 Jan Petersen
Jan Petersen Anxhela Rredhi1†
Anxhela Rredhi1† Tilman Kottke
Tilman Kottke Maria Mittag
Maria Mittag- 1Matthias Schleiden Institute of Genetics, Bioinformatics and Molecular Botany, Friedrich Schiller University, Jena, Germany
- 2Experimental Molecular Biophysics, Department of Physics, Freie Universität Berlin, Berlin, Germany
- 3Department of Chemistry, Bielefeld University, Bielefeld, Germany
- 4Biophysical Chemistry and Diagnostics, Medical School OWL, Bielefeld University, Bielefeld, Germany
Algae are photosynthetic eukaryotic (micro-)organisms, lacking roots, leaves, and other organs that are typical for land plants. They live in freshwater, marine, or terrestrial habitats. Together with the cyanobacteria they contribute to about half of global carbon fixation. As primary producers, they are at the basis of many food webs and they are involved in biogeochemical processes. Algae are evolutionarily distinct and are derived either by primary (e.g., green and red algae) or secondary endosymbiosis (e.g., diatoms, dinoflagellates, and brown algae). Light is a key abiotic factor needed to maintain the fitness of algae as it delivers energy for photosynthesis, regulates algal cell- and life cycles, and entrains their biological clocks. However, excess light can also be harmful, especially in the ultraviolet range. Among the variety of receptors perceiving light information, the cryptochromes originally evolved as UV-A and blue-light receptors and have been found in all studied algal genomes so far. Yet, the classification, biophysical properties, wavelength range of absorbance, and biological functions of cryptochromes are remarkably diverse among algal species, especially when compared to cryptochromes from land plants or animals.
Introduction
What Are Algae?
Algae are photosynthetic eukaryotes defined primarily by their lack of roots, leaves, and the stem that are typical for higher plants (Parker et al., 2008). They are divided into microscopically small microalgae that are a major part of phytoplankton and macroalgae, including seaweeds. Algae are found anywhere from soil to lakes, rivers and oceans and they are crucial to food webs (Steele, 1974). Algal activities can even influence biogeochemical processes as observed with the Greenland ice sheet (McCutcheon et al., 2021). Together with cyanobacteria, aquatic algae are responsible for about 50% of global carbon fixation (Field et al., 1998) and thus are of high ecological relevance. Evolutionarily, algae can be divided into two major groups, with one group derived from primary endosymbiosis and the other one from secondary endosymbiosis (Keeling et al., 2005). For primary endosymbionts, a unicellular eukaryotic cell engulfed a cyanobacterium to become a chloroplast. In the case of secondary endosymbionts, a unicellular eukaryotic cell engulfed either a green alga (secondary green) or a red alga (secondary red). Recently, it was shown that tertiary and possibly even higher-order endosymbiotic events occurred (Sibbald and Archibald, 2020), but these will not be part of this review. Primary endosymbionts (named Archaeplastida) include glaucophytes (algae with cyanelle plastids), rhodophytes (red algae), chlorophytes (green algae), and all land plants together with ferns and mosses (Bowman et al., 2007; Adl et al., 2012). Green algae and land plants are also grouped as Chloroplastida. All these groups possess plastids surrounded by two envelope membranes. In the lineages with secondary endosymbionts, plastids are encircled by either four (e.g., diatoms, brown algae, haptophytes, and cryptophytes) or three membranes (euglenophytes, dinoflagellates; Gentil et al., 2017). Some of these lineages still contain the nucleus of the engulfed green or red alga, but in a strongly reduced form, called nucleomorph (e.g., in cryptophytes). Others such as euglenophytes, diatoms, dinoflagellates, or brown algae have completely lost this nucleus (Gentil et al., 2017). In this review, we focus solely on algal groups with cryptochromes that have been so far phylogenetically described based on existing genomes or have been characterized by their photoreceptors.
The Influence of Light on Algal Life and the Diversity of Algal Photoreceptors
Light is an important source of energy and information for algal life on our planet (Figure 1). As for all photosynthetic organisms, algae transform the radiation energy of the sunlight into chemical energy (Eberhard et al., 2008). They perceive light via pigments or specialized photoreceptors. Light regulates their photosynthesis and balances their photosynthetic apparatus, controls their behavior (photoorientation), entrains their circadian clocks, and influences their cell and sexual cycles as well as developmental processes (reviewed in Hegemann, 2008; Kianianmomeni and Hallmann, 2014; Kottke et al., 2017; Rredhi et al., 2021).
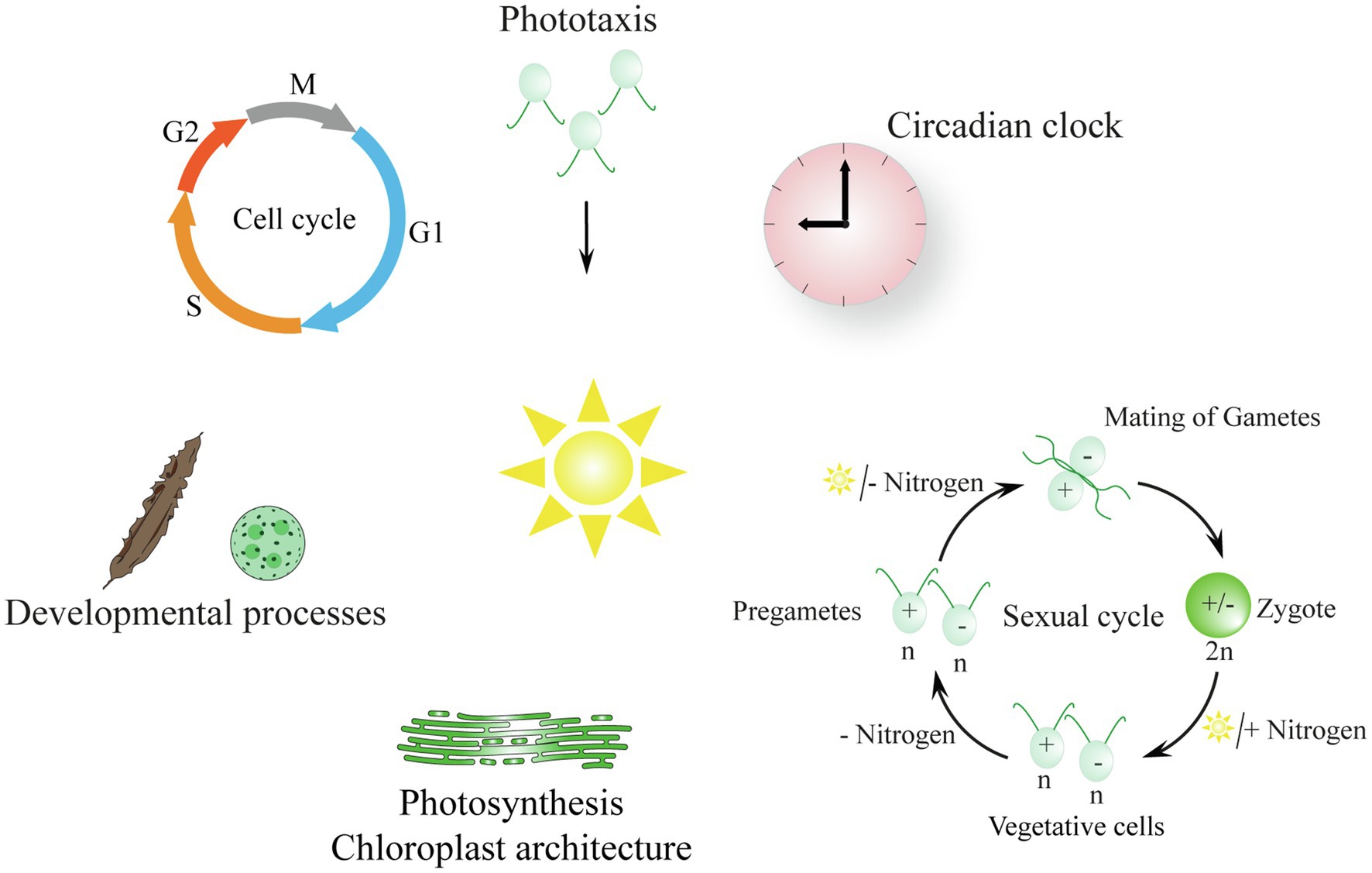
Figure 1. Biological processes in algae that are regulated by light (reviewed in Hegemann, 2008; Kianianmomeni and Hallmann, 2014; Kottke et al., 2017; Sasso et al., 2018). The description of the symbolized processes begins at the top of the scheme and moves in a clockwise manner: Photoorientation of flagellate algae: tactic movement to the light is shown; phobic movement occurs with strong light (not shown). ǀ The algal circadian clock is entrained by light–dark cycles. ǀ The sexual cycle of Chlamydomonas: vegetative cells turn into pregametes in the dark without a nitrogen source and then become gametes in the light in the absence of a nitrogen source; gametes fuse and a resilient zygote is formed that needs a nitrogen source and light to undergo meiosis. ǀ Light is needed for photosynthesis and influences the chloroplast architecture; thylakoid membranes are shown exemplarily (Rredhi et al., 2021) ǀ Light influences developmental processes in the multicellular algae Saccharina and Volvox (Kianianmomeni and Hallmann, 2015; Yang et al., 2020). ǀ The cell cycle is symbolized; algal cells are synchronized by light–dark cycles; M, mitosis; G1, gap1; S, synthesis; G2, gap2 (Coesel et al., 2009; Cross and Umen, 2015).
Cyanobacteria, also known as prokaryotic blue-green algae, and eukaryotic algae have an amazing repertoire of different photoreceptors compared to land plants or animals. For many members, we still do not know their detailed functions and/or mechanisms (Kianianmomeni and Hallmann, 2014; Greiner et al., 2017; Jaubert et al., 2017; Kottke et al., 2017; Kroth et al., 2017; König et al., 2017b; Rockwell and Lagarias, 2020). In addition to the classically known photoreceptors from land plants which absorb in the red/far red (phytochrome) and blue region of the visible spectrum (cryptochrome, phototropin, Zeitlupe) as well as in the UV-B (UVR8; Kim et al., 2007; Fankhauser and Christie, 2015; Ahmad, 2016; Wang and Lin, 2020; Podolec et al., 2021), algae bear several novel types of light sensors. Moreover, cyanobacteria and algae also possess a larger variety of the above-mentioned “classical” photoreceptors with new properties and biological functions. For example, cyanobacteriochromes, which are phytochromes in cyanobacteria, absorb strongly shifted from the well known red/far-red range to shorter wavelengths of visible light (Rockwell et al., 2012). Moreover, new types of cryptochromes are present in algae with novel biophysical properties such as red-light absorption and will be detailed in the chapters below.
Numerous photoreceptors are unique to algae. For example, aureochrome photoreceptors are only present in a single group of algae, the photosynthetic stramenopiles, including Xanthophyceae (yellow-green algae), diatoms, and brown algae (Kroth et al., 2017). These aureochrome photoreceptors contain a light-oxygen-voltage (LOV) domain for light reception and a basic region leucine zipper (bZIP) domain for DNA binding and act as light-driven transcription factors. In the green unicellular model alga Chlamydomonas reinhardtii (Cr), even 18 different types of photoreceptors have been reported, including two channelrhodopsins that are fundamental to its photoorientation and several histidine kinase rhodopsins (HKRs) whose functions are largely unknown (Greiner et al., 2017; Luck and Hegemann, 2017). HKRs contain a His-kinase-, response regulator-, and a rhodopsin domain. Some of the HKRs even have guanylyl cyclase activity that has been recently used for application as an optogenetic tool (Tian et al., 2021). The field of optogenetics is self-evolving since knowledge on the exceptional properties of Cr channelrhodopsins became available (Hegemann and Nagel, 2013).
Further types of (possibly) novel light sensors were recently discovered in phytoplankton in the open ocean (Coesel et al., 2021). The genomes of these marine microalgae encode light-sensing proteins with new combinations of known domain structures or even fusions of different types of photoreceptors. For example, LOV domains were found within one protein together with different DNA-binding domains such as a homeobox or a heat shock factor domain or with signal-transduction motifs (e.g., an EF-hand; Coesel et al., 2021). A new type of dual cryptochrome that is fused to another photoreceptor was also found in these genomes (Makita et al., 2021) and is included in this review.
It should also be stated that the functions of photoreceptors in algae may differ from those in higher plants. For example, Cr phototropin influences the sexual cycle (Huang and Beck, 2003), the development of the eyespot, a primitive visual system of the alga (Trippens et al., 2012), and mediates the feedback regulation of photosynthesis in the green alga (Petroutsos et al., 2016).
Blue-light-activated adenyl cyclases represent another type of light sensors in algae. They mediate photoavoidance in the secondary green alga Euglena gracilis (Iseki et al., 2002) and contain blue-light sensor using flavin adenine dinucleotide (BLUF) domains (Ito et al., 2010). In the green alga Cr as well as in animal spermatozoa, the BLUF domain is directly associated with dynein and involved in ciliary motility (Kutomi et al., 2021).
Variety of Algal Cryptochromes
In this review, we focus on the variety of cryptochromes found in algae. Cryptochromes are flavoproteins that were first described as blue-light receptors in plants and animals (Chaves et al., 2011). They are derived from the blue-light-dependent DNA repair enzymes called photolyases that can be found in pro- and eukaryotes (Ahmad and Cashmore, 1993; Sancar, 2003; Chaves et al., 2011). As primary light sensors, cryptochromes and photolyases carry a non-covalently bound flavin adenine dinucleotide (FAD) molecule within their photolyase homology domain (PHR). Additionally, they may bind an antenna chromophore like 5,10-methenyltetrahydrofolate (MTHF) or 8-hydroxy-7,8-didemethyl-5-deazaflavin (8-HDF). The antenna chromophore harvests additional light and transfers the energy to the FAD (Hoang et al., 2008). Photoexcitation of the FAD may lead to a change of its redox state that can be either oxidized (FADox), semireduced as an anion radical (FAD•−), semireduced as a neutral radical (FADH•) or fully reduced (FADH−; Chaves et al., 2011). Conformational changes are promoted within the cryptochrome protein structure depending on the FAD redox state, resulting in different signaling properties. Most cryptochromes possess a C-terminal extension (CCT) whose structure is rather undefined and only shows poor conservation. The CCT varies in length depending on the cryptochrome. Nevertheless, the CCT is of great importance for cryptochrome function and downstream signaling (Chaves et al., 2011). The first cryptochromes were analyzed as blue-light receptors in the land plant Arabidopsis thaliana (At CRY1 and At CRY2; Ahmad and Cashmore, 1993: Chaves et al., 2011) and in the fly Drosophila melanogaster (dCRY) as an animal-type I CRY (Emery et al., 1998). Later, it was found that the cryptochromes from mice or humans group to animal type II CRYs. They do not act as light sensors but instead have a function in the central oscillator of the circadian clock (Okamura et al., 1999; Todo, 1999; Chaves et al., 2011).
Cryptochrome Categories in Algae
To date, four different classes of cryptochromes are known in algae based on phylogenetic analyses and further characterizations. Moreover, a fifth class was recently described consisting of a fusion of a cryptochrome with another photoreceptor. The following section will present details on the five different classes:
1. The classical plant cryptochromes (pCRYs) present in land plants (At CRY1 and At CRY2) are found in some but not in all algae. Plant CRYs, like all cryptochromes, share the conserved PHR at the N-terminus binding FAD (Chaves et al., 2011), and contain a comparatively long CCT (Reisdorph and Small, 2004; Müller et al., 2017). Cr pCRY has the longest known extension with about 500 amino acids (Figure 2). Plant CRYs may also bind MTHF as antenna chromophore as derived from their homology to CPD (cyclobutane pyrimidine dimer) photolyases (Chaves et al., 2011; Beel et al., 2012). There is no indication that pCRYs still have photolyase activity (Lin and Shalitin, 2003). Plant CRYs are found in many investigated Chloroplastida (Figure 3) but seem to be absent in the green picoalga Ostreococcus tauri (Ot), in the red macroalga Porphyra umbilicalis (Pum) or in the diatom Phaeodactylum tricornutum (Pt; Figure 3).
2. The cryptochrome–photolyase family (CPF1) of animal-like cryptochromes. For a long time, it was thought that all cryptochromes have lost photolyase activity. However, it was recently discovered that some algal animal-like CRYs from Pt, Ot, and Cr have maintained photolyase activity (Coesel et al., 2009; Heijde et al., 2010; Franz et al., 2018). These animal-like cryptochromes group by their phylogeny closely to the cryptochrome (6-4)–photolyase family (CPF1) and are thus phylogenetically located in between the animal-type I and -type II cryptochromes from animals (Beel et al., 2012). They exert further biological functions including light perception and circadian clock control as well as the control of the algal Cr life cycle (Coesel et al., 2009; Heijde et al., 2010; Beel et al., 2012; Zou et al., 2017). Cr aCRY binds 8-HDF as antenna chromophore (Franz et al., 2018; Figure 2). The CCTs of the animal-like CRYs are usually shorter as compared to the pCRYs. Notably, CPF animal-like CRYs are widely distributed in algae but are missing in land plants (Figure 3).
3. The CRY-DASH (Drosophila, Arabidopsis, Synechocystis, Homo) cryptochromes. CRY-DASH (-like) proteins are found in many organisms from bacteria to vertebrates (Brudler et al., 2003; Kiontke et al., 2020). They group closely to CPD photolyases (Beel et al., 2012; Fortunato et al., 2015). Bacterial, plant, and fungal CRY-DASHs can repair CPD in single-stranded and even in double-stranded DNA, as reported for some members of the mucoromycotina fungi (Selby and Sancar, 2006; Pokorny et al., 2008; Tagua et al., 2015; Navarro et al., 2020). Studies on repair activities by members of the cryptochrome/photolyase family in algal systems are rare. It was found that two out of three putative CRY-DASHs of the red picoalga Cm repair CPD lesions, but only in single-stranded and not in double-stranded DNA (Asimgil and Kavakli, 2012). Interestingly, CRY-DASH proteins can be located in organelles. The land plant At CRY3 was found in the chloroplast and in mitochondria (Kleine et al., 2003), while the algal Cr CRY-DASH1 protein was only found in the chloroplast (Rredhi et al., 2021; Figure 2). Recently, algal CRY-DASHs were shown to be involved in further biological functions beside DNA repair, supporting their additional or alternative role as photoreceptors (Yang et al., 2020; Rredhi et al., 2021). CRY-DASH proteins were found in all selected algal systems (Figure 3), including an Antarctic Chlamydomonas sp. ICE-L in which its highest transcript expression level is at 5°C and a salinity of 32% (Zhang et al., 2020).
4. The plant-like CryP, found in algae and metazoans. Instead of a classical pCRY, the genome of Pt encodes a plant-like CRY, named CryP (Juhas et al., 2014; König et al., 2017b) that was later identified in metazoan genomes (Oliveri et al., 2014). CryP binds FAD and MTHF as chromophores and has a CCT of about 70 residues (Figure 2). As in CRY-DASH proteins, the FAD in CryP is present in the neutral radical state after isolation as opposed to the oxidized state found in pCRYs (see below; Juhas et al., 2014). A plant-like CRY is also present in the picoalga Ot that lacks a classical pCRY. Interestingly, the red picoalga Cyanidioschyzon merolae (Cm) and the green picoalga Pycnococcus provasolii (Pp) have both plant-like CRYs and plant CRYs (Figure 3).
5. Dualchrome, a new dual-photoreceptor chimera. Recently, a new type of photoreceptor from the marine picoalga Pp was discovered in metagenome data of ocean picoplankton, named dualchrome (DUC1, Figures 2, 3). DUC1 bears a fusion of a phytochrome and a plant CRY (Makita et al., 2021) and was not found in any other alga or other organisms so far.
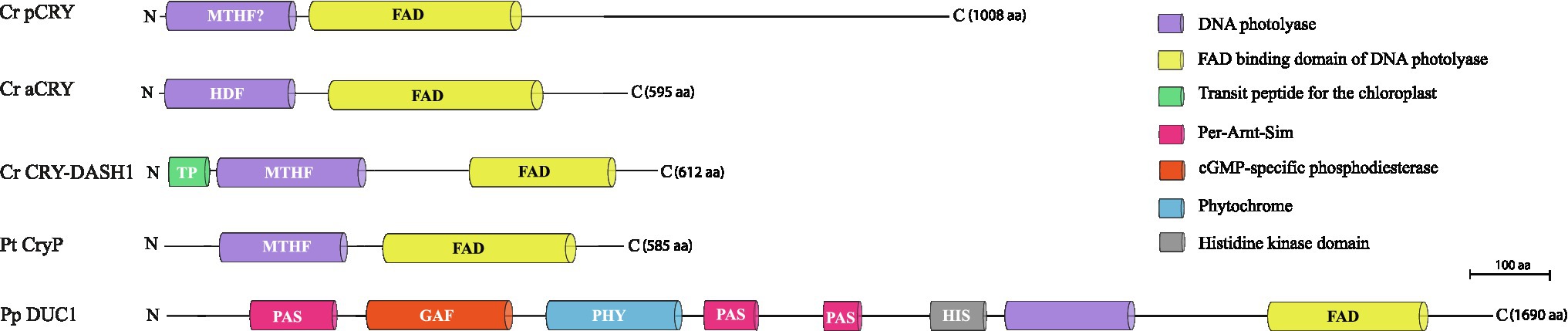
Figure 2. Cryptochrome categories in algae (Juhas et al., 2014; Kottke et al., 2017; Makita et al., 2021; Rredhi et al., 2021). A representative member of each of the five known categories is shown, starting with the classical plant cryptochrome of the green flagellate alga Chlamydomonas reinhardtii (Cr pCRY) and its long C-terminal extension. It is followed by the Cr animal-like CRY of the cryptochrome/photolyase family (Cr aCRY), the Cr CRY-DASH1 with its chloroplast transit peptide, the plant-like CryP of the diatom Phaeodactylum tricornutum and the dual cryptochrome DUC1 from the green picoalga Pycnococcus provasolii, a fusion of a phytochrome and a cryptochrome. Domains are indicated, including the names of the chromophores (FAD) and antenna chromophores (MTHF, 8-HDF); FAD, flavin adenine dinucleotide; MTHF, 5,10-methenyltetrahydrofolate; 8-HDF, 8-hydroxy-7,8-didemethyl-5-deazaflavin.
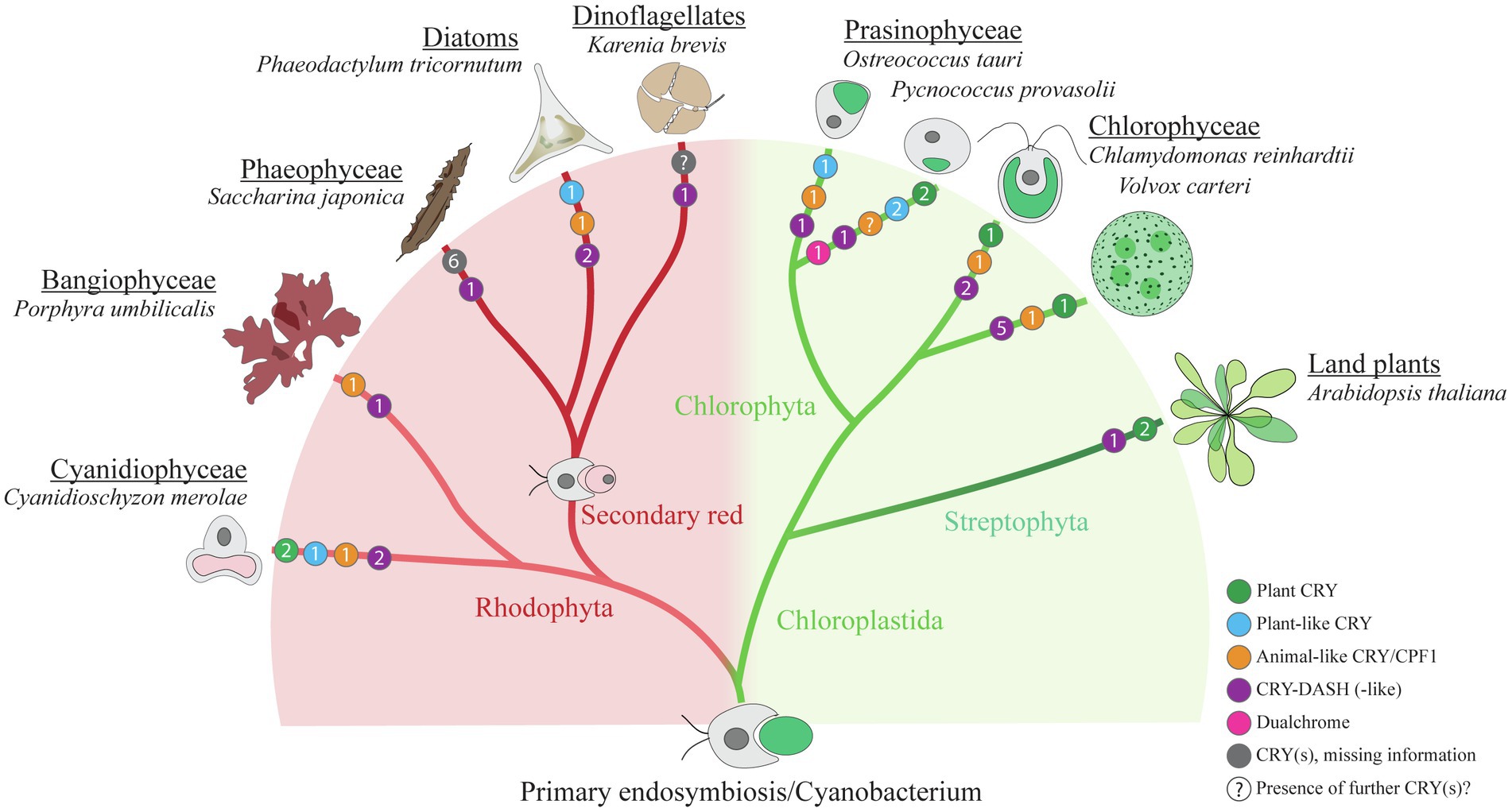
Figure 3. Cryptochrome distribution within selected algal species. Simplified schematic cladogram of cryptochrome (CRY) distribution within selected algal species in comparison with the land plant Arabidopsis thaliana. Colored circles indicate the numbers (presented in each circle) and representatives of different types of CRYs within the presented species (green, plant CRY; blue, plant-like CRY; orange, animal-like CRY/CPF1; purple, CRY-DASH and CRY-DASH-like; magenta, dualchrome; gray, CRY(s) that are currently not assigned to the former groups; “?,” indicating that there may be further potential CRY(s) in this organism that have not been identified so far). Information for the presented data was taken from the following references: Cyanidioschyzon merolae (Cm; Brawley et al., 2017), Porphyra umbilicalis (Pum; Brawley et al., 2017), Saccharina japonica (Sj; Deng et al., 2012; Yang et al., 2020), Phaeodactylum tricornutum (Pt; Juhas et al., 2014; Oliveri et al., 2014; Fortunato et al., 2015), Karenia brevis (Kb; Brunelle et al., 2007), Ostreococcus tauri (Ot; Heijde et al., 2010; Kottke et al., 2017), Pycnococcus provasolii (Pp; Makita et al., 2021), Chlamydomonas reinhardtii (Cr; Beel et al., 2012; Kottke et al., 2017), Volvox carteri (Vc; Kottke et al., 2017) and Arabidopsis thaliana (At; Chaves et al., 2011). Please note that three putative CRY-DASHs are predicted for Cm in the analysis of Asimgil and Kavakli (2012). For the genus Karenia, three putative CRY/photolyase members are predicted (Shikata et al., 2019). The number of cryptochrome/photolyase members from an additional dinoflagellate and from three stramenopile radiophytes, which all form noxious red tides (Shikata et al., 2019), has been omitted from the figure for clarity. As there is no data available of CRYs from the secondary green line, this line was omitted. The cladogram was modified after Handrich et al. (2017).
Spatiotemporal Expression of Algal Cryptochromes
Expression Profiles in Light–Dark Cycles
The expression of algal cryptochromes was so far mainly studied under light–dark cycles (diurnal conditions). In most cases, it is still unknown if their expression is also controlled by the circadian clock. Algal cryptochromes are routinely expressed in a diurnal way whereby the amplitude of the rhythm and the phase of the peak can vary. The green algal Cr pCRY protein (formerly known as Chlamydomonas photolyase homolog 1, CPH1) is nearly absent during the day, but accumulates strongly (amplitude increase up to tenfold) during the night (Reisdorph and Small, 2004; Müller et al., 2017). Cr pCRY protein abundance seems to be independent of the circadian clock (Müller et al., 2017). The protein is known to be rapidly degraded by light via the proteasome pathway. Transcript levels of Cr pCRY also increase during the night (Kottke et al., 2017), which is also the case for a plant CRY from the chlorophyte Haematococcus pluvialis under the tested high light conditions (Hang et al., 2020). These findings along with the negative regulation of Cr pCRY in gametogenesis (see below) suggests a “dark” function of green algal pCRYs (Müller et al., 2017). In contrast to Cr pCRY, the Cr aCRY protein is rather consistently expressed over the light–dark (LD) cycle (chosen cycle of 12h light:12h darkness), being highest from the beginning until the middle of the day and lowest at the beginning of the night (change in amplitude about 2-fold; Beel et al., 2012; Kottke et al., 2017). The third CRY from Cr, Cr CRY-DASH1 reaches its maximal protein abundance at midday and is lowest at the beginning of the night (change in amplitude about 4-fold; Rredhi et al., 2021). Taken together, the acrophases of peak expression vary with all three investigated Cr CRYs, ranging from early day until late night. For some other algal cryptochromes, only transcript abundances have been determined so far. In the kelp Saccharina japonica (Sj), the maximum transcript level of a CRY-DASH is reached at midday in a LD 12h:12hcycle (Yang et al., 2020). Transcript levels of the marine picoalga Ot CPF1, as determined by microarrays, peak during the day, while levels of Ot CPF2, belonging to the CRY-DASH family, peak at the beginning of the day (Heijde et al., 2010). Also in Pt, all studied transcript levels of different CRYs (studied by qPCR) had rhythmic expression in an LD cycle of 16h:8h with peaks in different acrophases, indicating a possible functional diversification within the diurnal cycle (Oliveri et al., 2014).
Localization of Cryptochromes
Depending on the expression and function, the subcellular localization of CRYs may vary. The green algal Cr aCRY plays a role in vegetative cells as well as in pregametes and gametes (Zou et al., 2017). During daytime (LD4 being 4h after light was switched on in a 12h:12h LD cycle), Cr aCRY is localized to a significant extent in the nucleus of vegetative cells, but is mainly distributed over the cell body (most likely in the cytosol) at late night (LD22 being 10h after light was switched off). In sexual cells (pregametes, gametes, and dark inactivated gametes) Cr aCRY was never found in the nucleus, but always distributed over the cell body. The presence of Cr aCRY in the nucleus of vegetative cells at daytime is in congruence with its activity as (6-4) photolyase, repairing UV-B induced damages (Franz et al., 2018). Thus, Cr aCRY localization varies depending on its function. In contrast, the subcellular localization of Pt CPF1 in the nucleus was not differentially regulated by darkness and light (blue or ultraviolet) treatment (Coesel et al., 2009).
For Cr pCRY, experimental data is lacking, but it is predicted to be localized in the nucleus (Kottke et al., 2017).
In the case of the colony-forming alga Volvox carteri (Vc) that consists of somatic cells and sexually active gonidia, transcript levels were determined for different photoreceptors in the different cell types. Vc aCRY and Vc pCRY were found to accumulate during final cellular differentiation (Kianianmomeni and Hallmann, 2015). Some Vc photoreceptor genes, including Vc pCRY, are highly expressed in the somatic cells. These data give first insights into the possibility that the different CRYs may also be involved in developmental cycles.
As mentioned above, CRY-DASH proteins like CRY3 from A. thaliana have been shown to be localized in organelles; CRY3 is found in chloroplasts and mitochondria (Kleine et al., 2003). Localization studies revealed the presence of CRY-DASH from the dinoflagellate Karenia brevis (Kb) only in chloroplasts (Brunelle et al., 2007). The green algal Cr CRY-DASH1 bearing a chloroplast transit peptide was also found solely in the chloroplast but not in mitochondria, using biochemical fractionation (Rredhi et al., 2021). Its localization is thus in close relation with its biological function, as detailed below.
Localization of the novel type dualchrome was analyzed heterologously in tobacco cells in fusion with GFP. It was primarily found in the nucleus and its localization was not changed by light treatment (Makita et al., 2021).
Biological Functions of Algal Cryptochromes
Although a large repertoire of algal cryptochromes is known (Figure 3), only a few members have been studied in depth regarding their biological function(s). These studies were mostly done in model algal species that can be transformed and where overexpressing lines and knockdown or knockout mutants can be generated. Table 1 summarizes the main functions of these cryptochromes. Like land plant CRYs, algal CRYs are involved in the regulation of several cellular functions.
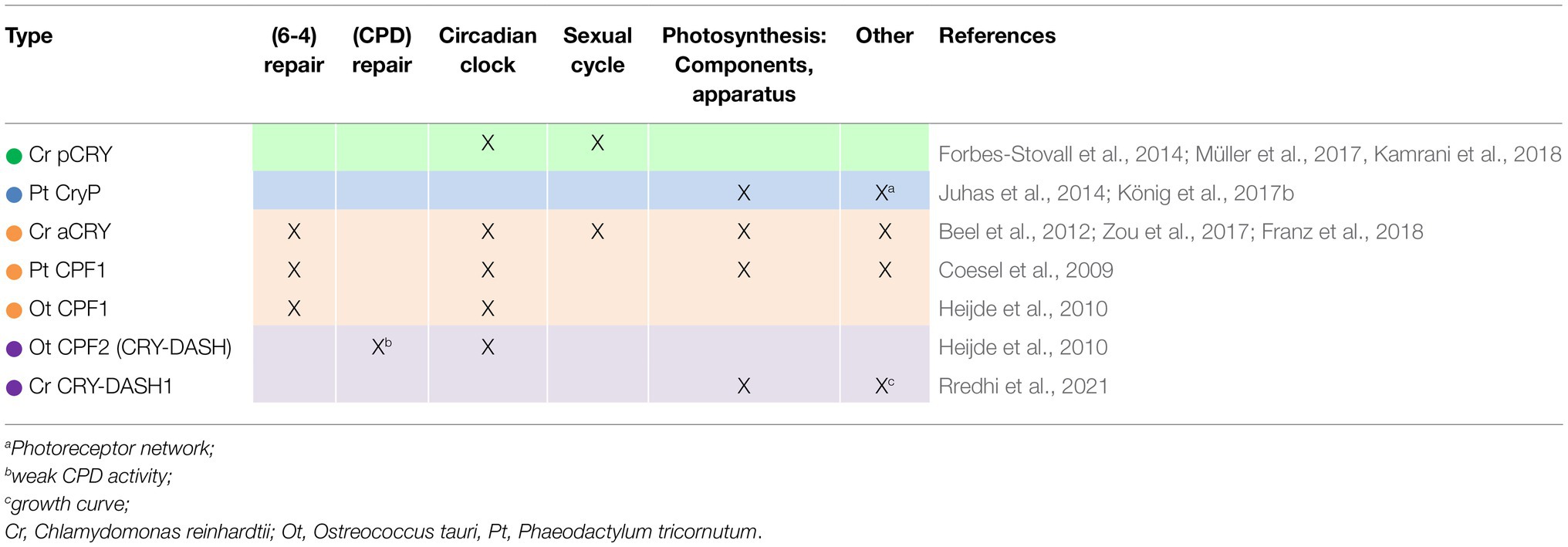
Table 1. Biological functions of algal CRYs, including comparative data from transgenic lines.
Dual Function Cryptochromes: DNA Repair and More
The first evidence for an algal animal-like cryptochrome with repair activity came from a marine diatom. Pt CPF1 was found to be a member of the cryptochrome (6-4) photolyase family. In contrast to other animal and land plant cryptochromes, Pt CPF1 shows (6-4) photoproduct repair activity (Coesel et al., 2009). Using a Pt CPF1-overexpressing line, blue-light regulated transcript levels of wild type were compared to that line. The analyses suggest that Pt CPF1 has a regulatory role in controlling the photosynthetic light harvesting complex and photoprotection as well as cell-cycle progression in addition to its photolyase activity (Coesel et al., 2009). Moreover, a clock function was postulated as detailed in the section below. Similarly, Ot CPF1 from the marine picoalga Ot exerts (6-4) photolyase activity and is involved in circadian rhythms (Heijde et al., 2010). Also the freshwater green alga Cr that lives mainly in wet soil (Sasso et al., 2018) has a dual function cryptochrome, Cr aCRY. It repairs (6-4) photoproducts; a knockdown mutant is less resistant to UV-B treatment than wild type (Franz et al., 2018). In addition, aCRY is involved in regulating transcript levels that are controlled by blue, yellow or red light, including those that encode a light harvesting protein (LHCBM6), glutamine synthetase 1, an enzyme of nitrogen metabolism or the circadian clock component C3, a subunit of the RNA-binding protein CHLAMY1 (Beel et al., 2012).
Algal CRYs and the Biological Clock Machinery
As indicated above, dual function animal-like cryptochromes are also involved in the circadian clock machinery. For Pt and Ot CPF1, the dual function was shown by positively testing their repressor activities in a heterologous system. The transcription factors CLOCK and BMAL were used for this purpose. They form a dimer and are part of the mammalian clock-controlled feedback loop. The inhibition of the CLOCK-BMAL-mediated transcription by Pt and Ot CPF1 was positively verified in COS cells using an E-box bearing luciferase reporter system (Coesel et al., 2009; Heijde et al., 2010). Moreover, Ot CPF1 knockdown lines were generated along with a luciferase reporter that was put under the control of a circadian promoter. In several of these lines, period lengthening (of 1 up to 5h) was observed compared to wild type, and the amplitude was dampened (Heijde et al., 2010). In a Cr aCRY knockdown line, light-induced induction of the C3 subunit of the circadian RNA-binding protein CHLAMY1 that influences the acrophase was altered (Beel et al., 2012). These experiments suggest that dual function CPF1/aCRYs play an important role within the circadian system.
For the green algal Cr pCRY, an involvement in the circadian clock was also found, using knockdown lines and a representative circadian clock-controlled process named photoaccumulation or phototaxis. Detection of this rhythm is automated (Forbes-Stovall et al., 2014; Müller et al., 2017). Phase response curves (PRCs) with blue-light pulses taken from wild type and a Cr pCRY knockdown line revealed the influence of Cr pCRY on PRC behavior. The largest differences in phase resetting of about 10h between wild type and the mutant were observed in early subjective night (Müller et al., 2017). These data suggest a role of pCRY in the entrainment of the circadian clock. Moreover, the Cr pCRY knockdown line exerts a significantly lengthened period of about 27.9h compared to wild type (24.5h in average); it becomes arrhythmic after a few days under free-running conditions that are used to analyze clock properties. It was also suggested that pCRY positively regulates the clock component, ROC75, a putative transcription factor (Kamrani et al., 2018). These data support the model that Cr pCRY is not only involved in circadian input but is also linked to the central oscillator.
CRYs and Life Cycle Regulation
The sexual cycle of the unicellular alga Cr, which is regulated by light and nitrogen availability [reviewed in Kottke et al. (2017); Sasso et al. (2018)], is well studied. Cr vegetative cells of both mating types (mt+ and mt−) turn into pregametes in the dark upon the removal of the nitrogen source. In the presence of light and without nitrogen source, pregametes become gametes. This process is controlled by three photoreceptors. It is promoted by phototropin and inhibited by two algal cryptochromes, aCRY and pCRY (Huang and Beck, 2003; Müller et al., 2017; Zou et al., 2017). It is assumed that the inhibition process allows gametes to reconvert to pregametes and vegetative cells if a nitrogen source becomes available short term. Sexually active gametes finally fuse to form a quadriflagellated cell that is converted to a resilient zygote (Figure 1). Zygotes can survive in the dark and without a nitrogen source for months (Sasso et al., 2018). Upon light and nitrogen availability, germination is induced, and a tetrad of haploid vegetative cells is formed which results in individual vegetative cells. Activation of germination is again mediated by the above-mentioned three photoreceptors (phototropin, aCRY, and pCRY), but in this case all three act in concert as positive regulators (Huang and Beck, 2003; Müller et al., 2017; Zou et al., 2017). Thus, both aCRY and pCRY play central roles in the sexual cycle of Cr either as negative regulators (gamete formation) or as positive regulators (germination). In this context, it is also of interest that blue light is the main trigger for gamete formation and germination. However, red light also has a significant albeit smaller influence on germination (Zou et al., 2017). aCRY with its property to absorb also in the red spectral region in addition to blue (Beel et al., 2012, see also chapter on properties below) seems to be the relevant receptor for red light because pCRY and phototropin do not absorb in this range.
CRYs and the Photosynthetic Apparatus Regulation
CRYs have a broad influence on the photosynthetic machinery by regulating transcript levels and/or protein abundance of photosynthetic components, pigments or the entire photosynthetic machinery. Here, we will present exemplarily the effects of two representative CRYs, the plant-like CryP from Pt and CRY-DASH1 from Cr, but it should be mentioned that also Pt CPF1 and Cr aCRY were shown to be involved in the regulation of transcript levels of photosynthetic compounds (Coesel et al., 2009; Beel et al., 2012).
Plastids and the photosynthetic apparatus of diatoms are different from that of Chloroplastida in several aspects. Diatoms belong to the secondary red endosymbionts (Figure 3). Their plastids are surrounded by four membranes, and they possess additional fucoxanthin pigments that give them an orange, brownish color. Based on a Pt CryP knockdown line, effects of this plant-like CRY on proteins of the photosynthetic apparatus were studied. Here, the most prominent members of the diatom light harvesting chlorophyll-fucoxanthin family (Lhcf) as well as proteins involved in their photoprotection (Lhcx) were analyzed. Lhcf1 -Lhcf11 protein abundance was enhanced in the CryP knockdown strains in comparison to wild type cells (Juhas et al., 2014). Contrarily, the protein level of Lhcx was decreased in the knockdown lines (Juhas et al., 2014). Changes were also observed in transcript levels under slightly different conditions, but were not always in agreement with the protein levels suggesting that posttranscriptional regulation plays a role (König et al., 2017a). Taken together, these data show that CryP contributes significantly to changes in photosynthetically relevant compounds.
Characterization of the green algal Cr CRY-DASH1 protein represents a first functional in-depth study of an algal CRY-DASH protein. The Cr CRY-DASH1 knockout line showed a significantly reduced growth rate (Rredhi et al., 2021) in contrast to Pt CryP knockdown lines that were influenced in photosynthesis components, but had a similar growth rate to wild type (Juhas et al., 2014). Intriguingly, the content of chlorophyll a and b as well as of the carotenoids was increased in the CRY-DASH1 knockout line. This was even visible by the naked eye; cultures of the knockout line were of a darker green than wild type (Rredhi et al., 2021). The increase in pigments went hand in hand with hyper-stacking of thylakoid membranes and an increase in two of the central proteins of photosystem II, D1 and the antenna protein CP43 in the mutant line (Rredhi et al., 2021). CRY-DASH1 thus acts as a repressor that prevents the synthesis of excessive pigments and membranes and thus balances the photosynthetic machinery. Its regulatory role seems to be exerted at the posttranscriptional/translational level as the transcript levels of the genes encoding D1 and CP43 are not altered in the mutant compared to wild type (Rredhi et al., 2021). It is postulated that the observed reduction in growth is due to the higher pigment amount resulting in a shading effect. Indeed, light intensity within a culture flask of the knockout line was reduced compared to that of a wild-type-containing-flask (Rredhi et al., 2021). Notably, Cr CRY-DASH1 absorbs primarily in the UV-A range (see chapter below), where the photosynthetic pigments absorb only to a smaller extent.
Others
The recently described dualchrome (DUC1) represents a chimera consisting of a phytochrome and a cryptochrome in the rather unknown green picoalga Pp (Figure 2). It was found that the Pp genome encodes also further homologue proteins of the model plant A. thaliana involved in light signaling such as phototropin, CONSTITUTIVE PHOTOMORPHOGENIC 1 (COP1), or ELONGATED HYPOCOTYL 5 (HY5) in addition to the mentioned CRY variants (Figure 3). Thus, it possesses a gene set necessary for adapting to various light conditions (Makita et al., 2021).
The presence of several CRYs and other photoreceptors in algae elicits the question of if they form a photoreceptor network influencing each other. Indeed, it was found in Pt CryP knockdown lines that the transcript levels of other photoreceptors (Pt phytochrome and Pt CPF1) are influenced by the reduction of Pt CryP. These data suggest exactly such a scenario (König et al., 2017a).
Biophysical Properties of Algal Cryptochromes
The physiological responses to light are the result of a chain of events on the molecular level. First, light is absorbed by a chromophore bound to the receptor, which then leads to a photochemical reaction. This reaction causes a change in structure and/or conformation of the protein, which then may change the interaction of the receptor with a signaling partner such as a specific binding protein or DNA. Finally, a signal transduction cascade leads to a change in the physiology of cells and organisms.
Light-Induced Oligomerization of Cryptochromes
An interesting aspect of the cryptochrome response to light is that the photochemical reaction of the FAD may cause a change in the oligomerization state, i.e., in the number of identical receptors that associate within a dynamic complex. A formation of large complexes called photobodies of At CRY2 has been observed in plant cells (Mas et al., 2000), which is in agreement with a homooligomerization to clusters of receptors upon blue-light illumination (Bugaj et al., 2013). These clusters are reversible in the dark and dissociate within minutes. Accordingly, the reversible clustering has drawn much attention to the development of optogenetic tools to localize fusion proteins to one spot within mammalian cells by illumination (Bugaj et al., 2013; Taslimi et al., 2014). Recent structural characterization by cryo-electron tomography and X-ray crystallography has revealed that At CRY2 forms tetramers of the PHR upon illumination (Ma et al., 2020; Shao et al., 2020; Palayam et al., 2021).
Oligomerization of algal cryptochromes has been studied on some selected members and light-induced changes in oligomerization have been found. A similar clustering behavior to At CRY2 has been observed for the light-sensitive domain of algal Cr pCRY as a fusion protein labeled with a fluorescence protein. The light-induced degradation of Cr pCRY in C. reinhardtii prevents a study of a potential functional role of this clustering in vivo (Reisdorph and Small, 2004; Müller et al., 2017). However, it has been shown that a complex containing Cr pCRY is formed at the end of the night in C. reinhardtii, possibly in context with its dark-related function. It remains open whether this complex includes homooligomers of pCRY or if it is a heteromeric complex with yet unknown partner(s) (Müller et al., 2017).
The oligomerization state has been characterized for isolated Cr aCRY in great detail. Here, red light induces the transition from a dimer in the dark to a tetramer in the light (Oldemeyer et al., 2016). Interestingly, the presence of the CCT is required for the formation of the dimer in the dark. Truncation of the CCT produces a monomer, which forms a dimer upon illumination only to a small extent (Oldemeyer et al., 2016). Some deviating results on oligomerization states of Cr aCRY can be explained by the different exclusion volumes of the columns used for gel filtration (Franz-Badur et al., 2019).
In summary, light-induced changes in oligomerization state have been observed in algal cryptochromes but differ between pCRY and aCRY. In general, little is known about complex formation of algal cryptochromes with signaling partners or other photoreceptors. These signaling partners need to be identified to link the changes in oligomerization state to the signal transduction cascade.
Strong Variations in the Absorption Spectra Define the Different Roles of Algal Cryptochromes
The absorption spectrum of a photoreceptor in the dark is decisive for the region of the sun’s spectrum, by which this receptor is activated. Ideally, the absorption spectrum matches the action spectrum that was recorded for the function in vivo. The absorption spectrum of cryptochromes reflects the redox state of the FAD cofactor, i.e., whether it binds oxidized FAD (FADox), the anion radical (FAD•−), the neutral radical (FADH•), or fully reduced FAD (FADH−). Different redox states of FAD can be stabilized in the dark by the specific protein environment of a cryptochrome. Accordingly, a cryptochrome acts as a blue and UV-A light receptor, if (FADox) is the stable redox state in the dark.
The interpretation of the absorption spectra of cryptochromes is complicated by the fact that usually the recording is performed after isolation from a heterologous expression system, which might lead to an oxidation of the FAD. In addition, an antenna chromophore is bound to some cryptochromes, which might be lost upon purification or might not even be produced by the heterologous expression system. The antenna molecule, which has a higher extinction coefficient as compared to flavin, serves the purpose to absorb the light and to transfer energy to the FAD cofactor, thereby increasing the probability of light absorption.
The plant cryptochrome Cr pCRY shows a typical spectrum for a blue-light receptor carrying oxidized FAD (FADox) with maxima in the blue and UV-A region (Figure 4A; Immeln et al., 2007). The presence of the CCT does not influence the absorption spectrum (Goett-Zink et al., 2021). Some residual MTHF has been identified after purification from bacterial cells, but a functional role as an antenna chromophore and binding of MTHF in the alga is not likely. FADox in pCRY is even present in the strongly reducing cytosol of living Escherichia coli cells (Goett-Zink et al., 2021), which is a strong argument for FADox being the native chromophore of pCRY in Chlamydomonas in the dark similar to findings for pCRYs from land plants (Bouly et al., 2007).
Members of the animal-like CRY/CPF1 family such as Ot CPF1, Pt CPF1 and Cr aCRY bind FADox after purification (Coesel et al., 2009; Heijde et al., 2010; Beel et al., 2012). The more unusual was the finding that Cr aCRY uses the flavin neutral radical (FADH•) as the dark state for a sensory function thereby accessing almost the complete visible spectral region including yellow and red light (Beel et al., 2012; Figure 4A). This fact suggests that FADH• is the predominant form of FAD in the alga in the dark, which is supported by a very efficient photoactivation reaction from FADox to FADH• in vitro (Lacombat et al., 2019). The tight binding of an antenna molecule is another difference of aCRY/CPF1 to plant cryptochromes. The binding of 8-HDF was suggested for Ot CPF1 from structural considerations (Brazard et al., 2012) and then identified for Cr aCRY after implementing the synthesis of 8-HDF in E. coli (Franz et al., 2018). The antenna 8-HDF strongly absorbs light in the blue region of the visible spectrum (Figure 4A) and additionally stabilizes the FADH• in the dark, preventing oxidation (Oldemeyer et al., 2020).
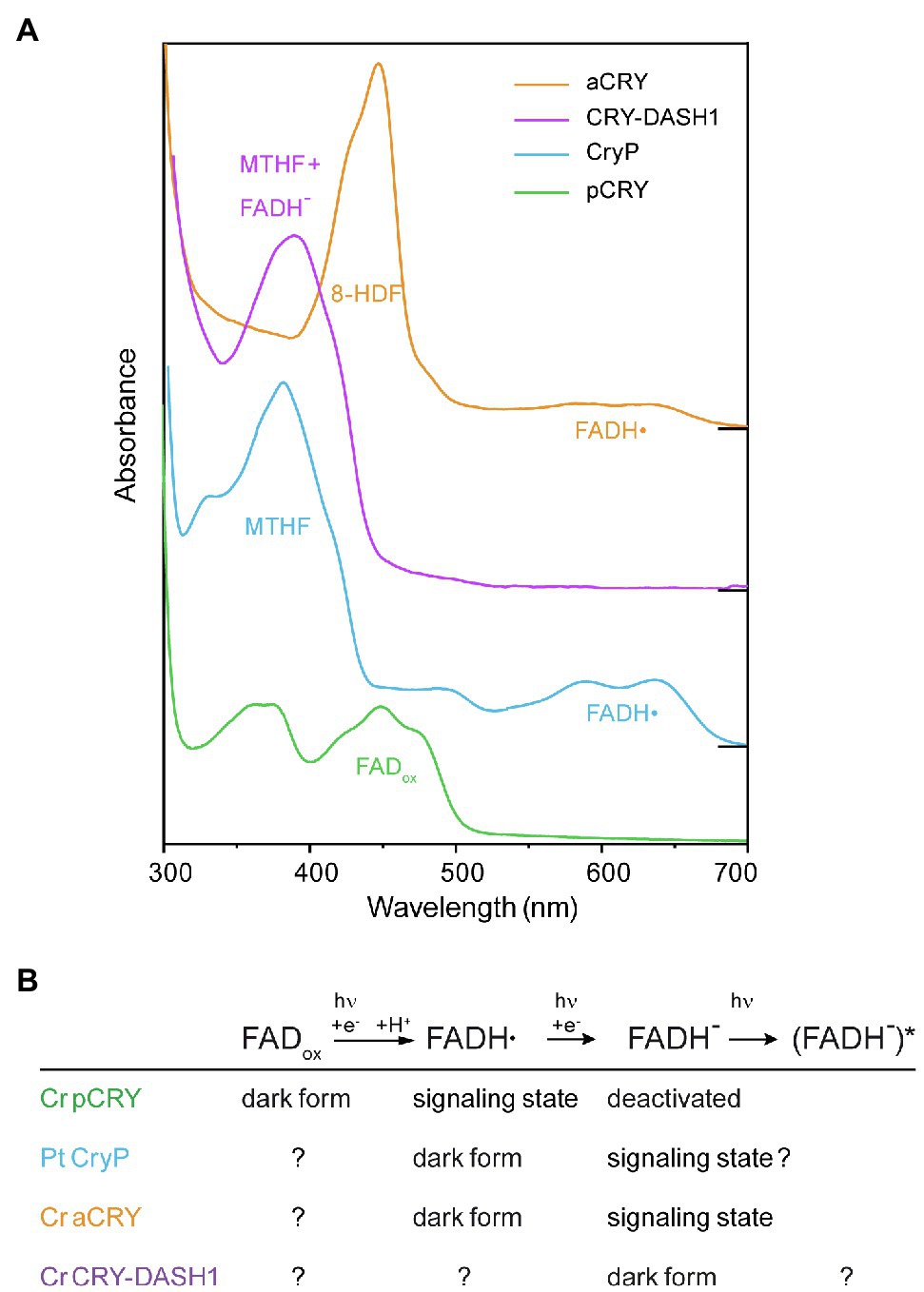
Figure 4. Absorption spectra of different cryptochromes from algae recorded in vitro directly after isolation and the postulated roles of the redox states of FAD in these cryptochromes. (A) The absorption spectra of cryptochromes are characterized by the different oxidation states of the FAD chromophore and the binding of antenna molecules. The plant cryptochrome Cr pCRY binds FADox and acts as blue-light receptor. Plant-like cryptochrome Pt CryP and animal-like cryptochrome Cr aCRY both absorb almost in the full visible spectral region because of the absorption of FADH•. However, Pt CryP binds MTHF as antenna, whereas Cr aCRY binds 8-HDF leading to different absorption maxima in the UV-A and blue region, respectively. The UV-A-receptor Cr CRY-DASH1 absorbs mainly at around 380nm dominated by the contribution of MTHF, whereas its chromophore FADH− absorbs only weakly. For simplicity, the absorption of FADox, FADH• and FADH− is indicated at the maximum with highest wavelength, but all FAD species also contribute to the absorption at lower wavelengths. Spectra were displaced vertically for better visibility. (B) The redox states of FAD play different roles in the algal cryptochromes, as postulated on the basis of the absorption spectra and functional studies of representative members. The dark form indicates the redox state present in vivo in the dark. Light absorption induces a reduction of the FAD. The signaling state is associated with conformational changes and/or changes in oligomerization state of the receptor that drive signal transduction.
Much less is known about the plant-like subfamily and its member Pt CryP. The binding of stable FADH• and MTHF in the dark after purification might indicate a light response similar to the aCRY family, with the distinct difference that the antenna MTHF absorbs shifted to the UV-A region compared to 8-HDF (Figure 4A; Juhas et al., 2014). Functional studies have focused so far only on the blue-light effects of CryP on Pt physiology (König et al., 2017a). It would be of high interest to study also potential responses of Pt CryP to green and red light.
The large subfamily of CRY-DASH proteins (Brudler et al., 2003) is also well represented with several members in algal genomes (Figure 3). Few members have been isolated and characterized by absorption spectroscopy. A typical behavior for members of CRY-DASH has been found for Ot CPF2 in vitro, which showed a dominant absorbance by the antenna MTHF and some contribution from FADox (Heijde et al., 2010). Similar findings have been presented for Cm PHR5 (Asimgil and Kavakli, 2012). Recent investigations on Cr CRY-DASH1 revealed that FAD is present in the fully reduced state (FADH−) directly after purification, which together with the photochemical results indicates a presence of FADH− in Cr in vivo (Rredhi et al., 2021). As a result, Cr CRY-DASH1 mainly absorbs light in the UV-A-region (Figure 4A), supported by functional studies as a UV-A receptor in Cr. Of note, the UV-A absorption of CRY-DASH1 is maximal in a spectral region where the typical blue-light receptors pCRY and phototropin as well as photosystems I and II show only weak absorption. Accordingly, CRY-DASH1 may complement the other receptors in the alga by the spectral range in which it is activated.
In summary, the strong differences in the absorption spectra of cryptochromes in the dark reflect the variety of functions for which these receptor subfamilies evolved (Kottke et al., 2017; Figure 4B). Cryptochromes act in algae as UV-A receptor (Cr CRY-DASH1), as UV-A/blue-light receptor (Cr pCRY) and as “white light” receptor covering almost the full spectral region (Cr aCRY), defined by the redox state of FAD cofactor in the dark. The FAD is reduced after the absorption of light and forms the signaling state, accompanied by conformational changes and/or changes in oligomerization state of the receptor. The identification of the signaling state and the mechanism of signal propagation within the receptor are important biophysical questions to be answered for each member. The details of the different pathways identified to date go beyond the scope of this review. Even more differences in the light responses will be revealed as more and more candidates of algal cryptochromes are being characterized biophysically.
Outlook
It is expected that metagenome analysis from phytoplankton will lead to the discovery of even more novel types of photoreceptors, including new cryptochrome variants such as the recently identified DUC1. The high number of different light sensors in aquatic organisms is correlated with the fact that light quality and light quantity for aquatic algae change dynamically. The light conditions depend on the depth within the water column where the algae occur at a given time in the ocean, on the presence of solved particles and on other light-absorbing organisms at this location. The challenge will be to analyze the role of all these photoreceptors and their mutual interaction partners in the in vivo systems.
Algae represent a rich source for novel cryptochrome photoreceptors. These might be used as optogenetic tools to generate light switches in algal cells for controlling gene expression or in combination with signaling molecules such as cAMP or Ca2+ (Kianianmomeni and Hallmann, 2014). They might trigger light-dependent adaptations of organismal physiology, development, and behavior in nature (Losi et al., 2018) and thus create potent tools for biotechnological approaches. They may also be used in heterologous systems for neurobiology and in medicine, as it was recently done with an algal rhodopsin to partly recover vision in blind people (Sahel et al., 2021). The land plant At CRY2 was already used for optogenetical protein clustering (Park et al., 2017) and for regulating gene expression in mammalian cells (Hernández-Candia et al., 2019). In general, the high variety of algal CRYs offers a high potential for engineering new sensor tools.
Currently, we are only beginning to understand the biophysical properties and the involved signal transduction pathways as well as the biological functions of the different algal cryptochromes. Their interplay within one algal system and along with the other photoreceptors of this system remains largely enigmatic, even in the today selected model algal organisms. Double-, triple-, or even higher-order mutants need to be produced to gain further insights in their complex photobiology. Yet, the basis of these light signaling events is the key to understand (i) algal fitness and thus (ii) their central contribution as photosynthetic organisms to life on Earth. As our knowledge on the role and function of algal cryptochromes improves, it will be possible to manipulate growth, reproduction and the development of algae as well as to optimize photosynthesis, for example by application of targeted illumination protocols.
Author Contributions
JP, AR, JS, and MM designed and drew Figures 1–3. SO and TK created Figure 4. JP and MM arranged Table 1. JP, AR, TK, and MM wrote the paper, with input from all authors. All authors contributed to the article and approved the submitted version.
Funding
Our work was supported by the Deutsche Forschungsgemeinschaft by DFG grants Mi373/16-1 to MM and Ko3580/4-2 and Ko3580/7-1 to TK. SO was funded by the DFG Sonderforschungsbereich SFB1078.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s Note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Acknowledgments
We thank Claudia Büchel, Ellen McPherson and the referees for helpful comments on the review.
References
Adl, S. M., Simpson, A. G. B., Lane, C. E., Lukeš, J., Bass, D., Bowser, S. S., et al. (2012). The revised classification of eukaryotes. J. Eukaryot. Microbiol. 59, 429–514. doi: 10.1111/j.1550-7408.2012.00644.x
Ahmad, M. (2016). Photocycle and signaling mechanisms of plant cryptochromes. Curr. Opin. Plant Biol. 33, 108–115. doi: 10.1016/j.pbi.2016.06.013
Ahmad, M., and Cashmore, A. R. (1993). HY4 gene of A. thaliana encodes a protein with characteristics of a blue-light photoreceptor. Nature 366, 162–166. doi: 10.1038/366162a0
Asimgil, H., and Kavakli, I. H. (2012). Purification and characterization of five members of photolyase/cryptochrome family from Cyanidioschyzon merolae. Plant Sci. 185–186, 190–198. doi: 10.1016/j.plantsci.2011.10.005
Beel, B., Prager, K., Spexard, M., Sasso, S., Weiss, D., Müller, N., et al. (2012). A flavin binding cryptochrome photoreceptor responds to both blue and red light in Chlamydomonas reinhardtii. Plant Cell 24, 2992–3008. doi: 10.1105/tpc.112.098947
Bouly, J. P., Schleicher, E., Dionisio-Sese, M., Vandenbussche, F., Van Der Straeten, D., Bakrim, N., et al. (2007). Cryptochrome blue light photoreceptors are activated through interconversion of flavin redox states. J. Biol. Chem. 282, 9383–9391. doi: 10.1074/jbc.M609842200
Bowman, J. L., Floyd, S. K., and Sakakibara, K. (2007). Green genes-comparative genomics of the green branch of life. Cell 129, 229–234. doi: 10.1016/j.cell.2007.04.004
Brawley, S. H., Blouin, N. A., Ficko-Blean, E., Wheeler, G. L., Lohr, M., Goodson, H. V., et al. (2017). Insights into the red algae and eukaryotic evolution from the genome of Porphyra umbilicalis (Bangiophyceae, Rhodophyta). Proc. Natl. Acad. Sci. U. S. A. 114, E6361–E6370. doi: 10.1073/pnas.1703088114
Brazard, J., Ley, C., Lacombat, F., Plaza, P., Mony, L., Heijde, M., et al. (2012). Photoantenna in two cryptochrome-photolyase proteins from O. tauri: presence, nature and ultrafast photoinduced dynamics. J. Photochem. Photobiol. A Chem. 234, 135–145. doi: 10.1016/j.jphotochem.2012.01.012
Brudler, R., Hitomi, K., Daiyasu, H., Toh, H., Kucho, K. I., Ishiura, M., et al. (2003). Identification of a new cryptochrome class: structure, function, and evolution. Mol. Cell 11, 59–67. doi: 10.1016/S1097-2765(03)00008-X
Brunelle, S. A., Hazard, E. S., Sotka, E. E., and Van Dolah, F. M. (2007). Characterization of a dinoflagellate cryptochrome blue-light receptor with a possible role in circadian control of the cell cycle. J. Phycol. 43, 509–518. doi: 10.1111/j.1529-8817.2007.00339.x
Bugaj, L. J., Choksi, A. T., Mesuda, C. K., Kane, R. S., and Schaffer, D. V. (2013). Optogenetic protein clustering and signaling activation in mammalian cells. Nat. Methods 10, 249–252. doi: 10.1038/nmeth.2360
Chaves, I., Pokorny, R., Byrdin, M., Hoang, N., Ritz, T., Brettel, K., et al. (2011). The cryptochromes: blue light photoreceptors in plants and animals. Annu. Rev. Plant Biol. 62, 335–364. doi: 10.1146/annurev-arplant-042110-103759
Coesel, S. N., Durham, B. P., Groussman, R. D., Hu, S. K., Caron, D. A., Morales, R. L., et al. (2021). Diel transcriptional oscillations of light-sensitive regulatory elements in open-ocean eukaryotic plankton communities. Proc. Natl. Acad. Sci. 118:e2011038118. doi: 10.1073/pnas.2011038118
Coesel, S., Mangogna, M., Ishikawa, T., Heijde, M., Rogato, A., Finazzi, G., et al. (2009). Diatom PtCPF1 is a new cryptochrome/photolyase family member with DNA repair and transcription regulation activity. EMBO Rep. 10, 655–661. doi: 10.1038/embor.2009.59
Cross, F. R., and Umen, J. G. (2015). The Chlamydomonas cell cycle. Plant J. 82, 370–392. doi: 10.1111/tpj.12795
Deng, Y., Yao, J., Wang, X., Guo, H., and Duan, D. (2012). Transcriptome sequencing and comparative analysis of Saccharina japonica (Laminariales, phaeophyceae) under blue light induction. PLoS One 7:e39704. doi: 10.1371/journal.pone.0039704
Eberhard, S., Finazzi, G., and Wollman, F. A. (2008). The dynamics of photosynthesis. Annu. Rev. Genet. 42, 463–515. doi: 10.1146/annurev.genet.42.110807.091452
Emery, P., So, W. V., Kaneko, M., Hall, J. C., and Rosbash, M. (1998). Cry, a Drosophila clock and light-regulated cryptochrome, is a major contributor to circadian rhythm resetting and photosensitivity. Cell 95, 669–679. doi: 10.1016/S0092-8674(00)81637-2
Fankhauser, C., and Christie, J. M. (2015). Plant phototropic growth. Curr. Biol. 25, R384–R389. doi: 10.1016/j.cub.2015.03.020
Field, C. B., Behrenfeld, M. J., Randerson, J. T., and Falkowski, P. (1998). Primary production of the biosphere: integrating terrestrial and oceanic components. Science 281, 237–240. doi: 10.1126/science.281.5374.237
Forbes-Stovall, J., Howton, J., Young, M., Davis, G., Chandler, T., Kessler, B., et al. (2014). Chlamydomonas reinhardtii strain CC-124 is highly sensitive to blue light in addition to green and red light in resetting its circadian clock, with the blue-light photoreceptor plant cryptochrome likely acting as negative modulator. Plant Physiol. Biochem. 75, 14–23. doi: 10.1016/j.plaphy.2013.12.002
Fortunato, A. E., Annunziata, R., Jaubert, M., Bouly, J.-P., and Falciatore, A. (2015). Dealing with light: the widespread and multitasking cryptochrome/photolyase family in photosynthetic organisms. J. Plant Physiol. 172, 42–54. doi: 10.1016/j.jplph.2014.06.011
Franz, S., Ignatz, E., Wenzel, S., Zielosko, H., Ngurah Putu, E. P. G., Maestre-Reyna, M., et al. (2018). Structure of the bifunctional cryptochrome aCRY from Chlamydomonas reinhardtii. Nucleic Acids Res. 46, 8010–8022. doi: 10.1093/nar/gky621
Franz-Badur, S., Penner, A., Straß, S., von Horsten, S., Linne, U., and Essen, L. O. (2019). Structural changes within the bifunctional cryptochrome/photolyase CraCRY upon blue light excitation. Sci. Rep. 9:9896. doi: 10.1038/s41598-019-45885-7
Gentil, J., Hempel, F., Moog, D., Zauner, S., and Maier, U. G. (2017). Review: origin of complex algae by secondary endosymbiosis: a journey through time. Protoplasma 254, 1835–1843. doi: 10.1007/s00709-017-1098-8
Goett-Zink, L., Toschke, A. L., Petersen, J., Mittag, M., and Kottke, T. (2021). C-terminal extension of a plant cryptochrome dissociates from the β-sheet of the flavin-binding domain. J. Phys. Chem. Lett. 12, 5558–5563. doi: 10.1021/acs.jpclett.1c00844
Greiner, A., Kelterborn, S., Evers, H., Kreimer, G., Sizova, I., and Hegemann, P. (2017). Targeting of photoreceptor genes in Chlamydomonas reinhardtii via zinc-finger nucleases and CRISPR/Cas9. Plant Cell 29, 2498–2518. doi: 10.1105/tpc.17.00659
Handrich, M., de Vries, J., Gould, S. B., Serôdio, J., and Christa, G. (2017). Ulvophyceaen photophysiology and research opportunities. Perspect. Phycol. 4, 83–92. doi: 10.1127/pip/2017/0074
Hang, W., Gujar, A., Zhang, H., Xu, W., Zhao, C., Zhu, X., et al. (2020). Cloning, expression, and characterization of a novel plant type cryptochrome gene from the green alga Haematococcus pluvialis. Protein Expr. Purif. 172:105633. doi: 10.1016/j.pep.2020.105633
Hegemann, P. (2008). Algal sensory photoreceptors. Annu. Rev. Plant Biol. 59, 167–189. doi: 10.1146/annurev.arplant.59.032607.092847
Hegemann, P., and Nagel, G. (2013). From channelrhodopsins to optogenetics. EMBO Mol. Med. 5, 173–176. doi: 10.1002/emmm.201202387
Heijde, M., Zabulon, G., Corellou, F., Ishikawa, T., Brazard, J., Usman, A., et al. (2010). Characterization of two members of the cryptochrome/photolyase family from Ostreococcus tauri provides insights into the origin and evolution of cryptochromes. Plant Cell Environ. 33, 1614–1626. doi: 10.1111/j.1365-3040.2010.02168.x
Hernández-Candia, C. N., Wysoczynski, C. L., and Tucker, C. L. (2019). Advances in optogenetic regulation of gene expression in mammalian cells using cryptochrome 2 (CRY2). Methods 164–165, 81–90. doi: 10.1016/j.ymeth.2019.03.011
Hoang, N., Schleicher, E., Kacprzak, S., Bouly, J. P., Picot, M., Wu, W., et al. (2008). Human and Drosophila cryptochromes are light activated by flavin photoreduction in living cells. PLoS Biol. 6:e160. doi: 10.1371/journal.pbio.0060160
Huang, K., and Beck, C. F. (2003). Phototropin is the blue-light receptor that controls multiple steps in the sexual life cycle of the green alga Chlamydomonas reinhardtii. Proc. Natl. Acad. Sci. U. S. A. 100, 6269–6274. doi: 10.1073/pnas.0931459100
Immeln, D., Schlesinger, R., Heberle, J., and Kottke, T. (2007). Blue light induces radical formation and autophosphorylation in the light-sensitive domain of Chlamydomonas cryptochrome. J. Biol. Chem. 282, 21720–21728. doi: 10.1074/jbc.M700849200
Iseki, M., Matsunaga, S., Murakami, A., Ohno, K., Shiga, K., Yoshida, K., et al. (2002). A blue-light-activated adenylyl cyclase mediates photoavoidance in Euglena gracilis. Nature 415, 1047–1051. doi: 10.1038/4151047a
Ito, S., Murakami, A., Iseki, M., Takahashi, T., Higashi, S., and Watanabe, M. (2010). Differentiation of photocycle characteristics of flavin-binding BLUF domains of α-And β-subunits of photoactivated adenylyl cyclase of Euglena gracilis. Photochem. Photobiol. Sci. 9, 1327–1335. doi: 10.1039/c0pp00130a
Jaubert, M., Bouly, J.-P., Ribera d’Alcalà, M., and Falciatore, A. (2017). Light sensing and responses in marine microalgae. Curr. Opin. Plant Biol. 37, 70–77. doi: 10.1016/j.pbi.2017.03.005
Juhas, M., von Zadow, A., Spexard, M., Schmidt, M., Kottke, T., and Büchel, C. (2014). A novel cryptochrome in the diatom Phaeodactylum tricornutum influences the regulation of light-harvesting protein levels. FEBS J. 281, 2299–2311. doi: 10.1111/febs.12782
Kamrani, Y. Y., Matsuo, T., Mittag, M., and Minagawa, J. (2018). ROC75 is an attenuator for the circadian clock that controls LHCSR3 expression. Plant Cell Physiol. 59, 2602–2607. doi: 10.1093/pcp/pcy179
Keeling, P. J., Burger, G., Durnford, D. G., Lang, B. F., Lee, R. W., Pearlman, R. E., et al. (2005). The tree of eukaryotes. Trends Ecol. Evol. 20, 670–676. doi: 10.1016/j.tree.2005.09.005
Kianianmomeni, A., and Hallmann, A. (2014). Algal photoreceptors: in vivo functions and potential applications. Planta 239, 1–26. doi: 10.1007/s00425-013-1962-5
Kianianmomeni, A., and Hallmann, A. (2015). Transcriptional analysis of Volvox photoreceptors suggests the existence of different cell-type specific light-signaling pathways. Curr. Genet. 61, 3–18. doi: 10.1007/s00294-014-0440-3
Kim, W. Y., Fujiwara, S., Suh, S. S., Kim, J., Kim, Y., Han, L., et al. (2007). ZEITLUPE is a circadian photoreceptor stabilized by GIGANTEA in blue light. Nature 449, 356–360. doi: 10.1038/nature06132
Kiontke, S., Göbel, T., Brych, A., and Batschauer, A. (2020). DASH-type cryptochromes – solved and open questions. Biol. Chem. 401, 1487–1493. doi: 10.1515/hsz-2020-0182
Kleine, T., Lockhart, P., and Batschauer, A. (2003). An Arabidopsis protein closely related to Synechocystis cryptochrome is targeted to organelles. Plant J. 35, 93–103. doi: 10.1046/j.1365-313X.2003.01787.x
König, S., Eisenhut, M., Bräïutigam, A., Kurz, S., Weber, A. P. M., and Büchel, C. (2017a). The influence of a cryptochrome on the gene expression profile in the diatom Phaeodactylum tricornutum under blue light and in darkness. Plant Cell Physiol. 58, 1914–1923. doi: 10.1093/pcp/pcx127
König, S., Juhas, M., Jäger, S., Kottke, T., and Büchel, C. (2017b). The cryptochrome—photolyase protein family in diatoms. J. Plant Physiol. 217, 15–19. doi: 10.1016/j.jplph.2017.06.015
Kottke, T., Oldemeyer, S., Wenzel, S., Zou, Y., and Mittag, M. (2017). Cryptochrome photoreceptors in green algae: unexpected versatility of mechanisms and functions. J. Plant Physiol. 217, 4–14. doi: 10.1016/j.jplph.2017.05.021
Kroth, P. G., Wilhelm, C., and Kottke, T. (2017). An update on aureochromes: phylogeny – mechanism – function. J. Plant Physiol. 217, 20–26. doi: 10.1016/j.jplph.2017.06.010
Kutomi, O., Yamamoto, R., Hirose, K., Mizuno, K., Nakagiri, Y., Imai, H., et al. (2021). A dynein-associated photoreceptor protein prevents ciliary acclimation to blue light. Sci. Adv. 7:eabf3621. doi: 10.1126/sciadv.abf3621
Lacombat, F., Espagne, A., Dozova, N., Plaza, P., Müller, P., Brettel, K., et al. (2019). Ultrafast oxidation of a tyrosine by proton-coupled electron transfer promotes light activation of an animal-like cryptochrome. J. Am. Chem. Soc. 141, 13394–13409. doi: 10.1021/jacs.9b03680
Lin, C., and Shalitin, D. (2003). Cryptochrome structure and signal transduction. Annu. Rev. Plant Biol. 54, 469–496. doi: 10.1146/annurev.arplant.54.110901.160901
Losi, A., Gardner, K. H., and Möglich, A. (2018). Blue-light receptors for optogenetics. Chem. Rev. 118, 10659–10709. doi: 10.1021/acs.chemrev.8b00163
Luck, M., and Hegemann, P. (2017). The two parallel photocycles of the Chlamydomonas sensory photoreceptor histidine kinase rhodopsin 1. J. Plant Physiol. 217, 77–84. doi: 10.1016/j.jplph.2017.07.008
Ma, L., Guan, Z., Wang, Q., Yan, X., Wang, J., Wang, Z., et al. (2020). Structural insights into the photoactivation of Arabidopsis CRY2. Nat. Plants 6, 1432–1438. doi: 10.1038/s41477-020-00800-1
Makita, Y., Suzuki, S., Fushimi, K., Shimada, S., Suehisa, A., Hirata, M., et al. (2021). Identification of a dual orange/far-red and blue light photoreceptor from an oceanic green picoplankton. Nat. Commun. 12:3593. doi: 10.1038/s41467-021-23741-5
Mas, P., Devlin, P. F., Panda, S., and Kay, S. A. (2000). Functional interaction of phytochrome B and cryptochrome 2. Nature 408, 207–211. doi: 10.1038/35041583
McCutcheon, J., Lutz, S., Williamson, C., Cook, J. M., Tedstone, A. J., Vanderstraeten, A., et al. (2021). Mineral phosphorus drives glacier algal blooms on the Greenland ice sheet. Nat. Commun. 12:570. doi: 10.1038/s41467-020-20627-w
Müller, N., Wenzel, S., Zou, Y., Künzel, S., Sasso, S., Weiß, D., et al. (2017). A plant cryptochrome controls key features of the Chlamydomonas circadian clock and its life cycle. Plant Physiol. 174, 185–201. doi: 10.1104/pp.17.00349
Navarro, E., Niemann, N., Kock, D., Dadaeva, T., Gutiérrez, G., Engelsdorf, T., et al. (2020). The DASH-type cryptochrome from the fungus Mucor circinelloides is a canonical CPD-photolyase. Curr. Biol. 30, 4483.e4–4490.e4. doi: 10.1016/j.cub.2020.08.051
Okamura, H., Miyake, S., Sumi, Y., Yamaguchi, S., Yasui, A., Muijtjens, M., et al. (1999). Photic induction of mPer1 and mPer2 in cry-deficient mice lacking a biological clock. Science 286, 2531–2534. doi: 10.1126/science.286.5449.2531
Oldemeyer, S., Franz, S., Wenzel, S., Essen, L.-O., Mittag, M., and Kottke, T. (2016). Essential role of an unusually long-lived tyrosyl radical in the response to red light of the animal-like cryptochrome aCRY. J. Biol. Chem. 291, 14062–14071. doi: 10.1074/jbc.M116.726976
Oldemeyer, S., Haddad, A. Z., and Fleming, G. R. (2020). Interconnection of the antenna pigment 8-HDF and flavin facilitates red-light reception in a bifunctional animal-like cryptochrome. Biochemistry 59, 594–604. doi: 10.1021/acs.biochem.9b00875
Oliveri, P., Fortunato, A. E., Petrone, L., Ishikawa-Fujiwara, T., Kobayashi, Y., Todo, T., et al. (2014). The cryptochrome/photolyase family in aquatic organisms. Mar. Genomics 14, 23–37. doi: 10.1016/j.margen.2014.02.001
Palayam, M., Ganapathy, J., Guercio, A. M., Tal, L., Deck, S. L., and Shabek, N. (2021). Structural insights into photoactivation of plant cryptochrome-2. Commun. Biol. 4:28. doi: 10.1038/s42003-020-01531-x
Park, H., Kim, N. Y., Lee, S., Kim, N., Kim, J., and Do Heo, W. (2017). Optogenetic protein clustering through fluorescent protein tagging and extension of CRY2. Nat. Commun. 8:30. doi: 10.1038/s41467-017-00060-2
Parker, M. S., Mock, T., and Armbrust, E. V. (2008). Genomic insights into marine microalgae. Annu. Rev. Genet. 42, 619–645. doi: 10.1146/annurev.genet.42.110807.091417
Petroutsos, D., Tokutsu, R., Maruyama, S., Flori, S., Greiner, A., Magneschi, L., et al. (2016). A blue-light photoreceptor mediates the feedback regulation of photosynthesis. Nature 537, 563–566. doi: 10.1038/nature19358
Podolec, R., Demarsy, E., and Ulm, R. (2021). Perception and signaling of ultraviolet-B radiation in plants. Annu. Rev. Plant Biol. 72, 793–822. doi: 10.1146/annurev-arplant-050718-095946
Pokorny, R., Klar, T., Hennecke, U., Carell, T., Batschauer, A., and Essen, L. O. (2008). Recognition and repair of UV lesions in loop structures of duplex DNA by DASH-type cryptochrome. Proc. Natl. Acad. Sci. U. S. A. 105, 21023–21027. doi: 10.1073/pnas.0805830106
Reisdorph, N. A., and Small, G. D. (2004). The CPH1 gene of Chlamydomonas reinhardtii encodes two forms of cryptochrome whose levels are controlled by light-induced proteolysis. Plant Physiol. 134, 1546–1554. doi: 10.1104/pp.103.031930
Rockwell, N. C., and Lagarias, J. C. (2020). Phytochrome evolution in 3D: deletion, duplication, and diversification. New Phytol. 225, 2283–2300. doi: 10.1111/nph.16240
Rockwell, N. C., Martin, S. S., and Lagarias, J. C. (2012). Red/green cyanobacteriochromes: sensors of color and power. Biochemistry 51, 9667–9677. doi: 10.1021/bi3013565
Rredhi, A., Petersen, J., Schubert, M., Li, W., Oldemeyer, S., Li, W., et al. (2021). DASH cryptochrome 1, a UV-A receptor, balances the photosynthetic machinery of Chlamydomonas reinhardtii. New Phytol. 232, 610–624. doi: 10.1111/nph.17603
Sahel, J. A., Boulanger-Scemama, E., Pagot, C., Arleo, A., Galluppi, F., Martel, J. N., et al. (2021). Partial recovery of visual function in a blind patient after optogenetic therapy. Nat. Med. 27, 1223–1229. doi: 10.1038/s41591-021-01351-4
Sancar, A. (2003). Structure and function of DNA photolyase and cryptochrome blue-light photoreceptors. Chem. Rev. 103, 2203–2237. doi: 10.1021/cr0204348
Sasso, S., Stibor, H., Mittag, M., and Grossman, A. R. (2018). The natural history of model organisms from molecular manipulation of domesticated Chlamydomonas reinhardtii to survival in nature. eLife 7:e39233. doi: 10.7554/eLife.39233
Selby, C. P., and Sancar, A. (2006). A cryptochrome/photolyase class of enzymes with single-stranded DNA-specific photolyase activity. Proc. Natl. Acad. Sci. U. S. A. 103, 17696–17700. doi: 10.1073/pnas.0607993103
Shao, K., Zhang, X., Li, X., Hao, Y., Huang, X., Ma, M., et al. (2020). The oligomeric structures of plant cryptochromes. Nat. Struct. Mol. Biol. 27, 480–488. doi: 10.1038/s41594-020-0420-x
Shikata, T., Takahashi, F., Nishide, H., Shigenobu, S., Kamei, Y., Sakamoto, S., et al. (2019). RNA-seq analysis reveals genes related to photoreception, nutrient uptake, and toxicity in a noxious red-tide raphidophyte Chattonella antiqua. Front. Microbiol. 10:1764. doi: 10.3389/fmicb.2019.01764
Sibbald, S. J., and Archibald, J. M. (2020). Genomic insights into plastid evolution. Genome Biol. Evol. 12, 978–990. doi: 10.1093/gbe/evaa096
Steele, J. H. (ed.) (1974). “Marine food webs” in The Structure of Marine Ecosystems (Cambridge MA, USA: Harvard University Press), 9–28.
Tagua, V. G., Pausch, M., Eckel, M., Gutiérrez, G., Miralles-Durán, A., Sanz, C., et al. (2015). Fungal cryptochrome with DNA repair activity reveals an early stage in cryptochrome evolution. Proc. Natl. Acad. Sci. U. S. A. 112, 15130–15135. doi: 10.1073/pnas.1514637112
Taslimi, A., Vrana, J. D., Chen, D., Borinskaya, S., Mayer, B. J., Kennedy, M. J., et al. (2014). An optimized optogenetic clustering tool for probing protein interaction and function. Nat. Commun. 5:4925. doi: 10.1038/ncomms5925
Tian, Y., Nagel, G., and Gao, S. (2021). An engineered membrane-bound guanylyl cyclase with light-switchable activity. BMC Biol. 19:54. doi: 10.1186/s12915-021-00978-6
Todo, T. (1999). Functional diversity of the DNA photolyase/blue light receptor family. Mutat. Res. 434, 89–97. doi: 10.1016/S0921-8777(99)00013-0
Trippens, J., Greiner, A., Schellwat, J., Neukam, M., Rottmann, T., Lu, Y., et al. (2012). Phototropin influence on eyespot development and regulation of phototactic behavior in Chlamydomonas reinhardtii. Plant Cell 24, 4687–4702. doi: 10.1105/tpc.112.103523
Wang, Q., and Lin, C. (2020). Mechanisms of cryptochrome-mediated photoresponses in plants. Annu. Rev. Plant Biol. 71, 103–129. doi: 10.1146/annurev-arplant-050718-100300
Yang, X., Li, L., Wang, X., Yao, J., and Duan, D. (2020). Non-coding RNAs participate in the regulation of CRY-DASH in the growth and early development of Saccharina japonica (Laminariales, Phaeophyceae). Int. J. Mol. Sci. 21:309. doi: 10.3390/ijms21010309
Zhang, X., Zheng, Z., He, Y., Liu, L., Qu, C., and Miao, J. (2020). Molecular cloning and expression of a cryptochrome gene CiCRY-DASH1 from the antarctic microalga Chlamydomonas sp. ICE-L. Mol. Biotechnol. 62, 91–103. doi: 10.1007/s12033-019-00225-y
Keywords: blue-light receptor, Chlamydomonas, flavin, Ostreococcus, Phaeodactylum, photoreceptor, photosynthetic microorganisms
Citation: Petersen J, Rredhi A, Szyttenholm J, Oldemeyer S, Kottke T and Mittag M (2021) The World of Algae Reveals a Broad Variety of Cryptochrome Properties and Functions. Front. Plant Sci. 12:766509. doi: 10.3389/fpls.2021.766509
Edited by:
Margaret Ahmad, Université Paris-Sorbonne, FranceReviewed by:
Oren Levy, Bar-Ilan University, IsraelAlfred Batschauer, University of Marburg, Germany
Copyright © 2021 Petersen, Rredhi, Szyttenholm, Oldemeyer, Kottke and Mittag. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Maria Mittag, bS5taXR0YWdAdW5pLWplbmEuZGU=
ORCID: Jan Petersen,1orcid.org/0000-0002-8469-6679
Anxhela Rredhi,1 orcid.org/0000-0001-9787-7711
Julie Szyttenholm,1 orcid.org/0000-0001-9373-0608
Sabine Oldemeyer, 2orcid.org/0000-0001-7139-7218
Tilman Kottke, 3orcid.org/0000-0001-8080-9579
Maria Mittag, 1orcid.org/0000-0003-3414-9850