 Lijin Lin
Lijin Lin Jiangyue Wang2
Jiangyue Wang2 Ming’an Liao
Ming’an Liao Yi Tang
Yi Tang- 1Institute of Pomology and Olericulture, Sichuan Agricultural University, Chengdu, China
- 2College of Horticulture, Sichuan Agricultural University, Chengdu, China
- 3Institute of Sichuan Edible Fungi, Chengdu, China
Soil in most areas of the world is selenium (Se) deficient, which results a low Se content in agricultural products. To improve the fruit tree Se accumulation, the effects of different Artemisia argyi water extract concentrations (0, 100, 200, 300, and 400-fold dilutions) on the growth and Se accumulation of peach seedlings were studied by a pot experiment. A 300- and 400-fold dilution of A. argyi water extract increased the root and shoot biomass (dry weight), leaf chlorophyll a content, superoxide dismutase (SOD) activity, and peroxidase (POD) activity of peach seedlings, but decreased the leaf chlorophyll a/b. Different A. argyi water extract concentrations had no significant effects on peach leaf chlorophyll a content of peach seedlings, but increased the leaf carotenoid content, catalase (CAT) activity, and soluble protein content. Different A. argyi water extract concentrations increased the total Se, inorganic Se, and organic contents in roots and shoots of peach seedlings to some extent. Furthermore, A. argyi water extract concentration exhibited a linear relationship with the root and shoot total Se contents. Compared with the control, the 100-, 200-, 300-, and 400-fold dilutions of A. argyi water extract increased the shoot total Se content by 18.95%, 31.31%, 39.32%, and 51.59%, respectively. Different A. argyi water extract concentrations also increased the leaf Se metabolism-related enzyme activities of peach seedlings, including the activities of adenosine triphosphate sulfurase (ATPS), adenosine 5’-phosphosulfate reductase (APR), and serine acetyltransferase (SAT), as well as selenocysteine methyltransferase (SMT) to some extent. Moreover, correlation and grey relational analyses revealed the root total Se content, CAT activity, and ATPS activity to be closely associated with the total shoot Se content. Therefore, applying A. argyi water extract can thus promote the growth and Se uptake of peach seedlings, and the future study should focus on the application effects of Se uptake in peach fruits.
Introduction
Selenium (Se) is an essential trace element in the human body and is extremely important for human health, having anti-cancer, anti-oxidation, and immunity enhancing functions. Se deficiency in the human body causes diseases such as Keshan disease and Kashin-Beck disease (Zhang et al., 2007; Qu et al., 2010; Méplan and Hesketh, 2012). Most areas in the world are Se-deficient, resulting in an extremely low Se content in agricultural products (Sun et al., 2017). The Se in humans and animals mainly derive from crops, while crop Se mainly derives from the soil; crops can convert inorganic Se into organic Se after being absorbed from the soil (Chen et al., 2017; Natasha et al., 2018). So, crop Se sources are the best option for human Se supplementation, and screening out methods to improve crop (especially fruit) Se uptake are urgently needed.
External Se fertilizer application is mostly used to increase crop Se content, but the continuous use or excessive application of Se fertilizer can easily become toxic for crops (Mostofa et al., 2017). Crop straw returning is an agronomic measure commonly used in agricultural production, which can increase crop yield and improve crop quality, because crop straw contains nitrogen (N), phosphorus (P), and potassium (K), as well as other elements and organic matter, to improve soil fertility (Lu and Xie, 2011; Li et al., 2018; Qin, 2021). Moreover, crop straw can release the allelochemicals during decay, which may produce the allelopathy to affect the growth and nutrient absorption of other crop (Ghnaya et al., 2016). The allelopathy of crop straw also affects the heavy metal accumulation in crops (Gu et al., 2017; Liu et al., 2018a). Under cadmium (Cd) contaminated soil, crop straw decreases the Cd accumulation in other crops by effectively reducing soil available Cd concentration (Wang et al., 2019; Lian et al., 2020). When peanut, rice, and maize straws are returned to the field, different wheat growth effects and Cd accumulation occur. Peanut straw decreases wheat Cd content, while increasing arsenic (As) uptake; rice straw does not significantly affect wheat Cd content, but decreases As uptake; maize straw decreases both Cd and As uptake in wheat (Cao et al., 2019). Other studies on heavy metal hyperaccumulators and accumulators, and fruit trees (Lin et al., 2014; Lin et al., 2015; He et al., 2017) have shown similar effects to wheat. These results may be related to the different allelochemicals released from the various straws, which may activate or complex the heavy metal in soil (Lin et al., 2015; Cao et al., 2019; Wang et al., 2019). For Se uptake, Pterocypsela laciniata straw application promotes grape seedling Se uptake, but inhibits grape seedling growth to a certain extent (Liu et al., 2021), which may be related to the crop straw increasing soil selenite bioavailability by activating soil Se (Wang, 2019a). However, no other reports exist regarding crop straw returning effects on other crop Se uptake. Thus, crop straw returning may affect plant Se uptake, but this requires screening of more crop straw and living crop combinations.
Peach (Prunus persica) is a global distribution fruit tree with delicious fruits. Its main rootstock materials are wild peach (P. persica) or mountain peach (P. davidiana) (Yang et al., 2011). Artemisia argyi is a perennial herb used for medicine, food, and dyes, as well as many other uses. There main components of A. argyi extract are terpenes, flavonoids, volatile oils, and trace elements, which can produce the allelopathy on the other crops (Li et al., 2016). If A. argyi extract is applied to wild peach (rootstock material), its growth and Se uptake may be promoted, and the improvement of Se contents in peach fruits also can improve its commercial value (Zhang, 2011). Therefore, in this experiment, different A. argyi water extract concentration effects on the growth and Se accumulation characteristics of peach seedlings were studied. The aim of this study was to investigate whether A. argyi water extract could promote the growth and Se uptake of peach seedlings, and to determine the best A. argyi water extract concentration for Se-enriched peach production.
Materials and methods
Materials
Fluvo-aquic soil was collected from a farmland near the Chengdu Campus of the Sichuan Agricultural University (30°42′N, 103°51′E). Its basic physicochemical properties have previously been described (Liu et al., 2022).
The shoots of A. argyi were also collected from a farmland near the Chengdu Campus of the Sichuan Agricultural University in May 2021. These shoots were dried in a drying oven at 75°C until constant weight, and cut into pieces about 1 cm in length. The preparation method of A. argyi water extract was according to the boiling method of traditional Chinese medicine. A total of 10 g dried A. argyi pieces were added to 1500 mL distilled water, boiled, and then simmered on a low heat for 30 min. After cooling, the A. argyi water extract was added to a final volume of 1000 mL (the water was evaporated about 600 mL during heating), resulting in an A. argyi:water ratio of 1:100 (w/v); this solution was stored in a -18°C refrigerator until required.
Wild peach seeds were collected from a 5-year-old peach tree on a farm near the Chengdu Campus of the Sichuan Agricultural University in September 2021. The seeds were stored in moist sand until germination. In March 2022, the germinated seeds were sown into plug trays (50 holes) filled with perlite for seedling culturing. The plug trays were irrigated with Hoagland’s nutrient solution every three days. One month later, the peach seedlings were transplanted into pots when they grew to 10 cm in height.
Experimental design
The experiment was conducted at a canopy of the Chengdu Campus of Sichuan Agricultural University from March to May 2022. In March 2022, each plastic pot (21 cm diameter and 20 cm depth) was filled with 3.0 kg of air-dried and crushed soil. Analytically pure Na2SeO3 was added and mixed into the soil to make a final soil Se concentration of 5 mg kg−1 (Hu et al., 2019). Each pot’s soil was watered every day to make sure that its soil moisture content was kept at 80% field capacity. In April 2022, four uniform peach seedlings (10 cm in height) from the plug trays cultured seedlings were transplanted into each pot, and evenly distributed in all four directions. Hereafter, different A. argyi water extract concentrations (0, 100, 200, 300, and 400-fold dilutions) were used to irrigate the pots. Each pot was irrigated 50 mL A. argyi water extract, and each treatment was repeated in triplicate (three pots) using a completely randomized design. Hereafter, pots were irrigated every seven days for a total of four irrigations. The pots were watered with tap water every day to maintain an adequate water supply for plant growth.
Determination of indicators
One month after the initial A. argyi water extract irrigation (May 2022), mature leaves, each at the same position (middle part of the peach stem) were collected to determine the contents of photosynthetic pigments (chlorophyll a, chlorophyll b, and carotenoids), antioxidant enzyme [superoxide dismutase (SOD), catalase (CAT), and peroxidase (POD)] activities, soluble protein content, and Se metabolism-related enzyme [adenosine triphosphate sulfurase (ATPS), adenosine 5’-phosphosulfate reductase (APR), serine acetyltransferase (SAT), and selenocysteine methyltransferase (SMT)] activities. Photosynthetic pigment contents were extracted using the acetone-ethanol extraction method and determined at 663, 645, and 470 nm wavelengths using a spectrophotometer (Summit, Shanghai, China) according to Hao et al. (2004), and the chlorophyll a/b was calculated as the chlorophyll a content/chlorophyll b content. SOD, CAT, and POD activities were determined using the nitrotetrazole chloride reduction, potassium permanganate titration, and guaiacol methods, respectively, as described by Lin et al. (2020) and Hao et al. (2004). The Coomassie brilliant blue method was used for soluble protein content determination according to Hao et al. (2004). The activities of Se metabolism-related enzymes were determined using the enzyme linked immunosorbent assay (ELISA) kits (Shanghai Enzyme Link Biotechnology Co., Ltd., Shanghai, China) following the manufacturer’s instructions. Hereafter, whole plants were harvested and treated following the methods described by Li et al. (2022). Roots and shoots were divided, and their dry weights (biomass) were measured using an electronic balance. The dried samples were finely ground and digested with nitrate and perchloric acids, and reduced in hydrochloric acid to determine total Se content. Total Se content was subsequently measured using hydride generation-atomic fluorescence spectrometry (AFS-9700, Beijing Haiguang Instrument Co., Ltd., Beijing, China), as described by Li et al. (2022). Inorganic Se content was determined using the hydrochloric acid extraction method as described by Li et al. (2022). Organic Se content and translocation factor were determined using the following formulae: organic Se content = total Se content - inorganic Se content (Li et al., 2022); translocation factor (TF) = shoot Se content/root Se content (Rastmanesh et al., 2010).
Statistical analysis
The data were analyzed using the SPSS 20.0.0 software (IBM, Chicago, IL, USA) with three repetitions. Data were normalized and subjected to a homogeneity test, followed by a one-way analysis of variance and a Duncan’s Multiple Range Test (P < 0.05). Moreover, the linear relationship between A. argyi water extract concentration, and root and shoot total Se content was analyzed using regression analysis. Pearson’s correlation was used to determine the relationships among biomass, total Se content, photosynthetic pigment content, antioxidant enzyme activity, soluble protein content, and Se metabolism-related enzyme activities. The grey relational analysis method was used to analyze biomass, root total Se content, photosynthetic pigment content, antioxidant enzyme activity, soluble protein content, and Se metabolism-related enzyme activity relationships with shoot total Se content, as described by Wang (2019b) and Ma et al. (2022).
Results
Peach biomass
Both peach root and shoot biomass increased as the A. argyi water extract fold dilution increased (Figures 1A, B). Compared with the control, the 200-, 300-, and 400-fold dilutions of A. argyi water extract increased root biomass by 9.61%, 15.65%, and 26.34%, respectively, and the 300- and 400-fold dilutions increased shoot biomass by 15.94% and 25.44%, respectively. The other fold dilutions did not significantly affect root or shoot biomass.
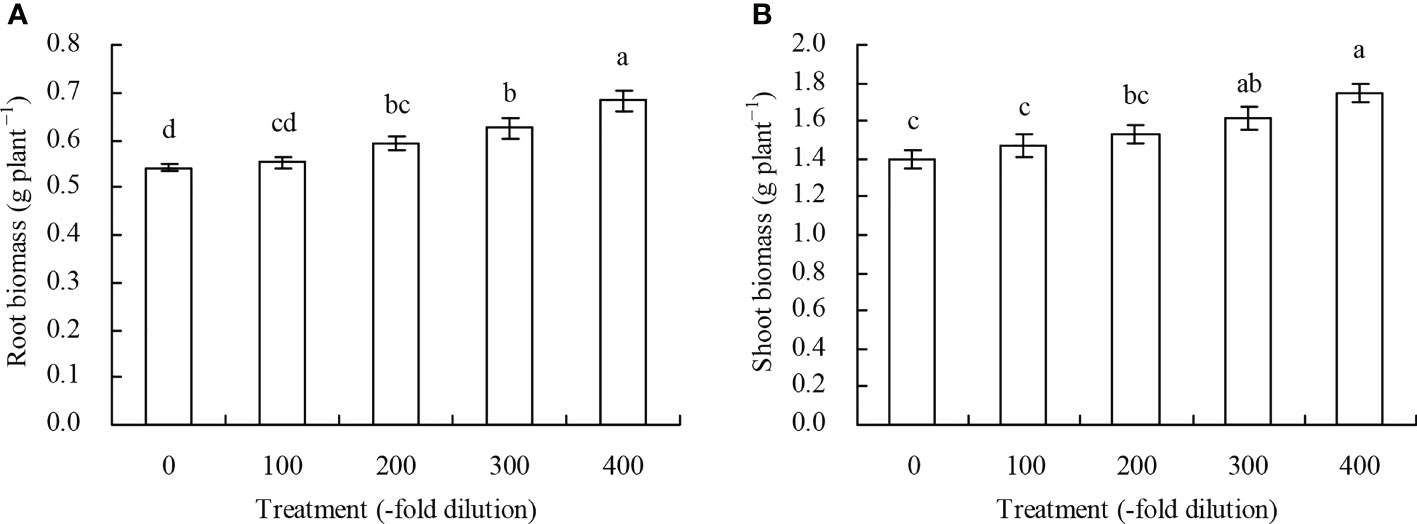
Figure 1 Biomass of peach seedlings. (A): root biomass; (B): shoot biomass. Values are means ± SD of three replicates. Different lowercase letters indicate significant differences among the treatments (Duncan’s Multiple Range Test, P < 0.05).
Photosynthetic pigment content in peach leaves
The different A. argyi water extract concentration did not significantly affect chlorophyll a content in the peach leaves (Figure 2A). The 300- and 400-fold dilution of A. argyi water extract increased peach leaf chlorophyll b content, but decreased chlorophyll a/b, compared with their respective controls, while the 100- and 200-fold dilutions did not significantly affect chlorophyll b content and chlorophyll a/b (Figures 2B, C). However, different A. argyi water extract concentrations increased peach leaf carotenoid content (Figure 2D).
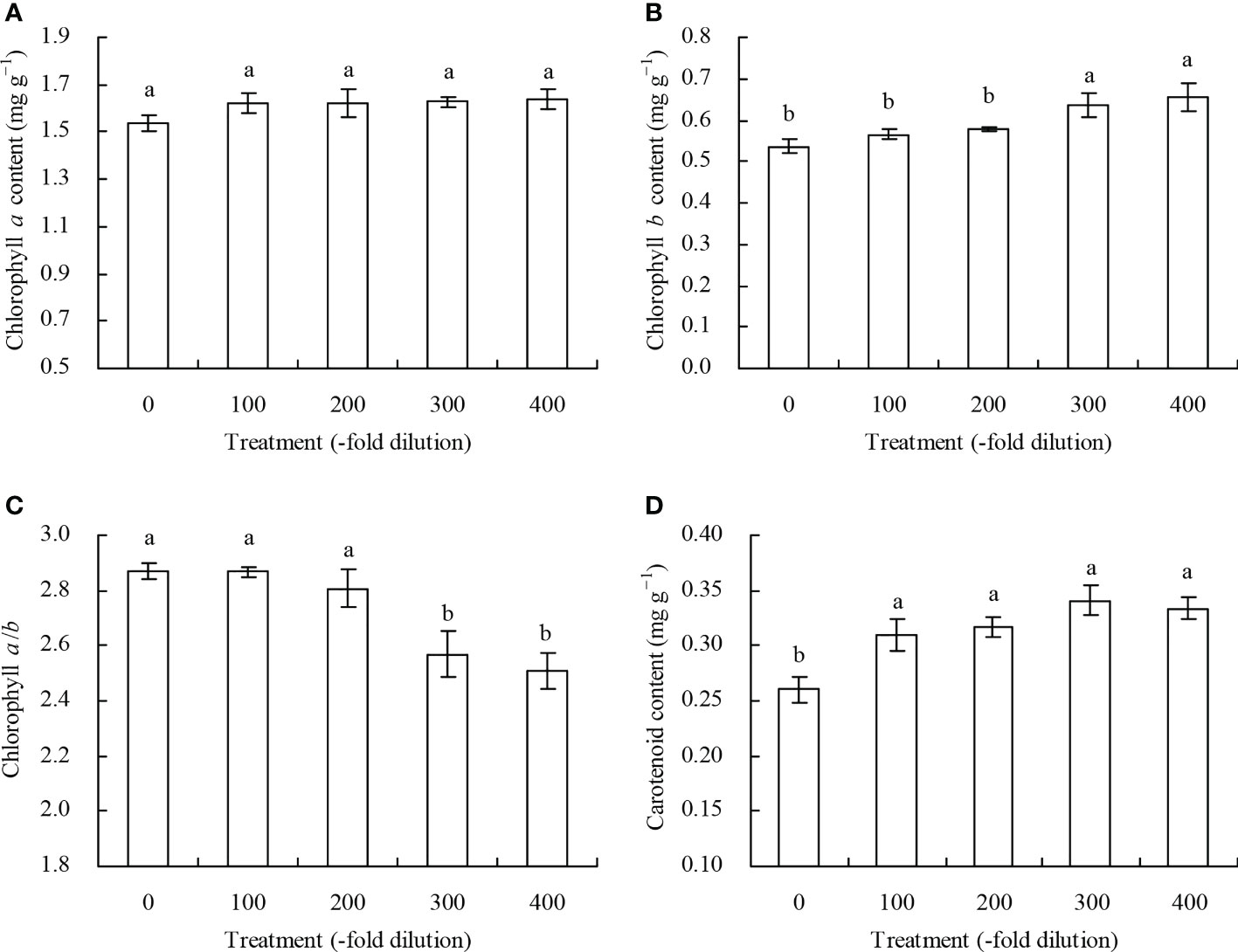
Figure 2 Photosynthetic pigment content in leaves of peach seedlings. (A): chlorophyll a content; (B): chlorophyll b content; (C): chlorophyll a/b; (D): carotenoid content. Values are means ± SD of three replicates. Different lowercase letters indicate significant differences among the treatments (Duncan’s Multiple Range Test, P < 0.05).
Peach leaf antioxidant enzyme activity and soluble protein content
The 100- and 200-fold dilutions of A. argyi water extract did not significantly affect peach leaf SOD activity, while the 300- and 400-fold dilutions increased SOD activity by 8.87% and 27.33%, respectively (Figure 3A). The 100-fold dilution did not significantly affect POD activity, while the 200-, 300-, and 400-fold dilutions increased POD activity by 32.28%, 51.97%, and 127.57%, respectively (Figure 3B). The different A. argyi water extract concentrations increased CAT activity and soluble protein content (Figures 3C, D).
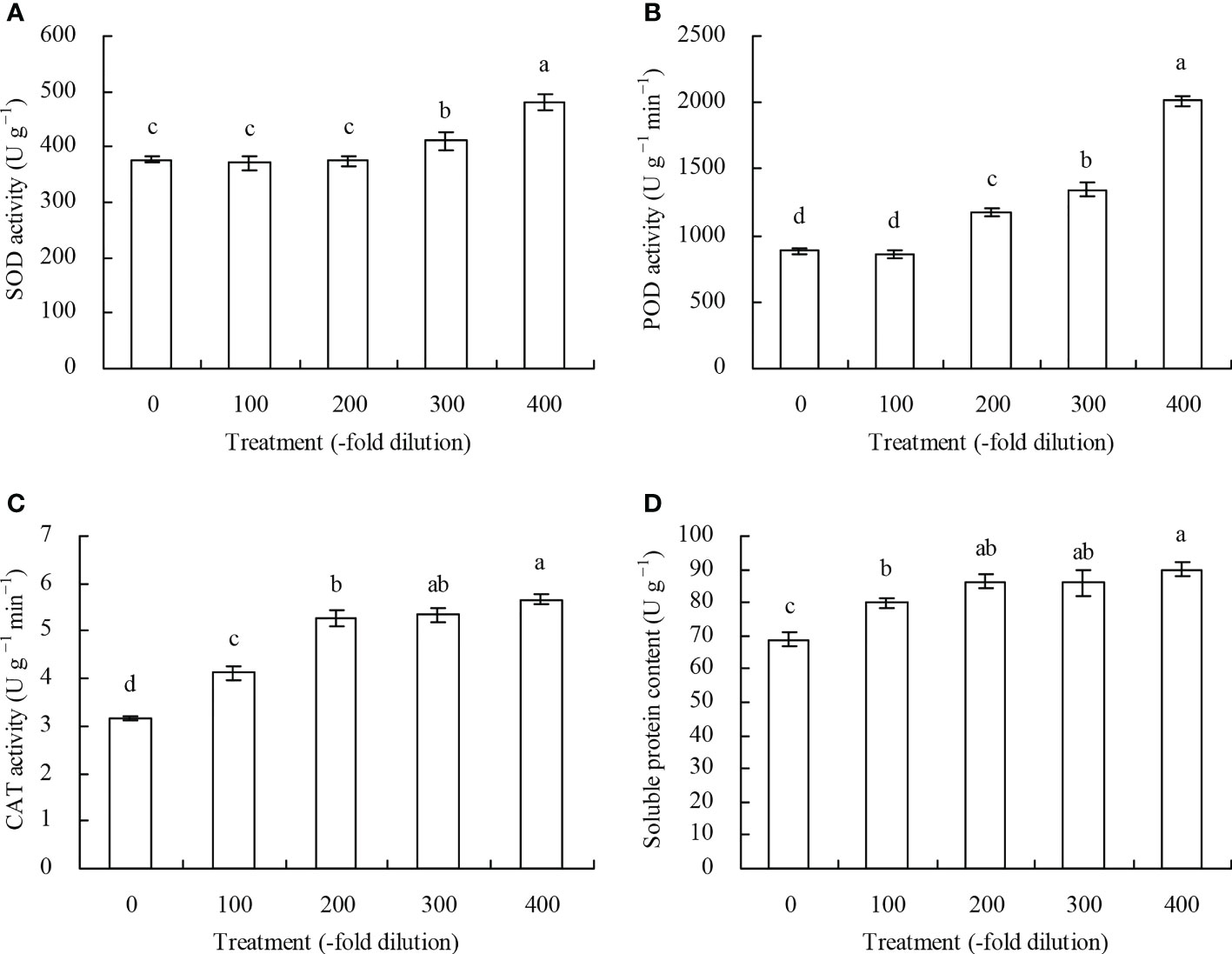
Figure 3 Antioxidant enzyme activity and soluble protein content of leaves of peach seedlings. (A): SOD activity; (B): POD activity; (C): CAT activity; (D): soluble protein content. Values are means ± SD of three replicates. Different lowercase letters indicate significant differences among the treatments (Duncan’s Multiple Range Test, P < 0.05).
Different Se forms and their transport
Peach root and shoot total Se content increased as the A. argyi water extract fold dilutions increased (Figures 4A, B). The 100-, 200-, 300-, and 400-fold dilutions increased root total Se content by 20.22%, 39.78%, 55.22%, and 75.72%, respectively, and increased shoot total Se content by 18.95%, 31.31%, 39.32%, and 51.59%, respectively. Moreover, regression analysis showed that A. argyi water extract concentration was linearly related to both the root total Se content (y = 0.022x + 11.848, R2 = 0.987, P = 0.000) and shoot total Se content (y = 0.002x + 1.900, R2 = 0.967, P = 0.000). Root and shoot inorganic Se content, and root and shoot organic Se content had similar fold dilution changes as A. argyi water extract increased, and were the same as total Se content (Table 1). The different A. argyi water extract concentrations decreased the root inorganic Se proportion, but increased the root organic Se proportion (Figure 5A). However, the different A. argyi water extract concentrations increased the shoot inorganic Se proportion, but decreased the shoot organic Se proportion (Figure 5B). The different A. argyi water extract concentrations decreased the TFs of total Se and organic Se to some extent (Figures 6A, B), while increasing the TF of inorganic Se, compared with their respective controls (Figure 6C).
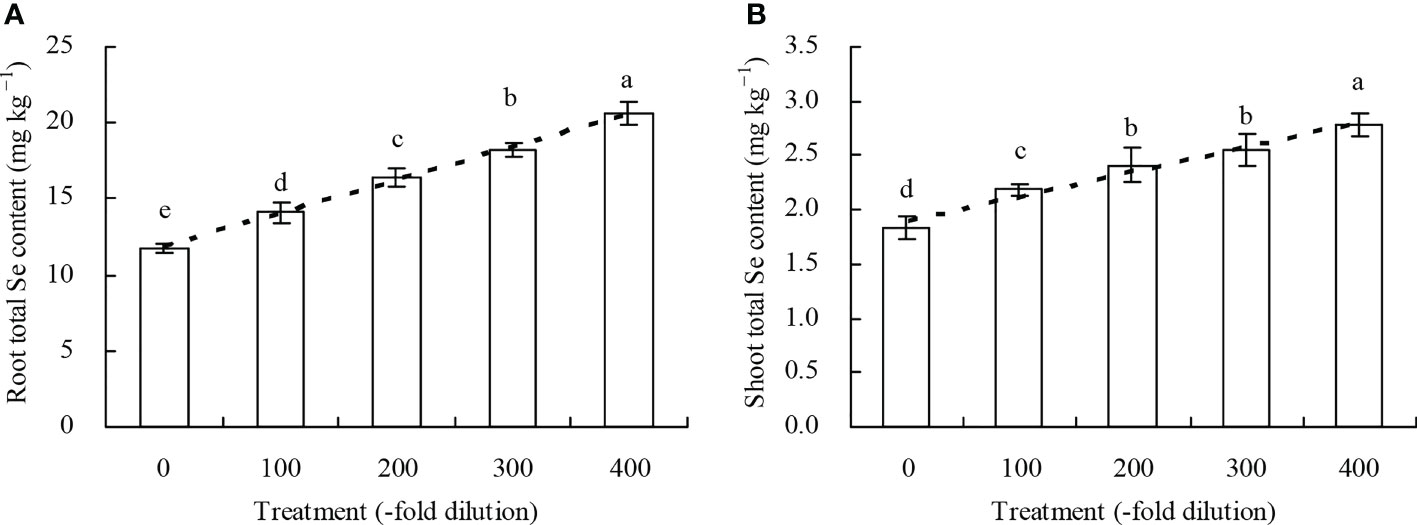
Figure 4 Total Se content in peach seedlings. (A): root total Se content; (B): shoot total Se content. Values are means ± SD of three replicates. Different lowercase letters indicate significant differences among the treatments (Duncan’s Multiple Range Test, P < 0.05).
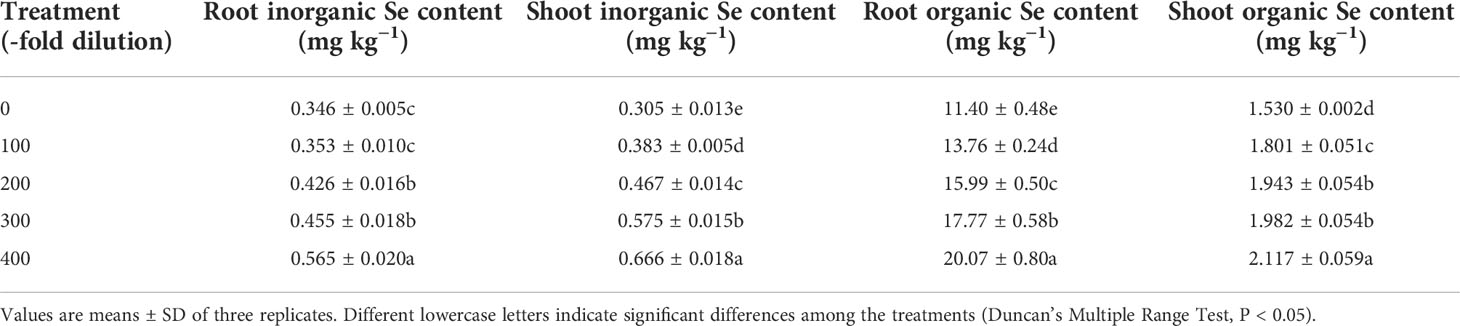
Table 1 Inorganic Se and organic Se contents in peach seedlings.
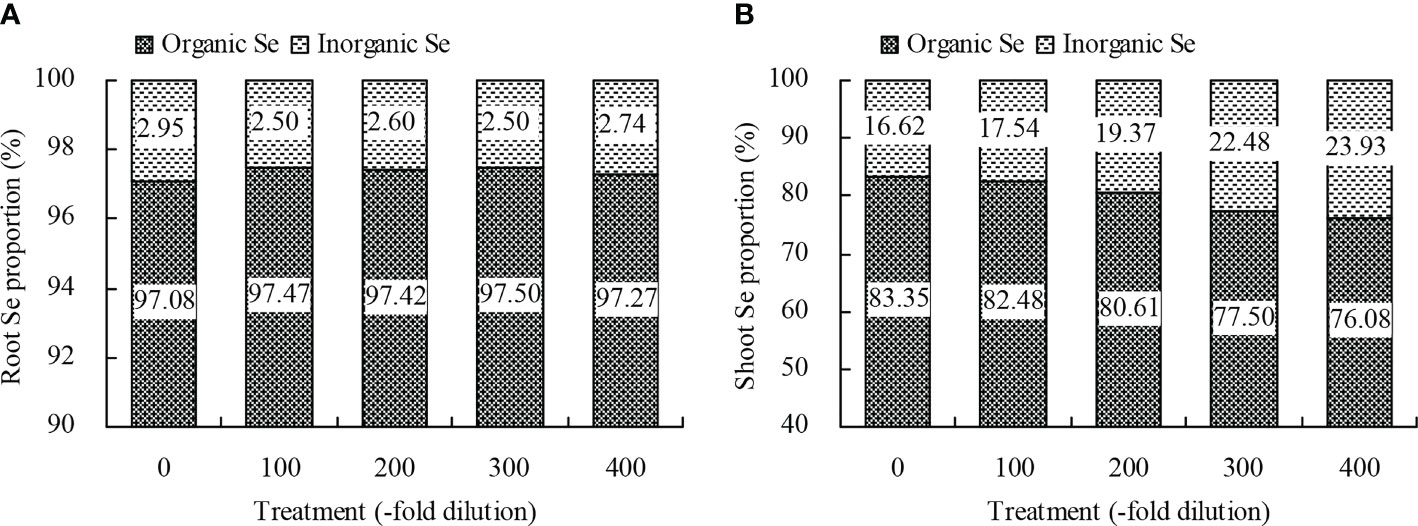
Figure 5 Proportions of organic Se and inorganic Se of peach seedlings. (A): root Se proportion; (B): shoot Se proportion.
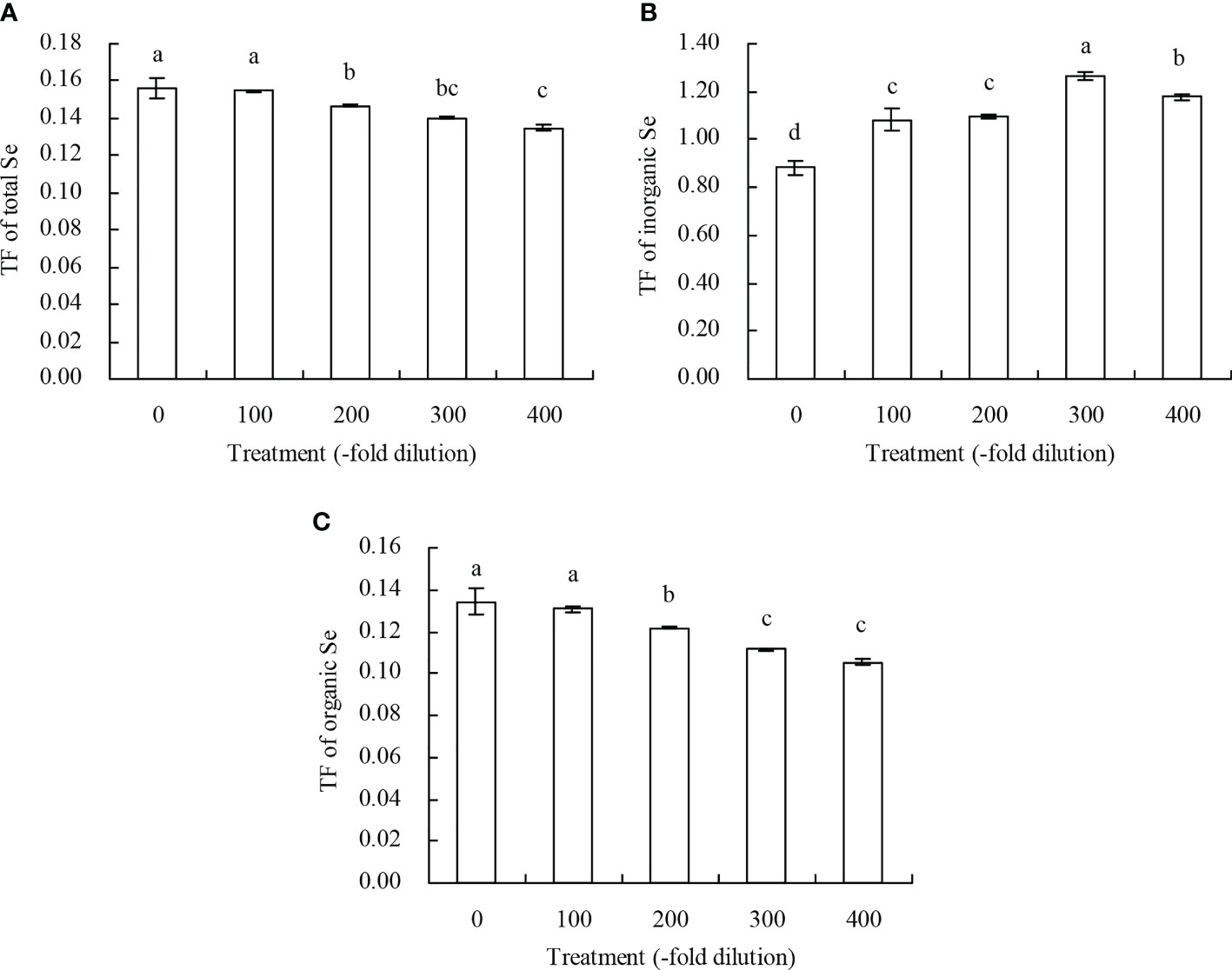
Figure 6 TF of Se of peach seedlings. (A): TF of total Se; (B): TF of inorganic Se; (C): TF of organic Se. Values are means ± SD of three replicates. Different lowercase letters indicate significant differences among the treatments (Duncan’s Multiple Range Test, P < 0.05). Translocation factor (TF) = Se content in shoots/Se content in roots.
Se metabolism-related enzyme activities of peach leaves
Peach leaf Se metabolism-related enzyme (ATPS, APR, SAT, and SMT) activities increased as the A. argyi water extract fold dilutions increased (Table 2). The 100-, 200-, 300-, and 400-fold dilutions of A. argyi water extract increased ATPS activity by 41.67%, 46.67%, 55.09%, and 64.26%, respectively, increased APR activity by 13.88%, 12.31%, 19.09%, and 23.21%, respectively, and increased SAT activity by 16.16%, 13.63%, 31.30%, and 36.05%, respectively. For SMT activity, only the 300- and 400-fold dilutions affected SMT activity, while the 100- and 200-fold dilutions had no significant effects.
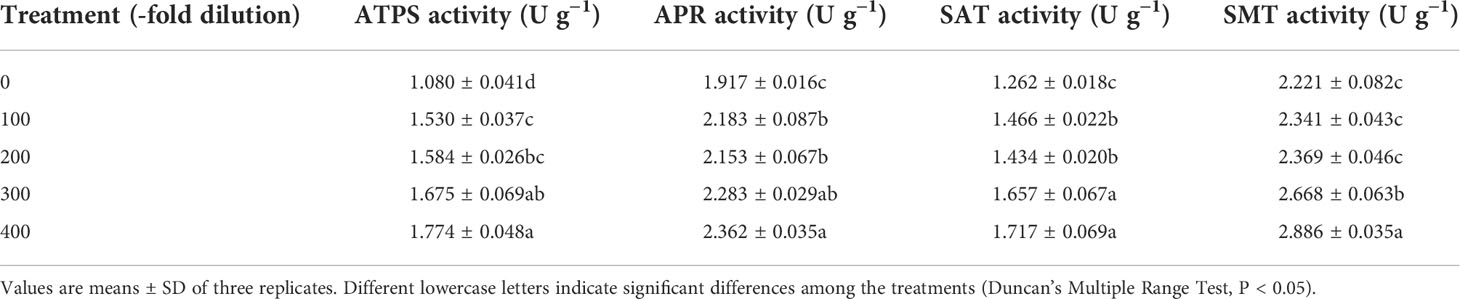
Table 2 Se metabolism-related enzyme activity of leaves of peach seedlings.
Correlation and grey relational analyses
Highly significant (P < 0.01) positive, or significant (0.01 ≤ P < 0.05) positive correlations existed among root biomass, shoot biomass, root total Se content, shoot total Se content, chlorophyll b content, carotenoid content, SOD activity, POD activity, CAT activity, soluble protein content, ATPS activity, APR activity, SAT activity, and SMT activity (Table 3). Chlorophyll a/b was highly significantly (P < 0.01) and negatively correlated with all of the other indicators. The grey relational analysis showed that biomass, root total Se content, photosynthetic pigment content, antioxidant enzyme activity, soluble protein content, and Se metabolism-related enzyme activities were all correlated with shoot total Se content (Figure 7). The top three largest indicators of the grey correlation coefficients were root total Se content, CAT activity, and ATPS activity with shoot total Se content, and were also the top three closest relationships.
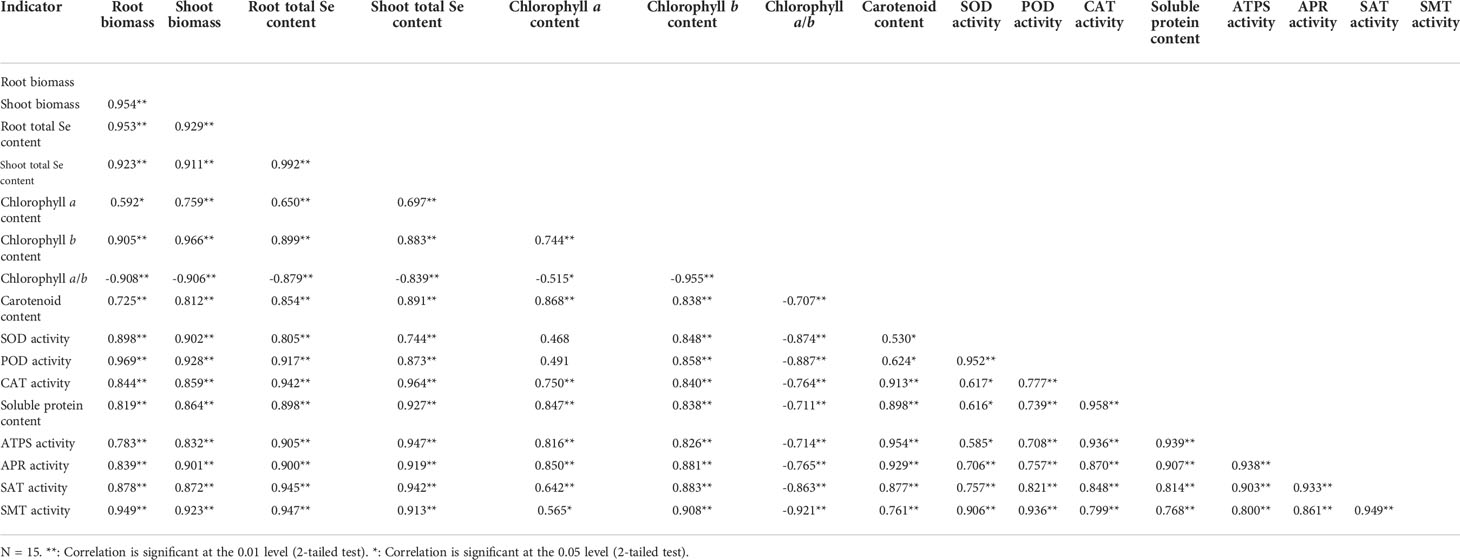
Table 3 Correlations among the biomass, total Se content, photosynthetic pigment content, antioxidant enzyme activity, soluble protein content, and Se metabolism-related enzyme activity.
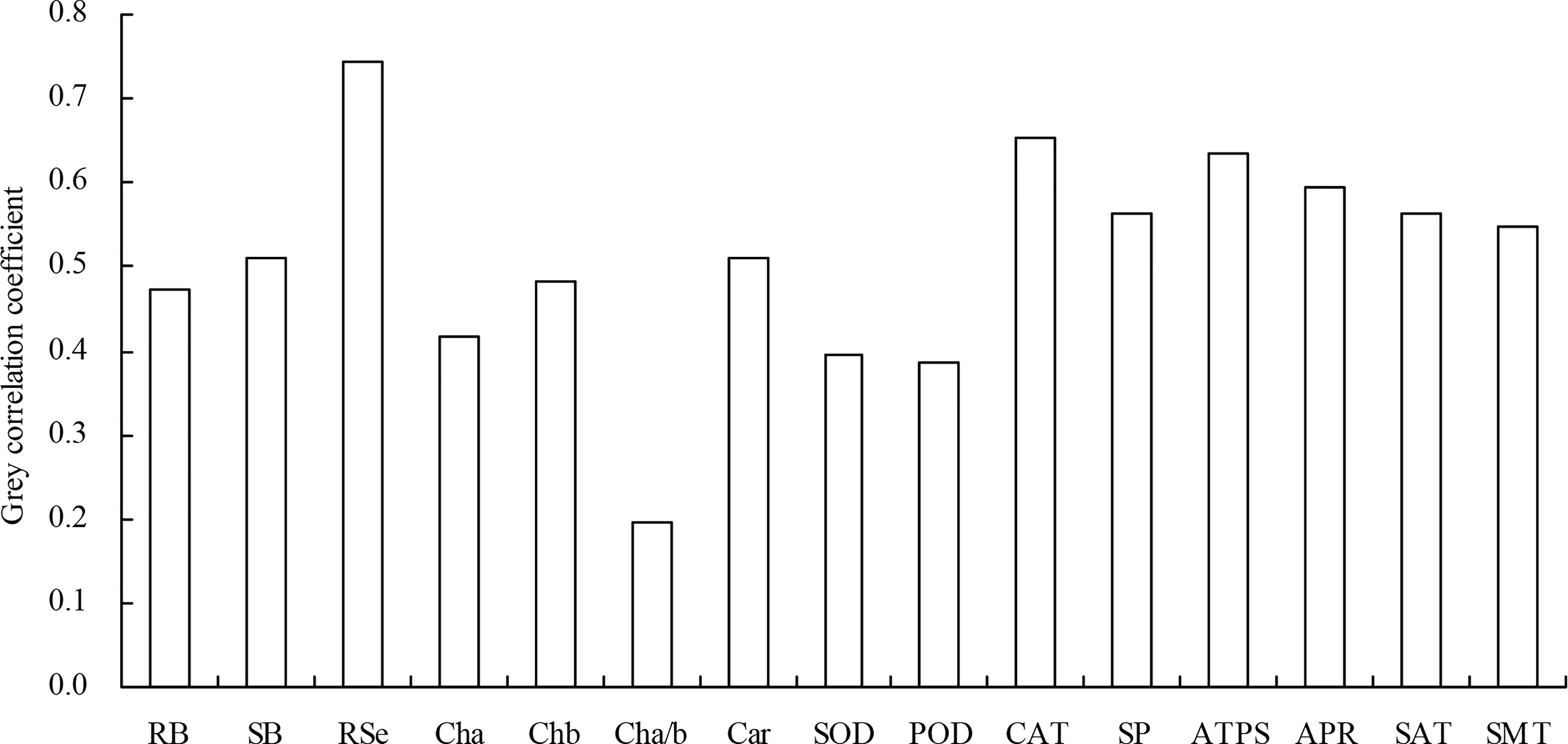
Figure 7 Grey correlation coefficient of the biomass, root total Se content, photosynthetic pigment content, antioxidant enzyme activity, soluble protein content, and Se metabolism-related enzyme activity with the shoot total Se content. RB, root biomass; SB, shoot biomass; RSe, root total Se content; Cha, chlorophyll a content; Chb, chlorophyll b content; Cha/b, chlorophyll a/b; Car, carotenoid content; SOD, SOD activity; POD, POD activity; CAT, CAT activity; SP, soluble protein content; ATPS, ATPS activity; APR, APR activity; SAT, SAT activity; SMT, SMT activity.
Discussion
Crop straw returning not only improves soil nutrient supplies (Zhang et al., 2022), but also releases allelochemicals during decay and decomposition, which has an allelopathy on the other crops (Qi et al., 2015). For crop seeds and seedlings, maize seeds treated with millet straw water extract promoted seedling root and shoot growth (Dong et al., 2016), while garlic straw water extract inhibited pepper and tomato seed germination rates (Liu et al., 2015). Cotton straw extract has an allelopathy promoting effect on lettuce seedling height and fresh weight (Liu et al., 2018b), while rape straw water extract increases rice seedling height and fresh weight (Zou et al., 2022). Moreover, A. argyi water extract inhibits radish and rape seedling growth, and Artemisia lavandulifolia water extract also inhibits Tricium aestivum, Brassica chinensis, Glycine soja, Melilotus suaveolens, Setaria chondrachne, and Solidago decurrens seedling growth (Zhang et al., 2006; Jiang et al., 2009). In this experiment, the 200-, 300-, and 400-fold dilutions of A. argyi water extract increased peach root biomass, while the 300- and 400-fold dilutions increased shoot biomass. The other A. argyi water extract concentrations did not significantly affect peach root or shoot biomass. These results are consistent with previous reports on straw water extracts from other plants (Dong et al., 2016; Liu et al., 2018b; Zou et al., 2022), but contrast with A. argyi water extract effects (Jiang et al., 2009). These results indicate that a low A. argyi water extract concentration (i.e., high fold dilution, such as 300- and 400-fold) could promote peach growth, while high concentrations (low fold dilution) had no effects, which may be related to A. argyi flavonoids (Li et al., 2016). This is because flavonoids can inhibit or promote plant growth at different concentrations (Zong et al., 2017). A. argyi water extract fold dilutions less than 100 may inhibit peach growth. Therefore, since A. argyi water extract inhibited the growth of other crops (Jiang et al., 2009), it may be related to high concentrations. This further suggests that a low A. argyi water extract concentration (high fold dilution) could promote peach growth.
Photosynthetic pigment content reflects plant photosynthetic capacity and affects plant growth (Tholen et al., 2007; Wang et al., 2013). Chlorophyll is related to the rate of photosynthesis and plays a crucial role in crop photosynthesis, and its content affects the absorption, transmission, and conversion of light energy by crop leaves (Scandalios, 1993; Jackson et al., 1996; Wang et al., 2008). Carotenoids are important light-absorbing auxiliary pigments in plant photosynthesis, which can absorb excess light energy and protect photosynthetic organelles (Lu et al., 2022). Different corn straw water extract concentrations are known to inhibit chlorophyll content in wheat seedlings, and this inhibitory effect is strengthened as the concentration increases (Zhen et al., 2015). However, pepper straw promotes the synthesis of photosynthetic pigments in watermelon seedlings (Wang and Hao, 2021), while Artemisia anethifolia water extract also promotes the synthesis of photosynthetic pigments in wheat (Guan, 2014). In this study, different A. argyi water extract concentrations did not significantly affect peach leaf chlorophyll a content, but increased carotenoid content. Only the 300- and 400-fold dilutions of A. argyi water extract increased chlorophyll b content, and decreased peach leaf chlorophyll a/b. These results differ from corn straw water extract effects on wheat seedlings (Zhen et al., 2015), but are, however, consistent with other studies (Wang and Hao, 2021; Guan, 2014). These results also indicate that A. argyi water extract could produce an allelopathic effect that promotes peach photosynthetic pigment synthesis to some extent, which may be related to A. argyi flavonoids content (Li et al., 2016). Therefore, A. argyi water extract had various effects on peach chlorophyll a, chlorophyll b, and carotenoid synthesis. Its mechanism therefore requires further study.
Antioxidant enzymes, including SOD, CAT, and POD, can remove excess reactive oxygen species (ROS) produced in plants (Fu et al., 2007; Li et al., 2007; Gan et al., 2010), thereby reducing or avoiding cell damage caused by ROS (Jiang et al., 2013). Soluble protein is an important osmotic regulating substance, which plays a protective role in cell substances and biofilms, and is one of the indicators of resistance (Shan, 2020). Previous studies showed that mustard straw decomposition treatment increased cowpea SOD, POD, and CAT activities (Chen et al., 2019), while maize straw water extract decreased wheat POD activity (Zhen et al., 2015). Additionally, wheat straw water extract increased the soluble protein content in Salvia miltiorrhiza and Forsythia suspensa (Hua and Li, 2019). In this study, only the 300- and 400-fold dilutions of A. argyi water extract increased peach SOD activity, while the 200-, 300-, and 400-fold dilutions increased POD activity. The different A. argyi water extract concentrations increased peach CAT activity and soluble protein content. These results are consistent with previous studies (Chen et al., 2019; Hua and Li, 2019), indicating that A. argyi water extract could improve peach resistance to some extent. Flavonoids can improve plant oxidative stress cause by the environment (Nakabayashi et al., 2014). Therefore, improving peach resistance may be related to A. argyi flavonoids (Li et al., 2016), which require further study.
Se is a trace element that promotes plant growth (Tang et al., 2022). Plants Se is mainly absorbed from the soil through the roots, and Se exists as various forms in the soil, such as selenate, selenite, elemental selenium, and organic selenium compounds (Wang et al., 2014). Selenite migrates with difficulty in plants, and most of the selenite absorbed by plant roots is directly assimilated into organic selenium within the roots, where it also accumulates (Li et al., 2008; Chen et al., 2014). Plant Se metabolism is a series of reactions completed by several related enzymes and proteins (Ma et al., 2017). Selenite is converted to selenide within the plant body after absorption, and selenide is then converted to selenocysteine by O-acetylserine lyase (OAS-TL) and acetylserine transferase (Wallenberg et al., 2010; Yarmolinsky et al., 2013). Increased selenium content in the environment can promote acetylserine transferase gene expression, which in turn increases selenocysteine conversion (Van Hoewyk et al., 2008). In this study, different A. argyi water extract concentrations increased the various peach Se forms (total, inorganic, and organic Se), Se metabolism-related enzyme (ATPS, APR, SAT, and SMT) activities, and inorganic Se TF to some extent, while decreasing total Se and organic Se TFs to some extent. The different A. argyi water extract concentrations decreased root inorganic Se, but increased the root organic Se, while increasing shoot inorganic Se and decreasing shoot organic Se. These results are consistent with a previous study (Liu et al., 2021), as well as other straw studies on crop nutrient uptake (Li, 2007; Zhang et al., 2015; Huang et al., 2017; Zhou et al., 2019), which indicate that A. argyi water extract could promote Se peach uptake. The reason may be that the allelopathic effects of A. argyi stimulated an increase in plant Se metabolism-related enzyme activities, as well as an increased Se resistance (Nakabayashi et al., 2014). Additionally, correlation and grey relational analyses showed that root total Se content, CAT activity, and ATPS activity were the three most closely associated factors with total shoot Se content, indicating that root total Se content, CAT activity, and ATPS activity significantly promoted peach Se uptake. However, their mechanisms require further study.
Conclusion
The different A. argyi water extract concentrations increased peach carotenoid content, CAT activity, and soluble protein content, and only the 300- and 400-fold dilutions increased biomass, chlorophyll a content, SOD activity, and POD activity. The different A. argyi water extract concentrations also promoted peach total Se, inorganic Se, and organic content uptake to some extent. A. argyi water extract concentration exhibited a linear relationship with root and shoot total Se content by increasing Se metabolism-related enzyme activities. Notably, root total Se content, CAT activity, and ATPS activity were all closely associated with total shoot Se content. So, A. argyi water extract can promote the growth and Se uptake of peach seedlings. Future studies should focus on replicating these findings in peach fields to study the application effects of, and Se uptake in, peach fruits.
Data availability statement
The original contributions presented in the study are included in the article/supplementary material. Further inquiries can be directed to the corresponding author.
Author contributions
LL and ML conceived and designed the research and checked and revised the manuscript. JW, RH, QD, and ZW performed the experiments. XW and YT analyzed the data. LL prepared and wrote the manuscript. All authors contributed to this article and approved the submitted version.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Cao, D., Bai, Y., Li, W. (2019). Effects of different straw returning on wheat growth and heavy metal absorption and accumulation. J. Anhui Agr. Sci. 47 (19), 93–95. doi: 10.3969/j.issn.0517-6611.2019.19.027
Chen, J., Liu, Y., Zeng, C., Pan, L., Lu, S., Lan, X., et al. (2017). Advance on uptake and transformation of selenium from soil to plants. Curr. Biotechnol. 7 (5), 421–427. doi: 10.19586/j.2095-2341.2017.0104
Chen, S., Sun, G., Chen, Z., Chen, F., Zhu, Y. (2014). Progresses on selenium metabolism and interaction with heavy metals in higher plants. Plant Physiol. Jounal. 50 (5), 612–624. doi: 10.13592/j.cnki.ppj.2014.1007
Chen, Y., Zhang, F., Fan, S., Wang, F., Wang, Q., Wu, C. (2019). Effects of crop straw decomposition on the soil properties of the continuous cropping cowpea and the seedling physiological indexes. J. Nucl. Agric. Sci. 3 (7), 1472–1479. doi: 10.11869/j.issn.100-8551.2019.07.1472
Dong, S., Yuan, X., Wen, Y., Guo, P., Hu, C., Nie, L., et al. (2016). Allelopathic effects of the water extracts of foxtail millet straw on maize growth at seedling stage. J. China Agric. University. 21 (8), 35–42. doi: 10.11841/j.issn.1007-4333.2016.08.05
Fu, S., Zong, L., Zhang, L., Sun, J. (2007). Influence of cd, Pb on germination and antioxidant system of Rodix isatis. Seed. 26 (3), 14–17. doi: 10.16590/j.cnki.1001-4705.2007.03.042
Gan, H., Liu, K., Fu, C., Lei, X., Mei, Z., Xiao, Y., et al. (2010). Antioxidative responses to NaCl stress in the leaves of different cultivar of citrus rootstocks. Ecol. Environ. Sci. 19 (1), 183–187. doi: 10.16258/j.cnki.1674-5906.2010.01.039
Ghnaya, A. B., Hamrouni, L., Amri, I., Ahoues, H., Hanana, M., Romane, A. (2016). Study of allelopathic effects of Eucalyptus erythrocorys l. crude extracts against germination and seedling growth of weeds and wheat. Natural Product Res. 30 (18), 2058–2064. doi: 10.1080/14786419.2015.1108973
Guan, C. (2014). Allelopathy of the aqueous extracts from aerial part of artemisia anethifolia (Changchun, China: Northeast Normal University). Master Thesis.
Gu, X., Shi, J., Wu, S., Cheng, L., Li, Y. (2017). Effects of applying crop straw in soil on photosynthetic characteristics of rice under heavy metal stress. J. Xichang Univ. (Natural Sci. Edition). 31 (2), 4–6. doi: 10.16104/j.issn.1673-1891.2017.02.002
Hao, Z., Cang, J., Xu, Z. (2004). Plant physiology experiment. 2nd ed (Harbin, China: Harbin Institute of Technology Press).
He, J., Lin, L., Ma, Q., Liao, M., Wang, X., Liang, D., et al. (2017). Effects of mulching accumulator straw on growth and cadmium accumulation of Cyphomandra betacea seedlings. Environ. Prog. Sustain. Energy. 36 (2), 366–371. doi: 10.1002/ep.12463
Hua, Z., Li, X. (2019). Allelopathy of wheat straw extracts on Salvia miltiorrhiza and Forsythia suspensa. Guizhou Agric. Sci. 47 (5), 110–115. doi: 10.3969/j.issn.1001-3601.2019.05.023
Huang, L., Zhao, Z., Liu, X., Li, M., Zhang, W., Zhang, Z. (2017). Effects of different straw return years on soil humus compositions and enzyme activity of vineyards. Hubei Agric. Sci. 56 (19), 3640–3645. doi: 10.14088/j.cnki.issn0439-8114.2017.19.012
Hu, R., Zhang, Z., Lin, L., Liao, M., Tang, Y., Liang, D., et al. (2019). Intercropping with hyperaccumulator plants decreases the cadmium accumulation in grape seedlings. Acta Agricult. Scandinavica Section B-Soil Plant Science. 69 (4), 304–310. doi: 10.1080/09064710.2018.1564786
Jackson, R. B., Canadell, J., Ehleringer, J. R., Mooney, H. A., Sala, O. E., Schulze, E. D. (1996). A global analysis of root distributions for terrestrial biomes. Oecologia. 108, 389–411. doi: 10.1007/BF00333714
Jiang, C., Shen, X., Zhou, Q., Li, C. (2013). The effect of Pb-cd compound stress on antioxidant enzyme system of three herbaceous flowers. Chin. Agric. Sci. Bulletin. 29 (28), 134–137. doi: 10.3969/j.issn.1000-6850.2013.28.026
Jiang, G., Yang, D., Huang, Z., Lin, H. (2009). Effect of allelopathy of some plants water extract on seedling growth of Rrassica chinensis l. and Raphanus sativus l. J. Changjiang Vegetables. 26 (24), 71–73. doi: 10.3865/j.issn.1001-3547.2009.24.026
Li, F. (2007). Patterns of nutrient release from straws returned to the field and the allelopathic effect of rice straw (Ya’an, China: Sichuan Agricultural University). Master Thesis.
Lian, H., Li, H., Lin, L. (2020). Effects of mulching with Solanum photeinocarpum straw on cadmium accumulation of Cyphomandra beracea seedlings. J. Henan Agric. Sci. 49 (2), 111–115. doi: 10.15933/j.cnki.1004-3268.2020.02.014
Li, S., Cheng, Z., Yang, X., Zhu, Y., Li, Y. (2007). Physiological response of Trichosanthes kirilowii seedlings to cd and zn combined pollution in soil. Acta Botanica Boreali-Occidentalia Sinica. 27 (6), 1191–1196. doi: 10.3321/j.issn:1000-4025.2007.06.021
Li, Z., Fan, R., Peng, X., Shu, J., Liu, L., Wang, J., et al. (2022). Salicylic acid alleviates selenium stress and promotes selenium uptake of grapevine. Physiol. Mol. Biol. Plants. 28, 625–635. doi: 10.1007/S12298-022-01169-5
Li, X., Han, J., Wu, W., Zhang, J., Zhao, Y., Feng, H. (2018). Effects of different straw returning methods on soil microbial biomass carbon, nitrogen and winter wheat yield in guanzhong plain. J. Soil Water Conserv. 32 (4), 170–176. doi: 10.13870/j.cnki.stbcxb.2018.04.027
Li, Z., Lv, J., Zhang, L., Zhu, H. (2016). Chemical constituents and pharmacology activities of Artemisia argyi: research advances. J. Int. Pharm. Res. 43 (6), 1059–1066. doi: 10.13220/j.cnki.jipr.2016.06.007
Li, H., McGrath, S. P., Zhao, F. J. (2008). Selenium uptake, translocation and speciation in wheat supplied with selenate or selenite. New Phytologist. 178 (1), 92–102. doi: 10.1111/j.1469-8137.2007.02343.x
Lin, L., Liao, M., Ren, Y., Luo, L., Zhang, X., Yang, D., et al. (2014). Effects of mulching tolerant plant straw on soil surface on growth and cadmium accumulation of Galinsoga parviflora. PLos One 9 (12), e114957. doi: 10.1371/journal.pone.0114957
Lin, L., Wu, C., Jiang, W., Liao, M., Tang, Y., Wang, J., et al. (2020). Grafting increases cadmium accumulation in the post-grafting generations of the potential cadmium-hyperaccumulator Solanum photeinocarpum. Chem. Ecology. 36 (7), 685–704. doi: 10.1080/02757540.2020.1760853
Lin, L., Yang, D., Tang, F., Luo, L., Liao, M., Yuan, L. (2015). Effects of applying accumulator straw in soil on cadmium accumulation of Capsella bursa-pastoris. Chin. J. Soil Science. 46 (2), 483–488. doi: 10.19336/j.cnki.trtb.2015.02.035
Liu, P., Huang, K., Dai, J., Li, R., Wang, J., Cheng, Q., et al. (2021). Effects of applying Pterocypsela laciniata straw on growth and selenium accumulation of grape seedlings. J. Hunan Agric. Univ. (Natural Sciences). 47 (1), 35–39. doi: 10.13331/j.cnki.jhau.2021.01.006
Liu, Y., Li, H., Xiao, Y., Xu, X., Lin, L., Liao, M., et al. (2022). Exogenous melatonin treatments promote the growth and nutrient uptake of peach (Prunus davidiana) seedlings. J. Plant Nutr. 45 (15), 2292–2302. doi: 10.1080/01904167.2022.2063737
Liu, Y., Xi, K., Wang, H., Wang, M., Wu, L. (2018b). Allelopathy effect of cotton straw extracts on lettuce. J. Shanxi Agric. Sci. 46 (10), 1643–1645. doi: 10.3969/j.issn.1002-2481.2018.10.15
Liu, S., Xu, J., Liu, Q., Wei, H. (2015). Study of allelopathy on straw aqueous extracts of garlic (Allium sativum l.). Northern Horticulture. 39 (9), 10–13. doi: 10.11937/bfyy.201509003
Liu, L., Yue, F., Li, J., Wang, Y. (2018a). Effects of stalk-derived biochar and chicken manure on the growth and physiological characters of maize under lead stress. J. Soil Water Conserv. 32 (4), 262–267. doi: 10.13870/j.cnki.stbcxb.2018.04.041
Lu, C., Gao, Y., Huang, H., Dai, S. (2022). Carotenoid metabolism and regulation in plants. Acta Hortic. Sinica. doi: 10.16420/j.issn.0513-353x.2021-0531
Lu, X., Xie, Y. (2011). Soil and fertilizer science. 2nd ed (Beijing, China: China Agricultural University Press).
Ma, Q., Fasih, U. H., Muhammad, F., Muhammad, A., Noman, S., Wu, J., et al. (2022). Selenium treated foliage and biochar treated soil for improved lettuce (Lactuca sativa l.) growth in cd-polluted soil. J. Cleaner Prod. 335, 130267. doi: 10.1016/J.JCLEPRO.2021.130267
Ma, Y., Jia, G., Zhang, S., Yao, Q., Hao, H., Xu, Z. (2017). Effect of sulfur and selenium combination on uptake and key metabolism enzyme activity of sulfur and selenium in flue cured tobacco. J. Agric. Sci. Technology. 19 (3), 17–25. doi: 10.13304/j.nykjdb.2016.386
Méplan, C., Hesketh, J. (2012). The influence of selenium and selenoprotein gene variants on colorectal cancer risk. Mutagenesis 27 (2), 177–186. doi: 10.1093/mutage/ger058
Mostofa, M. G., Hossain, M. A., Siddiqui, M. N., Fujita, M., Tran, L. S. P. (2017). Phenotypical, physiological and biochemical analyses provide insight into selenium-induced phytotoxicity in rice plants. Chemosphere. 178, 212–223. doi: 10.1016/j.chemosphere.2017.03.046
Nakabayashi, R., Yonekura-Sakakibara, K., Urano, K., Suzuki, M., Yamada, Y., Nishizawa, T., et al. (2014). Enhancement of oxidative and drought tolerance in arabidopsis by over accumulation of antioxidant flavonoids. Plant J. 77 (3), 367–379. doi: 10.1111/tpj.12388
Natasha, Shahid, M., Niazi, N. K., Khalid, S., Murtaza, B., Bibi, I., et al. (2018). A critical review of selenium biogeochemical behavior in soil-plant system with an inference to human health. Environ. Pollution. 234, 915–934. doi: 10.1016/j.envpol.2017.12.019
Qin, T. (2021). Effects of pepper straw returning amount on rhizosphere soil microenvironment and quality of watermelon. J. Henan Agric. Sci. 50 (9), 79–86. doi: 10.15933/j.cnki.1004-3268.2021.09.010
Qi, Y., Zhen, W., Li, H. (2015). Allelopathy of decomposed maize straw products on three soil born diseases of wheat and the analysis by GC-MS. J. Integr. Agricult. 14 (1), 88–97. doi: 10.1016/S2095-3119(14)60795-4
Qu, L., Yang, S., Zhuo, S., Wang, H., Lu, X. (2010). Preliminary discussion on the effects of microelement selenium. Chin. Agric. Sci. Bulletin. 26 (7), 94–97.
Rastmanesh, F., Moore, F., Keshavarzi, B. (2010). Speciation and phytoavailability of heavy metals in contaminated soils in sarcheshmeh area, kerman province, Iran. Bull. Environ. Contamination Toxicol. 85, 515–519. doi: 10.1007/s00128-010-0149-z
Scandalios, J. G. (1993). Oxygen stress and superoxide dismutase. Plant Physiol. 101, 7–12. doi: 10.1104/pp.101.1.7
Shan, X. (2020). Effects of heat preservation and water treatment on physiological and biochemical traits of zoysia japonica steud. under winter stress (Beijing, China: Beijing Forestry University). Master Thesis.
Sun, G., Li, Y., Li, G., Chen, Z., Zhu, Y. (2017). Climatic causes of the selenium-deficient soil belt in China. Curr. Biotechnol. 7 (5), 387–394. doi: 10.19586/j.2095-2341.2017.0061
Tang, M., Yang, K., Zhang, S., Chen, L., Liu, Y., Zhang, X., et al. (2022). Effects of selenium on the activities of antioxidant protective enzymes and fruit quality of walnut. J. Nanjing Forestry Univ. (Natural Sci. Edition). 46 (5), 127–134. doi: 10.12302/J.ISSN.1000-2006.202012032
Tholen, D., Pons, T. L., Voesenek, L. A. C. J., Hendrik, P. (2007). Ethylene insensitivity results in down-regulation of RUBISCO expression and photosynthetic capacity in tobacco. Plant Physiol. 144 (3), 1305–1315. doi: 10.1104/pp.107.099762
Van Hoewyk, D., Takahashi, H., Inoue, E., Hess, A., Tamaoki, M., Pilon-Smits, E. A. H. (2008). Transcriptome analyses give insights into selenium-stress responses and selenium tolerance mechanisms in Arabidopsis. Physiol. Plant 132 (2), 236–253. doi: 10.1111/j.1399-3054.2007.01002.x
Wallenberg, M., Olm, E., Hebert, C., Björnstedt, M., Fernandes, A. P. (2010). Selenium compounds are substrates for glutaredoxins: a novel pathway for selenium metabolism and a potential mechanism for selenium-mediated cytotoxicity. Biochem. J. 429 (1), 85–93. doi: 10.1042/BJ20100368
Wang, D. (2019a). Effect of straw amendment on selenium bioavailability and the mechanisms in soil (Yangling, China: Northwestern Agriculture and Forestry University). Doctor Thesis.
Wang, X. (2019b). Application of grey relation analysis theory to choose high reliability of the network node. J. Physics: Conf. Series. 1237, 32056. doi: 10.1088/1742-6596/1237/3/032056
Wang, X., Chen, S., Luo, Z., Huang, Q., Qiao, Y., Sun, H., et al. (2014). Mechanisms of selenium uptake, translocation and chemical speciation transformation in plants. J. Agric. Resour. Environment. 31 (6), 539–544. doi: 10.13254/j.jare.2014.0250
Wang, X., Hao, S. (2021). Effect of pepper straw composite matrix on watermelon seedling quality. China Cucurbits Vegetables. 34 (3), 31–35. doi: 10.16861/j.cnki.zggc.2021.0055
Wang, J., Li, H., Liu, X., Wang, J., Liu, L., Wang, T., et al. (2019). Effects of accumulator plant straw mulching on cadmium accumulation in grape seedlings. J. Gansu Agr. Univ. 54 (4), 92–99. doi: 10.13432/j.cnki.jgsau.2019.04.013
Wang, W., Song, T., Wang, Y., Dong, X., Chai, J., Xue, Y. (2013). Correlation analysis on chlorophyll content and spad value in tomato leaves. Northern Horticulture. 37 (23), 12–15. doi: 10.3969/j.issn.1673-6257.2008.06.006
Wang, S., Yang, J., Han, X., Liu, X., Zhan, X., Liu, S. (2008). Effect of fertilizer application on photosynthetic traits of spring maize. Soil Fertilizer Sci. China. 45 (6), 23–27. doi: 10.3969/j.issn.1673-6257.2008.06.006
Yang, J., Liu, L., Li, X. (2011). Current status and prospects of peach and nectarine production of import and export trade in China. Agric. Outlook. 7 (3), 48–52.
Yarmolinsky, D., Brychkova, G., Fluhr, R., Sagi, M. (2013). Sulfite reductase protects plants against sulfite toxicity. Plant Physiol. 161 (2), 725–743. doi: 10.1104/pp.112.207712
Zhang, H. (2011). Effects of selenium to physiological indexes of peach, jujube and strawberry (Beijing, China: Beijing Forestry University). Doctor Thesis.
Zhang, J., Chen, Q., Liu, W., Li, Q., Gao, X., Ma, J. (2022). Research progress on the effect of straw returning on soil and crop growth in saline-alkali land. Jiangsu Agric. Sci. 50 (11), 13–22. doi: 10.15889/j.issn.1002-1302.2022.11.002
Zhang, G., Liu, R., Yang, C., Yang, F. (2015). Allelopathic effects of cotton straw extract on seed germination and seedling growth of wheat. J. Triticeae Crops. 35 (4), 555–562. doi: 10.7606/j.issn.1009-1041.2015.04.17
Zhang, Z., Wu, Z., Wu, S., Mu, X. (2007). Research progress of essential trace element selenium. J. Anim. Sci. Veterinary Med. 26 (1), 44–46. doi: 10.3969/j.issn.1001-2443.2006.06.017
Zhang, Z., Zhang, X., Liu, H., Shao, J., Yang, K., Zhang, X. (2006). Studies on the allelopathic effects of Artemisia lavandulaefolia. J. Anhui Normal Univ. (Natural Science). 29 (6), 579–581.
Zhen, X., Qian, L., Li, X. (2015). Effects of maize straw water extract on seed germination and seedling growth of three kinds of wheat. Jiangsu Agric. Sci. 43 (7), 93–95.
Zhou, X., Zhu, D., Wang, S., Shao, W., Zhnag, W. (2019). Effects of straw returning methods on rape growth and nutrient absorption. China Agric. Technol. Extension. 35 (S1), 29–31.
Zong, C., Su, X., Wang, Y. (2017). Application of flavonoids in agricultural production. Modern Agric. Sci. Technology. 46 (5), 120–122. doi: 10.3969/j.issn.1007-5739.2017.05.080
Keywords: allelopathy, fruit tree, plant physiology, selenium transportation, growth
Citation: Lin L, Wang J, Liao M, Hu R, Deng Q, Wang Z, Wang X and Tang Y (2022) Artemisia argyi water extract promotes selenium uptake of peach seedlings. Front. Plant Sci. 13:1014454. doi: 10.3389/fpls.2022.1014454
Received: 08 August 2022; Accepted: 27 October 2022;
Published: 10 November 2022.
Edited by:
Massimo Zacchini, National Research Council (CNR), ItalyReviewed by:
Shahbaz Khan, National Agricultural Research Center, PakistanMurat Dikilitas, Harran University, Turkey
Copyright © 2022 Lin, Wang, Liao, Hu, Deng, Wang, Wang and Tang. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Ming’;an Liao, bG1hbkBzaWNhdS5lZHUuY24=