 Xi Chen
Xi Chen Xin Chen
Xin Chen Dongming Fang
Dongming Fang- Jiyang College of Zhejiang A&F University, Zhuji, Zhejiang, China
In recent decades, Moso bamboo has been largely increasing in the subtropical area of China, raising ecological concerns about its invasion into other native forest ecosystems. One concern is whether the invasion of Moso bamboo significantly simplifies forest community composition and structure and declines biomass. This study adopted the space-for-time method to investigate a secondary coniferous and broad-leaved mixed forest (SF) being invaded by an adjacent Moso bamboo forest (MB) in the Wuxie forest reserve, Zhejiang Province. Three plots were established in each SF, MB, and transitional forest. The results showed that the species composition and species dominance of the arborous layer changed significantly (P < 0.05), which was indicated by the significantly decreased species richness (Margalef index, Shannon–Wiener index, and Simpson index) and evenness (Pielou evenness index). In contrast, the species richness of the shrub and herbaceous layers had two divergent indications (increasing or unchanged), and the evenness remained unchanged. The total and arborous-layer aboveground biomass of the forest community has had no noticeable change (P < 0.05). However, the biomass of the shrub and herbaceous layers showed an increasing trend (shrub significant but herbaceous not), but they only occupied a small proportion (∼1%) of the total biomass. Finally, the aboveground biomass and the diversity index had no significant correlation in each layer and overall stands. We hope that the findings could provide a theoretical basis for the invasion mechanism and ecological consequences of the Moso bamboo invasion.
Introduction
The invasion of alien plant species into native plant communities has become a common phenomenon worldwide over the past several decades (Ramula and Pihlaja, 2012; Wang et al., 2017, 2019). Alien plant species become the dominant species of the new communities, for instance, due to their growth advantage or lack of enemies when they invade the new ecosystem (van Kleunen et al., 2010). Alien plant species inhibit the growth of other local plants, which eventually reduces biodiversity (Davies, 2011; Vilà et al., 2011) and affects the stability of community biomass (Bai et al., 2007). Similarly, native plants may become increasingly dominant in their original range, a phenomenon known as overabundance (Garrott et al., 1993), i.e., native plant invasion. The invasion of native plants into new ecosystems in the same region or country may also significantly impact biodiversity (Ouyang et al., 2016). Compared with the extensive studies that draw attention to the impact of alien plant invasion, relatively few studies consider the effects of native plant invasion (Nackley et al., 2017; Warren et al., 2017; Shi et al., 2018).
Generally, a forest with a closed canopy can resist the invasion of other species due to the weak light conditions under the canopy, which could weaken the growth of invading seedlings. Without human intervention, native species will not encroach on adjacent natural forests. However, when shade-tolerant tree species invade, the complete forest may only provide weak resistance (Martin et al., 2009). Moso bamboo (Phyllostachys pubescens) is widely distributed in southern China (Shi et al., 2020) and has competitive strengths via its strong rhizome-root system and leaf functional traits (Huang et al., 2019, 2020; Guo et al., 2021). Native Moso bamboo may invade the surrounding natural forest by itself (Bai et al., 2016) because the elongation of bamboo shoots may depend on the carbohydrate supply of bamboo rhizomes. The reserve stored in bamboo rhizomes can hold a large amount of energy and resources for shoot growth (Xue et al., 2021), largely independent of the light environment (Wang et al., 2016).
The invasion of Moso bamboo into subtropical evergreen forests has become a significant problem in many areas of southern China (Tian et al., 2020). In the past, Moso bamboo was the essential bamboo species for both shoot and timber production in China. Additionally, the Moso bamboo forest (MB) has a comparable net primary productivity (8.86 ± 3.46 Mg C ha–1 year–1), which enables it to be a vast potential carbon pool (Lin et al., 2017). Therefore, managing pure Moso bamboo forests can bring considerable income to bamboo farmers. Thus, a large area of subtropical evergreen forest in southern China has been transformed into the MB by farmers (Chen et al., 2009). However, in recent years, due to the increase in Moso bamboo production capacity and the price decline, the management level of MB in many production areas has decreased. As a result, a large area of MB has been abandoned (Jiang et al., 2015), which has exacerbated the invasion of Moso bamboo. Previous studies have shown that Moso bamboo invasion may lead to a series of ecological and environmental problems, such as simplifying forest community composition and structure (Ouyang et al., 2016), declining species diversity (Okutomi et al., 1996; Nakai and Kisanuki, 2006; Yang et al., 2011; Akutsu et al., 2012; Bai et al., 2013; Zhang and Xue, 2018), and making the forest biomass change obviously (Yang et al., 2011; Fukushima et al., 2015; Lin et al., 2017; Song et al., 2017).
As the largest terrestrial ecosystem on earth, the forest has about half of the carbon reserves on land, which plays a vital role in maintaining the ecological balance of the whole world and human survival (Pan et al., 2011). The subtropical forest is the vegetation type mainly distributed in China, with a broad distribution area and rich biodiversity (Corlett, 2013). The relationship between species diversity and aboveground biomass of natural forest communities is still controversial (Lasky et al., 2014; Bracken et al., 2017; Silva Pedro et al., 2017). Studies have shown that species richness positively affects biomass or productivity of subtropical forests (Barrufol et al., 2013; Bruelheide et al., 2014; Cavanaugh et al., 2014) or has no effect (Wu et al., 2015). The different results may be due to the forest ecosystem’s different restoration or succession times. It may also be due to the complexity of community structure and the resource effectiveness of habitat conditions (Ali and Yan, 2017; Mori et al., 2017; Ratcliffe et al., 2017; van der Sande et al., 2017). Plant invasion can significantly change the species diversity and biomass of biological communities. However, the relationship between biodiversity changes and biomass of forest communities during the Moso bamboo invasion remains largely unknown.
Therefore, we selected a typical site where Moso bamboo has been expanding into the coniferous and broad-leaved mixed forest to investigate the community characteristics. We applied the “Space for Time” method (Pickett, 1989), which assumes that spatial and temporal variation are equivalent, so the different temporal stages of the Moso bamboo invasion were simultaneously compared in the same area. The species diversity and aboveground biomass of three adjacent forest types were investigated, i.e., (1) the native secondary coniferous and broad-leaved mixed forest (SF), (2) transition tree-Moso-bamboo forest (TF), and (3) nearly pure MB. We attempted to verify three assumptions: (1) The invasion of Moso bamboo changed the species composition and reduced the diversity of the local ecosystem; (2) Moso bamboo invasion may reduce the aboveground biomass of the forest community; (3) The changes in species diversity of forest community may have correlations with the aboveground biomass during Moso bamboo invasion. We anticipated that the findings might provide theoretical and management references to local farmers and the government.
Materials and methods
Study site
The study was performed at Wuxie Nature Reserve (120⋅2′40″E; 294⋅4′15″N) in Zhuji, Zhejiang Province, China. The area has a subtropical monsoon climate with a mean annual temperature of 16.3°C and an annual precipitation of 1,573 mm. The area has many soil types, mainly hilly red soil, with good soil fertility. The zonal vegetation is SF and MB. Due to the strong expansion ability of the Moso bamboo rhizome, a mixed forest of coniferous and broad-leaved trees and bamboos is formed between the two types of forests.
Sampling design
In May 2020, we selected three transects spanning from a SF, via a transition zone, to a MB, representing three typical stages of the Moso bamboo invasion into subtropical forests. The altitude of the plot is 210–230 m, containing SF, representing the forest stage not invaded by Moso bamboos. The dominant species in the arborous layer of SF was Schima superba, with a forest age of 30–40 years. The associated species were Liquidambar formosana and Pinus massoniana. The average height of the arborous layer was 15.6 m, and the average diameter breast height (DBH) was 20.5 cm. The TF represented the forest stage moderately invaded by Moso bamboo, and the ratio of bamboo to wood is about 4:1. The average height and DBH of Moso bamboo were 16.3 m and 11.5 cm, respectively. Finally, the MB represented the forest stage heavily invaded by Moso bamboos, and the bamboos were the species with an absolute advantage in the arborous layers. The average height of Moso bamboo was 16.9 m, and the average DBH was 12.1 cm (Table 1).

Table 1. The height and diameter breast height (DBH) class of Moso bamboo in secondary coniferous and broad-leaved mixed forest (SF), transitional forest, and Moso bamboo forest (MB) (mean ± SE).
Each transect had a width of 30 m and a length of 80 m. The utmost 5 m surrounding each horizontal transect was set as a buffer zone, and the middle area was used for setting up three 20 m × 20 m sampling plots with a 5-m spacing between them. In each sampling plot, biomass and diversity of the arborous layer were investigated. Furthermore, two 5 m × 5 m quadrats were set at the diagonal position of each sample plot and were used for shrub layer species investigation. Additionally, one 1 m × 1 m quadrats were placed in the center, and another four were established in the four corners of the sample plot, which were used for herbaceous layer species investigation. We measured the DBH of trees (>5 cm) from the arborous layer and recorded the species, DBH, height, and abundance. Woody plants with DBH less than 5 cm were measured in the shrub layer, including saplings and shrubs, and we recorded species names, ground diameter, height, abundance, and so on.
Aboveground biomass
The tree height, DBH, and bamboo age were measured through quadrat investigation. Then the standing volume of trees in the sample plot was estimated according to the Chinese standing volume table (Zeng, 2018), and the aboveground biomass of the arborous layer per unit area was calculated through the volume biomass model (Wang et al., 2009; Shen and Tang, 2019). The formulas are as follows:
where V is the standing volume, D is the diameter at breast height (DBH), H is the tree height, ci is the parameters (i = 0, 1, 2), ε is the error, B is the tree biomass, v is the volume per unit area, a and b are the constants of the corresponding forest type, W is the bamboo stem biomass, and A is the bamboo age.
The aboveground biomass of the shrub and herbaceous layers was obtained using the harvest method. We randomly selected a 5 m × 5 m investigation quadrat in each sample plot as the harvest quadrat of the shrub layer. Three vegetation survey quadrats with diagonal directions in the standard plot for the herbaceous layer were selected. The aboveground shrub and herbaceous layers were harvested in the set harvesting quadrat and brought back to the laboratory. The aboveground biomass of shrubs and herbs was measured after drying at 85°C to constant weight. We calculated the aboveground biomass of the shrub/herbaceous layer per unit area of each stand.
Species diversity
Margalef index, Shannon–Wiener index, Simpson index, and Pielou evenness index were used to estimate α diversity of community species. The formulas are as follows:
where S is the total number of species, N is the sum of the number of individuals of all species, Pi is the proportion of the number of individuals of species i to the total number of individuals in the community, that is, pi = ni/N, ni is the number of individuals of species i.
Statistical analyses
SAS 9.4 (SAS Institute Inc., Cary, NC, United States) and R Statistical Software (V4.2; R Core Team, 2022) were used to process, analyze, and graph the data.
First, the diversity indexes and biomass of the arborous layer in each sampling plot were calculated and measured. At the same time, the diversity indexes and biomass in each quadrat (5 m × 5 m for the shrub layer and 1 m × 1 m for the herbaceous layer) were calculated and measured. Then, the diversity indexes and biomass of these quadrats (two and five quadrats for the shrub and herbaceous layers, respectively) per sampling plot were averaged. In this case, we got three replicates of the diversity indexes and biomass in shrub and herbaceous layers, and they were further used for comparison between the three forest types.
Considering the limited three replicates per layer that may not meet the requirement of normal distribution by the analysis of variance (ANOVA), we selected a nonparametric test (Wilcoxon rank-sum test) to conduct the comparison in diversity indexes and biomass among SF, TF, and MB. The linear correlation between diversity index and biomass of arborous, shrub, and herb layers was separately analyzed.
Specifically, R package vegan (Dixon, 2003) was used to examine the species composition difference among the three forest types (MB, TF, and SF) with PerMANOVA (permutational multivariate ANOVA).
Results
Effects of Moso bamboo invasion on species composition and quantitative characteristics
The number of standing bamboo and trees in the three stands varied greatly. After the expansion of Moso bamboo, the number of standing bamboo and total standing trees in the MB increased significantly, increasing to 2,783 and 2,575 culms per hectare, respectively. The number of standing stems of other tree species decreased significantly; 208 plants/ha were reduced (Table 2). These results indicated that the number of standing bamboos increased during the expansion of Moso bamboo was significantly greater than the number of coniferous and broad-leaved mixed species decreased, resulting in a significant increase in the total number of standing individuals in the community.

Table 2. Effect of Moso bamboo invasion on stem number [ind./ha; (N = 3 in each group)].
Except in the herbaceous layer (P > 0.05), the species composition in the arborous, shrub, and whole layers was significantly different among the three communities (P < 0.05; Figure 1 and Table 3).
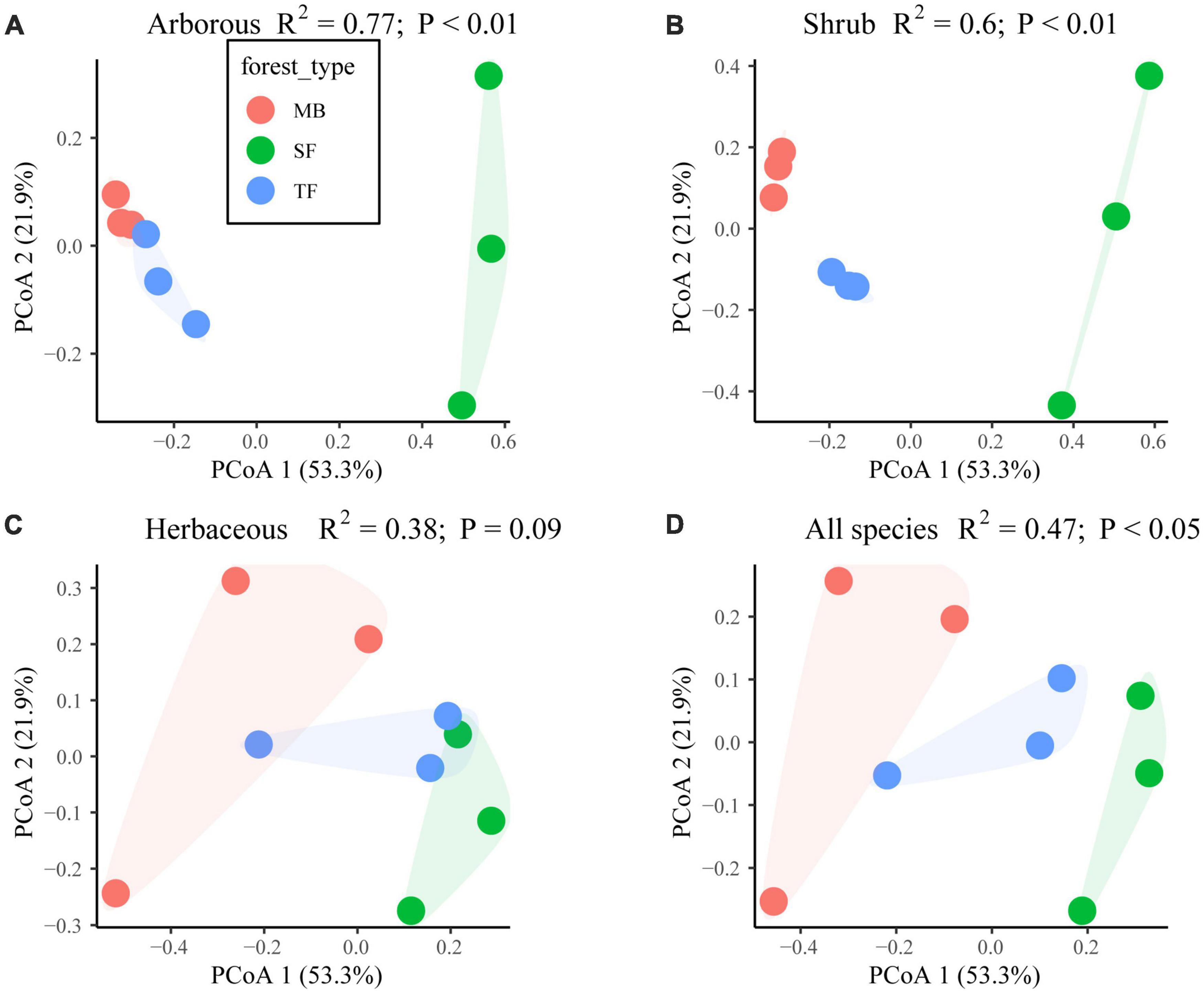
Figure 1. Effect of the Moso bamboo invasion on species composition difference of each layer among the three forest types [Moso bamboo forest (MB), transition tree-Moso-bamboo forest (TF), and secondary coniferous and broad-leaved mixed forest (SF)] in (A) arborous, (B) shrub, (C) herbaceous layers, and (D) the compound layers with all arborous, shrub, herbaceous species. Significant differences examined with PerMANOVA (permutational multivariate ANOVA) are indicated by P < 0.05 or 0.01. PCoA means principal co-ordinates analysis. PCoA1 and PCoA2 in the figures represented two components of PCoA with largest explaining power to the variation of the species composition difference.
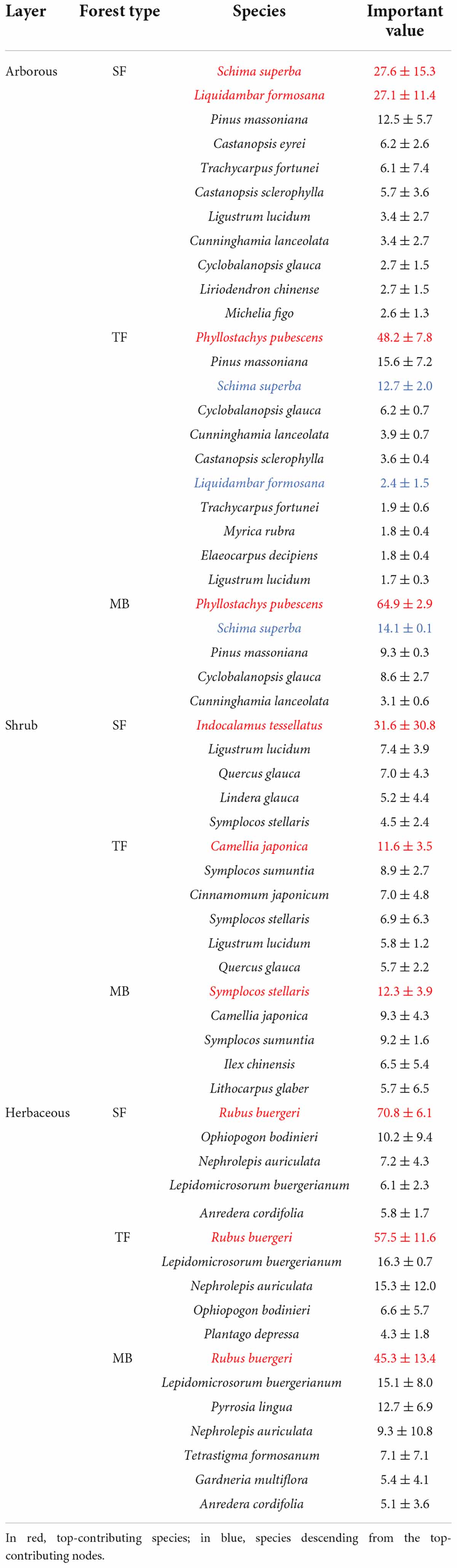
Table 3. Effects of Moso bamboo invasion on main species composition and importance value (mean ± SD).
For the arborous layer, species richness decreased from 11 in SF and TF to 5 in MB. Only three species stayed in all three communities, i.e., S. superba, P. massoniana, and Cyclobalanopsis glauca, which corresponded to dominant, inferior, and minority roles in SF with important values of 27.6, 12.5, and 2.7, respectively (Table 3). The dominant species changed from S. superba and L. formosana in SF (important values of 27.6 and 27.1, respectively) to Moso bamboo in TF and MB (important values of 48.2 and 64.9, respectively). The top contributing species S. superba in SF descended to an inferior role in TF and MB (important values of 12.7 and 14.1, respectively), and L. formosana descended to an ignorable status in TF (important value of 2.4) and disappeared in MB. P. massoniana stayed relatively stable in all three communities, with important values of 12.5, 15.6, and 9.3 in SF, TF, and MB, respectively. C. glauca is always kept as a minority, although its important values increased over the invasion stages (2.7, 6.2, and 8.6 in SF, TF, and MB, respectively).
The shrub layer’s species richness was relatively stable (5–6 species) over three communities, but none of them grew in all three communities (Table 3). In addition, the species composition differed significantly among the three forest types (P < 0.05; Figure 1 and Table 3). The dominant species were Indocalamus tessellatus, Camellia japonica, and Symplocos stellaris in SF, TF, and MB, respectively (important values of 31.6, 11.6, and 12.3, respectively). Compared with SF, the species composition was more even in both TF (important values ranging from 5.7 to 11.6) and MB (important values ranging from 5.7 to 12.3).
In the herbaceous layer of the three communities, the species richness was close to each other (five in SF and TF, and seven in MB), and the three communities had three of the same species (Rubus buergeri, Nephrolepis auriculata, and Lepidomicrosorum buergerianum). The statistical results indicated similar species composition among the communities (P > 0.05; Figure 1 and Table 3). Furthermore, the dominant species were the same, i.e., R. buergeri, but its important values dropped with the invasion progression (70.8, 57.5, and 45.3 in SF, TF, and MB, respectively).
Effects of the Moso bamboo invasion on species diversity
Changes in species diversity in the arborous layer
During the invasion of Moso bamboo into SF, species diversity in the arborous layer showed a significant decreasing trend, indicated by the significantly decreasing Shannon–Wiener index and increasing Simpson index over SF, TF, and MB (P < 0.05; Figure 2). Changes in the Margalef index seemed to support the same trend, but its values were not significantly different between the SF and TF (P > 0.05; Figure 2). Moreover, as the invasion progressed, the species’ evenness got worse, as indicated by the reducing Pielou index (P < 0.05; Figure 2).

Figure 2. Effect of the Moso bamboo invasion on species diversity of each layer. Different letters in the same column indicate significant differences at P < 0.05.
Changes in species diversity in the shrub layer
The Margalef index and Simpson index suggested no significant difference in the shrub layers among the three communities (P > 0.05; Figure 2), although the values seemed to point to increasing biodiversity over the invasion. In contrast, the Shannon–Wiener index indicated significantly higher species diversity in TF and MB than in SF (P < 0.05; Figure 2). The species evenness had no significant difference in the shrub layers among the three communities, as indicated by the Pielou index (P > 0.05; Figure 2).
Changes in species diversity in the herbaceous layer
The Margalef and Shannon–Wiener indexes suggested no difference in the herbaceous layers among the three communities (P > 0.05), while the Simpson index indicated higher diversity in MB than in SF (P < 0.05; Figure 2). Finally, species evenness had no significant difference in the herbaceous layers among the three communities, as indicated by the Pielou index (P > 0.05; Figure 2).
Effects of Moso bamboo invasion on aboveground biomass
During the invasion of Moso bamboo into SF, the aboveground biomass showed no significant differences in arborous and herbaceous layers and the overall stand between the three forest types (P > 0.05; Table 4). In contrast, the aboveground biomass in the shrub layers was significantly higher in TF and MB than in SF (P < 0.05; Table 4). The proportions of aboveground biomass in each layer to the total biomass had no significant difference among the three forest types (P > 0.05; Table 4). The arborous layers occupied the largest proportion (∼99%) of the aboveground biomass in all three forest types. With the invasion progress, the aboveground biomass of Moso bamboo in the community gradually increased to 3,538.63 ± 1,305.82 and 7,482.72 ± 2,532.43 g m–2 in the TF and MB, which correspondingly occupied 17.90 ± 6.41% and 41.56 ± 8.21% of the total aboveground biomass, respectively.

Table 4. Effect of the Moso bamboo invasion on aboveground biomass (mean ± SD; N = 3 in each group) in each layer and their contribution to the total aboveground biomass.
Relationship between diversity and aboveground biomass
The correlation between plant diversity and aboveground biomass was insignificant in the arborous, shrub, and herbaceous layers (P > 0.05; Table 5). It was also not significant in the overall stand when all layers were considered (P > 0.05; Table 5).
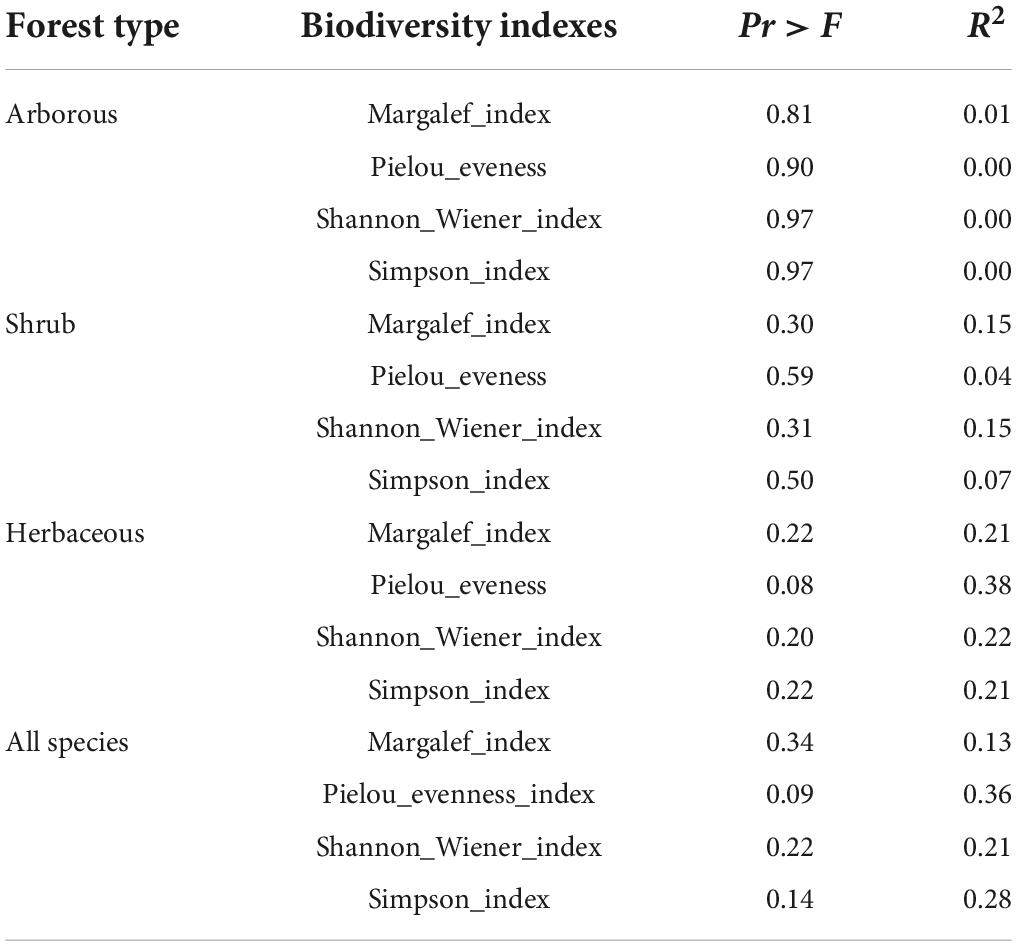
Table 5. Effect of the Moso bamboo invasion on the relationships between aboveground biomass and biodiversity indexes in each layer and the whole stand comprised of three layers (N = 9 in each group).
Discussion
Effects of Moso bamboo invasion on species composition and diversity
Species composition is not only the basis of community structure and biodiversity but also the root of forest ecosystem processes (Tilman et al., 1997). Existing studies show that both alien and native plant invasions have a certain impact on the community’s species composition (Garrott et al., 1993; Davies, 2011; Lima et al., 2012). This study found that the invasion of Moso bamboo significantly changed the species composition in the arborous and shrub layers but not in the herbaceous layer (Figures 1A–C and Table 3), which also changed the overall species composition significantly (Figure 1D). For example, the number of species had been reduced from 11 in SF to 5 in MB in the arborous layer. The important values of the main dominant arborous species S. superba, L. formosana, and P. massoniana decreased from 27.6, 27.1, and 12.5 before expansion to 14.1, 0, and 9.3, respectively (Table 3). On the contrary, Moso bamboo gradually dominated the arborous layers with important values of 48.2 and 64.9 in TF and MB, respectively. The interchanged dominance between trees and Moso bamboo in the arborous layer was consistent with several other research results (Bai et al., 2013; Yang et al., 2015; Ouyang et al., 2016). The findings supported our first hypothesis, i.e., that the invasion of Moso bamboo changed species composition, especially in the arborous and shrub layers. Although a few herbaceous species disappeared or newly appeared with the invasion, the species composition in the herbaceous layer was not significantly changed (Figure 1D and Table 3). Three herbaceous species (R. buergeri, N. auriculata, and L. buergerianum) have always been in the SF, TF, and MB, and R. buergeri has always been the most dominant species in the herbaceous layers of the three communities. The finding may indicate that the invasion of Moso bamboo did not alter the near-ground light or soil conditions suiting the local shade-tolerant herbaceous species.
Regarding species diversity of the three layers, the arborous layer was the most influenced by the invasion in this study. Compared with the pre-invaded SF, MB had significantly lower diversity, as indicated by all three diversity indexes (Margalef, Shannon–Wiener, and Simpson index; P < 0.05; Figure 2). At the same time, the species evenness was smaller in MB than in SF (P < 0.05; Figure 2). This result is consistent with several previous studies (Bai et al., 2013; Yang et al., 2015; Ouyang et al., 2016) and partly supports our first hypothesis that invasion decreases plant diversity. However, the first hypothesis was not fully supported by the results regarding the shrub and herbaceous layers. Among the three diversity indexes, only one of them (Shannon–Wiener and Simpson indexes for the shrub and herbaceous layers, respectively) indicated a significantly increased diversity. In contrast, the other two indexes inversely meant that the diversities of these two layers were unchanged (Figure 2). The finding was consistent with the results observed in the invasion progress of Moso bamboo into Chinese fir (Xu et al., 2019). In contrast, compared with the invaded coniferous and broad-leaved mixed forests (Bai et al., 2013; Yang et al., 2015; Ouyang et al., 2016) and the secondary evergreen broad-leaved forest (Ouyang et al., 2016), MB had lower diversity in both shrub and herbaceous layers. In another study on the invaded coniferous and broad-leaved mixed forests (Bai et al., 2013), species diversity was reduced in the shrub layers but improved in the herbaceous layer.
Species diversity varied in different studies, which may be related to the short observation time and the different invasion stages because understory species diversity is a dynamic process. The increase of species diversity in shrub and herb layers during the invasion of Moso bamboo into SF may result from increased light intensity under the forest (Liu et al., 2011) because better light conditions may reduce light competition to improve the species diversity of understory vegetation (Hautier et al., 2009). However, it should be noted that different plants have different adaptability to light, and the influence of light intensity on species diversity may be restricted by the light requirement of plants (Boothroyd-Roberts et al., 2013). Much higher or much lower light intensity is not conducive to increasing the species diversity of understory plants (Tao and Feng, 1998). On the contrary, Moso bamboo forests have low stem density and soil-water content than other forest types (Shinohara and Otsuki, 2015), which may benefit understory diversity. In addition, Moso bamboo invasion increased soil bacterial diversity, soil pH, and soil organic carbon concentrations (Lin et al., 2014; Liu et al., 2021; Ouyang et al., 2022), which may also be beneficial to increasing the biodiversity under the forest. However, there is not enough direct evidence to support this argument. The mechanism of Moso bamboo invasion needs further study.
Effects of Moso bamboo invasion on aboveground biomass and its relationship with species diversity
Although the response relationship between the change of community species composition and forest ecosystem function is complex, many studies have shown that the loss of community species will lead to the decline of forest productivity and resistance and increase the instability of the system (Lavorel and Garnier, 2002; Barrufol et al., 2013; Bruelheide et al., 2014; Cavanaugh et al., 2014). In particular, the replacement of the dominant species in a community will affect the community environment on which other tree species depend, leading to the transformation of plant functional groups (Hooper et al., 2005), thus affecting the composition, structure, and function of the whole ecosystem (Lima et al., 2012). Contrary to our second hypothesis, the aboveground biomass in the entire stand and the arborous layer was not significantly reduced in TF and MB compared to the SF, which may be attributed to the increasing stem number of Moso bamboo (Table 2). This finding is consistent with the research results of Fukushima et al. (2015).
Previous studies have shown that Moso bamboo may obtain more nutrients and light resources by increasing the DBH and height to promote rapid growth and expansion during the invasion (Bai et al., 2012; Liu et al., 2016; Xu et al., 2019). However, in this study, the average DBH and height of Moso bamboo decreased during the invasion of Moso bamboo into SF (Table 1). This result may be due to the different invasion sites and forests (Liu et al., 2019). It may also be that the intraspecific competition of standing bamboo in the MB may be more intense, so it is necessary to obtain more light and nutrients by increasing height and DBH to get a competitive advantage. Its internal mechanism needs to be further studied.
The proportion of understory vegetation biomass in the total biomass of the forest ecosystem is small. Still, it plays a vital role in maintaining the succession and functional stability of the whole forest ecosystem (Sun et al., 2015). This study found that the aboveground biomass of the shrub and herbaceous layers and their proportion of the total aboveground biomass increased during the invasion of Moso bamboo to SF, but the increment in the herbaceous layer was not statistically significant (Table 4). Although the shrub and herb layers of the three stands contribute less to the biomass of the community, the understory often has more abundant species (Table 3). These species play an important role in the environment under plastic afforestation, affecting forest dynamics, soil carbon pool, and nutrient cycle (Nilsson and Wardle, 2005). Therefore, the change of species biomass in the understory should not be ignored when evaluating the ecological aftereffects of the Moso bamboo invasion.
The results showed no significant correlation between aboveground biomass and the diversity index of each layer and the whole stand during the Moso bamboo invasion (P > 0.05; Table 5), which indicated that the Moso bamboo invasion had no impact on the relationship. This result does not verify our third hypothesis that the positive correlation between species diversity and biomass is common across forest layers (Zhang et al., 2017). This insignificant relationship may be attributed to the intensity and direction of the correlation between biodiversity and ecosystem function changing under different environmental conditions (Cardinale et al., 2000). According to the stress gradient hypothesis, species are more competitive in the ideal environment but more mutual aid in an adverse environment. Therefore, it is more likely to find a positive correlation between species richness and aboveground biomass in adverse environmental conditions (Maestre et al., 2009). In this study, the arborous layer is rich in light resources. Therefore, Moso bamboo contributes a lot of biomass and excludes the survival of other species, which leads to no significant positive relationship between species richness and biomass.
Conclusion
Although the Moso bamboo invasion has drawn considerable attention to its impact, especially on biodiversity, few studies have reported the effects of the Moso bamboo invasion on forest biomass and the forest biodiversity–biomass relationship. By using the space-for-time method, we investigated the dynamics of species composition, diversity, and aboveground biomass of arborous, shrub, and herbaceous layers during Moso bamboo invasion into a SF. As the most significant part of the aboveground biomass (>99%), the arborous layer showed a decreasing trend in the aboveground biomass with the invasion progress. Still, the decrease was not statistically significant, which may be attributed to the increased five-fold stems. Moreover, the aboveground biomass in the arborous layer had no significant correlation with the significantly decreasing diversity. In the understory layers, shrubs had significantly increased aboveground biomass, while herbs did not. Similar to the arborous layer, both shrub and herbaceous layers did not significantly correlate their aboveground biomass and diversity indexes. However, these findings were concluded based on three plots per forest type and one transitional status, which may not cover enough variation in both biodiversity and aboveground biomass. Additionally, belowground biomass is also an essential part of the forest, which should be considered. We hope the findings of this study contribute to some fundamentals for future research and understanding of Moso bamboo invasion.
Data availability statement
The original contributions presented in this study are included in the article/supplementary material, further inquiries can be directed to the corresponding author/s.
Author contributions
XiC: methodology, software, investigation, visualization, writing – original draft and review, and editing. XinC and SH: data curation and investigation. DF: methodology, writing – review and editing, conceptualization, and investigation. All authors contributed to the article and approved the submitted version.
Funding
This research was supported by the Talent Project of Jiyang College of Zhejiang A&F University (Grant numbers 05251700035 and 05251700038).
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Akutsu, H., Aizawa, M., Matsue, K., and Ohkubo, T. (2012). Distribution and invasion of Phyllostachys pubescens stands into neighboring forests in Nasukarasuyama, Tochigi Prefecture. Bull. Utsunomiya Univ. For. 48, 139–152.
Ali, A., and Yan, E.-R. (2017). The forest strata-dependent relationship between biodiversity and aboveground biomass within a subtropical forest. For. Ecol. Manag. 401, 125–134. doi: 10.1016/j.foreco.2017.06.056
Bai, S., Wang, Y., Conant, R. T., Zhou, G., Xu, Y., Wang, N., et al. (2016). Can native clonal Moso bamboo encroach on adjacent natural forest without human intervention? Sci. Rep. 6:31504. doi: 10.1038/srep31504
Bai, S.-B., Zhou, G.-M., Wang, Y.-X., Liang, Q.-Q., Chen, J., Cheng, Y.-Y., et al. (2013). Plant species diversity and dynamics in forests invaded by Moso bamboo (Phyllostachys edulis) in Tianmu mountain nature reserve. Biodiv. Sci. 21, 288–295. doi: 10.3724/SP.J.1003.2013.08258
Bai, S.-B., Zhou, G.-M., Wang, Y.-X., Yu, S.-Q., Li, Y.-H., and Fang, F.-Y. (2012). Stand structure change of Phyllostachys pubescens forest expansion in tianmushan national nature reserve. J. West China For. Sci. 41, 77–82.
Bai, Y.-F., Wu, J.-G., Pan, Q.-M., Huang, J.-H., Wang, Q.-B., Li, F.-S., et al. (2007). Positive linear relationship between productivity and diversity: Evidence from the Eurasian Steppe. J. Appl. Ecol. 44, 1023–1034. doi: 10.1111/j.1365-2664.2007.01351.x
Barrufol, M., Schmid, B., Bruelheide, H., Chi, X., Hector, A., Ma, K., et al. (2013). Biodiversity promotes tree growth during succession in subtropical forest. PLoS One 8:e81246. doi: 10.1371/journal.pone.0081246
Boothroyd-Roberts, K., Gagnon, D., and Truax, B. (2013). Can hybrid poplar plantations accelerate the restoration of forest understory attributes on abandoned fields? For. Ecol. Manag. 287, 77–89. doi: 10.1016/j.foreco.2012.09.021
Bracken, M. E., Douglass, J. G., Perini, V., and Trussell, G. C. (2017). Spatial scale mediates the effects of biodiversity on marine primary producers. Ecology 98, 1434–1443. doi: 10.1002/ecy.1812
Bruelheide, H., Nadrowski, K., Assmann, T., Bauhus, J., Both, S., Buscot, F., et al. (2014). Designing forest biodiversity experiments: General considerations illustrated by a new large experiment in subtropical China. Methods Ecol. Evol. 5, 74–89. doi: 10.1111/2041-210X.12126
Cardinale, B. J., Nelson, K., and Palmer, M. A. (2000). Linking species diversity to the functioning of ecosystems: On the importance of environmental context. OIKOS 91, 175–183. doi: 10.1034/j.1600-0706.2000.910117.x
Cavanaugh, K. C., Gosnell, J. S., Davis, S. L., Ahumada, J., Boundja, P., Clark, D. B., et al. (2014). Carbon storage in tropical forests correlates with taxonomic diversity and functional dominance on a global scale. Glob. Ecol. Biogeogr. 23, 563–573. doi: 10.1111/geb.12143
Chen, Z.-Y., Yao, X.-M., and Tian, X.-M. (2009). The regression analysis between fertilization and the output, the economic benefit in Phyllostachys pubescens for culm-producing. Chin. Agric. Sci. Bull. 25, 166–169.
Davies, K. W. (2011). Plant community diversity and native plant abundance decline with increasing abundance of an exotic annual grass. Oecologia 167, 481–491. doi: 10.1007/s00442-011-1992-2
Fukushima, K., Usui, N., Ogawa, R., and Tokuchi, N. (2015). Impacts of Moso bamboo (Phyllostachys pubescens) invasion on dry matter and carbon and nitrogen stocks in a broad-leaved secondary forest located in Kyoto, western Japan. Plant Species Biol. 30, 81–95. doi: 10.1111/1442-1984.12066
Garrott, R. A., White, P., and White, C. A. V. (1993). Overabundance: An issue for conservation biologists? Conserv. Biol. 7, 946–949. doi: 10.1046/j.1523-1739.1993.740946.x
Guo, X.-C., Shi, P.-J., Niinemets, Ü., Hölscher, D., Wang, R., Liu, M.-D., et al. (2021). “Diminishing returns” for leaves of five age-groups of Phyllostachys edulis culms. Am. J. Bot. 108, 1662–1672. doi: 10.1002/ajb2.1738
Hautier, Y., Niklaus, P. A., and Hector, A. (2009). Competition for light causes plant biodiversity loss after eutrophication. Science 324, 636–638. doi: 10.1126/science.1169640
Hooper, D. U., Chapin Iii, F. S., Ewel, J. J., Hector, A., Inchausti, P., Lavorel, S., et al. (2005). Effects of biodiversity on ecosystem functioning: A consensus of current knowledge. Ecol. Monogr. 75, 3–35. doi: 10.1890/04-0922
Huang, W.-W., Reddy, G. V., Li, Y.-Y., Larsen, J. B., and Shi, P.-J. (2020). Increase in absolute leaf water content tends to keep pace with that of leaf dry mass—evidence from bamboo plants. Symmetry 12, 1345. doi: 10.3390/sym12081345
Huang, W.-W., Su, X.-F., Ratkowsky, D. A., Niklas, K. J., Gielis, J., and Shi, P.-J. (2019). The scaling relationships of leaf biomass vs. leaf surface area of 12 bamboo species. Glob. Ecol. Conserv. 20:e00793. doi: 10.1016/j.gecco.2019.e00793
Jiang, L.-H., Mao, C.-M., Wu, H.-Z., Duan, Q.-X., and Fang, Z.-X. (2015). Investigation and analysis of Moso forest management in songyang county, Zhejiang province. World Bamboo Rattan 13, 28–34.
Lasky, J. R., Uriarte, M., Boukili, V. K., Erickson, D. L., John Kress, W., and Chazdon, R. L. (2014). The relationship between tree biodiversity and biomass dynamics changes with tropical forest succession. Ecol. Lett. 17, 1158–1167. doi: 10.1111/ele.12322
Lavorel, S., and Garnier, É. (2002). Predicting changes in community composition and ecosystem functioning from plant traits: Revisiting the Holy Grail. Funct. Ecol. 16, 545–556. doi: 10.1046/j.1365-2435.2002.00664.x
Lima, R. A., Rother, D. C., Muler, A. E., Lepsch, I. F., and Rodrigues, R. R. (2012). Bamboo overabundance alters forest structure and dynamics in the Atlantic Forest hotspot. Biol. Conserv. 147, 32–39. doi: 10.1016/j.biocon.2012.01.015
Lin, M.-Y., Hsieh, I., Lin, P.-H., Laplace, S., Ohashi, M., Chen, T.-H., et al. (2017). Moso bamboo (Phyllostachys pubescens) forests as a significant carbon sink? A case study based on 4-year measurements in central Taiwan. Ecol. Res. 32, 845–857. doi: 10.1007/s11284-017-1497-5
Lin, Y.-T., Tang, S.-L., Pai, C.-W., Whitman, W. B., Coleman, D. C., and Chiu, C.-Y. (2014). Changes in the soil bacterial communities in a cedar plantation invaded by Moso bamboo. Microb. Ecol. 67, 421–429. doi: 10.1007/s00248-013-0291-3
Liu, C.-X., Zhou, Y., Qin, H., Liang, C.-F., Shao, S., Fuhrmann, J. J., et al. (2021). Moso bamboo invasion has contrasting effects on soil bacterial and fungal abundances, co-occurrence networks and their associations with enzyme activities in three broadleaved forests across subtropical China. For. Ecol. Manag. 498:119549. doi: 10.1016/j.foreco.2021.119549
Liu, S., Zhou, G.-M., and Bai, S.-B. (2011). Light intensity changes on Cunninghamia lanceolata in mixed stands with different concentrations of Phyllostachys pubescens. J. Zhejiang A&F Univ. 28, 550–554.
Liu, X., Siemann, E., Cui, C., Liu, Y., Guo, X., and Zhang, L. (2019). Moso bamboo (Phyllostachys edulis) invasion effects on litter, soil and microbial PLFA characteristics depend on sites and invaded forests. Plant Soil 438, 85–99. doi: 10.1007/s11104-019-04010-3
Liu, X.-Z., Fan, S.-H., Liu, G.-L., and Peng, C. (2016). Changing characteristics of main structural indexes of community during the expansion of Moso bamboo forests. Chin. J. Ecol. 35, 3165–3171.
Maestre, F. T., Callaway, R. M., Valladares, F., and Lortie, C. J. (2009). Refining the stress-gradient hypothesis for competition and facilitation in plant communities. J. Ecol. 97, 199–205. doi: 10.1111/j.1365-2745.2008.01476.x
Martin, P. H., Canham, C. D., and Marks, P. L. (2009). Why forests appear resistant to exotic plant invasions: Intentional introductions, stand dynamics, and the role of shade tolerance. Front. Ecol. Environ. 7:142–149. doi: 10.1890/070096
Mori, A. S., Osono, T., Cornelissen, J. H. C., Craine, J., and Uchida, M. (2017). Biodiversity–ecosystem function relationships change through primary succession. OIKOS 126, 1637–1649. doi: 10.1111/oik.04345
Nackley, L. L., West, A. G., Skowno, A. L., and Bond, W. J. (2017). The nebulous ecology of native invasions. Trends Ecol. Evol. 32, 814–824. doi: 10.1016/j.tree.2017.08.003
Nakai, A., and Kisanuki, H. (2006). Effect of the bamboo culm density on the tree species diversity and density of understory at a forest stand on a major bed along the Miya river downstream [Japan]. Bull. Faculty Bioresour. Mie Univ. (apan) 21–28.
Nilsson, M.-C., and Wardle, D. A. (2005). Understory vegetation as a forest ecosystem driver: Evidence from the northern Swedish boreal forest. Front. Ecol. Environ. 3:421–428. doi: 10.1890/1540-9295(2005)003[0421:UVAAFE]2.0.CO;2
Okutomi, K., Shinoda, S., and Fukuda, H. (1996). Causal analysis of the invasion of broad-leaved forest by bamboo in Japan. J. Veg. Sci. 7, 723–728. doi: 10.2307/3236383
Ouyang, M., Tian, D., Pan, J.-M., Chen, G.-P., Su, H.-J., Yan, Z.-B., et al. (2022). Moso bamboo (Phyllostachys edulis) invasion increases forest soil pH in subtropical China. CATENA 215:106339. doi: 10.1016/j.catena.2022.106339
Ouyang, M., Yang, Q.-P., Chen, X., Yang, G.-Y., Shi, J.-M., and Fang, X.-M. (2016). Effects of the expansion of Phyllostachys edulis on species composition, structure and diversity of the secondary evergreen broad-leaved forests. Biodiv. Sci. 24, 649–657. doi: 10.17520/biods.2015290
Pan, Y.-D., Birdsey, R. A., Fang, J.-Y., Houghton, R., Kauppi, P. E., Kurz, W. A., et al. (2011). A large and persistent carbon sink in the world’s forests. Science 333, 988–993. doi: 10.1126/science.1201609
Pickett, S. T. (1989). “Space-for-time substitution as an alternative to long-term studies,” in Long-term studies in ecology, ed. G. E. Likens (New York, NY: Springer), 110–135. doi: 10.1007/978-1-4615-7358-6_5
R Core Team (2022). R: A language and environment for statistical computing. Vienna: R Foundation for Statistical Computing.
Ramula, S., and Pihlaja, K. (2012). Plant communities and the reproductive success of native plants after the invasion of an ornamental herb. Biol. Invasions 14, 2079–2090. doi: 10.1007/s10530-012-0215-z
Ratcliffe, S., Wirth, C., Jucker, T., Van Der Plas, F., Scherer-Lorenzen, M., Verheyen, K., et al. (2017). Biodiversity and ecosystem functioning relations in European forests depend on environmental context. Ecol. Lett. 20, 1414–1426. doi: 10.1111/ele.12849
Shen, Q.-Y., and Tang, M.-P. (2019). Stem biomass models of Phyllostachysedulis in Zhejiang province. Sci. Silvae Sin. 55, 181–188.
Shi, P.-J., Gao, J., Song, Z.-P., Liu, Y.-H., and Hui, C. (2018). Spatial segregation facilitates the coexistence of tree species in temperate forests. Forests 9:768. doi: 10.3390/f9120768
Shi, P.-J., Preisler, H. K., Quinn, B. K., Zhao, J., Huang, W.-W., Röll, A., et al. (2020). Precipitation is the most crucial factor determining the distribution of Moso bamboo in Mainland China. Glob. Ecol. Conserv. 22:e00924. doi: 10.1016/j.gecco.2020.e00924
Shinohara, Y., and Otsuki, K. (2015). Comparisons of soil-water content between a Moso bamboo (Phyllostachys pubescens) forest and an evergreen broadleaved forest in western Japan. Plant Species Biol. 30, 96–103. doi: 10.1111/1442-1984.12076
Silva Pedro, M., Rammer, W., and Seidl, R. (2017). Disentangling the effects of compositional and structural diversity on forest productivity. J. Veg. Sci. 28, 649–658. doi: 10.1111/jvs.12505
Song, Q.-N., Lu, H., Liu, J., Yang, J., Yang, G.-Y., and Yang, Q.-P. (2017). Accessing the impacts of bamboo expansion on NPP and N cycling in evergreen broadleaved forest in subtropical China. Sci. Rep. 7:40383. doi: 10.1038/srep40383
Sun, Y.-J., Ma, W., and Liu, Y.-H. (2015). Biomass of Larix olgensis plantations based on species diversity analyses in Heilongjiang, China. Acta Ecol. Sin. 35, 3329–3338. doi: 10.5846/stxb201306201752
Tao, F.-L., and Feng, Z.-W. (1998). Studies on plant species diversity under the canopy of Larix kaempferi community in west mountain area of Henan Province. Chin. J. Ecol. 17, 1–6.
Tian, X.-K., Wang, M.-Y., Meng, P., Zhang, J.-S., Zhou, B.-Z., Ge, X.-G., et al. (2020). Native bamboo invasions into subtropical forests alter microbial communities in litter and soil. Forests 11:314. doi: 10.3390/f11030314
Tilman, D., Knops, J., Wedin, D., Reich, P., Ritchie, M., and Siemann, E. (1997). The influence of functional diversity and composition on ecosystem processes. Science 277, 1300–1302. doi: 10.1126/science.277.5330.1300
van der Sande, M. T., Peña-Claros, M., Ascarrunz, N., Arets, E. J., Licona, J. C., Toledo, M., et al. (2017). Abiotic and biotic drivers of biomass change in a Neotropical forest. J. Ecol. 105, 1223–1234. doi: 10.1111/1365-2745.12756
van Kleunen, M., Weber, E., and Fischer, M. (2010). A meta-analysis of trait differences between invasive and non-invasive plant species. Ecol. Lett. 13, 235–245. doi: 10.1111/j.1461-0248.2009.01418.x
Vilà, M., Espinar, J. L., Hejda, M., Hulme, P. E., Jarošík, V., Maron, J. L., et al. (2011). Ecological impacts of invasive alien plants: A meta-analysis of their effects on species, communities and ecosystems. Ecol. Lett. 14, 702–708. doi: 10.1111/j.1461-0248.2011.01628.x
Wang, B., Liu, M., and Zhang, B. (2009). Dynamics of net production of chinese forest vegetation based on forest inventory data. For. Resour. Manag. 1, 35–43.
Wang, Y. J., Müller-Schärer, H., Van Kleunen, M., Cai, A. M., Zhang, P., Yan, R., et al. (2017). Invasive alien plants benefit more from clonal integration in heterogeneous environments than natives. New Phytol. 216, 1072–1078. doi: 10.1111/nph.14820
Wang, Y.-J., Chen, D., Yan, R., Yu, F.-H., and Van Kleunen, M. (2019). Invasive alien clonal plants are competitively superior over co-occurring native clonal plants. Perspect. Plant Ecol. Evol. Syst. 40:125484. doi: 10.1016/j.ppees.2019.125484
Wang, Y.-X., Bai, S.-B., Binkley, D., Zhou, G.-M., and Fang, F.-Y. (2016). The independence of clonal shoot’s growth from light availability supports Moso bamboo invasion of closed-canopy forest. For. Ecol. Manag. 368, 105–110. doi: 10.1016/j.foreco.2016.02.037
Warren, R. J., King, J. R., Tarsa, C., Haas, B., and Henderson, J. (2017). A systematic review of context bias in invasion biology. PLoS One 12:e0182502. doi: 10.1371/journal.pone.0182502
Wu, X., Wang, X.-P., Tang, Z.-Y., Shen, Z.-H., Zheng, C.-Y., Xia, X.-L., et al. (2015). The relationship between species richness and biomass changes from boreal to subtropical forests in China. Ecography 38, 602–613.
Xu, D.-W., Liu, J.-F., He, Z.-S., and Zheng, S.-Q. (2019). Community species diversity after Phyllostachys edulis expansion to Cunninghamia lanceolata forest. J. For. Environ. 39, 37–41.
Xue, W., Shen, J. X., Guo, Z. W., Lei, J. P., Li, J. M., Yu, F. H., et al. (2021). Shoot removal interacts with soil temperature to affect survival, growth and physiology of young ramets of a bamboo. For. Ecol. Manage. 481:118735.
Yang, Q.-P., Wang, B., Guo, Q.-R., Zhao, G.-D., Fang, K., and Liu, Y.-Q. (2011). Effects of Phyllostachys edulis expansion on carbon storage of evergreen broad-leaved forest in Dagangshan Mountain, Jiangxi. Acta Agric. Univ. Jiangxiensis 33, 0529–0536.
Yang, Q.-P., Yang, G.-Y., Song, Q.-N., Shi, J.-M., Ouyang, M., Qi, H.-Y., et al. (2015). Ecological studies on bamboo expansion: Process, consequence and mechanism. Chin. J. Plant Ecol. 39, 110–124. doi: 10.17521/cjpe.2015.0012
Zeng, W. (2018). Validation of two-variable tree volume tables for main tree species in China. For. Resour. Manage. 5, 37–43.
Zhang, H.-N., and Xue, J.-H. (2018). Spatial pattern and competitive relationships of Moso bamboo in a native subtropical rainforest community. Forests 9:774. doi: 10.3390/f9120774
Keywords: biomass, Moso bamboo invasion, Phyllostachys pubescens, secondary coniferous and broad-leaved mixed forest, species diversity
Citation: Chen X, Chen X, Huang S and Fang D (2022) Impacts of Moso bamboo (Phyllostachys pubescens) invasion on species diversity and aboveground biomass of secondary coniferous and broad-leaved mixed forest. Front. Plant Sci. 13:1001785. doi: 10.3389/fpls.2022.1001785
Received: 24 July 2022; Accepted: 31 August 2022;
Published: 30 September 2022.
Edited by:
Guolei Li, Beijing Forestry University, ChinaReviewed by:
Peijian Shi, Nanjing Forestry University, ChinaDaxing Gu, Guangxi Institute of Botany (CAS), China
Keitaro Fukushima, Fukushima University, Japan
Fei-Hai Yu, Taizhou University, China
Hiroyuki Shima, University of Yamanashi, Japan
Copyright © 2022 Chen, Chen, Huang and Fang. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Dongming Fang, ZG1mYW5nQHphZnUuZWR1LmNu