 Xiang Yao
Xiang Yao Haoran Wang1
Haoran Wang1 Maria Oosthuizen
Maria Oosthuizen Wanrong Wei
Wanrong Wei
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Plant Sci. , 30 December 2022
Sec. Functional Plant Ecology
Volume 13 - 2022 | https://doi.org/10.3389/fpls.2022.1036438
Introduction: Burrowing activities of plateau pikas cause widespread bare patches in alpine meadows on the Qinghai-Tibet Plateau, affecting the plant community composition and forage production. However, it is not clear how these bare patches influence the main forage composition in alpine meadows.
Methods: Therefore, we investigated the plant communities in bare patches (BP) and neighboring control plots (CK) in alpine meadows in Maqu county in the Gannan region of China.
Results: Our results showed that plant communities in the CK plots differed from those in the BP plots. The sedge cover, number of sedge species and number of grass species were all significantly higher in the CK plots compared to the BP plots. However, grass cover and its dry weight were significantly higher in the BP plots. Grass cover and the grass dry weight in the BP plots were 1.859 times and 1.802 times higher than that in the CK plots across the five sites, respectively. Grasses also had a significantly higher cover and dry weight than sedge in the BP plots, grass cover was 66.5 times higher than the sedge cover, and the grass dry weight was 68.242 times that of the sedge dry weight. Therefore, bare patches resulting from plateau pika burrowing activity significantly increase the grass/sedge ratio in alpine meadows.
Discussion: A potential explanation is that grasses have a stronger reproductive potential than sedges in bare soil. This has implications for pasture yields since grasses have a higher biomass per unit area compared to sedges in alpine meadows.
The Qinghai-Tibet Plateau (QTP) plays important ecological and economic roles such as climate regulation, supporting wild animals and plants and animal husbandry (Kang et al., 2007). It also serves as the source region of the two most important rivers in China, the Yangtze and Yellow rivers, and is therefore very important for the conservation of water and soil (Sun et al., 2017). The habitat of the QTP consists primarily of alpine meadows and alpine grasslands. Alpine meadows are at higher elevations, between 3200 and 4800 m on the QTP in China (Qiao and Duan, 2016), and are dominated by sedges (Kobresia spp., Cyperaceae) accompanied by grasses and other forbs. At lower elevations, alpine grasslands are dominated by grasses (Gramineae) that are accompanied by forbs (He and Richards, 2015; Lu et al., 2015). Both sedges and grasses are high quality forage for livestock in alpine meadows, but these plant types have different traits. Grasses have a higher plant height and higher biomass per unit area compared to sedges in alpine meadows (Dong et al., 2007). Hence, the sedge/grass proportion will determine the pasture yield. Therefore, it is important to be mindful of the forage composition in alpine meadows.
Recent literature yielded a number of studies regarding the changes in plant community composition in alpine meadows. The plant community in alpine meadows is sensitive to changes in environmental factors such as grazing intensity, climate change, invasive plants, the refuge effect of unpalatable plants and rangeland degradation (Liu et al., 2018). Even light grazing of livestock (Yang et al., 2016) and grazing during the growth period of grasslands (Wei et al., 2022a) can increase plant species richness, while heavy grazing increases the proportion of unpalatable plants and decreases the proportion of palatable graminoids, the plant cover and biomass (Yang et al., 2016; Ji et al., 2020). The plant community composition and biomass are also highly sensitive to climate change in alpine meadows (Yang et al., 2011). For example, higher temperatures increase plant biomass but decreases plant diversity (Baldwin et al., 2014). The proportion of grasses increased while sedges and forbs decreased in alpine meadows following climate warming on the Qinghai-Tibet Plateau (Liu et al., 2018). In addition, another study showed that experimental warming altered the graminoid composition by increasing the frequency of sedges but reducing the frequency of grasses in a subalpine meadow of the Rocky Mountains, USA (Rudgers et al., 2014). However, herbivores can mitigate the response of plant communities to climate change through selective foraging (Olofsson et al., 2004; Post and Pedersen, 2008). Invasive plants primarily influence plant community composition due to their high resource-use efficiency, toxicity or other traits (Funk, 2013). The refuge effect of unpalatable plants increases the plant diversity (Yao et al., 2015), the graminoids diversity and the proportion of grass in alpine sedge meadows (Yao et al., 2019). Moreover, alpine meadows are widely degraded, and the level of degradation has different effects on the cover and biomass of different plant functional types such as sedges, grasses, legumes and forbs (Li et al., 2015).
Rodent activity is also known to regulate vegetation structure. For example, disturbances from prairie dogs (Cynomys ludovicianus) maintain grassland and savanna by preventing the establishment of woody species (Weltzin et al., 1997), while pocket gophers (Geomys bursarius) regulate the vegetation structure by facilitating seedling establishment in alpine meadows (Forbis, 2003). Plateau zokors (Myospalax fontanierii) reduce vegetation cover and height in alpine meadows (Zhang and Liu, 2003). As a habitat, alpine meadows support a large number of plateau pikas (Ochotona curzoniae). Pikas are some of the most representative wild animals in alpine meadows (Qu et al., 2013; Qu et al., 2019), and impact the plant community composition and forage production with their burrowing and feeding activities (Dorji et al., 2014; Zhang et al., 2020; Wei et al., 2022b).
Pika activities can have both positive and negative effects on alpine meadows. Plant species richness is higher in the patches where plateau pikas burrowed compared to control areas in the high-altitude arid rangelands of Trans-Himalaya (Bagchi et al., 2006), however, in bare patches following plateau pika burrowing activities, plant species richness was reported to be significantly lower than in neighboring undisturbed areas (Wei et al., 2007). Intermediate disturbance levels from plateau pika burrowing activities can be beneficial by increasing the forage quality and plant diversity in the alpine meadows (Guo et al., 2012; Liu et al., 2017; Pang and Guo, 2018). Plateau pika burrowing disturbances also benefit clonal growth of sedges (Wang et al., 2020) and the biomass of sedges in alpine meadows is positively correlated with increased burrow density (Liu et al., 2017). Pikas (O. princeps) are regarded as allogenic engineers, and increase the nutrient availability of soil to plants in North America (Aho et al., 1998). High-density plateau pika grazing decreases the biomass of palatable plants in alpine meadows (McIntire and Hik, 2005; Guo et al., 2012; Sun et al., 2015; Liu et al., 2017; Zhang et al., 2020), but increases the biomass and species richness of the other forbs (Zhang et al., 2020). The effects of plateau pikas on alpine meadows appear to be density dependent, since at low densities, plateau pikas have no significant effect on the plant species richness or diversity on winter pastures in alpine meadows (Wei et al., 2020a). The impact of plateau pikas on alpine grassland vegetation is also not uniform within the home range of pika families (Wei et al., 2019). The above examples illustrate the complex nature of plant communities and their interactions with biotic and abiotic factors. However, to date, it is not known how bare patches created by plateau pika burrowing activities influence the grass/sedge ratio in alpine meadows.
We predict that bare patches resulting from plateau pika burrowing will have an inconsistent influence on the biomass and coverage of sedges and grasses, which may change the sedge/grass ratio in alpine meadows. We conducted a study in a representative alpine meadow in Maqu county on the Gannan plateau of China, with the objective to determine the effect of bare patches from plateau pika burrowing activities on the ground cover and biomass of grasses and sedges.
This study was performed in Maqu county in the Gannan Tibetan autonomous prefecture located on the north-eastern Qinghai-Tibet Plateau (33° 40′ N, 101° 54′ E, 3430 m). The research was conducted in a summer grazing rangeland dominated by sedges (Kobresia pygmaea and K. kansuensis) and grasses (Poa crymophila and Elymus nutans). This region has a short warm season from June to September and a long cool season from October to May. The mean annual temperature is 1.2°C, the mean annual precipitation is 564 mm (Wei et al., 2019). The Plateau pika is the most prominent small mammal herbivore that is supported by the alpine meadows.
In order to investigate the effect of bare patches from plateau pika burrowing activity on the plant community, specifically graminoid species (grasses and sedges), therefore bare patches were selected on the condition that graminoid species occur on it. Five sites were selected using the five-point sampling method (Figure 1A) and the sites were approximately 100 m apart. Each site consisted of a 20 m x 20 m square and contained four replicates of a 0.5 m x 0.5 m control plot (CK) and a 0.5 m x 0.5 m bare patch plot (BP) (Figure 1B). Control plots (Figure 1C) and experimental plots (Figure 1D) were in close proximity (less than 1m) to avoid environmental differences of habitat such as edaphic and climatic conditions. Our survey showed that the active burrows density at the five sites was between 800 and 900 burrows per ha, which falls in the high burrow density range according to Wei et al. (2020b).
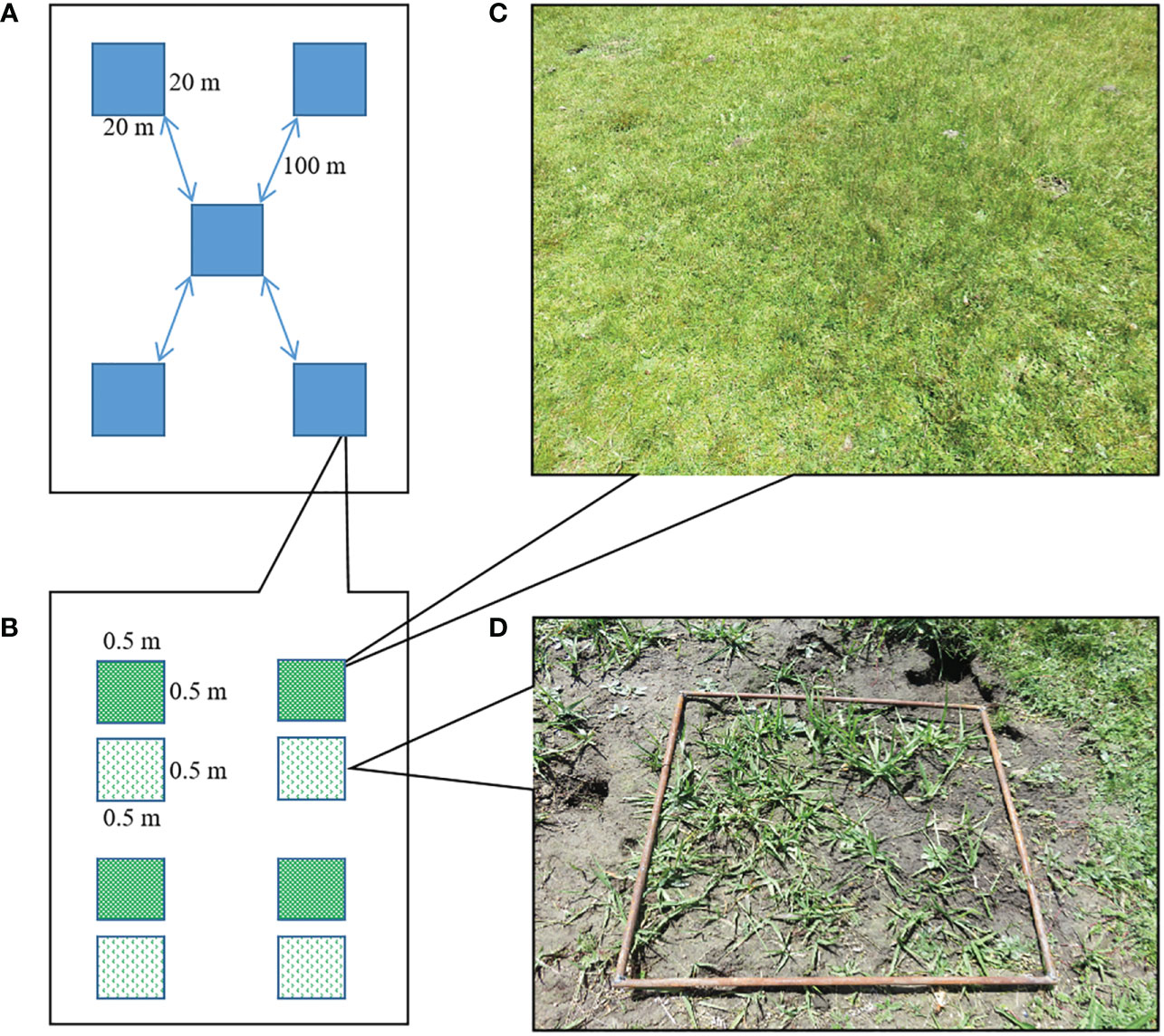
Figure 1 Study design. (A) Five sites were selected, (B) each site had two types of plots, (C) a control plot, (D) and a bare patch plot created by plateau pika burrowing activity.
Plant cover, height and aboveground biomass for each species were determined in August 2019. These indices were then compared for each plant species in the CK plots and the BP plots. Our study focused on differences in graminoid species (in particular Poaceae and Cyperaceae) in the different plots, because they are dominant plant species and important forage for livestock (yak and sheep) in alpine meadows. Livestock had free access to all the plots. The plant cover and plant height of each species were recorded in situ. Plant cover was measured by using the point frame method (Godínez-Alvarez et al., 2009). Plant height was calculated by the average height of 10 randomly selected individual plants per species per plot. All plant species were then clipped to ground level for dry weight measurement. The dry weight of each plant was measured in the laboratory after being oven-dried at 75°C for 48 h.
Principal coordinates analysis (PCoA) was applied to visualize the difference of plant community composition between the CK plots and the BP plots. For PCoA analysis, the plant cover of each plant species was used. Diversity was calculated with the Shannon–Wiener index (Spellerberg and Fedor, 2003).
Data analysis was performed with IBM SPSS 25.0. A nonparametric, Mann-Whitney U test was used to test statistical significance across the whole data set, because the data of cover, plant height and aboveground biomass of some species were not normally distributed. The effect of bare patches from pika burrowing activity on plant communities was assessed using a mixed-effect ANOVA by treating “study site” as a random effect (Verbeke and Lesaffre, 1996). The statistical significance was defined at the 95% confidence level (P = 0.05).
In the control plots, the most abundant plant species was Kobresia humilis, K. kansuensis, Elymus nutans, Poa crymophila and Potentilla fragarioides, whereas the five most abundant plant species in the bare patch plots were E. nutans, P. anserina, Saussurea japonica, Medicago falcata and Lancea tibetica. The complete plant species list and their cover are presented in Supplementary Table S1. Based on the PCoA analysis, it was clear that plant communities in the CK plots were different from plant communities in the BP plots (Figure 2A). Plant communities in the CK plots were highly similar in all the five sites while plant community in the BP plots had more variation (Figure 2A). Graminoid communities also showed high similarity in CK plots while there was more variation in the BP plots (Figure 2B). PCoA 1 explained 77.7% of the variation in plant communities (Figure 2A) and 92.5% of the variation in graminoid communities (Figure 2B).
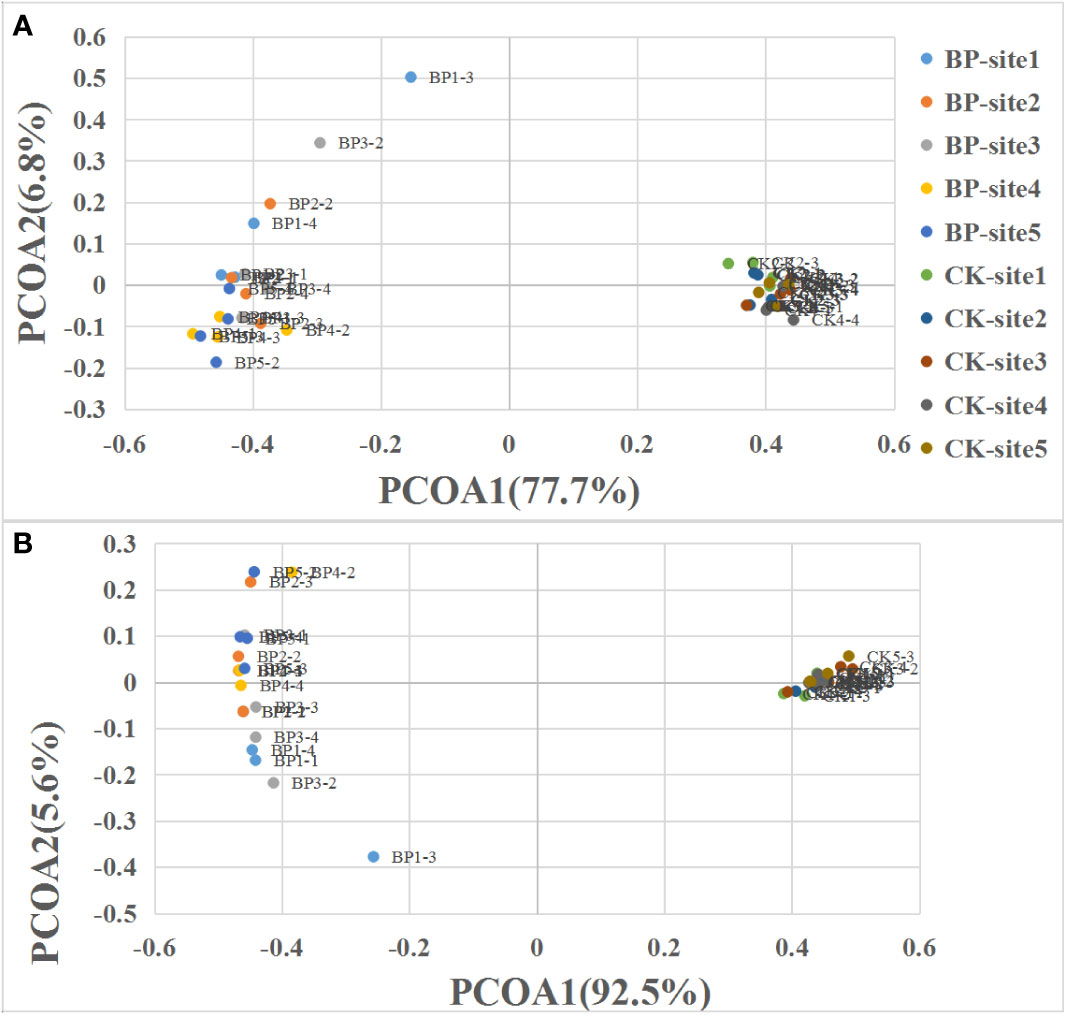
Figure 2 Principal coordinates analysis (PCoA) of (A) the plant community and (B) the graminoid community. Graminoids included grasses and sedges.
Plant communities had a significantly higher total cover (Z= -2.611, P =0.009), total dry weight (Z= -2.611, P =0.009) and a higher plant diversity (Z= -2.611, P =0.009) across the five sites in the control (CK) plots compared to the bare patch (BP) plots (Table 1). There was a significantly higher cover of sedge (Z= -2.611, P =0.009), a higher number of sedge species (Z= -2.668, P =0.008) and higher number of grass species (Z= -2.835, P =0.005) in the CK plots than in the BP plots across the five sites (Table 1). There was a significantly higher cover (Z= -2.402, P =0.016) and dry weight of grass (Z= -2.402, P =0.016) (mainly Elymus nutans in this study) in the BP plots compared to the CK plots across the five sites (Table 1). The grass cover and grass dry weight in the BP plots were 1.859 times and 1.802 times higher than in the CK plots across the five sites (Table 1), respectively. The mixed-effect ANOVA revealed that bare patches significantly influenced the above-mentioned indices in all the five sites (Table 2).
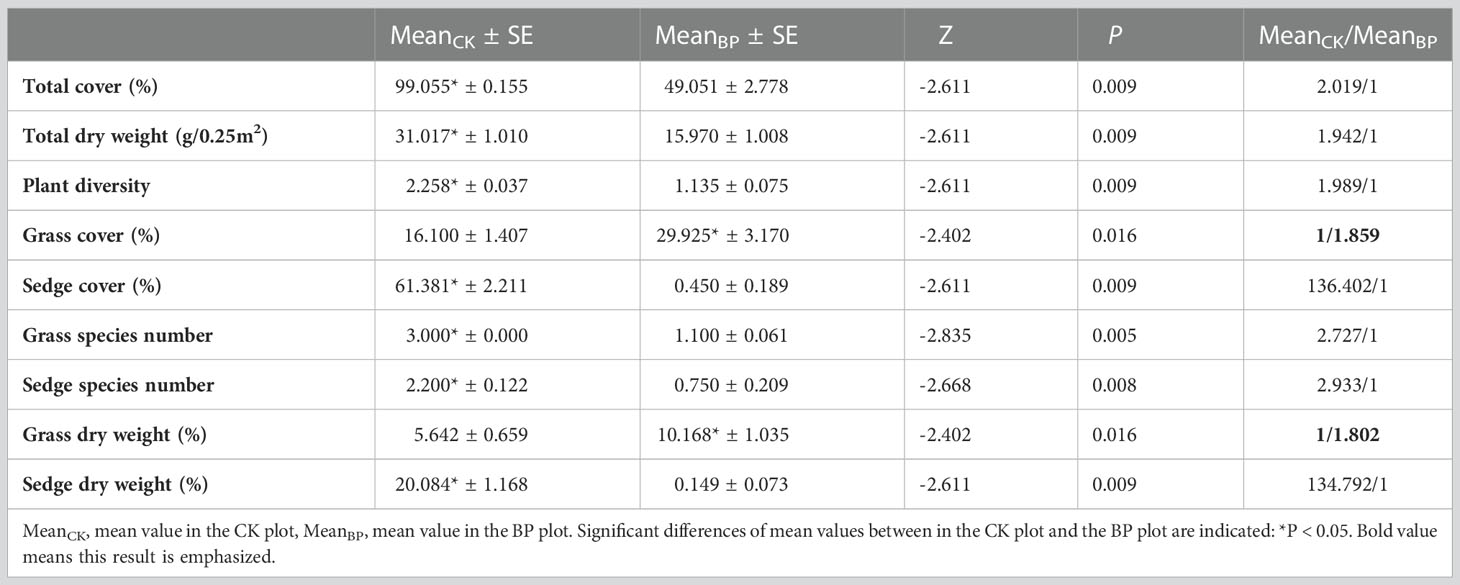
Table 1 Mean value of plant indices across the five sites.
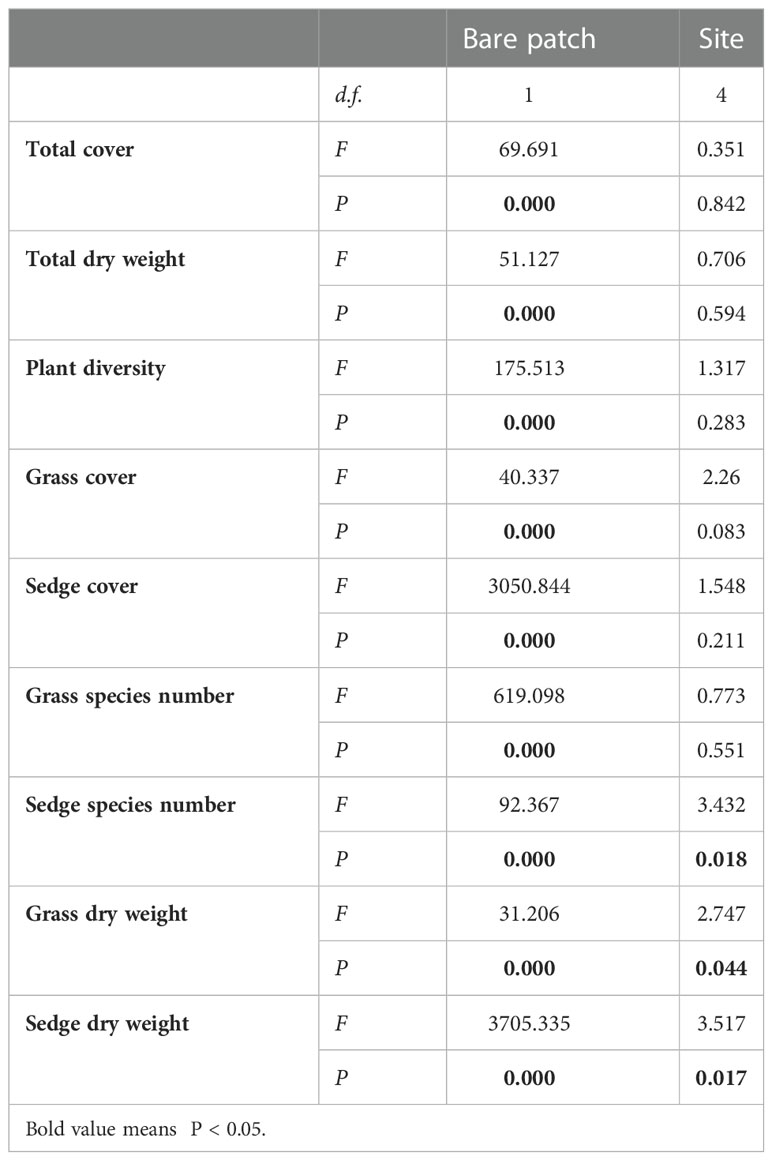
Table 2 Effects of bare patches resulting from plateau pika burrowing activity and sites on plant community as analysed by a mixed-effect ANOVA.
Sedges had a significantly higher cover (Z= -2.611, P =0.009) and higher dry weight (Z= -2.611, P =0.009) than grasses in the CK plots in all the five sites (Table 1). In the CK plot, the sedge cover/grass cover ratio was 3.812, and the sedge dry weight/grass dry weight ratio was 3.560. Grasses had significantly higher cover (Z= -2.611, P =0.009) and a higher dry weight (Z= -2.611, P =0.009) than sedges in the BP plots in all the five sites. In the BP plot, the grass cover/sedge cover ratio was 66.5, and the grass dry weight/sedge dry weight ratio was 68.242.
There are several factors that affect the plant composition in alpine meadows, including grazing density (Yang et al., 2016; Ji et al., 2020), temperature and precipitation (Yang et al., 2011; Baldwin et al., 2014), invasive plants (Funk, 2013), refuge effect of unpalatable plants (Yao et al., 2015; Yao et al., 2019), and burrowing activities of rodents (Dorji et al., 2014; Pang and Guo, 2018; Wang et al., 2020). However, it is not clear how bare patches resulting from plateau pika burrowing activities influence the grass/sedge ratio in alpine meadows. The plateau pika is one of the most widely distributed small animals in alpine meadows. Their continuous burrow digging activity during the day and night has earned them the title of ecosystem engineers (Smith and Foggin, 1999; Lai and Smith, 2003; Wei et al., 2019), that has a large effect on the vegetation structure. In this study, we investigated the plant communities in and around bare patches created by plateau pikas with their burrowing activities in alpine sedge meadows, and found an increased grass:sedge ratio in the bare patches.
Plant communities are more similar in the CK plots than in the BP plots. This is likely because plant communities in the CK plots have been stable for many years while the BP plots are disturbed and modified to open habitats available for plant colonization. The plant communities in the BP plots are largely influenced by factors such as herbivore grazing and the native plant community (Shang and Long, 2007). Similar to our results, a previous study reported that control plots are not significantly different from each other, but variance in the bare patch plots is approximately twice that of the control plots, and can be attributed to the lower species richness in bare patch plots (Smith et al., 2019). In addition, we showed that plant communities have a higher total cover and total dry weight in the CK plots compared to the BP plots. That is probably because newly colonized plants have a lower underground biomass or alternatively, that bare soil is less fertile. Furthermore, alpine meadows have a low mean annual temperature (1.2°C) that is not optimal for plant growth, therefore it is difficult for plants to get established within one or two years in the BP plots.
Previous research indicated that the biomass of sedges increases while the biomass of grasses initially increases and then decreases with increasing burrowing density (Guo et al., 2012; Liu et al., 2017). Moderate disturbances by plateau pikas increases the proportion of graminoids, including grasses and sedges in meadows by increasing graminoid bud density, while a high level of disturbance deteriorates alpine meadows by reducing the graminoid bud density (Wang et al., 2018). In addition, disturbance by plateau pikas benefits clonal growth of sedges in alpine meadows (Wang et al., 2020). In this study, we found that the BP plots have significantly lower sedge cover, a lower number of sedge species and a lower number of grass species than the CK plots, and grasses have higher cover and more biomass than sedges in the BP plots. This indicates that bare patches resulting from plateau pika burrowing initially show an increase in grasses and not sedges. Consistent with our results, a previous study showed that grasses increase while sedges decrease in alpine meadows following climate warming on the Qinghai-Tibet Plateau (Liu et al., 2018). That is possibly because grasses have stronger sexual reproduction than sedges, and can colonize the bare soil more easily and faster. For example, no seedlings of the sedge K. pyagmaea were found on exposed soil in the control experiment and no seedlings of K. myosuroides were recorded in alpine meadows (Forbis, 2003), while Kobresia humilis seedlings were occasionally observed in alpine meadows (Deng et al., 2001). In contrast, the germination rate of Elymus spp. exceeds 80% in alpine meadows on the Qinghai-Tibet Plateau (Bu et al., 2008). This suggests that grasses can colonize bare soil resulting from plateau pika burrowing much faster and with more ease than sedges. Besides, there could also be additional factors that affect the grass/sedge ratio in this study, for example a change in soil properties and change of soil seed banks of bare patches. These factors could be included in future research. A previous study reported that toxic bunchgrass (Achnatherum inebrians) was beneficial for seed production of grasses i.e. sexual reproduction in grazing alpine sedge meadows since it deters livestock (Yao et al., 2019). This promotes the increase of grasses in the alpine sedge meadows.
The plateau pika is one of the most representative wild animals that inhabit the alpine meadows (Wang et al., 2020). In recent years, their densities increased as the meadows degraded. Bare soil resulting from plateau pika burrowing activities accounts for 62.6% of the degraded rangeland in the Qinghai province, and for 25% of the degraded rangeland on the Qinghai-Tibet Plateau (Dong et al., 2013). High densities of plateau pikas will increase the effect on the grass/sedge ratio in alpine meadows.
Grasses and sedges are the most important forages for herbivores, but they have different plant traits. Grasses (Elymus nutans) have a higher plant height and more biomass than Kobresia spp. (Dong et al., 2007), and are widely distributed, from low to very high elevations (1000-5000 m) (Lu, 1993). Kobresia spp. plants are generally distributed at higher elevations from 3000 m to 5960 m (Miehe et al., 2008; Wu et al., 2011). In recent years, the climate is warming, causing the proportion of grasses to increase in alpine meadows on the Qinghai-Tibet Plateau (Liu et al., 2018). Grasses may be able to adapt better to environmental changes, and the increased grass proportion in alpine meadows may render a higher pasture yield.
Plant communities in the CK plots were different from plant communities in the BP plots. There was a significantly higher cover of sedge in the CK plots compared to the BP plots. However, there was significantly higher grass cover in the BP plots than in the CK plots. Grass had significantly higher cover and higher dry weight than sedge in the BP plots. Therefore, we suggest that the bare patches resulting from plateau pika burrowing activities significantly increased the grass/sedge ratio in alpine meadows. That is possibly because grasses have stronger sexual reproduction than sedges in bare soil, promoting the fast colonization of grasses in disturbed and modified habitats. Hence, the pasture yield may be increased since grasses have more biomass per unit area than sedges in alpine meadows.
The original contributions presented in the study are included in the article/Supplementary Material. Further inquiries can be directed to the corresponding author.
XY Wrote the original draft. XY and WW performed the experiment. MO, WW, HW, SZ and YH reviewed and edited the article. All authors contributed to the article and approved the submitted version.
This research was supported by Sichuan Natural Science Foundation of China (23NSFSC5406), the Open Project Funding of the Key Laboratory of Southwest China Wildlife Resource Conservation (Ministry of Education) (XNYB, 22-06) and the Research and Demonstration on Crucial Technologies of Wetland Eco-environment Protection Under the Influence of Highway Engineering on the Western Sichuan Plateau (2019-ZL-19).
Authors SZ and YH were employed by the company Sichuan Jiuma Expressway Co. Ltd.
The remaining authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
RE-JQ declared a past collaboration with the author/s XY, HW to the handling editor at the time of review Chinese Academy of Sciences.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fpls.2022.1036438/full#supplementary-material
Aho, K., Huntly, N., Moen, J., Oksanen, T. (1998). Pikas (Ochotona princeps : Lagomorpha) as allogenic engineers in an alpine ecosystem. Oecologia 114, 405–409. doi: 10.1007/s004420050463
Bagchi, S., Namgail, T., Ritchie, M. E. (2006). Small mammalian herbivores as mediators of plant community dynamics in the high-altitude arid rangelands of trans-himalaya. Biol. Conserv. 127, 438–442. doi: 10.1016/j.biocon.2005.09.003
Baldwin, A. H., Jensen, K., Schönfeldt, M. (2014). Warming increases plant biomass and reduces diversity across continents, latitudes, and species migration scenarios in experimental wetland communities. Global Change Biol. 20, 835–850. doi: 10.1111/gcb.12378
Bu, H., Xu, X., Liu, K., Jia, P., Wen, S., Sun, D., et al. (2008). Seed germinating characteristics of 54 gramineous species in the alpine meadow on the eastern qinghai-Tibet plateau. Front. Biol. China 3, 187–193. doi: 10.17521/cjpe.2006.0082
Deng, Z., Xie, X., Zhou, X. (2001). Primary study on reproductive strategies of Kobresia humilis population in alpine meadow. Chin. J. Ecol. 20, 68–70.
Dong, S. K., Kang, M. Y., Yun, X. J., Yun, X. J., Long, R. J., Hu, Z. Z., et al. (2007). Economic comparison of forage production from annual crops, perennial pasture and native grassland in the alpine region of the qinghai-Tibetan plateau, China. Grass. Forage. Sci. 62, 405–415. doi: 10.1111/j.1365-2494.2007.00594.x
Dong, Q. M., Zhao, X. Q., Wu, G. L., Shi, J. J., Ren, G. H. (2013). A review of formation mechanism and restoration measures of “black-soil-type” degraded grassland in the qinghai-Tibetan plateau. Environ. Earth Sci. 70, 2359–2370. doi: 10.1007/s12665-013-2338-7
Dorji, T., Moe, S. R., Klein, J. A., Totland, Ø. (2014). Plant species richness, evenness, and composition along environmental gradients in an alpine meadow grazing ecosystem in central Tibet, China. Arct. Antarct. Alp. Res. 46, 308–326. doi: 10.1657/1938-4246-46.2.308
Forbis, T. A. (2003). Seedling demography in an alpine ecosystem. Am. J. Bot. 90, 1197–1206. doi: 10.3732/ajb.90.8.1197
Funk, J. L. (2013). The physiology of invasive plants in low-resource environments. Conserv. Physiol. 1, 17. doi: 10.1093/conphys/cot026
Godínez-Alvarez, H., Herrick, J. E., Mattocks, M., Toledo, D., Van Zee, J. (2009). Comparison of three vegetation monitoring methods: Their relative utility for ecological assessment and monitoring. Ecol. Indic. 9, 1001–1008. doi: 10.1016/j.ecolind.2008.11.011
Guo, Z. G., Li, X. F., Liu, X. Y., Zhou, X. R.. (2012). Response of alpine meadow communities to burrow density changes of plateau pika (Ochotona curzoniae) in the qinghai-Tibet plateau. Acta Ecol. Sin. 32, 44–49. doi: 10.1016/j.chnaes.2011.12.002
He, S., Richards, K. (2015). Impact of meadow degradation on soil water status and pasture management–a case study in Tibet. Land. Degrad. Dev. 26, 468–479. doi: 10.1002/ldr.2358
Ji, L., Qin, Y., Jimoh, S. O., Hou, X., Zhang, N., Gan, Y., et al. (2020). Impacts of livestock grazing on vegetation characteristics and soil chemical properties of alpine meadows in the eastern qinghai-Tibetan plateau. Ecoscience 27, 107–118. doi: 10.1080/11956860.2019.1710908
Kang, L., Han, X., Zhang, Z., Sun, O. J. (2007). Grassland ecosystems in China: review of current knowledge and research advancement. Phil. Trans. R. Soc B. 362, 997–1008. doi: 10.1098/rstb.2007.2029
Lai, C. H., Smith, A. T. (2003). Keystone status of plateau pikas (Ochotona curzoniae): effect of control on biodiversity of native birds. Biodivers. Conserv. 12, 1901–1912. doi: 10.1023/A:1024161409110
Liu, Y., Fan, J., Shi, Z., Yang, X., Harris, W. (2017). Relationships between plateau pika (Ochotona curzoniae) densities and biomass and biodiversity indices of alpine meadow steppe on the qinghai–Tibet plateau China. Ecol. Eng. 102, 509–518. doi: 10.1016/j.ecoleng.2017.02.026
Liu, H., Mi, Z., Lin, L., Wang, Y., Zhang, Z., Zhang, F., et al. (2018). Shifting plant species composition in response to climate change stabilizes grassland primary production. P. Natl. Acad. Sci. U.S.A. 115, 4051–4056. doi: 10.1073/pnas.1700299114
Li, J., Zhang, F., Lin, L., Li, H., Du, Y., Li, Y., et al. (2015). Response of the plant community and soil water status to alpine Kobresia meadow degradation gradients on the qinghai–Tibetan plateau, China. Ecol. Res. 30, 589–596. doi: 10.1007/s11284-015-1258-2
Lu, B. R. (1993). Meiotic studies of Elymus nutans and E. jacquemontii (Poaceae, triticeae) and their hybrids with Pseudoroegneria spicata and seventeen Elymus species. Plant Syst. Evol. 186, 193–212. doi: 10.1007/BF00940798
Lu, H., Cong, J., Liu, X., Wang, X., Tang, J., Li, D., et al. (2015). Plant diversity patterns along altitudinal gradients in alpine meadows in the three river headwater region, China. Acta Pratacult. Sin. 24, 197–204. doi: 10.1007/bf00940798
McIntire, E. J. B., Hik, D. S. (2005). Influences of chronic and current season grazing by collared pikas on above-ground biomass and species richness in subarctic alpine meadows. Oecologia 145, 287–296. doi: 10.11686/cyxb2014347
Miehe, G., Miehe, S., Kaiser, K., Liu, J., Zhao, X. (2008). Status and dynamics of Kobresia pygmaea ecosystem on the Tibetan plateau. Ambio 37, 272–279. doi: 10.1579/0044-7447(2008)37[272:Sadotk]2.0.Co;2
Olofsson, J., Hulme, P. E., Oksanen, L., Suominen, O. (2004). Importance of large and small mammalian herbivores for the plant community structure in the forest tundra ecotone. Oikos 106, 324–334. doi: 10.1111/j.0030-1299.2004.13224.x
Pang, X. P., Guo, Z. G. (2018). Effects of plateau pika disturbance levels on the plant diversity and biomass of an alpine meadow. Grassl. Sci. 64, 159–166. doi: 10.1111/grs.12199
Post, E., Pedersen, C. (2008). Opposing plant community responses to warming with and without herbivores. P. Natl. Acad. Sci. U.S.A. 105, 12353–12358. doi: 10.1073/pnas.0802421105
Qiao, Y., Duan, Z. (2016). “Understanding alpine meadow ecosystems,” in Landscape and ecosystem diversity, dynamics and management in the yellow river source zone. Eds. Brierley, G. J., Li, X., Cullum, C., Gao, J. (Cham: Springer International Publishing), 117–135.
Qu, J., Li, W., Yang, M., Ji, W., Zhang, Y. (2013). Life history of the plateau pika (Ochotona curzoniae) in alpine meadows of the Tibetan plateau. Mamm. Biol. 78, 68–72. doi: 10.1016/j.mambio.2012.09.005
Qu, J., Réale, D., Fletcher, Q. E., Zhang, Y. (2019). Among-population divergence in personality is linked to altitude in plateau pikas (Ochotona curzoniae). Front. Zoo. 16, 1–7. doi: 10.1186/s12983-019-0329-6
Rudgers, J. A., Kivlin, S. N., Whitney, K. D., Price, M. V., Waser, N. M., Harte, J., et al. (2014). Responses of high-altitude graminoids and soil fungi to 20 years of experimental warming. Ecology 95, 1918–1928. doi: 10.1890/13-1454.1
Shang, Z., Long, R. (2007). Formation causes and recovery of the “Black soil type” degraded alpine grassland in qinghai-Tibetan plateau. Front. Agric. China 1, 197–202. doi: 10.11733/j.issn.1007-0435.2018.01.001
Smith, A. T., Badingqiuying, Wilson, M. C., Hogan, B. (2019). Functional-trait ecology of the plateau pika Ochotona curzoniae in the qinghai–Tibetan plateau ecosystem. Integr. Zool. 14, 87–103. doi: 10.1111/1749-4877.12300
Smith, A. T., Foggin, J. M. (1999). The plateau pika (Ochotona curzoniae) is a keystone species for biodiversity on the Tibetan plateau. Anim. Conserv. 2, 235–240. doi: 10.1017/S1367943099000566
Spellerberg, I. F., Fedor, P. J. (2003). A tribute to Claude Shannon, (1916–2001) and a plea for more rigorous use of species richness, species diversity and the ‘Shannon–wiener’ index. Global Ecol. Biogeogr. 12, 177–179. doi: 10.1046/j.1466-822X.2003.00015.x
Sun, F., Chen, W., Liu, L., Liu, W., Cai, Y., Smith, P., et al. (2015). Effects of plateau pika activities on seasonal plant biomass and soil properties in the alpine meadow ecosystems of the Tibetan plateau. Grassl. Sci. 61, 195–203. doi: 10.1111/grs.12101
Sun, Q., Li, B., Zhou, C., Li, F., Zhang, Z., Ding, L., et al. (2017). A systematic review of research studies on the estimation of net primary productivity in the three-river headwater region, China. J. Geogr. Sci. 27, 161–182. doi: 10.1007/s11442-017-1370-z
Verbeke, G., Lesaffre, E. (1996). A linear mixed-effects model with heterogeneity in the random-effects population. J. Am. Stat. Assoc. 91, 217–221. doi: 10.1080/01621459.1996.10476679
Wang, Q., Yu, C., Pang, X. P., Jin, S. H., Zhang, J., Guo, Z. Z., et al. (2018). The disturbance and disturbance intensity of small and semi-fossorial herbivores alter the belowground bud density of graminoids in alpine meadows. Ecol. Eng. 113, 35–42. doi: 10.1016/j.ecoleng.2018.01.003
Wang, Y., Zhang, X., Sun, Y., Zhang, Z., Sun, Y. (2020). Pika burrow and zokor mound density and their relationship with grazing management and sheep production in alpine meadow. Ecosphere 11, e03088. doi: 10.1002/ecs2.3088
Wei, W., An, S., Zheng, Q., Qin, M., Chen, T. (2022a). Structural changes in vegetation coincident with reseeding Elymus nutans can increase perceived predation risk of plateau pikas (Ochotona curzoniae). Appl. Anim. Behav. Sci. 255, 105745. doi: 10.1016/j.applanim.2022.105745
Wei, W. R., He, J. D., Zheng, Q. Y. (2020a). Plateau pikas (Ochotona curzoniae) at low densities have no destructive effect on winter pasture in alpine meadows. Rangeland. J. 42, 55–61. doi: 10.1071/RJ19042
Wei, W., He, J., Zheng, Q., He, Y., Oosthuizen, M. K. (2020b). A comparison of density estimation methods in plateau pika populations in an alpine meadow ecosystem. J. Mamma. 101, 1091–1096. doi: 10.1093/jmammal/gyaa049
Wei, W., Knops, J. M. H., Zhang, W. (2019). The impact of plateau pikas (Ochotona curzoniae) on alpine grassland vegetation and soil is not uniform within the home range of pika families. Plant Ecol. Divers. 12, 417–426. doi: 10.1080/17550874.2019.1628113
Wei, X., Li, S., Yang, P., Chen, H.. (2007). Soil erosion and vegetation succession in alpine Kobresia steppe meadow caused by plateau pika–a case study of nagqu county, Tibet. Chin. Geogr. Sci. 17, 75–81. doi: 10.1007/s11769-007-0075-0
Wei, W., Zhen, Q., Deng, J., Yue, H., Qin, M., Oosthuizen, M. K., et al. (2022b). Grazing during the grassland greenup period promotes plant species richness in alpine grassland in winter pastures. Front. Plant Sci. 13. doi: 10.3389/fpls.2022.973662
Weltzin, J. F., Archer, S., Heitschmidt, R. K. (1997). Small-mammal regulation of vegetation structure in a temperate savanna. Ecology 78, 751–763. doi: 10.1890/0012-9658(1997)078[0751:SMROVS]2.0.CO;2
Wu, Y., Wu, J., Deng, Y., Tan, H., Du, Y., Gu, S., et al. (2011). Comprehensive assessments of root biomass and production in a Kobresia humilis meadow on the qinghai-Tibetan plateau. Plant Soil 338, 497–510. doi: 10.1007/s11104-010-0562-4
Yang, H., Wu, M., Liu, W., Zhang, Z., Zhang, N., Wan, S., et al. (2011). Community structure and composition in response to climate change in a temperate steppe. Global Change Biol. 17, 452–465. doi: 10.1111/j.1365-2486.2010.02253.x
Yang, Z. A., Xiong, W., Xu, Y., Jiang, L., Zhu, E., Zhan, W., et al. (2016). Soil properties and species composition under different grazing intensity in an alpine meadow on the eastern Tibetan plateau, China. Environ. Monit. Assess. 188, 678. doi: 10.1007/s10661-016-5663-y
Yao, X., Chai, Q., Chen, T., Chen, Z., Wei, X., Bao, G., et al. (2019). Disturbance by grazing and the presence of rodents facilitates the dominance of the unpalatable grass Achnatherum inebrians in alpine meadows of northern China. Rangeland. J. 41, 301–312. doi: 10.1071/rj18096
Yao, X., Christensen, M. J., Bao, G., Zhang, C., Li, X., Li, C., et al. (2015). A toxic endophyte-infected grass helps reverse degradation and loss of biodiversity of over-grazed grasslands in northwest China. Sci. Rep-UK. 5, 18527. doi: 10.1038/srep18527
Zhang, Y., Liu, J. (2003). Effects of plateau zokors (Myospalax fontanierii) on plant community and soil in an alpine meadow. J. Mamma. 8, 644–651. doi: 10.1644/1545-1542(2003)084<0644:EOPZMF>2.0.CO;2
Keywords: bare soil, grass, sedge, graminoid, pasture yield, plateau pika
Citation: Yao X, Wang H, Zhang S, Oosthuizen M, Huang Y and Wei W (2022) Impact of plateau pika burrowing activity on the grass/sedge ratio in alpine sedge meadows in China. Front. Plant Sci. 13:1036438. doi: 10.3389/fpls.2022.1036438
Received: 04 September 2022; Accepted: 15 December 2022;
Published: 30 December 2022.
Edited by:
Rubén Retuerto, Universidade de Santiago de Compostela, SpainReviewed by:
Julia Sanchez Vilas, Cardiff University, United KingdomCopyright © 2022 Yao, Wang, Zhang, Oosthuizen, Huang and Wei. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Wanrong Wei, d2Vpd3IxOEAxMjYuY29t
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.