
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
REVIEW article
Front. Plant Sci., 17 December 2021
Sec. Plant Symbiotic Interactions
Volume 12 - 2021 | https://doi.org/10.3389/fpls.2021.791033
 Beena Alam1†Jùnwén Lǐ1†Qún Gě1†
Beena Alam1†Jùnwén Lǐ1†Qún Gě1† Mueen Alam Khan2‡
Mueen Alam Khan2‡ Jǔwǔ Gōng1‡Shahid Mehmood3
Jǔwǔ Gōng1‡Shahid Mehmood3 Yǒulù Yuán1*‡
Yǒulù Yuán1*‡ Wànkuí Gǒng1*‡
Wànkuí Gǒng1*‡
Endophytic fungi (EF) are a group of fascinating host-associated fungal communities that colonize the intercellular or intracellular spaces of host tissues, providing beneficial effects to their hosts while gaining advantages. In recent decades, accumulated research on endophytic fungi has revealed their biodiversity, wide-ranging ecological distribution, and multidimensional interactions with host plants and other microbiomes in the symbiotic continuum. In this review, we highlight the role of secondary metabolites (SMs) as effectors in these multidimensional interactions, and the biosynthesis of SMs in symbiosis via complex gene expression regulation mechanisms in the symbiotic continuum and via the mimicry or alteration of phytochemical production in host plants. Alternative biological applications of SMs in modern medicine, agriculture, and industry and their major classes are also discussed. This review recapitulates an introduction to the research background, progress, and prospects of endophytic biology, and discusses problems and substantive challenges that need further study.
Plants, especially perennials, are colonized by many types of endophytic microorganisms (Stone et al., 2004; Demain, 2014), which live inside plant tissues either throughout their lives or during a certain period of their life cycles without causing visible damage or morphological changes in their hosts. These microorganisms include both fungi and bacteria (Zhang et al., 2006; Porras-Alfaro and Bayman, 2011; Gouda et al., 2016), and usually coexist with pathogens. According to their colonizing behaviors, endophytic microflora can be sorted into facultative and obligate categories. Facultative endophytes colonize plants at certain stages of their life cycles, but they may also reside outside the plant at other stages to form an association with the immediate rhizosphere soil of host plants (Abreu-Tarazi et al., 2010). In contrast, obligate strains live in plants throughout their entire life cycles. They usually proliferate across plant generations through vertical transmission and use or alter the metabolic machinery and products of plants for their own survival (Hardoim et al., 2008; Gouda et al., 2016).
Among these endophytic microorganisms, endophytic fungi (EFs) have attracted much research interest because they have provided not only novel sources of cytotoxic compounds, such as anticarcinogenic molecules (Uzma et al., 2018) and antibacterial substances (Radic and Strukelj, 2012), but also biostimulants for essential oil biosynthesis (El Enshasy et al., 2019). They may enhance nutrient solubilization in the plant rhizosphere (Mehta et al., 2019), promote plant growth (Poveda et al., 2021), act as biological control agents (Poveda and Baptista, 2021), or activate plant systemic resistances to biotic (Poveda et al., 2020a) or abiotic (Cui et al., 2021) stresses. In this review, we provide a comprehensive overview of the biological aspects of EFs that are related to their diversity, their distribution, and their multidimensional interactions with multiple players, including host plants, epiphytes, and pathogens, in their communities. We also examine the role of secondary metabolites (SMs) in these multidimensional interactions and how SMs are biosynthesized through gene expression regulation and through the mimicry or alteration of phytochemical production in host plants. Finally, we identify the principal categories of SMs, characterize their attributes, and review their alternative biological potential. This review provides readers with a profound understanding of EFs and SMs.
The geographical distribution of biota is characterized as a continuous gradient distribution of traits. Biodiversities at various levels, including species, function, and phylogenesis, are the basis of this continuous distribution (Violle et al., 2014). Although little is known about the mechanisms of biodiversity formation in a particular geographical habitat, fungal strains mediate many processes and may play a crucial role in their habitats (Vandenkoornhuyse et al., 2002). Plant tissue is arranged in multi-layers, forming a spatial and temporal supportive refuge, like a natural habitat, for various endophytic microorganisms. Based on an accepted estimation of a 1:4 or 1:5 ratio of vascular plants to fungal strains, there could be more than one million strains of EFs remaining to be discovered (Sun and Guo, 2012). However, our limited recognition of EF diversity renders the ratio a biased estimation, because EFs thrive ubiquitously in species diversity, while rare species and those that are characterized as non-sporulating, non-culturable, or asceptic cannot be examined properly in current laboratory isolation and fermentation attempts (Stone et al., 2004; Zhang et al., 2006; Ding et al., 2017; Alvear-Daza et al., 2021).
According to the reproductive pattern and host occurrence, EF communities can be sorted into two categories: the Clavicipitaceous/Balansiaceous group (C-group) and the non-Clavicipitaceous/non-Balansiaceous group (NC-group). C-group EFs infect the ovules of host plants and transmit vertically from parents to progenies through host seeds. The target tissues for their colonization are living rhizomes and shoots of host plants, but the host range is restricted to grass species (Poaceae). C-group species are typical obligate endophytes (Carroll, 1988), which protect their hosts from herbivore attacks or enable the hosts to survive under drought conditions by secreting defensive or supportive bioactive metabolites, respectively (Roberts and Lindow, 2014; Poveda, 2021). NC-group EFs, which are non-grass-host related (Ascomycota, Basidiomycota), have a wide biodiversity and distribution from tropical to polar areas, with their hosts including nonvascular, vascular, and woody plant communities. They transmit sexually or asexually by producing spores or conidia, which contribute to horizontal propagation, i.e., the induction of symbiosis. These EFs are not closely associated the host plants because they can exist in a quiescent state until they sense the chemical changes from host plants suffering injuries, wounds, or other environmental stresses (Carroll, 1988; Rodriguez et al., 2008; Mishra et al., 2021). The colonization of the NC-group in aerial organs is usually local, restricted, limited, and mainly intercellular, but the colonization of this group in roots or the rhizosphere is extensive, organized, systematic, intercellular, and intracellular. Some illustrative endophytic mycobiomes of the NC-group include Fusarium spp., Piriformospora indica, and dark septate mycobiota (Varma et al., 2000; Schulz et al., 2002).
Recent progress in molecular techniques, such as metagenomic sequencing, DNA fingerprinting, and phylogenetic analysis, has been successfully employed to detect and identify the species, community composition, and diversity of EFs. These technologies provide more precise methods of fungal identification and accommodation of the asceptic strains in situ than conventional isolation attempts (Arnold and Lutzoni, 2007; Tao et al., 2008; Tejesvi et al., 2009; Bahram et al., 2018; Jiang et al., 2018; Huang et al., 2019; Zhang J. et al., 2019). Studies have demonstrated that the host range of EFs includes algae (Tan and Zou, 2001), liverworts, mosses, hornworts (Davey and Currah, 2006; Aly et al., 2010; Sun and Guo, 2012), grasses (Muller and Krauss, 2005; Su et al., 2010), lycophytes, ferns, equisetopsida (Aly et al., 2010), shrubs, deciduous and coniferous trees (Gao et al., 2008; Albrectsen et al., 2010; Mohamed et al., 2010; Sun et al., 2011), gymnosperms, angiosperms, and annual/perennial herbaceous and broad-leaved plants. EFs are also distributed in a broad range of geographic habitats, such as tropical, temperate, arctic tundra, alpine, aquatic, and xerophytic ecosystems (Tan and Zou, 2001; Stone et al., 2004; Zhang et al., 2006) for more than 400 million years (Sun and Guo, 2012).
During their biogenesis and the establishment of symbiosis, EFs encounter specific host groups (Stone et al., 2004; Sun et al., 2008; Chadha et al., 2014), non-host plant communities, epiphytes, and pathogens. These multiple encounters prompt EFs to develop multidimensional interactions with the organisms they encounter (Figure 1). Major evolutionary and ecological novelties may direct or correlate to these interactions.
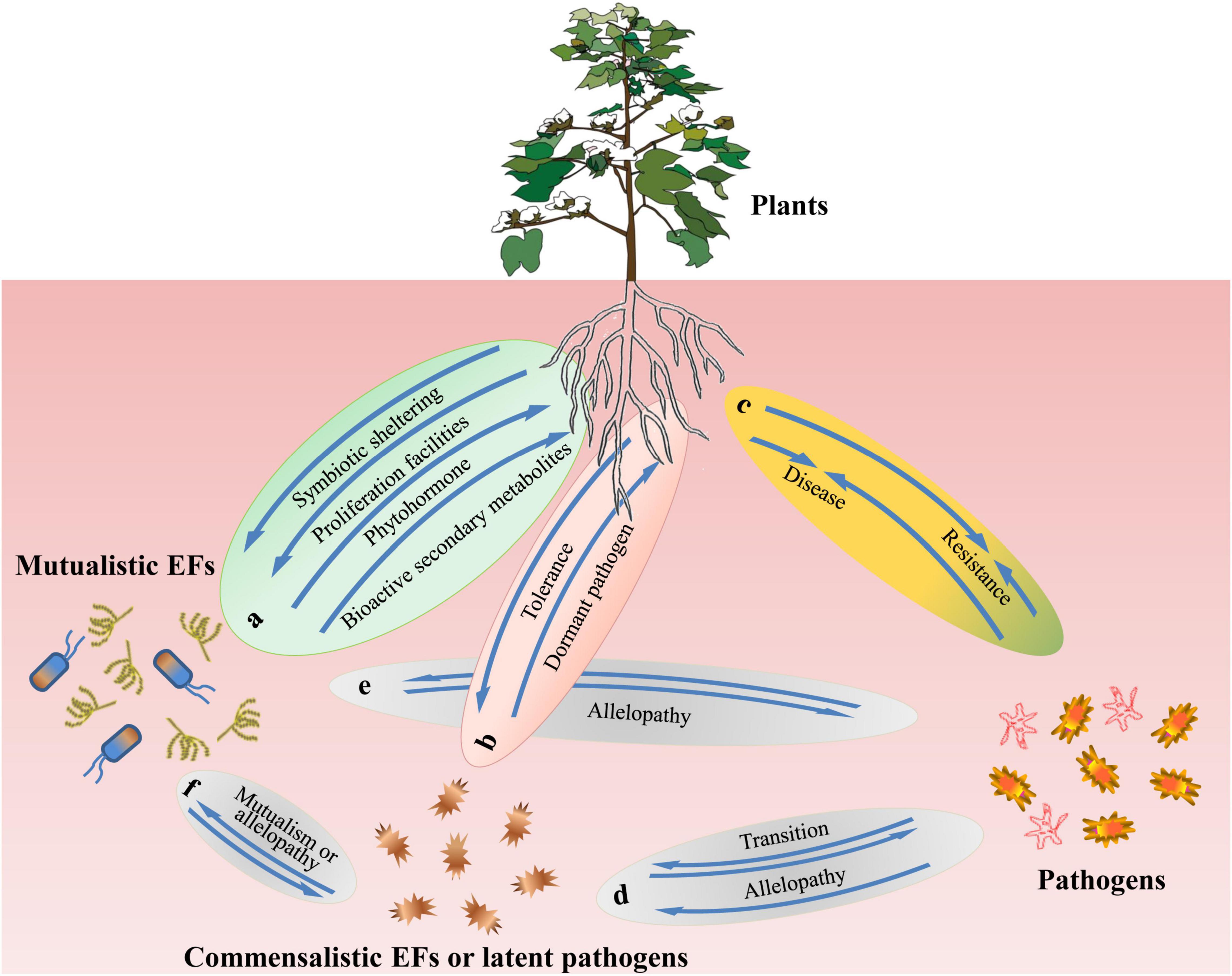
Figure 1. A schematic model of plant and microbiome interactions. The interactions include: (a) interactions between plants and mutualistic endophytic fungi (EFs), (b) interactions between plants and commensalistic EFs or latent pathogens, (c) interactions between Plants and pathogens, and (d–f) interactions between microbiomes.  Mutualistic endophytic fungus (EF),
Mutualistic endophytic fungus (EF),  commensalistic
commensalistic  EF,
EF,  pathogens.
pathogens.
Fungi can colonize the intercellular or intracellular spaces of plants, but systematic and extensive colonization is most likely to occur in the roots rather than in the aerial leaves or stems. Colonization in aerial organs primarily depends on the host’s apoplastic fluid as the nutrient source to support the normal reproduction of EFs in aerial organs (Schulz et al., 2002). During infection, fungi form three corresponding types of interactions with hosts: mutualistic (beneficial endophytes), commensalistic (non-beneficial/virulent endophytes), and pathogenic (virulent pathogens) (Figure 1), depending on the physiological status or specific circumstances that host plants experience. According to these three modes of action, fungal strains can increase, have no palpable effects on, or decrease host fitness (Kogel et al., 2006).
In mutualistic symbiosis, both partners of EFs and host plants enjoy this beneficial symbiotic continuum (Jia et al., 2016) and eventually achieve evolutionary and ecological success. EFs alter the metabolic mechanism of host plants, improve metal and drought tolerance, enhance growth, and promote nutrient acquisition (Hunt and Newman, 2005; Rodriguez and Redman, 2008; Mejia et al., 2014; Poveda et al., 2021). They also enhance the defense efficiency of host plants against herbivorous animals and pests as well as against infections of pathogenic microorganisms (Cui et al., 2021). In response to these benefits, host plants provide symbiotic shelters and other proliferation facilities, such as an adequate nutrient supply and protection, to enable fungi to safely complete their life cycles during colonization (Figure 1a). The exact mechanism of mutual interaction between EFs and plants remains to be elucidated (Haas and Défago, 2005), but EFs offer these potential advantages to host plants in several possible ways. One of these ways is to enhance the plant’s immune system by producing a plethora of bioactive SMs as protective agents. It is speculated that increasing the number of SMs causes physiological changes in the infected host plant that further stimulate the plant immune system (Poveda et al., 2020a). Various experiments on endophytic and pathogenic fungi and monitoring their comparative effects on host plants suggest that EFs promote the defense mechanisms of host plants by synthesizing bioactive SMs and herbicidal metabolites in hosts (Figueiredo et al., 2008). In an in vitro tripartite interaction assay, it was observed that Fusarium oxysporum rapidly killed A. thaliana plants, whereas the presence of Paraconiothyrium variabile reduced plant death by up to 85% (Bärenstrauch et al., 2020). This hypothesis was confirmed in the following experiments. When mosquito larva were assayed with ethanol extracts from Poa ampla Merr. (big bluegrass), the results showed that only the extracts from the plants that were inoculated with Neotyphodium typhnium were effective against the insect, whereas the extracts from the plants that were not inoculated with the fungus were inactive (Ju et al., 1998). Another way that an EF bestows host plant benefits is that EFs promote plant growth by producing and providing phytohormones, including auxins, cytokines, and gibberellins. This has been confirmed by the discovery of a common gibberellin biosynthesis pathway in fungi and higher plants (Tan and Zou, 2001; Kumar et al., 2013). Studies have demonstrated that endophytes can improve the growth and proliferation of plants by enhancing their defensive systems, like ethylene and jasmonic acid do (Kunkel and Brooks, 2002; Van der Ent et al., 2009; Di et al., 2016; Verbon and Liberman, 2016; Forni et al., 2017; Yang et al., 2019), or by interacting with ethylene-targeted transcription factors (Camehl et al., 2010). Neotyphodium, an EF, colonizes in tall fescue ryegrass and confers protection and stability on host plants in hostile conditions; in return, ryegrass provides facilities to promote fungal proliferation through ryegrass seeds infected with fungal hyphae (Tan and Zou, 2001). The resulting competitive advantage provides both hosts and symbionts with greater potential for growth and survival than the non-symbiotic plants and fungi.
In a commensalistic or latent pathogenic relationship, EFs sporulate rapidly and interact with host plants through a relationship with or without any significant beneficial effects on plant physiology (Stone et al., 2004; Hiruma et al., 2016). Studies have reported that these endophytes exist as latent pathogens in hosts under normal conditions (Brown et al., 1998; Photita et al., 2004; Rodriguez and Redman, 2008; Gorzynska et al., 2018; Figure 1b), while some studies have described various species and genera of EFs from host plants as active pathogens under unusual physiological stresses (Photita et al., 2004; Figure 1c). Fungi that have been identified as endophytes that are also possible pathogens include Cladosporium, Fusarium, Colletotrichum, Cordana, Deightoniella, Periconiella, Verticillium, Curvularia, Nigrospora, Guignardia, and Phoma (Photita et al., 2004; Cui et al., 2021). These EFs stay in latent or dormant state in the tissue of their host plants long before the outbreak of disease symptoms. In such cases, the dormancy phase is essential because it determines the time when the fungus is harmless as an endophyte and when it is virulent as a pathogen. In the virulent phase, EFs cause obvious symptoms and change the morphology and physiology of host plants under adverse conditions (Figure 1d). It is precisely these hostile conditions, including malnutrition, disruption of ontogenetic state (Sieber, 2007; Rodriguez and Redman, 2008), biotic stresses, drastic climate changes (such as elevated temperature and excessive humidity), and senescence, that break the balance between EFs and their hosts and lead to the transition of EFs from latent mode to active virulent pathogens, although there are no obvious disease symptoms before transition (Romero et al., 2001; Photita et al., 2004; Poveda et al., 2020b). There are also endemic fungal species, which typically include the majority of F. oxysporum strains, that live in host tissues without causing disease symptoms. Some strains even confer beneficial effects (Imazaki and Kadota, 2015; Di et al., 2016), such as C. tofieldiae, which promotes plant growth and fertility as an endophyte under phosphorus-deficient conditions (Hiruma et al., 2016). It is assumed that a combination of effectors, enzymes, and secondary metabolites determines the outcome of an interaction; that is, whether it is endophytic or pathogenic (Di et al., 2016; Poveda et al., 2020b). Nutrient status may have facilitated the transition of C. tofieldiae from pathogenicity to symbiosis (Hiruma et al., 2016).
As one of the numerous microbial players in the endophyte–host continuum, EFs inevitably have dynamic and complex interactions with other plant microbial communities, including endosphere-associated fungal and bacterial strains, regardless of whether they are pathogenic or symbiotic under natural conditions (Strobel, 2018; Figures 1d–f). Studies have observed that fungal endophytic metabolomic profiles can be affected by pathogen infection (Combes et al., 2012), indicating that antagonistic effects, or chemical communications, exist between the two microorganisms (Combes et al., 2012; Saikkonen et al., 2013). Endophytic fungi may also harbor a variety of bacterial species (endohyphal bacteria) in their hyphae (Hoffman and Arnold, 2010). When a foliar endophyte hosts the endohyphal bacterium Luteibacter sp., its indole acetic acid (IAA) production is significantly enhanced. However, the axenic culture of the bacterium does not show IAA production (Hoffman et al., 2013). Host plants may provide direct interfaces facilitating interactions between EFs and bacterial microflora. It is observed that two mutualistic EFs, Neotyphodium sp. and Epichloë sp., protect the leaves of fescue grasses from herbivores by producing loline alkaloids. On the leaf surface of fescue grass that is not infected with such endophytes or on the leaf surface of other plants that are infected with endophytes incapable of producing loline alkaloids, there is no accumulation of loline-consuming bacteria (Roberts and Lindow, 2014). In some cases, indirect interactions between EFs and other microorganisms may involve the participation of a third organism. A typical example is the interaction between the EF N. coenophialum and grass yellow dwarf virus (Hunt and Newman, 2005). N. coenophialum protects its host plants from aphids, and aphids are primary vectors of viruses (Grafton et al., 1982; Malmstrom et al., 2005; Yi and Gray, 2020). The N. coenophialum strains that provide tall fescue with better prevention against bird-cherry-oat aphids also inhibit the spread of the virus (Hunt and Newman, 2005).
When different microorganisms occupy the same habitat, they must compete for the resources of that habitat for nutrition, living space, reproduction, and other needs throughout their life cycles. Compared to microorganisms that have poor adaptability, adaptable microorganisms are more likely to obtain adequate resources and increase their abundance when the resources are insufficient to meet the needs of the community. As a result, the former may not be able to survive in adverse conditions. In order to survive, organisms have developed two effective strategies to compete. One is to produce allelochemicals that inhibit the growth of their competitors and eliminate toxic effects produced by their competitors in the vicinity (Macías-Rubalcava et al., 2008; Konarzewska et al., 2020; Poveda, 2021). The other is to produce allelochemicals that help their producers form alliances through symbiotic relationships with symbionts or hosts. These symbiotic relationships enable both parties to survive and reproduce safely, even in extremely adverse environments (Macías-Rubalcava et al., 2008). According to the hypothesis of long-term coevolution within biological communities (Ji et al., 2009), this mutual orientation of EFs and their hosts leads to each EF having developed a specific range of host species, enabling them to accumulate in a specific eukaryotic host group (Figure 2a).
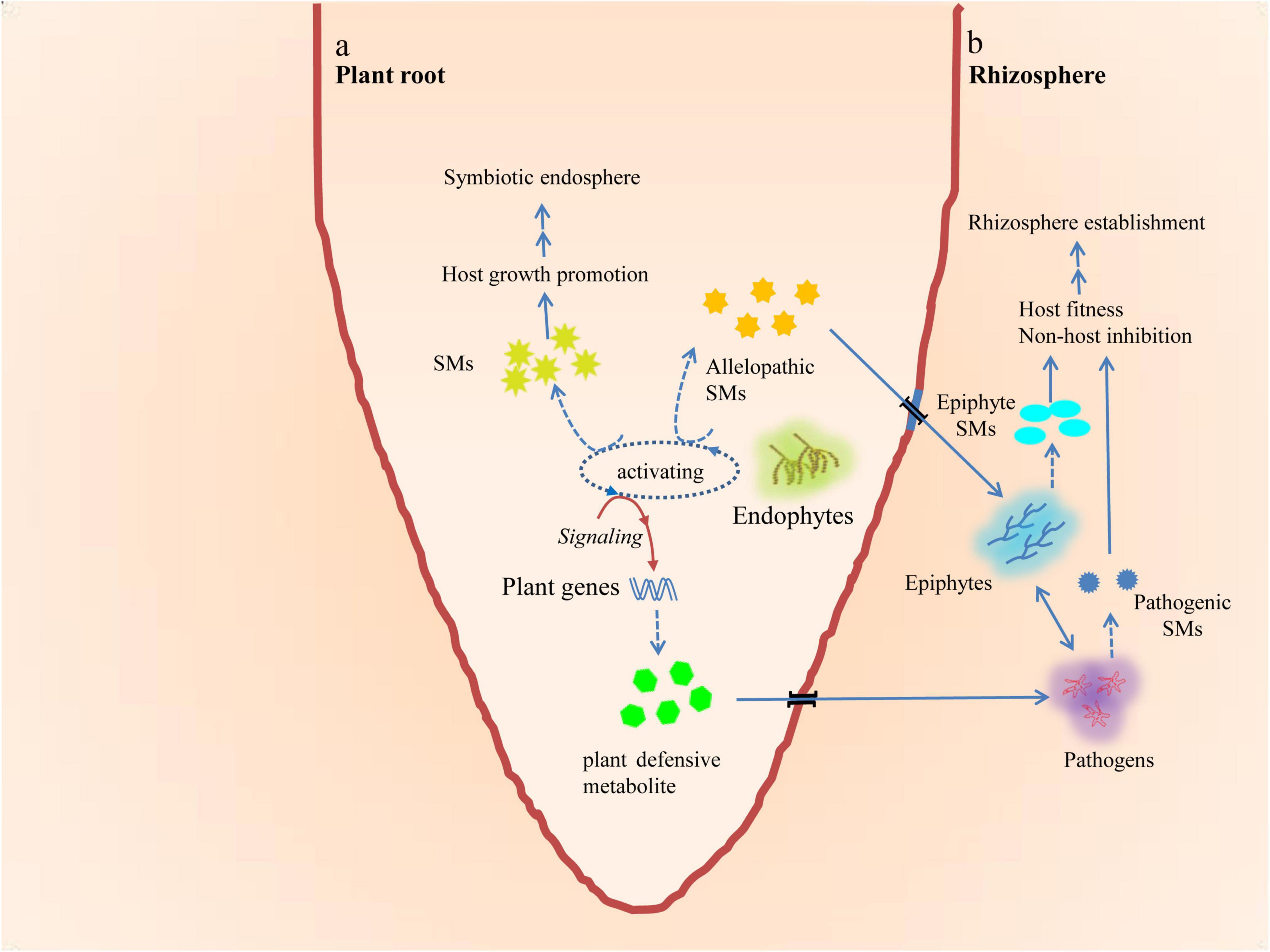
Figure 2. Mutual orientation of microbiome and plant (a) and multidirectional crosstalks or chemical communications among plants, endophytes, epiphytes, and pathogens under natural ecological conditions (b).  Endophytic fungus (EF),
Endophytic fungus (EF),  EF secondary metabolite (SM),
EF secondary metabolite (SM),  allelopathic SMs,
allelopathic SMs,  pathogens, pathogenic SM,
pathogens, pathogenic SM,  epiphytes,
epiphytes,  epiphytic SM,
epiphytic SM,  plant defensive metabolite.
plant defensive metabolite.
Most of these allelochemicals are SMs, a variety of usually low-molecular-weight and amazingly heterogeneous chemicals that were previously thought to have no direct functional effect on the growth, development, and reproduction of the organisms that produce them (Keller et al., 2005; Yu and Keller, 2005; Fox and Howlett, 2008; Shwab and Keller, 2008). Volatile organic compounds (VOCs) are a large group of such chemicals that allow their producers (including plants and microorganisms) to defend themselves against attacks by pests or pathogens, or to convey warnings intra- or inter-specifically during such attacks (for details of VOCs, please refer to Poveda, 2021). Historically, the term “secondary” used for such natural metabolites has been associated with their “inessentiality,” but they have been demonstrated to play important roles in the growth and development of organisms in indirect ways (Pichersky and Gang, 2000; Deepika et al., 2016).
By preventing competition from other organisms, SMs help the organisms that produce them survive. They may also cause harm to other individuals of the same species. Examples of these SMs include those that help organisms access limited resources and survive in a specific adverse niche and those that inhibit competitors (Williams et al., 1989; Vaishnav and Demain, 2010; Xie et al., 2019; Figure 2b). In an in vitro competition experiment, naphthoquinone spiroketals, isolated from a newly identified EF strain, exhibited an allelochemical inhibitory effect against other EFs, such as Colletotrichum sp., Phomopsis sp. (Wang et al., 2016), and G. manguifera (Macías-Rubalcava et al., 2008; Macías-Rubalcava et al., 2014). In another in vitro dual culture experiment, it was observed that P. variabile, an EF, exhibited direct antagonism against F. oxysporum, a phytopathogen, by secreting the induced metabolite hydroperoxin oxylipin, which decreased the concentration of pathogenic mycotoxins, whereas none of the pure axenic cultures showed an increase or decrease of this metabolite (Combes et al., 2012). Further study has revealed that the biosynthesis of hydroperoxy oxylipins is catalyzed by lipoxygenases and the two lipoxygenase genes (pvlox1 and pvlox2) in P. variabile, and that only pvlox2 is specifically up-regulated during the interaction (Bärenstrauch et al., 2020).
The concept of coevolution has been applied to characterize the biochemical interactions between EFs and hosts. It is thought that the coevolution of endophytes and their host plants shapes the production of SMs, which play important roles in endophyte-host communication for mutual adaptation and their orientation to different environments (Debbab et al., 2011; Lind et al., 2017). There are three main schools of thought to interpret the relationship between the biosynthesis pathway of common SMs and the evolution of symbiosis between endophytes and their hosts. According to the hypothesis of the first school, this may occur in the era of coevolution when a responsive relationship has been established between EFs and host plants. EFs and host plants were adapted to each other, leading them to share common biosynthetic pathways of natural active SMs. In this case, the endophyte–host association is imperative because it may be the critical factor affecting the secretion of bioactive metabolites (Vaishnav and Demain, 2010). The environmental factors that affect symbiosis formation will also affect SM biosynthesis. The second school of thought suggests that endophytes gradually acquired adaptations to the internal microenvironment of hosts by horizontal gene transfer (HGT) between endophytes and plants during the primeval period (Slot and Rokas, 2011; Soanes and Richards, 2014). In such genetic variations, fungal strains insert some fragments of their genetic materials into plant genomes, or uptake some plant genetic fragments into their own genomes (see section “Horizontal Gene Transfer”). The third school of thought has argued that both endophytes and plants synthesize these common metabolites and transfer them to their corresponding symbiotic systems (Zhang et al., 2006).
Some investigations support the hypothesis that phytoendophytes originate from phytopathogens. Phylogenetic analysis has revealed the interconnections between phytoendophytes and phytopathogens in various environments (Arnold et al., 2009). SMs might also be involved in such interconnections (Di et al., 2016). Recent studies reported that the transient evolutionary roots of two fungal endophytic communities, Clavicipitaceous sp. N. coenophialum, an endophyte of tall fescue, and Harpophora oryzae, a beneficial endosymbiont of wild rice, originated from insect parasitoid and phytopathogenic ancestors, respectively (Spatafora et al., 2007; Xu et al., 2014). The exact phylogenetic mechanism of endophytes from apparent pathogens still remains unclear. It has been observed that the transfer of one specific F. oxysporum f. sp. lycopersici chromosome, which contains most of its effector genes and a secondary metabolite coding gene cluster, confers pathogenicity to an endophytic strain (Ma et al., 2010; Di et al., 2016). Studies suggest that environmental stresses, sudden drastic climate changes, or senescence may facilitate the conversion of endophytes into pathogens in the host to adapt to these ecological changes (Romero et al., 2001; Photita et al., 2004). They may also play a role in phylogenesis from pathogens to endophytes (Cui et al., 2021). Given such a prolonged virulent contradiction between pathogens and plants, the loss-of-function mutations of the virulence genes in pathogenic strains and the alteration of the SM biosynthetic pathway and accumulation of SMs eventually convert such pathogen–host interactions into mutual symbiotic relationships that are beneficial to both parties (Tan and Zou, 2001; Kemen et al., 2015). It has been observed that plants infected with EFs have more bioactive chemical compounds than those infected with pathogenic fungi (Schulz et al., 2002). Generally, when facultative EFs begin to colonize host plants, they face three kinds of competitors: epiphytes, pathogens, and the host defense systems. This may explain why plants infected with EFs produce more defensive chemicals than plants infected with pathogens (Schulz et al., 2002; Hunt and Newman, 2005; Figure 2b). These examples illustrate the significance of metabolic communications for the construction of multiple interactions among EFs, host plants, and other plant microbiomes, and for the identification of diverse metabolic chemicals secreted by plants or EFs during their interaction (Friesen et al., 2011; Quambusch et al., 2014; Hardoim et al., 2015).
In competitive and unfavorable ecological environments, plant species modify their biological systems by producing various defense reactions, which are mainly manifested in the synthesis of defensive SMs. It is estimated that plant genomes may contain more than 70,000 genes (Wang et al., 2018), of which 15–25% are involved in encoding enzymes that participate in secondary metabolism (Bevan et al., 1998; Somerville and Somerville, 1999). This indicates that defensive SMs are often highly diverse within and across populations. EFs also take part in the establishment of host defense mechanisms through SMs to enhance the host capacity to adapt to a wide range of biogeographical ecosystems (Violle et al., 2014). One reason for the diversity of the SMs of an EF species is the diverse arrangement of coding genes in the SM synthetic pathway. As the coding genes of SM synthetic pathways in EFs are usually in clusters (Andersen et al., 2013), the divergent rearrangements of the clusters within an EF species or across EF species may also contribute to the diversity of SMs. Recently, at least five divergent types of variation in SM gene clusters in the fungal species Aspergillus fumigatus were identified, revealing its diverse secondary metabolism (Lind et al., 2017). Although their mechanism is still poorly understood, SMs may cause the morphological or physiological alteration of host plants (Braga et al., 2018). These alterations are believed to be linked to plant diversity, which means that the colonization of EFs enhances the host adaptability and thus its viability under various environmental conditions (Hyde and Soytong, 2008); in return, the survival of EFs may also be enhanced through their host diversifications.
Analyzing the gene expression regulation mechanism of EFs, their hosts, and their symbiosis contributes greatly to further understanding the interactions between the two symbiotic partners (Deepika et al., 2016). The expression of SM coding genes in a symbiotic continuum may be regulated by factors including gene clustering, transcription factors, the presence of EFs, and HGT.
The diversity of EF genomes in eukaryotic systems and fungal genetic studies has revealed that genes coding specific SM biosynthesis pathways are clustered. In some cases, such as when the pathway function requires specific transcription factors or transporters, the genes encoding these factors (enzymes) are also located in these clusters (Nützmann et al., 2018). Two findings suggest that the architecture or structure of fungal secondary metabolic clusters varies with the situation. First, some highly complex SMs are synthesized through collaboration between different clusters (Chiang et al., 2016; Henke et al., 2016). Second, in some cases clusters of different pathways are adjacent to each other in the genome (Nützmann et al., 2018). These clusters are usually found in the dynamic regions of chromosomes or near telomeres. Subtelomeric regions are well-known hotspots in chromosomal recombinations and segmental duplications (Field et al., 2011; Andersen et al., 2013; Lind et al., 2017). The mechanism of SM-coding gene clusters in unstable DNA regions is still unclear, but an acceptable explanation is that it may be related to gene expression regulation (Osbourn, 2010a). The proximity of clustered genes may be a necessary condition for the synthesis of bioactive products related to a pathway, because it keeps the pathway genes closer during genomic rearrangements (Deepika et al., 2016). This clustering genetic format may facilitate the co-inheritance of favorable combinations of alleles at these multigene loci (Chu et al., 2011; Field et al., 2011). It may also monitor the synchronization of the clustered gene expression by altering the arrangement of chromatins (Hurst et al., 2004; Sproul et al., 2005; Field and Osbourn, 2008; Osbourn and Field, 2009; Osbourn, 2010b; Field et al., 2011) and by exchanging the non-contiguous regulatory elements, which are harbored in the clusters (Hurst et al., 2004; Sproul et al., 2005; Osbourn and Field, 2009; Osbourn, 2010b). In the genomes of higher plants, the functionally correlated genes interspersed in the genome may form clusters at the transcriptional level through helix-loop-helix domains (Nützmann et al., 2018). The formation of DNA loops causes cis-elements to be located adjacent to each other and creates high local concentrations of transcription factors that are close to the transcription initiation sites of the genes, thereby initiating transcription (Mendes et al., 2013). Any interference with the transcription of clustered genes will not only cause the loss of the coding products of these genes, but may also cause toxic intermediates to accumulate in the biochemical pathways (Hurst et al., 2004; Sproul et al., 2005). Evidence indicates that gene regulation at chromatin levels is important for the expression of secondary metabolic gene clusters (Osbourn, 2010b).
The activities of these clustered genes in the secondary metabolism are further regulated by two main groups of transcription factors: narrow domain transcription factors (NDTFs) (Table 1) and broad domain transcription factors (BDTFs) (Table 2; Keller et al., 2005). NDTFs act on the genes in the cluster, and may also act on the clustered genes at different genomic locations from NDTFs themselves. This can be illustrated by the typical NDTF AflR, which is a well-characterized Zn2Cys6 transcription factor that regulates the aflatoxin and sterigmatocystin gene clusters through binding to the palindromic sequence 5′-TCG(N5)CGA-3′, an 11-bp motif in the promoter regions of coding genes in a few Aspergillus species (Slot and Rokas, 2011). It also regulates three more genes outside the aflatoxin metabolite gene cluster (Woloshuk et al., 1994; Yu et al., 1996; Cánovas et al., 2017). Some typical NDTFs are shown in Table 1. BDTFs, or global transcription factors, are upper hierarchical-level control systems that respond to external stimuli that are not directly related to secondary biochemical gene clusters (Keller et al., 2005; Yu and Keller, 2005; Young et al., 2006; Hoffmeister and Keller, 2007; Fox and Howlett, 2008; Shwab and Keller, 2008; Deepika et al., 2016). Studies have revealed that EF signals interfere preferentially with ethylene-targeted transcription factors (Camehl et al., 2010). It is well-accepted that the biosynthesis of SMs depends on a combination of developmental competence and the stimulation of environmental factors, such as nutrient availability, illumination, pH, injury, infection, and developmental changes during the host life cycle (Bayram and Braus, 2012; Xie et al., 2019). BDTFs play an essential role in the transmission of environmental stimuli to the genome. They create and regulate the signaling transduction from environmental cues to cellular responses in the formation of specific SMs (Keller et al., 2005). Some typical BDTFs are shown in Table 2.
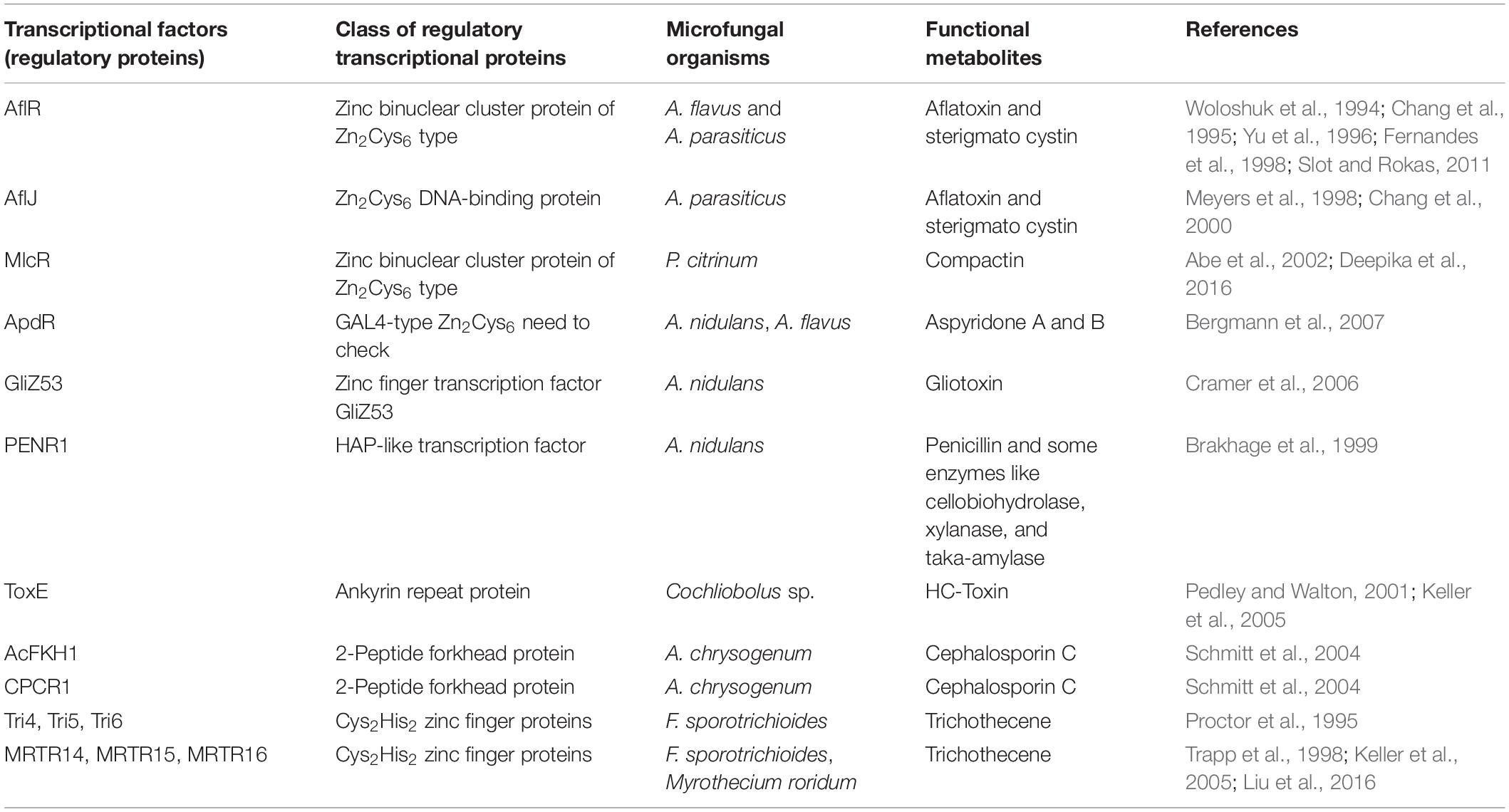
Table 1. Narrow domain transcription factors (NDTFs).
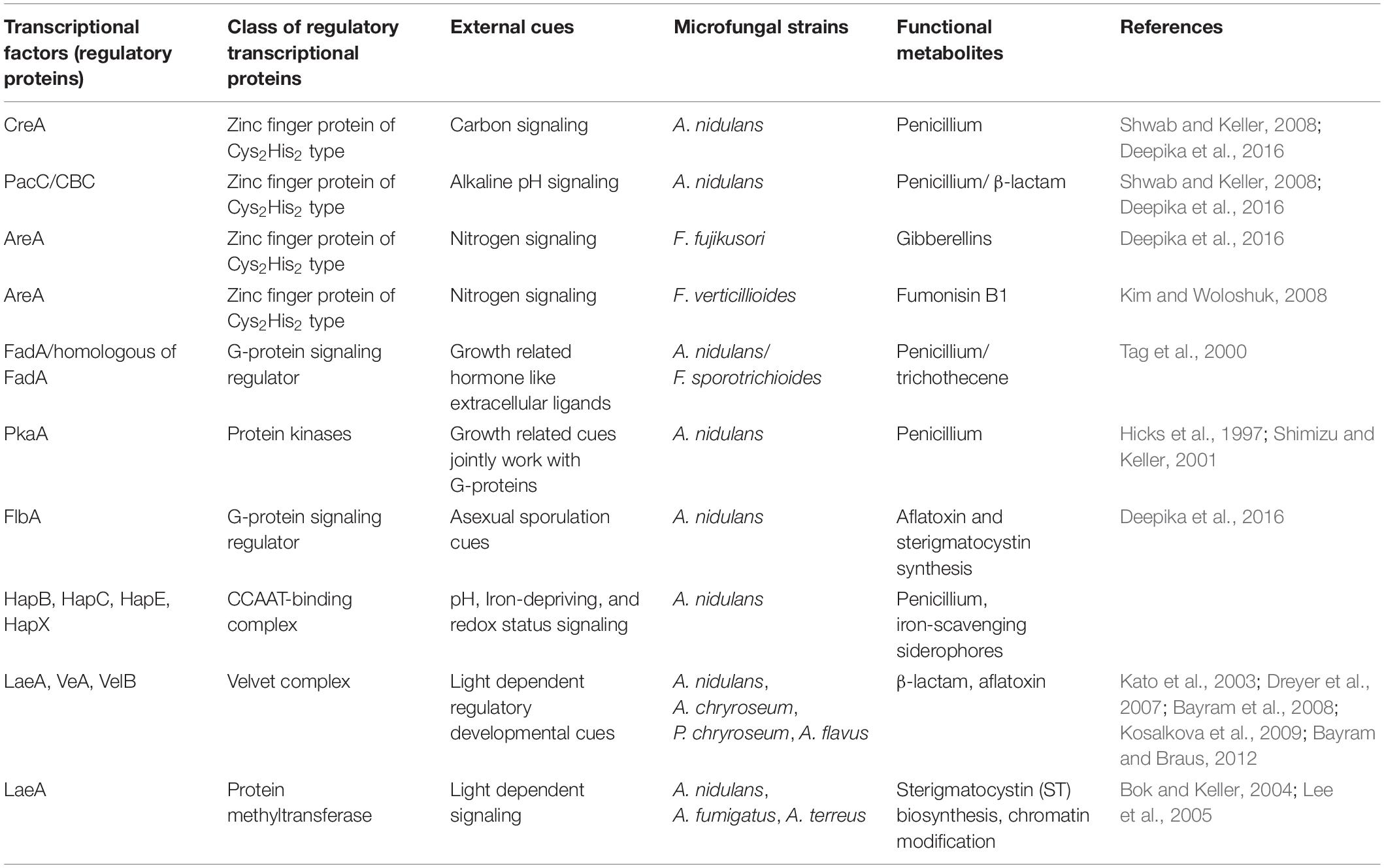
Table 2. Broad domain transcription factors (BDTFs)/global transcription factors.
The close relationship between NDTFs and BDTFs has been extensively reviewed by various reports, which have explained how BDTFs or global transcription factors perceive the environmental and developmental cues and transduct these external messages to NDTFs through chromatin and histone modification or through specific biochemical cascade reactions, including methylation, phosphorylation, and acetylation. These reactions are essential for activating the silent clustered genes associated with specific SMs that are required in particular cellular metabolisms, growth stages, or environmental conditions (Figure 3; Foss et al., 1993; Kouzminova and Selker, 2001; Tamaru and Selker, 2001; Calvo et al., 2002; Yu and Keller, 2005; Shwab and Keller, 2008).
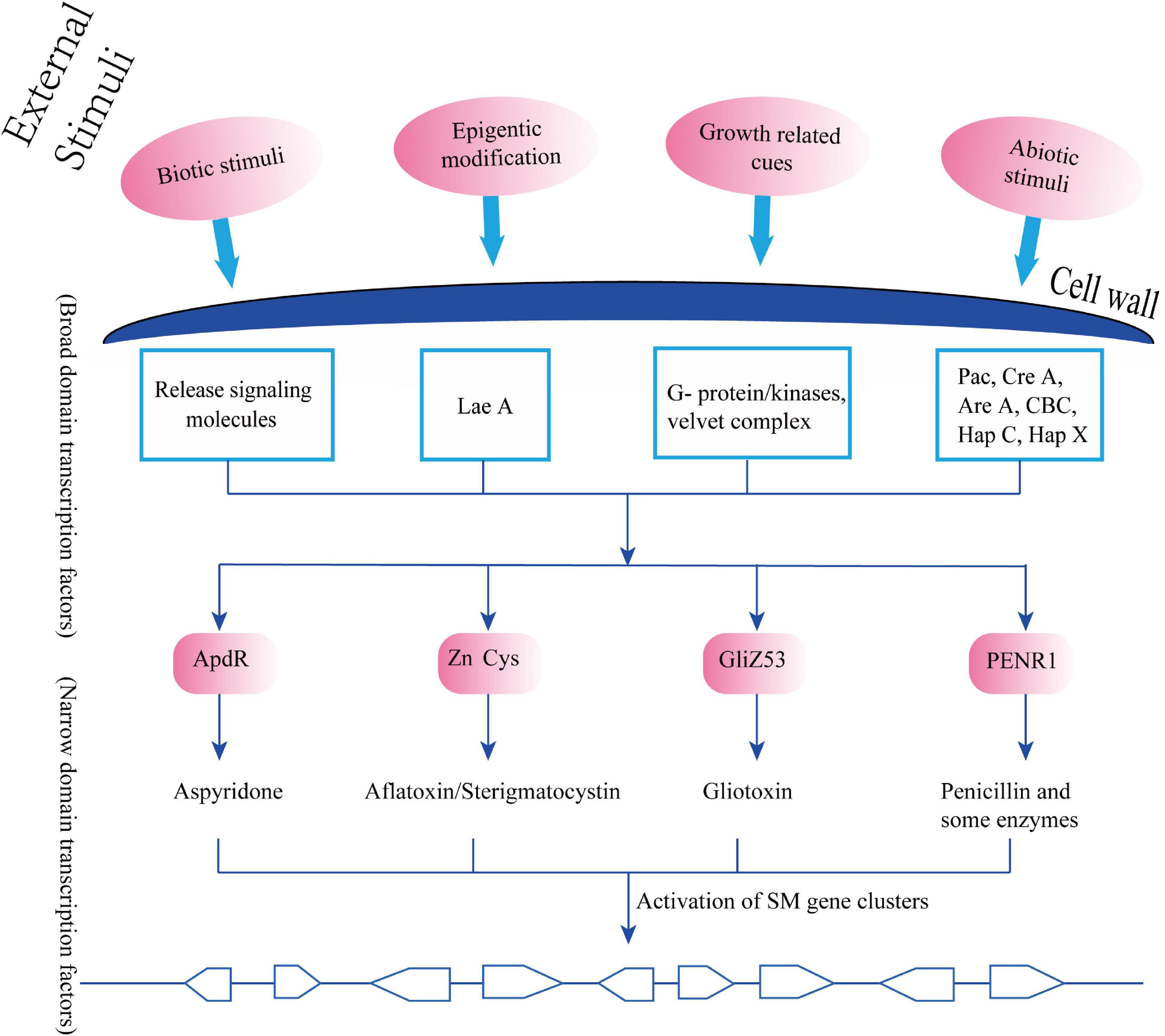
Figure 3. Synchronized regulatory model of secondary metabolites (SMs) biosynthesis and external/morphological indicators.
Plant phenotypes depend not only on the genetic makeup of the plant, but also on the activity of microbiome and environmental factors (Jia et al., 2016). It has been observed that the presence of EFs enhances the resistance potential of host plants to biotic and abiotic stresses (Rodriguez et al., 2008; Shoresh et al., 2010; Busby et al., 2017; Yan et al., 2019) or improves nutrient uptake (Xie et al., 2019). Although the exact mechanism still remains to be elucidated, evidence has shown that plant genetic expression profiles are altered in the presence of EFs (Mejia et al., 2014; Cui et al., 2017; Liao et al., 2019; Xie et al., 2019). Endophytes reprogram the host genomic expression through epigenetic interactions with the host. In epigenetic events, DNA methylation and demethylation induced by endophytes enhance the host’s defensive processes (Kouzminova and Selker, 2001; Strobel, 2018; Geng et al., 2019). EFs stimulate the immune systems of plants and increase the quantity of SMs (Cui et al., 2017; Xie et al., 2019), which may induce some physiological changes in infected host plants. Bailey et al. (2006) regarded these altered or differentiated gene expressions in the presence of EFs as a complex system of genetic crosstalk between EFs and hosts. During the crosstalk, the genomic expressions of both endophytes and hosts are altered. Additional examples are given in Table 3.
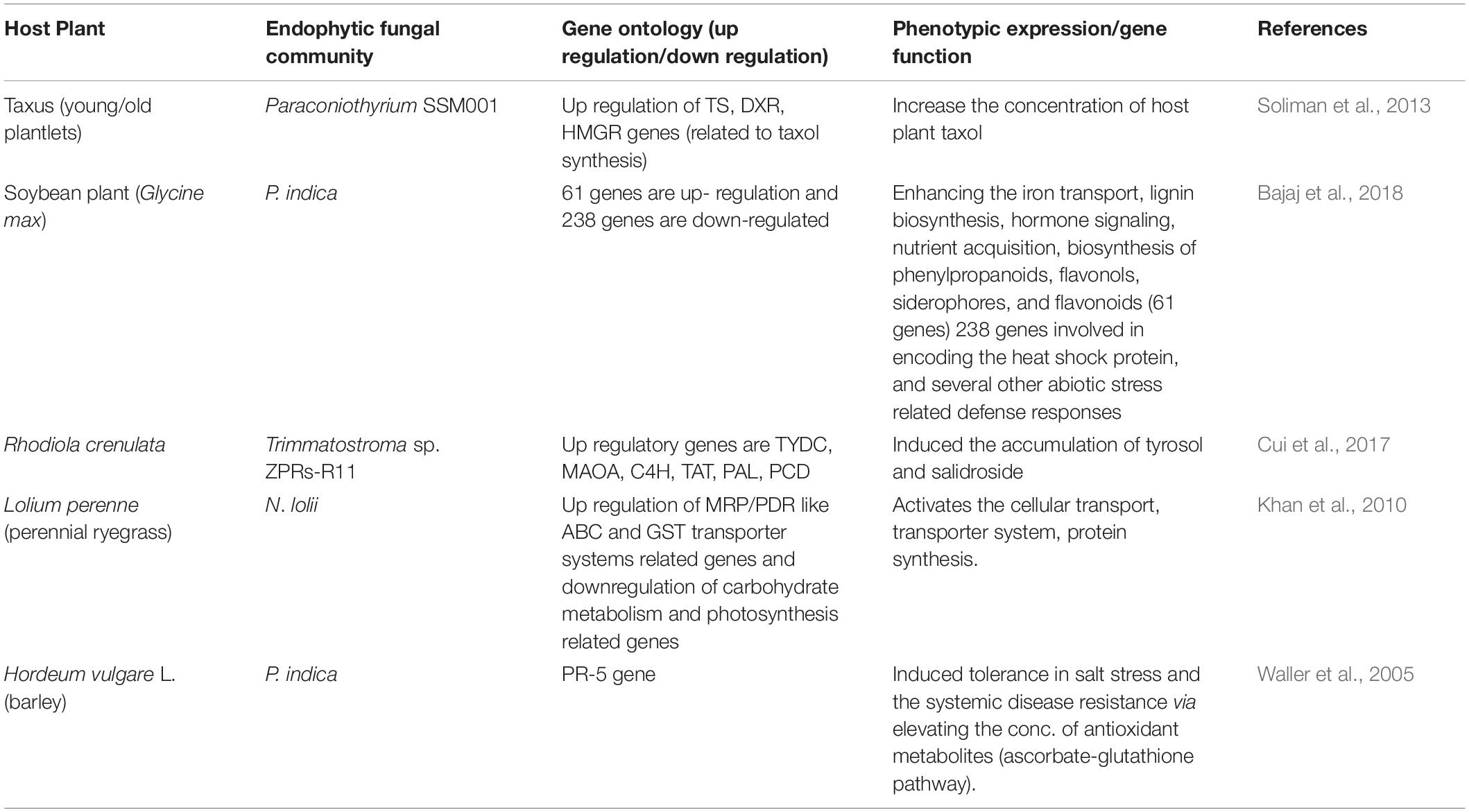
Table 3. Selected examples of plant gene alteration in the presence of endophytic fungi (EFs) and their resulted beneficial expression.
Horizontal gene transfer refers to the exchange of genetic materials between isolated lineages through asexual processes. A growing body of evidence has suggested that frequent HGT occurs between independent organisms and between EFs and their hosts (Richards et al., 2011; Bapteste et al., 2012). Most HGTs identified in microbiota are associated with invading, degrading, and manipulating hosts (Soanes and Richards, 2014), which supports the hypothesis that HGT may be a robust source of novel adaptive traits of EFs and that HGT is a strong driving force for fungal adaptive evolution (Feurtey and Stukenbrock, 2018). As the coding genes of a metabolic pathway are always located in a cluster, HGT of an intact metabolic cluster may enable the recipient organism to obtain a new and complete metabolic pathway (Slot and Rokas, 2011). Bapteste et al. (2012) identified a particular evolutionary unit of genetic materials that introgress into different host structures and propagate within these structures. These evolutionary units may leave recognizable patterns in resemblance networks. This finding suggests that HGT also plays an important role in the emergence of new fungal strains (Feurtey and Stukenbrock, 2018).
The expression of horizontally transferred genes in recipient organisms is the first step for HGT to play a role, and determine the colonization mode and capacity of EFs in host plants (Soanes and Richards, 2014). The mechanism of how horizontally transferred genes initiate their expression in recipient organisms remains to be discussed. Slot and Rokas (2011) demonstrated that when the gene cluster encoding the biosynthesis of sterigmatocystin (a toxic SM) was transferred from an Aspergillus species to Podospora anserine, the transferred cluster was functional because it was expressed in the latter. Many studies have demonstrated that EFs can produce host-mediated compounds, whereas the latter can also produce some EF-originated metabolites. For instance, djalonensone has been detected as a fungal metabolite in various Alternaria fungi, but it was first identified in extracts from cauliflower plants. Similarly, the EF metabolite aureonitol produced by Chaetomium sp. can also be detected in the extract of Helichrysum aureonitens. Many reported EF-originated compounds, such as alternariol, alternariol monomethyl ether, altenusin, macrosporin, and methylalaternin, were further detected in their host plants, Polygonum senegalense and Urospermum picroides. These results provide evidence for HGT or genetic recombination during the coevolution between hosts and endophytes, thus forming the genetic regulation mechanism of SM biosynthesis (Aly et al., 2010; Kozyrovska, 2013).
When HGT occurs between prokaryotes and eukaryotes, the integrated genes are more likely to be expressed in a modified manner in the recipient organism due to the fundamentally distinct gene expression mechanisms between the two genomes. The failure of HGT expression may lead to the appearance of pseudogenes in recipient organisms (Andersson, 2009).
The wide range of biotic and abiotic stresses that plants constantly face in natural or agricultural environments lead to specific transcriptional variations at the individual gene level, with a high degree of variability and stress specificity (Zhang and Sonnewald, 2017). This was supported by a study of plant responses to combined heat and drought stress, in which the genes involved in secondary metabolism were significantly up-regulated (Prasch and Sonnewald, 2013). In symbiosis, EFs mimic the production of SMs in hosts using primary phytocompounds as precursors. In contrast, proteins secreted by EFs may potentially act as effectors altering host metabolism (Spanu et al., 2010; Soanes and Richards, 2014; Kemen et al., 2015), with the specificity of each effector targeting a distinct hormone signaling pathway (Di et al., 2016).
A “xenohormesis” hypothesis proposed by Howitz and Sinclair (2008) suggests that heterotrophic organisms (animals and microbes) may sense plant signals or molecules induced under stress. By doing so, the heterotrophic organisms may take advantage of the defensive responses of autotrophs to increase their own survival chances. EFs may also have the ability to sense chemical cues in plants, and begin to produce similar SMs (Kusari et al., 2012). As ecological adaptions and survival elements, SMs are not synthesized throughout the whole life cycle, but only when necessary, especially in the early differentiation or late senescence stages of an organism (Deepika et al., 2016; Stahl et al., 2018; Siddhardha and Meena, 2020). In general, their synthesis is minimal when the organism grows rapidly and is maximal when the growth of the organism ceases. For an organism, the core indicator point of its synthesis of SMs is that it is in a state of rapid response rather than in a state of rapid growth (Deepika et al., 2016). In such circumstances, the genetic materials of both EFs and their hosts are cross-activated under hostile conditions (Stierle et al., 1993; Zhou et al., 2010). The best example, which well supports the previous hypothesis, is the pioneering identification of the anti-carcinogenic drug Taxol (paclitaxel), which is a worldwide selling cancer drug, with annual sales of million USD, from an EF, Taxomyces andreanae. This EF was originally identified from Taxus brevifolia, the host plant of T. andreanae, indicating that both the EF and its host produce the same SM in response to environmental stimuli. Since its initial discovery in the past few decades, Taxol has been proven to be more efficiently produced in the EF than in the host. Studies also show that Taxol can be effectively isolated from several other EF strains and Pacific yew plants (Stierle et al., 1993; Zhou et al., 2010; Chandra, 2012; Zaiyou et al., 2017; Uzma et al., 2018).
Primary metabolites are the end products of primary metabolic pathways, including carbohydrates, amino acids, proteins, and lipids. They play a primary metabolic role in the construction and development of an organism. Without them, the growth and development of the organism are at extreme risk for defects. An important role of primary metabolism is that the products of some key steps provide precursors for the synthesis of SMs. Endophytes and their host plants share these precursors in their respective SM biosynthesis pathways. The biosynthetic pathway of SMs in EFs may be the result of their mimicking of the host pathways (Kirby and Keasling, 2009; Deepika et al., 2016). The synthetic pathways of some phytochemicals, including ergot alkaloids, aflatoxin, and lovastatin, have been studied by blocking mutant and radio labeling techniques (Keller et al., 2005). It has been revealed that although diverse, SMs are produced by a few common biosynthetic pathways and the metabolomic pathways of endophytic fungal communities and their host plants are similar. At this stage, the question still remains of whether these low molecular weight phytochemicals are synthesized by plants or as a result of symbiosis with microorganisms living inside their tissues. The combination of some possible inducers has promoted the accumulation of bioactive metabolites in EFs and plants, indicating that EFs play a significant role in the biosynthesis of SMs. These inducers include nutrient deficiencies, morphological development, and growth rates (Kirby and Keasling, 2009; Deepika et al., 2016).
EFs are considered to be rich sources of diverse bioactive SMs and phytohormones to support plant growth and enable plants to survive under biotic or abiotic stresses (Tan and Zou, 2001; Chadha et al., 2014). Some of the most important commercially exploited SMs, including antibiotics, anticarcinogenics, cytotoxics, insecticides, and allelopathic compounds, can also be biosynthesized by EFs (Schneider et al., 2008). They have broad, promising commercial prospects in the pharmaceutical, medical, agricultural, nutraceutical, cosmetic, flavor, and fragrance industries, making EFs an attractive topic in the field of endophytism research (Hyde and Soytong, 2008; Table 4). Therefore, exploring the benign symbiotic relationship that synthesizes these bioactive SMs between EFs and plants and its impact on the genetic materials of EFs and plants will provide a promising framework for the discovery and development of new bioactive SMs through metabolomic and genetic engineering in the future (Kirby and Keasling, 2009; Deepika et al., 2016). Of the numerous SMs biosynthesized by EFs and hosts, most can be sorted into the following classes: alkaloids, terpenoids, polyketides, phenylpropanoids and lignins, flavonoids, saponins, phenols and phenolic acids, aliphatic, and chlorinated metabolites, peptides, and steroids. To facilitate the comparison of these SMs, they are presented in tabular form in Table 5 and Figure 4 after a brief description of their EF sources, chemical structures, and biological application potential.
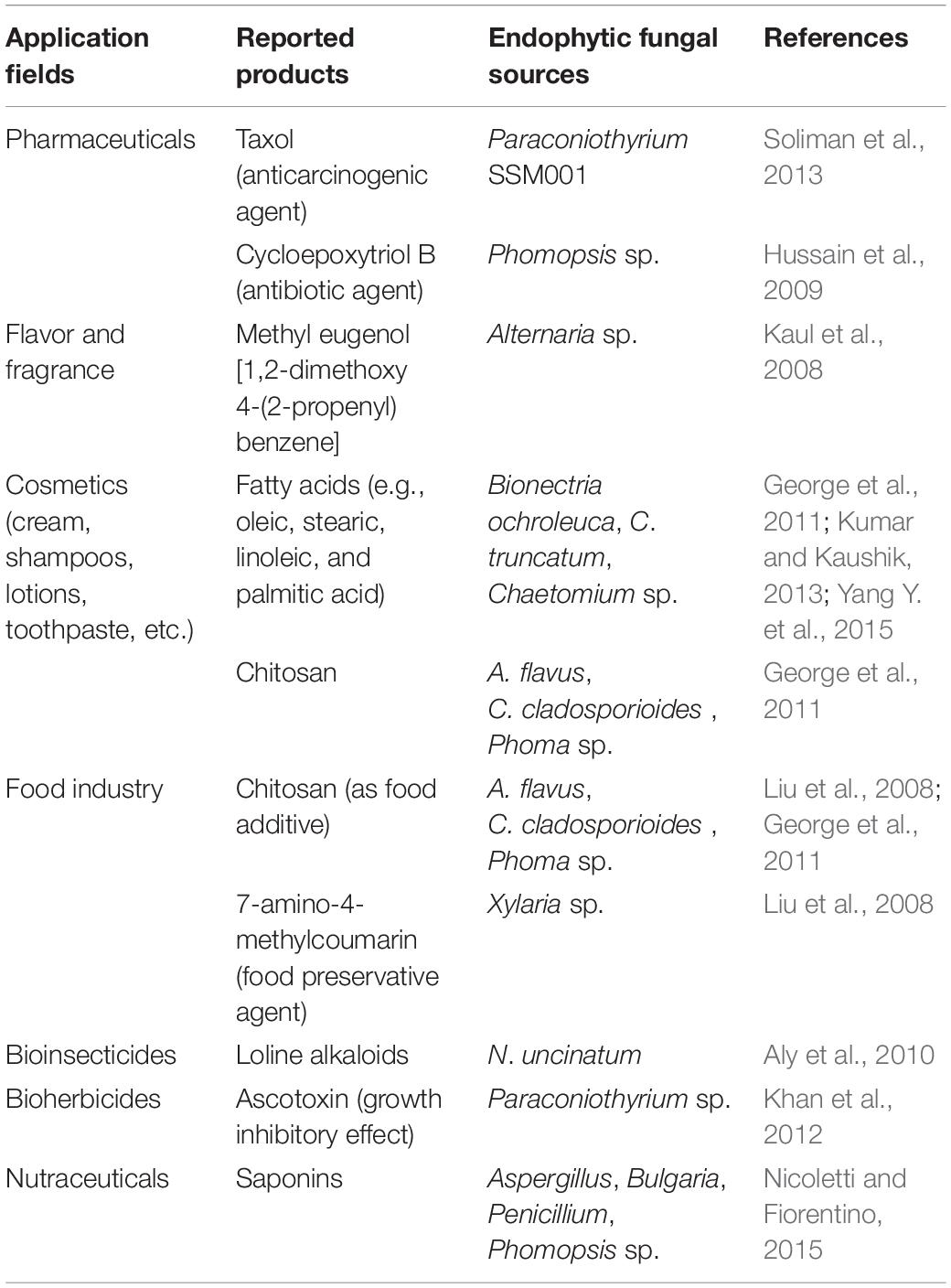
Table 4. Commercial applications of bioactive natural products with endophytic fungi (EFs)-based biogenesis.
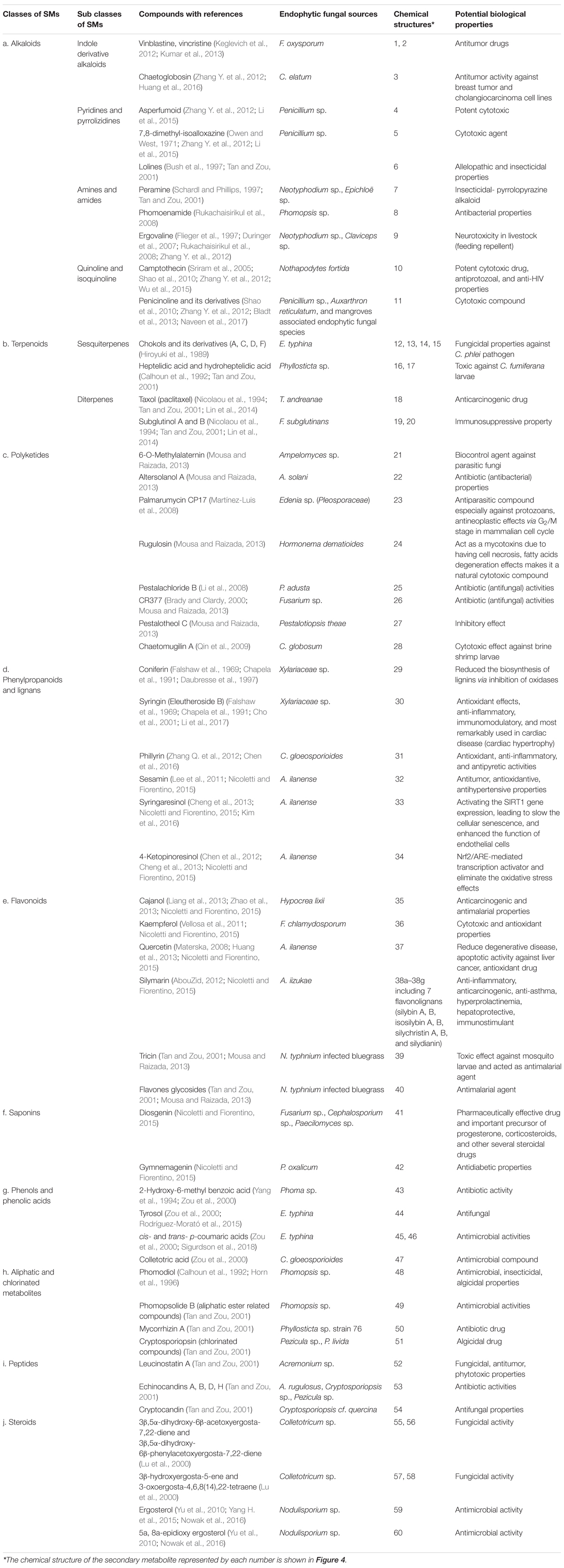
Table 5. Classes of endophytic fungal secondary metabolites (SMs) with biological potential activities.
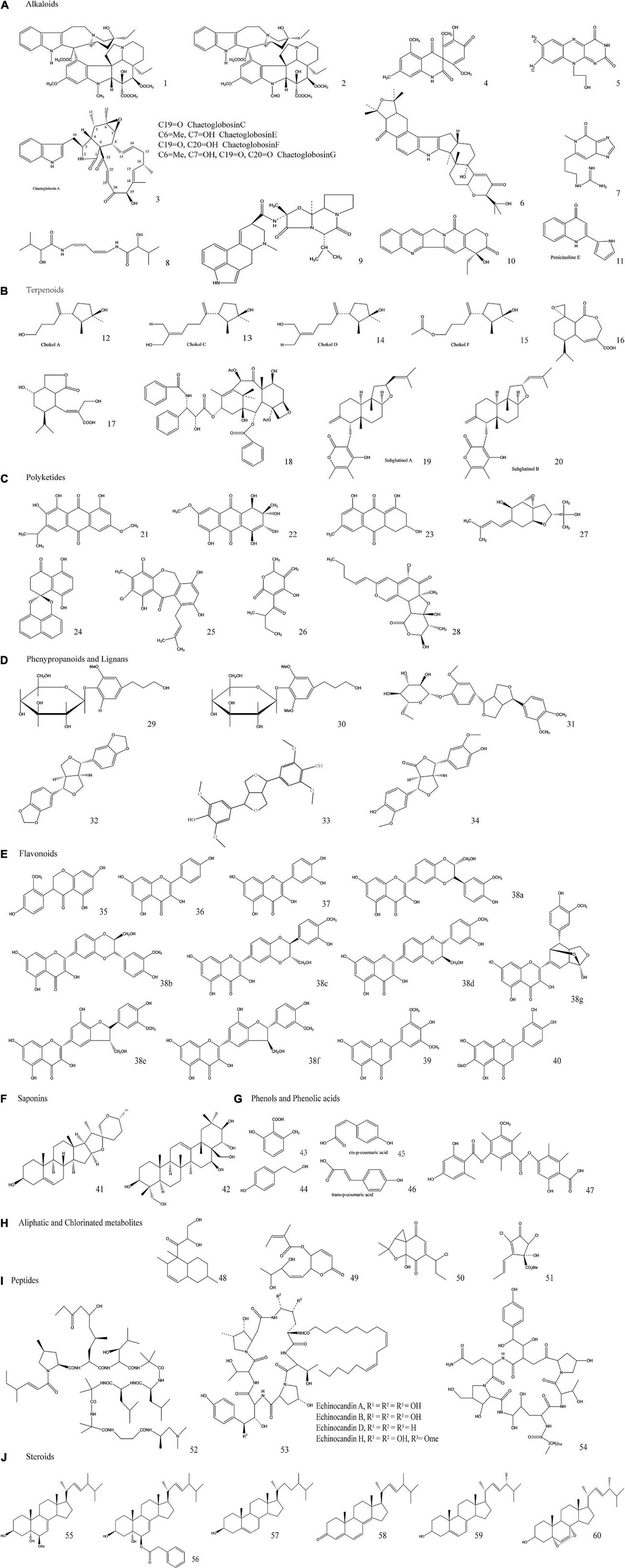
Figure 4. Molecular structures of some typical secondary metabolites (SMs). The classes, subclasses, compounds with references, endophytic fungal sources, together with their potential biological properties of the SMs are briefed in Table 5.
a. Alkaloids
Alkaloids include indole derivatives, pyrimidine and pyrrolizidine, quinoline and isoquinoline, amines, and amides (Zhang Y. et al., 2012). They are isolated from EFs that colonize in grass (Clavicipitaceous genera: Epichloë sp.) (Guerre, 2015), and because of their toxicity, they enable hosts to inhibit insect and herbivore attacks. In addition, their concentration levels are mainly dependent on EF strains, species, or genotypes and less dependent on environmental factors and host genotypes (Zhang et al., 2006; Gundel et al., 2018). Alkaloids are composed of bioactive compounds. In addition to complex chemical properties, alkaloids have bioactive properties, such as fungicidal, antibiotic, and antitumor activities, making them the main focus of numerous drug research and development projects (Zhang et al., 2006; Song et al., 2020).
b. Terpenoids
Terpenoids are the second major group of plant SMs. They also are obtained from endophytic mycobiota. Their primary characteristics include defensive agents, pollinator attractors (mainly odoriferous or color components), and allelochemicals in competitive environments. Among terpenoids, sesquiterpenoids, and diterpenoids can be easily isolated from endophytic cultures without degradation (Jakubczyk and Dussart, 2020).
c. Polyketides
Polyketides are the most copious and well-characterized fungal metabolites, and are exclusively detected in mycospecies. Polyketide synthases play the most important role in their biosynthesis. These synthases are similar to those of eukaryotic fatty acid, but their products are different due to the selective reduction of the β-carbon in polyketides rather than the compulsive reduction of that position in fatty acids (Keller et al., 2005; Jakubczyk and Dussart, 2020).
d. Phenylpropanoids and lignins
These types of SMs are produced in wounds and lesion conditions to protect the host from pathogenic attacks. Lignin compounds play a role in initiating wound healing (Yadav et al., 2020). EFs also produce these phytochemicals to improve the adaptability of host plants to adversity. These compounds can be isolated from both infected plants and separately cultured EFs (Dixon and Paiva, 1995; Sampangi-Ramaiah et al., 2020).
e. Flavonoids
Flavonoids are essential plant natural products that have polyphenolic moieties, but they are also EF derived. Due to their antioxidant, anti-inflammatory, antimicrobial, antimutagenic, and anticarcinogenic effects, they have significant miscellaneous biochemical, cosmetic, nutritional, and pharmaceutical applications for the treatment of various ailments, such as Alzheimer’s disease, cancer, and atherosclerosis. In addition, flavonoids are basic ingredients in the field of cosmetics (Panche et al., 2016).
f. Saponins
Saponins are a kind of glycoside compound in which the sugar moiety attaches to sapogenin through a glycosidic bond. Because of their antimicrobial properties, saponins play a defensive role in their symbiosis with host plants. Other applications are related to their anticancer, antinutritive, and anticholesterol properties (Nicoletti and Fiorentino, 2015).
g. Phenols and phenolic acids
Phenols and phenolic acids are a diversified class of SMs that are principally synthesized by plants and have also been isolated and identified from EF sources. Their major function is to act as signal factors in plant–microbial associations. They also act as defensive agents, promoting plant growth in nutrient-limited soil during symbiosis (Singh et al., 2011). This may be due to the higher concentrations of phenolic acids, phenols, and their derivatives in host plants inoculated with EFs (Mandal et al., 2010).
h. Aliphatic and chlorinated metabolites
Aliphatic and chlorinated metabolites have simple chemical structures but are considered xenobiotic compounds. These metabolites are widely biosynthesized by several forest and wood litter-degrading fungal species and endophytes. They have antibiotic activities against pathogenic microorganisms, insects, and algae, but they are also carcinogenic and genotoxic to animals and humans (Lin et al., 2011).
i. Peptides
Endophytic fungal peptides are another class of SMs that act as defense agents. They are protein forms with a molecular weight of less than 10 kDa (Ng, 2004). Significant scientific efforts have been directed toward identifying and isolating peptides as candidate drugs due to their high degree of interactions with their specific targets. Studies have reported endophyte isolates as potential sources of peptide-based drugs for the treatment of a variety of illnesses (Abdalla and Matasyoh, 2014). The most important group of anticarcinogenic and antifungal peptides, Leucinostatin, has been extracted from the fungal endophyte Acremonium sp., which is isolated from T. baccata (European yew plant) (Abdalla and Matasyoh, 2014).
j. Steroids
Steroids are natural chemical substances that are abundantly produced not only in plants and animals, but also in microbial communities. Steroids are bio-lipid-based terpenoids that have a structure of four fused carbon skeleton-based rings, which is characterized as the steroid nucleus or sterane. Steroids vary in structure and function because of the different oxidation states of functional groups attached to the rings (Theodorakidou et al., 2018).
It has already been demonstrated that endophytic steroids such as ergosterol exhibit extra pharmaceutical activities and natural roles in their producers. The 5a- and 8a-epidioxy ergosterols that have been isolated from Nodulisporium sp. have potent antimicrobial activities against a series of pathogenic microbial strains (Yu et al., 2010; Yang H. et al., 2015; Nowak et al., 2016).
At present, endogenous biology is receiving increasing attention due to the great application potential of the chemicals secreted by EF–host symbiotic associations in sustainable agriculture and biomedicine. Scientists are interested in understanding the underlying mechanisms of endophytism and its biological and ecological roles. Its importance is highlighted by the large number of studies in the field of endophytic biological research (Hyde and Soytong, 2008; Deepika et al., 2016; Nützmann et al., 2018; Fang et al., 2019; Huang et al., 2019). Although EF research has attracted great attention, this field still faces substantial challenges to be addressed in the coming decades, including:
• The selection of suitable host plants and their healthy organs or tissues to identify and isolate new EFs and to dissect their related mutualistic or antagonistic signaling mechanisms during symbiosis (Strobel, 2018).
• The complexities in the process of artificial culture due to the aseptic or non-culturable characteristics of some fungal strains. It is important to introduce de novo bioengineering systems or to modify conventional isolation techniques to address this challenge (Stone et al., 2004; Zhang et al., 2006; Zhang P. et al., 2019).
• The biosynthesis of natural products of EFs, especially SMs, requires inducing stimuli from host plants or from symbiosis. In artificial axenic culture, EFs may not be able to synthesize the same chemicals as they do in a symbiotic continuum due to the absence of these plant-mediated stimuli or signals. For the EFs that have been successfully isolated and cultured, their SM production decreases with successive subcultures under axenic monoculture conditions (Cánovas et al., 2017). Therefore, it is also a challenge to monitor the in vivo stimuli of successfully isolated EFs under quasi-natural conditions and maintain their “competency” in culture.
• Fungal SMs play critical roles in understanding the endosymbiotic mechanisms between EFs and hosts. This endosymbiosis can be indirectly monitored by the transformation biogenesis of SMs under artificial culture conditions. Considering the significant impact of different types and concentrations of nutrients in artificial media in EF culture (Ruiz et al., 2010), this is not an easy job.
• The degradability of SMs extracted from target EFs needs to be addressed. Compounds isolated from a symbiotic continuum may be highly unstable in vitro or in axenic culture. Thus, it is difficult to obtain these required novel compounds in an artificial medium.
• The number of EFs that have been explored is limited. It is estimated that only 1-2% of about 300,000 plant species have been investigated, and at present, little information has been obtained from hydro-ecosystems, which means that the vast majority of EF symbiotic relationships remain elusive (Strobel, 2018).
In the current “omics” era, tools including genomics, epigenomics, transcriptomics, proteomics, and their related meta-omics (metagenomics, metatranscriptomics, and metaproteomics) (Zhang J. et al., 2019), will be extraordinarily supportive in illuminating the gray areas of myco-endophytisms and in tackling the aforementioned challenges to reveal complementary information on these symbionts and their interactions inside the internal niches of host plants (Peršoh, 2015; Nowack and Weber, 2018; Zhang J. et al., 2019). Furthermore, collaborative research between omics tools and other disciplines, such as combinatorial chemistry, will be more effective and fruitful in constructing novel molecular models of these EF–host interactions (Nowack and Weber, 2018).
Secondary metabolites play a pivotal role in mediating biochemical communications between EFs and host plants. These biochemical communications guide the multidimensional interactions among EFs, host plants, and pathogens in their community and determine the host range of an EF and endophyte populations in a plant host. The biosynthesis of SMs during symbiosis is precisely regulated by several genetic mechanisms, including gene clustering, transcription factors, the altering of the genetic makeup of the host in the presence of EFs, and HGT. These regulation mechanisms may have coevolved with the initiation of EF–host symbiosis. Recently, SMs have attracted widespread research efforts due to their great biological potential in the discovery of modern medicines, sustainable agriculture, and industry. Further investigations at the molecular level may still be needed for gaining a better understanding the endophyte–host relationship in natural ecosystems at the genomic level, and for efficiently identifying the hidden genes involved in the biosynthesis of SMs and new compounds in axenic culture.
BA and WG: conceive the idea and write the initial draft of the manuscript. JL and QG: help organizing and editing the manuscript. QG and WG: perform the network construction and figure presentation. MK, JG, and SM: proofreading the manuscript. YY and WG: contribute to the final editing of the manuscript. All authors contribute in the interpretation of the manuscript and approve it.
This work was funded by the Natural Science Foundation of China (31621005), the National Key R&D Program of China (2017YFD0101603-11, 2016YFD0100500, and 2016YFD0101401), the Agricultural Science and Technology Innovation Program for CAAS (CAAS-ASTIP-ICRCAAS), the National High Technology Research and Development Program of China (2012AA101108 and 2009AA101104), and the Central Level of the Scientific Research Institutes for Basic R&D Special Fund Business (1610162014008).
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
We acknowledge SciLang for its linguistic assistance during the preparation of this manuscript and Baocai Zhang, Institute of Genetics and Developmental Biology, Chinese Academy of Sciences, for his critical review and valuable suggestions to the manuscript.
Abdalla, M. A., and Matasyoh, J. C. (2014). Endophytes as producers of peptides: an overview about the recently discovered peptides from endophytic microbes. Nat. Prod. Bioprospect. 4, 257–270. doi: 10.1007/s13659-014-0038-y
Abe, Y., Ono, C., Hosobuchi, M., and Yoshikawa, H. (2002). Functional analysis of mlcR, a regulatory gene for ML-236B (compactin) biosynthesis in Penicillium citrinum. Mol. Genet. Genomics 268, 352–361. doi: 10.1007/s00438-002-0755-5
AbouZid, S. (2012). “Silymarin, natural flavonolignans from milk thistle,” in Phytochemicals-A Global Perspective of Their Role in Nutrition and Health, ed. V. Rao (London: IntechOpen).
Abreu-Tarazi, M. F., Navarrete, A. A., Andreote, F. D., Almeida, C. V., Tsai, S. M., and Almeida, M. (2010). Endophytic bacteria in long-term in vitro cultivated “axenic” pineapple microplants revealed by PCR–DGGE. World J. Microbiol. Biotechnol. 26, 555–560. doi: 10.1007/s11274-009-0191-3
Albrectsen, B. R., Witzell, J., Robinson, K. M., Wulff, S., Luquez, V. M. C., Ågren, R., et al. (2010). Large scale geographic clines of parasite damage to Populus tremula L. Ecography 33, 483–493. doi: 10.1111/j.1600-0587.2009.05982.x
Alvear-Daza, J. J., García-Barco, A., Osorio-Vargas, P., Gutiérrez-Zapata, H. M., Sanabria, J., and Rengifo-Herrera, J. A. (2021). Resistance and induction of viable but non culturable states (VBNC) during inactivation of E. coli and Klebsiella pneumoniae by addition of H2O2 to natural well water under simulated solar irradiation. Water Res. 188:116499. doi: 10.1016/j.watres.2020.116499
Aly, A. H., Debbab, A., Kjer, J., and Proksch, P. (2010). Fungal endophytes from higher plants: a prolific source of phytochemicals and other bioactive natural products. Fungal Divers. 41, 1–16.
Andersen, M. R., Nielsen, J. B., Klitgaard, A., Petersen, L. M., Zachariasen, M., Hansen, T. J., et al. (2013). Accurate prediction of secondary metabolite gene clusters in filamentous fungi. Proc. Natl. Acad. Sci. U.S.A. 110, E99–E107. doi: 10.1073/pnas.1205532110
Andersson, J. O. (2009). Gene transfer and diversification of microbial eukaryotes. Annu. Rev. Microbiol. 63, 177–193. doi: 10.1146/annurev.micro.091208.073203
Arnold, A. E., and Lutzoni, F. (2007). Diversity and host range of foliar fungal endophytes: are tropical leaves biodiversity hotspots? Ecology 88, 541–549.
Arnold, A. E., Miadlikowska, J., Higgins, K. L., Sarvate, S. D., Gugger, P., Way, A., et al. (2009). A phylogenetic estimation of trophic transition networks for ascomycetous fungi: are lichens cradles of symbiotrophic fungal diversification? Syst. Biol. 58, 283–297. doi: 10.1093/sysbio/syp001
Bahram, M., Hildebrand, F., Forslund, S. K., Anderson, J. L., Soudzilovskaia, N. A., Bodegom, P. M., et al. (2018). Structure and function of the global topsoil microbiome. Nature 560, 233–237. doi: 10.1038/s41586-018-0386-6
Bailey, B. A., Bae, H., Strem, M. D., Roberts, D. P., Thomas, S. E., Crozier, J., et al. (2006). Fungal and plant gene expression during the colonization of cacao seedlings by endophytic isolates of four Trichoderma species. Planta 224, 1449–1464. doi: 10.1007/s00425-006-0314-0
Bajaj, R., Huang, Y., Gebrechristos, S., Mikolajczyk, B., Brown, H., Prasad, R., et al. (2018). Transcriptional responses of soybean roots to colonization with the root endophytic fungus Piriformospora indica reveals altered phenylpropanoid and secondary metabolism. Sci. Rep. 8:10227. doi: 10.1038/s41598-018-26809-3
Bapteste, E., Lopez, P., Bouchard, F., Baquero, F., McInerney, J. O., and Burian, R. M. (2012). Evolutionary analyses of non-genealogical bonds produced by introgressive descent. Proc. Natl. Acad. Sci. U.S.A. 109, 18266–18272. doi: 10.1073/pnas.1206541109
Bärenstrauch, M., Mann, S., Jacquemin, C., Bibi, S., Sylla, O. K., Baudouin, E., et al. (2020). Molecular crosstalk between the endophyte Paraconiothyrium variabile and the phytopathogen Fusarium oxysporum – modulation of lipoxygenase activity and beauvericin production during the interaction. Fungal Genet. Biol. 139:103383. doi: 10.1016/j.fgb.2020.103383
Bayram, Ö, and Braus, G. H. (2012). Coordination of secondary metabolism and development in fungi: the velvet family of regulatory proteins. FEMS Microbiol. Rev. 36, 1–24. doi: 10.1111/j.1574-6976.2011.00285.x
Bayram, O., Krappmann, S., Ni, M., Bok, J. W., Helmstaedt, K., Valerius, O., et al. (2008). VelB/VeA/LaeA complex coordinates light signal with fungal development and secondary metabolism. Science 320, 1504–1506. doi: 10.1126/science.1155888
Bergmann, S., Schumann, J., Scherlach, K., Lange, C., Brakhage, A. A., and Hertweck, C. (2007). Genomics-driven discovery of PKS-NRPS hybrid metabolites from Aspergillus nidulans. Nat. Chem. Biol. 3, 213–217. doi: 10.1038/nchembio869
Bevan, M., Bancroft, I., Bent, E., Love, K., Goodman, H., Dean, C., et al. (1998). Analysis of 1.9 Mb of contiguous sequence from chromosome 4 of Arabidopsis thaliana. Nature 391, 485–488. doi: 10.1038/35140
Bladt, T., Frisvad, J., Knudsen, P., and Larsen, T. (2013). Anticancer and antifungal compounds from Aspergillus, Penicillium and other filamentous fungi. Molecules 18, 11338–11376.
Bok, J. W., and Keller, N. P. (2004). LaeA, a regulator of secondary metabolism in Aspergillus spp. Eukaryot. Cell 3, 527–535. doi: 10.1128/ec.3.2.527-535.2004
Brady, S. F., and Clardy, J. (2000). CR377, a new pentaketide antifungal agent isolated from an endophytic fungus. J. Nat. Prod. 63, 1447–1448.
Braga, R. M., Padilla, G., and Araújo, W. L. (2018). The biotechnological potential of Epicoccum spp.: diversity of secondary metabolites. Crit. Rev. Microbiol. 44, 759–778. doi: 10.1080/1040841x.2018.1514364
Brakhage, A. A., Andrianopoulos, A., Kato, M., Steidl, S., Davis, M. A., Tsukagoshi, N., et al. (1999). HAP-Like CCAAT-binding complexes in filamentous fungi: implications for biotechnology. Fungal Genet. Biol. 27, 243–252. doi: 10.1006/fgbi.1999.1136
Brown, K. B., Hyde, K. D., and Guets, D. I. (1998). Preliminary studies on endophytic fungal communities of Musa acuminata species comples in Hong Kong and Australia. Fungal Divers. 1, 27–51.
Busby, P. E., Soman, C., Wagner, M. R., Friesen, M. L., Kremer, J., Bennett, A., et al. (2017). Research priorities for harnessing plant microbiomes in sustainable agriculture. PLoS Biol. 15:e2001793. doi: 10.1371/journal.pbio.2001793
Bush, L. P., Wilkinson, H. H., and Schardl, C. L. (1997). Bioprotective alkaloids of grass-fungal endophyte symbioses. Plant Physiol. 114, 1–7.
Calhoun, L. A., Findlay, J. A., Miller, J. D., and Whitney, N. J. (1992). Metabolites toxic to spruce budworm from balsam fir needle endophytes. Mycol. Res. 96, 281–286.
Calvo, A. M., Wilson, R. A., Woo, B. J., and Keller, N. P. (2002). Relationship between secondary metabolism and fungal development. Microbiol. Mol. Biol. Rev. 66, 447–459.
Camehl, I., Sherameti, I., Venus, Y., Bethke, G., Varma, A., Lee, J., et al. (2010). Ethylene signalling and ethylene-targeted transcription factors are required to balance beneficial and nonbeneficial traits in the symbiosis between the endophytic fungus Piriformospora indica and Arabidopsis thaliana. New Phytol. 185, 1062–1073. doi: 10.1111/j.1469-8137.2009.03149.x
Cánovas, D., Studt, L., Marcos, A. T., and Strauss, J. (2017). High-throughput format for the phenotyping of fungi on solid substrates. Sci. Rep. 7:4289. doi: 10.1038/s41598-017-03598-9
Carroll, G. (1988). Fungal endophytes in stems and leaves: from latent pathogen to mutualistic symbiont. Ecology 69, 2–9.
Chadha, N., Mishra, M., Prasad, R., and Varma, A. (2014). Root endophytic fungi: research update. J. Biol. Life Sci. 5, 135–158. doi: 10.5296/jbls.v5i2.5960
Chandra, S. (2012). Endophytic fungi: novel sources of anticancer lead molecules. Appl. Microbiol. Biotechnol. 95, 47–59. doi: 10.1007/s00253-012-4128-7
Chang, P. K., Ehrlich, K. C., Yu, J., Bhatnagar, D., and Cleveland, T. E. (1995). Increased expression of Aspergillus parasiticus aflR, encoding a sequence-specific DNA-binding protein, relieves nitrate inhibition of aflatoxin biosynthesis. Appl. Environ. Microbiol. 61, 2372–2377.
Chang, P. K., Yu, J., Bhatnagar, D., and Cleveland, T. E. (2000). Characterization of the Aspergillus parasiticus major nitrogen regulatory gene, area. Biochim. Biophys. Acta 1491, 263–266. doi: 10.1016/s0167-4781(00)00004-x
Chapela, I., Petrini, O., and Hagmann, L. (1991). Monolignol glucosides as specific recognition messengers in fungus-plant symbioses. Physiol. Mol. Plant Pathol. 39, 289–298.
Chen, H.-H., Chen, Y.-T., Huang, Y.-W., Tsai, H.-J., and Kuo, C.-C. (2012). 4-Ketopinoresinol, a novel naturally occurring ARE activator, induces the Nrf2/HO-1 axis and protects against oxidative stress-induced cell injury via activation of PI3K/AKT signaling. Free Radic. Biol. Med. 52, 1054–1066.
Chen, J., Chen, Q., Yu, F., Huang, H., Li, P., Zhu, J., et al. (2016). Comprehensive characterization and quantification of phillyrin in the fruits of Forsythia suspensa and its medicinal preparations by liquid chromatography-ion trap mass spectrometry. Acta Chromatogr. 28, 145–157.
Cheng, M.-J., Wu, M.-D., Chen, J.-J., Hsieh, S.-Y., Yuan, G.-F., Chen, I.-S., et al. (2013). Secondary metabolites from the endophytic fungus of Annulohypoxylon ilanense. Chem. Nat. Compd. 49, 523–525.
Chiang, Y. M., Ahuja, M., Oakley, C. E., Entwistle, R., Asokan, A., Zutz, C., et al. (2016). Development of genetic dereplication strains in Aspergillus nidulans results in the discovery of aspercryptin. Angew. Chem. 55, 1662–1665. doi: 10.1002/anie.201507097
Cho, J. Y., Nam, K. H., Kim, A. R., Park, J., Yoo, E. S., Baik, K. U., et al. (2001). In-vitro and in-vivo immunomodulatory effects of syringin. J. Pharm. Pharmacol. 53, 1287–1294.
Chu, H. Y., Wegel, E., and Osbourn, A. (2011). From hormones to secondary metabolism: the emergence of metabolic gene clusters in plants. Plant J. 66, 66–79. doi: 10.1111/j.1365-313X.2011.04503.x
Combes, A., Ndoye, I., Bance, C., Bruzaud, J., Djediat, C., Dupont, J., et al. (2012). Chemical communication between the endophytic fungus Paraconiothyrium variabile and the phytopathogen Fusarium oxysporum. PLoS One 7:e47313. doi: 10.1371/journal.pone.0047313
Cramer, R. A. Jr., Gamcsik, M. P., Brooking, R. M., Najvar, L. K., Kirkpatrick, W. R., Patterson, T. F., et al. (2006). Disruption of a nonribosomal peptide synthetase in Aspergillus fumigatus eliminates gliotoxin production. Eukaryot. Cell 5, 972–980. doi: 10.1128/EC.00049-06
Cui, J. L., Wang, Y. N., Jiao, J., Gong, Y., Wang, J. H., and Wang, M. L. (2017). Fungal endophyte-induced salidroside and tyrosol biosynthesis combined with signal cross-talk and the mechanism of enzyme gene expression in Rhodiola crenulata. Sci. Rep. 7:12540. doi: 10.1038/s41598-017-12895-2
Cui, R., Lu, X., Chen, X., Malik, W. A., Wang, D., Wang, J., et al. (2021). A novel raffinose biological pathway is observed by symbionts of cotton=Verticillium dahliae to improve salt tolerance genetically on cotton. J. Agron. Crop Sci. 207, 956–969. doi: 10.1111/jac.12556
Daubresse, N., Chupeau, Y., Francesch, C., Lapierre, C., Pollet, B., and Rolando, C. (1997). Rational design, synthesis and biological evaluation of the first inhibitor of lignin polymerization. Chem. Commun. 1489–1490.
Davey, M. L., and Currah, R. S. (2006). Interactions between mosses (Bryophyta) and fungi. Can. J. Bot. 84, 1509–1519.
Debbab, A., Aly, A. H., and Proksch, P. (2011). Bioactive secondary metabolites from endophytes and associated marine derived fungi. Fungal Divers. 49, 1–12. doi: 10.1007/s13225-011-0114-0
Deepika, V. B., Murali, T. S., and Satyamoorthy, K. (2016). Modulation of genetic clusters for synthesis of bioactive molecules in fungal endophytes: a review. Microbiol. Res. 182, 125–140. doi: 10.1016/j.micres.2015.10.009
Demain, A. L. (2014). Importance of microbial natural products and the need to revitalize their discovery. J. Ind. Microbiol. Biotechnol. 41, 185–201.
Di, X., Takken, F. L., and Tintor, N. (2016). How phytohormones shape interactions between plants and the soil-borne fungus Fusarium oxysporum. Front. Plant Sci. 7:170. doi: 10.3389/fpls.2016.00170
Ding, T., Suo, Y., Xiang, Q., Zhao, X., Chen, S., Ye, X., et al. (2017). Significance of viable but nonculturable Escherichia coli: induction, detection, and control. J. Microbiol. Biotechnol. 27, 417–428. doi: 10.4014/jmb.1609.09063
Dixon, R. A., and Paiva, N. L. (1995). Stress-induced phenylpropanoid metabolism. Plant Cell 7, 1085–1097.
Dreyer, J., Eichhorn, H., Friedlin, E., Kurnsteiner, H., and Kuck, U. (2007). A homologue of the Aspergillus velvet gene regulates both cephalosporin C biosynthesis and hyphal fragmentation in Acremonium chrysogenum. Appl. Environ. Microbiol. 73, 3412–3422. doi: 10.1128/AEM.00129-07
Duringer, J., DeLorme, M., Lehner, A., and Craig, A. (2007). “A review of the ergot alkaloids found in endophyte-infected tall fescue and perennial ryegrass and their metabolism after ingestion by livestock,” in Proceedings of the 6th International Symposium on Fungal Endophytes of Grasses, (Christchurch), 377–382.
El Enshasy, H. A., Hanapi, S. Z., Malek, R. A., Abdelgalil, S. A., and Leng, O. M. (2019). “Endophytic fungi: the desired biostimulants for essential oil production,” in Advances in Endophytic Fungal Research: Present Status and Future Challenges, ed. B. P. Singh (Cham: Springer International Publishing), 211–232.
Falshaw, C. P., Ollis, W. D., Ormand, K. L., Mongkolsuk, S., and Podimuang, V. (1969). The spectroscopic identification of coniferin. Phytochemistry 8, 913–915. doi: 10.1016/s0031-9422(00)85883-5
Fang, C., Fernie, A. R., and Luo, J. (2019). Exploring the diversity of plant metabolism. Trends Plant Sci. 24, 83–98. doi: 10.1016/j.tplants.2018.09.006
Fernandes, M., Keller, N. P., and Adams, T. H. (1998). Sequence-specific binding by Aspergillus nidulans AflR, a C6 zinc cluster protein regulating mycotoxin biosynthesis. Mol. Microbiol. 28, 1355–1365.
Feurtey, A., and Stukenbrock, E. H. (2018). Interspecific gene exchange as a driver of adaptive evolution in fungi. Annu. Rev. Microbiol. 72, 377–398. doi: 10.1146/annurev-micro-090817-062753
Field, B., and Osbourn, A. E. (2008). Metabolic diversification–independent assembly of operon-like gene clusters in different plants. Science 320, 543–547. doi: 10.1126/science.1154990
Field, B., Fiston-Lavier, A. S., Kemen, A., Geisler, K., Quesneville, H., and Osbourn, A. E. (2011). Formation of plant metabolic gene clusters within dynamic chromosomal regions. Proc. Natl. Acad. Sci. U.S.A. 108, 16116–16121. doi: 10.1073/pnas.1109273108
Figueiredo, A. C., Barroso, J. G., Pedro, L. G., and Scheffer, J. J. C. (2008). Factors affecting secondary metabolite production in plants: volatile components and essential oils. Flavour Fragr. J. 23, 213–226. doi: 10.1002/ffj.1875
Flieger, M., Wurst, M., and Shelby, R. (1997). Ergot alkaloids—sources, structures and analytical methods. Folia Microbiol. 42, 3–30.
Forni, C., Duca, D., and Glick, B. R. (2017). Mechanisms of plant response to salt and drought stress and their alteration by rhizobacteria. Plant Soil 410, 335–356.
Foss, H. M., Roberts, C. J., Claeys, K. M., and Selker, E. U. (1993). Abnormal chromosome behavior in Neurospora mutants defective in DNA methylation. Science 262, 1737–1741. doi: 10.1126/science.7505062
Fox, E. M., and Howlett, B. J. (2008). Secondary metabolism: regulation and role in fungal biology. Curr. Opin. Microbiol. 11, 481–487. doi: 10.1016/j.mib.2008.10.007
Friesen, M. L., Porter, S. S., Stark, S. C., von Wettberg, E. J., Sachs, J. L., and Martinez-Romero, E. (2011). Microbially mediated plant functional traits. Annu. Rev. Ecol. Evol. Syst. 42, 23–46. doi: 10.1146/annurev-ecolsys-102710-145039
Gao, L. D., Huang, G. R., and Wang, Y. (2008). Seasonal and tissue age influences on endophytic fungi of Pinus tabulaeformis (Pinaceae) in the Dongling Mountains, Beijing. J. Integr. Plant Biol. 50, 997–1003. doi: 10.1111/j.1744-7909.2008.00394x
Geng, S., Kong, X., Song, G., Jia, M., Guan, J., Wang, F., et al. (2019). DNA methylation dynamics during the interaction of wheat progenitor Aegilops tauschii with the obligate biotrophic fungus Blumeria graminis f. sp. tritici. New Phytol. 221, 1023–1035. doi: 10.1111/nph.15432
George, T. S., Guru, K. S. S., Vasanthi, N. S., and Kannan, K. P. (2011). Extraction, purification and characterization of chitosan from endophytic fungi isolated from medicinal plants. World J. Sci. Technol. 1, 43–48.
Gorzynska, K., Slachetka, M., Ryszka, P., Turnau, K., Plachno, B. J., and Lembicz, M. (2018). Incidence, identification, and mycoparasitic activity of clonostachys Epichloë, a hyperparasite of the fungal endophyte Epichloë typhina. Plant Dis. 102, 1973–1980. doi: 10.1094/PDIS-02-18-0320-RE
Gouda, S., Das, G., Sen, S. K., Shin, H. S., and Patra, J. K. (2016). Endophytes: a treasure house of bioactive compounds of medicinal importance. Front. Microbiol. 7:1538. doi: 10.3389/fmicb.2016.01538
Grafton, K. F., Poehlman, J. M., Sehgal, O. P., and Sechler, D. T. (1982). Tall fescue as a natural host and aphid vectors of barley yellow dwarf virus in Missouri. Plant Dis. 66, 318–320.
Guerre, P. (2015). Ergot alkaloids produced by endophytic fungi of the genus Epichloë. Toxins 7, 773–790. doi: 10.3390/toxins7030773
Gundel, P. E., Seal, C. E., Biganzoli, F., Martínez-Ghersa, M. A., Molina-Montenegro, M. A., Vázquez-de-Aldana, B. R., et al. (2018). Occurrence of alkaloids in grass seeds symbiotic with vertically-transmitted Epichloë fungal endophytes and its relationship with antioxidants. Front. Ecol. Evol. 6:211. doi: 10.3389/fevo.2018.00211
Haas, D., and Défago, G. (2005). Biological control of soil-borne pathogens by fluorescent pseudomonads. Nat. Rev. Microbiol. 3, 307–319. doi: 10.1038/nrmicro1129
Hardoim, P. R., van Overbeek, L. S., and van Elsas, J. D. (2008). Properties of bacterial endophytes and their proposed role in plant growth. Trends Microbiol. 16, 463–471. doi: 10.1016/j.tim.2008.07.008
Hardoim, P. R., van Overbeek, L. S., Berg, G., Pirttila, A. M., Compant, S., Campisano, A., et al. (2015). The hidden world within plants: ecological and evolutionary considerations for defining functioning of microbial endophytes. Microbiol. Mol. Biol. Rev. 79, 293–320. doi: 10.1128/MMBR.00050-14
Henke, M. T., Soukup, A. A., Goering, A. W., McClure, R. A., Thomson, R. J., Keller, N. P., et al. (2016). New aspercryptins, lipopeptide natural products, revealed by HDAC inhibition in Aspergillus nidulans. ACS Chem. Biol. 11, 2117–2123. doi: 10.1021/acschembio.6b00398
Hicks, J. K., Yu, J. H., Keller, N. P., and Adams, T. H. (1997). Aspergillus sporulation and mycotoxin production both require inactivation of the FadA G alpha protein-dependent signaling pathway. EMBO J. 16, 4916–4923. doi: 10.1093/emboj/16.16.4916
Hiroyuki, K., Satoshi, T., Shun-ichi, T., Yoshihara, T., Sakamura, S., Shimanuki, T., et al. (1989). New fungitoxic sesquiterpenoids, chokols AG, from stromata of Epichloë typhina and the absolute configuration of chokol E. Agric. Biol. Chem. 53, 789–796.
Hiruma, K., Gerlach, N., Sacristan, S., Nakano, R. T., Hacquard, S., Kracher, B., et al. (2016). Root endophyte Colletotrichum tofieldiae confers plant fitness benefits that are phosphate status dependent. Cell 165, 464–474. doi: 10.1016/j.cell.2016.02.028
Hoffman, M. T., and Arnold, A. E. (2010). Diverse bacteria inhabit living hyphae of phylogenetically diverse fungal endophytes. Appl. Environ. Microbiol. 76, 4063–4075. doi: 10.1128/AEM.02928-09
Hoffman, M. T., Gunatilaka, M. K., Wijeratne, K., Gunatilaka, L., and Arnold, A. E. (2013). Endohyphal bacterium enhances production of indole-3-acetic acid by a foliar fungal endophyte. PLoS One 8:e73132. doi: 10.1371/journal.pone.0073132
Hoffmeister, D., and Keller, N. P. (2007). Natural products of filamentous fungi: enzymes, genes, and their regulation. Nat. Prod. Rep. 24, 393–416. doi: 10.1039/b603084j
Horn, W. S., Simmonds, M. S., Schwartz, R. E., and Blaney, W. M. (1996). Variation in production of phomodiol and phomopsolide B by Phomopsis spp. Mycologia 88, 588–595.
Howitz, K. T., and Sinclair, D. A. (2008). Xenohormesis: sensing the chemical cues of other species. Cell 133, 387–391. doi: 10.1016/j.cell.2008.04.019
Huang, A. C., Jiang, T., Liu, Y. X., Bai, Y. C., Reed, J., Qu, B., et al. (2019). A specialized metabolic network selectively modulates Arabidopsis root microbiota. Science 364:eaau6389. doi: 10.1126/science.aau6389
Huang, S., Chen, H., Li, W., Zhu, X., Ding, W., and Li, C. (2016). Bioactive chaetoglobosins from the mangrove endophytic fungus Penicillium chrysogenum. Mar. Drugs 14:172.
Huang, W., Wan, C., and Zhou, S. (2013). Quercetin-A flavonoid compound from Sarcopyramis bodinieri var delicate with potential apoptotic activity in HepG2 liver cancer cells. Trop. J. Pharm. Res. 12, 529–533.
Hunt, M. G., and Newman, J. A. (2005). Reduced herbivore resistance from a novel grass-endophyte association. J. Appl. Ecol. 42, 762–769. doi: 10.1111/j.1365-2664.2005.01061.x
Hurst, L. D., Pal, C., and Lercher, M. J. (2004). The evolutionary dynamics of eukaryotic gene order. Nat. Rev. Genet. 5, 299–310. doi: 10.1038/nrg1319
Hussain, H., Akhtar, N., Draeger, S., Schulz, B., Pescitelli, G., Salvadori, P., et al. (2009). New bioactive 2,3-epoxycyclohexenes and isocoumarins from the endophytic fungus Phomopsis sp. from Laurus azorica. Eur. J. Organ. Chem. 2009, 749–756. doi: 10.1002/ejoc.200801052
Imazaki, I., and Kadota, I. (2015). Molecular phylogeny and diversity of Fusarium endophytes isolated from tomato stems. FEMS Microbiol. Ecol. 91:fiv098. doi: 10.1093/femsec/fiv098
Jakubczyk, D., and Dussart, F. (2020). Selected fungal natural products with antimicrobial properties. Molecules 25:911. doi: 10.3390/molecules25040911
Ji, H. F., Li, X. J., and Zhang, H. Y. (2009). Natural products and drug discovery. Can thousands of years of ancient medical knowledge lead us to new and powerful drug combinations in the fight against cancer and dementia? EMBO Rep. 10, 194–200. doi: 10.1038/embor.2009.12
Jia, M., Chen, L., Xin, H. L., Zheng, C. J., Rahman, K., Han, T., et al. (2016). A friendly relationship between endophytic fungi and medicinal plants: a systematic review. Front. Microbiol. 7:906. doi: 10.3389/fmicb.2016.00906
Jiang, X., Zerfass, C., Feng, S., Eichmann, R., Asally, M., Schafer, P., et al. (2018). Impact of spatial organization on a novel auxotrophic interaction among soil microbes. ISME J. 12, 1443–1456. doi: 10.1038/s41396-018-0095-z
Ju, Y., Sacalis, J. N., and Still, C. C. (1998). Bioactive flavonoids from endophyte-infected blue grass (Poa ampla). J. Agric. Food Chem. 46, 3785–3788.
Kato, N., Brooks, W., and Calvo, A. M. (2003). The expression of sterigmatocystin and penicillin genes in Aspergillus nidulans is controlled by veA, a gene required for sexual development. Eukaryot. Cell 2, 1178–1186. doi: 10.1128/ec.2.6.1178-1186.2003
Kaul, S., Wani, M., Dhar, K. L., and Dhar, M. K. (2008). Production and GC-MS trace analysis of methyl eugenol from endophytic isolate of Alternaria from rose. Ann. Microbiol. 58:443.
Keglevich, P., Hazai, L., Kalaus, G., and Szántay, C. (2012). Modifications on the basic skeletons of vinblastine and vincristine. Molecules 17, 5893–5914.
Keller, N. P., Turner, G., and Bennett, J. W. (2005). Fungal secondary metabolism – from biochemistry to genomics. Nat. Rev. Microbiol. 3, 937–947. doi: 10.1038/nrmicro1286
Kemen, A. C., Agler, M. T., and Kemen, E. (2015). Host-microbe and microbe-microbe interactions in the evolution of obligate plant parasitism. New Phytol. 206, 1207–1228. doi: 10.1111/nph.13284
Khan, A. L., Hamayun, M., Hussain, J., Kang, S.-M., and Lee, I.-J. (2012). The newly isolated endophytic fungus Paraconiothyrium sp. LK1 produces ascotoxin. Molecules 17, 1103–1112.
Khan, A., Bassett, S., Voisey, C., Gaborit, C., Johnson, L., Christensen, M., et al. (2010). Gene expression profiling of the endophytic fungus Neotyphodium lolii in association with its host plant perennial ryegrass. Aust. Plant Pathol. 39, 467–476.
Kim, H., and Woloshuk, C. P. (2008). Role of AREA, a regulator of nitrogen metabolism, during colonization of maize kernels and fumonisin biosynthesis in Fusarium verticillioides. Fungal Genet. Biol. 45, 947–953. doi: 10.1016/j.fgb.2008.03.007
Kim, J., Cho, S. Y., Kim, S. H., Kim, S., Park, C.-W., Cho, D., et al. (2016). Ginseng berry and its biological effects as a natural phytochemical. Nat. Prod. Chem. Res. 4:2.
Kirby, J., and Keasling, J. D. (2009). Biosynthesis of plant isoprenoids: perspectives for microbial engineering. Annu. Rev. Plant Biol. 60, 335–355. doi: 10.1146/annurev.arplant.043008.091955
Kogel, K. H., Franken, P., and Heückelhoven, R. (2006). Endophyte or parasite—–what decides? Curr. Opin. Plant Biol. 9, 358–363. doi: 10.1016/j.pbi.2006.05.001
Konarzewska, Z., Śliwińska-Wilczewska, S., Felpeto, A. B., Vasconcelos, V., and Latala, A. (2020). Assessment of the allelochemical activity and biochemical profile of different phenotypes of picocyanobacteria from the genus Synechococcus. Mar. Drugs 18:179. doi: 10.3390/md18040179
Kosalkova, K., Garcia-Estrada, C., Ullan, R. V., Godio, R. P., Feltrer, R., Teijeira, F., et al. (2009). The global regulator LaeA controls penicillin biosynthesis, pigmentation and sporulation, but not roquefortine C synthesis in Penicillium chrysogenum. Biochimie 91, 214–225. doi: 10.1016/j.biochi.2008.09.004
Kouzminova, E., and Selker, E. U. (2001). dim-2 encodes a DNA methyltransferase responsible for all known cytosine methylation in Neurospora. EMBO J. 20, 4309–4323. doi: 10.1093/emboj/20.15.4309
Kozyrovska, N. O. (2013). Crosstalk between endophytes and a plant host within information-processing networks. Biopolym. Cell 29, 234–243. doi: 10.7124/bc.00081D
Kumar, A., Patil, D., Rajamohanan, P. R., and Ahmad, A. (2013). Isolation, purification and characterization of vinblastine and vincristine from endophytic fungus Fusarium oxysporum isolated from Catharanthus roseus. PLoS One 8:e71805. doi: 10.1371/journal.pone.0071805
Kumar, S., and Kaushik, N. (2013). Endophytic fungi isolated from oil-seed crop Jatropha curcas produces oil and exhibit antifungal activity. PLoS One 8:e56202. doi: 10.1371/journal.pone.0056202
Kunkel, B. N., and Brooks, D. M. (2002). Cross talk between signaling pathways in pathogen defense. Curr. Opin. Plant Biol. 5, 325–331.
Kusari, S., Hertweck, C., and Spiteller, M. (2012). Chemical ecology of endophytic fungi: origins of secondary metabolites. Chem. Biol. 19, 792–798. doi: 10.1016/j.chembiol.2012.06.004
Lee, C.-C., Liu, K.-J., Wu, Y.-C., Lin, S.-J., Chang, C.-C., and Huang, T.-S. (2011). Sesamin inhibits macrophage-induced vascular endothelial growth factor and matrix metalloproteinase-9 expression and proangiogenic activity in breast cancer cells. Inflammation 34, 209–221.
Lee, D. Y., Teyssier, C., Strahl, B. D., and Stallcup, M. R. (2005). Role of protein methylation in regulation of transcription. Endocr. Rev. 26, 147–170. doi: 10.1210/er.2004-0008
Li, E., Jiang, L., Guo, L., Zhang, H., and Che, Y. (2008). Pestalachlorides A–C, antifungal metabolites from the plant endophytic fungus Pestalotiopsis adusta. Bioorg. Med. Chem. 16, 7894–7899.
Li, F., Zhang, N., Wu, Q., Yuan, Y., Yang, Z., Zhou, M., et al. (2017). Syringin prevents cardiac hypertrophy induced by pressure overload through the attenuation of autophagy. Int. J. Mol. Med. 39, 199–207.
Li, S., Li, Z., Qian, L., Xiao-Wen, L., Ju-Qun, X., Gui-Mei, K., et al. (2015). Fumigaclavine I, a new alkaloid isolated from endophyte Aspergillus terreus. Chin. J. Nat. Med. 13, 937–941.
Liang, L., Gao, C., Luo, M., Zhao, C., Wang, W., Gu, C., et al. (2013). The phytoestrogenic compound Cajanol from Pigeonpea roots is associated with the activation of estrogen receptor α−dependent signaling pathway in human prostate cancer cells. Phytother. Res. 27, 1834–1841.
Liao, H. L., Bonito, G., Rojas, J. A., Hameed, K., Wu, S., Schadt, C. W., et al. (2019). Fungal endophytes of Populus trichocarpa alter host phenotype, gene expression, and rhizobiome composition. Mol. Plant Microbe Interact. 32, 853–864.
Lin, C., Yang, L., Xu, G., and Wu, J. (2011). Biodegradation and metabolic pathway of β-chlorinated aliphatic acid in Bacillus sp. CGMCC no. 4196. Appl. Microbiol. Biotechnol. 90, 689–696. doi: 10.1007/s00253-010-3081-6
Lin, R., Kim, H., Hong, J., and Li, Q.-J. (2014). Biological evaluation of subglutinol A as a novel immunosuppressive agent for inflammation intervention. ACS Med. Chem. Lett. 5, 485–490.
Lind, A. L., Wisecaver, J. H., Lameiras, C., Wiemann, P., Palmer, J. M., Keller, N. P., et al. (2017). Drivers of genetic diversity in secondary metabolic gene clusters within a fungal species. PLoS Biol. 15:e2003583. doi: 10.1371/journal.pbio.2003583
Liu, H. X., Liu, W. Z., Chen, Y. C., Sun, Z. H., Tan, Y. Z., Li, H. H., et al. (2016). Cytotoxic trichothecene macrolides from the endophyte fungus Myrothecium roridum. J. Asian Nat. Prod. Res. 18, 684–689. doi: 10.1080/10286020.2015.1134505
Liu, X., Dong, M., Chen, X., Jiang, M., Lv, X., and Zhou, J. (2008). Antimicrobial activity of an endophytic Xylaria sp.YX-28 and identification of its antimicrobial compound 7-amino-4-methylcoumarin. Appl. Microbiol. Biotechnol. 78, 241–247. doi: 10.1007/s00253-007-1305-1
Lu, H., Zou, W. X., Meng, J. C., Hu, J., and Tan, R. X. (2000). New bioactive metabolites produced by Colletotrichum sp., an endophytic fungus in Artemisia annua. Plant Sci. 151, 67–73.
Ma, L.-J., van der Does, H. C., Borkovich, K. A., Coleman, J. J., Daboussi, M.-J., Di Pietro, A., et al. (2010). Comparative genomics reveals mobile pathogenicity chromosomes in Fusarium. Nature 464, 367–373. doi: 10.1038/nature08850
Macías-Rubalcava, M. L., Hernández-Bautista, B. E., Jiménez-Estrada, M., González, M. C., Glenn, A. E., Hanlin, R. T., et al. (2008). Naphthoquinone spiroketal with allelochemical activity from the newly discovered endophytic fungus Edenia gomezpompae. Phytochemistry 69, 1185–1196.
Macías-Rubalcava, M. L., Sobrino, M. E. R.-V., Meléndez-González, C., and Hernández-Ortega, S. (2014). Naphthoquinone spiroketals and organic extracts from the endophytic fungus Edenia gomezpompae as potential herbicides. J. Agric. Food Chem. 62, 3553–3562.
Malmstrom, C. M., McCullough, A. J., Johnson, H. A., Newton, L. A., and Borer, E. T. (2005). Invasive annual grasses indirectly increase virus incidence in California native perennial bunchgrasses. Oecologia 145, 153–164. doi: 10.1007/s00442-005-0099-z
Mandal, S. M., Chakraborty, D., and Dey, S. (2010). Phenolic acids act as signaling molecules in plant-microbe symbioses. Plant Signal. Behav. 5, 359–368. doi: 10.4161/psb.5.4.10871
Martínez-Luis, S., Della-Togna, G., Coley, P. D., Kursar, T. A., Gerwick, W. H., and Cubilla-Rios, L. (2008). Antileishmanial constituents of the Panamanian endophytic fungus Edenia sp. J. Nat. Prod. 71, 2011–2014.
Materska, M. (2008). Quercetin and its derivatives: chemical structure and bioactivity-a review. Pol. J. Food Nutr. Sci. 58, 407–413.
Mehta, P., Sharma, R., Putatunda, C., and Walia, A. (2019). “Endophytic fungi: role in phosphate solubilization,” in Advances in Endophytic Fungal Research: Present Status and Future Challenges, ed. B. P. Singh (Cham: Springer International Publishing), 183–209.
Mejia, L. C., Herre, E. A., Sparks, J. P., Winter, K., Garcia, M. N., Van Bael, S. A., et al. (2014). Pervasive effects of a dominant foliar endophytic fungus on host genetic and phenotypic expression in a tropical tree. Front. Microbiol. 5:479. doi: 10.3389/fmicb.2014.00479
Mendes, M. A., Guerra, R. F., Berns, M. C., Manzo, C., Masiero, S., Finzi, L., et al. (2013). MADS domain transcription factors mediate short-range DNA looping that is essential for target gene expression in Arabidopsis. Plant Cell 25, 2560–2572. doi: 10.1105/tpc.112.108688
Meyers, D. M., Obrian, G., Du, W. L., Bhatnagar, D., and Payne, G. A. (1998). Characterization of aflJ, a gene required for conversion of pathway intermediates to aflatoxin. Appl. Environ. Microbiol. 64, 3713–3717.
Mishra, Y., Sharma, L., Dhiman, M., and Sharma, M. M. (2021). “Endophytic fungal diversity of selected medicinal plants and their bio-potential applications,” in Fungi Bio-Prospects in Sustainable Agriculture, Environment and Nano-Technology, Chap. 10, eds V. K. Sharma, M. P. Shah, S. Parmar, and A. Kumar (Cambridge, MA: Academic Press), 227–283.
Mohamed, R., Jong, P. L., and Zali, M. S. (2010). Fungal diversity in wounded stems of Aquilaria malaccensis. Fungal Divers. 43, 67–74.
Mousa, W. K., and Raizada, M. N. (2013). The diversity of anti-microbial secondary metabolites produced by fungal endophytes: an interdisciplinary perspective. Front. Microbiol. 4:65.
Muller, C. B., and Krauss, J. (2005). Symbiosis between grasses and asexual fungal endophytes. Curr. Opin. Plant Biol. 8, 450–456. doi: 10.1016/j.pbi.2005.05.007
Naveen, B., Ommi, N. B., Mudiraj, A., Mallikarjuna, T., Babu, P. P., and Nagarajan, R. (2017). Total synthesis of penicinoline E, marinamide, methyl marinamide and their antimalarial activity. ChemistrySelect 2, 3256–3261.
Ng, T. B. (2004). Peptides and proteins from fungi. Peptides 25, 1055–1073. doi: 10.1016/j.peptides.2004.03.013
Nicolaou, K., Yang, Z., Liu, J., Ueno, H., Nantermet, P., Guy, R., et al. (1994). Total synthesis of taxol. Nature 367:630.
Nicoletti, R., and Fiorentino, A. (2015). Plant bioactive metabolites and drugs produced by endophytic fungi of Spermatophyta. Agriculture 5, 918–970.
Nowack, E. C. M., and Weber, A. P. M. (2018). Genomics-informed insights into endosymbiotic organelle evolution in photosynthetic eukaryotes. Annu. Rev. Plant Biol. 69, 51–84. doi: 10.1146/annurev-arplant-042817-040209
Nowak, R., Drozd, M., Mendyk, E., Lemieszek, M., Krakowiak, O., Kisiel, W., et al. (2016). A new method for the isolation of ergosterol and peroxyergosterol as active compounds of Hygrophoropsis aurantiaca and in vitro antiproliferative activity of isolated ergosterol peroxide. Molecules 21:946.
Nützmann, H. W., Scazzocchio, C., and Osbourn, A. (2018). Metabolic gene clusters in eukaryotes. Annu. Rev. Genet. 52, 159–183. doi: 10.1146/annurev-genet-120417-031237
Osbourn, A. (2010a). Gene clusters for secondary metabolic pathways: an emerging theme in plant biology. Plant Physiol. 154, 531–535. doi: 10.1104/pp.110.161315
Osbourn, A. (2010b). Secondary metabolic gene clusters: evolutionary toolkits for chemical innovation. Trends Genet. 26, 449–457. doi: 10.1016/j.tig.2010.07.001
Osbourn, A. E., and Field, B. (2009). Operons. Cell. Mol. Life Sci. 66, 3755–3775. doi: 10.1007/s00018-009-0114-3
Owen, E., and West, D. (1971). “Isolation and identification of 7, 8-dimethyl-10-(2’-hydroxyethyl) isoalloxazine from natural sources,” in Methods in Enzymology, eds D. B. McCormick and L. D. Wright (Amsterdam: Elsevier), 574–579.
Panche, A. N., Diwan, A. D., and Chandra, S. R. (2016). Flavonoids: an overview. J. Nutr. Sci. 5:e47. doi: 10.1017/jns.2016.41
Pedley, K. F., and Walton, J. D. (2001). Regulation of cyclic peptide biosynthesis in a plant pathogenic fungus by a novel transcription factor. Proc. Natl. Acad. Sci. U.S.A. 98, 14174–14179.
Peršoh, D. (2015). Plant-associated fungal communities in the light of meta’omics. Fungal Divers. 75, 1–25. doi: 10.1007/s13225-015-0334-9
Photita, W., Lumyong, S., Lumyong, P., McKenzie, E. H. C., and Hyde, K. D. (2004). Are some fungi isolated as endophytes of Musa acuminata latent pathogens? Fungal Divers. 16, 131–140.
Pichersky, E., and Gang, D. R. (2000). Genetics and biochemistry of secondary metabolites in plants: an evolutionary perspective. Trends Plant Sci. 5, 439–445.
Porras-Alfaro, A., and Bayman, P. (2011). Hidden fungi, emergent properties: endophytes and microbiomes. Annu. Rev. Phytopathol. 49, 291–315. doi: 10.1146/annurev-phyto-080508-081831
Poveda, J. (2021). Beneficial effects of microbial volatile organic compounds (MVOCs) in plants. Appl. Soil Ecol. 168:104118. doi: 10.1016/j.apsoil.2021.104118
Poveda, J., Abril-Urias, P., and Escobar, C. (2020a). Biological control of plant-parasitic nematodes by filamentous fungi inducers of resistance: Trichoderma, Mycorrhizal and endophytic fungi. Front. Microbiol. 11:992. doi: 10.3389/fmicb.2020.00992
Poveda, J., Zabalgogeazcoa, I., Soengas, P., Rodríguez, V. M., Cartea, M. E., Abilleira, R., et al. (2020b). Brassica oleracea var. acephala (kale) improvement by biological activity of root endophytic fungi. Sci. Rep. 10:20224. doi: 10.1038/s41598-020-77215-7
Poveda, J., and Baptista, P. (2021). Filamentous fungi as biocontrol agents in olive (Olea europaea L.) diseases: Mycorrhizal and endophytic fungi. Crop Prot. 146:105672. doi: 10.1016/j.cropro.2021.105672
Poveda, J., Eugui, D., Abril-Urías, P., and Velasco, P. (2021). Endophytic fungi as direct plant growth promoters for sustainable agricultural production. Symbiosis 85, 1–19. doi: 10.1007/s13199-021-00789-x
Prasch, C. M., and Sonnewald, U. (2013). Simultaneous application of heat, drought, and virus to Arabidopsis plants reveals significant shifts in signaling networks. Plant Physiol. 162, 1849–1866. doi: 10.1104/pp.113.221044
Proctor, R. H., Hohn, T. M., McCormick, S. P., and Desjardins, A. E. (1995). Tri6 encodes an unusual zinc finger protein involved in regulation of trichothecene biosynthesis in Fusarium sporotrichioides. Appl. Environ. Microbiol. 61, 1923–1930.
Qin, J.-C., Zhang, Y.-M., Gao, J.-M., Bai, M.-S., Yang, S.-X., Laatsch, H., et al. (2009). Bioactive metabolites produced by Chaetomium globosum, an endophytic fungus isolated from Ginkgo biloba. Bioorg. Med. Chem. Lett. 19, 1572–1574.
Quambusch, M., Pirttilä, A. M., Tejesvi, M. V., Winkelmann, T., and Bartsch, M. (2014). Endophytic bacteria in plant tissue culture: differences between easy-and difficult-to-propagate Prunus avium genotypes. Tree Physiol. 34, 524–533.
Radic, N., and Strukelj, B. (2012). Endophytic fungi: the treasure chest of antibacterial substances. Phytomedicine 19, 1270–1284. doi: 10.1016/j.phymed.2012.09.007
Richards, T. A., Leonard, G., Soanes, D. M., and Talbot, N. J. (2011). Gene transfer into the fungi. Fungal Biol. Rev. 25, 98–110. doi: 10.1016/j.fbr.2011.04.003
Roberts, E., and Lindow, S. (2014). Loline alkaloid production by fungal endophytes of Fescue species select for particular epiphytic bacterial microflora. ISME J. 8, 359–368. doi: 10.1038/ismej.2013.170
Rodriguez, R. J., Henson, J., Van Volkenburgh, E., Hoy, M., Wright, L., Beckwith, F., et al. (2008). Stress tolerance in plants via habitat-adapted symbiosis. ISME J. 2, 404–416.
Rodriguez, R., and Redman, R. (2008). More than 400 million years of evolution and some plants still can’t make it on their own: plant stress tolerance via fungal symbiosis. J. Exp. Bot. 59, 1109–1114. doi: 10.1093/jxb/erm342
Rodríguez-Morató, J., Xicota, L., Fitó, M., Farré, M., Dierssen, M., and de la Torre, R. (2015). Potential role of olive oil phenolic compounds in the prevention of neurodegenerative diseases. Molecules 20, 4655–4680.
Romero, A., Carrion, G., and Rico-Gray, V. (2001). Fungal latent pathogens and endophytes from leaves of Parthenium hysterophorus (Asteraceae). Fungal Divers. 7, 81–87.
Ruiz, B., Chavez, A., Forero, A., Garcia-Huante, Y., Romero, A., Sanchez, M., et al. (2010). Production of microbial secondary metabolites: regulation by the carbon source. Crit. Rev. Microbiol. 36, 146–167. doi: 10.3109/10408410903489576
Rukachaisirikul, V., Sommart, U., Phongpaichit, S., Sakayaroj, J., and Kirtikara, K. (2008). Metabolites from the endophytic fungus Phomopsis sp. PSU-D15. Phytochemistry 69, 783–787.
Saikkonen, K., Gundel, P. E., and Helander, M. (2013). Chemical ecology mediated by fungal endophytes in grasses. J. Chem. Ecol. 39, 962–968. doi: 10.1007/s10886-013-0310-3
Sampangi-Ramaiah, M. H., Dey, P., Jambagi, S., Kumari, M. V., Oelmüller, R., Nataraja, K. N., et al. (2020). An endophyte from salt-adapted Pokkali rice confers salt-tolerance to a salt-sensitive rice variety and targets a unique pattern of genes in its new host. Sci. Rep. 10:3237. doi: 10.1038/s41598-020-59998-x
Schardl, C. L., and Phillips, T. D. (1997). Protective grass endophytes: where are they from and where are they going? Plant Dis. 81, 430–438.
Schmitt, E. K., Hoff, B., and Kück, U. (2004). AcFKH1, a novel member of the forkhead family, associates with the RFX transcription factor CPCR1 in the cephalosporin C-producing fungus Acremonium chrysogenum. Gene 342, 269–281.
Schneider, P., Misiek, M., and Hoffmeister, D. (2008). In vivo and in vitro production options for fungal secondary metabolites. Mol. Pharm. 5, 234–242. doi: 10.1021/mp7001544
Schulz, B., Boyle, C., Draeger, S., Römmert, A. K., and Krohn, K. (2002). Endophytic fungi: a source of novel biologically active secondary metabolites. Mycol. Res. 106, 996–1004.
Shao, C.-L., Wang, C.-Y., Gu, Y.-C., Wei, M.-Y., Pan, J.-H., Deng, D.-S., et al. (2010). Penicinoline, a new pyrrolyl 4-quinolinone alkaloid with an unprecedented ring system from an endophytic fungus Penicillium sp. Bioorg. Med. Chem. Lett. 20, 3284–3286.
Shimizu, K., and Keller, N. P. (2001). Genetic involvement of a cAMP-dependent protein kinase in a G protein signaling pathway regulating morphological and chemical transitions in Aspergillus nidulans. Genetics 157, 591–600.
Shoresh, M., Harman, G. E., and Mastouri, F. (2010). Induced systemic resistance and plant responses to fungal biocontrol agents. Annu. Rev. Phytopathol. 48, 21–43. doi: 10.1146/annurev-phyto-073009-114450
Shwab, E. K., and Keller, N. P. (2008). Regulation of secondary metabolite production in filamentous ascomycetes. Mycol. Res. 112(Pt 2), 225–230. doi: 10.1016/j.mycres.2007.08.021
Siddhardha, B., and Meena, H. (2020). “Microbial secondary metabolites: natural benediction elements for plants during abiotic and biotic stress conditions,” in Microbial Services in Restoration Ecology, eds J. S. Singh and Vimal, S. R. (Elsevier), 99–107.
Sieber, T. N. (2007). Endophytic fungi in forest trees: are they mutualists? Fungal Biol. Rev. 21, 75–89.
Sigurdson, G., Tang, P., and Giusti, M. (2018). Cis–trans configuration of coumaric acid acylation affects the spectral and colorimetric properties of anthocyanins. Molecules 23:598.
Singh, L. P., Gill, S. S., and Tuteja, N. (2011). Unraveling the role of fungal symbionts in plant abiotic stress tolerance. Plant Signal. Behav. 6, 175–191. doi: 10.4161/psb.6.2.14146
Slot, J. C., and Rokas, A. (2011). Horizontal transfer of a large and highly toxic secondary metabolic gene cluster between fungi. Curr. Biol. 21, 134–139. doi: 10.1016/j.cub.2010.12.020
Soanes, D., and Richards, T. A. (2014). Horizontal gene transfer in eukaryotic plant pathogens. Annu. Rev. Phytopathol. 52, 583–614. doi: 10.1146/annurev-phyto-102313-050127
Soliman, S. S., Trobacher, C. P., Tsao, R., Greenwood, J. S., and Raizada, M. N. (2013). A fungal endophyte induces transcription of genes encoding a redundant fungicide pathway in its host plant. BMC Plant Biol. 13:93. doi: 10.1186/1471-2229-13-93
Somerville, C., and Somerville, S. (1999). Plant functional genomics. Science 285, 380–383. doi: 10.1126/science.285.5426.380
Song, Q., Li, F., Nan, Z., Coulter, J. A., and Wei, W. (2020). Do Epichloë endophytes and their grass symbiosis only producetoxic alkaloids to insects and livestock? J. Agric. Food Chem. 68, 1169–1185. doi: 10.1021/acs.jafc.9b06614
Spanu, P. D., Abbott, J. C., Amselem, J., Burgis, T. A., Soanes, D. M., Stüber, K., et al. (2010). Genome expansion and gene loss in powdery mildew fungi reveal tradeoffs in extreme parasitism. Science 330, 1543–1546. doi: 10.1126/science.1194573
Spatafora, J. W., Sung, G. H., Sung, J. M., Hywel-Jones, N. L., and White, J. F. J. (2007). Phylogenetic evidence for an animal pathogen origin of ergot and the grass endophytes. Mol. Ecol. 16, 1701–1711. doi: 10.1111/j.1365-294X.2007.03225.x
Sproul, D., Gilbert, N., and Bickmore, W. A. (2005). The role of chromatin structure in regulating the expression of clustered genes. Nat. Rev. Genet. 6, 775–781. doi: 10.1038/nrg1688
Sriram, D., Yogeeswari, P., Thirumurugan, R., and Ratan Bal, T. (2005). Camptothecin and its analogues: a review on their chemotherapeutic potential. Nat. Prod. Res. 19, 393–412.
Stahl, E., Hilfiker, O., and Reymond, P. (2018). Plant-arthropod interactions: who is the winner? Plant J. 93, 703–728. doi: 10.1111/tpj.13773
Stierle, A., Strobel, G., and Stierle, D. (1993). Taxol and taxane production by Taxomyces andreanae, an endophytic fungus of Pacific yew. Science 260, 214–216. doi: 10.1126/science.8097061
Stone, J., Polishook, J. D., and White, J. F. (2004). “Endophytic fungi,” in Biodiversity of Fungi, eds G. M. Mueller, G. F. Bills, and J. F. White (Amsterdam: Elsevier), 241–270.
Strobel, G. (2018). The emergence of endophytic microbes and their biological promise. J. Fungi 4:57. doi: 10.3390/jof4020057
Su, Y. Y., Guo, L. D., and Hyde, K. D. (2010). Response of endophytic fungi of Stipa grandis to experimental plant function group removal in Inner Mongolia steppe, China. Fungal Divers. 43, 93–101.
Sun, J., Guo, L., Zang, W., Ping, W., and Chi, D. (2008). Diversity and ecological distribution of endophytic fungi associated with medicinal plants. Sci. China C Life Sci. 51, 751–759. doi: 10.1007/s11427-008-0091-z
Sun, X., and Guo, L.-D. (2012). Endophytic fungal diversity: review of traditional and molecular techniques. Mycology 3, 65–76. doi: 10.1080/21501203.2012.656724
Sun, X., Guo, L. D., and Hyde, K. D. (2011). Comunity composition of endophytic fungi in Acer truncatum and their role in decomposition. Fungal Divers. 47, 85–95.
Tag, A., Hicks, J., Garifullina, G., Ake, C. Jr., Phillips, T. D., Beremand, M., et al. (2000). G-protein signalling mediates differential production of toxic secondary metabolites. Mol. Microbiol. 38, 658–665.
Tamaru, H., and Selker, E. U. (2001). A histone H3 methyltransferase controls DNA methylation in Neurospora crassa. Nature 414, 277–283. doi: 10.1038/35104508
Tan, R. X., and Zou, W. X. (2001). Endophytes: a rich source of functional metabolites. Nat. Prod. Rep. 18, 448–459.
Tao, G., Liu, Z. Y., Hyde, K. D., Liu, X. Z., and Yu, Z. N. (2008). Whole rDNA analysis reveals novel and endophytic fungi in Bletilla ochracea (Orchidaceae). Fungal Divers. 33, 101–112.
Tejesvi, M. V., Tamhankar, S. A., Kini, K. R., Rao, V. S., and Prakash, H. S. (2009). Phylogenetic analysis of endophytic Pestalotiopsis species from ethnopharmaceutically important medicinal trees. Fungal Divers. 38, 167–183.
Theodorakidou, M., Nikola, O.-A., and Lambrou, G. I. (2018). The multiple roles of steroids and anabolic steroids and its relations to cardiovasular and musculoskeletal pathology: a brief review. J. Res. Pract. Musculoskelet. Syst. 2, 22–30. doi: 10.22540/jrpms-02-022
Trapp, S. C., Hohn, T. M., McCormick, S., and Jarvis, B. B. (1998). Characterization of the gene cluster for biosynthesis of macrocyclic trichothecenes in Myrothecium roridum. Mol. Gen. Genet. 257, 421–432.
Uzma, F., Mohan, C. D., Hashem, A., Konappa, N. M., Rangappa, S., Kamath, P. V., et al. (2018). Endophytic fungi-alternative sources of cytotoxic compounds: a review. Front. Pharmacol. 9:309. doi: 10.3389/fphar.2018.00309
Vaishnav, P., and Demain, A. L. (2010). Unexpected applications of secondary metabolites. Biotechnol. Adv. 29, 223–229. doi: 10.1016/j.biotechadv.2010.11.006
Van der Ent, S., Van Wees, S. C., and Pieterse, C. M. (2009). Jasmonate signaling in plant interactions with resistance-inducing beneficial microbes. Phytochemistry 70, 1581–1588.
Vandenkoornhuyse, P., Baldauf, S. L., Leyval, C., Straczek, J., and Young, J. P. (2002). Extensive fungal diversity in plant roots. Science 295:2051. doi: 10.1126/science.295.5562.2051
Varma, A., Rai, M., Sahay, S., and Sahay, N. S. (2000). “Microbial-biotechnology: new-paradigms and role in sustainable agriculture,” in Microbial Biotechnology for Sustainable Development and Productivity, ed. R. C. Rajak (India: Scientific Publishers), 22–37.
Vellosa, J. C. R., Regasini, L. O., Khalil, N. M., Bolzani, V. D. S., Khalil, O. A., Manente, F. A., et al. (2011). Antioxidant and cytotoxic studies for kaempferol, quercetin and isoquercitrin. Eclética Quim. 36, 7–20.
Verbon, E. H., and Liberman, L. M. (2016). Beneficial microbes affect endogenous mechanisms controlling root development. Trends Plant Sci. 21, 218–229.
Violle, C., Reich, P. B., Pacala, S. W., Enquist, B. J., and Kattge, J. (2014). The emergence and promise of functional biogeography. Proc. Natl. Acad. Sci. U.S.A. 111, 13690–13696. doi: 10.1073/pnas.1415442111
Waller, F., Achatz, B., Baltruschat, H., Fodor, J., Becker, K., Fischer, M., et al. (2005). The endophytic fungus Piriformospora indica reprograms barley to salt-stress tolerance, disease resistance, and higher yield. Proc. Natl. Acad. Sci. U.S.A. 102, 13386–13391. doi: 10.1073/pnas.0504423102
Wang, M., Tu, L., Yuan, D., Zhu, D., Shen, C., Li, J., et al. (2018). Reference genome sequences of two cultivated allotetraploid cottons, Gossypium hirsutum and Gossypium barbadense. Nat. Genet. 51, 224–229. doi: 10.1038/s41588-018-0282-x
Wang, R., Liu, G., Yang, M., Wang, M., and Zhou, L. (2016). Total synthesis and antifungal activity of palmarumycin CP17 and its methoxy analogues. Molecules 21:600. doi: 10.3390/molecules21050600
Williams, D. H., Stone, M. J., Hauck, P. R., and Rahman, S. K. (1989). Why are secondary metabolites (natural products) biosynthesized? J. Nat. Prod. 52, 1189–1208.
Woloshuk, C. P., Foutz, K. R., Brewer, J. F., Bhatnagar, D., Cleveland, T. E., and Payne, G. A. (1994). Molecular characterization of aflR, a regulatory locus for aflatoxin biosynthesis. Appl. Environ. Microbiol. 60, 2408–2414.
Wu, D., Zhang, S.-Y., Liu, Y.-Q., Wu, X.-B., Zhu, G.-X., Zhang, Y., et al. (2015). Synthesis, biological activities, and quantitative structure–activity relationship (QSAR) study of novel camptothecin analogues. Molecules 20, 8634–8653.
Xie, W., Hao, Z., Yu, M., Wu, Z., Zhao, A., Li, J., et al. (2019). Improved phosphorus nutrition by arbuscular mycorrhizal symbiosis as a key factor facilitating glycyrrhizin and liquiritin accumulation in Glycyrrhiza uralensis. Plant Soil 439, 243–257. doi: 10.1007/s11104-018-3861-9
Xu, X. H., Su, Z. Z., Wang, C., Kubicek, C. P., Feng, X. X., Mao, L. J., et al. (2014). The rice endophyte Harpophora oryzae genome reveals evolution from a pathogen to a mutualistic endophyte. Sci. Rep. 4:5783. doi: 10.1038/srep05783
Yadav, V., Wang, Z., Wei, C., Amo, A., Ahmed, B., Yang, X., et al. (2020). Phenylpropanoid pathway engineering: an emerging approach towards plant defense. Pathogens 9:312. doi: 10.3390/pathogens9040312
Yan, L., Zhu, J., Zhao, X., Shi, J., Jiang, C., and Shao, D. (2019). Beneficial effects of endophytic fungi colonization on plants. Appl. Microbiol. Biotechnol. 103, 3327–3340. doi: 10.1007/s00253-019-09713-2
Yang, H., Tong, J., Lee, C. W., Ha, S., Eom, S. H., and Im, Y. J. (2015). Structural mechanism of ergosterol regulation by fungal sterol transcription factor Upc2. Nat. Commun. 6:6129.
Yang, H.-R., Yuan, J., Liu, L.-H., Zhang, W., Chen, F., and Dai, C.-C. (2019). Endophytic Pseudomonas fluorescens induced sesquiterpenoid accumulation mediated by gibberellic acid and jasmonic acid in Atractylodes macrocephala Koidz plantlets. Plant Cell Tissue Organ Cult. 138, 445–457.
Yang, X., Strobel, G., Stierle, A., Hess, W., Lee, J., and Clardy, J. (1994). A fungal endophyte-tree relationship: Phoma sp. in Taxus wallachiana. Plant Sci. 102, 1–9.
Yang, Y., Jin, Z., Jin, Q., and Dong, M. (2015). Isolation and fatty acid analysis of lipid-producing endophytic fungi from wild Chinese Torreya Grandis. Microbiology 84, 710–716.
Yi, X. U., and Gray, S. M. (2020). Aphids and their transmitted potato viruses: a continuous challenges in potato crops. J. Integr. Agric. 19, 367–375.
Young, C. A., Felitti, S., Shields, K., Spangenberg, G., Johnson, R. D., Bryan, G. T., et al. (2006). A complex gene cluster for indole-diterpene biosynthesis in the grass endophyte Neotyphodium lolii. Fungal Genet. Biol. 43, 679–693. doi: 10.1016/j.fgb.2006.04.004
Yu, H., Zhang, L., Li, L., Zheng, C., Guo, L., Li, W., et al. (2010). Recent developments and future prospects of antimicrobial metabolites produced by endophytes. Microbiol. Res. 165, 437–449.
Yu, J. H., and Keller, N. (2005). Regulation of secondary metabolism in filamentous fungi. Annu. Rev. Phytopathol. 43, 437–458. doi: 10.1146/annurev.phyto.43.040204.140214
Yu, J. H., Butchko, R. A., Fernandes, M., Keller, N. P., Leonard, T. J., and Adams, T. H. (1996). Conservation of structure and function of the aflatoxin regulatory gene aflR from Aspergillus nidulans and A. flavus. Curr. Genet. 29, 549–555.
Zaiyou, J., Li, M., and Xiqiao, H. (2017). An endophytic fungus efficiently producing paclitaxel isolated from Taxus wallichiana var. mairei. Medicine 96:e7406. doi: 10.1097/MD.0000000000007406
Zhang, H. W., Song, Y. C., and Tan, R. X. (2006). Biology and chemistry of endophytes. Nat. Prod. Rep. 23, 753–771. doi: 10.1039/b609472b
Zhang, H., and Sonnewald, U. (2017). Differences and commonalities of plant responses to single and combined stresses. Plant J. 90, 839–855.
Zhang, J., Liu, Y. X., Zhang, N., Hu, B., Jin, T., Xu, H., et al. (2019). NRT1.1B is associated with root microbiota composition and nitrogen use in field-grown rice. Nat. Biotechnol. 37, 676–684. doi: 10.1038/s41587-019-0104-4
Zhang, P., Yuan, X.-L., Du, Y.-M., Zhang, H.-B., Shen, G.-M., Zhang, Z.-F., et al. (2019). Angularly prenylated indole alkaloids with antimicrobial and insecticidal activities from an endophytic fungus Fusarium sambucinum TE-6L. J. Agric. Food Chem. 67, 11994–12001. doi: 10.1021/acs.jafc.9b05827
Zhang, Q., Wei, X., and Wang, J. (2012). Phillyrin produced by Colletotrichum gloeosporioides, an endophytic fungus isolated from Forsythia suspensa. Fitoterapia 83, 1500–1505.
Zhang, Y., Han, T., Ming, Q., Wu, L., Rahman, K., and Qin, L. (2012). Alkaloids produced by endophytic fungi: a review. Nat. Prod. Commun. 7, 963–968.
Zhao, J., Li, C., Wang, W., Zhao, C., Luo, M., Mu, F., et al. (2013). Hypocrea lixii, novel endophytic fungi producing anticancer agent cajanol, isolated from pigeon pea (C ajanus cajan [L.] M illsp.). J. Appl. Microbiol. 115, 102–113.
Zhou, X., Zhu, H., Liu, L., Lin, J., and Tang, K. (2010). A review: recent advances and future prospects of taxol-producing endophytic fungi. Appl. Microbiol. Biotechnol. 86, 1707–1717. doi: 10.1007/s00253-010-2546-y
Keywords: symbiosis, endophytic fungi, endophytic fungi-host interaction, genetic regulation, secondary metabolites
Citation: Alam B, Lǐ J, Gě Q, Khan MA, Gōng J, Mehmood S, Yuán Y and Gǒng W (2021) Endophytic Fungi: From Symbiosis to Secondary Metabolite Communications or Vice Versa? Front. Plant Sci. 12:791033. doi: 10.3389/fpls.2021.791033
Received: 07 October 2021; Accepted: 29 November 2021;
Published: 17 December 2021.
Edited by:
Sabine Dagmar Zimmermann, Délégation Languedoc Roussillon, Center for the National Scientific Research (CNRS), FranceReviewed by:
Jorge Poveda, Public University of Navarre, SpainCopyright © 2021 Alam, Lǐ, Gě, Khan, Gōng, Mehmood, Yuán and Gǒng. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Wànkuí Gǒng, Z29uZ3dhbmt1aUBjYWFzLmNu; Yǒulù Yuán, eXVhbnlvdWx1QGNhYXMuY24=
†These authors have contributed equally to this work
‡ORCID: Mueen Alam Khan, orcid.org/0000-0003-3261-9724; Jǔwǔ Gōng, orcid.org/0000-0002-9708-943X; Yǒulù Yuán, orcid.org/0000-0003-0644-553X; Wànkuí Gǒng, orcid.org/0000-0002-8556-0669
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.