 (1), 161–172. doi: 10.1111/j.1365-313X.2012.04981.x
Chen, Y. C., Holmes, E. C., Rajniak, J., Kim, J. G., Tang, S., Fischer, C. R., et al. (2018). N-hydroxy-pipecolic acid is a mobile metabolite that induces systemic disease resistance in arabidopsis. Proc. Natl. Acad. Sci. United States America 115 (21), E4920–E4929. doi: 10.1073/pnas.1805291115
Chisholm, S. T., Coaker, G., Day, B., Staskawicz, B. J. (2006). Host-microbe interactions: shaping the evolution of the plant immune response. Cell 124 (4), 803–814. doi: 10.1016/j.cell.2006.02.008
Clarke, J. D., Volko, S. M., Ledford, H., Ausubel, F. M., Dong, X. (2000). Roles of salicylic acid, jasmonic acid, and ethylene in cpr-induced resistance in arabidopsis. Plant Cell 12 (11), 2175–2190. doi: 10.1105/tpc.12.11.2175
Coll, N. S., Epple, P., Dangl, J. L. (2011). Programmed cell death in the plant immune system. Cell Death Differ. 18 (8), 1247–1256. doi: 10.1038/cdd.2011.37
Cui, J., Bahrami, A. K., Pringle, E. G., Hernandez-Guzman, G., Bender, C. L., Pierce, N. E., et al. (2005). Pseudomonas syringae manipulates systemic plant defenses against pathogens and herbivores. Proc. Natl. Acad. Sci. United States America 102 (5), 1791–1796. doi: 10.1073/pnas.0409450102
Day, B., Dahlbeck, D., Staskawicz, B. J. (2006). NDR1 interaction with RIN4 mediates the differential activation of multiple disease resistance pathways in arabidopsis. Plant Cell 18 (10), 2782–2791. doi: 10.1105/tpc.106.044693
Delaney, T. P., Friedrich, L., Ryals, J. A. (1995). Arabidopsis signal transduction mutant defective in chemically and biologically induced disease resistance. Proc. Natl. Acad. Sci. United States America 92 (14), 6602–6606. doi: 10.1073/pnas.92.14.6602
Dempsey, D. A., Klessig, D. F. (2012). SOS - too many signals for systemic acquired resistance? Trends Plant Sci. 17 (9), 538–545. doi: 10.1016/j.tplants.2012.05.011
Dewdney, J., Reuber, T. L., Wildermuth, M. C., Devoto, A., Cui, J., Stutius, L. M., et al. (2000). Three unique mutants of arabidopsis identify eds loci required for limiting growth of a biotrophic fungal pathogen. Plant J. 24 (2), 205–218. doi: 10.1046/j.1365-313x.2000.00870.x
Ding, Y., Shaholli, D., Mou, Z. (2015). A large-scale genetic screen for mutants with altered salicylic acid accumulation in arabidopsis. Front. Plant Sci. 5. doi: 10.3389/fpls.2014.00763
Durrant, W. E., Dong, X. (2004). Systemic acquired resistance. Annu. Rev. Phytopathol. 42, 185–209. doi: 10.1146/annurev.phyto.42.040803.140421
Falk, A., Feys, B. J., Frost, L. N., Jones, J. D., Daniels, M. J., Parker, J. E. (1999). EDS1, an essential component of r gene-mediated disease resistance in arabidopsis has homology to eukaryotic lipases. Proc. Natl. Acad. Sci. United States America 96 (6), 3292–3297. doi: 10.1073/pnas.96.6.3292
Fu, Z. Q., Yan, S., Saleh, A., Wang, W., Ruble, J., Oka, N., et al. (2012). NPR3 and NPR4 are receptors for the immune signal salicylic acid in plants. Nature 486 (7402), 228–232. doi: 10.1038/nature11162
Garcion, C., Lohmann, A., Lamodière, E., Catinot, J., Buchala, A., Doermann, P., et al. (2008). Characterization and biological function of the ISOCHORISMATE SYNTHASE2 gene of arabidopsis. Plant Physiol. 147 (3), 1279–1287. doi: 10.1104/pp.108.119420
Geng, X., Cheng, J., Gangadharan, A., Mackey, D. (2012). The coronatine toxin of pseudomonas syringae is a multifunctional suppressor of arabidopsis defense. Plant Cell 24 (11), 4763–4774. doi: 10.1105/tpc.112.105312
Glazebrook, J. (2005). Contrasting mechanisms of defense against biotrophic and necrotrophic pathogens. Annu. Rev. Phytopathol. 43, 205–227. doi: 10.1146/annurev.phyto.43.040204.135923
Greenberg, J. T., Silverman, F. P., Liang, H. (2000). Uncoupling salicylic acid-dependent cell death and defense-related responses from disease resistance in the arabidopsis mutant acd5. Genetics 156 (1), 341–350. doi: 10.1093/genetics/156.1.341
Guttman, D. S., Greenberg, J. T. (2001). Functional analysis of the type III effectors AvrRpt2 and AvrRpm1 of pseudomonas syringae with the use of a single-copy genomic integration system. Mol. Plant-Microbe Interact. 14 (2), 145–155. doi: 10.1094/MPMI.2001.14.2.145
Haney, C. H., Wiesmann, C. L., Shapiro, L. R., Melnyk, R. A., O'Sullivan, L. R., Khorasani, S., et al. (2018). Rhizosphere-associated pseudomonas induce systemic resistance to herbivores at the cost of susceptibility to bacterial pathogens. Mol. Ecol. 27 (8), 1833–1847. doi: 10.1111/mec.14400
Han, S. W., Jung, H. W. (2013). Molecular sensors for plant immunity; pattern recognition receptors and race-specific resistance proteins. J. Plant Biol. 56, 357–366. doi: 10.1007/s12374-013-0323-z
Hartmann, M., Kim, D., Bernsdorff, F., Ajami-Rashidi, Z., Scholten, N., Schreiber, S., et al. (2017). Biochemical principles and functional aspects of pipecolic acid biosynthesis in plant immunity. Plant Physiol. 174 (1), 124–153. doi: 10.1104/pp.17.00222
Hartmann, M., Zeier, T., Bernsdorff, F., Reichel-Deland, V., Kim, D., Hohmann, M., et al. (2018). Flavin monooxygenase-generated n-hydroxypipecolic acid is a critical element of plant systemic immunity. Cell 173 (2), 456–469.e16. doi: 10.1016/j.cell.2018.02.049
Henty-Ridilla, J. L., Shimono, M., Li, J., Chang, J. H., Day, B., Staiger, C. J. (2013). The plant actin cytoskeleton responds to signals from microbe-associated molecular patterns. PloS Pathog. 9 (4), e1003290. doi: 10.1371/journal.ppat.1003290
Jelenska, J., Davern, S. M., Standaert, R. F., Mirzadeh, S., Greenberg, J. T. (2017). Flagellin peptide flg22 gains access to long-distance trafficking in arabidopsis via its receptor, FLS2. J. Exp. Bot. 68 (7), 1769–1783. doi: 10.1093/jxb/erx060
Jiang, S. C., Engle, N. L., Banday, Z. Z., Cecchini, N. M., Jung, H. W., Tschaplinski, T. J., et al. (2021). ALD1 accumulation in arabidopsis epidermal plastids confers local and non-autonomous disease resistance. J. Exp. Bot. 72 (7), 2710–2726. doi: 10.1093/jxb/eraa609
Jirage, D., Tootle, T. L., Reuber, T. L., Frost, L. N., Feys, B. J., Parker, J. E., et al. (1999). Arabidopsis thaliana PAD4 encodes a lipase-like gene that is important for salicylic acid signaling. Proc. Natl. Acad. Sci. United States America 96 (23), 13583–13588. doi: 10.1073/pnas.96.23.13583
Jones, J. D., Dangl, J. L. (2006). The plant immune system. Nature 444 (7117), 323–329. doi: 10.1038/nature05286
Jones, J. D., Vance, R. E., Dangl, J. L. (2016). Intracellular innate immune surveillance devices in plants and animals. Sci. (New York N.Y.) 354 (6316), aaf6395. doi: 10.1126/science.aaf6395
Jung, H. W., Tschaplinski, T. J., Wang, L., Glazebrook, J., Greenberg, J. T. (2009). Priming in systemic plant immunity. Sci. (New York N.Y.) 324 (5923), 89–91. doi: 10.1126/science.1170025
Kang, Y., Jelenska, J., Cecchini, N. M., Li, Y., Lee, M. W., Kovar, D. R., et al. (2014). HopW1 from pseudomonas syringae disrupts the actin cytoskeleton to promote virulence in arabidopsis. PloS Pathog. 10 (6), e1004232. doi: 10.1371/journal.ppat.1004232
Katagiri, F., Tsuda, K. (2010). Understanding the plant immune system. Mol. Plant-Microbe Interact. 23 (12), 1531–1536. doi: 10.1094/MPMI-04-10-0099
Katsir, L., Schilmiller, A. L., Staswick, P. E., He, S. Y., Howe, G. A. (2008). COI1 is a critical component of a receptor for jasmonate and the bacterial virulence factor coronatine. Proc. Natl. Acad. Sci. United States America 105 (19), 7100–7105. doi: 10.1073/pnas.0802332105
Koornneef, A., Pieterse, C. M. (2008). Cross talk in defense signaling. Plant Physiol. 146 (3), 839–844. doi: 10.1104/pp.107.112029
Kunkel, B. N., Bent, A. F., Dahlbeck, D., Innes, R. W., Staskawicz, B. J. (1993). RPS2, an arabidopsis disease resistance locus specifying recognition of pseudomonas syringae strains expressing the avirulence gene avrRpt2. Plant Cell 5 (8), 865–875. doi: 10.1105/tpc.5.8.865
Laluk, K., Luo, H., Chai, M., Dhawan, R., Lai, Z., Mengiste, T. (2011). Biochemical and genetic requirements for function of the immune response regulator BOTRYTIS-INDUCED KINASE1 in plant growth, ethylene signaling, and PAMP-triggered immunity in arabidopsis. Plant Cell 23 (8), 2831–2849. doi: 10.1105/tpc.111.087122
Larionov, A., Krause, A., Miller, W. (2005). A standard curve based method for relative real time PCR data processing. BMC Bioinf. 6, 62. doi: 10.1186/1471-2105-6-62
Lee, M. W., Lu, H., Jung, H. W., Greenberg, J. T. (2007). A key role for the arabidopsis WIN3 protein in disease resistance triggered by pseudomonas syringae that secrete AvrRpt2. Mol. Plant-Microbe Interact. 20 (10), 1192–1200. doi: 10.1094/MPMI-20-10-1192
Lin, N. C., Martin, G. B. (2005). An avrPto/avrPtoB mutant of pseudomonas syringae pv. tomato DC3000 does not elicit pto-mediated resistance and is less virulent on tomato. Mol. Plant-Microbe Interact. 18 (1), 43–51. doi: 10.1094/MPMI-18-0043
Liu, L., Sonbol, F. M., Huot, B., Gu, Y., Withers, J., Mwimba, M., et al. (2016). Salicylic acid receptors activate jasmonic acid signalling through a non-canonical pathway to promote effector-triggered immunity. Nat. Commun. 7, 13099. doi: 10.1038/ncomms13099
Lu, H., Salimian, S., Gamelin, E., Wang, G., Fedorowski, J., LaCourse, W., et al. (2009). Genetic analysis of acd6-1 reveals complex defense networks and leads to identification of novel defense genes in arabidopsis. Plant J. 58 (3), 401–412. doi: 10.1111/j.1365-313X.2009.03791.x
Macho, A. P., Zipfel, C. (2014). Plant PRRs and the activation of innate immune signaling. Mol. Cell 54 (2), 263–272. doi: 10.1016/j.molcel.2014.03.028
Melotto, M., Underwood, W., Koczan, J., Nomura, K., He, S. Y. (2006). Plant stomata function in innate immunity against bacterial invasion. Cell 126 (5), 969–980. doi: 10.1016/j.cell.2006.06.054
Mersmann, S., Bourdais, G., Rietz, S., Robatzek, S. (2010). Ethylene signaling regulates accumulation of the FLS2 receptor and is required for the oxidative burst contributing to plant immunity. Plant Physiol. 154 (1), 391–400. doi: 10.1104/pp.110.154567
Métraux, J. P., Signer, H., Ryals, J., Ward, E., Wyss-Benz, M., Gaudin, J., et al. (1990). Increase in salicylic acid at the onset of systemic acquired resistance in cucumber. Sci. (New York N.Y.) 250 (4983), 1004–1006. doi: 10.1126/science.250.4983.1004
Michelmore, R. W., Christopoulou, M., Caldwell, K. S. (2013). Impacts of resistance gene genetics, function, and evolution on a durable future. Annu. Rev. Phytopathol. 51, 291–319. doi: 10.1146/annurev-phyto-082712-102334
Mishina, T. E., Zeier, J. (2006). The arabidopsis flavin-dependent monooxygenase FMO1 is an essential component of biologically induced systemic acquired resistance. Plant Physiol. 141 (4), 1666–1675. doi: 10.1104/pp.106.081257
Mishina, T. E., Zeier, J. (2007). Pathogen-associated molecular pattern recognition rather than development of tissue necrosis contributes to bacterial induction of systemic acquired resistance in arabidopsis. Plant J. 50 (3), 500–513. doi: 10.1111/j.1365-313X.2007.03067.x
Mittal, S., Davis, K. R. (1995). Role of the phytotoxin coronatine in the infection of arabidopsis thaliana by pseudomonas syringae pv. tomato. Mol. Plant-Microbe Interact. 8 (1), 165–171. doi: 10.1094/mpmi-8-0165
Mudgett, M. B., Staskawicz, B. J. (1999). Characterization of the pseudomonas syringae pv. tomato AvrRpt2 protein: demonstration of secretion and processing during bacterial pathogenesis. Mol. Microbiol. 32 (5), 927–941. doi: 10.1046/j.1365-2958.1999.01403.x
Munch, D., Rodriguez, E., Bressendorff, S., Park, O. K., Hofius, D., Petersen, M. (2014). Autophagy deficiency leads to accumulation of ubiquitinated proteins, ER stress, and cell death in arabidopsis. Autophagy 10 (9), 1579–1587. doi: 10.4161/auto.29406
Mur, L. A., Kenton, P., Lloyd, A. J., Ougham, H., Prats, E. (2008). The hypersensitive response; the centenary is upon us but how much do we know? J. Exp. Bot. 59 (3), 501–520. doi: 10.1093/jxb/erm239
Návarová, H., Bernsdorff, F., Döring, A. C., Zeier, J. (2012). Pipecolic acid, an endogenous mediator of defense amplification and priming, is a critical regulator of inducible plant immunity. Plant Cell 24 (12), 5123–5141. doi: 10.1105/tpc.112.103564
Nawrath, C., Métraux, J. P. (1999). Salicylic acid induction-deficient mutants of arabidopsis express PR-2 and PR-5 and accumulate high levels of camalexin after pathogen inoculation. Plant Cell 11 (8), 1393–1404. doi: 10.1105/tpc.11.8.1393
Ngou, B., Ahn, H. K., Ding, P., Jones, J. (2021). Mutual potentiation of plant immunity by cell-surface and intracellular receptors. Nature 592 (7852), 110–115. doi: 10.1038/s41586-021-03315-7
Ng, G., Seabolt, S., Zhang, C., Salimian, S., Watkins, T. A., Lu, H. (2011). Genetic dissection of salicylic acid-mediated defense signaling networks in arabidopsis. Genetics 189 (3), 851–859. doi: 10.1534/genetics.111.132332
Nicaise, V., Roux, M., Zipfel, C. (2009). Recent advances in PAMP-triggered immunity against bacteria: pattern recognition receptors watch over and raise the alarm. Plant Physiol. 150 (4), 1638–1647. doi: 10.1104/pp.109.139709
Panchal, S., Roy, D., Chitrakar, R., Price, L., Breitbach, Z. S., Armstrong, D. W., et al. (2016). Coronatine facilitates pseudomonas syringae infection of arabidopsis leaves at night. Front. Plant Sci. 7. doi: 10.3389/fpls.2016.00880
Raffaele, S., Rivas, S., Roby, D. (2006). An essential role for salicylic acid in AtMYB30-mediated control of the hypersensitive cell death program in arabidopsis. FEBS Lett. 580 (14), 3498–3504. doi: 10.1016/j.febslet.2006.05.027
Rate, D. N., Greenberg, J. T. (2001). The arabidopsis aberrant growth and death2 mutant shows resistance to pseudomonas syringae and reveals a role for NPR1 in suppressing hypersensitive cell death. Plant J. 27 (3), 203–211. doi: 10.1046/j.0960-7412.2001.1075umedoc.x
Rosebrock, T. R., Zeng, L., Brady, J. J., Abramovitch, R. B., Xiao, F., Martin, G. B. (2007). A bacterial E3 ubiquitin ligase targets a host protein kinase to disrupt plant immunity. Nature 448 (7151), 370–374. doi: 10.1038/nature05966
Ryals, J. A., Neuenschwander, U. H., Willits, M. G., Molina, A., Steiner, H. Y., Hunt, M. D. (1996). Systemic acquired resistance. Plant Cell 8 (10), 1809–1819. doi: 10.1105/tpc.8.10.1809
Sambrook, J., Fritsch, E. R., Maniatis, T. (1989). Molecular cloning: A laboratory manual. 2nd ed (Cold Spring Harbor, NY: Cold Spring Harbor Laboratory Press).
Seguel, A., Jelenska, J., Herrera-Vásquez, A., Marr, S. K., Joyce, M. B., Gagesch, K. R., et al. (2018). PROHIBITIN3 forms complexes with ISOCHORISMATE SYNTHASE1 to regulate stress-induced salicylic acid biosynthesis in arabidopsis. Plant Physiol. 176 (3), 2515–2531. doi: 10.1104/pp.17.00941
Serrano, M., Robatzek, S., Torres, M., Kombrink, E., Somssich, I. E., Robinson, M., et al. (2007). Chemical interference of pathogen-associated molecular pattern-triggered immune responses in arabidopsis reveals a potential role for fatty-acid synthase type II complex-derived lipid signals. J. Biol. Chem. 282 (9), 6803–6811. doi: 10.1074/jbc.M608792200
Seskar, M., Shulaev, V., Raskin, I. (1998). Endogenous methyl salicylate in pathogen-inoculated tobacco plants. Plant Physiol. 116 (1), 387–392. doi: 10.1104/pp.116.1.387
Sobolev, V., Edelman, M., Dym, O., Unger, T., Albeck, S., Kirma, M., et al. (2013). Structure of ALD1, a plant-specific homologue of the universal diaminopimelate aminotransferase enzyme of lysine biosynthesis. acta crystallographica. Section F Struct. Biol. Crystallization Commun. 69 (Pt 2), 84–89. doi: 10.1107/S1744309112050270
Sohn, K. H., Hughes, R. K., Piquerez, S. J., Jones, J. D., Banfield, M. J. (2012). Distinct regions of the pseudomonas syringae coiled-coil effector AvrRps4 are required for activation of immunity. Proc. Natl. Acad. Sci. United States America 109 (40), 16371–16376. doi: 10.1073/pnas.1212332109
Song, J. T., Lu, H., Greenberg, J. T. (2004a). Divergent roles in arabidopsis thaliana development and defense of two homologous genes, aberrant growth and death2 and AGD2-LIKE DEFENSE RESPONSE PROTEIN1, encoding novel aminotransferases. Plant Cell 16 (2), 353–366. doi: 10.1105/tpc.019372
Song, J. T., Lu, H., McDowell, J. M., Greenberg, J. T. (2004b). A key role for ALD1 in activation of local and systemic defenses in arabidopsis. Plant J. 40 (2), 200–212. doi: 10.1111/j.1365-313X.2004.02200.x
Spoel, S. H., Dong, X. (2008). Making sense of hormone crosstalk during plant immune responses. Cell Host Microbe 3 (6), 348–351. doi: 10.1016/j.chom.2008.05.009
Tateda, C., Zhang, Z., Shrestha, J., Jelenska, J., Chinchilla, D., Greenberg, J. T. (2014). Salicylic acid regulates arabidopsis microbial pattern receptor kinase levels and signaling. Plant Cell 26 (10), 4171–4187. doi: 10.1105/tpc.114.131938
Tsuda, K., Mine, A., Bethke, G., Igarashi, D., Botanga, C. J., Tsuda, Y., et al. (2013). Dual regulation of gene expression mediated by extended MAPK activation and salicylic acid contributes to robust innate immunity in arabidopsis thaliana. PloS Genet. 9 (12), e1004015. doi: 10.1371/journal.pgen.1004015
Venugopal, S. C., Jeong, R. D., Mandal, M. K., Zhu, S., Chandra-Shekara, A. C., Xia, Y., et al. (2009). Enhanced disease susceptibility 1 and salicylic acid act redundantly to regulate resistance gene-mediated signaling. PloS Genet. 5 (7), e1000545. doi: 10.1371/journal.pgen.1000545
Vernooij, B., Friedrich, L., Morse, A., Reist, R., Kolditz-Jawhar, R., Ward, E., et al. (1994). Salicylic acid is not the translocated signal responsible for inducing systemic acquired resistance but is required in signal transduction. Plant Cell 6 (7), 959–965. doi: 10.1105/tpc.6.7.959
Wildermuth, M. C., Dewdney, J., Wu, G., Ausubel, F. M. (2001). Isochorismate synthase is required to synthesize salicylic acid for plant defence. Nature 414 (6863), 562–565. doi: 10.1038/35107108
Wu, Y., Zhang, D., Chu, J. Y., Boyle, P., Wang, Y., Brindle, I. D., et al. (2012). The arabidopsis NPR1 protein is a receptor for the plant defense hormone salicylic acid. Cell Rep. 1 (6), 639–647. doi: 10.1016/j.celrep.2012.05.008
Yi, S. Y., Min, S. R., Kwon, S. Y. (2015). NPR1 is instrumental in priming for the enhanced flg22-induced MPK3 and MPK6 activation. Plant Pathol. J. 31 (2), 192–194. doi: 10.5423/PPJ.NT.10.2014.0112
Yi, S. Y., Shirasu, K., Moon, J. S., Lee, S. G., Kwon, S. Y. (2014). The activated SA and JA signaling pathways have an influence on flg22-triggered oxidative burst and callose deposition. PloS One 9 (2), e88951. doi: 10.1371/journal.pone.0088951
Yuan, M., Jiang, Z., Bi, G., Nomura, K., Liu, M., Wang, Y., et al. (2021). Pattern-recognition receptors are required for NLR-mediated plant immunity. Nature 592 (7852), 105–109. doi: 10.1038/s41586-021-03316-6
Zeidler, D., Dubery, I. A., Schmitt-Kopplin, P., Von Rad, U., Durner, J. (2010). Lipopolysaccharide mobility in leaf tissue of arabidopsis thaliana. Mol. Plant Pathol. 11 (6), 747–755. doi: 10.1111/j.1364-3703.2010.00638.x
Zeng, W., He, S. Y. (2010). A prominent role of the flagellin receptor FLAGELLIN-SENSING2 in mediating stomatal response to pseudomonas syringae pv tomato DC3000 in arabidopsis. Plant Physiol. 153 (3), 1188–1198. doi: 10.1104/pp.110.157016
Zhang, X., Chen, S., Mou, Z. (2010). Nuclear localization of NPR1 is required for regulation of salicylate tolerance, isochorismate synthase 1 expression and salicylate accumulation in arabidopsis. J. Plant Physiol. 167 (2), 144–148. doi: 10.1016/j.jplph.2009.08.002
Zhang, C., Gutsche, A. T., Shapiro, A. D. (2004). Feedback control of the arabidopsis hypersensitive response. Mol. Plant-Microbe Interact. 17 (4), 357–365. doi: 10.1094/MPMI.2004.17.4.357
Keywords: Arabidopsis ALD1, avirulent Pseudomonas, plant immune response, petiole exudates, salicylic acid
Citation: Yoo S-J, Choi HJ, Noh SW, Cecchini NM, Greenberg JT and Jung HW (2022) Genetic requirements for infection-specific responses in conferring disease resistance in Arabidopsis. Front. Plant Sci. 13:1068438. doi: 10.3389/fpls.2022.1068438
Received: 12 October 2022; Accepted: 09 November 2022;
Published: 29 November 2022.
Edited by:
Yuelin Zhang, University of British Columbia, CanadaReviewed by:
Kenichi Tsuda, Huazhong Agricultural University, ChinaZhonglin Mou, University of Florida, United States
Copyright © 2022 Yoo, Choi, Noh, Cecchini, Greenberg and Jung. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Ho Won Jung, hwjung@dau.ac.kr
†Present addresses: Sung-Je Yoo, Sangju Agricultural Technology Center, Sangju, South Korea
Hyo Ju Choi, Food Microbiology Division, Food Safety Evaluation Department, National Institute of Food and Drug Safety Evaluation Director General, Cheongju, South Korea