 Nardana Esmaeili
Nardana Esmaeili Guoxin Shen
Guoxin Shen Hong Zhang
Hong Zhang
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
REVIEW article
Front. Plant Sci. , 21 September 2022
Sec. Plant Abiotic Stress
Volume 13 - 2022 | https://doi.org/10.3389/fpls.2022.1011985
This article is part of the Research Topic Molecular Basis of Stress Resistant Signal Transduction in Plants: A Biotechnological Intervention to Develop Climate-resilient Crops View all 8 articles
Abiotic stresses are major limiting factors that pose severe threats to agricultural production. Conventional breeding has significantly improved crop productivity in the last century, but traditional breeding has reached its maximum capacity due to the multigenic nature of abiotic stresses. Alternatively, biotechnological approaches could provide new opportunities for producing crops that can adapt to the fast-changing environment and still produce high yields under severe environmental stress conditions. Many stress-related genes have been identified and manipulated to generate stress-tolerant plants in the past decades, which could lead to further increase in food production in most countries of the world. This review focuses on the recent progress in using transgenic technology and gene editing technology to improve abiotic stress tolerance in plants, and highlights the potential of using genetic engineering to secure food and fiber supply in a world with an increasing population yet decreasing land and water availability for food production and fast-changing climate that will be largely hostile to agriculture.
About 40 years ago, Boyer wrote that the negative impacts of environmental factors could reduce crop yield by around 70%, which would be a disaster for this planet (Boyer, 1982). He also proposed exploring crops’ genetic potential to improve their yields. The adverse effects of abiotic stresses such as heat, drought, and salinity that are accelerated by climate change and global warming are becoming serious threats to today’s world (Esmaeili et al., 2019). It is projected that by mid-21st century the global temperature will increase by around 4 °C above 20th century, which will pose a great risk to global food security (IPCC, 2014). According to a new report from United Nations the world population is expected to reach 9.8 billion by 2050 and 11.2 billion by 2100 (World population prospects: the 2017 revision, UN Department of Economic and Social Affairs). To feed the growing population on earth, an increase of 44 million metric tons of grains per year is required (Tester and Langridge, 2010). Reports show that about 10% of arable land can be classified as stress-free zones, indicating that crops growing in the remaining 90% of arable land are facing some types of environmental stresses (Dita et al., 2006), with some severely reducing agricultural productivity annually. Many crop improvement strategies such as conventional breeding, tissue culture, chemical priming, and genetic engineering have been deployed to overcome the threats posed by abiotic stresses (Kumar et al., 2020; Rivero et al., 2022).
Plants are subjected to various stresses due to their sessile nature. Thus, plants have evolved several strategies and elaborate mechanisms to perceive, respond, and adapt to adverse environments. Plant response to unfavorable environments is manifested by triggering molecular networks, including signal transduction, up-regulation of stress-related genes, and production of proteins and metabolites that help plants to handle adverse conditions. In many cases, plants show a similar response to different environmental stresses. For instance, plant responses to salt, drought, and cold stresses share similar genes that are triggered by these stresses (Chinnusamy et al., 2007). The polygenic nature of plant response to abiotic stresses makes plant improvement very difficult (Dita et al., 2006). Despite numerous efforts in studying the underlying mechanism of plant response to environmental stresses, the plant stress response is still not adequately understood (Reguera et al., 2012). In recent years the “omics” approach (e.g., proteomics, genomics, and metabolomics) assisted scientists in unraveling the signaling pathways that regulate plant response to stresses, which could result in a large gain in crops productivity (Van Emon, 2016). Using model plants such as Arabidopsis thaliana provided a fundamental platform in the plant biotechnology arena. Since most of these studies aiming at improving plant stress tolerance were conducted with the model plants and remained in the model systems, a very limited number of transgenic crops were created and tested in the field, and only a few transgenic crops were released commercially (Umezawa et al., 2006). Most studies with model plants were conducted in the lab under artificial conditions mimicking the environment, but plants growing in the field respond to complex environmental conditions, which vary in time, duration, and intensity. Therefore, it is crucial to focus on different aspects of combined stresses to successfully develop crops that can withstand multiple stresses in the field (Mittler, 2006). The advantages of ‘stacking’ or ‘pyramiding’ of stress-related genes in crops offer a great potential to prepare futural crops for a fast-changing environment (Esmaeili et al., 2019; Wijewardene et al., 2020; Esmaeili et al., 2021; Balasubramaniam et al., 2022).
Although conventional breeding has improved crop yield considerably, it was not very successful in enhancing abiotic stress tolerance in crops (Hu and Xiong, 2014; Kumar et al., 2020; Rivero et al., 2022). This lack of success is partially due to breeders’ preference to test their genetic materials under optimum conditions. The complex nature of abiotic stresses and variability in plants’ sensitivity to different stresses during their life cycle further complicate the selection criteria for increased stress tolerance in conventional breeding. Therefore, it is imperative to adopt an alternative approach that could be employed to improve abiotic stress tolerance and enhance crop yield and quality. Genetic engineering is an alternative strategy to generate transgenic crops that can withstand the fast-changing environment. In the last two decades, transgenic plants have been generated by altering the expression of different genes responsible for a specific trait via different transformation methods (Ashraf and Akram, 2009). According to the International Service for the Acquisition of Agro-Biotech Applications (ISAAA, 2019), the United States, Brazil, Argentina, Canada, and India are the top five countries that grow genetically engineered crops. The major transgenic crops adopted by GMO-growing countries in 2019 were soybean, maize, cotton, and canola (ISAAA, 2019). In this review, we overview recent advances in developing transgenic plants with improved abiotic stress tolerance and discuss the roadmap for further enhancing abiotic stress tolerance in crops.
The in-depth study of plant stress response involves a coalition of pathways in terms of signal perception and transduction cascades, activation of transcription factors, expression or modulation of stress-related genes, production of various functional proteins and enzymes, production of osmolytes, increased antioxidation capacities against reactive oxygen species (ROS), and alterations to biochemical, physiological, and cellular aspects of plant cellular metabolisms. In this section, we will discuss the employment of the transgenic approach in the alteration of single gene expression, which improves plant performance under major abiotic stress conditions.
Since around 25% of the earth’s outermost layer is affected by salt, salinity is considered a growing agricultural problem threatening global food security (Rao et al., 2006; Abdelraheem et al., 2019). Soil salinization is caused by both human and natural processes. Accumulation of salt in soil inhibits water uptake by roots, disrupts ion homeostasis, and inversely affects plant growth and development (Zhu, 2001). To combat the adverse effects of salinity, plants have developed different strategies, including salt exclusion and compartmentalization into vacuoles. The Na+ accumulation in vacuoles and export out of cells are controlled by the activities of proton pumps and antiporters operating at the tonoplast and the plasma membrane (Gaxiola et al., 2001; Shi et al., 2003). The Na+/H+ antiporter exchanges vacuolar H+ for cytosolic Na+ and thus sequesters Na+ ions into vacuoles (Apse et al., 1999) or exports Na+ ions out of cells, while the proton gradient force generated by vacuolar membrane-bound H+-pyrophosphatase (V-PPase) and ATPase and plasma membrane-bound ATPase contribute to these processes (Gaxiola et al., 2001; Zhu, 2001).
Studies have shown that overexpression of several stress-related genes to reduce the uptake of toxic ions such as Na+ in the cytosol could improve plant salt tolerance (Table 1). Since the first report by Gaxiola et al. (2001), researchers have overexpressed the vacuolar H+-pyrophosphatase (V-PPase) genes in different crops such as alfalfa (Bao et al., 2009), rice (Kim et al., 2020), sugarcane (Kumar et al., 2014), cotton (Lv et al., 2008; Pasapula et al., 2011), peanut (Qin et al., 2013), wheat (Lv et al., 2015), and tobacco (Gao et al., 2006) to improve the salt tolerance and yield in those crops. The V-PPase-overexpressing plants also produce more robust root systems, which is due to more active auxin polar transport, thereby leading to more efficient water and nutrient absorption (Li et al., 2005). Extensive research has also been conducted on overexpression of the plant vacuolar Na+/H+ antiporter gene, NHX, to improve salt tolerance in transgenic plants. For instance, transgenic tomato plants overexpressing the Arabidopsis vacuolar Na+/H+ antiporter gene 1 (i.e., AtNHX1) could grow, flower, and produce fruits under 200 mM NaCl. At the same time, high Na+ and Cl- contents were detected in leaves, but not in the fruits (Zhang and Blumwald, 2001). Overexpression of AtNHX1 in cotton also improved salt tolerance, increased photosynthetic rate, and enhanced fiber production in transgenic lines after salt treatment (He et al., 2005). Transgenic canola, wheat, and maize plants overexpressing AtNHX1 demonstrated improved salt tolerance and produced more biomass and grain yield under saline conditions (Zhang et al., 2001; Xue et al., 2004; Yin et al., 2004). Furthermore, overexpression of AtNHX1 orthologs such as AtNHX5, OsNHX1, MdNHX1, TaNHX2, PgNHX1, and LeNHX2 also improved salt tolerance in crops including rice, eggplant, soybean, apple, and tomato. (Fukuda et al., 2004; Chen et al., 2007a; Verma et al., 2007; Li et al., 2010; Cao et al., 2011; Li et al., 2011a; Yarra et al., 2012; Huertas et al., 2013; Yarra and Kirti, 2019).
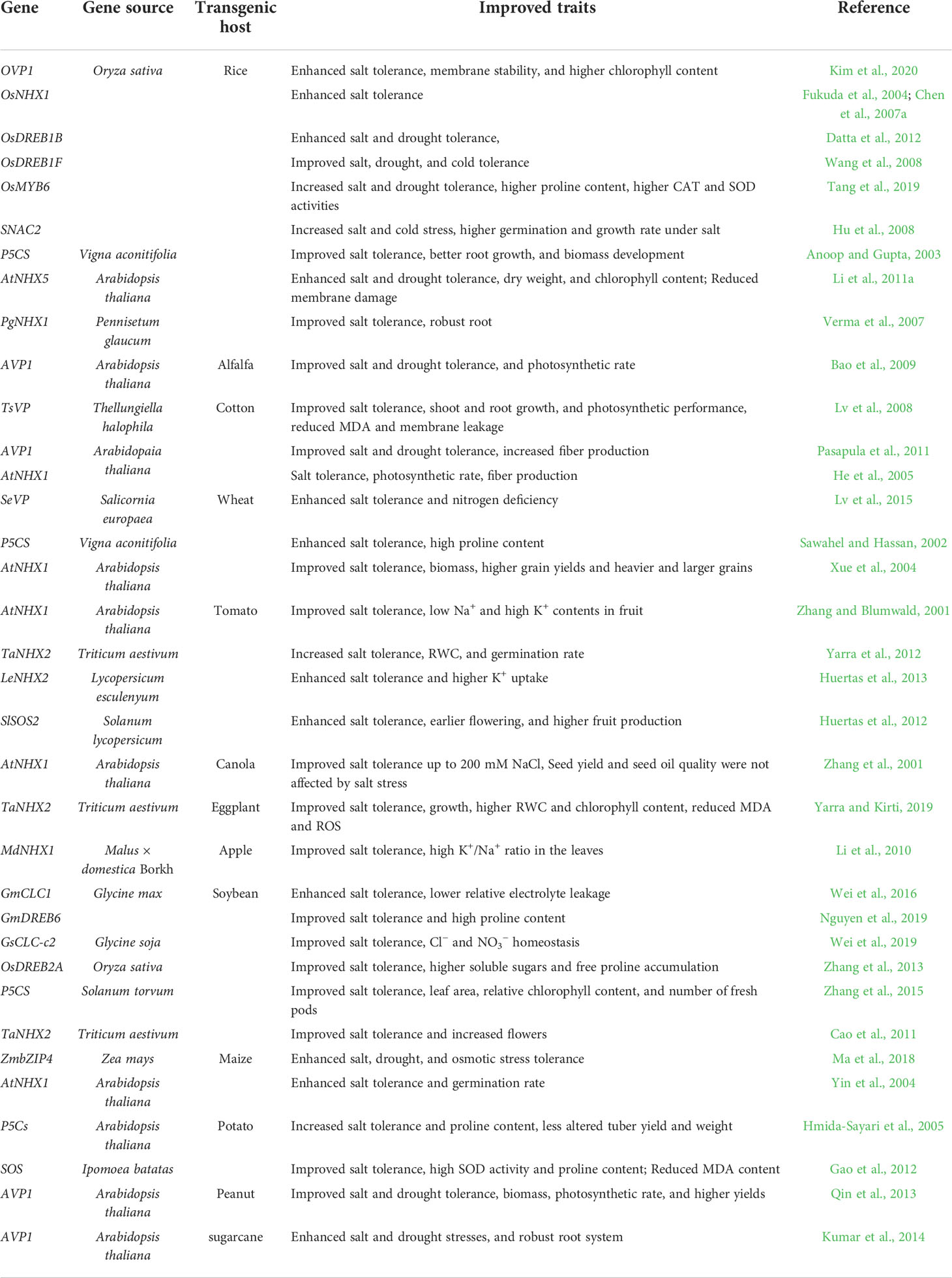
Table 1 Improving plant salt stress tolerance through genetic engineering.
The genes in the salt overly sensitive (SOS) signaling pathway, SOS1, SOS2, and SOS3, play a substantial role in salt tolerance in plants by excluding Na+ ions at the cellular level and maintaining ion homeostasis in root cells (Shi et al., 2000). Overexpression of SOS2 that encodes a calcineurin-interacting protein kinase from Solanum lycopersicum in tomato increased salt tolerance (Huertas et al., 2012). In a study by Yue et al. (2012), transgenic tobacco plants overexpressing the plasma membrane N+/H+ antiporter gene SOS1 showed enhanced salt tolerance, and they grew much better than wild-type plants when irrigated with 150 mM NaCl. A similar result was obtained by Gao et al. (2012) when SOS genes were overexpressed in sweet potato plants. Compartmentalization of Cl- ions in vacuole using chloride channel proteins (CLCs) is another mechanism to counter salt stress in plants to decrease Cl- levels in the cytosol, thereby maintaining ion homeostasis in plant cells. Overexpression of chloride channel protein genes GmCLC1 and GsCLC-c2 in soybean improved salt stress tolerance in transgenic plants (Wei et al., 2016; Wei et al., 2019), while silencing GhCLCg-1 in upland cotton compromised salt stress tolerance, indicating the importance of CLCs in plant response to salt stress (Liu et al., 2021).
Plant response to environmental stresses is regulated by a series of stress-related genes that are modulated by specific transcription factors. The functions of some transcription factors, e.g., MYC, bZIP, WRKY, NAC, and AP2, in salt signaling pathways were identified (Golldack et al., 2011). Overexpression of the dehydration responsive element binding protein genes, DREB1B and OsDREB1F in rice (Wang et al., 2008; Datta et al., 2012), GmDREB2 in tobacco (Chen et al., 2007b), GmDREB6 and OsDREB2A in soybean (Zhang et al., 2013; Nguyen et al., 2019), and DREB1A in potato (Behnam et al., 2006) improved salt stress tolerance. The transgenic rice overexpressing the stress-responsive NAC transcription factor gene SNAC2 showed improved salinity tolerance (Hu et al., 2008). The myeloblastosis oncogene encoded proteins (MYBs) are another group of transcription factors whose function in abiotic stress response in plants is well understood. Recently, Tang et al. (2019) showed that overexpression of the rice gene OsMYB6 improves salinity and drought stress tolerance in transgenic rice. OsMYB6-overexpressing rice plants also produced a larger amount of proline, and they showed increased catalase (CAT) and superoxide dismutase (SOD) activities (Tang et al., 2019). Also, overexpression of ZmbZIP4 in maize improved salt and drought tolerance at the seedling stage and enhanced osmotic stress adjustments (Ma et al., 2018).
Osmotic adjustment is a vital plant response to abiotic stresses. The accumulation of osmolytes under abiotic stress conditions is well documented. These compatible osmolytes include amino acids and their derivatives (e.g., proline and glycine betaine), soluble sugars (e.g., trehalose and mannitol), and sugar alcohols (Suprasanna et al., 2016). Several reports showed that manipulating genes that control the production of low molecular weight metabolites such as proline (an essential amino acid in plants) improves plant tolerance to abiotic stresses, including salinity and drought. Transgenic soybean overexpressing the proline biosynthetic gene P5CS that encodes the Δ′-pyrroline-5-carboxylate synthetase (P5CS) demonstrated increased salt tolerance with higher proline content (Zhang et al., 2015). This is consistent with previous results from transgenic wheat, potato, and indica rice plants overexpressing P5CS (Sawahel and Hassan, 2002; Anoop and Gupta, 2003; Hmida-Sayari et al., 2005). Glycine betaine (GB) is an essential osmolyte that protects plants against osmotic stress by stabilizing membrane and photosynthetic machinery under salt, drought, and cold stresses. Transgenic rice was developed by overexpressing the choline oxidase A gene, codA, in chloroplast and cytosol. In both cases, higher production of GB and improved salt stress tolerance were observed. Since rice does not produce GB, expression of codA in transgenic rice is essential for the increased abiotic stress tolerance in rice (Mohanty et al., 2002).
Plants have developed mechanisms such as antioxidant molecules and enzymes to respond to the overproduction of reactive oxygen species (ROS) generated under abiotic stress conditions (Grene, 2002; Devireddy et al., 2021). The antioxidant molecules such as ascorbate and glutathione can interact directly with ROS and therefore reduce ROS content in plant cells. The antioxidant enzymes such as superoxide dismutase (SOD), catalase (CAT), ascorbate peroxidase (APX), and glutathione reductase (GR) can scavenge ROS efficiently in plant cells (Anjum et al., 2016). Numerous studies on overexpression of antioxidant genes to improve plant tolerance to abiotic stresses were reported (Gill and Tuteja, 2010). For instance, overexpression of Cu/Zn SOD and APX in potatoes increased salt stress tolerance (Yan et al., 2016).
Drought is a major stress that results in huge crop loss. Around one-third of the lands on earth are located in arid and semiarid regions where drought severely limits agricultural production (Abdelraheem et al., 2019). Therefore, developing crops that can grow and produce high yields under water deficit conditions is urgently needed. Plant response to drought stress is very complex which involves different pathways, and the intensity and span of drought stress alter both their interaction and individual responses. In the past few decades, many studies have focused on generating transgenic crops with improved drought tolerance (Table 2). The first commercialized drought-tolerant corn known as DroughtGard, MON87460, which expresses the cold shock protein B gene from Bacillus subtilis, was developed by Monsanto (Monsanto Co., St. Louis, MO, USA) and released in 2009 (Nemali et al., 2014; Liang, 2016a). However, its success in the market is not very clear.
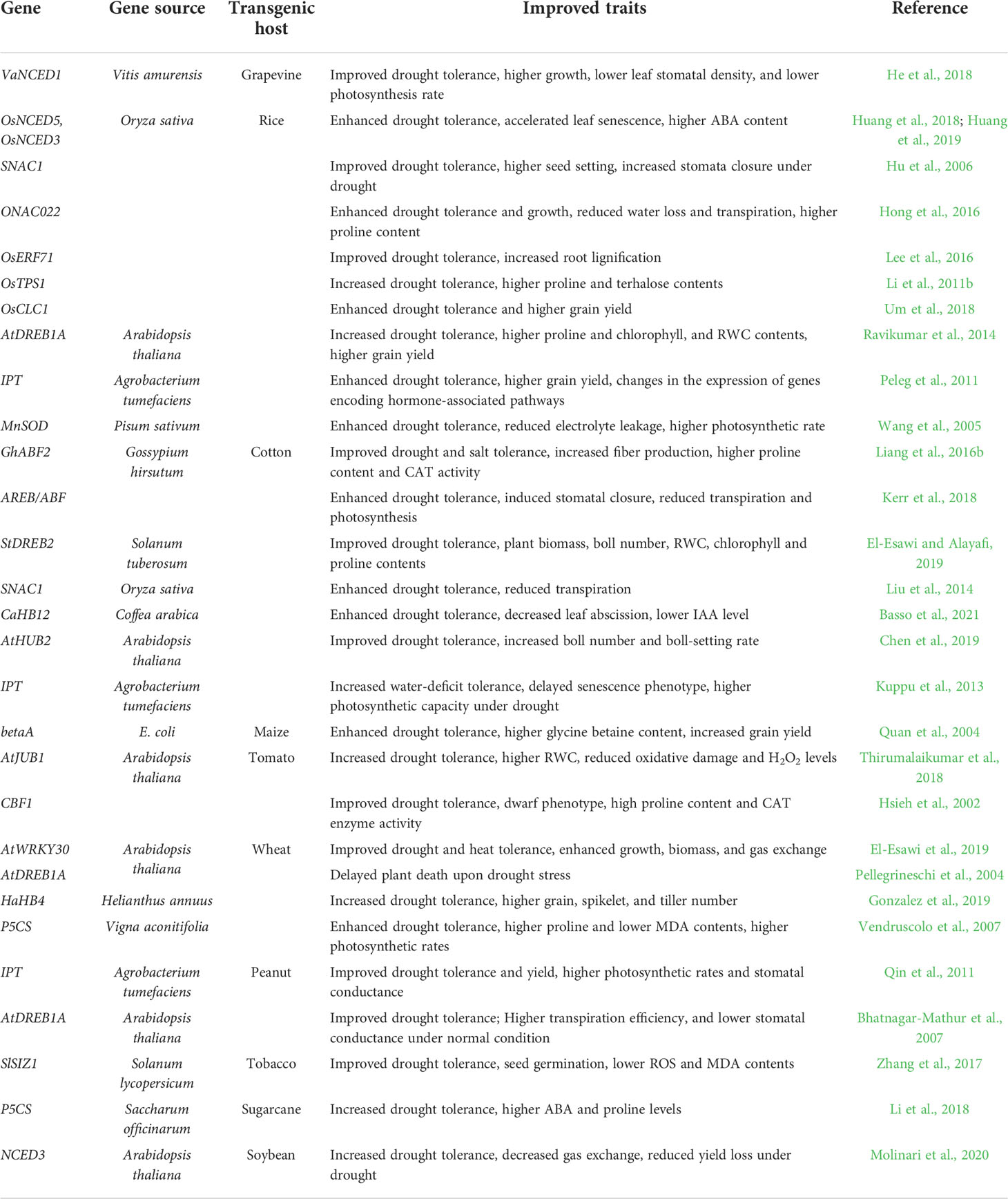
Table 2 Improving drought stress tolerance in transgenic crops.
Phytohormones are known to play a crucial role in plant response to environmental stresses including drought stress. Water deprivation alters biosynthesis of different phytohormones such as abscisic acid (ABA), auxins (IAA), gibberellins (GAs), jasmonic acid (JA), ethylene (ET), salicylic acid (SA), brassinosteroids (BRs), cytokinins (CKs). Although ABA is the major phytohormone whose production is induced by drought stress, the crosstalk between other hormones improves drought stress response in plants (Ullah et al., 2018). Two ABA-dependent and three ABA-independent regulatory pathways are involved in plant response to drought stress (Shinozaki and Yamaguchi-Shinozaki, 2007). It was shown that ABA induces the expression of most drought-related genes (e.g., NCED, RD22, ABREs, and RD29), and the up-regulation can be 40 times higher under drought stress conditions compared to under normal growth conditions (Shinozaki et al., 2003). The enzyme 9-cis-epoxycarotenoid dioxygenase (NCED) is a key enzyme that converts epoxy-carotenoid precursor to xanthonin, which is consequently converted to ABA. Overexpression of VaNCED1 in grapevine (He et al., 2018), OsNCED3 and OsNCED5 in rice (Huang et al., 2018; Huang et al., 2019), and AtNCED3 in soybean (Molinari et al., 2020) significantly improved drought tolerance in transgenic plants. The drought-responsive element (DRE)-binding proteins such as DREB1 and DREB2 are transcription factors that bind to the promoter region of dehydration-responsive genes such as RD29A and induce their expression in response to environmental stresses, including drought (Shinozaki and Yamaguchi-Shinozaki, 2007). The stress-inducible expression of transcriptional factor gene DREB1A in peanut and rice improved drought stress tolerance in transgenic plants, while its expression in wheat delayed plant death upon water deprivation (Pellegrineschi et al., 2004; Bhatnagar-Mathur et al., 2007; Ravikumar et al., 2014). Hsieh et al. (2002) showed that the ectopic expression of the Arabidopsis gene CBF1 (C repeat/dehydration-responsive element binding factor 1) in tomatoes improved plant tolerance to water deprivation, and transgenic plants accumulated more proline and showed higher CAT activity. Moreover, ectopically overexpressing AREB/ABF genes (coding for ABA binding factor/ABA-responsive element binding proteins) in cotton enhanced drought tolerance through stomatal regulation (Kerr et al., 2018). Transgenic cotton plants overexpressing the potato DREB2 gene, StDREB2, showed improved drought tolerance with increased biomass and boll number (El-Esawi and Alayafi, 2019).
Hu et al. (2006) reported an improved drought tolerance in transgenic rice plants by overexpressing the SNAC1 gene, which was attributed to the function of this gene in regulating stomata closure and water use efficiency. Overexpression of SNAC1 in cotton improved salt and drought stress tolerance, enhanced the rooting system, and reduced the transpiration rate in transgenic plants (Liu et al., 2014). Furthermore, overexpression of ONAC022 enhanced drought and salt tolerance with higher ABA biosynthesis in transgenic rice (Hong et al., 2016). Overexpression of the JUNGBRUNNEN1 gene, AtJUB1 (a member of the NAC family) in tomatoes increased drought tolerance associated with higher relative water content and lower H2O2 levels (Thirumalaikumar et al., 2018). Silencing the Gossypium barbadense MYB gene, GbMYB5, compromised drought tolerance in cotton, while its overexpression in tobacco enhanced drought stress response (Chen et al., 2015). It was also shown that the activities of antioxidant enzymes SOD, CAT, and POD (peroxidase) were lower and higher in silenced cotton and transgenic tobacco, respectively (Chen et al., 2015). The transcription factor gene AtWRKY30 was overexpressed in wheat, leading to increased drought and heat tolerance (El-Esawi et al., 2019). Overexpression of WRKY30 also induced the transcript levels of several stress-related genes such as DREB1, DREB3, WRKY19, and TIP2 (El-Esawi et al., 2019). Furthermore, overexpression of the drought-responsive AP2/ERF transcription factor gene OsERF71 enhanced drought tolerance in transgenic rice by increasing the lignification level in roots, resulting in root architecture alteration (Lee et al., 2016). Transgenic cotton plants overexpressing the Coffea arabica HB12 gene, CaHB12, and Gossypium hirsutum bZIP transcription factor gene, GhABF2, independently, showed improved drought tolerance by up-regulation of genes in the ABA-dependent signaling pathway (Liang et al., 2016b; Basso et al., 2021). Recently, González et al. (2019) overexpressed the sunflower gene HaHB4 that encodes a protein in the homeodomain-leucine zipper I family in wheat, and they showed that transgenic wheat grown in the field under drought conditions outperformed wild-type wheat.
Proline is an essential amino acid and metabolite with functions in osmotic adjustment and free radical scavenging in plants under stress conditions (Ghosh et al., 2022). Transgenic wheat and sugarcane overexpressing the P5CS gene showed improved drought stress tolerance associated with higher proline production (Vendruscolo et al., 2007; Li et al., 2018). As a nonreducing disaccharide, trehalose plays a vital role in plant cellular metabolism and plant response to environmental stresses (Grennan, 2007). Overexpression of the rice trehalose-6-phosphate synthase gene, OsTPS1, enhanced salt and drought tolerance in transgenic rice by increasing trehalose and proline contents (Li et al., 2011b). Previously, Jang et al. (2003) transformed rice with a gene encoding a bifunctional fusion enzyme of the TPS synthase and TPS phosphatase from Escherichia coli, and they observed a higher accumulation of trehalose with improved drought, salt, and cold stress tolerance. Also, overexpression of the betaA gene from E. coli in maize elevated glycine betaine levels in transgenic plants and improved drought tolerance and grain yield (Quan et al., 2004).
Overexpression of the isopentenyl transferase gene, IPT (a key gene in the biosynthesis of cytokinin), led to increased drought tolerance in transgenic peanuts (Qin et al., 2011) and rice (Peleg et al., 2011). A similar result was observed in IPT-transgenic cotton (Kuppu et al., 2013); however, an extended study by Zhu et al. (2018) showed that the increased drought tolerance of IPT-transgenic cotton depends on the timing of the occurrence of water deficit stress: if drought stress occurs after cotton starts to flower, the increased drought tolerance is lost. Transgenic rice plants overexpressing MnSOD showed enhanced drought tolerance (Wang et al., 2005). Recently Chen et al. (2019) showed that overexpression of AtHUB2 that encodes a histone H2B monoubiquitination E3 ligase significantly improves boll number and boll-setting rate under drought stress conditions in transgenic cotton. Overexpression of the tomato SlSIZ1 gene in tobacco confers increased drought tolerance, which was attributed to the accumulation of proline and SUMO conjugates (Zhang et al., 2017). Also, transgenic tobacco plants showed improved seed germination and growth while they accumulated lower amounts of ROS and malondialdehyde (Zhang et al., 2017). In rice, the root-specific expression of the chloride channel gene, OsCLC1, enhanced drought tolerance and resulted in higher grain yield in transgenic plants, whereas chloride channel mutant osclc1 exhibited compromised drought tolerance and produced less yield than wild-type plants (Um et al., 2018).
The average temperature of the earth is rising (Battisti and Naylor, 2009). Changes in ambient temperature accelerated by global warming affect rainfall and drought patterns across the globe, thus negatively impacting agricultural production. According to the Inter-Governmental Panel on Climatic Change (IPCC), the global average surface temperature by the end of 21st century will be 0.3 - 1.7 °C under the Representative Concentraion Pathway (RCP) 2.6, 1.1 - 2.6 °C under RCP 4.5, 1.4 - 3.1 °C under RCP 6.0, and 2.6 - 4.8 °C under RCP 8.5 (IPCC, 2014). Plants’ capacity to cope with changing environments, including temperature fluctuations, varies among different species due to variations in basal and acquired thermotolerance (Grover et al., 2013). Therefore, implementation of transgenic technology is a practical solution for sustainable agriculture where transgenic crops can grow and reproduce under extreme temperatures with minimum or no damage to their cells (Mishra et al., 2017; Esmaeili et al., 2021). Here we summarize the recent advances in developing transgenic crops with improved heat tolerance (Table 3).
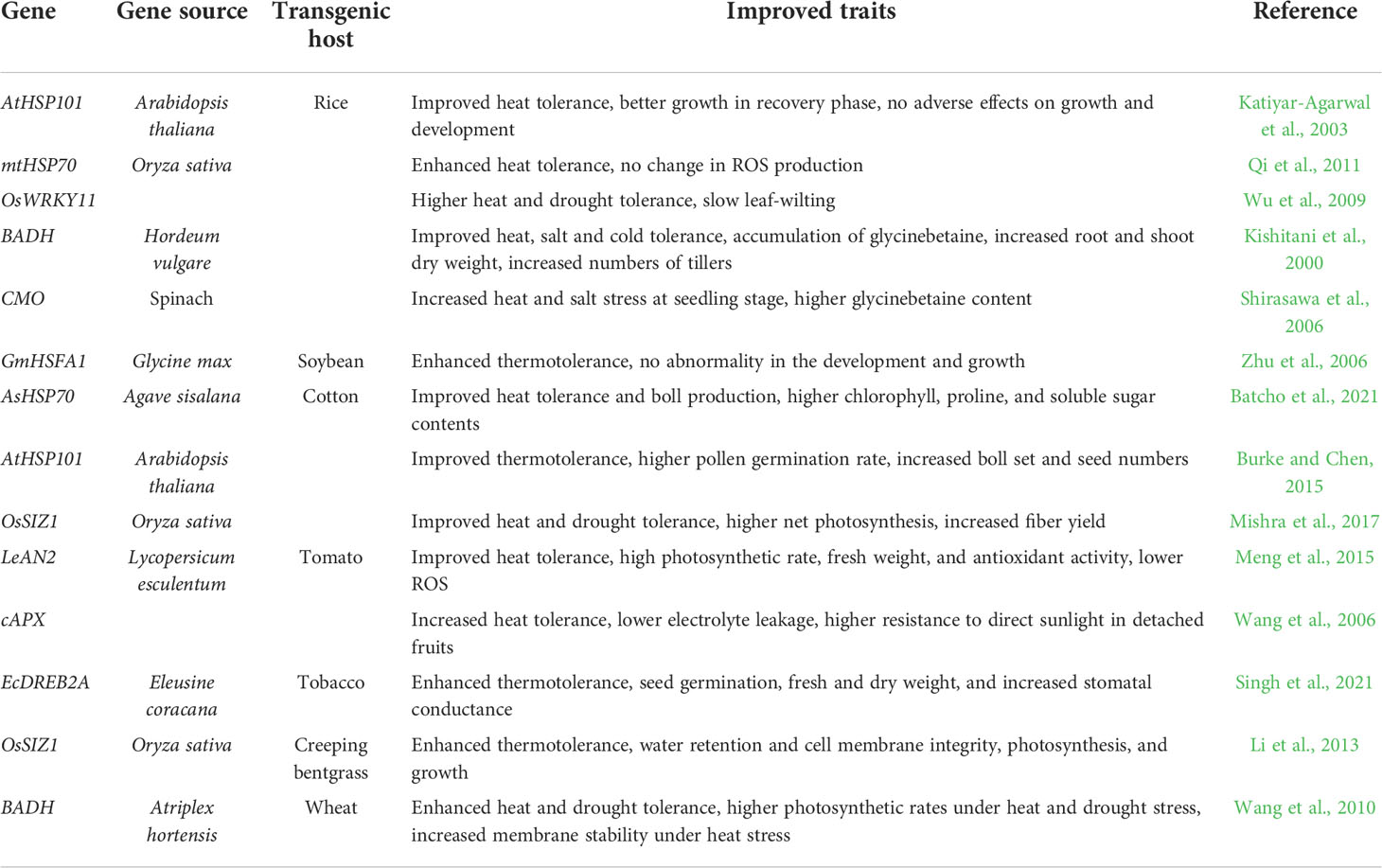
Table 3 Improving heat stress tolerance in transgenic crops.
Upon heat stress, cellular proteins lose their biological activities due to aggregation and misfolding of proteins. As a primary response to increased temperature, plants have evolved molecular chaperone and protein degradation machinery to minimize the heat-related damage in cells. The upregulation of heat shock protein (HSP) genes is an important event associated with heat stress response which results in the accumulation of HSPs as molecular chaperones to stabilize, repair, and re-fold denatured proteins, thus protecting cells against heat stress-related damages (Mittler et al., 2012). Several reports showed that overexpression of HSP genes in plants improves heat stress tolerance. For instance, overexpression of the mitochondrial HSP70 gene, mtHSP70, and HSP101 in rice (Katiyar-Agarwal et al., 2003; Qi et al., 2011), the Glycine max heat shock transcription factor gene, GmHsfA1, in soybean (Zhu et al., 2006) increased heat stress tolerance in transgenic plants. Transgenic cotton overexpressing AtHSP101 (Burke and Chen, 2015) and AsHSP70 (Batcho et al., 2021) demonstrated improved heat stress tolerance and produced more boll and seeds under high temperatures.
LeAN2 is an anthocyanin-associated R2R3-MYB transcription factor, and it was shown that overexpression of LeAN2 in tomatoes up-regulated transcripts of several genes in the anthocyanin biosynthetic pathway and caused enhanced heat stress tolerance (Meng et al., 2015). WRKY transcription factors function as repressors and activators of gene expression, and when the WRKY11 gene was overexpressed in rice, transgenic rice showed improved tolerance to heat and drought stresses (Wu et al., 2009). Although DREB transcription factors were first reported to be involved in plant response to drought and cold stresses, some research indicated that DREBs and HSFs could interact with each other in response to extreme heat (Grover et al., 2013). Recently Singh et al. (2021) showed that overexpression of EcDREB2A in tobacco improves heat stress tolerance through increasing ROS scavenging capacity in transgenic plants.
Accumulating osmolytes during the heatwave is an adaptive mechanism to protect protein’s structure in plant cells (Suprasanna et al., 2016). Overexpression of the barely peroxisomal betaine aldehyde dehydrogenase gene, BADH, in rice improved tolerance to heat, cold, and salt stresses (Kishitani et al., 2000). Transgenic wheat overexpressing BADH from Atriplex hortensis showed enhanced thermotolerance, which was attributed to a more stable membrane (Wang et al., 2010). Accumulating glycine betaine in transgenic rice overexpressing the choline monooxygenase gene CMO resulted in higher thermotolerance (Shirasawa et al., 2006). Enhanced plant biomass production and heat stress tolerance were achieved in transgenic Medicago plants overexpressing the TPS1-TPP2 genes from yeast (Suárez et al., 2009).
Oxidative stress results from ROS accumulation in plant cells, which can be caused by many environmental stress conditions such as heat stress. Thus, utilization of genes involved in antioxidation metabolism could lead to enhanced thermotolerance in transgenic plants (Grover et al., 2013). Transgenic tomato plants overexpressing a cytosolic peroxidase gene, cAPX, increased tolerance to heat stress (Wang et al., 2006). SUMOylation is an essential post-translational modification process in plants that is also involved in abiotic stress response and the ABA-signaling pathway. Overexpression of the rice SUMO E3 ligase gene, OsSIZ1, drastically increased plant tolerance to heat and drought stresses (Li et al., 2013; Mishra et al., 2017). Transgenic cotton plants overexpressing OsSIZ1 produced more fiber and maintained higher photosynthetic rates under heat stress conditions (Mishra et al., 2017).
Plants are usually exposed to a combination of different stresses in the field; thus, plant tolerance to multiple stresses differs from their response to single stress (Zandalinas et al., 2022a and Zandalinas et al., 2022b). Therefore, more studies are required to discover the molecular mechanism of plant response to multiple stresses. In addition, crop improvement studies should focus on stress combinations that mimic field conditions (Mittler, 2006; Tian et al., 2021; Zandalinas et al., 2021a; Zandalinas et al., 2021b). Improving plant stress tolerance to complex environments such as combined drought, heat, and salt stresses is unlikely achievable if only a single gene is altered. Thus, the gene stacking strategy is a potential solution to improve crops’ tolerance to multiple stresses.
In recent years several studies have reported enhanced tolerance to multiple stresses (Table 4). Co-overexpression of AVP1 and AtNHX1 in cotton enhanced tonoplast Na+/H+ antiporter activity, leading to improved salt and drought tolerance (Shen et al., 2015). These AVP1/AtNHX1 co-overexpressing cotton plants outperformed AVP1-overexpressing and AtNHX1-overexpressing cotton plants and produced more bolls and higher fiber yield under 200 mM NaCl. Furthermore, they produced more fiber than wild-type, AVP1-overexpressing, and AtNHX1-overexpressing cotton plants under low-irrigation and dryland conditions in the field (Shen et al., 2015). Improved salt stress tolerance was also achieved in transgenic cotton co-overexpressing AtNHX1 and TsVP (Cheng et al., 2018). The AtNHX1/TsVP co-overexpressing cotton plants produced higher seed yield when grown in saline soil, which was attributed to the accumulation of Na+, K+, and Ca2+ in leaves and better cellular ion homeostasis in transgenic plants (Cheng et al., 2018). Transgenic tomato plants co-overexpressing LeNHX2 and SlSOS2 showed enhanced salt tolerance and produced more yield with enhanced fruit quality (Baghour et al., 2019). Recently, several studies employed the stacking approach to co-overexpress multiple genes in model plants, and the results are very promising. For instance, co-overexpression of AVP1 and PP2A-C5 in Arabidopsis increased tolerance to multiple stresses, including salt stress, drought stress, and phosphorous deficiencies (Sun et al., 2018). Wijewardene et al. (2020) showed that co-overexpression of the AVP1 and the creosote Rubisco activase gene RCA in Arabidopsis leads to improved tolerance to drought, heat, and salt stresses. In addition, transgenic plants performed significantly better under combined drought and heat stresses, as well as under combined salt, drought, and heat stresses, and the seed yield was increased dramatically in transgenic plants. These results suggest that co-overexpression of AVP1 and RCA could increase tolerance to combined drought and heat stresses and increase crop yield in light of climate changes (Wijewardene et al., 2020). In a study by Balasubramaniam et al. (2022), three genes, AVP1, PP2A-C5, and AtCLCc, were co-overexpressed in Arabidopsis to improve salt and drought tolerance, and they were able to increase the salt tolerance level up to 300 mM NaCl, a level never reached before via genetic engineering approach. We demonstrated that transgenic Arabidopsis and cotton plants co-overexpressing AVP1 and OsSIZ1 showed significantly improved tolerance to combined stresses such as heat with drought or drought with salt (Esmaeili et al., 2019; Esmaeili et al., 2021). In particular, in field trials under dryland conditions, transgenic cotton co-overexpressing AVP1 and OsSIZ1 produced 133% and 81% more fiber in two consecutive years (Esmaeili et al., 2021). These findings demonstrate that co-overexpression of multiple genes that can improve plant tolerance to abiotic stresses without imposing drawbacks on plant growth and development could be employed as an effective strategy to achieve significant tolerance towards environmental stresses.
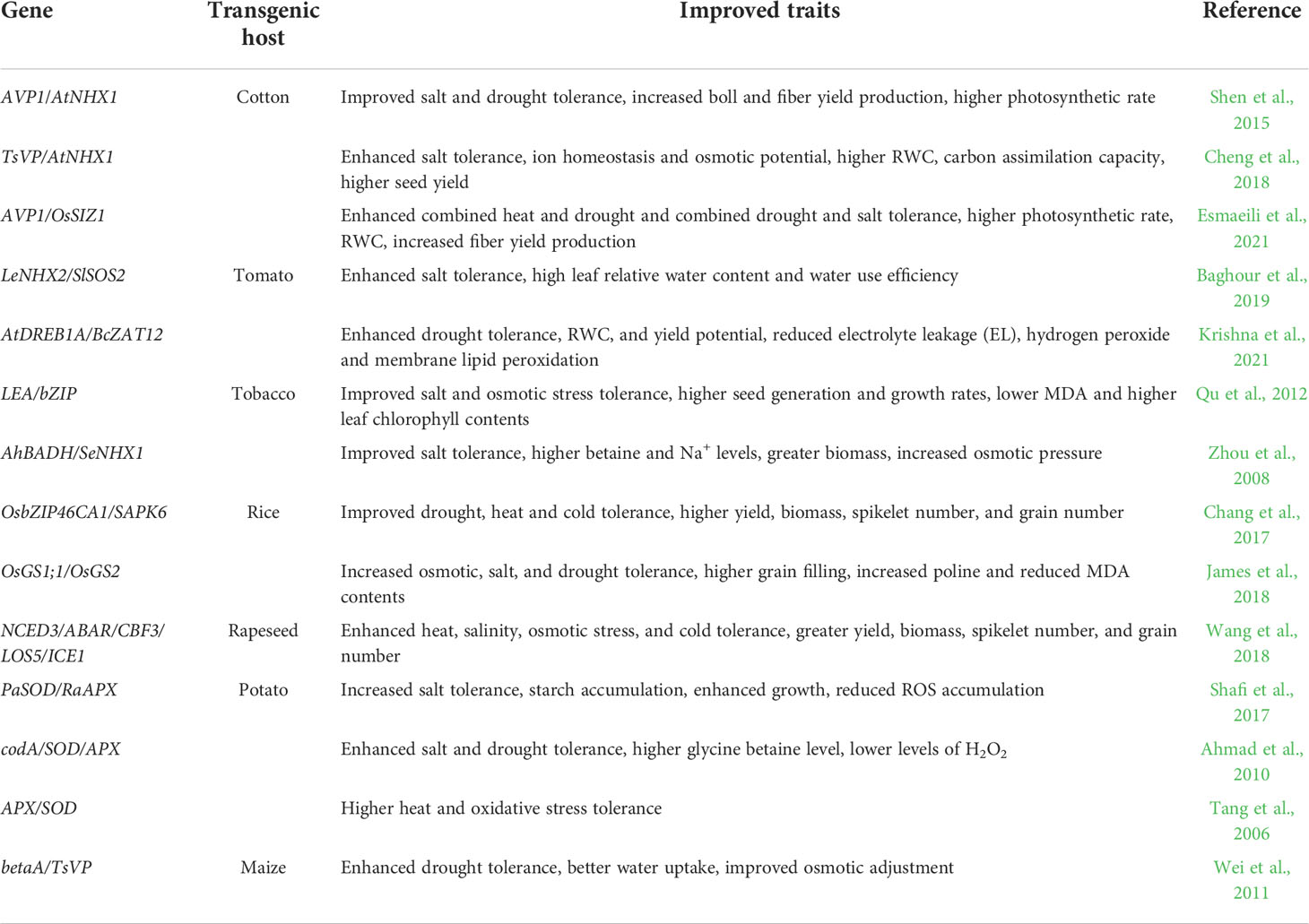
Table 4 Enhanced tolerance to multiple stresses using gene pyramiding approach.
Gene pyramiding of the Arabidopsis gene AtDREB1A with the Brassica Zinc finger transcription factor gene BcZAT12 in tomato improved drought tolerance and fruit production (Krishna et al., 2021). These transgenic tomato plants also showed an elevated water use efficiency with higher proline content, while electrolyte leakage, hydrogen peroxide level, and membrane lipid peroxidation were significantly reduced (Krishna et al., 2021). Co-overexpression of the late embryogenesis abundant protein gene LEA and the basic leucine zipper transcriptional factor gene bZIP in tobacco enhanced tolerance to salt and osmotic stresses, resulting in increased seed production (Qu et al., 2012). Previously, co-overexpression of the constitutively active form of a bZIP transcription factor gene OsbZIP46CA1 and a protein kinase gene SAPK6 in rice enhanced tolerance to drought, heat, and cold stresses (Chang et al., 2017). The gene pyramiding approach was also employed to generate transgenic rapeseed (Brassica napus) with novel traits (Wang et al., 2018). Five stress-related genes, NCED3 (nine-cis-epoxycarotenoid dioxygenase 3), ABAR (ABA receptor, magnesium-chelatase subunit chlH), CBF3 (c-repeat binding factor 3), LOS5 (molybdenum cofactor sulfurase), and ICE1 (interactor of little elongation complex ELL subunit 1) were simultaneously expressed in rapeseed plants, which led to improved tolerance to multiple stresses such as heat, salinity, osmotic stress, and cold (Wang et al., 2018).
Glutamine synthetase (GS) is known for its function in the nitrogen metabolism, in which its activity and expression are affected by different abiotic stresses (Ji et al., 2019). Recently, James et al. (2018) developed transgenic rice plants that co-overexpress the rice cytosolic GS1 and chloroplastic GS2 genes, OsGS1;1 and OsGS2, and they observed improved osmotic, salt, and drought tolerance in transgenic plants. In addition, the grain filling rate in transgenic rice was dramatically higher than that in control plants under salinity and drought stresses, which was associated with higher proline and lower MDA contents (James et al., 2018). In transgenic potatoes, enhanced heat stress tolerance and increased antioxidation capacity were achieved by co-overexpressing a Cu/Zn superoxide dismutase gene SOD and an ascorbate peroxidase gene APX in the chloroplast (Tang et al., 2006). Co-overexpression of ROS scavenging enzymes genes PaSOD and RaAPX in potato increased starch accumulation and enhanced growth under salt stress conditions, while it reduced accumulation of ROS because of the higher APX and SOD activities in transgenic plants (Shafi et al., 2017). Transgenic potato plants co-overexpressing the genes codA, SOD, and APX showed improved salt stress tolerance (Ahmad et al., 2010).
Co-overexpression of the Atriplex betaine synthesis gene BADH with the Salicornia NHX1 gene SeNHX1 in tobacco resulted in a higher accumulation of betaine and Na+ in transgenic lines under salinity stress, and these BADH/SeNHX1 co-overexpressing plants produced more biomass and maintained increased osmotic pressure (Zhou et al., 2008). Wei et al. (2011) employed a similar approach to simultaneously overexpress betaA from Escherichia coli and TsVP from Thellungiella halophila in maize. Their study showed that betaA/TsVP co-overexpressing maize lines performed better than wild-type plants under drought stress conditions, with an enhanced osmotic adjustment that facilitates water uptake by roots (Wei et al., 2011).
Although the gene stacking approach is an effective and promising strategy to improve plant tolerance to abiotic stresses, there might be some drawbacks associated with this approach. Overexpression of single target genes such as those encoding specific enzymes or transporters could improve plant performance against individual stresses in most cases, or two related stresses such as drought and salt stresses, while manipulating the upstream regulatory genes such as transcriptional factor genes could result in enhanced tolerance against multiple abiotic stresses. However, due to the complex cross-talks among regulatory and metabolic pathways in plants, altering upstream genes might also lead to undesired agronomical traits, including growth retardation. Therefore, as mentioned above, an appropriate combination of genes to ensure the synergistic interactions of the genes is critical. Since promoters can significantly affect the outcomes from a transgenic alteration, selecting a proper combination of promoters for the genes is also crucial.
Recently, clustered regularly interspaced short palindromic repeats-associated protein 9 (CRISPR/Cas9) technique has emerged as a promising gene editing technology with a great potential for precise genetic modification, aiming to improve abiotic stress tolerance in plants (Li et al., 2022). Unlike zinc-finger nucleases (ZFNs) and transcription activator-like endonucleases (TALENs), the CRISPR/Cas9 endonuclease system is a very fast, accurate, and highly efficient genome editing tool to introduce desirable traits in plants for crop improvement (Rao and Wang, 2021). Genome editing using the CRISPR/Cas9 knock-out system has produced several crops with enhanced environmental stress tolerance. The simultaneous knock-out of three abscisic acid receptor genes, OsPYL1, OsPYL4, and OsPYL6, improved heat stress tolerance in rice, which leads to increased yield production (Miao et al., 2018). In plants, mitogen-activated protein kinases (MAPKs) are highly conserved serine and threonine protein kinases that are involved in plant development, hormone regulation, and response to abiotic stresses. In a study by Yu et al. (2019), the SlMAPK3 gene, a member of the MAPK family in tomatoes, was knocked out using the CRISPR/Cas9 technique. The slmapk3 mutants showed an improved heat stress tolerance, and the transcripts of several HSP and HSF genes were upregulated under stress conditions (Yu et al., 2019). Recently, Zeng et al. (2020) employed CRISPR/Cas9 technology to generate three knock-out mutants in rice by editing OsPIN5b, GS3, and OsMYB30, and they showed that the osmyb30 (a cold-responsive R2R3-type MYB gene) mutants displayed improved cold tolerance, while ospin5b (a gene involved in balance and transport of auxin) and gs3 mutants showed increased panicle length and enlarged grain size, respectively. In addition, the simultaneous knockout of all three genes resulted in enhanced cold tolerance and higher yield production compared to WT plants (Zeng et al., 2020). The auxin response factor (ARF) regulates the auxin-responsive genes in plants. It was shown that the knockouts of SlARF4 in tomato improved tolerance to drought stress and increased rehydration ability via upregulation of ABA signaling pathway genes such as SlABI5/ABF and SCL3 (Chen et al., 2021). In addition, arf4 mutants showed a higher level of antioxidant enzyme activities compared to WT plants, and no significant decrease in photosynthetic efficiency was observed (Chen et al., 2021). Furthermore, the antisense down-regulation of SlARF4 in tomato plants enhanced salinity and osmotic stress tolerance. In addition, plants showed higher levels of soluble sugars and chlorophyll contents and enhanced root growth under stress conditions (Bouzroud et al., 2020). The ARF4 antisense plants also maintained higher relative water content in leaves and ABA content under both normal and stress conditions, which was attributed to the upregulation of several ABA biosynthesis genes (Bouzroud et al., 2020). The ARGOS8 is considered a negative regulator of ethylene responses in maize plants, but its mRNA level is relatively low in maize. In an effort by Shi et al. (2017), novel ARGOS8 variants harboring native maize GOS2 promoter were generated using the CRISPR-Cas9 technology. The results showed that the ARGOS8 transcripts level was elevated in ARGOS8 variants. In addition, the ARGOS8 variants produced significantly higher grain yield than WT plants when grown under drought stress conditions in the field (Shi et al., 2017). Furthermore, the targeted mutagenesis of the Rice Enhanced Response to ABA1 gene, OsERA1, improved ABA response and drought stress tolerance in osera1 rice mutants (Ogata et al., 2020).
The CRISPR/Cas9 system was also employed to enhance salt stress tolerance in rice at the seedling stage via targeted mutagenesis of the transcription factor gene OsRR22, and no significant differences in agronomy traits were observed between the osrr22 mutant and WT plants (Zhang et al., 2019). Tran et al. (2021) showed that the negative-response domain in SlHyPRP1 was precisely removed in tomato hybrid plants using the CRISPR/Cas9 method. The tomato hybrid proline-rich protein 1, HyPRP1, is considered a negative regulator of salt stress; however, its elimination in tomato plants resulted in an increased salt stress tolerance at both germination and vegetation stages (Tran et al., 2021). The ABA-induced transcription repressors (AITRs) are members of the transcription factor family involved in the ABA signaling pathway. It was reported that the targeted mutation of GmAITR in soybean via the CRISPR/Cas9 gene editing strategy leads to improved salinity tolerance in mutants grown in the field (Wang et al., 2021). In a study by Lou et al. (2017), the functional properties of osmotic stress/ABA–activated protein kinase 2 (SAPK2) in rice was investigated. The sapk2 mutants generated via CRISPR/Cas 9 showed ABA-insensitive phenotypes during germination and reduced tolerance to drought stress, indicating that SAPK2 is involved in drought stress response in rice (Lou et al., 2017).
The nonexpressor of pathogenesis-related gene 1 (NPR1) is a salicylic acid receptor, and its function in plant response to pathogens has been well documented (Wu et al., 2012). However, there is a poor understanding of NPR1’s role in regulating plant response to abiotic stresses (Li et al., 2019). Recently, CRISPR/Cas9 technique was employed to generate mutations in the NPR1 gene in tomato plants. The slnpr1 mutants showed reduced drought stress tolerance with a significant decrease in the transcript level of several drought-related genes, including SlGST, SlDHN, and SlDREB (Li et al., 2019). In rice, the leaf morphology is a critical agronomical trait where rolled leaves show reduced water loss and improved drought tolerance compared to semi-rolled leaves. CRISPR/Cas9-based mutagenesis of semi-rolled leaf1 and leaf2 genes, SRL1 and SRL2, in rice resulted in rolled leaf mutants with enhanced drought stress tolerance and grain filling percentage compared to WT plants (Liao et al., 2019).
The CRISPR from Prevotella and Francisella 1 (Cpf1) is a single RNA-guided endonuclease with several advantages that sets it apart from the Cas9 system. Unlike CRISPR/Cas9 system, the CRISPR-Cpf1 results in higher transformation efficiency because it does not require large constructs to express multiple sgRNA cassettes (Wang et al., 2017). With high efficiency, engineered CRISPR-Cpf1 was recently used for multiplex gene editing in rice plants. The Francisella novicida Cpf1 (FnCpf1) was used to edit four members of rice Related to receptor-like kinases (OsRLKs) and Lachnospiraceae bacterium ND2006 Cpf1 (LbCpf1) was employed to edit four members of the rice Bentazon‐sensitive‐lethal (OsBELs). The analysis of T0 transgenic rice plants showed successful multiplex gene editing, indicating that engineered CRISPR-Cpf1 could be utilized as a powerful tool to target multiple members of a gene family (Wang et al., 2017). Overall, CRISPR/Cas9 gene editing technique involves simple designing and cloning methods that plant biotechnologists could use as an alternative tool for crop improvement. This technique has been employed in several different crop species, such as rice, wheat, maize, tomato, and soybean, to improve their yield and/or their response to biotic and abiotic stresses. Although it is a highly efficient and fast tool, CRISPR/Cas9 is relatively new and has been modified in many functional studies, further improvements are likely needed (Jaganathan et al., 2018).
This review briefly summarizes the recent advances in developing transgenic crops to combat major abiotic stresses, including heat, drought, and salinity. Despite the progress in improving abiotic stress tolerance in crops, the assessment of stress tolerance in transgenic plants has been largely carried out in the laboratory and/or greenhouse under controlled conditions where plants are not exposed to other stress conditions related to the field. In many cases, laboratory stress tolerance assays use nutrient-rich media containing sucrose, which shows no relationship with field conditions. Therefore, the abiotic stress tolerance of transgenic crops must be evaluated in the field, and more importantly, the yield potentials of these transgenic crops should be assessed. Evaluating the performance of transgenic plants in the field is challenging due to the complexity and variability of stresses in the field (Esmaeili et al., 2021). The concurrence of multiple stresses and the potential for interactions with other field factors such as soil fertility, light intensity, soil pH, presence of different salts and toxic elements, temperature, humidity, mechanical stress from strong wind, and transpiration and water loss make plant stress evaluation in the field very difficult (Yamaguchi and Blumwald, 2005). Therefore, developing multi-stress tolerant crops, particularly those that could quickly adapt to the changing environments, should be prioritized.
Furthermore, most studies on developing abiotic stress-tolerant plants have been carried out on model plants such as Arabidopsis, tobacco, and rice. Thus, it is critical to generate transgenic crops using the knowledge we learned from studying model plants (Mittler, 2006). Although rice is an important crop in addition to being a model plant for monocots, transforming other monocots has been time-consuming, expensive, and challenging (Yamaguchi and Blumwald, 2005). Despite the tremendous progress in improving crops through the biotechnology approach, a better understanding of plant stress response and the tolerance mechanism is urgently needed (Zandalinas et al., 2022a; Zandalinas et al., 2022b). Indeed, a more precise and comprehensive understanding of the underlying mechanisms of plant response to stresses will help us to design climate-resilient crops for the future (Dita et al., 2006; Tian et al., 2021; Rivero et al., 2022). The emergence of several functional tools over the past decades has assisted researchers in unraveling the underlying mechanisms of stress tolerance in plants. For instance, marker-assisted selection (MAS) enabled researchers to construct associated gene maps and identify quantitative trait loci (QTL) responsible for improved stress tolerance. The genome-wide association study in crops covering the whole genome could detect major QTLs in crops responsible for enhancing abiotic stress tolerance (Abdelraheem et al., 2019). Other emerging technologies, including gene editing tools such as CRISPR/Cas9 and Transcription Activator-Like Effector Nucleases (TALEN), are examples of new technologies that give promise to the future of crop biotechnology. The advantage of CRISPR/Cas9 in genome editing involves alteration of a few nucleotides in the original DNA of an organism without introduction of a large foreign DNA fragment, which could help the acceptance of genetically modified organisms (GMO) in the rest of the world as only minimal changes are made in the crop genomes.
Crops lack many beneficial traits of their wild-type relative species, such as disease resistance and abiotic stress tolerance, due to extensive breeding and domestication that occurred over millennia (Zsögön et al., 2018). Recent studies on the domestication of crops show that only a limited number of genes have been altered through this process, and in fact, some of these genes are highly conserved among different species. This collective evidence has driven an increasing interest in de novo domestication (dnD) and re-domestication of crops (Fernie and Yan, 2019). Recent advances in gene editing technology and the de novo domestication approach have opened promising avenues to generating crops by altering domestication-related genes in wild species. The de novo domestication platform uses CRISPR–Cas9 targeted genome editing technology to manipulate crops’ wild relatives by targeting specific genes linked to stress tolerance and/or nutritional quality (Zsögön et al., 2017). Although there are not many reports on using de novo domestication, Zsögön et al. (2017) examined the domestication of the world’s six main crops, including maize, rice, and wheat. They suggested that the key monogenic traits could be introduced into wild relatives of crops via gene editing technique (Zsögön et al., 2017). Recently, the wild relative of the present-day tomato crop, wild Solanum pimpinellifolium was domesticated de novo via editing six loci. The engineered lines produced fruits threefold and tenfold larger in size and number, respectively, compared with the wild parent (Zsögön et al., 2018). Therefore, the genetic diversity of wild species can be utilized in molecular breeding for the domestication of wild plants by targeting agronomically valuable traits such as improved stress tolerance, nutritional features, and enhanced yields (Gasparini et al., 2021).
On the other hand, the most globally consumed crops, including rice, maize, wheat, soybean, sugarcane, potato, and tomato, are mainly exotic species. Recent reports demonstrate that re-domestication of these plant species through gene editing or selective breeding could be an alternative approach to growing exotic species (Fernie and Yan, 2019). It has been proved that the gene editing strategy is a valuable and reliable technique to improve target traits accurately and rapidly in plant species. Recent progress in knowledge-driven re-domestication and de novo domestication of crops opens up promising doors to achieve improved crop tolerance and yield production.
Reports suggest that “gene discovery” is an important limiting factor in the genetic engineering of plants. Whole-genome sequencing, along with omics technology (e.g., genomics, proteomics, and metabolomics), will likely lead to identifying different genes expressed under stress conditions. Such novel genes could be used as potential candidates to enhance plant stress tolerance. Identification of stress-related metabolites in crops can be essential in improving plant stress tolerance. Because overexpression of single genes in crops leads to a limited increase in stress tolerance, the gene pyramiding approach in which several functionally related genes are simultaneously overexpressed appears to be a more logical strategy. Nevertheless, recent progress in improving plant tolerance against a combination of abiotic stresses via multi-gene assembly raises a solid hope to tackle the negative impacts of the abiotic stresses on agricultural production. Thus, the main goal in attaining sustainable agriculture is to gather and implement the knowledge we gained to develop crops that can grow and reproduce successfully in a complex environment.
NE wrote the first draft, GS and HZ participated in discussion and revision of the manuscript. All authors approved the final manuscript.
This work was partly supported by grants to Guoxin Shen from the Key Technologies R & D Program for Crop Breeding of Zhejiang Province (2021C02072-5) and the Natural Science Foundation of China (31402140).
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Abdelraheem, A., Esmaeili, N., O’Connell, M., Zhang, J. (2019). Progress and perspective on drought and salt stress tolerance in cotton. Ind. Crops Prod. 130, 118–129. doi: 10.1016/j.indcrop.2018.12.070
Ahmad, R., Kim, Y. H., Kim, M. D., Kwon, S. Y., Cho, K., Lee, H. S., et al. (2010). Simultaneous expression of choline oxidase, superoxide dismutase and ascorbate peroxidase in potato plant chloroplasts provides synergistically enhanced protection against various abiotic stresses. Physiol. Plant 138 (4), 520–533. doi: 10.1111/j.1399-3054.2010.01348.x
Anjum, N. A., Sharma, P., Gill, S. S., Hasanuzzaman, M., Khan, E. A., Kachhap, K., et al. (2016). Catalase and ascorbate peroxidase-representative H2O2-detoxifying heme enzymes in plants. Environ. Sci. pollut. Res. 23 (19), 19002–19029. doi: 10.1007/s11356-016-7309-6
Anoop, N., Gupta, A. K. (2003). Transgenic indica rice cv IR-50 over-expressing Vigna aconitifolia Δ1-pyrroline-5-carboxylate synthetase cDNA shows tolerance to high salt. J. Plant Biochem. Biotechnol. 12 (2), 09–116. doi: 10.1007/BF03263170
Apse, M. P., Aharon, G. S., Snedden, W. A., Blumwald, E. (1999). Salt tolerance conferred by overexpression of a vacuolar Na+/H+ antiport in Arabidopsis. Science 285 (5431), 1256–1258. doi: 10.1126/science.285.5431.1256
Ashraf, M., Akram, N. A. (2009). Improving salinity tolerance of plants through conventional breeding and genetic engineering: An analytical comparison. Biotechnol. Adv. 27 (6), 744–752. doi: 10.1016/j.biotechadv.2009.05.026
Baghour, M., Gálvez, F. J., Sánchez, M. E., Aranda, M. N., Venema, K., Rodríguez-Rosales, M. P. (2019). Overexpression of LeNHX2 and SlSOS2 increases salt tolerance and fruit production in double transgenic tomato plants. Plant Physiol. Biochem. 135, 77–86. doi: 10.1016/j.plaphy.2018.11.028
Balasubramaniam, T., Wijewardene, I., Hu, R., Shen, G., Zhang, J., Zhang, H. (2022). Co-Overexpression of AVP1, PP2A-C5, and AtCLCc in arabidopsis thaliana greatly increases tolerance to salt and drought stresses. Environ. Exp. Bot. 104934. doi: 10.1016/j.envexpbot.2022.104934
Bao, A. K., Wang, S. M., Wu, G. Q., Xi, J. J., Zhang, J. L., Wang, C. M. (2009). Overexpression of the Arabidopsis h+-PPase enhanced resistance to salt and drought stress in transgenic alfalfa (Medicago sativa l.). Plant Sci. 176 (2), 232–240. doi: 10.1016/j.plantsci.2008.10.009
Basso, M. F., Costa, J. A., Ribeiro, T. P., Arraes, F. B. M., Lourenço-Tessutti, I. T., Macedo, A. F., et al. (2021). Overexpression of the CaHB12 transcription factor in cotton (Gossypium hirsutum) improves drought tolerance. Plant Physiol. Biochem. 165, 80–93. doi: 10.1016/j.plaphy.2021.05.009
Batcho, A. A., Sarwar, M. B., Rashid, B., Hassan, S., Husnain, T. (2021). Heat shock protein gene identified from agave sisalana (AsHSP70) confers heat stress tolerance in transgenic cotton (Gossypium hirsutum). Theor. Exp. Plant Physiol. 33 (2), 141–156. doi: 10.1007/s40626-021-00200-6
Battisti, D. S., Naylor, R. L. (2009). Historical warnings of future food insecurity with unprecedented seasonal heat. Science 323, 240–244. doi: 10.1126/science.1164363
Behnam, B., Kikuchi, A., Celebi-Toprak, F., Yamanaka, S., Kasuga, M., Yamaguchi-Shinozaki, K., et al. (2006). The Arabidopsis DREB1A gene driven by the stress-inducible rd29A promoter increases salt-stress tolerance in proportion to its copy number in tetrasomic tetraploid potato (Solanum tuberosum). Plant Biotechnol. 23, 169–177. doi: 10.5511/plantbiotechnology.23.169
Bhatnagar-Mathur, P., Devi, M. J., Reddy, D. S., Lavanya, M., Vadez, V., Serraj, R., et al. (2007). Stress-inducible expression of AtDREB1A in transgenic peanut (Arachis hypogaea l.) increases transpiration efficiency under water-limiting conditions. Plant Cell Rep. 26 (12), 2071–2082. doi: 10.1007/s00299-007-0406-8
Bouzroud, S., Gasparini, K., Hu, G., Barbosa, M. A. M., Rosa, B. L., Fahr, M., et al. (2020). Down regulation and loss of auxin response factor 4 function using CRISPR/Cas9 alters plant growth, stomatal function and improves tomato tolerance to salinity and osmotic stress. Genes 11 (3), 272. doi: 10.3390/genes11030272
Boyer, J. S. (1982). Plant productivity and environment. Science. 218, 443–448. doi: 10.1126/science.218.4571.443
Burke, J. J., Chen, J. (2015). Enhancement of reproductive heat tolerance in plants. PloS One 10 (4), e0122933. doi: 10.1371/journal.pone.0122933
Cao, D., Hou, W., Liu, W., Yao, W., Wu, C., Liu, X., et al. (2011). Overexpression of TaNHX2 enhances salt tolerance of ‘composite’ and whole transgenic soybean plants. Plant Cell Tissue Organ Cult. 107 (3), 541–552. doi: 10.1007/s11240-011-0005-9
Chang, Y., Nguyen, B. H., Xie, Y., Xiao, B., Tang, N., Zhu, W., et al. (2017). Co-Overexpression of the constitutively active form of OsbZIP46 and ABA-activated protein kinase SAPK6 improves drought and temperature stress resistance in rice. Front. Plant Sci. 8. doi: 10.3389/fpls.2017.01102
Chen, H., An, R., Tang, J. H., Cui, X. H., Hao, F. S., Chen, J., et al. (2007a). Over-expression of a vacuolar Na+/H+ antiporter gene improves salt tolerance in an upland rice. Mol. Breed. 19 (3), 215–225. doi: 10.1007/s11032-006-9048-8
Chen, H., Feng, H., Zhang, X., Zhang, C., Wang, T., Dong, J. (2019). An Arabidopsis E3 ligase HUB 2 increases histone H2B monoubiquitination and enhances drought tolerance in transgenic cotton. Plant Biotechnol. J. 17 (3), 556–568. doi: 10.1111/pbi.12998
Chen, T., Li, W., Hu, X., Guo, J., Liu, A., Zhang, B. (2015). A cotton MYB transcription factor, GbMYB5, is positively involved in plant adaptive response to drought stress. Plant Cell Physiol. 56 (5), 917–929. doi: 10.1093/pcp/pcv019
Chen, M., Wang, Q. Y., Cheng, X. G., Xu, Z. S., Li, L. C., Ye, X. G., et al. (2007b). GmDREB2, a soybean DRE-binding transcription factor, conferred drought and high-salt tolerance in transgenic plants. Biochem. Biophys. Res. Commun. 353 (2), 299–305. doi: 10.1016/j.bbrc.2006.12.027
Chen, M., Zhu, X., Liu, X., Wu, C., Yu, C., Hu, G., et al. (2021) 3347. Knockout of auxin response factor SlARF4 improves tomato resistance to water deficit. Int. J. Mol. Sci. 22 (7), 3347. doi: 10.3390/ijms22073347
Cheng, C., Zhang, Y., Chen, X., Song, J., Guo, Z., Li, K., et al. (2018). Co-Expression of AtNHX1 and TsVP improves the salt tolerance of transgenic cotton and increases seed cotton yield in a saline field. Mol. Breed. 38, 19. doi: 10.1007/s11032-018-0774-5
Chinnusamy, V., Zhu, J., Zhu, J. K. (2007). Cold stress regulation of gene expression in plants. Trends Plant Sci. 12 (10), 444–451. doi: 10.1016/j.tplants.2007.07.002
Datta, K., Baisakh, N., Ganguly, M., Krishnan, S., Yamaguchi Shinozaki, K., Datta, S. K. (2012). Overexpression of Arabidopsis and rice stress genes’ inducible transcription factor confers drought and salinity tolerance to rice. Plant Biotechnol. J. 10 (5), 579–586. doi: 10.1111/j.1467-7652.2012.00688.x
Devireddy, A. R., Zandalinas, S. I., Fichman., Y., Mittler, R. (2021). Integration of reactive oxygen species and hormone signaling during abiotic stress. Plant J. 105, 459–476. doi: 10.1111/tpj.15010
Dita, M. A., Rispail, N., Prats, E., Rubiales, D., Singh, K. B. (2006). Biotechnology approaches to overcome biotic and abiotic stress constraints in legumes. Euphytica 147 (1), 1–24. doi: 10.1007/s10681-006-6156-9
El-Esawi, M. A., Alayafi, A. A. (2019). Overexpression of StDREB2 transcription factor enhances drought stress tolerance in cotton (Gossypium barbadense l.). Genes 10 (2), 142. doi: 10.3390/genes10020142
El-Esawi, M. A., Al-Ghamdi, A. A., Ali, H. M., Ahmad, M. (2019). Overexpression of AtWRKY30 transcription factor enhances heat and drought stress tolerance in wheat (Triticum aestivum l.). Genes 10 (2), 163. doi: 10.3390/genes10020163
Esmaeili, N., Cai, Y., Tang, F., Zhu, X., Smith, J., Mishra, N., et al. (2021). Towards doubling fibre yield for cotton in the semiarid agricultural area by increasing tolerance to drought, heat and salinity simultaneously. Plant Biotechnol. J. 19, 462–476. doi: 10.1111/pbi.13476
Esmaeili, N., Yang, X., Cai, Y., Sun, L., Zhu, X., Shen, G., et al. (2019). Co-Overexpression of AVP1 and OsSIZ1 in Arabidopsis substantially enhances plant tolerance to drought, salt, and heat stresses. Sci. Rep. 9 (1), 1–15. doi: 10.1038/s41598-019-44062-0
Fernie, A. R., Yan, J. (2019). De novo domestication: an alternative route toward new crops for the future. Mol. Plant 12 (5), 615–631. doi: 10.1016/j.molp.2019.03.016
Fukuda, A., Nakamura, A., Tagiri, A., Tanaka, H., Miyao, A., Hirochika, H., et al. (2004). Function, intracellular localization and the importance in salt tolerance of a vacuolar Na+/H+ antiporter from rice. Plant Cell Physiol. 45 (2), 146–159. doi: 10.1093/pcp/pch014
Gao, F., Gao, Q., Duan, X., Yue, G., Yang, A., Zhang, J. (2006). Cloning of an h+-PPase gene from Thellungiella halophila and its heterologous expression to improve tobacco salt tolerance. J. Exp. Bot. 57 (12), 3259–3270. doi: 10.1093/jxb/erl090
Gao, S., Yuan, L., Zhai, H., Liu, >C.l., He, S. Z., Liu, Q. C. (2012). Overexpression of SOS genes enhanced salt tolerance in sweetpotato. J. Integr. Agric. 11 (3), 378–386. doi: 10.1016/S2095-3119(12)60022-7
Gasparini, K., dos Reis Moreira, J., Peres, L. E. P., Zsögön, A. (2021). De novo domestication of wild species to create crops with increased resilience and nutritional value. Curr. Opin. Plant Biol. 60, 102006. doi: 10.1016/j.pbi.2021.102006
Gaxiola, R. A., Li, J., Undurraga, S., Dang, L. M., Allen, G. J., Alper, S. L., Fink, G. R. (2001). Drought- and salt-tolerant plants result from overexpression of the AVP1 h+-pump. Proc. Natl. Acad. Sci. U. S. A. 98 (20), 11444–11449. doi: 10.1073/pnas.191389398
Ghosh, U. K., Islam, M. N., Siddiqui, M. N., Cao, X., Khan, M. A. R. (2022). Proline, a multifaceted signalling molecule in plant responses to abiotic stress: understanding the physiological mechanisms. Plant Biol. 24 (2), 227–239. doi: 10.1111/plb.13363
Gill, S. S., Tuteja, N. (2010). Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiol. Biochem. 48, 909–930. doi: 10.1016/j.plaphy.2010.08.016
Golldack, D., Lüking, I., Yang, O. (2011). Plant tolerance to drought and salinity: stress regulating transcription factors and their functional significance in the cellular transcriptional network. Plant Cell Rep. 30 (8), 1383–1391. doi: 10.1007/s00299-011-1068-0
González, F. G., Capella, M., Ribichich, K. F., Curín, F., Giacomelli, J. I., Ayala, F., et al. (2019). Field-grown transgenic wheat expressing the sunflower gene HaHB4 significantly outyields the wild type. J. Exp. Bot. 70 (5), 1669–1681. doi: 10.1093/jxb/erz037
Grene, R. (2002). Oxidative stress and acclimation mechanisms in plants. Arabidopsis Book (Derwood, MD: American Society of Plant Biologists), 1, e0036. doi: 10.1199/tab.0036.1
Grennan, A. K. (2007). The role of trehalose biosynthesis in plants. Plant Physiol. 144 (1), 3–5. doi: 10.1104/pp.104.900223
Grover, A., Mittal, D., Negi, M., Lavania, D. (2013). Generating high temperature tolerant transgenic plants: achievements and challenges. Plant Sci. 205, 38–47. doi: 10.1016/j.plantsci.2013.01.005
He, C., Yan, J., Shen, G., Fu, L., Holaday, A. S., Auld, D., et al. (2005). Expression of an Arabidopsis vacuolar sodium/proton antiporter gene in cotton improves photosynthetic performance under salt conditions and increases fiber yield in the field. Plant Cell Physiol. 46 (11), 1848–1854. doi: 10.1093/pcp/pci201
He, R., Zhuang, Y., Cai, Y., Agüero, C. B., Liu, S., Wu, J., et al. (2018). Overexpression of 9-cis-epoxycarotenoid dioxygenase cis gene in grapevine increases drought tolerance and results in pleiotropic effects. Front. Plant Sci. 9. doi: 10.3389/fpls.2018.00970
Hmida-Sayari, A., Gargouri-Bouzid, R., Bidani, A., Jaoua, L., Savouré, A., Jaoua, S. (2005). Overexpression of Δ1-pyrroline-5-carboxylate synthetase increases proline production and confers salt tolerance in transgenic potato plants. Plant Sci. 169 (4), 746–752. doi: 10.1016/j.plantsci.2005.05.025
Hong, Y., Zhang, H., Huang, L., Li, D., Song, F. (2016). Overexpression of a stress-responsive NAC transcription factor gene ONAC022 improves drought and salt tolerance in rice. Front. Plant Sci. 7. doi: 10.3389/fpls.2016.00004
Hsieh, T. H., Lee, J. T., Charng, Y. Y., Chan, M. T. (2002). Tomato plants ectopically expressing Arabidopsis CBF1 show enhanced resistance to water deficit stress. Plant Physiol. 130 (2), 618–626. doi: 10.1104/pp.006783
Hu, H., Dai, M., Yao, J., Xiao, B., Li, X., Zhang, Q., et al. (2006). Overexpressing a NAM, ATAF, and CUC (NAC) transcription factor enhances drought resistance and salt tolerance in rice. Proc. Natl. Acad. Sci. U. S. A. 103 (35), 12987–12992. doi: 10.1073/pnas.0604882103
Huang, Y., Guo, Y., Liu, Y., Zhang, F., Wang, Z., Wang, H., et al. (2018). 9-cis-epoxycarotenoid dioxygenase 3 regulates plant growth and enhances multi-abiotic stress tolerance in rice. Front. Plant Sci. 9. doi: 10.3389/fpls.2018.00162
Huang, Y., Jiao, Y., Xie, N., Guo, Y., Zhang, F., Xiang, Z., et al. (2019). OsNCED5, a 9-cis-epoxycarotenoid dioxygenase gene, regulates salt and water stress tolerance and leaf senescence in rice. Plant Sci. 287, 110188. doi: 10.1016/j.plantsci.2019.110188
Huertas, R., Olias, R., Eljakaoui, Z., Gálvez, F. J., Li, J. U. N., De Morales, P. A., et al. (2012). Overexpression of SlSOS2 (SlCIPK24) confers salt tolerance to transgenic tomato. Plant Cell Environ. 35 (8), 1467–1482. doi: 10.1111/j.1365-3040.2012.02504.x
Huertas, R., Rubio, L., Cagnac, O., Garcia-Sanchez, M. J., Alché, J. D. D., Venema, K., et al. (2013). The K+/H+ antiporter LeNHX2 increases salt tolerance by improving k+ homeostasis in transgenic tomato. Plant Cell Environ. 36 (12), 2135–2149. doi: 10.1111/pce.12109
Hu, H., You, J., Fang, Y., Zhu, X., Qi, Z., Xiong, L. (2008). Characterization of transcription factor gene SNAC2 conferring cold and salt tolerance in rice. Plant Mol. Biol. 67 (1), 169–181. doi: 10.1007/s11103-008-9309-5
Hu, H., Xiong, L. (2014). Genetic engineering and breeding of drought-resistant crops. Annu. Rev. Plant Biol. 65, 715–741. doi: 10.1146/annurev-arplant-050213-040000
IPCC, Core Writing Team (2014). Climate change 2014: Synthesis report. contribution of working groups I, II and III to the fifth assessment report of the intergovernmental panel on climate change. Eds. Pachauri, R. K., Meyer, L. A. (Geneva, Switzerland: IPCC), 151 pp.
ISAAA (2019). Global Status of Commercialized Biotech/GM Crops in 2019: Biotech Crops Drive SocioEconomic Development and Sustainable Environment in the New Frontier. The International Service for the Acquisition of Agri-biotech Applications (ISAAA). Brief No. 55. (ISAAA: Ithaca, NY).
Jaganathan, D., Ramasamy, K., Sellamuthu, G., Jayabalan, S., Venkataraman, G. (2018). CRISPR for crop improvement: an update review. Front. Plant Sci. 9. doi: 10.3389/fpls.2018.00985
James, D., Borphukan, B., Fartyal, D., Ram, B., Singh, J., Manna, et al. (2018). Concurrent overexpression of OsGS1;1 and OsGS2 genes in transgenic rice (Oryza sativa l.): impact on tolerance to abiotic stresses. Front. Plant Sci. 9. doi: 10.3389/fpls.2018.00786
Jang, I. C., Oh, S. J., Seo, J. S., Choi, W. B., Song, S. I., Kim, C. H., et al. (2003). Expression of a bifunctional fusion of the Escherichia coli genes for trehalose-6-phosphate synthase and trehalose-6-phosphate phosphatase in transgenic rice plants increases trehalose accumulation and abiotic stress tolerance without stunting growth. Plant Physiol. 131 (2), 516–524. doi: 10.1104/pp.007237
Ji, Y., Li, Q., Liu, G., Selvaraj, G., Zheng, Z., Zou, J., et al. (2019). Roles of cytosolic glutamine synthetases in Arabidopsis development and stress responses. Plant Cell Physiol. 60 (3), 657–671. doi: 10.1093/pcp/pcy235
Katiyar-Agarwal, S., Agarwal, M., Grover, A. (2003). Heat-tolerant basmati rice engineered by over-expression of. hsp101. Plant Mol. Biol. 51 (5), 677–686. doi: 10.1023/A:1022561926676
Kerr, T. C., Abdel-Mageed, H., Aleman, L., Lee, J., Payton, P., Cryer, D., et al. (2018). Ectopic expression of two AREB/ABF orthologs increases drought tolerance in cotton (Gossypium hirsutum). Plant Cell Environ. 41 (5), 898–907. doi: 10.1111/pce.12906
Kim, J.-J., Park, S.-I., Kim, Y.-H., Park, H.-M., Kim, Y.-S., Yoon, H.-S. (2020). Overexpression of a proton pumping gene OVP1 enhances salt stress tolerance, root growth and biomass yield by regulating ion balance in rice (Oryza sativa l.). Environ. Exp. Bot. 175, 104033. doi: 10.1016/j.envexpbot.2020.104033
Kishitani, S., Takanami, T., Suzuki, M., Oikawa, M., Yokoi, S., Ishitani, M., et al. (2000). Compatibility of glycine betaine in rice plants: evaluation using transgenic rice plants with a gene for peroxisomal betaine aldehyde dehydrogenase from barley. Plant Cell Environ. 23 (1), 107–114. doi: 10.1046/j.1365-3040.2000.00527.x
Krishna, R., Ansari, W. A., Jaiswal, D. K., Singh, A. K., Verma, J. P., Singh, M. (2021). Co-Overexpression of AtDREB1A and BcZAT12 increases drought tolerance and fruit production in double transgenic tomato (Solanum lycopersicum) plants. Environ. Exp. Bot. 184, 104396. doi: 10.1016/j.envexpbot.2021.104396
Kumar, K., Gambhir, G., Dass, A., Tripathi, A. K., Singh, A., Jha, A. K., et al. (2020). Genetically modified crops: current status and future prospects. Planta 251, 91. doi: 10.1007/s00425-020-03372-8
Kumar, T., Uzma, Khan, M. R., Abbas, Z., Muhammad Ali, ,. G. (2014). Genetic improvement of sugarcane for drought and salinity stress tolerance using Arabidopsis vacuolar pyrophosphatase (AVP1) gene. Mol. Biotechnol. 56, 199–209. doi: 10.1007/s12033-013-9695-z
Kuppu, S., Mishra, N., Hu, R., Sun, L., Zhu, X., Shen, G., et al. (2013). Water-deficit inducible expression of a cytokinin biosynthetic gene IPT improves drought tolerance in cotton. PloS One 8 (5), e64190. doi: 10.1371/journal.pone.0064190
Lee, D. K., Jung, H., Jang, G., Jeong, J. S., Kim, Y. S., Ha, S. H., et al. (2016). Overexpression of the OsERF71 transcription factor alters rice root structure and drought resistance. Plant Physiol. 172 (1), 575–588. doi: 10.1104/pp.16.00379
Li, Z., Hu, Q., Zhou, M., Vandenbrink, J., Li, D., Menchyk, N., et al. (2013). Heterologous expression of OsSIZ 1, a rice SUMO e 3 ligase, enhances broad abiotic stress tolerance in transgenic creeping bentgrass. Plant Biotechnol. J. 11 (4), 432–445. doi: 10.1111/pbi.12030
Li, M., Lin, X., Li, H., Pan, X., Wu, G. (2011a). Overexpression of AtNHX5 improves tolerance to both salt and water stress in rice (Oryza sativa l.). Plant Cell Tissue Organ Cult. 107 (2), 283–293. doi: 10.1007/s11240-011-9979-6
Li, R., Liu, C., Zhao, R., Wang, L., Chen, L., Yu, W., et al. (2019). CRISPR/Cas9-mediated SlNPR1 mutagenesis reduces tomato plant drought tolerance. BMC Plant Biol. 19 (1), 1–13. doi: 10.1186/s12870-018-1627-4
Li, J., Phan, T. T., Li, Y. R., Xing, Y. X., Yang, L. T. (2018). Isolation, transformation and overexpression of sugarcane SoP5CS gene for drought tolerance improvement. Sugar Tech 20 (4), 464–473. doi: 10.1007/s12355-017-0568-9
Li, X., Xu, S., Fuhrmann-Aoyagi, M. B., Yuan, S., Iwama, T., Kobayashi, M., et al. (2022). CRISPR/Cas9 technique for temperature, drought, and salinity stress responses. Curr. Issues Mol. Biol. 44 (6), 2664–2682. doi: 10.3390/cimb44060182
Li, J., Yang, H., Ann Peer, W., Richter, G., Blakeslee, J., Bandyopadhyay, A., et al. (2005). Arabidopsis h+-PPase AVP1 regulates auxin-mediated organ development. Science 310 (5745), 121–125. doi: 10.1126/science.1115711
Li, H. W., Zang, B. S., Deng, X. W., Wang, X. P. (2011b). Overexpression of the trehalose-6-phosphate synthase gene OsTPS1 enhances abiotic stress tolerance in rice. Planta 234 (5), 1007–1018. doi: 10.1007/s00425-011-1458-0
Li, Y., Zhang, Y., Feng, F., Liang, D., Cheng, L., Ma, F., et al. (2010). Overexpression of a malus vacuolar Na+/H+ antiporter gene (MdNHX1) in apple rootstock m. 26 and its influence on salt tolerance. Plant Cell Tissue Organ Cult. 102 (3), 337–345. doi: 10.1007/s11240-010-9738-0
Liang, C. (2016a). Genetically modified crops with drought tolerance: achievements, challenges, and perspectives. In Drought Stress Tolerance Plants Vol 2, (pp. 531–547). doi: 10.1007/978-3-319-32423-4_19
Liang, C., Meng, Z., Meng, Z., Malik, W., Yan, R., Lwin, K. M., et al. (2016b). GhABF2, a bZIP transcription factor, confers drought and salinity tolerance in cotton (Gossypium hirsutum l.). Sci. Rep. 6 (1), 1–14. doi: 10.1038/srep35040
Liao, S., Qin, X., Luo, L., Han, Y., Wang, X., Usman, B., et al. (2019). CRISPR/Cas9-induced mutagenesis of semi-rolled leaf1, 2 confers curled leaf phenotype and drought tolerance by influencing protein expression patterns and ROS scavenging in rice (Oryza sativa l.). Agronomy 9 (11), 728. doi: 10.3390/agronomy9110728
Liu, W., Feng, J., Ma, W., Zhou, Y., Ma, Z. (2021). GhCLCg-1, a vacuolar chloride channel, contributes to salt tolerance by regulating ion accumulation in upland cotton. Front. Plant Sci. 2336. doi: 10.3389/fpls.2021.765173
Liu, G., Li, X., Jin, S., Liu, X., Zhu, L., Nie, Y., et al. (2014). Overexpression of rice NAC gene SNAC1 improves drought and salt tolerance by enhancing root development and reducing transpiration rate in transgenic cotton. PloS One 9 (1), e86895. doi: 10.1371/journal.pone.0086895
Lou, D., Wang, H., Liang, G., Yu, D. (2017). OsSAPK2 confers abscisic acid sensitivity and tolerance to drought stress in rice. Front. Plant Sci. 8. doi: 10.3389/fpls.2017.00993
Lv, S., Jiang, P., Nie, L., Chen, X., Tai, F., Wang, D., et al. (2015). H+-pyrophosphatase from Salicornia europaea confers tolerance to simultaneously occurring salt stress and nitrogen deficiency in Arabidopsis and wheat. Plant Cell Environ. 38 (11), 2433–2449. doi: 10.1111/pce.12557
Lv, S., Zhang, K., Gao, Q., Lian, L., Song, Y., Zhang, J. (2008). Overexpression of an h+-PPase gene from Thellungiella halophila in cotton enhances salt tolerance and improves growth and photosynthetic performance. Plant Cell Physiol. 49 (8), 1150–1164. doi: 10.1093/pcp/pcn090
Ma, H., Liu, C., Li, Z., Ran, Q., Xie, G., Wang, B., et al. (2018). ZmbZIP4 contributes to stress resistance in maize by regulating ABA synthesis and root development. Plant Physiol. 178 (2), 753–770. doi: 10.1104/pp.18.00436
Meng, X., Wang, J. R., Wang, G. D., Liang, X. Q., Li, X. D., Meng, Q. W. (2015). An R2R3-MYB gene, LeAN2, positively regulated the thermo-tolerance in transgenic tomato. J. Plant Physiol. 175, 1–8. doi: 10.1016/j.jplph.2014.09.018
Miao, C., Xiao, L., Hua, K., Zou, C., Zhao, Y., Bressan, R. A., et al. (2018). Mutations in a subfamily of abscisic acid receptor genes promote rice growth and productivity. Proc. Natl. Acad. Sci. U. S. A. 115 (23), 6058–6063. doi: 10.1073/pnas.1804774115
Mishra, N., Sun, L., Zhu, X., Smith, J., Srivastava, P. A., Yang, X., et al. (2017). Overexpression of the rice SUMO E3 ligase gene OsSIZ1 in cotton enhances drought and heat tolerance, and substantially improves fiber yields in the field under reduced irrigation and rainfed conditions. Plant Cell Physiol. 58, 735–746. doi: 10.1093/pcp/pcx032
Mittler, R. (2006). Abiotic stress, the field environment and stress combination. Trends Plant Sci. 11 (1), 15–19. doi: 10.1016/j.tplants.2005.11.002
Mittler, R., Finka, A., Goloubinoff, P. (2012). How do plants feel the heat? Trends Biochem. Sci. 37 (3), 118–125. doi: 10.1016/j.tibs.2011.11.007
Mohanty, A., Kathuria, H., Ferjani, A., Sakamoto, A., Mohanty, P., Murata, N., et al. (2002). Transgenics of an elite indica rice variety pusa basmati 1 harbouring the codA gene are highly tolerant to salt stress. Theor. Appl. Genet. 106 (1), 51–57. doi: 10.1007/s00122-002-1063-5
Molinari, M. D. C., Fuganti-Pagliarini, R., Marin, S. R. R., Ferreira, L. C., Barbosa, D. D. A., Marcolino-Gomes, J., et al. (2020). Overexpression of AtNCED3 gene improved drought tolerance in soybean in greenhouse and field conditions. Genet. Mol. Biol. 43 (3), e20190292. doi: 10.1590/1678-4685-GMB-2019-0292
Nemali, K. S., Bonin, C., Dohleman, F. G., Stephens, M., Reeves, W. R., Nelson, D. E., et al. (2014). Physiological responses related to increased grain yield under drought in the first biotechnology-derived drought-tolerant maize. Plant Cell Environ. 38 (9), 1866–1880. doi: 10.1111/pce.12446
Nguyen, Q. H., Vu, L. T. K., Nguyen, L. T. N., Pham, N. T. T., Nguyen, Y. T. H., Le, S. V., et al. (2019). Overexpression of the GmDREB6 gene enhances proline accumulation and salt tolerance in genetically modified soybean plants. Sci. Rep. 9 (1), 1–8. doi: 10.1038/s41598-019-55895-0
Ogata, T., Ishizaki, T., Fujita, M., Fujita, Y. (2020). CRISPR/Cas9-targeted mutagenesis of OsERA1 confers enhanced responses to abscisic acid and drought stress and increased primary root growth under nonstressed conditions in rice. PloS One 15 (12), e0243376. doi: 10.1371/journal.pone.0243376
Pasapula, V., Shen, G., Kuppu, S., Paez-Valencia, J., Mendoza, M., Hou, P., et al. (2011). Expression of an Arabidopsis vacuolar h+-pyrophosphatase gene (AVP1) in cotton improves drought- and salt tolerance and increases fibre yield in the field conditions. Plant Biotechnol. J. 9, 88–99. doi: 10.1111/j.1467-7652.2010.00535.x
Peleg, Z., Reguera, M., Tumimbang, E., Walia, H., Blumwald, E. (2011). Cytokinin-mediated source/sink modifications improve drought tolerance and increase grain yield in rice under water-stress. Plant Biotechnol. J. 9 (7), 747–758. doi: 10.1111/j.1467-7652.2010.00584.x
Pellegrineschi, A., Reynolds, M., Pacheco, M., Brito, R. M., Almeraya, R., Yamaguchi-Shinozaki, K., et al. (2004). Stress-induced expression in wheat of the Arabidopsis thaliana DREB1A gene delays water stress symptoms under greenhouse conditions. Genome 47 (3), 493–500. doi: 10.1139/g03-140
Qi, Y., Wang, H., Zou, Y., Liu, C., Liu, Y., Wang, Y., et al. (2011). Over-expression of mitochondrial heat shock protein 70 suppresses programmed cell death in rice. FEBS Lett. 585 (1), 231–239. doi: 10.1016/j.febslet.2010.11.051
Qin, H., Gu, Q., Kuppu, S., Sun, L., Zhu, X., Mishra, N., et al. (2013). Expression of the Arabidopsis vacuolar h+-pyrophosphatase gene AVP1 in peanut to improve drought and salt tolerance. Plant Biotechnol. Rep. 7 (3), 345–355. doi: 10.1007/s11816-012-0269-5
Qin, H., Gu, Q., Zhang, J., Sun, L., Kuppu, S., Zhang, Y., et al. (2011). Regulated expression of an isopentenyltransferase gene (IPT) in peanut significantly improves drought tolerance and increases yield under field conditions. Plant Cell Physiol. 52, 1904–1914. doi: 10.1093/pcp/pcr125
Qu, G. Z., Zang, L., Xilin, H., Zheng, T., Li, K. L. (2012). Co-Transfer of LEA and bZip genes from Tamarix confers additive salt and osmotic stress tolerance in transgenic tobacco. Plant Mol. Biol. Rep. 30, 512–518. doi: 10.1007/s11105-011-0371-9
Quan, R., Shang, M., Zhang, H., Zhao, Y., Zhang, J. (2004). Engineering of enhanced glycine betaine synthesis improves drought tolerance in maize. Plant Biotechnol. J. 2 (6), 477–486. doi: 10.1111/j.1467-7652.2004.00093.x
Rao, K. M., Raghavendra, A., Reddy, K. J. (2006). Physiology and molecular biology of stress tolerance in plants. (Springer: Dordrecht, Nertherlands). doi: 10.1007/1-4020-4225-6
Rao, M. J., Wang, L. (2021). CRISPR/Cas9 technology for improving agronomic traits and future prospective in agriculture. Planta 254 (4), 1–16. doi: 10.1007/s00425-021-03716-y
Ravikumar, G., Manimaran, P., Voleti, S. R., Subrahmanyam, D., Sundaram, R. M., Bansal, K. C., et al. (2014). Stress-inducible expression of AtDREB1A transcription factor greatly improves drought stress tolerance in transgenic indica rice. Transgenic Res. 23 (3), 421–439. doi: 10.1007/s11248-013-9776-6
Reguera, M., Peleg, Z., Blumwald, E. (2012). Targeting metabolic pathways for genetic engineering abiotic stress-tolerance in crops. Biochim. Biophys. Acta Gene Regul. Mech. 1819, 186–194. doi: 10.1016/j.bbagrm.2011.08.005
Rivero, R. M., Mittler, R., Blumwald, E., Zandalinas, S. I. (2022). Developing climate resilient crops: improving plant tolerance to stress combination. Plant J. 109, 373–389. doi: 10.1111/tpj.15483
Sawahel, W. A., Hassan, A. H. (2002). Generation of transgenic wheat plants producing high levels of the osmoprotectant proline. Biotechnol. Lett. 24 (9), 721–725. doi: 10.1023/A:1015294319114
Shafi, A., Pal, A. K., Sharma, V., Kalia, S., Kumar, S., Ahuja, P. S., et al. (2017). Transgenic potato plants overexpressing SOD and APX exhibit enhanced lignification and starch biosynthesis with improved salt stress tolerance. Plant Mol. Biol. Rep. 35 (5), 504–518. doi: 10.1007/s11105-017-1041-3
Shen, G., Kuppu, S., Sun, L., Payton, P., Zhang, H. (2015). Double overexpression of AVP1 and AtNHX1 in cotton further improves drought- and salt-tolerance in transgenic cotton plants. Plant Mol. Biol. Rep. 33, 167–177. doi: 10.1007/s11105-014-0739-8
Shi, J., Gao, H., Wang, H., Lafitte, H. R., Archibald, R. L., Yang, M., et al. (2017). ARGOS 8 variants generated by CRISPR-Cas9 improve maize grain yield under field drought stress conditions. Plant Biotechnol. J. 15 (2), 207–216. doi: 10.1111/pbi.12603
Shi, H., Ishitani, M., Kim, C., Zhu, J. K. (2000). The Arabidopsis thaliana salt tolerance gene SOS1 encodes a putative Na+/H+ antiporter. Proc. Natl. Acad. Sci. U. S. A. 97 (12), 6896–6901. doi: 10.1073/pnas.120170197
Shi, H., Lee, B. H., Wu, S. J., Zhu, J. K. (2003). Overexpression of a plasma membrane Na+/H+ antiporter gene improves salt tolerance in Arabidopsis thaliana. Nat. Biotechnol. 21 (1), 81–85. doi: 10.1038/nbt766
Shinozaki, K., Yamaguchi-Shinozaki, K. (2007). Gene networks involved in drought stress response and tolerance. J. Exp. Bot. 58, 221–227. doi: 10.1093/jxb/erl164
Shinozaki, K., Yamaguchi-Shinozaki, K., Seki, M. (2003). Regulatory network of gene expression in the drought and cold stress responses. Curr. Opin. Plant Biol. 6 (5), 410–417. doi: 10.1016/S1369-5266(03)00092-X
Shirasawa, K., Takabe, T., Takabe, T., Kishitani, S. (2006). Accumulation of glycinebetaine in rice plants that overexpress choline monooxygenase from spinach and evaluation of their tolerance to abiotic stress. Ann. Bot. (Oxford U. K.) 98 (3), 565–571. doi: 10.1093/aob/mcl126
Singh, S., Chopperla, R., Shingote, P., Chhapekar, S. S., Deshmukh, R., Khan, S., et al. (2021). Overexpression of EcDREB2A transcription factor from finger millet in tobacco enhances tolerance to heat stress through ROS scavenging. J. Biotechnol. 336, 10–24. doi: 10.1016/j.jbiotec.2021.06.013
Suárez, R., Calderón, C., Iturriaga, G. (2009). Enhanced tolerance to multiple abiotic stresses in transgenic alfalfa accumulating trehalose. Crop Sci. 49 (5), 1791–1799. doi: 10.2135/cropsci2008.09.0573
Sun, L., Pehlivan, N., Esmaeili, N., Jiang, W., Yang, X., Jarrett, P., et al. (2018). Co-Overexpression of AVP1 and PP2A-C5 in Arabidopsis makes plants tolerant to multiple abiotic stresses. Plant Sci. 274, 271–283. doi: 10.1016/j.plantsci.2018.05.026
Suprasanna, P., Nikalje, G. C., Rai, A. N. (2016). “Osmolyte accumulation and implications in plant abiotic stress tolerance,” in Osmolytes and plants acclimation to changing environment: emerging omics technologies. Eds. Iqbal, N., Nazar, R. A., Khan, N. (New Delhi: Springer). doi: 10.1007/978-81-322-2616-1_1
Tang, Y., Bao, X., Zhi, Y., Wu, Q., Guo, Y., Yin, X., et al. (2019). Overexpression of a MYB family gene, OsMYB6, increases drought and salinity stress tolerance in transgenic rice. Front. Plant Sci. 10. doi: 10.3389/fpls.2019.00168
Tang, L., Kwon, S. Y., Kim, S. H., Kim, J. S., Choi, J. S., Cho, K. Y., et al. (2006). Enhanced tolerance of transgenic potato plants expressing both superoxide dismutase and ascorbate peroxidase in chloroplasts against oxidative stress and high temperature. Plant Cell Rep. 25 (12), 1380–1386. doi: 10.1007/s00299-006-0199-1
Tester, M., Langridge, P. (2010). Breeding technologies to increase crop production in achanging world. Science 327, 818–822. doi: 10.1126/science.1183700
Thirumalaikumar, V. P., Devkar, V., Mehterov, N., Ali, S., Ozgur, R., Turkan, I., et al. (2018). NAC transcription factor JUNGBRUNNEN 1 enhances drought tolerance in tomato. Plant Biotechnol. J. 16 (2), 354–366. doi: 10.1111/pbi.12776
Tian, Z., Wang, J. W., Jiayang Li, J., Han, B. (2021). Designing future crops: challenges and strategies for sustainable agriculture. Plant J. 105, 1165–1178. doi: 10.1111/tpj.15107
Tran, M. T., Doan, D. T. H., Kim, J., Song, Y. J., Sung, Y. W., Das, S., et al. (2021). CRISPR/Cas9-based precise excision of SlHyPRP1 domain (s) to obtain salt stress-tolerant tomato. Plant Cell Rep. 40 (6), 999–1011. doi: 10.1007/s00299-020-02622-z
Ullah, A., Manghwar, H., Shaban, M., Khan, A. H., Akbar, A., Ali, U., et al. (2018). Phytohormones enhanced drought tolerance in plants: a coping strategy. Environ. Sci. pollut. Res. 25 (33), 33103–33118. doi: 10.1007/s11356-018-3364-5
Um, T. Y., Lee, S., Kim, J. K., Jang, G., Choi, Y. D. (2018). CHLORIDE CHANNEL 1 promotes drought tolerance in rice, leading to increased grain yield. Plant Biotechnol. Rep. 12 (4), 283–293. doi: 10.1007/s11816-018-0492-9
Umezawa, T., Fujita, M., Fujita, Y., Yamaguchi-Shinozaki, K., Shinozaki, K. (2006). Engineering drought tolerance in plants: discovering and tailoring genes to unlock the future. Curr. Opin. Plant Biol. 17, 113–122. doi: 10.1016/j.copbio.2006.02.002
United Nations, Department of Economic and Social Affairs, Population Division (2017). World Population Prospects: The 2017 Revision, Key Findings and Advance Tables. Working Paper No. ESA/P/WP/248. (New York: United Nations).
Van Emon, J. M. (2016). The omics revolution in agricultural research. J. Agric. Food Chem. 64, 36–44. doi: 10.1021/acs.jafc.5b04515
Vendruscolo, E. C. G., Schuster, I., Pileggi, M., Scapim, C. A., Molinari, H. B. C., Marur, C. J., et al. (2007). Stress-induced synthesis of proline confers tolerance to water deficit in transgenic wheat. J. Plant Physiol. 164 (10), 1367–1376. doi: 10.1016/j.jplph.2007.05.001
Verma, D., Singla-Pareek, S. L., Rajagopal, D., Reddy, M. K., Sopory, S. K. (2007). Functional validation of a novel isoform of Na+/H+ antiporter from Pennisetum glaucum for enhancing salinity tolerance in rice. J. Biosci. 32 (3), 621–628. doi: 10.1007/s12038-007-0061-9
Wang, Q., Guan, Y., Wu, Y., Chen, H., Chen, F., Chu, C. (2008). Overexpression of a rice OsDREB1F gene increases salt, drought, and low temperature tolerance in both Arabidopsis and rice. Plant Mol. Biol. 67 (6), 589–602. doi: 10.1007/s11103-008-9340-6
Wang, M., Mao, Y., Lu, Y., Tao, X., Zhu, J. K. (2017). Multiplex gene editing in rice using the CRISPR-Cpf1 system. Mol. Plant 10 (7), 1011–1013. doi: 10.1016/j.molp.2017.03.001
Wang, F. Z., Wang, Q. B., Kwon, S. Y., Kwak, S. S., Su, W. A. (2005). Enhanced drought tolerance of transgenic rice plants expressing a pea manganese superoxide dismutase. J. Plant Physiol. 162 (4), 465–472. doi: 10.1016/j.jplph.2004.09.009
Wang, Y., Wisniewski, M., Meilan, R., Cui, M., Fuchigami, L. (2006). Transgenic tomato (Lycopersicon esculentum) overexpressing cAPX exhibits enhanced tolerance to UV-b and heat stress. J. Appl. Hortic. 8, 87–90. doi: 10.37855/jah.2006.v08i02.21
Wang, T., Xun, H., Wang, W., Ding, X., Tian, H., Hussain, S., et al. (2021). Mutation of GmAITR genes by CRISPR/Cas9 genome editing results in enhanced salinity stress tolerance in soybean. Front. Plant Sci. 2752. doi: 10.3389/fpls.2021.779598
Wang, Z., Yang, C., Chen, H., Wang, P., Wang, P., Song, C., et al. (2018). Multi-gene co-expression can improve comprehensive resistance to multiple abiotic stresses in Brassica napus l. Plant Sci. 274, 410–419. doi: 10.1016/j.plantsci.2018.06.014
Wang, G. P., Zhang, X. Y., Li, F., Luo, Y., Wang, W. (2010). Overaccumulation of glycine betaine enhances tolerance to drought and heat stress in wheat leaves in the protection of photosynthesis. Photosynthetica 48 (1), 117–126. doi: 10.1007/s11099-010-0016-5
Wei, P., Che, B., Shen, L., Cui, Y., Wu, S., Cheng, C., et al. (2019). Identification and functional characterization of the chloride channel gene, GsCLC-c2 from wild soybean. BMC Plant Biol. 19 (1), 1–15. doi: 10.1186/s12870-019-1732-z
Wei, A., He, C., Li, B., Li, N., Zhang, J. (2011). The pyramid of transgenes TsVP and BetA effectively enhances the drought tolerance of maize plants. Plant Biotechnol. J. 9 (2), 216–229. doi: 10.1111/j.1467-7652.2010.00548.x
Wei, P. P., Wang, L. C., Liu, A. L., Yu, B. J., Lam, H. M. (2016). GmCLC1 confers enhanced salt tolerance through regulating chloride accumulation in soybean. Front. Plant Sci. 7, 1082. doi: 10.3389/fpls.2016.01082
Wijewardene, I., Mishra, N., Sun, L., Smith, J., Zhu, X., Payton, P., et al. (2020). Improving drought-, salinity-, and heat-tolerance in transgenic plants by co-overexpressing Arabidopsis vacuolar pyrophosphatase gene AVP1 and larrea rubisco activase gene RCA. Plant Sci. 296, 110499. doi: 10.1016/j.plantsci.2020.110499
Wu, X., Shiroto, Y., Kishitani, S., Ito, Y., Toriyama, K. (2009). Enhanced heat and drought tolerance in transgenic rice seedlings overexpressing OsWRKY11 under the control of HSP101 promoter. Plant Cell Rep. 28 (1), 21–30. doi: 10.1007/s00299-008-0614-x
Wu, Y., Zhang, D., Chu, J. Y., Boyle, P., Wang, Y., Brindle, I. D., et al. (2012). The arabidopsis NPR1 protein is a receptor for the plant defense hormone salicylic acid. Cell Rep. 1 (6), 639–647. doi: 10.1016/j.celrep.2012.05.008
Xue, Z. Y., Zhi, D. Y., Xue, G. P., Zhang, H., Zhao, Y. X., Xia, G. M. (2004). Enhanced salt tolerance of transgenic wheat (Tritivum aestivum l.) expressing a vacuolar Na+/H+ antiporter gene with improved grain yields in saline soils in the field and a reduced level of leaf na+. Plant Sci. 167 (4), 849–859. doi: 10.1016/j.plantsci.2004.05.034
Yamaguchi, T., Blumwald, E. (2005). Developing salt-tolerant crop plants: challenges and opportunities. Trends Plant Sci. 10 (12), 615–620. doi: 10.1016/j.tplants.2005.10.002
Yan, H., Li, Q., Park, S. C., Wang, X., Liu, Y. J., Zhang, Y. G., et al. (2016). Overexpression of CuZnSOD and APX enhance salt stress tolerance in sweet potato. Plant Physiol. Biochem. 109, 20–27. doi: 10.1016/j.plaphy.2016.09.003
Yarra, R., He, S. J., Abbagani, S., Ma, B., Bulle, M., Zhang, W. K. (2012). Overexpression of a wheat Na+/H+ antiporter gene (TaNHX2) enhances tolerance to salt stress in transgenic tomato plants (Solanum lycopersicum l.). Plant Cell Tissue Organ Cult. 111 (1), 49–57. doi: 10.1007/s11240-012-0169-y
Yarra, R., Kirti, P. B. (2019). Expressing class I wheat NHX (TaNHX2) gene in eggplant (Solanum melongena l.) improves plant performance under saline condition. Funct. Integr. Genomics 19 (4), 541–554. doi: 10.1007/s10142-019-00656-5
Yin, X. Y., Yang, A. F., Zhang, K. W., Zhang, J. R. (2004). Production and analysis of transgenic maize with improved salt tolerance by the introduction of AtNHX1 gene. Acta Bot. Sin. (Engl. Transl.) 46 (7), 854–861.c.
Yue, Y., Zhang, M., Zhang, J., Duan, L., Li, Z. (2012). SOS1 gene overexpression increased salt tolerance in transgenic tobacco by maintaining a higher K+/Na+ ratio. J. Plant Physiol. 169 (3), 255–261. doi: 10.1016/j.jplph.2011.10.007
Yu, W., Wang, L., Zhao, R., Sheng, J., Zhang, S., Li, R., et al. (2019). Knockout of SlMAPK3 enhances tolerance to heat stress involving ROS homeostasis in tomato plants. BMC Plant Biol. 19 (1), 1–13. doi: 10.1186/s12870-019-1939-z
Zandalinas, S. I., Balfagón, D., Gómez-Cadenas, A., Mittler, R. (2022a). Plant responses to climate change: metabolic changes under combined abiotic stresses. J. Exp. Bot. 73, 3339–3354. doi: 10.1093/jxb/erac073
Zandalinas, S. I., Fritschi, F. B., Mittler, R. (2021a). Global warming, climate change, and environmental pollution: recipe for a multifactorial stress combination disaster. Trend. Plant Sci. 26 (6), 588–599. doi: 10.1016/j.tplants.2021.02.011
Zandalinas, S. I., Mittler, R. (2022b). Plant responses to multifactorial stress combination. New Phytol. 234, 1161–1167. doi: 10.1111/nph.18087
Zandalinas, S. I., Sengupta, S., Fritschi, F. B., Azad, R. K., Nechushtai, R., Mittler, R. (2021b). The impact of multifactorial stress combination on plant growth and survival. New Phytol. 230, 1034–1048. doi: 10.1111/nph.17232
Zeng, Y., Wen, J., Zhao, W., Wang, Q., Huang, W. (2020) 1663. Rational improvement of rice yield and cold tolerance by editing the three genes OsPIN5b, GS3, and OsMYB30 with the CRISPR–Cas9 system. Front. Plant Sci. 10. doi: 10.3389/fpls.2019.01663
Zhang, H. X., Blumwald, E. (2001). Transgenic salt-tolerant tomato plants accumulate salt in foliage but not in fruit. Nat. Biotechnol. 19 (8), 765–768. doi: 10.1038/90824
Zhang, H. X., Hodson, J. N., Williams, J. P., Blumwald, E. (2001). Engineering salt-tolerant brassica plants: characterization of yield and seed oil quality in transgenic plants with increased vacuolar sodium accumulation. Proc. Natl. Acad. Sci. U. S. A. 98 (22), 2832–12836. doi: 10.1073/pnas.231476498
Zhang, A., Liu, Y., Wang, F., Li, T., Chen, Z., Kong, D., et al. (2019). Enhanced rice salinity tolerance via CRISPR/Cas9-targeted mutagenesis of the OsRR22 gene. Mol. Breed. 39 (3), 1–10. doi: 10.1007/s11032-019-0954-y
Zhang, X. X., Tang, Y. J., Ma, Q. B., Yang, C. Y., Mu, Y. H., Suo, H. C., et al. (2013). OsDREB2A, a rice transcription factor, significantly affects salt tolerance in transgenic soybean. PloS One 8 (12), e83011. doi: 10.1371/journal.pone.0083011
Zhang, S., Zhuang, K., Wang, S., Lv, J., Ma, N. N., Meng, Q. (2017). A novel tomato SUMO E3 ligase, SlSIZ1, confers drought tolerance in transgenic tobacco. J. Integr. Plant Biol. 59 (2), 102–117. doi: 10.1111/jipb.12514
Zhang, G. C., Zhu, W. L., Gai, J. Y., Zhu, Y. L., Yang, L. F. (2015). Enhanced salt tolerance of transgenic vegetable soybeans resulting from overexpression of a novel Δ1-pyrroline-5-carboxylate synthetase gene from Solanum torvum Swartz. Hortic. Environ. Biotechnol. 56 (1), 94–104. doi: 10.1007/s13580-015-0084-3
Zhou, S., Chen, X., Zhang, X., Li, Y. (2008). Improved salt tolerance in tobacco plants by co-transformation of a betaine synthesis gene BADH and a vacuolar Na+/H+ antiporter gene SeNHX1. Biotechnol. Lett. 30 (2), 369–376. doi: 10.1007/s10529-007-9548-6
Zhu, J. K. (2001). Plant salt tolerance. Trends Plant Sci. 6 (2), 66–71. doi: 10.1016/S1360-1385(00)01838-0
Zhu, X., Sun, L., Kuppu, S., Hu, R., Mishra, N., Smith, J., et al. (2018). The yield difference between wild-type cotton and transgenic cotton that expresses IPT depends on when water-deficit stress is applied. Sci. Rep. 8, 2538. doi: 10.1038/s41598-018-20944-7
Zhu, B., Ye, C., Lü, H., Chen, X., Chai, G., Chen, J., et al. (2006). Identification and characterization of a novel heat shock transcription factor gene, GmHsfA1, in soybeans (Glycine max). J. Plant Res. 119 (3), 247–256. doi: 10.1007/s10265-006-0267-1
Zsögön, A., Čermák, T., Naves, E. R., Notini, M. M., Edel, K. H., Weinl, S., et al. (2018). De novo domestication of wild tomato using genome editing. Nat. Biotechnol. 36 (12), 1211–1216. doi: 10.1038/nbt.4272
Keywords: abiotic stresses, crop production, drought stress, heat stress, salinity stress, transgenic plants
Citation: Esmaeili N, Shen G and Zhang H (2022) Genetic manipulation for abiotic stress resistance traits in crops. Front. Plant Sci. 13:1011985. doi: 10.3389/fpls.2022.1011985
Received: 04 August 2022; Accepted: 05 September 2022;
Published: 21 September 2022.
Edited by:
Dayong Zhang, Nanjing Agricultural University, ChinaReviewed by:
Zuo Ren Yang, State Key Laboratory of Cotton Biology (CAAS), ChinaCopyright © 2022 Esmaeili, Shen and Zhang. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Nardana Esmaeili, bmFyZGFuYWVAdXBlbm4uZWR1; Guoxin Shen, Z3VveGluc2hlbkBnbWFpbC5jb20=
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.