 Jovanka Vladejić
Jovanka Vladejić Fen Yang
Fen Yang Eva Dvořák Tomaštíková
Eva Dvořák Tomaštíková Jaroslav Doležel
Jaroslav Doležel Jan J. Palecek
Jan J. Palecek Ales Pecinka
Ales Pecinka- 1Institute of Experimental Botany (IEB), Czech Acad Sci, Centre of the Region Haná for Biotechnological and Agricultural Research (CRH), Olomouc, Czechia
- 2Department of Cell Biology and Genetics, Faculty of Science, Palacký University, Olomouc, Czechia
- 3National Centre for Biomolecular Research (NCBR), Faculty of Science, Masaryk University, Brno, Czechia
The integrity of plant genetic information is constantly challenged by various internal and external factors. Therefore, plants use a sophisticated molecular network to identify, signal and repair damaged DNA. Here, we report on the identification and analysis of four uncharacterized Arabidopsis BRCT5 DOMAIN CONTAINING PROTEINs (BCPs). Proteins with the BRCT5 domain are frequently involved in the maintenance of genome stability across eukaryotes. The screening for sensitivity to induced DNA damage identified BCP1 as the most interesting candidate. We show that BCP1 loss of function mutants are hypersensitive to various types of DNA damage and accumulate an increased number of dead cells in root apical meristems upon DNA damage. Analysis of publicly available sog1 transcriptomic and SOG1 genome-wide DNA binding data revealed that BCP1 is inducible by gamma radiation and is a direct target of this key DNA damage signaling transcription factor. Importantly, bcp1 plants showed a reduced frequency of somatic homologous recombination in response to both endogenous and induced DNA damage. Altogether, we identified a novel plant-specific DNA repair factor that acts downstream of SOG1 in homology-based repair.
Introduction
Genome stability is constantly threatened by internally and externally-induced DNA damage (Razqallah, 2008; Chatterjee and Walker, 2017). Among others, the presence of damaged DNA negatively affects DNA replication, transcription, and cell cycle progression. Therefore, living organisms developed a sophisticated safeguarding system that recognizes various types of DNA damage, signals their presence, and activates specific molecular effectors that repair the damaged site. This prevents the occurrence of potentially deleterious mutations. Once the repair is completed, the halted cellular processes are restarted and continued. Numerous studies demonstrated that DNA damage repair is essential for the normal growth and fertility of plants, similar to other organisms (Manova and Gruszka, 2015; Nisa et al., 2019) However, despite a generally high degree of evolutionary conservation of the eukaryotic DNA repair system, several unique DNA repair factors evolved in plants (Yoshiyama et al., 2013b; Hu et al., 2016).
Depending on the type of DNA damage, specific DNA repair pathways are activated. A common and highly toxic type of lesion is DNA double-strand break (DSB), which may be generated by external or internal factors. Its persistence in the genome may lead to a loss of genetic information, structural genome changes, and even cell death. The DSB repair begins with a recognition of the damaged site by the MRN (MRE11-RAD50-NBS1) complex and phosphorylation of histone variant H2A.X to produce gamma-H2A.X. This stimulates the binding of the transcription factor Breast cancer type 1 susceptibility protein (BRCA1), followed by signaling through Ataxia Telangiectasia Mutated (ATM) and/or ATM- and RAD3-related (ATR) kinases. The kinase activity of ATM and/or ATR activates the p53 transcription factor at the sites of DNA damage in metazoa and its functional homolog SUPPRESSOR OF GAMMA RADIATION 1 (SOG1) in plants (Preuss and Britt, 2003; Seton-Rogers, 2006; Yoshiyama et al., 2009; Hafner et al., 2019). During the following steps, these transcription factors orchestrate various responses, including pausing of the cell cycle, promotion repair by non-homologous end-joining (NHEJ) or homologous recombination (HR), or (in extreme cases) cell death. In contrast with the error-prone NHEJ, HR represents an error-free mechanism where an intact DNA molecule homologous to the damaged site is used as a template for repair (Heyer et al., 2010). Although the mechanism of HR is studied in great detail across the major branches of the tree of life, not all molecular factors taking place in this process are known.
A prominent group of proteins associated with cell cycle regulation and DNA damage repair contains the BRCA1 C-Terminus (BRCT) domain (Bork et al., 1997), which consists of approximately 100 amino acids and mediates protein-protein interactions by binding to the phosphate groups (Yu et al., 2003). Later studies in animals and yeasts suggested several structurally distinct types of BRCT domains (Wan et al., 2016) The best-studied examples of plant BRCT domain-containing proteins are the BRCA1 and its homolog BREAST CANCER ASSOCIATED RING 1 (BARD1). Both proteins are required for normal levels of somatic HR in plants, and their loss of function mutants are hypersensitive to DNA damage (Trapp et al., 2011). A conspicuous type of BRCT domain is the BRCT5 that was found in budding and fission yeast proteins Rtt107 and Brc1, respectively, and in human protein NSE5/SLF1 (Williams et al., 2010; Li et al., 2012; Räschle et al., 2015). These proteins represent species-specific cofactors involved in the loading of the evolutionary conserved DNA damage repair complex Structural maintenance of chromosomes 5/6 (SMC5/6) to chromatin (Leung et al., 2011; Räschle et al., 2015; Oravcová et al., 2019). However, none of the currently known plant SMC5/6 complex interactors contains this domain. Another example of BRCT5 domain-containing protein includes human Pax2 transactivation domain-interacting protein (PTIP) that performs ATM-dependent activation of p53 and thus promotes DSB repair in mammals (Yan et al., 2011). PTIP also lacks a functional homolog in plants. Therefore, BRCT5 domain proteins represent a little understood group in plants.
Our study demonstrates that analyzing plant proteins carrying BRCT5 domain-containing is an attractive route toward discovering new players involved in the control of plant genome stability. Thus we performed in silico identification of Arabidopsis BRCT5 DOMAIN CONTAINING PROTEINs (BCPs). Subsequently, loss of function mutants of four genes was characterized by the expression pattern and hypersensitivity to DNA damage. The most promising candidate BCP1 was analyzed as to its role in HR-based repair.
Materials and methods
Plant materials and growth conditions
Unless stated otherwise, all Arabidopsis thaliana (Arabidopsis) genotypes used in this study had Columbia (Col-0) background. T-DNA lines used in this study were: GK_301C08 (bcp1-1), SALK_001578C (bcp1-2), SALK_022790 (bcp1-3), GK_076D08 (bcp2-1), SALK_111173C (bcp3-1), SALK_038422 (bcp4-1), and SALK_123114C (smc6b-1). T-DNA mutant lines were obtained from the SALK institute (Alonso et al., 2003) and GABI-Kat (Kleinboelting et al., 2012) via the European Arabidopsis Stock Centre (NASC). Double mutants were generated by crossing homozygous single mutants and analyzing progeny in F2 generation by PCR-based genotyping for both mutations. HR reporter lines B11 in the C24 background (Swoboda et al., 1994) and IC9C (Puchta et al., 1995; Molinier et al., 2004) were crossed with bcp1-1. The resulting hybrids were grown into F4 generation and selected by PCR for double homozygous lines. The oligonucleotides used for genotyping are listed in Supplementary Table 1.
Plants used for phenotyping, seed generation, and crossing were grown in climate-controlled phytotron under long-day conditions (at 16 h light, 150 μmol m−2 s−1 intensity, 19°C during the day; 8 h at 18°C during the night). In vitro plant cultivation was done in an air-conditioned phytochamber with a long day regime (16h light, 150 µmol m-2 s-1, 21°C, 8h dark, 19°C).
Basic local alignment search tool and BRCT5 domain structure comparisons
New Arabidopsis BRCT5 domain-containing proteins were identified by the BLAST search (Altschul et al., 1990) using the fission yeast Schizosaccharomyces pombe Brc1 (SpBrc1) and human Homo sapiens NSE5 (HsNSE5) proteins against the Arabidopsis thaliana database (taxid: 3702). The retrieved BRCT5 domain sequences were manually aligned, and their AlphaFold structural models (Varadi et al., 2022) were compared.
Molecular cloning, plant transformation, and GUS assays
To develop the promoter-reporter line, a region 2000 bp upstream of the BCP1 transcription start site (ProBCP1) was amplified by PCR and cloned by Gateway Technology (ThermoFisher Scientific; Cat. nos.: 11789100, 12538120) into pDONR207 (Invitrogen) and then recombined into the binary vector pKGWFS7.0, containing uidA gene encoding β-glucuronidase (GUS). Plasmids carrying the ProBCP1::GUS fusion were transformed into Agrobacterium tumefaciens strain GV3101. Transformation of Arabidopsis Col-0 was performed using the floral dip method. (Zhang et al., 2006). The selection of transformed plants in T1 generation was carried out on a medium containing 100 μg/ml kanamycin (Sigma-Aldrich, Cat. no. 60615). Resistant plants were transferred to soil for seed production. The following generation (T2) of plants was selected based on the activation of the GUS reporter gene. The oligonucleotides used for genotyping or cloning are listed in Supplementary Table 1.
The expression pattern of BCP1 in highly dividing vegetative tissues, as well as the change of expression under DNA damage stress, was examined by using a ProBCP1::GUS reporter line. Plants grown for seven days on a solid medium were transferred for 24 h to liquid ½ MS medium with or without 10 μM mitomycin C (MMC, Cat. no. M0503), a genotoxic agent causing intrastrand DNA crosslinks. Following the treatments, plants were stained by GUS histochemical staining. GUS solution containing 10 mM EDTA (Sigma-Aldrich, Cat. no. E5134), 2 mM potassium ferrocyanide (Lachema, Cat. no. 68 4514), 2 mM potassium ferricyanide (Lachema), 100 mM disodium phosphate (Penta, Cat. no. 15150), 100 mM monosodium phosphate (Lachema, Cat. no. 68 4639), 0.1% Triton X-100 (Sigma-Aldrich, Cat. no. T8787) and 2 mM X-Gluc (Thermo Scientific, Cat. no. R0852) was prepared as described in (Baubec et al., 2009). Seedlings were transferred to 5 ml tubes and infiltrated with GUS staining solution under a vacuum. After five to ten minutes, the vacuum was released and tubes were placed at 37°C overnight. Subsequently, the GUS staining solution was removed and plants were cleared by incubation in 70% ethanol (v/v) at 37°C. Ethanol was changed 3 times, and after the last change, plants were left overnight at 4°C. Pictures were taken under a stereo-microscope (Olympus SZX16) and fluorescent microscope (Olympus BX60).
For the analysis of BCP1 expression in the reproductive tissues, inflorescences were fixed in 90% (v/v) acetone and incubated for 45 min at -20°C. Acetone was then removed, and samples were washed three times with 100 mM phosphate buffer, pH 7.2. After washing, the flowers were infiltrated with GUS staining solution under a vacuum for 10 min and left overnight at 37°C. The next morning the solution was removed, and samples were washed shortly with phosphate buffer and cleared in chloral hydrate solution containing eight parts chloral hydrate (Sigma-Aldrich, Cat. no. 23100) two parts water, and one part glycerol (Sigma-Aldrich, Cat. no. G516). Flowers were mounted on the microscope slide and dissected in the same solution. Pictures were taken under a stereo-microscope (Olympus SZX16) and fluorescent microscope (Olympus BX60).
Root sensitivity assays
For root sensitivity assays, surface sterilized and stratified seeds were grown on ½ MS growth medium with 0.6% agar (w/v) and 1% sucrose (w/v). Seeds were sterilized in 70% ethanol (v/v) for 5 min, followed by 8% sodium hypochlorite solution (v/v) for 6-10 min, and washed 3 times in sterile water. Seeds were stratified for 48h in 0.1% agarose solution (w/v) at 4°C in the dark. Stratified seeds were evenly distributed on Petri dishes containing ½ MS medium (mock) or ½ MS medium supplemented with 10 μM MMC (Sigma-Aldrich, Cat. no. M0503), 20 nM camptothecin (CPT; Sigma-Aldrich, Cat. no. C9911), 20 μM zebularine (Sigma-Aldrich, Cat. no. Z4775), or 50 nM bleomycin (Sigma-Aldrich, Cat. no. 203408-M). Plants grown for seven days in a horizontal position were then carefully pulled off the medium using tweezers and laid flat on a plate with agar. The length of the primary root was measured using the ImageJ plugin SmartRoot (Lobet et al., 2011). Experiments were performed in three biological replicates with typically 20 plants per replicate (minimum of 11 plants in one replicate). Statistical significance was tested with One-way ANOVA with posthoc Tukey HSD in Minitab.
Cell death assays
Sterilized and stratified seeds were grown vertically on plates with ½ MS medium with 0.8% agar (w/v) for five days and then transferred into liquid ½ MS medium for a 24 h treatment. Mock samples were grown in pure liquid ½ MS medium, while treated plants had medium supplemented with 10 μM MMC. Following the treatment, seedlings were stained with 10 mg.mL-1 propidium iodide solution (Sigma) on glass microscope slides. Visualization and photography were performed using Leica confocal microscope TCS SP8 (Leica, Wetzlar, Germany) and HC PL APO CS2 20x/0.75 DRY objective equipped with Leica LAS-X software with Leica Lightning module laser scanning confocal microscope (Leica). At least 13 plants for each group were analyzed. The means of the three replicates are depicted. Statistical significance was tested withKruskall-Wallis H-test with post hoc Conover-Iman test of multiple comparisons using rank sums with Benjamini-Hochberg procedure in R 4.2.1 (R Core Team, 2018).
Homologous recombination assays
The B11, B11 bcp1-1, IC9C, and IC9C bcp1-1 plants were grown on ½ MS medium with or without (mock) 1 μM MMC under sterile conditions. Ten days-old seedlings were histochemically stained using GUS as described above. Plants were transferred to a Petri dish containing ethanol and examined using a stereo-microscope (Olympus SZX16) for HR events identified as blue-stained cells or areas. The means of the three replicates are depicted. Statistical significance was tested withMann-Whitney U-tes in Minitab (www.minitab.com).
Fresh weight measurements
Plants were grown as described in homologous recombination assays were measured on an analytical scale. Measuring was done in triplicates, and each sample was composed of 60 seedlings. Mann-Whitney U-test (P < 0.05) was used to assess the significance of weight differences (www.minitab.com).
RNA-seq data analysis
RNA-seq data for wild-type and sog1-1 plants were obtained from a publicly available dataset (Bourbousse et al., 2018). The database contains gene expression values (fragment per kilobase per million reads, FPKM) in plants grown under normal and DNA damaging conditions at six-time points post gamma irradiation (20 min, 1.5, 3, 6, 12, and 24 h). From it, we acquired expression profiles of BCPs. The changes in gene expression were assessed as described in the results. For the assessment of statistical significance, we used a two-sample T-test with unequal variances.
Reverse transcription-quantitative polymerase chain reaction
T-DNA mutants lines’ seeds, sterilized and stratified, were grown on ½ MS medium with 0.6% agarose. Seven days old seedlings were sampled and flash-frozen in liquid nitrogen. RNA extraction was performed by RNeasy Mini Kit (Qiagen, Cat. no. 74104). cDNA was constructed with RevertAid H Minus First Strand cDNA Synthesis Kit (Thermo Scientific™, Cat. no. K1631). The qPCR was performed with the HOT FIREPol® EvaGreen® qPCR Mix Plus (Solis BioDyne, Cat. no. 08-24-0000S) in CFX96 Touch Real-Time PCR Detection System. Wild type and sog1-1 plants were grown for seven days in ½ MS 0.6% agarose and then transferred to liquid ½ MS with or without 40 μM MMC for a 1 h treatment. Following the treatment samples were flash-frozen in liquid nitrogen, and RNA extraction, cDNA synthesis and RT-qPCR were performed as described above. Mann-Whitney U-test was performed in Minitab to assess statistical significance of the data.
Accession numbers
Gene information and sequences used in this article can be found in TAIR under the following accession numbers: BCP1 (AT4G02110), BCP2 (AT2G41450), BCP3 (AT4G03130), BCP4 (AT3G21480), SMC6B (AT5G61460).
Results
Identification of Arabidopsis BRCT5 domain-containing proteins
To identify potential Arabidopsis BRCT5 domain-containing proteins, we performed a BLAST search (Altschul et al., 1990) using the BRCT5 domains of the fission yeast Schizosaccharomyces pombe Brc1 (SpBrc1) and human Homo sapiens NSE5 (HsNSE5) proteins against the Arabidopsis protein database. Three candidates, At4g02110, At4g03130, and At4G21070, were found as potential genes of interest. We hypothesized that the large phylogenetic distance between Arabidopsis versus yeast and human might have reduced the efficiency of such a screen and compromised the direct identification of some candidates. Therefore, we performed an additional search for the BRCT5 domain-containing proteins in the genome of moss Physcomitrium patens using the same query sequences from SpBrc1 and HsNSE5. The BRCT5 domains were found in the moss proteins Pp3c4_1630, Pp3c7_24750, Pp3c8_2040, and Pp3c11_4990. As the next step, the moss proteins were BLASTed against the Arabidopsis genome, which revealed three additional genes At1g04020, At2g41450, and At3g21480. The sequence comparison identified a conserved pattern of amino acids (Figure 1A) with different properties typical for the BRCT5 type domain that supported all the candidates identified via BLAST. The candidates At4g21070 and At1g04020 were previously described as Arabidopsis orthologs of human BREAST CANCER SUSCEPTIBILITY 1 (BRCA1) and its homolog BREAST CANCER ASSOCIATED RING 1 (BARD1), whose functions in plant DNA damage repair have been already documented (Lafarge and Montané, 2003; Reidt et al., 2006). Therefore, both BRCA1 and BARD1 were excluded from subsequent analyses. Based on this, we selected the remaining four candidate proteins for further analysis and named them BRCT5 DOMAIN-CONTAINING PROTEINs (BCPs): BCP1 (At4g02110), BCP2 (At2g41450), BCP3 (At4g03130) and BCP4 (At3g21480). The superimposition of computationally modeled BRCT5 domains of SpBrc1 and the four selected BCPs revealed their high structural similarity (Figure 1B). Based on Araport11 gene annotation, BCP1 is a cell cycle regulated transcriptional coactivator (Menges et al., 2002). Based on its two BRCT domains, it was also considered a possible candidate for plant homolog of the human DNA topoisomerase 2-binding protein 1 (TOPBP1), but the overall low sequence similarity did not allow for the drawing of a firm conclusion (Shultz et al., 2007). BCP2 is described as N-acetyltransferase (Araport11) and is expressed in the female gametophyte (Wuest et al., 2010). BCP3 and BCP4 are both described as BRCT domain-containing DNA repair proteins (Araport11), likely based on the presence of their C-terminal BRCT domains, with no further information. Hence, we identified six BRCT5 domain-containing candidates in Arabidopsis, with four of them representing uncharacterized Arabidopsis genes.
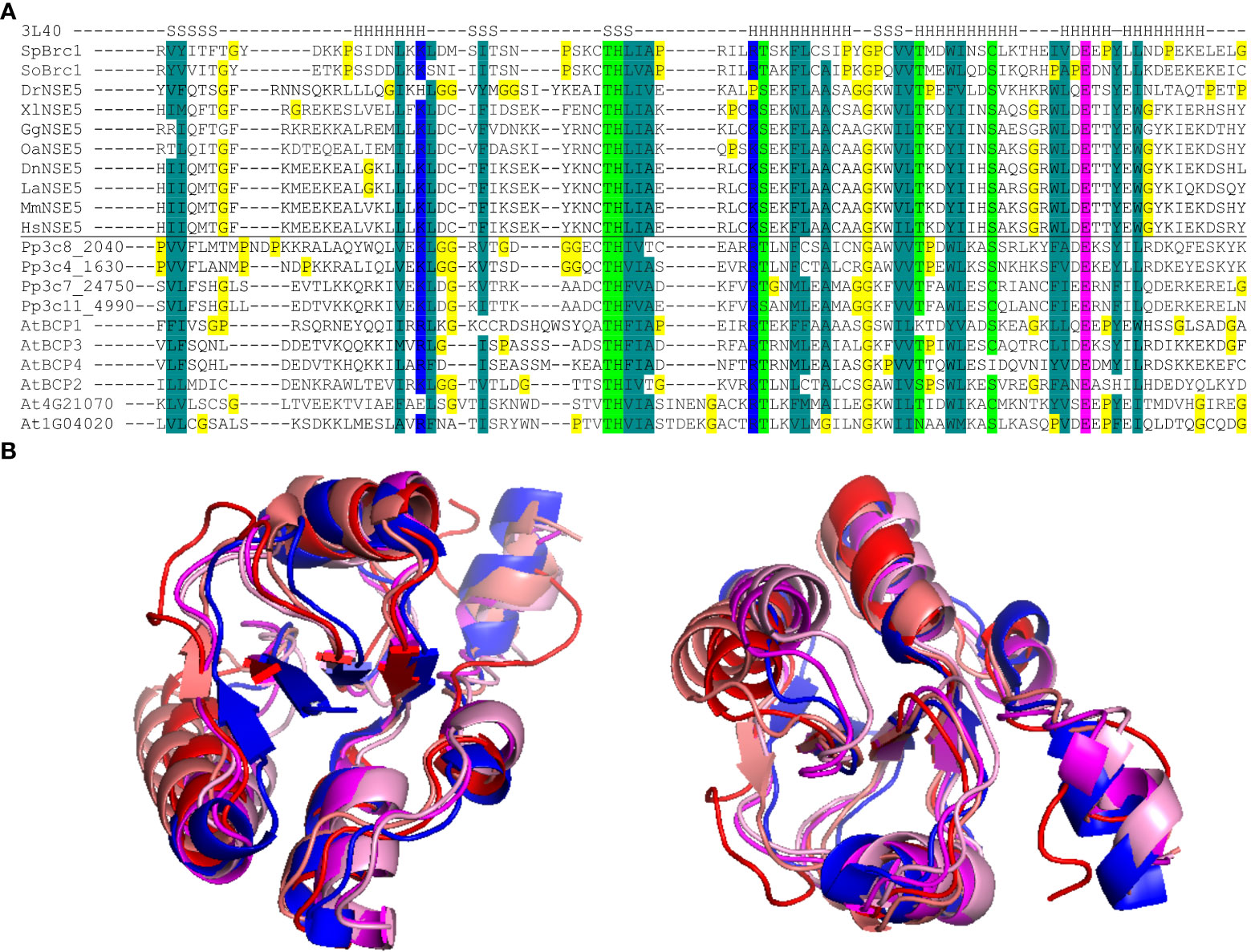
Figure 1 BRCT5 domain analysis. (A) Alignment of the core part of the BRCT5 domain with helical (H) and β-strand (S) segments above (from PDB: 3L40 structure; Williams et al., 2010). The Brc1 and NSE5 orthologs are from Schizosaccharomyces pombe (Sp), S. octosporus (So), Physcomitrium patens (Pp), Arabidopsis thaliana (At), Danio rerio (Dr), Xenopus laevis (Xl), Gallus gallus (Gg), Ornithorhynchus anatinus (Oa), Monodelphis domestica (Md), Dasypus novemcinctus (Dn), Loxodonta africana (La), Mus musculus (Mm), Homo sapiens (Hs). Coloring indicates amino acid groups conserved across the family: dark green, hydrophobic and aromatic; light green, polar; blue, acidic; pink, basic; all glycine and proline residues are highlighted in yellow. (B) Superimposition of modeled BRCT5 domains of At4g02110 (red), At2g41450 (deep pink), At4g03130 (coral), At3g21480 (pale pink), and crystal structure SpBrc1 (blue) shown from two views.
Loss of BCP1 causes sensitivity to DNA damage, and its transcription is SOG1 dependent
To test for the potential role of BCPs in DNA damage repair, we isolated their T-DNA insertional mutants (Figure 2A). All homozygous mutants were viable and did not show any obvious developmental defects during somatic development and/or sterility during the reproductive stage.

Figure 2 Phenotypes of mutants in uncharacterized Arabidopsis BRCT5 domain-containing genes. (A) Gene and protein structures of the BRCT5 CONTAINING PROTEINS (BCPs). The positions of T-DNAs in the used mutant alleles are indicated by black triangles above the gene models. Introns are indicated by a horizontal line and exons by green (untranslated regions) and purple (coding sequence) colors. Protein models under gene models (grey) show the position of known domains: BRCT - blue rectangles and GNAT (Gcn5-related N-acetyltransferase) - brown rectangle. (B) Representative phenotypes of seven days old wild-type (WT) and homozygous mutant plants grown on media containing 20 μM zebularine (ZEB), 10 μM mitomycin C (MMC), 20 nM camptothecin (CPT), 50nM nM bleocin (BLEO). The smc6b-1 served as a sensitive control. Scale bar = 1 cm. (C) Root length of WT and mutant plants under control (mock) conditions. Error bars indicate the standard deviation between the means of three biological replicates. The letters above columns indicate similarities between samples. The same letters indicate samples that were not significantly different in one-way ANOVA with post hoc Tukey’s test (P < 0.05). (D) Root length of WT and mutants from B under DNA damaging treatments relative to the growth of the same genotype under mock conditions. Error bars represent the standard deviation between three biological replicates, each with at least 15 plants. Statistics were performed as in (C).
As the first step, we performed an initial screening for mutant sensitivity to different types of DNA damage. The aim was to identify if any of the genes are important for DNA damage repair. To this end, we focused on the induction of all possible types of DNA damage including DSBs, DNA inter-strand, and DNA-protein crosslinks. Seeds were germinated, plants were grown on the genotoxin-containing media, and their root length was measured (Figure 2B). The smc6b-1 mutant allele of STRUCTURAL MAINTENANCE OF CHROMOSOMES 6B (SMC6B) served as a hypersensitive control. Under mock conditions, only the bcp4-1 mutant plants had significantly shorter roots (9.4 mm ± 1.5 mm in bcp1-4 compared to 13.23 mm ± 0.68 mm in WT control), while the root length of the remaining mutants was not significantly different from the wild type (Figure 2C). Under DNA damaging conditions, the bcp1-1 plants were hypersensitive to 10 μM DNA inter-strand cross-linker MMC, 20 nM DNA-protein cross-linker CPT and 20 nM radiomimetic agent bleomycin causing DNA strand breaks. The bcp1-1 plants also exhibited sensitivity to 20 μM type I DNA-protein cross-linker zebularine (Prochazkova et al., 2022). The bcp2-1, bcp3-1, and bcp4-1 mutant plants did not show significantly increased sensitivity to any of the genotoxic treatments (Figure 2D).
Absence of sensitivity in combination with non-coding sequence location of T-DNAs stimulated us to analyze the expression of BCPs in their corresponding T-DNA insertion mutant lines by RT-qPCR (Supplementary Figure 1). The bcp1-1 and bcp4-1 showed a very strongly reduced amount of transcript compared to their WT variants. Surprisingly, the bcp2-1 with T-DNA insertion in the first out of total 13 exons showed more than 90-fold over-expression of BCP2. This might be caused by the expression from the Cauliflower Mosaic Virus 35S promoter that is part of the T-DNA insertion. The bcp3-1 showed no significant difference in the amount of transcript compared to wild type. This suggests that bcp1-1 and bcp4-1 are loss of function mutants, bcp2-1 is a potential overexpressor line and bcp3-1 might not affect BCP3 gene function.
Next, we analyzed the expression of the BCP candidates using available transcriptomic data. Surprisingly, none of the selected candidates is represented on the Arabidopsis ATH1 expression array. RNA-sequencing-based atlas of Arabidopsis developmental stages (Klepikova et al., 2016) revealed that BCP2 and BCP3 were only weakly expressed throughout the whole plant development and that the expression slightly increased only in some floral parts (Supplementary Figure 2). In contrast, both BCP1 and BCP4 showed a low to moderate expression with the highest values observed in floral organs and seeds. Surprisingly, only weak expression was found in the root tissues. To find a potential involvement of BCPs in DNA damage response, we analyzed their expression after gamma-irradiation in wild-type and sog1 mutant background using a publicly available RNA-seq dataset (Bourbousse et al., 2018). Under ambient (mock) conditions, BCP1 was expressed stably at a basal level in both WT and sog1-1 plants (Supplementary Figure 3A). In response to gamma-irradiation, BCP1 was upregulated 3.2-fold already 20 min post-treatment, and the amount of transcript reached its 14-fold increase maximum 1.5 h post-irradiation (Figure 3A). The amount of transcript lowered over time and returned to mock levels 24 h after the treatment. In the sog1 plants, gamma radiation-induced expression was not observed, suggesting that transcriptional response of BCP1 to DNA damage is SOG1-dependent, and that BCP1 acts downstream of SOG1. This was confirmed also in RT-qPCR experiment where BCP1 was significantly up-regulated in response to MMC treatment in wild-type but not in sog1-1 mutant plants (Figure 3B). However, the same amount of BCP1 expression in mock-treated wild-type and sog1-1 plants indicates that basal BCP1 expression is SOG1 independent. The remaining genes BCP2, BCP3, and BCP4 showed only minor transcriptional changes that differed between wild-type and mutant plants, mostly at solitary time-points, suggesting that these genes are not gamma-irradiation inducible and their expression is not SOG1-dependent (Figure 3A).
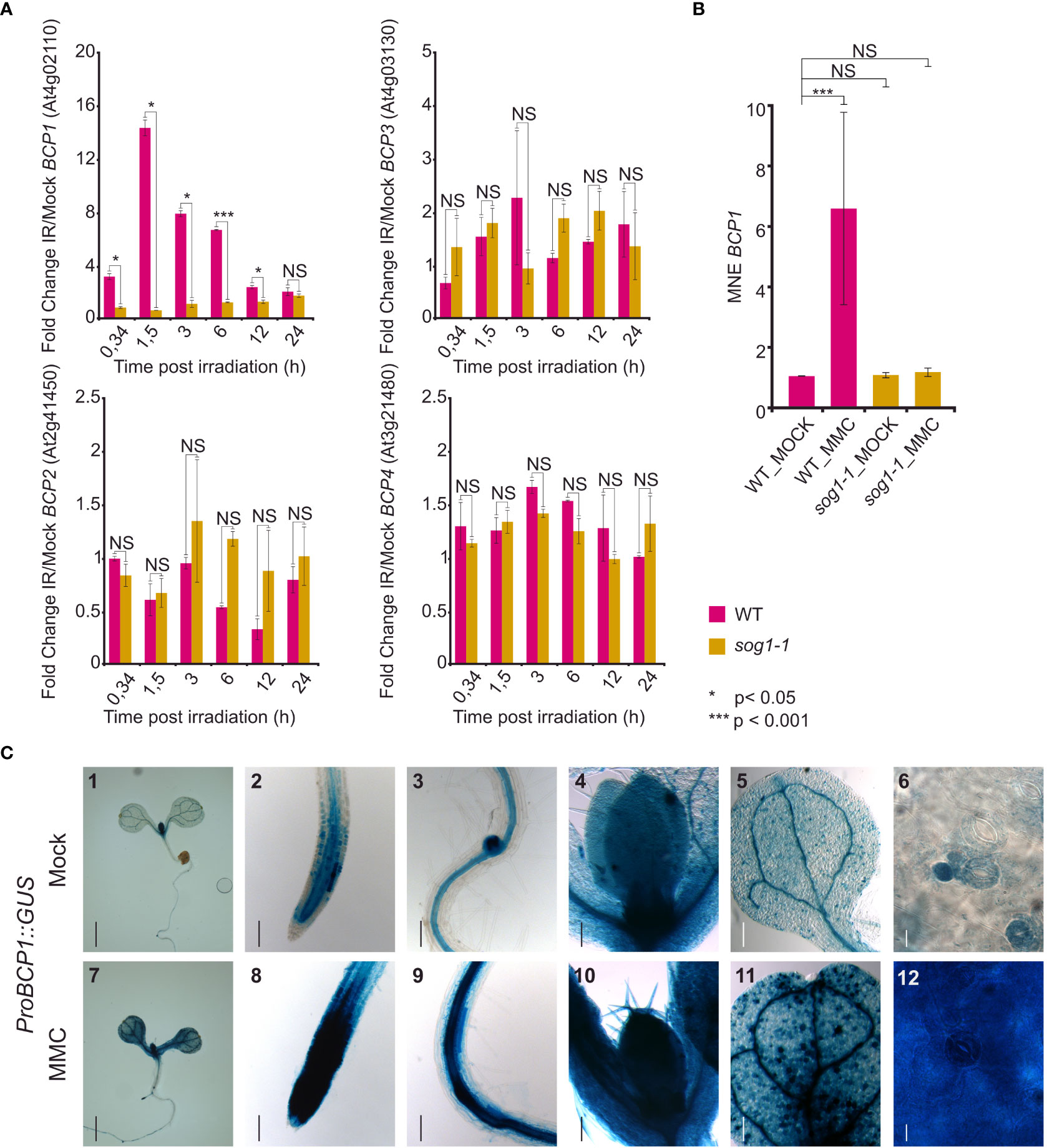
Figure 3 Expression of the BCP genes. (A) Relative expression of BCP1 to BCP4 in wild type (WT) and sog1-1 based on RNA-sequencing experiment of Bourbousse et al. (2018). The normalized read counts from RNA-sequencing (FPKM) were used to calculate fold change in expression after gamma irradiation pulse (IR) versus mock conditions (y-axis). The x-axis indicates the harvesting time after the irradiation treatment. Error bars represent the standard error of the mean. Asterisks represent significant differences in two sample T-test with unequal variance, * P < 0.05, *** P < 0.001, NS – not significantly different. (B) Reverse transcription qPCR analysis of BCP1 expression in wild-type (WT) and sog1-1 plants without (MOCK) and after 1 h treatment with 40 μM MMC (MMC). Y-axis shows mean normalized expression relative to PP2A. Error bars show three biological replicates. NS = not significantly different, *** statistically significantly different in Mann-Whitney U-test at P < 0.05. (C) In planta analysis of BCP1 promoter activity. Seven days old seedlings carrying ProBCP1::GUS were transferred to mock and DNA damaging conditions for 24h. BCP1 promoter activity was monitored using GUS histochemical staining. Representative stereo microscope pictures of tissues showing the gene expression. (1,7) Whole seedling, (2, 8) shoot apical meristem, (3, 9) lateral root meristem, (4, 10) shoot apical meristem with first real leaves, (5, 11) cotyledon, (6, 12) leaf blade cells. Scale bars: 1,7 = 1 mm; 2-5,8-11 = 100 μm, 6,12 = 50 μm.
To gain more insights into the transcriptional response of BCP1 to DNA damage, we generated stable Arabidopsis transformants carrying BCP1 promoter fused with the GUS reporter gene (ProBCP1::GUS). Under mock conditions, the BCP1 promoter was active in tissues with actively dividing cells, such as the root and shoot apical meristems, lateral root meristems, and vasculature (Figure 3C_2,3,4). The signals in true leaves had a peculiar dotted pattern. After inspection at the cellular level, it was obvious that these “dots” are represented by young stomata guard cells, stomatal lineage ground cells, and guard mother cells. The older (larger) stomata guard cells and pavement cells showed little or no GUS signals (Figure 3C_5,6). As the Arabidopsis transcriptomic atlas (Klepikova et al., 2016) data suggested the highest BCP1 transcript amount in reproduction (Supplementary Figure 2), we further examined the pattern of BCP1 expression in inflorescences (Supplementary Figures 4A–E). We found particularly strong GUS signals in pistils throughout the entire flower development, young stamen, filaments, and perianth of closed flowers.
To visually confirm that the BPC1 transcription is induced by DNA damage, as suggested by the transcriptomic data, we exposed seedlings of the ProBCP1::GUS reporter line to 10 μM MMC for 24 h and subsequently scored BCP1 promoter activity. Intense signals appeared in almost all parts of the plant, including the true leaves (Figure 3C_7-12). This strongly supports transcriptomic data and demonstrates that BCP1 transcription is inducible by DNA damage. Based on these experiments, we considered BCP1 as the most promising candidate for further analysis.
BCP1 is required for the repair of various types of DNA damage
To validate our initial findings based on a single mutant allele, we isolated two more BCP1 T-DNA-insertional mutants located in the 8th exon (bcp1-2) and the 7th intron (bcp1-3) (Figure 4A). Phenotypic analysis of all three homozygous mutant lines confirmed the absence of obvious developmental defects at four and six weeks of age (Supplementary Figure 5).
Next, we extended the sensitivity assays by exposing plants of all bcp1 mutant lines to 10 μM MMC, 20 μM zebularine, and 20 nM CPT (Figures 4B, D). In mock conditions bcp1-1 and bcp1-2 mutants showed no difference in root length compared to wild type, while bcp1-3 plants had slightly longer roots (Figure 4C). Both bcp1-1 and bcp1-2 alleles were significantly sensitive to all three drug treatments. In contrast, the intronic mutant bcp1-3 was hypersensitive only to 10 μM MMC (Figure 4D). This is in agreement with the amount of BCP1 transcript which was almost not detectable in bcp1-1 and bcp1-2 mutants but was not significantly reduced in bcp3-1 (Supplementary Figure 1). Specifically, MMC-treated wild-type plants reached 38.8 ± 3.6% of the standard root length compared to mock conditions, while it was only 21.8 ± 1.7%, 25.5 ± 2.5%, and 27 ± 3% of the mock-treated plant root length for bcp1-1, bcp1-2, and bcp1-3, respectively (all comparisons P <0.001 one-way ANOVA with post hoc Tukey HSD). Zebularine-treated wild-type plants reached 53.3 ± 1.7% of the mock control length. For zebularine treated bcp1-1, bcp1-2 and bcp1-3 plants it was 42.7 ± 5.6%, 41.6 ± 1.25% and 50.5. ± 4.0%, respectively (all comparisons P <0.001). Similarly, CPT-treated wild-type plants reached 43.2 ± 4% of the normal root length, but it was 25.5 ± 1.1%, 37.4 ± 1.4%, and 47.5 ± 2% for the individual bcp1 mutant alleles, respectively (all comparisons P <0.001 one-way ANOVA with post hoc Tukey HSD). The sensitive control smc6b-1 plants had massive root length reduction to 16 ± 2%, 10 ± 0.3%, and 20 ± 1% for MMC, zebularine, and CPT (all comparisons P <0.001 one-way ANOVA with post hoc Tukey HSD).
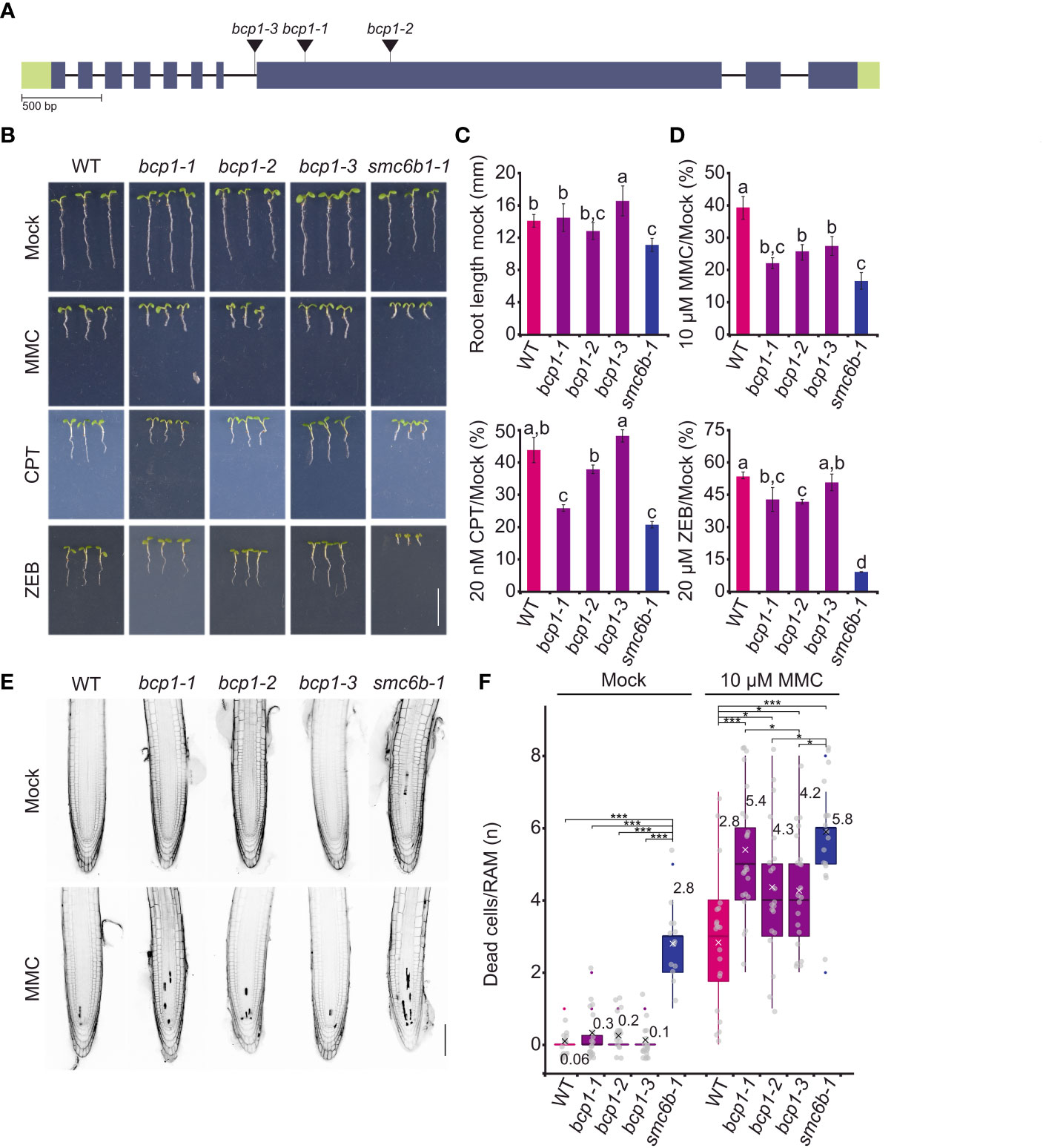
Figure 4 BCP1 is required for normal resistance to DNA damaging treatments. (A) Gene model of BCP1 with indicated positions of all used T-DNA insertional mutants. The style follows the description in Figure 2A. (B) DNA damage sensitivity assays of different bcp1 alleles. Representative phenotypes of seven days old wild-type (WT) and homozygous mutant plants grown on media containing 20 μM zebularine (ZEB), 10 μM mitomycin C (MMC), and 20 nM camptothecin (CPT). The smc6b-1 served as a sensitive control. Scale bar = 1 cm. (C) Root length of WT and mutant plants grown under control (mock) conditions. Error bars indicate the standard deviation between the means of nine biological replicates. The letters above columns indicate similarity between samples. The same letters indicate samples that were not significantly different in one-way ANOVA with posthoc Tukey’s test (P < 0.05). (D) Root length of WT and mutants under DNA damaging treatments relative to the growth of the same genotypes under mock conditions. Statistics were performed as in (C). (E) Representative confocal microscopy images of the primary roots stained by propidium iodide in cell death assays. Five days old seedlings of WT and bcp1 mutants were exposed to mock or 10 μM MMC treatments for 24 h, stained with propidium iodide, and analyzed to reveal dead cells that appear as dark sectors inside the roots. The smc6b-1 served as a control with increased cell death. Scale bar = 100 μm. (F) Quantification of dead cells per root apical meristem in different genotypes and treatments (complements E). Each gray dot indicates the number of dead cells per root (n = 13-22). The boxplots’ hinges are in the 1st and 3rd quartile, with a marked median. The mean is indicated by a cross with a numerical value. Whisker marks show the lowest or highest value within the 1.5 interquartile range below or above hinges. Statistical significance was tested by Kruskall-Wallis H-test with post hoc Conover-Iman test of multiple comparisons with Benjamini-Hochberg procedure (P < ½ α, α = 0.05). NS - not significant, * P < 0.025, *** P < 0.001.
To assess the extent of damage at the cellular level, we performed a cell death assay based on the staining of root apices with propidium iodide (PI), where PI marks the dead cells and is excluded from the living cells (Figures 4E, F). Wild-type and smc6b-1 were used as standard and hypersensitive controls. Mock-treated wild-type and bcp1 plants showed no significantly different mean values of less than one dead cell per root (Figures 4E, F). After the treatment with 10 μM MMC for 24 h, the median number of dead cells per root increased to three in wild type, four in bcp1-2 and bcp1-3, and five in bcp1-1 (Figure 4F). The values in all mutant lines were significantly higher compared to wild-type plants. This shows that loss of function from BCP1 makes Arabidopsis plants hypersensitive to diverse types of DNA damage and leads to increased cell death.
BCP1 is required for normal frequency of homologous recombination
Based on the SOG1-dependent transcriptional activation of BCP1 upon DNA damage and hypersensitivity of bcp1 plants to DNA damaging treatments, we hypothesized about a possible role of BCP1 in HR. To experimentally test this hypothesis, we generated double homozygous bcp1-1 B11 and bcp1-1 IC9C lines (Swoboda et al., 1994; Puchta et al., 1995; Molinier et al., 2004). Owing to the organization of the reporter regions, these lines allow locus-specific monitoring of the frequency of single-strand annealing (SSA) and synthesis-dependent strand annealing (SDSA) types of HR, respectively (Orel et al., 2003). The plants were germinated and grown on media without (mock) and with 1 μM MMC for 10 days and analyzed for HR events. There were no significant differences (Mann-Whitney U test, P > 0.05) in fresh weight between all genotypes (Supplementary Figure 6), indicating similar number of cells. Under mock conditions, we found on average 2.9 ± 2.2 SSA HR events per B11 wild type (n = 131) plant (Figure 5), while bcp1-1 B11 plants (n = 151) showed 52% less SSA HR events per plant (1.4 ± 1.5). This difference was statistically significantly different (P < 0.001 Mann-Whitney U-test), indicating a possible role of BCP1 in HR independent of exogenous DNA damage. In response to a mild DNA inter-strand crosslinking treatment by MMC, there were on average 34 ± 12 SSA HR events per B11 plant (n = 137 plants) and 21 ± 10.6 per bcp1-1 B11 plant (n = 151 plants), corresponding to a significant (P < 0.001Mann-Whitney U-test) 38% reduction in the mutant.

Figure 5 (A) Schematic model of constructs used to create B11 and IC9C HR reporter lines (Swoboda et al., 1994; Molinier et al., 2004). (B) Loss of BCP1 causes reduced frequency of somatic homologous recombination (HR). Wild type (WT) and bcp1-1 plants carrying genomic substrates for single-strand annealing (B11) and synthesis-dependent strand annealing (IC9C) types of HR were grown on mock and 1 μM containing MMC media for 10 days. Gray dots indicate HR events per plant. The boxplots’ hinges are in the 1st and 3rd quartile, with a marked median. Mean is represented by a cross with a numerical value. Whisker marks show the lowest or highest value within the 1.5 interquartile range below or above hinges. Asterisks represent significant differences in Mann-Whitney U-test *** P < 0.001, NS – not significantly different.
A similar pattern was observed in SDSA HR reporter line IC9C. Also here, we did not find significant differences (Mann-Whitney U-test, P > 0.05) in fresh weight between all genotypes (Supplementary Figure 6), indicating similar number of cells. The IC9C wild type and IC9C bcp1-1 lines showed a similar 0.2 ± 0.46 (n = 153 plants) and 0.28 ± 0.61 (n = 109 plants) SDSA events per plant under mock conditions, respectively. After treatment with 1 µM MMC, IC9C wild type showed an average of 1.59 ± 1.31 (n = 102 plants) SDSA HR events plant, while IC9C bcp1-1 line had 0.78 ± 0.85 (n = 158 plants) SDSA HR events per plant. This is a 50.5% decrease in the number of HR events in the bcp1-1 mutant under mild genotoxic stress. Collectively, this shows that BCP1 is needed for normal levels of SSA and SDSA HR in Arabidopsis and suggests an involvement of BCP1 in the HR repair.
Discussion
In this work, we found a new Arabidopsis protein BCP1 which contains the BRCT5 domain and contributes to DNA damage repair by homologous recombination in a SOG1-dependent manner.
To identify Arabidopsis BRCT5 domain-containing proteins, we performed a homology search using fission yeast SpBrc1 and human HsNSE5. These proteins were selected because they are known to mediate interactions of the conserved SMC5/6 DNA repair complex to chromatin (Li et al., 2012; Räschle et al., 2015). While human NSE5 directly contains the BRCT5 domain, the yeast NSE5 does not, but it binds BRCT5-containing Brc1 protein which targets it to DNA damage sites. The situation in Arabidopsis resembles yeast where none of the currently known SMC5/6 complex subunits harbors a BRCT domain (Yan et al., 2013). Hence, identification of the plant BRCT5 domain-containing proteins might lead to a plant-specific SMC5/6 cofactor mediating interaction with DNA repair complexes and/or chromatin.
Via two BLASTs, first against the moss Physcomitrium patens and then Arabidopsis, we found in total six Arabidopsis BRCT5 domain-containing proteins, including two already known DNA damage repair factors BRCA1 and BARD1. BRCA1 is a well-known tumor suppressor in humans that is evolutionarily conserved also in plants (Trapp et al., 2011). Studies in mammals and Arabidopsis revealed that BRCA1 and BARD1 frequently act as a heterodimer (Wu et al., 1996; Reidt et al., 2006). In Arabidopsis, both BRCA1 and BARD1 are necessary for resistance to DNA damage and also for normal levels of somatic homologous recombination (Reidt et al., 2006). Furthermore, the function of BARD1 seems to go beyond the regulation of genome stability because BARD1 was found to suppress the expression of WUSCHEL1, a master regulator homeobox gene controlling the stem cell pool (Mayer et al., 1998), in the shoot apical meristems and thus contributing to the meristem normal growth and organization during plant development (Han et al., 2008). Besides the established role of these two proteins in plant DNA damage repair, their exact molecular functions, including the binding targets of BRCT5 domains, remain unknown.
The BCP2, BCP3, and BCP4 proteins carry a pair of BRCT domains only at their C-termini. On contrary, BCP1 bears an additional pair of BRCT domains also at the N-terminus. The BRCT5 domain of all Arabidopsis BCPs shows a conserved pattern of specific amino acids with different properties. Furthermore, in silico-based modeling revealed a conserved structure of this domain in plants relative to the fission yeast Brc1. The only non-BRCT domain identified in BCPs was an N-terminally positioned Gcn5-related N-acetyltransferase domain (Uniprot) in BCP2. It implies that BCP2 might contribute to chromatin relaxation and/or transcription. However, none of the BCPs repeated the repertoire of domains in SpBrc1 and/or HsNSE5, suggesting that they are not direct Arabidopsis homologs, and biochemical studies will have to be conducted to explore their potential relationship at the protein-protein interaction level. BCP1 shows possible homologies to the human proteins PTIP and TOPIP1B. However, a significant homology is present only over the BRCT domain regions. Based on this, we conclude that all four identified BCPs represent novel plant-specific BRCT5 domain-containing proteins.
An important step toward the functional characterization of the BCPs was their response to DNA damage. The most promising candidate in DNA damage sensitivity assays was BCP1, while BCP2, BCP3, and BCP4 did not differ significantly from wild-type. However, analysis of additional mutant alleles for at least BCP2 and BCP3 is needed because the alleles tested in this study most likely do not represent loss of function mutants. The bcp2-1 allele may even be a BCP2 overexpressor line. BCP1 loss-of-function mutants were hypersensitive to DNA DSBs caused by bleocin, DNA-inter-strand crosslinks induced by MMC, and two types of DNA-protein crosslinks caused by zebularine and CPT. Hence, BCP1 emerged from our analyses as an important player in DNA repair of multiple types of DNA lesions, possibly through a mainstream DNA repair pathway. The possible role of BCP2, BCP3, and BCP4 in e.g. repair of other types of DNA damage is not excluded and should be a focus of future studies.
We made an exciting observation that BCP1 is transcriptionally upregulated in response to gamma-radiation and MMC treatments and that the activation is SOG1-dependent. SOG1 is a plant-specific transcription factor that is phosphorylated by ATM and ATR kinases and orchestrates downstream responses of the key set of genes involved in the maintenance of genome stability, including cell cycle and homologous recombination repair (Yoshiyama et al., 2013a; Yoshiyama, 2016; Ogita et al., 2018). Two recent studies defined the SOG1 consensus binding motif CTT(N)7AAG and found that SOG1 is physically binding to the cis-regulatory region of BCP1 in Arabidopsis (Bourbousse et al., 2018; Ogita et al., 2018). Surprisingly, we did not find any such a motif in the region upstream of the BCP1 transcription start site which suggests a presence of a non-canonical SOG1 binding motif in the BCP1 promoter.
The absence of BCP1 transcriptional upregulation in the sog1 mutant background also clearly places BCP1 downstream of SOG1 in the same DNA damage repair pathway. Although BCP1 transcription is enhanced by DNA damage, it is not fully dependent on it. This is apparent from the expression of BCP1 promoter in both somatic and floral meristems without any stress. Our analysis suggests that BCP1 is activated to a basal level in SOG1-independent and induced-DNA damage-independent manner. Whether this represents an activation induced by spontaneously occurring DNA damage (e.g. during DNA replication) remains to be studied. In summary, we identify BCP1 as an Arabidopsis BRCT5 domain-containing gene directly transcriptionally controlled by SOG1 during induced DNA damage.
The critical experiment was the analysis of somatic homologous recombination using genetically engineered HR trap lines. This experiment showed a significantly reduced frequency of HR in bcp1 mutant plants, strongly suggesting that BCP1 is needed for normal levels of HR. How BCP1 directly functions in this process is currently unknown. By its N- and C-terminal BRCT domains, it could bind two phosphorylated proteins and this way facilitate HR. Such interactors will be identified in the follow-up research.
In conclusion, out of four uncharacterized Arabidopsis BRCT5 domain-containing proteins, we identified BCP1 as a new Arabidopsis DNA damage repair factor that is directly controlled by SOG1 and ensures normal levels of homologous recombination.
Data availability statement
The datasets presented in this study can be found in online repositories. The names of the repository/repositories and accession number(s) can be found below: https://www.ncbi.nlm.nih.gov/geo/, GSE112773.
Author contributions
AP, JP, JD, and JV conceived and designed the study. JV, FY, and ET performed experiments. All authors analyzed data and interpreted the results. AP and JV wrote the paper. All authors contributed to the article and approved the submitted version.
Funding
This work was supported by a Purkyně Fellowship from the Czech Academy of Sciences and Czech Science Foundation project 22-00871S (all to AP), China Scholarship Council Fellowship (File No. 201604910685), and the Fisher Scholarship from the Palacký University in Olomouc (both to FY). JV, ET, and AP were additionally supported by the ERDF project “Plants as a tool for sustainable global development” (No. CZ.02.1.01/0.0/0.0/16_019/0000827). JJP was funded by the Czech Science Foundation project GA20-05095S.
Acknowledgments
We thank E. Jahnová, H. Tvardíková, and Z. Bursová for excellent technical assistance and Dr. Inna Lermontova for the donation of the pKGWFS7.0 vector and Dr. Julie Law for the FPKM values from DNA damage RNA-seq experiment.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fpls.2022.1023358/full#supplementary-material
References
Alonso, J. M., Stepanova, A. N., Leisse, T. J., Kim, C. J., Chen, H., Shinn, P., et al. (2003). Genome-wide insertional mutagenesis of arabidopsis thaliana. Science 301, 653–657. doi: 10.1126/science.1086391
Altschul, S. F., Gish, W., Miller, W., Myers, E. W., Lipman, D. J. (1990). Basic local alignment search tool. J. Mol. Biol. 215, 403–410. doi: 10.1016/S0022-2836(05)80360-2
Baubec, T., Pecinka, A., Rozhon, W., Mittelsten Scheid, O. (2009). Effective, homogeneous and transient interference with cytosine methylation in plant genomic DNA by zebularine. Plant J. 57, 542–554. doi: 10.1111/j.1365-313X.2008.03699.x
Bork, P., Hofmann, K., Bucher, P., Neuwald, A. F., Altschul, S. F., Koonin, E. V. (1997). A superfamily of conserved domains in DNA damage- responsive cell cycle checkpoint proteins. FASEB J. 11, 68–76. doi: 10.1096/fasebj.11.1.9034168
Bourbousse, C., Vegesna, N., Law, J. A. (2018). SOG1 activator and MYB3R repressors regulate a complex DNA damage network in arabidopsis. Proc. Natl. Acad. Sci. U.S.A. 115, E12453–E12462. doi: 10.1073/pnas.1810582115
Chatterjee, N., Walker, G. C. (2017). Mechanisms of DNA damage, repair, and mutagenesis. Environ. Mol. Mutagen. 58, 235–263. doi: 10.1002/em.22087
Hafner, A., Bulyk, M. L., Jambhekar, A., Lahav, G. (2019). The multiple mechanisms that regulate p53 activity and cell fate. Nat. Rev. Mol. Cell Biol. 20, 199–210. doi: 10.1038/s41580-019-0110-x
Han, P., Li, Q., Zhu, Y.-X. (2008). Mutation of arabidopsis BARD1 causes meristem defects by failing to confine WUSCHEL expression to the organizing center. Plant Cell 20, 1482–1493. doi: 10.1105/tpc.108.058867
Heyer, W.-D., Ehmsen, K. T., Liu, J. (2010). Regulation of homologous recombination in eukaryotes. Annu. Rev. Genet. 44, 113–139. doi: 10.1146/annurev-genet-051710-150955
Hu, Z., Cools, T., De Veylder, L. (2016). Mechanisms used by plants to cope with DNA damage. Annu. Rev. Plant Biol. 67, 439–462. doi: 10.1146/annurev-arplant-043015-111902
Kleinboelting, N., Huep, G., Kloetgen, A., Viehoever, P., Weisshaar, B. (2012). GABI-kat SimpleSearch: new features of the arabidopsis thaliana T-DNA mutant database. Nucleic Acids Res. 40, D1211–D1215. doi: 10.1093/nar/gkr1047
Klepikova, A. V., Kasianov, A. S., Gerasimov, E. S., Logacheva, M. D., Penin, A. A. (2016). A high resolution map of the arabidopsis thaliana developmental transcriptome based on RNA-seq profiling. Plant J. 88, 1058–1070. doi: 10.1111/tpj.13312
Lafarge, S., Montané, M.-H. (2003). Characterization of arabidopsis thaliana ortholog of the human breast cancer susceptibility gene 1: AtBRCA1 , strongly induced by gamma rays. Nucleic Acids Res. 31, 1148–1155. doi: 10.1093/nar/gkg202
Leung, G. P., Lee, L., Schmidt, T. I., Shirahige, K., Kobor, M. S. (2011). Rtt107 is required for recruitment of the SMC5/6 complex to DNA double strand breaks. J. Biol. Chem. 286, 26250–26257. doi: 10.1074/jbc.M111.235200
Li, X., Liu, K., Li, F., Wang, J., Huang, H., Wu, J., et al. (2012). Structure of c-terminal tandem BRCT repeats of Rtt107 protein reveals critical role in interaction with phosphorylated histone H2A during DNA damage repair. J. Biol. Chem. 287, 9137–9146. doi: 10.1074/jbc.M111.311860
Lobet, G., Pagès, L., Draye, X. (2011). A novel image-analysis toolbox enabling quantitative analysis of root system architecture. Plant Physiol. 157, 29–39. doi: 10.1104/pp.111.179895
Manova, V., Gruszka, D. (2015). DNA Damage and repair in plants – from models to crops. Front. Plant Sci. 6. doi: 10.3389/fpls.2015.00885
Mayer, K. F. X., Schoof, H., Haecker, A., Lenhard, M., Jürgens, G., Laux, T. (1998). Role of WUSCHEL in regulating stem cell fate in the arabidopsis shoot meristem. Cell 95, 805–815. doi: 10.1016/S0092-8674(00)81703-1
Menges, M., Hennig, L., Gruissem, W., Murray, J. A. H. (2002). Cell cycle-regulated gene expression inArabidopsis *. J. Biol. Chem. 277, 41987–42002. doi: 10.1074/jbc.M207570200
Molinier, J., Ries, G., Bonhoeffer, S., Hohn, B. (2004). Interchromatid and interhomolog recombination in arabidopsis thaliana. Plant Cell 16, 342–352. doi: 10.1105/tpc.019042
Nisa, M.-U., Huang, Y., Benhamed, M., Raynaud, C. (2019). The plant DNA damage response: signaling pathways leading to growth inhibition and putative role in response to stress conditions. Front. Plant Sci. 10. doi: 10.3389/fpls.2019.00653
Ogita, N., Okushima, Y., Tokizawa, M., Yamamoto, Y. Y., Tanaka, M., Seki, M., et al. (2018). Identifying the target genes of SUPPRESSOR OF GAMMA RESPONSE 1, a master transcription factor controlling DNA damage response in arabidopsis. Plant J. 94, 439–453. doi: 10.1111/tpj.13866
Oravcová, M., Gadaleta, M. C., Nie, M., Reubens, M. C., Limbo, O., Russell, P., et al. (2019)Brc1 promotes the focal accumulation and SUMO ligase activity of Smc5-Smc6 during replication stress. In: Molecular and cellular biology. Available at: https://journals.asm.org/doi/full/10.1128/MCB.00271-18 (Accessed May 11, 2022).
Orel, N., Kyryk, A., Puchta, H. (2003). Different pathways of homologous recombination are used for the repair of double-strand breaks within tandemly arranged sequences in the plant genome. Plant J. 35, 604–612. doi: 10.1046/j.1365-313X.2003.01832.x
Preuss, S. B., Britt, A. B. (2003). A DNA-damage-induced cell cycle checkpoint in arabidopsis. Genetics 164, 323–334. doi: 10.1093/genetics/164.1.323
Prochazkova, K., Finke, A., Tomaštíková, E. D., Filo, J., Bente, H., Dvořák, P., et al. (2022). Zebularine induces enzymatic DNA–protein crosslinks in 45S rDNA heterochromatin of arabidopsis nuclei. Nucleic Acids Res. 50, 244–258. doi: 10.1093/nar/gkab1218
Puchta, H., Swoboda, P., Hohn, B. (1995). Induction of intrachromosomal homologous recombination in whole plants. Plant J. 7, 203–210. doi: 10.1046/j.1365-313X.1995.7020203.x
R Core Team. (2018). R: A Language and Environment for Statistical Computing (Vienna:R Foundation for Statistical Computing). Available at: https://www.R-project.org.
Räschle, M., Smeenk, G., Hansen, R. K., Temu, T., Oka, Y., Hein, M. Y., et al. (2015). Proteomics reveals dynamic assembly of repair complexes during bypass of DNA cross-links. Science 348, 1253671. doi: 10.1126/science.1253671
Razqallah, H. (2008). DNA-Damage repair; the good, the bad, and the ugly. EMBO J. 27, 589–605. doi: 10.1038/emboj.2008.15
Reidt, W., Wurz, R., Wanieck, K., Ha Chu, H., Puchta, H. (2006). A homologue of the breast cancer-associated gene BARD1 is involved in DNA repair in plants. EMBO J. 25, 4326–4337. doi: 10.1038/sj.emboj.7601313
Shultz, R. W., Tatineni, V. M., Hanley-Bowdoin, L., Thompson, W. F. (2007). Genome-wide analysis of the core DNA replication machinery in the higher plants arabidopsis and rice. Plant Physiol. 144, 1697–1714. doi: 10.1104/pp.107.101105
Swoboda, P., Gal, S., Hohn, B., Puchta, H. (1994). Intrachromosomal homologous recombination in whole plants. EMBO J. 13, 484–489. doi: 10.1002/j.1460-2075.1994.tb06283.x
Trapp, O., Seeliger, K., Puchta, H. (2011). Homologs of breast cancer genes in plants. Front. Plant Sci. 2. doi: 10.3389/fpls.2011.00019
Varadi, M., Anyango, S., Deshpande, M., Nair, S., Natassia, C., Yordanova, G., et al. (2022). AlphaFold protein structure database: massively expanding the structural coverage of protein-sequence space with high-accuracy models. Nucleic Acids Res. 50, D439–D444. doi: 10.1093/nar/gkab1061
Wan, B., Hang, L. E., Zhao, X. (2016). Multi-BRCT scaffolds use distinct strategies to support genome maintenance. Cell Cycle 15, 2561–2570. doi: 10.1080/15384101.2016.1218102
Williams, J. S., Williams, R. S., Dovey, C. L., Guenther, G., Tainer, J. A., Russell, P. (2010). gammaH2A binds Brc1 to maintain genome integrity during s-phase. EMBO J. 29, 1136–1148. doi: 10.1038/emboj.2009.413
Wuest, S. E., Vijverberg, K., Schmidt, A., Weiss, M., Gheyselinck, J., Lohr, M., et al. (2010). Arabidopsis female gametophyte gene expression map reveals similarities between plant and animal gametes. Curr. Biol. 20, 506–512. doi: 10.1016/j.cub.2010.01.051
Wu, L. C., Wang, Z. W., Tsan, J. T., Spillman, M. A., Phung, A., Xu, X. L., et al. (1996). Identification of a RING protein that can interact in vivo with the BRCA1 gene product. Nat. Genet. 14, 430–440. doi: 10.1038/ng1296-430
Yan, W., Shao, Z., Li, F., Niu, L., Shi, Y., Teng, M., et al. (2011). Structural basis of γH2AX recognition by human PTIP BRCT5-BRCT6 domains in the DNA damage response pathway. FEBS Lett. 585, 3874–3879. doi: 10.1016/j.febslet.2011.10.045
Yan, S., Wang, W., Marqués, J., Mohan, R., Saleh, A., Durrant, W. E., et al. (2013). Salicylic acid activates DNA damage responses to potentiate plant immunity. Mol. Cell 52, 602–610. doi: 10.1016/j.molcel.2013.09.019
Yoshiyama, K. O. (2016). SOG1: a master regulator of the DNA damage response in plants. Genes Genet. Syst. 90, 209–216. doi: 10.1266/ggs.15-00011
Yoshiyama, K., Conklin, P. A., Huefner, N. D., Britt, A. B. (2009). Suppressor of gamma response 1 (SOG1) encodes a putative transcription factor governing multiple responses to DNA damage. Proc. Natl. Acad. Sci. 106, 12843–12848. doi: 10.1073/pnas.0810304106
Yoshiyama, K., Kobayashi, J., Nobuo, O., Minako, U., Kimura, S., Maki, H., et al. (2013a). ATM-Mediated phosphorylation of SOG1 is essential for the DNA damage response in arabidopsis. EMBO Rep. 14, 817–822. doi: 10.1038/embor.2013.112
Yoshiyama, K. O., Sakaguchi, K., Kimura, S. (2013b). DNA Damage response in plants: conserved and variable response compared to animals. Biology 2, 1338–1356. doi: 10.3390/biology2041338
Yu, X., Chini, C. C. S., He, M., Mer, G., Chen, J. (2003). The BRCT domain is a phospho-protein binding domain. Science 302, 639–642. doi: 10.1126/science.1088753
Keywords: DNA damage repair, genome stability, BRCT domain, BRCT5 domain, homologous recombination, Arabidopsis
Citation: Vladejić J, Yang F, Dvořák Tomaštíková E, Doležel J, Palecek JJ and Pecinka A (2022) Analysis of BRCT5 domain-containing proteins reveals a new component of DNA damage repair in Arabidopsis. Front. Plant Sci. 13:1023358. doi: 10.3389/fpls.2022.1023358
Received: 19 August 2022; Accepted: 14 November 2022;
Published: 12 December 2022.
Edited by:
Inna Lermontova, Leibniz Institute of Plant Genetics and Crop Plant Research (IPK), GermanyReviewed by:
Anne Bagg Britt, University of California, Davis, United StatesCécile Raynaud, UMR9213 Institut des Sciences des Plantes de Paris Saclay (IPS2), France
Copyright © 2022 Vladejić, Yang, Dvořák Tomaštíková, Doležel, Palecek and Pecinka. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Ales Pecinka, cGVjaW5rYUB1ZWIuY2FzLmN6