 Adalberto Alves Pereira Filho1
Adalberto Alves Pereira Filho1 Grasielle C. D‘Ávila Pessoa1
Grasielle C. D‘Ávila Pessoa1 Lydia F. Yamaguchi2
Lydia F. Yamaguchi2 Mariana Alves Stanton2
Mariana Alves Stanton2 Artur M. Serravite1
Artur M. Serravite1 Rafael H. M. Pereira1
Rafael H. M. Pereira1 Welber S. Neves2
Welber S. Neves2 Massuo Jorge Kato2*
Massuo Jorge Kato2*- 1Laboratório de Fisiologia de Insetos Hematófagos, Departamento de Parasitologia/ICB, Universidade Federal de Minas Gerais, Belo Horizonte, Brazil
- 2Laboratory of Natural Product Chemistry, Department of Fundamental Chemistry, University of São Paulo, São Paulo, Brazil
The continuous and indiscriminate use of insecticides has been responsible for the emergence of insecticide resistant vector insect populations, especially in Aedes aegypti. Thus, it is urgent to find natural insecticide compounds with novel mode of action for vector control. The goal of this study was to investigate the larvicidal activity of essential oils (EOs) from Piper species against A. aegypti characterized as resistant and susceptible strains to pyrethroids. The EOs from leaves of 10 Piper species were submitted to the evaluation of larvicidal activity in populations of A. aegypti in agreement with the (World Health Organization, 2005) guidelines. The resistance of the strains characterized by determining the lethal concentrations (LCs) with the insecticide deltamethrin (positive control). The major compounds of the EOs from Piper species was identified by GC-MS. The EOs from Piper aduncum, P. marginatum, P. gaudichaudianum, P. crassinervium, and P. arboreum showed activity of up to 90% lethality at 100 ppm (concentration for screening). The activities of the EOs from these 6 species showed similar LCs in both susceptible strain (Rockefeller) and resistant strains (Pampulha and Venda Nova) to pyrethroids. The major compounds identified in the most active EO were available commercially and included β-Asarone, (E)-Anethole, (E)-β-Caryophyllene, γ-Terpinene, p-Cymene, Limonene, α-Pinene, and β-Pinene. Dillapiole was purified by from EO of P. aduncum. The phenylpropanoids [Dillapiole, (E)-Anethole and β-Asarone] and monoterpenes (γ-Terpinene, p-Cymene, Limonene, α-Pinene, and β-Pinene) showed larvicidal activity with mortality between 90 and 100% and could account for the toxicity of these EOs, but the sesquiterpene (E)-β-Caryophyllene, an abundant component in the EOs of P. hemmendorffii and P. crassinervium, did not show activity on the three populations of A. aegypti larvae at a concentration of 100 ppm. These results indicate that Piper's EOs should be further evaluated as a potential larvicide, against strains resistant to currently used pesticides, and the identification of phenylpropanoids and monoterpenes as the active compounds open the possibility to study their mechanism of action.
Introduction
Aedes aegypti (Linnaeus, 1762) (Mattingly et al., 1962) is a mosquito species known to transmit arboviruses such as dengue, chikungunya, and zika virus worldwide. It is a diurnal mosquito extremely adapted to urban and domestic environments (Maciel-de-Freitas et al., 2012). The rapid increase in rates of urbanization in tropical regions, lack of basic infrastructure and limited or non-existent sanitation, associated with favorable climatic conditions for the mosquito's development, have contributed to the expansion of the occurrence range of arboviruses transmitted by females of A. aegypti (Rebêlo et al., 1999; Carvalho and Moreira, 2016). Therefore, we have witnessed the increasing transmission of Dengue (DENV), Zika (ZIKV), and Chikungunya (CHIKV) virus in these regions.
Despite the FDA approval of a dengue vaccine (Dengvaxia) in 2019, its efficiency is restricted to people who have been previously infected by dengue and not as disease prevention for a large portion of the population. Therefore, the control of A. aegypti populations still represents the best line of defense. This strategy has focused on controlling the mosquito's population by means of using insecticides such as the larvicide Pyriproxyfen (Juvenil Hormone Analog–JHA), the adulticides malathion (organophosphate) and Cielo®, an insecticide containing imidacloprid (neonicotinoid) and paletrine (pyrethroid) (Valle et al., 2019). The main larvicide used worldwide was the organophosphate temephos, but by the end of 1990's, it led to the development of resistance in A. aegypti. In fact, in the last decades, the indiscriminate use of synthetic insecticides (for example, domestic use of pyrethroid insecticides available in the retail market, especially in epidemic periods), together with the lack of coordinated programs in multi-endemic areas, have led to the emergence of populations of A. aegypti resistant to different insecticides used (Maciel-de-Freitas et al., 2012; Macoris et al., 2018).
The resistance of A. aegypti in Brazil studied from 2005 to 2012 was characterized by the frequency and distribution of the resistance of this vector (Valle et al., 2019). The phenotypes for populations resistant to pyrethroids throughout the country have been characterized and are associated to the changes in biochemical and target site mutations V410L, G923V, I1011M, V1016I, and F1534C (Brengues et al., 2003; Saavedra-Rodriguez et al., 2007; Martins et al., 2009a,b; Lima et al., 2011; Araújo et al., 2013; Lins et al., 2014; Maciel-de-Freitas et al., 2014; Bellinato et al., 2016; Collet et al., 2016; Dolabella et al., 2016; Haddi et al., 2017; Viana-Medeiros et al., 2017; Garcia et al., 2018; Valle et al., 2019; Costa et al., 2020).
In the case of larval resistance in A. aegypti, two studies were carried out in the city of Belo Horizonte (MG, Brazil) (Belinato et al., 2013; Valle et al., 2019). The resistance ratio 95 (RR95) to the insecticide Temephos was quantified in mosquito populations in 2005 (Belinato et al., 2013) and 2008 (Valle et al., 2019), in which resistance was observed (RR95 = 5.4 and RR95 = 10.8, respectively).
Essential oils (EOs) are odoriferous and volatile compounds found stored in plants structures, such as glands, secretory trichomes, secretory ducts, secretory cavities, or resin ducts (Ciccarelli et al., 2008; Bezić et al., 2009; Liolios et al., 2010; Morone-Fortunato et al., 2010). The production of these volatiles in plants is associated with the ecological role they display in nature, such as protecting plants against pathogens and herbivores and attraction of pollinating insects (Grodnitzky and Coats, 2002; Csekea et al., 2007; Bakkali et al., 2008). The emission of plant volatiles is associated with several messages they convey to the surrounding interacting organisms, such as volatiles used in the attraction of pollinating insects, kairomones in response to herbivores, attraction of parasitoids when damaged by herbivores, controlling the growth of pathogens in aerial parts or roots, and so forth (Grodnitzky and Coats, 2002; Csekea et al., 2007; Bakkali et al., 2008; Raveau et al., 2020). EOs from the plants Cymbopogon spp., Ocimum spp. and Eucalyptus spp. are well-known for their application as insect repellents and the active principles are associated with the presence of α-Pinene, Limonene, Citronellol, Citronellal, Camphor and Thymol (Nerio et al., 2010). Additionally, some plant species have been adopted in push-pull strategies for controlling insect pests in agricultures thanks to their emission of specific volatile compounds with repellent or attractive properties that lead pests away from cultivated plants and onto toxic “trap crops” (Cook et al., 2007; Alkema et al., 2019).
Several applications of volatiles of Piper species have been suggested because of their high potential for pest control, and due to the green technologies involved in the extraction process and the expected low environmental impact (da Silva et al., 2017; Salehi et al., 2019). The evaluation of the larvicidal activity of EOs of species of the genus Piper in A. aegypti has already been studied for the species: Piper humaytanum, P. permucronatum, P. hostmanianum, P. gaudichaudianum (de Morais et al., 2007), P. augustum, P. corrugatum, P. curtispicum, P. darienense, P. grande, P. hispidum, P. jacquemontianum, P. longispicum, P. multiplinervium, P. reticulatum, P. trigonum (Santana et al., 2016), P. marginatum (Autran et al., 2009; Santana et al., 2015), P. klotzschianum (Nascimento et al., 2013), P. aduncum (de Almeida et al., 2009; Oliveira et al., 2013; Santana et al., 2015; Scalvenzi et al., 2019), P. corcovadensis (da Silva et al., 2016), P. sarmentosum (Hematpoor et al., 2016), P. betle (Vasantha-Srinivasan et al., 2018; Martianasari and Hamid, 2019), P. arboreum (Santana et al., 2015), and P. capitarianum (França et al., 2021). Besides, non-volatile compounds from Piper species such as amides and lignans have also been described as larvicidal (Cabral et al., 2009; Kanis et al., 2018).
Despite the large number of studies with EOs from Piper species against larvae of A. aegypti, there is no assessment of their effect on strains of mosquitoes resistant to synthetic insecticides. Therefore, considering the limited number of safe chemical approaches for controlling A. aegypti as vectors in the field, the aim of this work is to investigate the larvicidal activity of essential oils of Piper species and to identify the active principle against populations of A. aegypti that are either susceptible or resistant to pyrethroids.
Materials and Methods
Plant Material
The leaves of 10 species of plants belonging to the genus Piper: Piper aduncum, P. marginatum, P. gaudichaudianum, P. crassinervium, P. arboreum, P. hemmendorffii, P. cernuum, P. lucaenum var. gradifolium, P. lindbergii, and P. amalago, were collected in the period of January to June 2018. The vouchers were deposited at the Herbarium of USP–University of São Paulo for identification (Table 1). All collections were made under permits #59161-1 and 010/2018-R from the Sistema de Autorização e Informação em Biodiversidade–SISBIO and Fundação Serra do Japi, respectively.
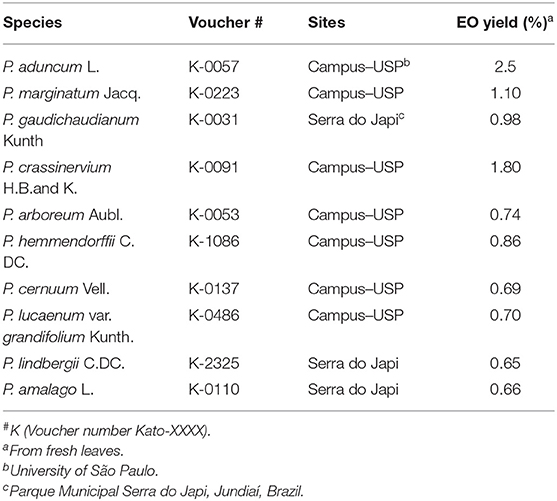
Table 1. Voucher number, sampling site, and yield of essential oil from Piper species.
Collection and Insect Rearing
In this study, three strains of A. aegypti larvae were used. The Rockefeller strain is a susceptible reference lineage (SRL) for all assays. Two other strains were collected in the regions of Pampulha (19° 51′ 04″ S; 43° 58′ 46″ W) and Venda Nova (20° 11′ 51″ S; 44° 1′ 40″ W) in Belo Horizonte, Minas Gerais, Brazil in the period of June 2018. The strains Pampulha (Pamp) and Venda Nova (VN) were evaluated and certified as resistant to pyrethroids.
The mosquitoes were kept and raised in the insectarium of the Laboratory of Physiology of Hematophagous Insects of the Federal University of Minas Gerais in accordance with the recommendations of the Ethics Committee (CEUA-UFMG) (protocol number 01/2017). The insects were maintained under controlled conditions of temperature (27 ± 1°C), photoperiod 12:12 h (L:D), and relative humidity (75%).
After the eggs hatched in dechlorinated water, the larvae and pupae were kept in plastic vats, containing fish food ad libitum. The adult insects were kept in cylindrical cages 30 × 90 cm with mesh on the top and with continuous access to cotton soaked in 10% sucrose solution. The females' blood meals were performed weekly on hamsters (Mesocricetus auratus) previously anesthetized with 0.2 mL of Thiopental® (50 mg/mL) and placed with the trichotomized abdomen on the screen of the cages so that the females could perform the blood meal for 1 h. The eggs were obtained 2 days after the meal using filter paper soaked in dechlorinated water in dark plastic pots, from which they were removed and kept in new plastic pots until the hatching time for testing.
Extraction of Essential Oils
The essential oils (EOs) were extracted from fresh leaves of each species, submitted to hydrodistillation in a Clevenger type apparatus for 4 h, using 300–500 g of fresh leaves and 500 mL of distilled water (Santos et al., 2012; Fanela et al., 2015). The EOs were collected and dried with anhydrous sodium sulfate and stored in amber bottles in a refrigerator at 4°C until the experiments were performed. The yield of EOs from Piper species are shown in Table 1.
Analysis of Essential Oils and Fractions by GC-MS
EOs samples were diluted 20 times in ethyl acetate (HPLC grade, Honeywell) and analyzed using a Shimadzu GCMS- QP2010 equipped with an HP-5ms column (length 30 m, ID 0.25 mm, film thickness 0.25 μm, Agilent) using Helium as a carrier gas (1.55 mL/min) and 1 μL of each sample was injected at 250°C with a 1:20 split. Detector temperature was set at 260°C with electron impact ionization energy of 70 eV and a scan range of m/z35-400 Da at 2500 spectra s−1. The oven program started at 40°C for 2 min, and the temperature was increased at 5°C min−1 to 260°C and held for 2 min. Individual volatile compound peaks were identified using extracted ion traces of three specific reference ions and quantified by the peak area of the most abundant ion trace per compound using a custom-made analysis method in the GC-MS Postrun Analysis software (Shimadzu). Relative % of each compound was calculated by comparing the % peak area in relation to the total sum of peak areas within a sample. The identification of compounds was conducted by calculating Arithmetic retention indexes (RIs) in relation to a series of alkane standards (C8-C40, Supelco) injected using the same GC-MS method as used for the samples, according to Van Den Dool and Kratz (1963). Compound mass spectra and RIs were compared to those available in the Adams and Wiley databases (Adams, 2007), in previous studies of these Piper species, and confirmed by comparison to authentic standards, when available.
LC50 and LC95 for Larvicidal Activity of Deltamethrin Insecticide
The characterization of the larvae to be used in the tests as susceptible or resistant was made by assaying them with the technical grade insecticide deltamethrin (Bayer Brazil, 99.1%). The dose-response tests were performed in the range of 10–90% mortality. Thirty L3-L4 larvae (F1 generation) were separated per dose (in triplicate), requiring a minimum of 8 doses to perform the curve. The larvae were placed in 500 ml cups containing 249 ml of dechlorinated water, along with 1 ml of the insecticide in the desired concentration, diluted in ethanol P.A. For the control group, 30 L3-L4 larvae were used in 250 mL of dechlorinated water. The concentrations of pyrethroid used for assaying Rockefeller, Venda Nova and Pampulha strains were 0.2–0, 9, 2.0–9.5, and 1.0–8.0 mg/mL, respectively.
Mortality was recorded every 10 min during the first hour and 24 h after the start of the test, as recommended by the World Health Organization (1981). Larvae that did not spontaneously move, even if subjected to mechanical stimulation, were considered dead. The LC calculation was performed using the Polo Plus program (see item on statistical analysis). The LC of the field population was divided by the LC of the Rockefeller strain to obtain the Resistance Ratio. The population was considered resistant when RR95 was >3 (Valle et al., 2019).
Qualitative Larvicidal Bioassays of Piper Essential Oils
The tests were performed in accordance with World Health Organization (2005), with modifications. The EOs of 10 species of Piper were diluted in a final volume of 100 ml of dechlorinated water with 2% dimethyl sulfoxide (DMSO), in a concentration of 100 ppm of each oil. Then, 30 larvae (L3–L4) from the three strains, Rockefeller, Venda Nova, and Pampulha were placed in the containers. Each experiment was carried out in three bottles (technical triplicate), being repeated five times on different days (five biological repetitions). The larvae of the control and vehicle control groups were exposed only to dechlorinated water and with 2% DMSO, respectively. Mortality was recorded at 24 h after the start of the test. Larvae not responding to mechanical stimulation were considered dead and the EOs with 90 to 100% larvicidal activity were considered active (Cheng et al., 2003; Dias et al., 2014; Intirach et al., 2016; Muturi et al., 2017).
Determination of LC50 and LC90 of Piper Essential Oils
EOs that showed preliminary larvicidal activity (90 to 100%) had their lethal concentrations (LCs) determined. Thus, 30 larvae L3-L4 from the three strains were submitted to different concentrations in a range of 10–90%, in a final volume of 100 mL of dechlorinated water. Each dose was assayed in duplicate, with three repetitions (biological triplicate) on different days. The larvae of the control and vehicle-control groups were exposed to dechlorinated water and with 2% DMSO, respectively, and the mortality was recorded 24 h after the start of the test.
Assays With the Major Compounds From the EOs
The major compounds from EOs characterized by GC-MS, E-Anethole (Sigma-Aldrich: 4180-23-8), γ-Asarone (Cayman Chemical: 11681) and (E)-β-Caryophyllene (Cayman Chemical: 21572), γ-Terpinene (Sigma-Aldrich: 86478), p-Cymene (Sigma-Aldrich: 121452), Limonene (Sigma-Aldrich: 45423), α-Pinene (Sigma-Aldrich: 147524), and β-Pinene (Sigma-Aldrich: 402753) were acquired commercially. Pure Dillapiole was obtained by fractionation using the Isolera Flash Chromatography system (Biotage INC). The EO of P. aduncum (0.5 ml) was loaded on the silica samplet and the flash chromatography was performed in the SNAP Ultra 25 g silica column using a gradient of hexane and ethyl acetate. The gradient started with 20% of ethyl acetate, after 2 min increased to 28% and in a linear increase reached 33% 12 min. Sixty fractions were collected and dillapiole was present in the fractions 30–35. The samples obtained from this fractionation were analyzed by GC-MS and a fraction with purity higher than 98% was selected for the assays.
For the larvicidal assays (qualitative bioassays), the pure compounds were diluted in dechlorinated water and 2% DMSO, in a final volume of 50 mL, to a final concentration of 100 ppm (screening concentration), containing 15 larvae of A. aegypti L3-L4 per cup. For the determination of LC50 and LC90, the methodology was used as described above, using 15 larvae of A. aegypti L3-L4 per cup.
Statistical Analysis
The data were organized in spreadsheets using Microsoft Excel software (Office 2007). Lethal concentrations (LC) 50%, 90%, 95% and slope were obtained through Probit analysis with the aid of the Polo Plus software (Raymond, 1985). Significant differences in the LC50 and LC90 values were based on non-overlap of 95% confidence intervals (Hematpoor et al., 2017; Wang et al., 2019).
Results
Determination of LC50 and LC95 for the Insecticide Deltamethrin
Based on the bioassays with Deltamethrin, the strains of Pampulha and Venda Nova were shown to be resistant to this insecticide, with the population of Pampulha (RR95 = 26.073) being more resistant than the population of Venda Nova (RR95 = 20.512). The resistance observed in these populations of A. aegypti for Deltamethrin is expected for pyrethroids in general, because of the similarity of the mode of action. The Rockefeller strain, defined as the susceptibility reference strain (LRS), has been maintained in the laboratory since 1881, without contact with insecticides and genetically isolated from external populations (Organização Pan-americana de Saúde (OPAS), 2005). The values of LC50 and LC95 with their 95% confidence intervals are listed in Table 2.
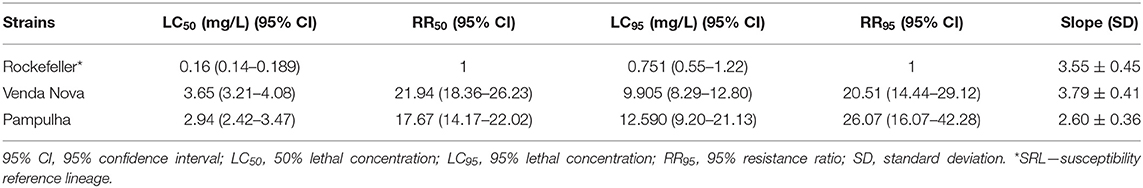
Table 2. LC50 and LC95 for the technical grade deltamethrin insecticide (Bayer Brazil, 99.1%) in larvae of Aedes aegypti.
Larvicidal Activity of Essential Oils
The EOs of the leaves of 10 species of the genus Piper obtained from hydrodistillation were tested against the three strains (resistant and susceptible) of A. aegypti. EOs from 5 out of 10 species were considered active: Piper aduncum, P. marginatum, P. gaudichaudianum, P. crassinervium, and P. arboreum, with larvicidal activity of 90–100% at 100 ppm (Table 3).
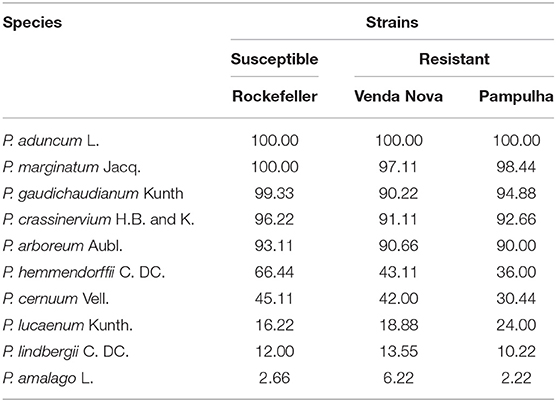
Table 3. Mortality percentage of Aedes aegypti larvae in resistant and susceptible strains to pyrethroids treated with essential oils of Piper species.
EOs of these five species had their lethal concentrations, LC50 and LC90 investigated. Thus, the L3-L4 larvae of the two populations and LRS were submitted to different concentrations to achieve larval mortality in a range of 10–90%. After 24 h of exposure the EOs from P. aduncum, P. gaudichaudianum, and P. marginatum had the lowest LC50 compared to P. crassinervium and P. arboreum. The EOs from P. aduncum was the most active with LC50 (23.50 ppm) for Rockefeller, and LC50 of 25.11 ppm and 26.39 ppm to Venda Nova and Pampulha, respectively (Table 4).
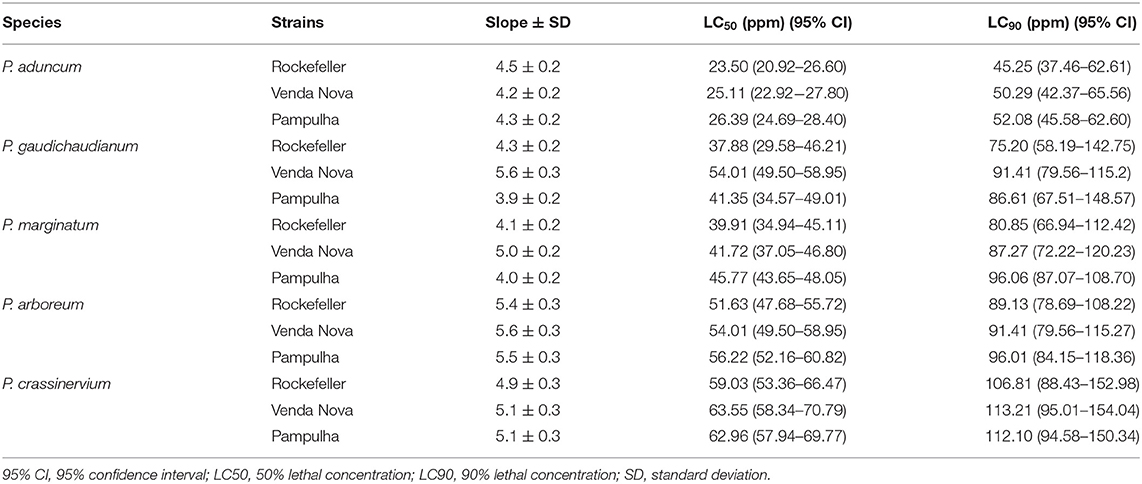
Table 4. Lethal concentrations of essential oil of Piper species against Aedes aegypti larvae resistant and susceptible strains to pyrethroids, during 24 h of exposure.
Identification of Essential Oil Compounds by GC-MS
The EOs of the 10 species were analyzed by GC-MS and main constituents were identified based on library search, retention index (RI), and use of standard compounds when available, and expressed as relative percentage of each constituent (Table 5). In summary, the major compounds were identified as phenylpropanoids, sesquiterpenes and monoterpenes. The complete list of all compounds, the retention indexes, and the relative percentage of each one, for all 10 species of Piper analyzed is shown in Table 6 and the GC-MS chromatograms for all species is shown in the Supplementary Figures 1–3.

Table 5. Major constituents of the EOs of Piper species.
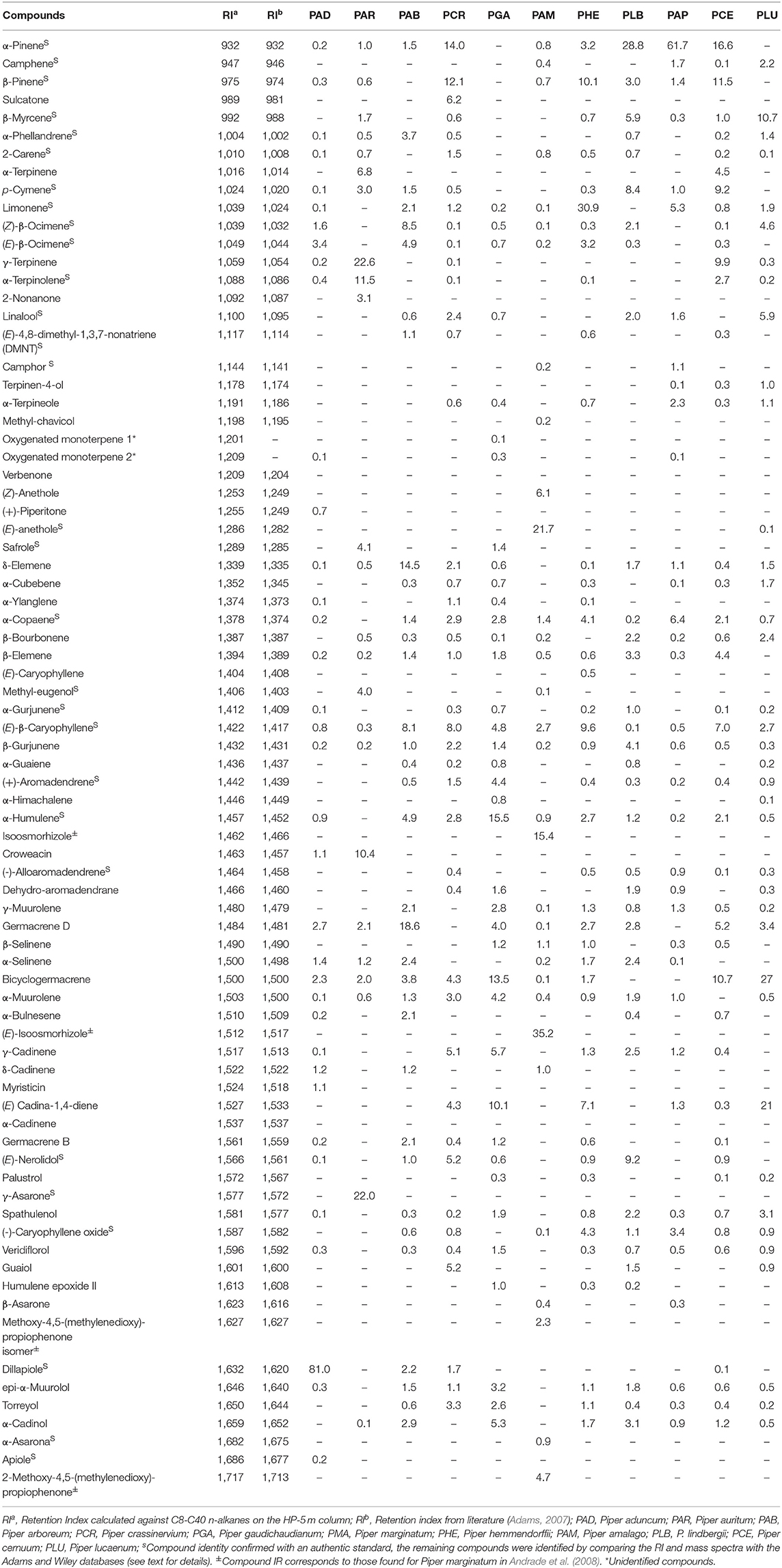
Table 6. Chemical composition of the essential oils of Piper species.
Evaluation of the Larvicidal Activity of the Main Compounds in the EOs
The commercially available compounds (E)-Anethole, β-Asarone, (E)-β-Caryophyllene, γ-Terpinene, p-Cymene, Limonene, α-Pinene and β-Pinene and Dillapiole, obtained by fractionation of EO of P. aduncum using flash chromatography were submitted to further evaluation to determine whether they were involved as the active compounds in the EOs. Thus, pure standards were diluted in water and 2% DMSO, in a final volume of 50 mL, to a final concentration of 100 ppm (screening concentration). Out of the nine compounds evaluated, only (E)-β-Caryophyllene did not show activity on A. aegypti larvae in the three strains at the screening concentration. Nevertheless, the phenylpropanoids (Dillapiole, (E)-Anethole and γ-Asarone) and monoterpenes (γ-Terpinene, p-Cymene, Limonene, α-Pinene and β-Pinene) showed larvicidal activity in the range of 90–100% (Table 7). Additionally, when comparing the LC50 and LC90 of the three phenylpropanoids, Dillapiole displayed the lowest LC50 for the three strains, followed by (E)-Anethole and γ-Asarone (Table 8). Among the five monoterpenes tested, Limonene and γ-Terpinene showed the lowest LC50 for the three strains (Table 8).
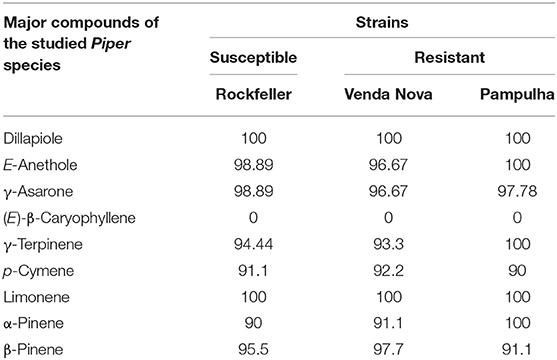
Table 7. Percentage of dead larvae after 24 h of exposure to major compounds of the studied Piper species at concentration 100 ppm.
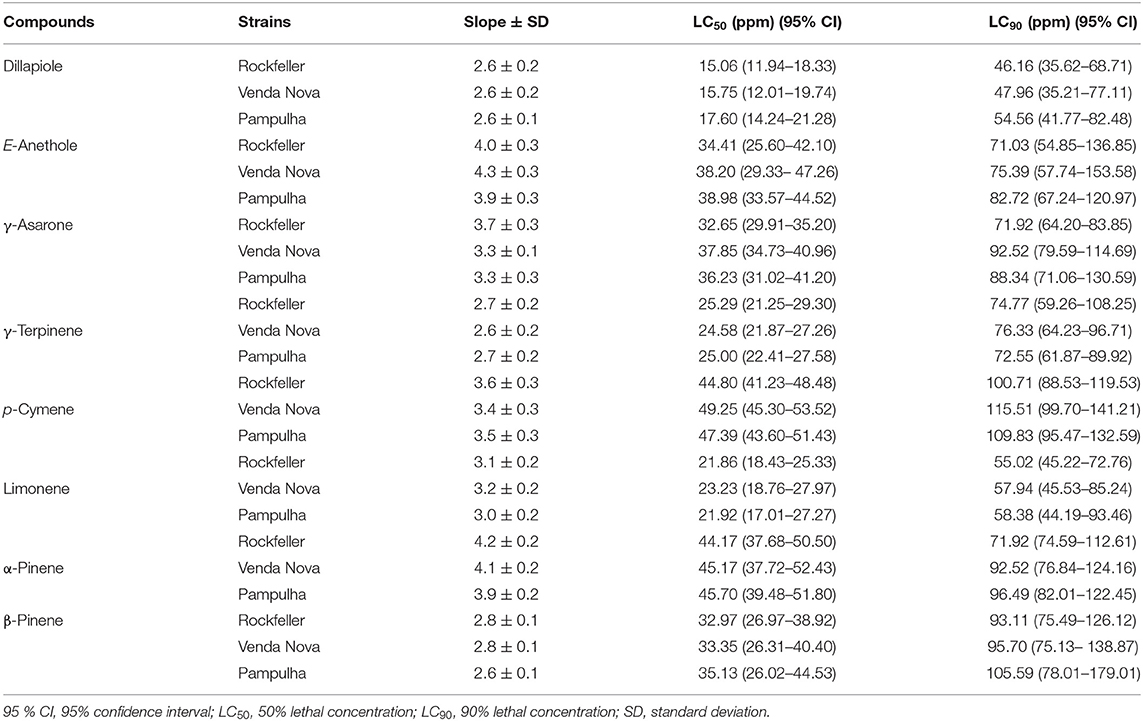
Table 8. Evaluation of lethal concentrations of major compounds in Aedes aegypti larvae of resistant and susceptible strains to pyrethroids during 24 h exposure to major compounds of Piper species.
Discussion
Studies focusing on the investigation of EOs from plants from the perspective of discovering new ovicides, larvicides, adulticides and repellents have been an important strategy for controlling agricultural pests, vectors of medical-veterinary importance or urban viruses (Santos et al., 2012; Phukerd and Soonwera, 2014; Govindarajan et al., 2016; Benelli et al., 2017; Muturi et al., 2017; Luz et al., 2020a). Regarding the urban diseases in tropical regions, A. aegypti is considered one of the main targets since it has great dispersal capacity, is the vector of DENV, ZIKV, CHIKV viruses, and has developed a remarkable resistance to commercially available insecticides (Smith et al., 2016).
The availability of two strains of A. aegypti resistant to Deltamethrin (Table 1) prompted us to seek alternatives to control these populations by screening bioactive EOs from plant species. Despite Deltamethrin not having been used to control A. aegypti in Brazil, it is a stable molecule with a well-known mechanism of action, and it is a standard pyrethroid in studies with insecticide resistance.
In this article, we adopted the WHO methodology (2005) to perform larvicidal tests against A. aegypti (Dias et al., 2015; Luz et al., 2020a). However, as the World Health Organization does not establish criteria to recognize larvicidal activity, in the present study, we choose the level of 90–100% of larvicidal activity for selecting active EOs as previously suggested (Cheng et al., 2003; Dias et al., 2014; Intirach et al., 2016; Muturi et al., 2017). Based on this criterion, EOs from five Piper out of 10 species tested showed larvicidal activity (Table 3). The efficiency of EOs from Piper species as botanical insecticides against various arthropods, including mosquito larvae of the species A. aegypti has been previously demonstrated. For instance, EOs of P. marginatum (Autran et al., 2009), P. aduncum (de Almeida et al., 2009; Oliveira et al., 2013), P. gaudichaudianum (de Morais et al., 2007), P. arboreum (Santana et al., 2015), and P. capitarianum (França et al., 2021) have displayed an efficient larvicidal action on A. aegypti. However, to the best of our knowledge, the present study is the first to demonstrate the bioactivity of EOs of the genus Piper and the main active compounds in essential oils in strains of pyrethroid-resistant A. aegypti larvae.
Among the five species that showed larvicidal activity against A. aegypti, P. aduncum had a lower LC50 compared to the other four Piper considered active, and previous reports for P. aduncum EO activity against larvae of A. aegypti led to variable values of LC50: 46 ppm (Santana et al., 2015); 50.9 ppm (de Almeida et al., 2009), and up to 289.9 ppm (Oliveira et al., 2013). Our average LC50 value of 25 ppm for EO from P. aduncum against the resistant strains (VN and PAMP) and SRL, is similar to that described by Scalvenzi et al., 2019 which was 23.73 ppm.
The analysis of the LC50 and LC90 of the five Piper species active against PAMP, VL, and SRL strains (Table 4) indicated comparable LCs values among them, indicating activity of Piper sp. EOs regardless of insect resistance to commercial pyrethroids. Such similar larvicidal activity, in populations of A. aegypti resistant and susceptible to the organophosphate temephos, was observed with EOs of Syzygium aromaticum (Myrtaceae) and Citrus sinensis (Rutaceae) (Araújo et al., 2016), while a study of EO from Petroselinum crispum (Apiaceae) showed no significant differences of the LC50 for EO larvicidal activity against the pyrethroid resistant and susceptible strains of A. aegypti (Intirach et al., 2016). Our results agree with previous studies of plant EOs and highlights their potential of acting as efficient larvicides on mosquito strains that are resistant to different types of insecticides, whose use has already led to the development of resistant populations in Brazil (for e.g.: Temephos–Valle et al., 2019; pyrethroids—this study and Costa et al., 2020) and elsewhere.
Although Piper species (e.g., P. hemmendorffii, P. lindbergii, P. amalago, P. cernuum) did not show any larvicidal activity, in previous studies their major compounds such as α-Pinene (Ali et al., 2014), β-Pinene (Lee and Ahn, 2013; Ali et al., 2014), (E)-Nerolidol (Ali et al., 2013), Limonene (Cheng et al., 2013; Lee and Ahn, 2013; Rocha et al., 2015; Nascimento et al., 2017), (E)-β-Caryophyllene (Ali et al., 2014, 2015), and β-Myrcene (Cheng et al., 2013; Lee and Ahn, 2013) presented larvicidal activity against A. aegypti. This suggests that minor compounds might negatively interfere with oil larvicidal activity, opening new possibilities to study synergisms between compounds, as their interactions are long-lasting and complex, especially because minor compounds often present biological effects.
Out of the major phenylpropanoid compounds tested, Dillapiole, (E)-Anethole and γ-Asarone (Table 7), only (E)-Anethole was previously reported as an active compound in essential oils against A. aegypti larvae. The LC50 interval found to (E)-anethole (28.0–30.6 ppm) (Rocha et al., 2015), overlaps the confidence interval found for the three strains in the present study (25.60–44.52 ppm) (Table 8). However, in a study carried out by Pandiyan et al. (2019) the LC50 confidence interval, was 48.89–51.50 ppm for the (E)-anethole, which is a higher value than that found in our study.
The sesquiterpene (E)-β-Caryophyllene was the only compound that did not show potential larvicidal activity (Table 7). In fact, this data is in accordance with that found by Luz et al. (2020b), but different to the LC50 values found by Ali et al. (2014) (26, μg/mL), Lee and Ahn (2013) (38.58 μg/mL), and Borrero-Landazabal et al. (2020) (29.28 μg/mL).
Larvae treated with Piper EOs that showed larvicidal activity, were completely damaged, compared with control groups, particularly in the chest and segments of the abdominal region. Specifically, the midgut region was destroyed, and content became dark. These visual observations after the exposure period to Piper's EOs or to major active compounds, indicate morphological (structural) changes in the larva. Therefore, the similar values of LCs in resistant and susceptible strains suggest a mode of action unrelated to the known biochemical and target site mutations in resistant strains.
The chemical profile of the compounds described in the EOs of the Piper species investigated here (Table 6) has already been described in previous studies. For instance, Dillapiole is a typical compound for P. aduncum (Pino et al., 2004; de Almeida et al., 2009; Guerrini et al., 2009; Volpe et al., 2016; Scalvenzi et al., 2019); Germacrene D for P. arboreum (Machado et al., 1994; Mundina et al., 1998; Navickiene et al., 2006; Perigo et al., 2016; Santana et al., 2016); α-Pinene, β-Pinene, and (E)-β-Caryophyllene for P. crassinervium (Morandim et al., 2010; Morandim-Giannetti et al., 2010; Perigo et al., 2016); α-Humulene and Bicyclogermacrene for P. gaudichaudianum (Von Poser et al., 1994; Andrade et al., 1998; Morandim-Giannetti et al., 2010; Sperotto et al., 2013); α-Pinene for P. amalago (Potzernheim et al., 2006; Perigo et al., 2016) and P. cernuum (Bernuci et al., 2016; Perigo et al., 2016).
In case of P. lucaenum, the major compound Bicyclogermacrene described in our study was replaced by α-pinene in another study (Marques et al., 2015). In fact, large chemical variability in EOs of Piper species has already been reported (Andrade et al., 2008). For instance, EOs from 22 samples of P. marginatum leaves collected in different areas and ecosystems of the Brazilian Amazon, separated by up to 1000 km, exhibited different major compounds depending on the place of origin. In our study, while the species P. marginatum had (E)-Anethole as a major compound, analysis of other specimens led to the characterization of 3,4-methylenedioxy propiophenone (Macêdo et al., 2020), and (Z)- or (E)-Asarone, and Patchouli alcohol (Autran et al., 2009) as major compounds. Such variability can result from different environmental conditions, soil composition, development, biotic factors, and plant genetic diversity (Gobbo-Neto and Lopes, 2007; Silva et al., 2019; Mollaei et al., 2020).
Conclusion
Our results suggest the promising role of the EOs of these five species of Piper as an alternative in controlling A. aegypti mosquito larvae of susceptible and insecticide resistant strains. The efficacy of these EOs suggest their use as alternative bioinsecticides in the management of insecticide resistant mosquitoes. Despite the ease of obtaining EOs by hydrodistillation, which is an advantage together the green appeal of such products, their high chemical variability may represent a potential drawback for product development unless a rigorous cultivation control or full understanding of the regulatory processes in the biosynthesis of these phenylpropanoids are achieved.
Data Availability Statement
The original contributions presented in the study are included in the article/Supplementary Material, further inquiries can be directed to the corresponding author.
Author Contributions
AP, MK, and GP designed the research carried. AP, MK, GP, LY, and MS interpreted the data and contributed to writing the manuscript. AS, RP, and WN contributed to methodology and investigation. All authors contributed to the article and approved the submitted version.
Funding
Support for this work was provided by the Brazilian agencies: Fundação de Amparo à Pesquisa do Estado de São Paulo (FAPESP) (2014/50316-7) and Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq).
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
The authors thank for Secretaria Municipal de Saúde de Belo Horizonte, MG, Brazil, for technical assistance in obtaining eggs of Aedes aegytpi from Venda Nova and Pampulha strains.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fpls.2021.685864/full#supplementary-material
References
Adams, R. P. (2007). Identification of Essential Oil Components by Gas Chromatography/Mass Spectrometry, 4.1 Edn. Carol Stream, IL: Allured Publishing Corporation.
Ali, A., Demirci, B., Kiyan, H. T., Bernier, U. R., Tsikolia, M., Wedge, D. E., et al. (2013). Biting deterrence, repellency, and larvicidal activity of Ruta chalepensis (Sapindales: Rutaceae) essential oil and its major individual constituents against mosquitoes. J. Med. Entomol. 50, 824–830. doi: 10.1603/ME12177
Ali, A., Tabanca, N., Demirci, B., Blythe, E. K., Ali, Z., Baser, K. H. C., et al. (2015). Chemical composition and biological activity of four salvia essential oils and individual compounds against two species of mosquitoes. J. Agric. Food Chem. 63, 447–456. doi: 10.1021/jf504976f
Ali, A., Tabanca, N., Kurkcuoglu, M., Duran, A., Blythe, E. K., Khan, I. A., et al. (2014). Chemical composition, larvicidal, and biting deterrent activity of essential oils of two subspecies of Tanacetum argenteum (Asterales: Asteraceae) and individual constituents against Aedes aegypti (Diptera: Culicidae). J. Med. Entomol. 51, 824–830. doi: 10.1603/ME13249
Alkema, J. T., Dicke, M., and Wertheim, B. (2019). Context-dependence and the development of push-pull approaches for integrated management of Drosophila suzukii. Insects 10:454. doi: 10.3390/insects10120454
Andrade, E. H. A., Carreira, L. M. M., da Silva, M. H. L., da Silva, J. D., Bastos, C. N., Sousa, P. J. C., et al. (2008). Variability in essential-oil composition of Piper marginatum sensu lato. Chem. Biodivers. 5, 197–208. doi: 10.1002/cbdv.200890011
Andrade, E. H. A., Zoghbi, M. G. B., Santos, A. S., and Maia, J. G. S. (1998). Essential Oils of Piper gaudichaudianum Kunth and P. regnellii (Miq.) C. DC. J. Essent. Oil Res. 10, 465–467. doi: 10.1080/10412905.1998.9700945
Araújo, A. F. O., Ribeiro-Paes, J. T., Deus, J. T., Cavalcanti, S. C. H., Nunes, R. S., Alves, P. B., et al. (2016). Larvicidal activity of Syzygium aromaticum (L.) Merr and Citrus sinensis (L.) Osbeck essential oils and their antagonistic effects with temephos in resistant populations of Aedes aegypti. Mem. Inst. Oswaldo Cruz. 111, 443–449. doi: 10.1590/0074-02760160075
Araújo, A. P., Diniz, D. F. A., Helvecio, E., de Barros, R. A., de Oliveira, C. M. F., Ayres, C. F. J., et al. (2013). The susceptibility of Aedes aegypti populations displaying temephos resistance to Bacillus thuringiensis israelensis: a basis for management. Parasit. Vectors 6:297. doi: 10.1186/1756-3305-6-297
Autran, E. S., Neves, I. A., Silva, C. S. B., Santos, G. K. N., Câmara, C. A. G., and Navarro, D. M. A. F. (2009). Chemical composition, oviposition deterrent and larvicidal activities against Aedes aegypti of essential oils from Piper marginatum Jacq. (Piperaceae). Bioresour. Technol. 100, 2284–2288. doi: 10.1016/j.biortech.2008.10.055
Bakkali, F., Averbeck, S., Averbeck, D., and Idaomar, M. (2008). Biological effects of essential oils—a review. Food Chem. Toxicol. 46, 446–475. doi: 10.1016/j.fct.2007.09.106
Belinato, T. A., Martins, A. J., Lima, J. B. P., and Valle, D. (2013). Effect of triflumuron, a chitin synthesis inhibitor, on Aedes aegypti, Aedes albopictus and Culex quinquefasciatus under laboratory conditions. Parasit. Vectors 6:83. doi: 10.1186/1756-3305-6-83
Bellinato, D. F., Viana-Medeiros, P. F., Araújo, S. C., Martins, A. J., Lima, J. B. P., and Valle, D. (2016). Resistance status to the insecticides temephos, deltamethrin, and diflubenzuron in Brazilian Aedes aegypti populations. Biomed. Res. Int. 2016:8603263. doi: 10.1155/2016/8603263
Benelli, G., Rajeswary, M., Vijayan, P., Senthilmurugan, S., Alharbi, N. S., Kadaikunnan, S., et al. (2017). Boswellia ovalifoliolata (Burseraceae) essential oil as an eco-friendly larvicide? toxicity against six mosquito vectors of public health importance, non-target mosquito fishes, backswimmers, and water bugs. Environ. Sci. Pollut. Res. Int. 25,10264–11027. doi: 10.1007/s11356-017-8820-0
Bernuci, K. Z., Iwanaga, C. C., Fernandez-Andrade, C. M. M., Lorenzetti, F. B., Torres-Santos, E. C., Faiões, V. D. S., et al. (2016). Evaluation of chemical composition and antileishmanial and antituberculosis activities of essential oils of Piper species. Molecules 21:1698. doi: 10.3390/molecules21121698
Bezić, N., Šamanić, I., Dunkić, V., Besendorfer, V., and Puizina, J. (2009). Essential Oil Composition and Internal Transcribed Spacer (ITS) Sequence variability of four South-Croatian Satureja species (Lamiaceae). Molecules 14, 925–938. doi: 10.3390/molecules14030925
Borrero-Landazabal, M. A., Duque, J. E., and andMendez-Sanchez, S. C. (2020). Model to design insecticides against Aedes aegypti using in silico and in vivo analysis of different pharmacological targets. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 229:108664. doi: 10.1016/j.cbpc.2019.108664
Brengues, C., Hawkes, N. J., Chandre, F., McCarroll, L., Duchon, S., Guillet, P., et al. (2003). Pyrethroid and DDT cross-resistance in Aedes aegypti is correlated with novel mutations in the voltage-gated sodium channel gene. Med. Vet. Entomol. 17, 87–94. doi: 10.1046/j.1365-2915.2003.00412.x
Cabral, M. M. O., Alencar, J. A., Guimaraes, A. E., and Kato, M. J. (2009). Larvicidal activity of grandisin against Aedes aegypty. J. Am. Mosq. Control Assoc. 25, 103–105. doi: 10.2987/08-5828.1
Carvalho, F., and Moreira, L. (2016). Why is Aedes aegypti Linnaeus so successful as a Species? Neotrop. Entomol. 46, 243–255. doi: 10.1007/s13744-017-0520-4
Cheng, S. S., Chang, H. T., Chang, S. T., Tsai, K. H., and Chen, W. J. (2003). Bioactivity of selected plant essential oils against the yellow fever mosquito Aedes aegypti larvae. Bioresour. Technol. 89, 99–102. doi: 10.1016/S0960-8524(03)00008-7
Cheng, S. S., Lin, C. Y., Chung, M. J., Liu, Y. H., Huang, C. G., and Chang, S. T. (2013). Larvicidal activities of wood and leaf essential oils and ethanolic extracts from Cunninghamia konishii Hayata against the dengue mosquitoes. Ind. Crops Prod. 47, 310–315. doi: 10.1016/j.indcrop.2013.03.016
Ciccarelli, D., Garbari, F., and Pagni, A. M. (2008). The flower of Myrtus communis (Myrtaceae): secretory structures, unicellular papillae, and their ecological role. Flora 203, 85–93. doi: 10.1016/j.flora.2007.01.002
Collet, M. L., Frizzo, C., Orlandin, E., Rona, L. D., Nascimento, J. C., Montano, M. A., et al. (2016). Frequency of the Val1016Ile mutation on the kdr gene in Aedes aegypti (Diptera: Culicidae) in South Brazil. Genet. Mol. Res. 15, 1–10. doi: 10.4238/gmr15048940
Cook, S. M., Khan, Z. R., and Pickett, J. A. (2007). The use of push-pull strategies in integrated pest management. Annu. Rev. Entomol. 52, 375–400. doi: 10.1146/annurev.ento.52.110405.091407
Costa, M. M., Campos, K. B., Brito, L. P., Roux, E., Rodovalho, C. M., Bellinato, D. F., et al. (2020). Kdr genotyping in Aedes aegypti from Brazil on a nation-wide scale from 2017 to 2018. Sci. Rep. 10:13267. doi: 10.1038/s41598-020-70029-7
Csekea, L. J., Kaufmanb, P. B., and Kirakosyanb, A. (2007). The biology of essential oils in the pollination of flowers. Nat. Prod. Commun. 2, 1317–1336. doi: 10.1177/1934578X0700201225
da Silva, J. K., da Trindade, R., Alves, N. S., Figueiredo, P. L., Maia, J. G. S., and Setzer, W. N. (2017). Essential oils from neotropical Piper species and their biological activities. Int. J. Mol. Sci. 18:2571. doi: 10.3390/ijms18122571
da Silva, M. F., Bezerra-Silva, P. C., de Lira, C. S., de Albuquerque, B. N. L., Agra Neto, A. C., Pontual, E. V., et al. (2016). Composition and biological activities of the essential oil of Piper corcovadensis (Miq.) C. DC (Piperaceae). Exp. Parasitol. 165, 64–70. doi: 10.1016/j.exppara.2016.03.017
de Almeida, R. R. P., Souto, R. N. P., Bastos, C. N., da Silva, M. H. L., and Maia, J. G. S. (2009). Chemical variation in Piper aduncum and biological properties of its dillapiole-rich essential oil. Chem. Biodivers. 6, 1427–1434. doi: 10.1002/cbdv.200800212
de Morais, S. M., Alves, V., Medeiros, L., Solon, E., and Cavalcanti, B. (2007). Chemical composition and larvicidal activity of essential oils from Piper species. Biochem. Syst. Ecol. 35, 670–675. doi: 10.1016/j.bse.2007.05.002
Dias, C. N., Alves, L. P., Rodrigues, K. A. F., Brito, M. C. A., Rosa, C. S., do Amaral, F. M. M., et al. (2015). Chemical composition and larvicidal activity of essential oils extracted from Brazilian Legal Amazon plants against Aedes aegypti L. (Diptera: Culicidae). Evid. Based Complement. Alternat. Med. 2015:490765. doi: 10.1155/2015/490765
Dias, C. N., Fernandes, D., and Moraes, C. (2014). Essential oils and their compounds as Aedes aegypti L, (Diptera: Culicidae) larvicides: a review. Parasitol. Res. 113, 565–592. doi: 10.1007/s00436-013-3687-6
Dolabella, S. S., Santos, R. L. C., Silva, M. C. N., Steffler, L. M., Ribolla, P. E. M., Cavalcanti, S. C. H., et al. (2016). Detection and distribution of V1016I kdr mutation in the voltage-gated sodium channel gene in Aedes aegypti (Diptera: Culicidae) populations from Sergipe State, Northeast Brazil. J. Med. Entomol. 53, 1–5. doi: 10.1093/jme/tjw053
Fanela, T., Baldin, E., Pannuti, L., Cruz, P., Crotti, A., Takeara, R., et al. (2015). Lethal and inhibitory activities of plant-derived essential oils against Bemisia tabaci Gennadius (Hemiptera: Aleyrodidae) Biotype B in Tomato. Neotrop. Entomol. 45, 201–210. doi: 10.1007/s13744-015-0356-8
França, L. P., Amaral, A. C. F., Ramos, A. S., Ferreira, J. L. P., Maria, A. C. B., Oliveira, K. M. T., et al. (2021). Piper capitarianum essential oil: a promising insecticidal agent for the management of Aedes aegypti and Aedes albopictus. Environ. Sci. Pollut. Res. 28, 9760–9776. doi: 10.1007/s11356-020-11148-6
Garcia, G. A., David, M. R., Martins, A. J., Maciel-de-Freitas, R., Lins, J. G. B., Araújo, S. C., et al. (2018). The impact of insecticide applications on the dynamics of resistance: the case of four Aedes aegypti populations from different Brazilian regions. PLoS Negl. Trop. Dis. 12:e0006227. doi: 10.1371/journal.pntd.0006227
Gobbo-Neto, L., and Lopes, N. P. (2007). Medicinal plants: factors of influence on the content of secondary metabolites. Quim. Nova 30, 374–381. doi: 10.1590/S0100-40422007000200026
Govindarajan, M., Kadaikunnan, S., Alharbi, N. S., and Benelli, G. (2016). Acute toxicity and repellent activity of the Origanum scabrum Boiss. and Heldr. (Lamiaceae) essential oil against four mosquito vectors of public health importance and its biosafety on non-target aquatic organisms. Environ. Sci. Pollut. Res. Int. Res. 23, 23228–23238. doi: 10.1007/s11356-016-7568-2
Grodnitzky, J., and Coats, J. (2002). QSAR evaluation of monoterpenoids' insecticidal activity. J. Agric. Food Chem. 50, 4576–4580. doi: 10.1021/jf0201475
Guerrini, A., Sacchetti, G., Rossi, D., Paganetto, G., Muzzoli, M., Andreotti, E., et al. (2009). Bioactivities of Piper aduncum L. and Piper obliquum Ruiz and Pavon (Piperaceae) essential oils from Eastern Ecuador. Environ. Toxicol. Pharmacol. 27, 39–48. doi: 10.1016/j.etap.2008.08.002
Haddi, K., Tomé, H. V. V., Du, Y., Valbon, W. R., Nomura, Y., Martins, G. F., et al. (2017). Detection of a new pyrethroid resistance mutation (V410L) in the sodium channel of Aedes aegypti: a potential challenge for mosquito control. Sci. Rep. 7:46549. doi: 10.1038/srep46549
Hematpoor, A., Liew, S. Y., Azirun, M. S., and Awang, K. (2017). Insecticidal activity and the mechanism of action of three phenylpropanoids isolated from the roots of Piper sarmentosum Roxb. Sci. Rep. 7:12576. doi: 10.1038/s41598-017-12898-z
Hematpoor, A., Liew, S. Y., Chong, W. L., Azirun, M. S., Lee, V. S., and Awang, K. (2016). Inhibition and larvicidal activity of phenylpropanoids from Piper sarmentosum on acetylcholinesterase against mosquito vectors and their binding mode of interaction. PLoS ONE 11:e0155265. doi: 10.1371/journal.pone.0155265
Intirach, J., Junkum, A., Lumjuan, N., Chaithong, U., Jitpakdi, A., Riyong, D., et al. (2016). Antimosquito property of Petroselinum crispum (Umbelliferae) against the pyrethroid resistant and susceptible strains of Aedes aegypti (Diptera: Culicidae). Environ. Sci. Pollut. Res. Int. 23, 23994–24008. doi: 10.1007/s11356-016-7651-8
Kanis, L. A., Rabelo, B. D., Moterle, D., Custódio, K. M., De Oliveira, J. G., De Lemos, A. B., et al. (2018). Piper ovatum (Piperaceae) extract/starch-cellulose films to control Aedes aegypti (Diptera: Culicidae) larvae. Ind. Crops Prod. 122, 148–155. doi: 10.1016/j.indcrop.2018.05.055
Lee, D. C., and Ahn, Y. J. (2013). Laboratory and simulated field bioassays to evaluate larvicidal activity of Pinus densiflora hydrodistillate, its constituents and structurally related compounds against Aedes albopictus, Aedes aegypti, and Culex pipienspallens in relation to their inhibitory effects on acetylcholinesterase activity. Insects 4, 217–229. doi: 10.3390/insects4020217
Lima, E. P., Paiva, M. H. S., de Araújo, A. P., da Silva, E. V. G., da Silva, U. M., de Oliveira, L. N., et al. (2011). Insecticide resistance in Aedes aegypti populations from Ceará, Brazil. Parasit. Vectors 4:5. doi: 10.1186/1756-3305-4-5
Lins, J. G. B., Brito, L. P., Garcia, G. A., Araki, A. S., Bruno, R. V., Lima, J. B. P., et al. (2014). Distribution and dissemination of the Val1016Ile and Phe1534Cys Kdr mutations in Aedes aegypti Brazilian natural populations. Parasit. Vectors 7, 1–11. doi: 10.1186/1756-3305-7-25
Liolios, C. C., Graikou, K., Skaltsa, E., and Chinou, I. (2010). Dittany of Crete: a botanical and ethnopharmacological review. J. Ethnopharmacol. 131, 229–241. doi: 10.1016/j.jep.2010.06.005
Luz, T. R. S. A., de Mesquita, L. S. S., Amaral, F. M. M., and Coutinho, D. F. (2020a). Essential oils and their chemical constituents against Aedes aegypti L. (Diptera: Culicidae) larvae. Acta Trop. 212:105705. doi: 10.1016/j.actatropica.2020.105705
Luz, T. R. S. A., Leite, J. A. C., de Mesquita, L. S. S., Bezerra, S. A., Silveira, D. P. B., de Mesquita, J. W. C., et al. (2020b). Seasonal variation in the chemical composition and biological activity of the essential oil of Mesosphaerum suaveolens (L.) Kuntze. Ind. Crops Prod. 153:112600. doi: 10.1016/j.indcrop.2020.112600
Macêdo, C. G., Fonseca, M. Y. N., Caldeira, A. D., Castro, S. P., Pacienza-Lima, W., Borsodi, M. P. G., et al. (2020). Leishmanicidal activity of Piper marginatum Jacq. from Santarém-PA against Leishmania amazonensis. Exp. Parasitol. 210:107847. doi: 10.1016/j.exppara.2020.107847
Machado, S. M. F., Militão, J. S. L. T., Facundo, V. A., Ribeiro, A., Morais, S. M., and Machado, M. I. L. (1994). Leaf oils of two Brazilian Piper Species: Piper arboretum Aublet var. latifolium (CDC) Yuncker and Piper hispidum Sw. J. Essent. Oil Res. 6, 643–644. doi: 10.1080/10412905.1994.9699360
Maciel-de-Freitas, R., Aguiar, R., Bruno, R. V., Guimarães, M. C., Lourenço-de-Oliveira, R., et al. (2012). Why do we need alternative tools to control mosquito-borne diseases in Latin America? Mem. Inst. Oswaldo Cruz 107, 828–829. doi: 10.1590/S0074-02762012000600021
Maciel-de-Freitas, R., Avendanho, F. C., Santos, R., Sylvestre, G., Araújo, S. C., Lima, J. B. P., et al. (2014). Undesirable consequences of insecticide resistance following Aedes aegypti control activities due to a dengue outbreak. PLoS ONE 9:92424. doi: 10.1371/journal.pone.0092424
Macoris, M. L., Martins, A. J., Andrighetti, M. T. M., Lima, J. B. P., and Valle, D. (2018). Pyrethroid resistance persists after ten years without usage against Aedes aegypti in governmental campaigns: lessons from São Paulo State, Brazil. PLoS Negl. Trop. Dis. 12:e0006390. doi: 10.1371/journal.pntd.0006390
Marques, A. M., Peixoto, A. C. C., de Paula, R. C., Nascimento, M. F. A., Soares, L. F., Velozo, L. S. M., et al. (2015). Phytochemical investigation of anti-plasmodial metabolites from Brazilian native Piper species. J. Essent. Oil Bear. Plants 18, 74–81. doi: 10.1080/0972060X.2014.974075
Martianasari, R., and Hamid, P. H. (2019). Larvicidal, adulticidal, and oviposition-deterrent activity of Piper betle L. essential oil to Aedes aegypti. Vet. World 12, 367–371. doi: 10.14202/vetworld.2019.367-371
Martins, A. J., Lima, J. B., Peixoto, A. A., and Valle, D. (2009b). Frequency of Val1016Ile mutation in the voltage-gated sodium channel gene of Aedes aegypti Brazilian populations. Trop. Med. Int. Health 14, 1351–1355. doi: 10.1111/j.1365-3156.2009.02378.x
Martins, A. J., Lins, R. M., Lins, J. G., Peixoto, A. A., and Valle, D. (2009a). Voltage-gated sodium channel polymorphism and metabolic resistance in pyrethroid-resistant Aedes aegypti from Brazil. Am. J. Trop. Med. Hyg. 81, 108–115. doi: 10.4269/ajtmh.2009.81.108
Mattingly, P. F., Stone, A., and Knight, K. L. (1962). Culex aegypti Linnaeus, 1762 (Insecta, Diptera); proposed validation and interpretation under the plenary powers of the species so named. Z.N.(S.) 1216. Bull. Zool. Nomencl. 19, 208–219.
Mollaei, S., Ebadi, M., Hazrati, S., Habibi, B., Gholami, F., and Sourestani, M. M. (2020). Essential oil variation and antioxidant capacity of Mentha pulegium populations and their relation to ecological factors. Biochem. System. Ecol. 91:104084. doi: 10.1016/j.bse.2020.104084
Morandim, A., Pin, A. R., Pietro, N. A. S., Alecio, A. C., Kato, M. J., et al. (2010). Composition and screening of antifungal activity against Cladosporium sphaerospermum and Cladosporium cladosporioides of essential oils of leaves and fruits of Piper species. Afr. J. Biotechnol. 9, 6135–6139. doi: 10.5897/AJB09.1956
Morandim-Giannetti, A. D. A., Pin, A. R., Pietro, N. A. S., Oliveira, H. C. D., Mendes-Giannini, M. J. S., Alecio, A. C., et al. (2010). Composition and antifungal activity against Candida albicans, Candida parapsilopisis, Candida krusei, and Cryptococcus neoformans of essential oils from leaves of Piper and Peperomia species. J. Med. Plant Res. 4, 1810–1814. doi: 10.5897/JMPR09.303
Morone-Fortunato, I., Montemurro, C., Ruta, C., Perrini, R., Sabetta, W., Blanco, A., et al. (2010). Essential oils, genetic relationships and in vitro establishment of Helichrysum italicum (Roth) G. Don ssp. italicum from wild Mediterranean germplasm. Ind. Crops Prod. 32, 639–649. doi: 10.1016/j.indcrop.2010.07.023
Mundina, M., Vila, R., Tomi, F., Gupta, M. P., Adzet, T., Casanova, J., et al. (1998). Leaf essential oils of three Panamanian Piper species. Phytochemistry 47, 1277–1282. doi: 10.1016/S0031-9422(97)00762-0
Muturi, E. J., Ramirez, J. L., Doll, K. M., and Bowman, M. J. (2017). Combined toxicity of three essential oils against Aedes aegypti (Diptera: Culicidae) Larvae. J. Med. Entomol. 54, 1684–1691. doi: 10.1093/jme/tjx168
Nascimento, A. M. D., Maia, T. D. S., Soares, T. E. S., Menezes, L. R. A., Scher, R., Costa, E. V., et al. (2017). Repellency and larvicidal activity of essential oils from Xylopia laevigata, Xylopia frutescens, Lippia pedunculosa, and their individual compounds against Aedes aegypti Linnaeus. Neotrop. Entomol. 46, 223–230. doi: 10.1007/s13744-016-0457-z
Nascimento, J. C. D., David, J. M., Barbosa, L. C., de Paula, V. F., Demuner, A. J., David, J. P., et al. (2013). Larvicidal activities and chemical composition of essential oils from Piper klotzschianum (Kunth) C. DC. (Piperaceae). Pest Manag. Sci. 69, 1267–1271. doi: 10.1002/ps.3495
Navickiene, H. M. D., Morandim, A., d,. A, Alécio, A. C., Regasini, L. O., Bergamo, D. C. B., et al. (2006). Composition and antifungal activity of essential oils from Piper aduncum, Piper arboreum and Piper tuberculatum. Quím. Nova 29, 467–470. doi: 10.1590/S0100-40422006000300012
Nerio, L. S., Olivero-Verbel, J., and Stashenko, E. (2010). Repellent activity of essential oils: A review. Bioresour. Technol. 101, 372–378. doi: 10.1016/j.biortech.2009.07.048
Oliveira, G. L., Cardoso, S. K., Lara Junior, C. R., Vieira, T. M., Guimarães, E. F., Figueiredo, L. S., et al. (2013). Chemical study and larvicidal activity against Aedes aegypti of essential oil of Piper aduncum L. (Piperaceae). An. Acad. Bras. Ciênc. 85, 1227–1234. doi: 10.1590/0001-3765201391011
Organização Pan-americana de Saúde (OPAS) (2005). Control of Yellow Fever—Field Guide. Publicação Científica n. 603, 2005. Available online at: www.paho.org (accessed March 12, 2018).
Pandiyan, G. N., Mathew, N., and Munusamy, S. (2019). Larvicidal activity of selected essential oil in synergized combinations against Aedes aegypti. Ecotoxicol. Environ. Saf. 174, 549–556. doi: 10.1016/j.ecoenv.2019.03.019
Perigo, C. V., Torres, R. B., Bernacci, L. C., Guimarães, E. F., Haber, L. L., Facanali, R., et al. (2016). The chemical composition and antibacterial activity of eleven Piper species from distinct rainforest areas in Southeastern Brazil. Ind. Crops Prod. 94, 528–539. doi: 10.1016/j.indcrop.2016.09.028
Phukerd, U., and Soonwera, M. (2014). Repellency of essential oils extracted from Thai native plants against Aedes aegypti (Linn.) and Culex quinquefasciatus (Say). Parasitol. Res. 113, 3333–3340. doi: 10.1007/s00436-014-3996-4
Pino, J. A., Marbot, R., Bello, A., and Urquiola, A. (2004). Essential oils of Piper peltata (L.) Miq. and Piper aduncum L. from Cuba. J. Essent. Oil Res. 16, 124–126. doi: 10.1080/10412905.2004.9698670
Potzernheim, M., Bizzo, H. R., Agostini-Costa, T. S., Vieira, R. F., Carvalho-Cilva, M., and Gracindo, L. A. M. B. (2006). Chemical characterization of seven Piper species (Piperaceae) from Federal District, Brazil, based on volatile oil constituents. Rev. Bras. Plantas Med. 8, 10–12.
Raveau, R., Fontaine, J., and Sahraoui, A., L.-H. (2020). Essential oils as potential alternative biocontrol products against plant pathogens and weeds: a review. Foods 9:365. doi: 10.3390/foods9030365
Raymond, M. (1985). Presentation d'unprogramme basic d'analyse log-probit pour micro-ordinateur. Cah. ORSTOM Ser. Entomol. Med. Parasitol. 23, 117–121.
Rebêlo, J. M. M., Costa, J. M. L., Silva, F. S., Pereira, Y. N. O., and Silva, J. M. D. (1999). Distribuição de Aedes aegypti e do dengue no Estado do Maranhão, Brasil. Cad. Saúde Pública 15, 477–486. doi: 10.1590/S0102-311X1999000300004
Rocha, D. K., Matos, O., Novo, M. T., Figueiredo, A. C., Delgado, M., and Moiteiro, C. (2015). Larvicidal activity against Aedes aegypti of Foeniculum vulgare essential oils from Portugal and Cape Verde. Nat. Prod. Commun. 10, 677–682. doi: 10.1177/1934578X1501000438
Saavedra-Rodriguez, K., Urdaneta-Marquez, L., Rajatileka, S., Moulton, M., Flores, A. E., Fernandez-Salas, I., et al. (2007). A mutation in the voltage-gated sodium channel gene associated with pyrethroid resistance in Latin American Aedes aegypti. Insect Mol. Biol. 16, 785–798. doi: 10.1111/j.1365-2583.2007.00774.x
Salehi, B., Zakaria, Z. A., Gyawali, R., Ibrahim, S. A., Rajkovic, J., Shinwari, Z. K., et al. (2019). Piper Species: a comprehensive review on their phytochemistry, biological activities, and applications. Molecules 24:1364. doi: 10.3390/molecules24071364
Santana, A. I., Vila, R., Cañigueral, S., and Gupta, M. P. (2016). Chemical composition and biological activity of essential oils from different species of Piper from Panama. Planta Med. 82, 986–991. doi: 10.1055/s-0042-108060
Santana, H. T., Trindade, F. T. T., Stabeli, R. G., Silva, A. A. E., Militão, J. S. T. L., and Facundo, V. A. (2015). Essential oils of leaves of Piper species display larvicidal activity against the dengue vector, Aedes aegypti (Diptera: Culicidae). Rev. Bras. Plant. Med. 17, 105–111. doi: 10.1590/1983-084X/13_052
Santos, G. K., Dutra, K. A., Barros, R. A., da Câmara, C. A., Lira, D. D., Gusmão, N. B., et al. (2012). Essential oils from Alpinia purpurata (Zingiberaceae): chemical composition, oviposition deterrence, larvicidal, and antibacterial activity. Ind. Crops Prod. 40, 254–260. doi: 10.1016/j.indcrop.2012.03.020
Scalvenzi, L., Radice, M., Toma, L., Severini, F., Boccolini, D., Bella, A., et al. (2019). Larvicidal activity of Ocimum campechianum, Ocotea quixo sand Piper aduncum essential oils against Aedes aegypti. Parasite 26:23. doi: 10.1051/parasite/2019024
Silva, A. C. A., Diodato, J. S., Castro, J. W., Matias, E. F. F., Silva, L. E., do Amaral, W., et al. (2019). Effect of the essential oils from Piper sp. and blue led lights in the enhancement of the antibiotic activity of drugs against mdr bacterial strains. J. Photochem. Photobiol. B. 199:111604. doi: 10.1016/j.jphotobiol.2019.111604
Smith, L. B., Kasai, S., and Scott, J. G. (2016). Pyrethroid resistance in Aedes aegypti and Aedes albopictus: important mosquito vectors of human diseases. Pestic. Biochem. Physiol. 133, 1–12. doi: 10.1016/j.pestbp.2016.03.005
Sperotto, A. R. M., Moura, D. J., Péres, V. F., Damasceno, F. C., Caramão, E. B., Henriques, J. A. P., et al. (2013). Cytotoxic mechanism of Piper gaudichaudianum Kunth essential oil and its major compound nerolidol. Food Chem. Toxicol. 57, 57–68. doi: 10.1016/j.fct.2013.03.013
Valle, D., Bellinato, D. F., Viana-Medeiros, P. F., Lima, J. B. P., and Martins Junior, A. J. (2019). Resistance to temephos and deltamethrin in Aedes aegypti from Brazil between 1985 and 2017. Mem. Inst. Oswaldo Cruz 114, 1–17. doi: 10.1590/0074-02760180544
Van Den Dool, H., and Kratz, D. J. (1963). A generalization of the retention index system including liner temperature programmed gas-liquid partition chromatography. J. Chromatogr. 11, 463–467. doi: 10.1016/S0021-9673(01)80947-X
Vasantha-Srinivasan, P., Thanigaivel, A., Edwin, E. S., Ponsankar, A., Senthil-Nathan, S., and Selin-Rani, S., et al. (2018). Toxicological effects of chemical constituents from Piper against the environment alburden Aedes aegypti Liston and their impact on non-target toxicity evaluation against biomonitoring aquatic insects. Environ. Sci. Pollut. Res. 25, 10434–10446. doi: 10.1007/s11356-017-9714-x
Viana-Medeiros, P. F., Bellinato, D. F., Martins, A. J., and Valle, D. (2017). Insecticide resistance, associated mechanisms, and fitness aspects in two Brazilian Stegomyia aegypti (=Aedes aegypti) populations. Med. Vet. Entomol. 31, 340–350. doi: 10.1111/mve.12241
Volpe, H. X. L., Fazolin, M., Garcia, R. B., Magnani, R. F., Barbosa, J. C., and Miranda, M. P. (2016). Efficacy of essential oil of Piper aduncum against nymphs and adults of Diaphorina citri. Pest Manag. Sci. 72, 1242–1249. doi: 10.1002/ps.4143
Von Poser, G. L., Rörig, L. R., Henriques, A. T., Lamaty, G., Menut, C., and Bessière, J. M. (1994). Aromatic Plants from Brazil. III. The chemical composition of Piper gaudichaudianum Kunth and P. mikanianum (Kunth) Steudel essential oils. J. Essent. Oil Res. 6, 337–340. doi: 10.1080/10412905.1994.9698395
Wang, Z., Perumalsamy, H., Wang, X., and Ahn, Y. J. (2019). Toxicity and possible mechanisms of action of honokiol from Magnolia denudata seeds against four mosquito species. Sci. Rep. 9:411. doi: 10.1038/s41598-018-36558-y
World Health Organization (1981). Instructions for Determining the Susceptibility or Resistance of Mosquito Larvae to Insecticides. Available online at: https://apps.who.int/iris/handle/10665/69615 (accessed March 12, 2018).
World Health Organization (2005). Guidelines for Laboratory and Field Testing of Mosquito Larvicides. Available online at: https://apps.who.int/iris/handle/10665/69101 (accessed March 12, 2018).
Keywords: essential oils, Piper, larvicides, vector control, Aedes aegypti, insecticide resistance
Citation: Pereira Filho AA, Pessoa GCD, Yamaguchi LF, Stanton MA, Serravite AM, Pereira RHM, Neves WS and Kato MJ (2021) Larvicidal Activity of Essential Oils From Piper Species Against Strains of Aedes aegypti (Diptera: Culicidae) Resistant to Pyrethroids. Front. Plant Sci. 12:685864. doi: 10.3389/fpls.2021.685864
Received: 25 March 2021; Accepted: 27 April 2021;
Published: 04 June 2021.
Edited by:
Rachid Lahlali, Ecole Nationale D'Agriculture de Meknès, MoroccoReviewed by:
Jose Tudela, University of Murcia, SpainOnilda Santos Da Silva, Federal University of Rio Grande Do Sul, Brazil
Copyright © 2021 Pereira Filho, Pessoa, Yamaguchi, Stanton, Serravite, Pereira, Neves and Kato. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Massuo Jorge Kato, bWFzc3Vvam9yZ2VAZ21haWwuY29t