 Pooja Jangir
Pooja Jangir Namita Mehra
Namita Mehra Neeraja Singh
Neeraja Singh Mamta Rani
Mamta Rani Rupam Kapoor
Rupam Kapoor- Department of Botany, University of Delhi, New Delhi, India
Fusarium oxysporum (Fo) is a notorious pathogen that significantly contributes to yield losses in crops of high economic status. It is responsible for vascular wilt characterized by the browning of conductive tissue, wilting, and plant death. Individual strains of Fo are host specific (formae speciales), and approximately, 150 forms have been documented so far. The pathogen secretes small effector proteins in the xylem, termed as Secreted in Xylem (Six), that contribute to its virulence. Most of these proteins contain cysteine residues in even numbers. These proteins are encoded by SIX genes that reside on mobile pathogenicity chromosomes. So far, 14 proteins have been reported. However, formae speciales vary in SIX protein profile and their respective gene sequence. Thus, SIX genes have been employed as ideal markers for pathogen identification. Acquisition of SIX-encoding mobile pathogenicity chromosomes by non-pathogenic lines, through horizontal transfer, results in the evolution of new virulent lines. Recently, some SIX genes present on these pathogenicity chromosomes have been shown to be involved in defining variation in host specificity among formae speciales. Along these lines, the review entails the variability (formae speciales, races, and vegetative compatibility groups) and evolutionary relationships among members of F. oxysporum species complex (FOSC). It provides updated information on the diversity, structure, regulation, and (a)virulence functions of SIX genes. The improved understanding of roles of SIX in variability and virulence of Fo has significant implication in establishment of molecular framework and techniques for disease management. Finally, the review identifies the gaps in current knowledge and provides insights into potential research landscapes that can be explored to strengthen the understanding of functions of SIX genes.
Introduction
Fusarium is a complex and an adaptive genus in Ascomycota that includes both pathogenic as well as non-pathogenic species (Mandeel and Baker, 1991; Gordon and Martyn, 1997). Under this genus, species Fusarium oxysporum Schlechtendal (1824) emend. Snyder and Hansen (1940) (Fo) represents the most pervasive, anamorphic, and polytypic soil-borne pathogen (O’Donnell and Cigelnik, 1997; O’Donnell et al., 1998) that is capable of infecting more than 150 plant species. The host range of Fo varies from vegetables (bottle gourd and tomato), flowers (tulips and carnations), field crops (cotton and chickpea) to plantation crops (banana, dates, and palms) (Pietro et al., 2003; Rana et al., 2017; Edel-Hermann and Lecomte, 2019). Despite showing a broad host range, strains of Fo are highly host specific and are genetically and morphologically distinct (Mandeel et al., 2005; Leslie and Summerell, 2008; Palmero et al., 2009; Edel-Hermann and Lecomte, 2019). Together, these host-specific forms constitute a consortium referred to as F. oxysporum species complex (FOSC) (Edel-Hermann and Lecomte, 2019). FOSC consists of causative agents of vascular wilt, stem-, root-, and crown-rot diseases of economically imperative crops worldwide (Weimer, 1944; Olivain and Alabouvette, 1999; Michielse and Rep, 2009; Gordon, 2017; Rana et al., 2017; Edel-Hermann and Lecomte, 2019). Based on its devastating impact on crop yield, Fo has been positioned fifth among the top 10 economically significant phytopathogenic fungi (Dean et al., 2012).
Genome-wide analysis conducted on Fo has revealed a two-speed genome organization; separating genomic regions required for normal development of the pathogen from relatively fast-evolving regions required for pathogenesis (Croll and McDonald, 2012; Raffaele and Kamoun, 2012; Dong et al., 2015; Fokkens et al., 2018). The host range and specificity of Fo are dictated by genes located on pathogenicity-associated genomic regions (Ma et al., 2010, 2015; Rep and Kistler, 2010; Williams et al., 2016). These pathogenicity-associated genes encode effector proteins, transcription factors (TFs), secreted enzymes, and proteins involved in secondary metabolism and signal transduction (Rep et al., 2002, 2004; Houterman et al., 2007; Van Der Does et al., 2008; Ma et al., 2010; Schmidt et al., 2013). Effector proteins either effectuate a compatible (virulence) response or, on interaction with their corresponding resistance (R) genes, result in incompatible (avirulence) reaction (Flor, 1971; Jones and Dangl, 2006). The horizontal transfer of host-specificity genes to otherwise genetically distinct lineages result in the rapid emergence of new pathogenic lines with a wider host range (Ma et al., 2010). Considering that non-pathogenic strains of Fo can colonize asymptomatic plants as endophytes (Kuldau and Yates, 2000), the potential of these strains to evolve into new virulent lines is a matter of major concern (Gordon and Martyn, 1997; Recorbet et al., 2003; Michielse and Rep, 2009). It is due to the evolution of new pathogens that management strategies for Fusarium wilt have not seen much success. On that account, a thorough understanding of the molecular basis of virulence in Fo is of primary importance as it will provide impetus to the development of efficient and effective disease control strategies. After providing an overview on the biology and variability of Fo pathogens, this review will focus, in particular, on Secreted in Xylem (SIX) genes, their diversity across formae speciales, their role in virulence and host specificity, and evolutionary relationships among Fo pathogens to better understand host–pathogen interactions and rapid emergence of new pathogenic strains.
Host–Pathogen Interaction
Fusarium wilt is a soil-borne disease that is characterized by wilted plants with yellow leaves and a marked reduction in crop yield. The pathogen thrives in warm climate and dry soil; hence, symptoms are severe at 25–30°C (Zitter, 1998; Joshi, 2018). Fusarium, being an anamorphic fungus, produces asexual spores, namely, microconidia, macroconidia, and chlamydospores (dormant propagules) (Gordon, 2017). Germination of these spores is triggered by secretion of exudates from host plant roots and sites of lateral root emergence or injury. Upon germination, the development of infection hypha is initiated that penetrates the root epidermis at the tip (Bishop and Cooper, 1983). Thereafter, the hypha progresses intercellularly via root cortical cells until it enters the xylem tissue. Upon reaching the vascular tissue, the fungus branches profusely and produces microconidia and macroconidia that are transported acropetally by the transpirational pull of plant system (Bishop and Cooper, 1983). Microconidia germinate, and the hyphae spread systemically throughout the host. However, contrasting results were obtained in a study by Michielse and Rep (2009), wherein neither conidiophores nor microconidia were observed in xylem vessels of infected tomato and Arabidopsis plants. These results did not align with the conventional idea that microconidia play an important role in colonization (Beckman, 1987). Generally, to prevent the spread of fungus, resistant plants produce antifungal compounds and occlude the lumen of the xylem vessel by tyloses (VanderMolen et al., 1987; Zhang et al., 1993). This response in susceptible hosts is generally delayed till later stages of infection. Blockage of xylem vessels eventually results in browning of the vascular tissue, a prominent symptom of Fusarium wilt. Disease progression over time leads to leaf bending, chlorosis, wilting, and eventual death of the host (Figure 1). At this stage, the fungus sporulates extensively on the surface of dead plant tissues. The disease spread to other hosts via infected plant parts, transplants or seeds, and contaminated soil (Bishop and Cooper, 1983; Michielse and Rep, 2009; Gordon, 2017; Joshi, 2018).
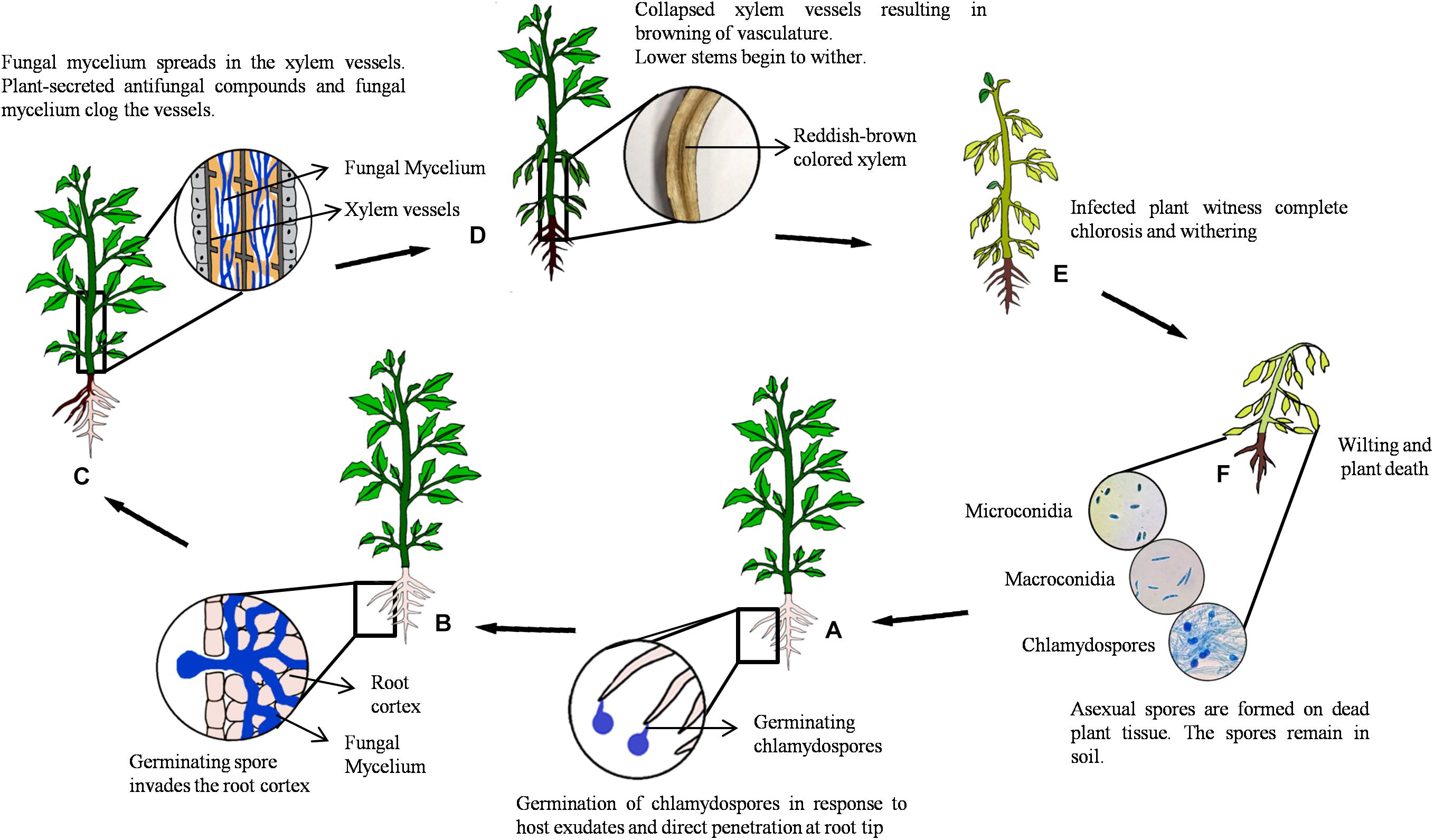
Figure 1. Disease cycle of Fusarium oxysporum. (A) Secretion of root exudates by host plant triggers spore germination and the development of infection hypha prompting penetration of the root epidermis at the tip. (B) The hypha progresses intercellularly via the root cortical cells until it enters the xylem tissue, parenchymal cells, and vessels, through xylem pits. (C) The pathogen colonizes vascular vessels causing blockage and browning as a result of excessive mycelial growth. (D) The initial stage of infection shows symptoms at the stem base and slowly advancing upward, triggering withering of young leaves. (E) Marginal yellowing or complete chlorosis in mature leaves is observed. (F) Disease progression results in wilting and death of the host plant. Fungal spores (microconidia, macroconidia, and chlamydospores) are formed on dead plant tissue and remain dispersed in soil.
Concept of Formae speciales, Races, and Vegetative Compatibility Groups
Pathogenic Fusarium isolates are differentiated at subspecies level into assemblages termed as formae speciales (ff. spp.) (Gordon, 1965; Armstrong and Armstrong, 1981; Baayen, 2000). A forma specialis (f. sp.) is composed of isolates capable of infecting a unique host. Individuals from a forma specialis are further subdivided into pathogenic races depending upon their varied virulence toward cultivars of the same host (Correll, 1991). New races within a forma specialis emerge as a result of mutations in pathogenicity-associated genes. For instance, in race 1 of F. oxysporum f. sp. lycopersici (Fol), whose isolates express three effector genes (AVR1, AVR2, and AVR3), deletion of AVR1 resulted in emergence of race 2 and point mutation(s) in AVR2 eventuated in the evolution of race 3 (Houterman et al., 2008, 2009; Takken and Rep, 2010; Biju et al., 2017). Based on the capability of the isolates to undergo heterokaryosis, they can be grouped as vegetative compatibility groups (VCGs) (Puhalla, 1985; Ploetz and Correll, 1988; Moore et al., 1993). The members of a particular VCG are clonal lineages and share similar pathological, physiological, and biological attributes (Caten and Jinks, 1966). The relationship between races and VCGs of a forma specialis varies from simple to relatively complex (Correll, 1991). In a rather simple relationship, isolates from one race (even from diverse geographical backgrounds) may correspond to a single VCG. For example, in F. oxysporum f. sp. niveum (Fon), all race 2 isolates belong to a single VCG (Correll, 1991; Epstein et al., 2017). On the other hand, occurrence of isolates of different races within a single VCG or isolates of a single race belonging to different VCGs may add to the complexity of the relationship thereof. For instance, three VCGs of F. oxysporum f. sp. cubense (Focub) (0124, 0125, and 0128) contain isolates of races 1 and 2, while isolates of race 1 of Focub belong to eight different VCGs (0123, 0124, 0125, 0128, 01210, 01217, 01218, and 01220) (Czislowski et al., 2018).
Polyphyletic Origin of Fusarium oxysporum Species Complex Members
Recent evolutionary studies have annulled the classical concept that most of the pathogenic isolates of Fo are monophyletic in origin. It is now well established that most formae speciales have evolved independently multiple times throughout the course of evolution pointing towards their para- or polyphyletic origin (O’Donnell et al., 1998, 1999). Remarkably, isolates from one forma specialis, race, or VCG may show close relatedness to the isolates of other formae speciales, races, or VCGs than their own members (Kistler, 1997; Lievens et al., 2009a). Conserved gene sequences and their combinations, mitochondrial or nuclear barcoding, and pathogenesis-related genes have been used to study the evolutionary relationships between different formae speciales, races, and VCGs. Studies based on IGS (intergenic spacer) region, vegetative compatibility, restriction fragment length polymorphism (RFLP), mitochondrial DNA (mtDNA) and isozyme polymorphism have demonstrated that Fol isolates may reflect genetically different evolutionary lines. For instance, phylogenetic analysis of Fol isolates by utilizing IGS rDNA sequences showed three well-supported clusters (A1, A2, and A3) (Kawabe et al., 2005). The major cluster A2 consisted of Fol isolates along with representatives of other formae speciales (melonis, batatas, and radicis-lycopersici). Lievens et al. (2009b) studied the evolutionary relationships between Fol and F. oxysporum f. sp. radicis-lycopersici (Forl) by constructing phylogenetic tree employing pgx4 (exo-polygalacturonase) and Translation Elongation Factor 1α (TEF-1α) gene data and concluded that Fol comprises of three independent clonal lineages. Phylogenetic tree developed by exploiting data from the IGS region of rDNA resolved the isolates of Fol and Forl into five distinct lineages (Cai et al., 2003). Interestingly, Fol VCG 0035 (lineage 5) isolates had more similarities to Forl isolates (lineage 4) compared with the isolates in other Fol lineages or VCGs (Cai et al., 2003). About a decade later, Nirmaladevi et al. (2016), through the ITS (internal transcribed spacer) region analysis, identified evolutionary relationships among Fol isolates and other formae speciales and concluded that Fol represents a polyphyletic forma specialis due to divergent evolution. A similar inference was drawn by employing mitochondrial small subunit (mtSSU) rDNA and TEF-1α-based studies in forma specialis cubense. These phylogenetic studies showed that Focub consists of four clades containing members from various polytypic species (O’Donnell et al., 1998). The results demonstrated a close relatedness of Focub isolates to other representative members of FOSC. Phylogenetic relationships between Focub and FOSC members, as well as between VCGs and Focub races, have revealed that the capacity of pathogens to trigger banana disease has evolved independently multiple times (Fourie et al., 2009). The ability of Focub to inflict disease on a particular cultivar of banana is a polyphyletic trait (Fourie et al., 2009). Multiple gene-genealogical studies established F. oxysporum f. sp. vasinfectum (Fov) as a polyphyletic forma specialis (Skovgaard et al., 2001). The phylogenetic tree obtained from integrated TEF-1α, NIR (nitrate reductase), PHO (acid phosphatase), and mtSSU rDNA sequences reported four different lineages of Fov that correlated with variations in their origin and virulence. The phylogenetic relationship deduced from TEF-1α data of Phoenix-specific F. oxysporum f. sp. canariensis (Focan) isolates reported the presence of three lineages, confirming that Focan in Australia evolved independently (Laurence et al., 2015). Depending on TEF-1α phylogenies, isolates belonging to F. oxysporum f. sp. cucumerinum (Foc) and F. oxysporum f. sp. radicis-cucumerinum (Forc) were reported as genetically diverse and resolved in clades separate from other non-cucurbit-infecting formae speciales (Lievens et al., 2007). Based on the phylogenetic tree obtained from 10 conserved gene dataset, Epstein et al. (2017) described F. oxysporum f. sp. apii as a polyphyletic forma specialis. Evaluation of genetic diversity among forma specialis betae isolates based on ITS, β-tubulin, and TEF-1α phylogenies reported the polyphyletic origin of this forma specialis (Hill et al., 2011). Isolates from F. oxysporum f. sp. melonis (Fom) were also reported to be polyphyletic based on the phylogenetic tree constructed using nuclear repetitive DNA sequences. The isolates were separated into different groups in the phylogenetic tree (Namiki et al., 1994; Gordon and Martyn, 1997). The mtSSU rRNA and TEF-1α phylogenies clustered F. oxysporum f. sp. vanillae isolates into different clades pointing toward a polyphyletic pattern of origin (Pinaria et al., 2015). Similarly, F. oxysporum f. sp. lactucae was described as polyphyletic based on IGS phylogeny and VCGs (Ogiso et al., 2002; Fujinaga et al., 2005; Pasquali et al., 2005).
Not all formae speciales are polyphyletic; a few monophyletic ones have also been reported. In a study by Baayen et al. (2000), TEF-1α, mtSSU rDNA, and amplified fragment length polymorphism (AFLP)-based phylogenies were assessed to identify the nature of origin in 89 isolates belonging to eight different formae speciales. The study revealed two formae speciales, tulipae and lilii, to be monophyletic and the remaining ones, asparagi, dianthi, gladioli, lini, opuntiarum, and spinaciae, to be polyphyletic in origin. Apart from lilii and tulipae (Baayen et al., 2000), ciceris is also considered as a monophyletic forma specialis (Jiménez-Gasco et al., 2002). Isolates of different F. oxysporum f. sp. ciceris (Focic) races shared similar sequences in the intronic region of TEF-1α, β-tubulin, calmodulin, actin, and histone 3 genes by virtue of which they clustered together, separated from other non-pathogenic isolates and formae speciales suggesting a monophyletic origin of Focic (Jiménez-Gasco et al., 2002). Identifying species boundary in FOSC is undeniably a challenge considering the lack of distinct morphological characters, ecological diversity, diverse genetic background, and dynamic host range of strains. FOSC members are devoid of sexual stages in their life cycle; however, horizontal gene transfer (HGT) within the complex may contribute to the observed genetic diversity.
Features of Fusarium oxysporum Genome
The genome sequences of 16 species (11 Fo and five Fusarium species) can be retrieved from the Joint Genome Institute (JGI) MycoCosm site, FungiDb (Fungi database), and GenBank National Centre for Biotechnology Information (NCBI) database (Ma et al., 2010, 2014; Ma L. J. et al., 2013; Williams et al., 2016; DeIulio et al., 2018). The complete genetic and physical maps of the pathogens provide an outstanding opportunity to investigate the variation in genome size and content within the genus (Ma et al., 2010; Ma L. J. et al., 2013). Comparative genomic studies among the members of the genus have provided insight on the variation in genome size within the genus; Fol strain 4287 (Fol-4287) has an average genome size of 61 Mb, whereas Fusarium verticillioides (Fv), Fusarium graminearum (Fg), and Fusarium solani (Fs) (syn. Nectria haematococca) have 42-, 51-, and 36-Mb genome sizes, respectively (Ma et al., 2010). Fv and Fg have comparable genome sizes, even though in the course of evolutionary diversification, Fol and Fv lineages share a common ancestor and have diverged earlier from the clade containing the Fg lineage (Ma L. J. et al., 2013; O’Donnell et al., 2013). Furthermore, genome sequencing and gene mapping of Fusarium species have revealed a variable chromosome count, fluctuating between four in Fg and 17 in Fs (Cuomo et al., 2007; Coleman et al., 2009; Ma et al., 2010).
The F. oxysporum genome is compartmentalized structurally and functionally into two components: a core genome that encodes housekeeping genes vital for survival and growth of the pathogen and an accessory genome encoding pathogenicity or virulence-associated genes (Ma et al., 2010; Croll and McDonald, 2012; Schmidt et al., 2013). Till date, the genome of Fol-4287 remains the most exhaustively studied genome that has been mapped into complete chromosome sequences. Therefore, the Fol-4287 strain is used as a key point of reference for subsequent studies. Out of 15 chromosomes mapped in the genome assembly of Fol-4287, 11 are designated as core chromosomes and four as accessory chromosomes. The core genome of Fol-4287 shows 80 and 90% similarity to Fg and Fv genomes, respectively, suggesting that they are highly syntenic across isolates and related species (Ma et al., 2010).
The accessory genome, also termed as conditionally dispensable (CD) chromosomes, supernumerary (SP) chromosomes, or lineage-specific (LS) region, encompasses 19 Mb of the total genome size and includes chromosomes 3, 6, 14, 15, scaffold 27 of core chromosome 1, and scaffold 31 of core chromosome 2 (Ma et al., 2010). Lineage-specific regions are rich in retro-elements including SINEs (short interspersed repeat elements), LINEs (long interspersed repeat elements), gypsy- and copia-like long terminal repeat retrotransposons, and DNA transposons [miniature inverted transposable elements (MITEs), hAT-like, Tc1-mariner, and Mutator-like] (Ma et al., 2010). The LS region contains 95% of DNA transposons and 74% of all transposable elements (TEs) present in the Fol-4287 genome (Ma et al., 2010) and may be specifically associated with pathogenic adaptation (Ma et al., 2010, 2015). The shared genomic region of Fol and Arabidopsis-infecting strain (Fo-5176) (55 Mb) amounts to less than 2% of sequence divergence. Intriguingly, counterparts of most of the Fol LS region are missing in Fo-5176 (Ma et al., 2010). Similarly, Fov also shows high sequence identity only to the core genomic region of Fol and not to the corresponding LS region (Ma et al., 2010). On this account, comparison among the genomes of Fo pathogens link LS regions to host adaptation (Ma et al., 2010, 2015).
The observed variation in genome size in the genus is attributed to the activity of TEs, horizontal chromosome transfer (HCT), and deletion or fusion of genomic regions (Kistler et al., 1995; Ma et al., 2010; Ma L. J. et al., 2013; Schmidt et al., 2013) (schematically represented in Figure 2). The activity of TEs can cause translocation, deletion, and complex arrangements of genetic material (Davière et al., 2001; Schmidt et al., 2013). The increase in genome size of Fol has been attributed to the acquisition of LS chromosomes from other Fusarium species through horizontal transfer (Ma et al., 2010).
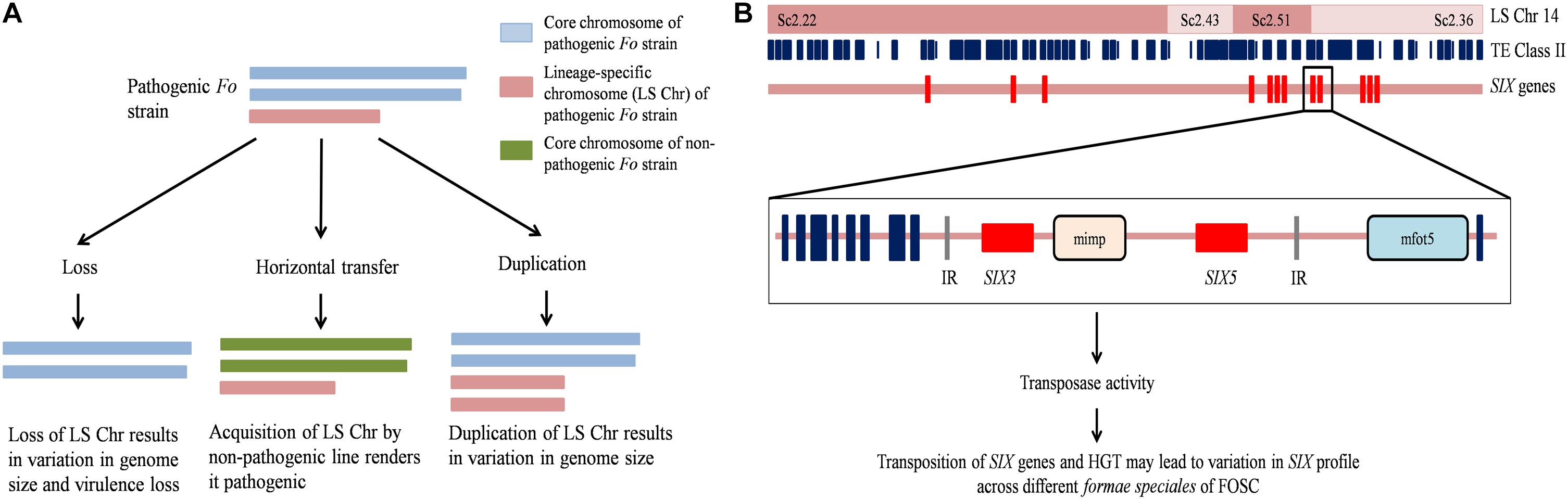
Figure 2. Schematic representation of Fusarium oxysporum genome features. (A) Horizontal transfer, deletion, and duplication of the lineage-specific (LS) chromosome 14 (LS Chr14) alter pathogenicity and genome size. (B) The LS Chr14 structure showing the presence of class II transposable elements (TE class II) and Secreted in Xylem (SIX) genes. TE class II elements present in the promoter region of SIX are miniature impala (mimp) and mfot5 elements. SIX genes might be trapped between the internal resolution (IR) sites of TEs and subsequently transposed together. The mobilization of SIX genes due to the activity of TEs and HGT could result in variation in SIX profile in FOSC members. Fo, Fusarium oxysporum; FOSC, F. oxysporum species complex; HGT, Horizontal gene transfer.
Horizontal Transfer of Mobile Pathogenicity Chromosome
Horizontal transfer provides a mechanism to transfer pathogenicity-associated genes/chromosomes from a pathogenic isolate to a non-pathogenic one, resulting in the generation of a new virulent lineage. Along these lines, the ability of Fol to cause disease on tomato has been presumed to be acquired through horizontal transfer of pathogenicity chromosome from other Fusarium species (Ma et al., 2010). This was experimentally demonstrated via co-incubation studies. Chromosome 14 of Fol pathogenic strain (Fol-007) marked with zeocin gene was co-incubated with a non-pathogenic Fo strain (Fo-47) labeled with the hygromycin gene. Transfer of chromosome 14 during this co-cultivation experiment rendered Fo-47 pathogenic to tomato (Ma et al., 2010). Similar outcomes in co-incubation experiments were obtained where LS chromosome were transferred from a pathogenic line (Fol-4287) to a non-pathogenic line that rendered it pathogenic (Vlaardingerbroek et al., 2016a). Studies have also reported HCT in formae speciales other than Fol. van Dam et al. (2017) through co-cultivation experiments assessed HCT between cucurbit-infecting strains, in which Forc-016 was chosen as the donor strain and Fo-47 as the recipient. All strains obtained from the experiment exhibited the karyotype of Fo-47 strain along with an additional chromosome presumed to have been transferred from Forc-016 as a result of HCT. Besides the members of FOSC, HGT between other Fusarium species and Fo was also reported (van Dam et al., 2017). Flower bulb-infecting strains, Fusarium proliferatum, Fusarium hostae, and Fusarium agapanthi, showed the presence of Fo-specific genes providing evidence for interspecific horizontal transfer due to shared habitat between their ancestors and F. oxysporum f. sp. hyacinthi or F. oxysporum f. sp. lilii strains. On the other hand, complete loss of chromosome 14 in pathogenic Fol-4287 strains compromised virulence on host plants (Ma et al., 2010), whereas no effect on pathogenicity was observed on the deletion of core chromosome (Vlaardingerbroek et al., 2016b). Contrarily, strains with partial deletion of Fol-4287 chromosome 14 regions including effector genes were still pathogenic, implicating that loss of individual or a few genes results in only fractional loss of virulence (Ma et al., 2010). Vlaardingerbroek et al. (2016b) further elaborated on these results. They showed that a part of the short arm (p arm) of the pathogenicity chromosome is adequate for inflicting disease on plants. Transfer of this portion of pathogenicity chromosome is sufficient to convert a non-pathogenic line to a pathogenic line. Interestingly, recipient strains of this portion of chromosome (short arm) were reported to be more virulent than strains that received complete pathogenicity chromosome (short and long arm). This suggested that the sequences present on the long arm (q arm) of the chromosome were possibly involved in suppressing virulence in non-pathogenic strains that received the complete chromosome (Vlaardingerbroek et al., 2016b). It was evident from these studies that LS chromosomes are significant for the development of new pathogenic lines. Owing to the limited availability of whole genome sequences of many formae speciales, it is difficult to trace the path of HCT between strains. More studies are needed to generate a curated database of genome assembly of pathogenic as well as non-pathogenic isolates. Analyses of the genome of isolates from different geographical backgrounds will shed light on how new pathogens evolve on the acquisition of mobile pathogenicity chromosomes from other lineages.
Pathogenicity Factors
Xylem-colonizing Fusarium pathogens employ both general and specific pathogenicity mechanisms to invade the host. While components of cell signaling pathways, such as cyclic adenosine monophosphate (cAMP), mitogen-activated protein kinase (MAPK), Ras (retrovirus-associated DNA sequences) proteins, G (guanine nucleotide-binding) protein, and cell wall-degrading enzymes, encompass the general factors regulating pathogenicity (Di Pietro et al., 2001; Jain et al., 2002, 2003, 2005; Ma L. J. et al., 2013; Guo et al., 2016; Liu et al., 2016), effectors and host-specific toxins attribute specificity to pathogens. Effectors secreted by the pathogen facilitate its colonization by modulating immune response in the host plant (Hogenhout et al., 2009). Secreted in Xylem (Six) proteins is one such example of effectors (Rep et al., 2002, 2004) whose detailed overview, structure, regulation, and diverse roles are dealt in further sections.
Secreted in Xylem Proteins
Rep et al. (2002) identified a small 12-kDa cysteine-rich fungal protein in the xylem sap proteome of tomato plants infected with Fol. Further structural analysis revealed that the observed 12-kDa protein corresponded to the central part (six of the eight cysteine residues) of the actual 30-kDa protein that they termed as Six1 (Rep et al., 2004). Later, Houterman et al. (2007) identified a 22-kDa propeptide of Six 1 protein along with three new Six proteins, namely, Six2, Six3, and Six4, that were approximately 24, 16, and 24 kDa in size, respectively, with eight, two, and six cysteine residues, respectively (Houterman et al., 2007). Van Der Does et al. (2008) and Schmidt et al. (2013), through genomic analysis, identified genes that encode Six5, Six6, and Six7; and Six8, Six9, Six10, Six11, Six12, Six13, and Six14 proteins, respectively. Hitherto, 14 Six proteins have been recognized in Fol. These are small secreted proteins, and most of them contain cysteine residues in even numbers (Rep et al., 2004; Rep, 2005; Houterman et al., 2007; Ma et al., 2010). Initially, SIX genes were considered to be limited to Fol, but later, homologs were identified in other formae speciales as well (Supplementary Table 1). It is noteworthy to mention that non-pathogenic strains of Fo share a set of conserved putative effector genes with the pathogenic strains but carry fewer SIX genes (van Dam et al., 2016; de Lamo and Takken, 2020).
Structure and Regulation of Secreted in Xylem Gene
The accessory chromosome 14 of Fol-4287 strain is dominated by TEs and has been predicted to predominantly harbor all 14 SIX genes (Ma et al., 2010). The presence of TEs in the genome was also associated with clustering of SIX genes observed in TE-rich regions. Occasionally, SIX genes present in the vicinity of IR (inverted repeats) sites of class II TEs might get trapped and translocated together to a new location within class II TE-rich chromosomal subregions (Schmidt et al., 2013). In accordance to that, the highly dynamic genomic location of AVR-Pita in rice blast fungus Magnaporthe oryzae was also attributed to the activity of TEs. Transposon insertion in AVR-Pita gene prevented the host from recognizing this avirulence protein (Zhou et al., 2007). Multiple translocation events of AVR-Pita resulted in a cycle of loss and gain of recognition by resistant rice cultivars (Chuma et al., 2011). Similarly, in Fol, deletion events caused by recombination between TEs led to the loss of an Fol avirulence gene (AVR1) that eventuated in overcoming of resistance mediated by the cognate resistance gene (Biju et al., 2017). Owing to the high density of TEs in LS regions of Fo pathogens, FOSC can prove useful as a model system to decipher relationships between virulence and TEs.
Structurally, SIX genes harbor two MITEs, namely, mimp (miniature impala; sized ≈220 nucleotides) and mFot5 (miniature Fot5 transposon) that vary in their distribution. While mfot5 has been reported to be present downstream of SIX9 or some mini-effector clusters, a portion of mimp is found consistently present in the promoter region of all SIX genes (Schmidt et al., 2013). Hence, mimp can be exploited as a diagnostic feature in the detection of putative SIX genes. Overall, 103 mimp elements have been reported in Fol-4287 genome. Among these, only four are present on core chromosomes, while 54 are located on accessory chromosome 14, and the remaining 45 are on other accessory chromosomes (Schmidt et al., 2013). Homologs of five SIX genes (SIX1, SIX2, SIX6, SIX7, and SIX11) and an avirulence gene (FomAVR2) were identified using mimp elements in melon-Fom pathosystem (van Dam and Rep, 2017). Similarly, mimp elements were utilized to predict effector candidates in Fol (Schmidt et al., 2013), legume-infecting strains such as Focic and F. oxysporum f. sp. pisi (Williams et al., 2016), F. oxysporum f. sp. cepae (Armitage et al., 2018), and race 1 and 4 of Focub (Chang et al., 2020). Interestingly, deletion of mimp element from the promoter region of SIX genes (SIX1, SIX3, and SIX5) neither altered gene expression nor affected pathogenicity of Fol ruling out the direct involvement of mimp in SIX gene expression (Schmidt et al., 2013).
Virulence in Fo is considered a polygenic trait and requires TFs for the regulation of pathogenicity-related genes (Husaini et al., 2018). The role of a TF Six gene expression 1 (SGE1) (situated on the core genome), in modulating the expression of SIX genes (SIX1, SIX2, SIX3, and SIX5) has been confirmed in a study by Michielse et al. (2009b) suggesting the dependency of SIX expression on the core chromosome. In compliance with its transcriptional role, deletion of SGE1 in tomato-infecting Fol resulted in reduced pathogenicity, which is attributable to the lost expression of effector genes. SGE1 deletion mutants of Fol also exhibited a quantitative reduction in conidiation, confirming the major role of SGE1 during parasitic growth of the pathogen (Michielse et al., 2009b). Various orthologs of SGE1 have been reported in fungi such as Fv and Candida albicans (Michielse et al., 2009a; Brown et al., 2014). The retention of this gene in Fol indicates that it is a conserved TF that has developed as a SIX gene regulator (Michielse et al., 2009b).
Two TFs, Fusarium transcription factors (FTF) 1 and 2, belonging to a Zn(II)2Cys6-type family factors, modulate the expression of SGE1 and SIX genes (Niño-Sánchez et al., 2016; Van Der Does et al., 2016). While multiple copies of FTF1 are present on chromosome 14 of Fol-4287 and virulent strains of F. oxysporum f. sp. phaseoli (Foph), a single copy of FTF2 is present in all filamentous Ascomycetes (de Vega-Bartol et al., 2011; Niño-Sánchez et al., 2015, 2016; Van Der Does et al., 2016). Studies on SGE1 reported that the expression of SIX genes is dependent on the core chromosome, but the presence of FTF on the pathogenicity chromosome suggested that the SIX gene expression may also be controlled by the chromosome itself (Michielse et al., 2009b; Schmidt et al., 2013). FTF1 resembles the SIX genes in terms of having mimp in its promoter region (Schmidt et al., 2013; Niño-Sánchez et al., 2016). Deletion mutants of FTF1 and FTF2 have implicated their role in the virulence of Foph; however, their functions as direct regulators of SGE1 and SIX genes need further validation (Niño-Sánchez et al., 2016).
Another important transcriptional regulator in Fom is FOW2 (F. oxysporum f. sp. melonis gene for wilt syndrome 2) (Imazaki et al., 2007). It is essentially required for the invasion and colonization of melon roots. Disruption of FOW2 induced loss of virulence in Fom; however, it had no recognizable effect on vegetative development, conidiation, and carbon source utilization (Imazaki et al., 2007).
Expression of SIX genes is very low in the absence of a living plant host (Michielse et al., 2009b). Under such conditions, the activity of SIX genes might be suppressed by the modification of chromatin to a closed/repressive state (Schmidt et al., 2013). The repressive state is achieved by TE silencing that is guided by small RNAs transcribed from TEs. In Solanaceae members, MITEs in the vicinity of resistance genes have been shown to encode small RNAs that recruit methylation machinery to silence TEs (Kuang et al., 2009). This strategy might serve as the first layer for SIX gene regulation where silencing of the MITEs in the vicinity of SIX genes creates a closed chromatin structure (Schmidt et al., 2013). Clustering of SIX genes in class II TE-rich subregions of the accessory chromosome might have facilitated a coordinated expression of SIX genes during infection. The captured genes share the same genomic environment, i.e., closed or open chromatin structure allowing simultaneous transcriptional regulation of these genes (Schmidt et al., 2013).
Roles of Secreted in Xylem Proteins/Genes
Secreted in Xylem Gene Profile Distinguishes Formae speciales and Races of Fusarium oxysporum
Fungicide treatment and soil solarization generally fail to control wilt infection in fields leaving use of resistant cultivar as the most reliable strategy of disease control (Nirmaladevi et al., 2016). Breeding of resistant cultivars requires a thorough understanding of different formae speciales and races of pathogen emerging in the field, which will provide timely information of genes relevant for breeding programs. Members of FOSC are devoid of discernable morphological characters and exhibit genetic heterogeneity attributed to the polyphyletic origin (Kistler, 1997) and horizontal transfer of pathogenicity-associated chromosomes (Ma et al., 2010). Discrimination between pathogenic and non-pathogenic isolates relies on pathogenicity assays that are both time consuming and strenuous owing to abundance of formae speciales and races in FOSC (Recorbet et al., 2003). On the other hand, standard molecular loci-based techniques used in fungal phylogenetics are also constrained by a weak correlation between pathogenicity and phylogenetic relations (Fraser-Smith et al., 2014).
The above challenges can be addressed by techniques that employ specific sequences of DNA closely associated with pathogenicity (Recorbet et al., 2003; van der Does and Rep, 2007; Lievens et al., 2008), such as SIX genes. In this regard, SIX genes can act as a sensitive and specific diagnostic marker as their array varies among members of different formae speciales and races (Lievens et al., 2008, 2009a) (diagramatically represented in Figure 3). For instance, SIX6 gene was used as a molecular marker to differentiate cotton-specific pathogenic Fov isolates from non-pathogenic ones collected from the same geographical regions in Australia (Chakrabarti et al., 2011). Likewise, three races (1, 2, and 3) of pathogenic Fol have been distinguished on the basis of specific array and number of SIX genes. Isolates of race 1 show the presence of three SIX genes (SIX4, SIX3, and SIX1), while race 2 and 3 show two (SIX3 and SIX1) and one (SIX1) genes, respectively (Rep et al., 2004; Houterman et al., 2008, 2009; Van Der Does et al., 2008; Lievens et al., 2009a; Takken and Rep, 2010; Kang et al., 2014). Furthermore, in Focub, race 1 (that infects Gros Michel cultivars of banana) and race 4 (pathogenic to Cavendish banana) were distinguished on the basis of presence, copy number, and sequence variability of SIX1. Three copies and four sequence variants were observed in race 4 compared with one copy and two variants in race 1 (Guo et al., 2014). Similarly, sequence variants of SIX8 have been used to further differentiate race 4 into tropical (TR4) and subtropical (STR4) races. TR4 race harbors four variants, unlike two in STR4 (Fraser-Smith et al., 2014).
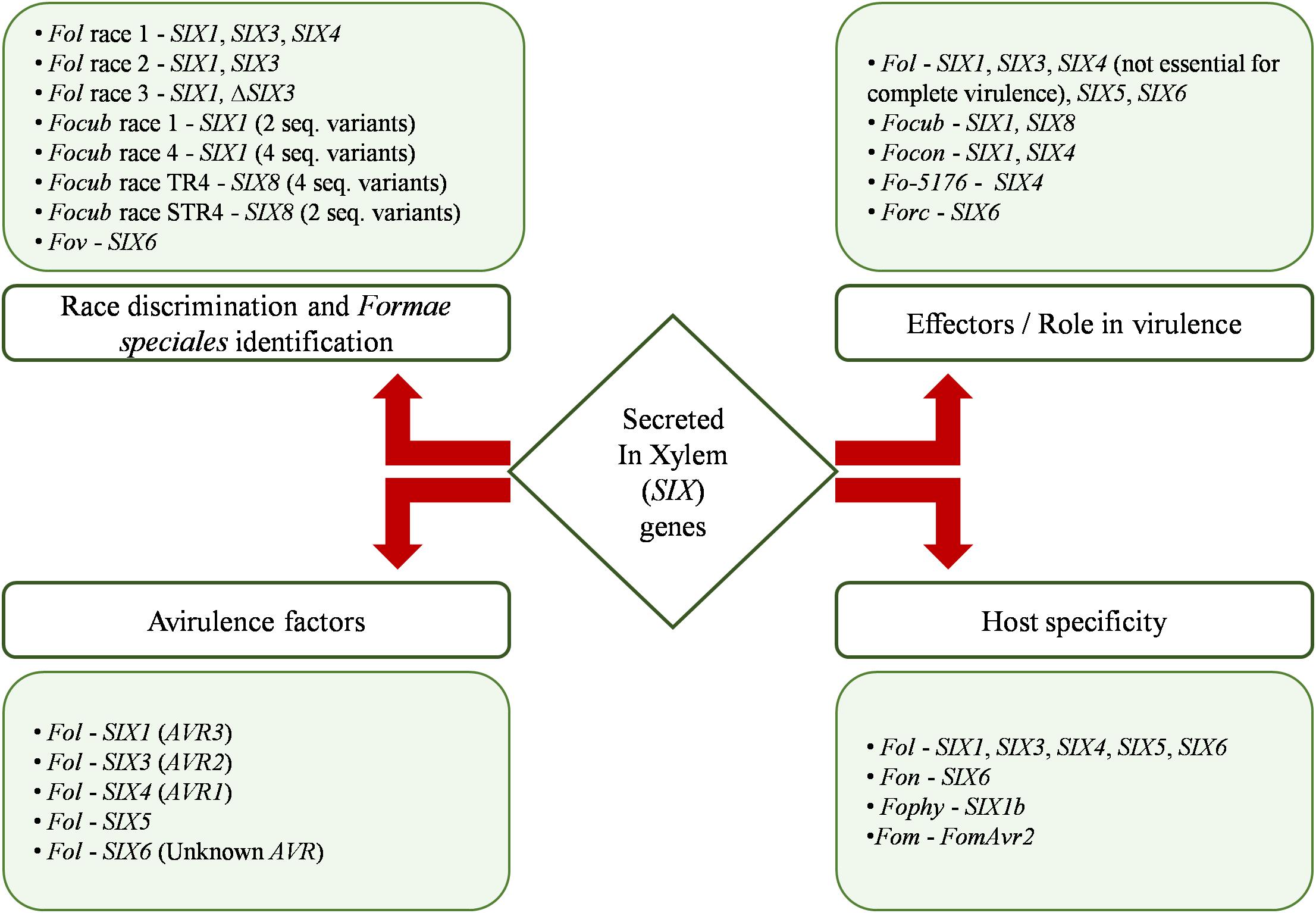
Figure 3. Diagrammatic summary of the roles of various Secreted in Xylem (SIX) genes in different formae speciales of Fusarium oxysporum species complex. Fol, Fusarium oxysporum f. sp. lycopersici; Focub, F. oxysporum f. sp. cubense; Fov, F. oxysporum f. sp. vasinfectum; Focon, F. oxysporum f. sp. conglutinans; Fo-5176, Arabidopsis-infecting strain; Forc, F. oxysporum f. sp. radicis-cucumerinum; Fon, F. oxysporum f. sp. niveum; Fophy, F. oxysporum f. sp. physalis; Fom, F. oxysporum f. sp. melonis; TR4, tropical race 4; STR4, subtropical race 4; AVR, avirulence gene; SIX, Secreted in Xylem gene.
An account of the variation in the arsenal of SIX genes in various formae speciales reported so far is given in Supplementary Table 1. The observed variation in SIX gene profile among formae speciales can be attributed to horizontal transfer of SIX genes among them. Since, LS chromosome 14 carries all SIX genes that reside within subregions of the chromosome rich in class II TEs (Ma et al., 2010; Schmidt et al., 2013), it has been observed that a few genes or a cluster of physically linked SIX genes can be transferred to other strains (Simbaqueba et al., 2018). The similarity in suite of effectors and low SIX sequence diversity in isolates of a forma specialis suggest that SIX genes have been transferred horizontally within and among formae speciales of Fo (Lievens et al., 2009a; Fraser-Smith et al., 2014; Laurence et al., 2015; Taylor et al., 2016; van Dam et al., 2016; Czislowski et al., 2018). In a recent study, the SIX genes profile of strains inhabiting asymptomatic banana plants differed from the known Focub SIX genes (Lyons et al., 2019). Thus, a significant prospect will be to explore the differences in effector gene profile of pathogenic isolates and endophytic strains colonizing asymptomatic plants. It would aid in the accurate detection of different formae speciales, races, as well as endophytic strains that will be contributory in the management of diseases caused by Fo. Moreover, if horizontal transfer can be traced, elucidation of whether the functions of the acquired genes remain conserved in both donor and recipient formae speciales is necessary.
Virulence Function of SIX Genes
The presence of effector genes in formae speciales has been widely documented, but their function in pathogenesis has been experimentally validated only in a few (diagrammatically represented in Figure 3). The presence of SIX1 has been reported to be a prerequisite for complete virulence of the pathogens F. oxysporum f. sp. conglutinans (Focon) and Fol on cabbage and tomato, respectively (Rep et al., 2004; Li et al., 2016). Widinugraheni et al. (2018) reported that SIX1 homolog contributes to virulence of Focub tropical race 4 toward the Cavendish banana. Like SIX1, the role of Fol-SIX3 in complete virulence on host plants has also been demonstrated (Van Der Does et al., 2008). The expression of SIX1 and SIX3 is spatiotemporally separated in Fol. While the expression of Fol-SIX1 is induced in the initial phases of root colonization, Fol-SIX3 is primarily expressed in the xylem during the later stages of hyphae growth (Van Der Does et al., 2008). Likewise, the importance of SIX4 in virulence has been demonstrated by deletion studies in different strains. In Focon, deletion of SIX4 led to a reduction in disease severity on both resistant and susceptible cabbage plants comparative to the SIX4-complemented and wild-type strains (Kashiwa et al., 2013). Similarly, in Fo-5176, SIX4 deletion mutants exhibited reduced fungal biomass that eventually resulted in reduced disease symptoms (Thatcher et al., 2012). Deletion studies carried out in Fol have also highlighted the role of SIX5 as an effector (Ma et al., 2015). Fol-ΔSIX5 displayed an apparent reduction in disease symptoms, and reintroduction of the gene restored pathogenicity in 75% of the mutants. Furthermore, knockout mutants of SIX6 in Fol and Forc exhibited compromised virulence confirming the role of SIX6 in pathogenicity (Gawehns et al., 2014; van Dam et al., 2017). SIX6 plays a role in virulence by inhibiting a hypersensitive response (HR) (Gawehns et al., 2014; De Wit, 2016). In Nicotiana benthamiana leaf cells (heterologous expression system), the transient expression of Fol-SIX6, without its signal peptide, suppressed ion leakage and cell death induced by Avr2-I-2 interaction (Gawehns et al., 2014). Like SIX1, SIX8 is also required for virulence of Focub TR4 on Cavendish banana (An et al., 2019). Similarly, SIX8 is involved in conferring virulence to Fo-5176 on Arabidopsis and cabbage plants (Ayukawa et al., 2020).
Functional annotation is absent for most SIX genes. Information on how they contribute to virulence is still obscure. Some evidence suggest that they facilitate virulence by modulating hormonal pathways or defense response cascades (Thatcher et al., 2012; Ma L. et al., 2013; Gawehns et al., 2014; Ma et al., 2015). The role of the SIX genes in virulence, their targets, and specific biological functions of their protein products warrant more research. The protein–protein interaction assay, such as co-immunoprecipitation, pull-down assays, and yeast two-hybrid, are used to identify putative targets of effector proteins (Alfano, 2009; Rao et al., 2014). In case of transient interaction between effector and their targets, in planta subcellular effector localization can provide hints on target identity. Next-generation sequencing (NGS) technologies can be used to obtain sequences of putative effectors that can be screened for polymorphisms (Alfano, 2009). Identification of effector targets and information on effector polymorphisms will improve our understanding on how the pathogen triggers disease or evade recognition by the host.
Secreted in Xylem Genes Act as Avirulence Determinants
Recognition of effector by cognate resistance (R) gene product of the plant results in induction of ETI (effector-triggered immunity) in the host (Jones and Dangl, 2006). Effectors secreted by the pathogen serve as a two-edged sword (Pradhan et al., 2020). While absence of the R gene in susceptible plants benefits the pathogen, their presence in tolerant plants triggers innate immunity characterized by HR-mediated cell death (Flor, 1971; Jones and Dangl, 2006). However, ETI-mediated resistance to vascular wilt pathogen (Fo) does not include HR response; rather, it involves accumulation of tyloses, gums, phenolic compounds, and callose plugs in the xylem vessels, to preclude systemic spread of the pathogen (Mes et al., 2000; Yadeta and Thomma, 2013).
Some SIX gene products such as Six1, Six3, and Six4 have been found to function as avirulence determinants in tomato-Fol pathosystem and, correspondingly, has been termed as Avr3, Avr2, and Avr1 proteins, respectively (Rep et al., 2004; Houterman et al., 2008, 2009; Takken and Rep, 2010; Ma et al., 2015). Four resistance genes have been identified in wild tomato cultivars (Solanum pimpinellifolium and Solanum pennellii) that confer resistance against Fol races. These resistance genes are I (Immunity), I-2, I-3, and I-7 (Bohn and Tucker, 1939; Alexander and Tucker, 1945; Stall and Walter, 1965; McGrath et al., 1987; Scott and Jones, 1989; Lim et al., 2006). Individual expression of these genes in commercial cultivars of tomato resulted in the development of resistance against race 1, race 2, and race 3. The I gene that encodes an LRR-RLP protein (a class of receptor-like protein) was found to provide resistance against Fol race 1 upon recognizing AVR1 (Houterman et al., 2008; Catanzariti et al., 2017). Both I-3 [a cell-surface S-receptor like kinase (SRLK)] and I-7 (LRR-RLP) proteins conferred resistance against race 3 by recognizing Avr3 and Avr7, respectively (Rep et al., 2004; Lim et al., 2006, 2008; Catanzariti et al., 2015; Gonzalez-Cendales et al., 2016). Likewise, I-2 (a cytoplasmic coiled-coil nucleotide-binding leucine-rich repeat protein) provided resistance against race 2 and race 1 by recognizing Avr2 (Simons et al., 1998; Houterman et al., 2009). Later, Di et al. (2017) demonstrated that I-2 confers resistance by recognizing a specific epitope of Avr2. Ma et al. (2015) observed a new variant of the gene-for-gene model, where they observed that interaction between SIX5 and AVR2(SIX3) is required for I-2-mediated resistance in tomato. While mutations in SIX5 led to evasion of recognition and also compromised the virulence of Fol, heterologous expression of AVR2 and I-2 in N. benthamiana leaves triggered I-2-mediated cell death (Gawehns et al., 2014; Ma et al., 2015). Cao et al. (2018), using the biomolecular fluorescence complementation assay, showed that Avr2 and Six5 interact at plasmodesmata, and Six5 facilitates cell-to-cell movement of Avr2, which in I-2-containing plants results in resistance. Interestingly, Avr1 also mediates the suppression of I-triggered responses. AVR1 gene in race 1 isolates enables the pathogen to overcome resistance response mediated by I-2 and I-3 despite the expression of AVR2 and AVR3 (Rep et al., 2005; Houterman et al., 2008). This strategy has enabled the pathogen to circumvent the emergence of new races carrying AVR1 and AVR3 that would retain the virulence function of AVR3 while avoiding I-3-initiated resistance (Catanzariti et al., 2017). However, this suppression effect is strain-specific suggesting the involvement of an unknown fungal factor (Houterman et al., 2008; Chellappan, 2014). Suppression of resistance response by AVR1 also established I as a gene of practical importance proposing that I-3-mediated resistance was safeguarded by deployment of I (Houterman et al., 2008). Nevertheless, AVR1 was not able to overcome resistance mediated by I-7 against race 3 (Gonzalez-Cendales et al., 2016). I-7 is EDS1 (enhanced disease susceptibility1)-dependent and I-2 and I-3 are EDS1 independent (Hu et al., 2005; Gonzalez-Cendales et al., 2016). The EDS1 signaling pathway is required for basal defense and systemic-acquired resistance (Catanzariti et al., 2017). The fact that AVR1 suppresses I-2 and I-3 and not I-7 suggests that AVR1 is not a general suppressor of basal resistance (Catanzariti et al., 2017). Recently, a new R-AVR interaction was recognized in melon-Fom pathosystem reported by Schmidt et al. (2016) wherein a novel AVR gene, FomAVR2, is recognized by FOM2 in resistant melon plants. Similar to SIX genes, FomAVR2 encodes a small secreted protein with two cysteine residues and is found associated with a mimp element in the promoter region (Schmidt et al., 2016).
Single nucleotide polymorphism serves as the source of genetic variation in SIX gene sequences. For instance, SIX genes in Focub were found to be present in multiple copies and showed the presence of sequence variants (Fraser-Smith et al., 2014). The gain of forms and variation in sequence of pathogenicity-associated genes are presumed as adaptations by the pathogen to respond to rapidly changing environment and host (Cuomo et al., 2007; Fraser-Smith et al., 2014; Maldonado et al., 2018). Houterman et al. (2009) demonstrated that Fol strains that are able to overcome I-2-mediated resistance carry specific point mutations in AVR2. These mutations in AVR2 resulted in amino acid change in the protein that led to the loss of its avirulence function. Till now, three AVR2 alleles have been described, each with one amino acid change at V41→M, R45→H, and R46→P in the protein (Houterman et al., 2009; Biju et al., 2017). Additionally, a race 3 isolate showed the presence of an AVR2 gene with deletion of threonine residue at position 50 of the protein. This deletion also resulted in loss of avirulence function of Avr2 (Biju et al., 2017). Di et al. (2017) analyzed the crystal structure of one of the AVR2 variants (AVR2R45H) that is able to evade recognition by I-2 while retaining its virulence function. They identified two threonine residues in Avr2 protein (T53 and T145) that are required for virulence of Avr2 but not for recognition by I-2. The study revealed that the site of recognition by I-2 differs from the site required for maintenance of virulence function of Avr2. Avr2(Six3) facilitates virulence by suppression of pattern-triggered immunity (PTI) response, mainly, MAPK cascade, ROS burst, callose deposition, and growth inhibition (Di et al., 2017). Similar to AVR2, AVR1, and AVR3 are presumed to be equally likely to undergo mutations that increase the probability of breakdown of resistance in tomato cultivars (Takken and Rep, 2010). Such studies should be extrapolated to the remaining SIX genes to investigate if any variation in their sequence implies structural changes in their corresponding proteins, which may potentially increase the likelihood of evasion of SIX gene recognition by cognate R gene. Thus, it becomes important to uncover the mode of recognition of effector proteins. Additionally, as no direct interaction of avirulence gene with the host R gene is documented in Fol, efforts toward mining the targets of effector and cognate R-gene proteins need more impetus to understand disease resistance in host-Fo pathosystems. It will also be interesting to understand how the plant’s response to other vascular wilt fungi varies from Fo.
Furthermore, physical linkage of certain SIX genes observed in various formae speciales is deemed as important for the functions of the interacting pair of genes. As mentioned above, the AVR2(SIX3)–SIX5 linkage is one such example. These genes reside as a minicluster on chromosome 14 and their expression is under the control of the same bidirectional promoter present on the shared upstream region (Schmidt et al., 2013). Another pair of effector genes, SIX8–PSE1, was identified in isolates capable of infecting Arabidopsis, and this pair was found to be associated with suppression of resistance in Arabidopsis (Ayukawa et al., 2020). The mode of action of SIX8–PSE1 potentially involves suppression of a phytoalexin called camalexin. The SIX8–PSE1 pair was found to be present in head-to-head orientation similar to the SIX3–SIX5 gene pair. However, unlike SIX3–SIX5, no direct interaction between SIX8–PSE1 was detected in yeast two-hybrid assays. Mutation in PSE1, and not the SIX8 gene, resulted in evasion of recognition by the corresponding resistance protein suggesting that PSE1 is required to avoid detection (Ayukawa et al., 2020). A conserved gene cluster SIX7/SIX10/SIX12 was also observed in formae speciales narcissi and gladioli (Simbaqueba et al., 2018). However, the presence of SIX7 alone in formae speciales cubense and lilii suggests that SIX7 may be functionally and physically separable from SIX10 and SIX12 (Simbaqueba et al., 2018). Clustering of SIX genes reflects cooperative interactions important to initiate (a)virulence functions. On this account, deletion studies either of any individual gene or a partial or a complete cluster can be done to elucidate the physical interactions among genes in a cluster and their individual roles in (a)virulence.
Secreted in Xylem Genes Confer Host Specificity
The capacity of fungal species to provoke disease on a specific host is referred to as host specificity. The basis of host specificity has been explained by molecular models like the guard and decoy hypothesis (Van der Biezen and Jones, 1998; Dangl and Jones, 2001; van der Hoorn and Kamoun, 2008; Zipfel and Rathjen, 2008). These models have tremendously contributed in deciphering the role of effectors on the virulence of the pathogen as well as understanding the underpinnings of host–pathogen interactions (van der Hoorn and Kamoun, 2008; Borah et al., 2018).
It is widely accepted that the factors that contribute to (a)virulence of a pathogen also determine its host specificity (Li et al., 2021). In this regard, there are studies where SIX genes have been implicated in imparting host specificity to the pathogen. Avirulence genes of Fol and Fom that confer resistance to races of tomato and melon, respectively, function as host-specific factors (Rep et al., 2004; Houterman et al., 2008, 2009; Catanzariti et al., 2015; Schmidt et al., 2016). SIX6 gene from Fon has been known to operate as an avirulence gene in watermelon-Fon pathosystem providing host specificity to the pathogen (Niu et al., 2016). The role of F. oxysporum f. sp. physalis (Fophy) SIX1 gene in specificity was demonstrated through complementation experiment where complementation strains of two homologs of Fophy-SIX1 (a and b) failed to overcome virulence loss in Fol-ΔSIX1 transformants (Simbaqueba et al., 2018). Interestingly, SIX1b complementation restored avirulence of Fol on IL7-3 transgenic tomato lines carrying I-3, demonstrating that Fophy-SIX1b is recognized by the resistance gene and functions as an avirulence factor. Similarly, complementation of Focon–ΔSIX1 mutant using Fol–SIX1 failed to rescue the virulence of Focon on cabbage suggesting a host-specific role of Fol–SIX1 (Li et al., 2016).
Formae speciales of Fo are generally host specific but Forc shows an exceptional host range. Forc infects cucumber, melon, watermelon, squash, and gourd (Vakalounakis, 1996; Punja and Parker, 2000; Cohen et al., 2015). Previous studies have demonstrated that forma specialis cucumerinum showed mild cross-pathogenicity toward melons (Cafri et al., 2005) that was later corroborated in the findings of a study by van Dam et al. (2016). They assessed cross-pathogenicity of cucurbit-infecting strains (cucumerinum, radicis-cucumerinum, melonis, and niveum) on resistant and susceptible cultivars of their corresponding hosts. The study revealed that Fom and Fon were highly host specific, whereas isolates of Forc displayed some degree of cross-pathogenicity toward musk melon (van Dam et al., 2016). The genetic mechanism underlying the difference in host range was examined, and results showed that effector genes present on the mobile pathogenicity chromosome of Forc and Fom limit host range (van Dam et al., 2017). A close comparison of the mobile pathogenicity chromosomes of Forc and Fom revealed that a single gene on mobile pathogenicity chromosome of Fom determined this difference in host range. This gene, upon introduction in Forc, rendered it non-pathogenic on cucumber suggesting that the gene functions as an avirulence factor (Li et al., 2021). Overall, studies devoted to discerning the role of SIX genes in determining host-specificity in members of FOSC are sparse (Table 1), obstructing our understanding of host specialization. Hence, more studies are required to identify gene(s) involved in limiting the host range of FOSC members.
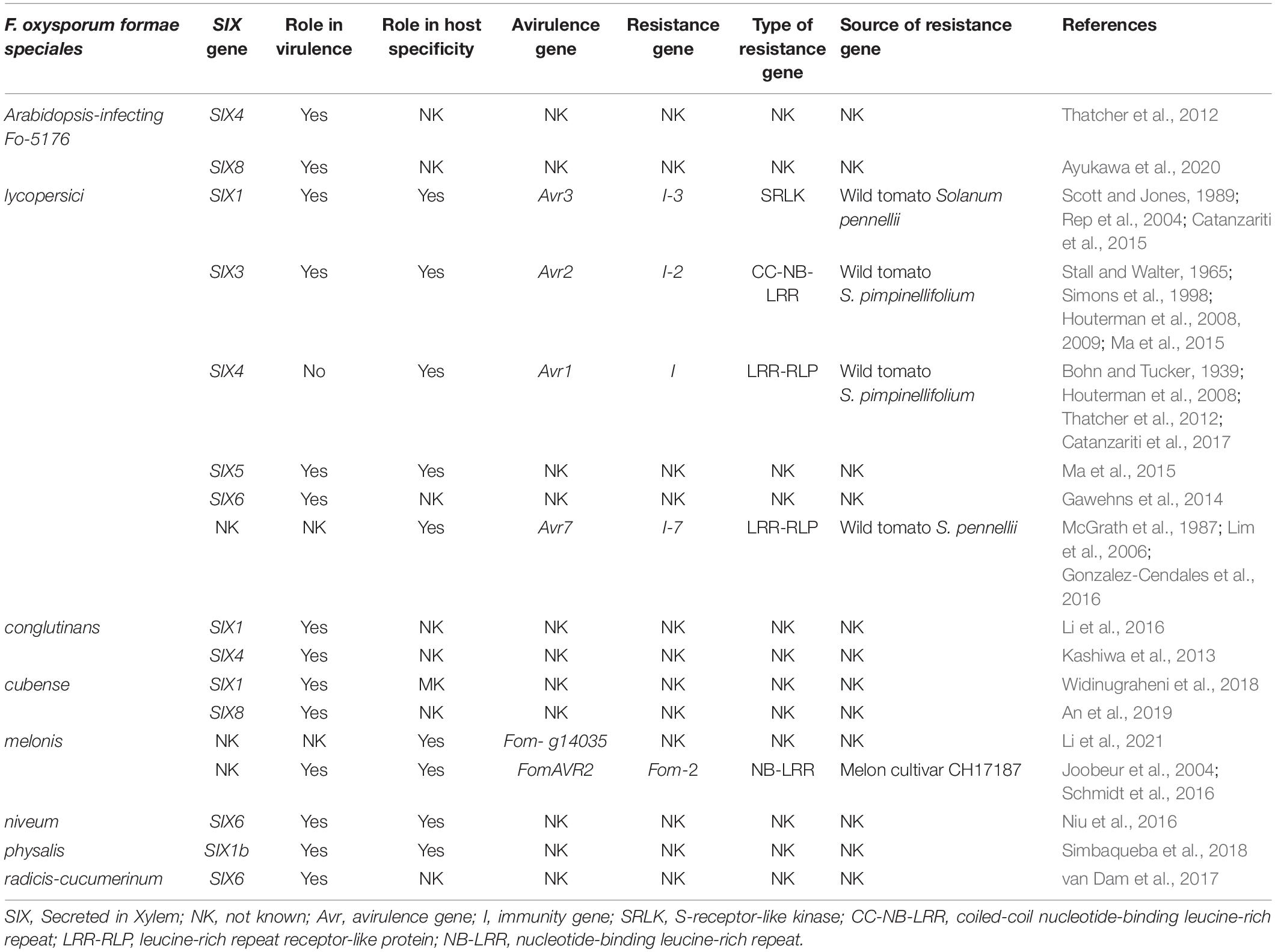
Table 1. Secreted in Xylem genes in formae speciales of Fusarium oxysporum, their role in (a) virulence and host specificity.
Conclusion
Even though F. oxysporum infects a variety of plant species, the investigations on the molecular basis of pathogenicity are restricted to a limited number of hosts, mainly tomato, banana, melons, cucurbits, and cabbage. Genome sequencing of tomato and other cucurbit-infecting pathogens has provided insights on host–pathogen interactions, but a large number of formae speciales are yet left unexplored. Genome-based approaches are needed to elucidate mechanisms and understand the evolution of the pathogenicity. Transcriptomic and proteomic studies in infected plant tissues, the role of transposons and HGT in genome structure modulation, and emergence of host-specific pathogenicity are particular areas of interest. A significant prospect is to explore the differences in effector gene profile between different formae speciales, races, and non-pathogenic isolates. It would aid in the molecular detection of different formae speciales and pathogenic races that will assist significantly in the management of diseases caused by Fo. In addition, the biological functions of effector genes are still under investigation and require exhaustive research.
Author Contributions
PJ and RK jointly conceptualized and wrote the manuscript, contributing major parts of the literature survey. All the authors have collectively reviewed the manuscript and approved it.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
The authors jointly acknowledge the Department of Biotechnology, Government of India, and Institution of Eminence, University of Delhi, India (Grant no. IoE/FRP/LS/2020/27). RK acknowledges the Research Council of the University of Delhi for the publication support. PJ and NS acknowledge the University Grants Commission, India, and NM, MR, and KS acknowledge the Council of Scientific and Industrial Research for Fellowships.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fpls.2021.628611/full#supplementary-material
References
Adusei-Fosu, K., and Dickinson, M. (2019). Development of pathogenicity assay and characterization of Fusarium oxysporum f. sp. elaeidis (FOE) based on Secreted In Xylem genes and EF-1α. J. Plant Pathol. 101, 1013–1024. doi: 10.1007/s42161-019-00332-4
Alexander, L. J., and Tucker, C. M. (1945). Physiological specialization in the Tomato wilt fungus Fusarium oxysporum f. sp. lycopersici. J. Agric. Res. 70, 303–313.
Alfano, J. R. (2009). Roadmap for future research on plant pathogen effectors. Mol. Plant Pathol. 10, 805–813. doi: 10.1111/j.1364-3703.2009.00588.x
An, B., Hou, X., Guo, Y., Zhao, S., Luo, H., He, C., et al. (2019). The effector six8 is required for virulence of Fusarium oxysporum f. sp. cubense tropical race 4 to cavendish banana. Fungal Biol. 123, 423–430. doi: 10.1016/j.funbio.2019.03.001
Armitage, A. D., Taylor, A., Sobczyk, M. K., Baxter, L., Greenfield, B. P., Bates, H. J., et al. (2018). Characterisation of pathogen-specific regions and novel effector candidates in Fusarium oxysporum f. sp. cepae. Sci. Rep. 8, 1–15. doi: 10.1038/s41598-018-30335-7
Armstrong, G. M., and Armstrong, J. K. (1981). “Formae speciales and races of Fusarium oxysporum causing wilt disease,” in Fusarium: Disease, Biology, and Taxonomy, eds P. E. Nelson, T. A. Toussoun, and R. J. Cook (University Park, PA: Pennsylvania State University Press), 391–399.
Ayukawa, Y., Asai, S., Gan, P., Tsushima, A., Ichihashi, Y., Shibata, A., et al. (2020). A pair of effectors encoded on a conditionally dispensable chromosome of Fusarium oxysporum suppress host-specific immunity. BioRxiv 329052. [preprint]. doi: 10.1101/2020.10.06.329052
Baayen, R. P. (2000). Diagnosis and detection of host-specific forms of Fusarium oxysporum. Bull OEPP 30, 489–491. doi: 10.1111/j.1365-2338.2000.tb00935.x
Baayen, R. P., O’Donnell, K., Bonants, P. J. M., Cigelnik, E., Kroon, L. P. N., Roebroeck, E. J. A., et al. (2000). Gene genealogies and AFLP analysis in the Fusarium oxysporum complex identify monophyletic and nonmonophyletic formae speciales causing wilt and rot disease. Phytopathology 90, 891–900. doi: 10.1094/PHYTO.2000.90.8.891
Batson, A. M., Fokkens, L., Rep, M., and du Toit, L. J. (2020). Putative effector genes distinguish two pathogenicity groups of Fusarium oxysporum f. sp. spinaciae. Mol. Plant Microbe Interact. 34, 141–156. doi: 10.1094/MPMI-06-20-0145-R
Beckman, C. H. (1987). The Nature of Wilt Diseases of Plants. St. Paul, MN: American Phytopathological Society Press.
Biju, V. C., Fokkens, L., Houterman, P. M., Rep, M., and Cornelissen, B. J. (2017). Multiple evolutionary trajectories have led to the emergence of races in Fusarium oxysporum f. sp. lycopersici. Appl. Environ. Microbiol. 83, 2548–2516e. doi: 10.1128/AEM.02548-16
Bishop, C. D., and Cooper, R. M. (1983). An ultrastructural study of root invasion in three vascular wilt diseases. Physiol. Plant Pathol. 22, 15–13. doi: 10.1016/0048-4059(83)90018-8
Bohn, G. W., and Tucker, C. M. (1939). Immunity to Fusarium wilt in the tomato. Science 89, 603–604. doi: 10.1126/science.89.2322.603
Borah, N., Albarouki, E., and Schirawski, J. (2018). Comparative methods for molecular determination of host-specificity factors in plant-pathogenic fungi. Int. J. Mol. Sci. 19:863. doi: 10.3390/ijms19030863
Brown, D. W., Busman, M., and Proctor, R. H. (2014). Fusarium verticillioides SGE1 is required for full virulence and regulates expression of protein effector and secondary metabolite biosynthetic genes. Mol. Plant Microbe Interact. 27, 809–823. doi: 10.1094/MPMI-09-13-0281-R
Cafri, D., Katan, J., and Katan, T. (2005). Cross−pathogenicity between formae speciales of Fusarium oxysporum, the pathogens of cucumber and melon. J. Phytopathol. 153, 615–622. doi: 10.1111/j.1439-0434.2005.01029.x
Cai, G., Gale, L. R., Schneider, R. W., Kistler, H. C., Davis, R. M., Elias, K. S., et al. (2003). Origin of race 3 of Fusarium oxysporum f. sp. lycopersici at a single site in California. Phytopathology 93, 1014–1022. doi: 10.1094/PHYTO.2003.93.8.1014
Cao, L., Blekemolen, M. C., Tintor, N., Cornelissen, B. J., and Takken, F. L. (2018). The Fusarium oxysporum Avr2-Six5 effector pair alters plasmodesmatal exclusion selectivity to facilitate cell-to-cell movement of Avr2. Mol. Plant 11, 691–705. doi: 10.1016/j.molp.2018.02.011
Carmona, S. L., Burbano-David, D., Gómez, M. R., Lopez, W., Ceballos, N., Castaño-Zapata, J., et al. (2020). Characterization of Pathogenic and Nonpathogenic Fusarium oxysporum Isolates Associated with Commercial Tomato Crops in the Andean Region of Colombia. Pathogens 9:70. doi: 10.3390/pathogens9010070
Catanzariti, A. M., Do, H. T., Bru, P., de Sain, M., Thatcher, L. F., Rep, M., et al. (2017). The tomato I gene for Fusarium wilt resistance encodes an atypical leucine−rich repeat receptor−like protein whose function is nevertheless dependent on SOBIR 1 and SERK 3/BAK 1. Plant J. 89, 1195–1209. doi: 10.1111/tpj.13458
Catanzariti, A. M., Lim, G. T., and Jones, D. A. (2015). The tomato I−3 gene: a novel gene for resistance to Fusarium wilt disease. N. Phytol. 207, 106–118. doi: 10.1111/nph.13348
Caten, C. E., and Jinks, J. L. (1966). Heterokaryosis: its significance in wild homothallic Ascomycetes and Fungi imperfecti. Trans. Br. Mycol. Soc. 49, 81–93. doi: 10.1016/S0007-1536(66)80038-4
Chakrabarti, A., Rep, M., Wang, B., Ashton, A., Dodds, P., and Ellis, J. (2011). Variation in potential effector genes distinguishing Australian and non−Australian isolates of the cotton wilt pathogen Fusarium oxysporum f. sp. vasinfectum. Plant Pathol. 60, 232–243. doi: 10.1111/j.1365-3059.2010.02363.x
Chang, W., Li, H., Chen, H., Qiao, F., and Zeng, H. (2020). Identification of mimp-associated effector genes in Fusarium oxysporum f. sp. cubense race 1 and race 4 and virulence confirmation of a candidate effector gene. Microbiol. Res. 232:126375. doi: 10.1016/j.micres.2019.126375
Chellappan, B. V. (2014). Evolution of Races within Fusarium oxysporum f. sp. lycopersici. Ph.D. thesis. Amsterdam: University of Amsterdam.
Chuma, I., Isobe, C., Hotta, Y., Ibaragi, K., Futamata, N., Kusaba, M., et al. (2011). Multiple translocation of the AVR-pita effector gene among chromosomes of the rice blast fungus Magnaporthe oryzae and related species. PLoS Pathog. 7:e1002147. doi: 10.1371/journal.ppat.1002147
Cohen, R., Orgil, G., Burger, Y., Saar, U., Elkabetz, M., Tadmor, Y., et al. (2015). Differences in the responses of melon accessions to fusarium root and stem rot and their colonization by F usarium oxysporum f. sp. radicis−cucumerinum. Plant Pathol. 64, 655–663. doi: 10.1111/ppa.12286
Coleman, J. J., Rounsley, S. D., Rodriguez-Carres, M., Kuo, A., Wasmann, C. C., Grimwood, J., et al. (2009). The genome of Nectria haematococca: contribution of supernumerary chromosomes to gene expansion. PLoS Genet. 5:e1000618. doi: 10.1371/journal.pgen.1000618
Correll, J. C. (1991). The relationship between formae speciales, races, and vegetative compatibility groups in Fusarium oxysporum. Phytopathology 81, 1061–1064.
Covey, P. A., Kuwitzky, B., Hanson, M., and Webb, K. M. (2014). Multilocus analysis using putative fungal effectors to describe a population of Fusarium oxysporum from sugar beet. Phytopathology 104, 886–896. doi: 10.1094/PHYTO-09-13-0248-R
Croll, D., and McDonald, B. A. (2012). The accessory genome as a cradle for adaptive evolution in pathogens. PLoS Pathog. 8:e1002608. doi: 10.1371/journal.ppat.1002608
Cuomo, C. A., Güldener, U., Xu, J. R., Trail, F., Turgeon, B. G., Di Pietro, A., et al. (2007). The Fusarium graminearum genome reveals a link between localized polymorphism and pathogen specialization. Science 317, 1400–1402. doi: 10.1126/science.1143708
Czislowski, E., Fraser−Smith, S., Zander, M., O’Neill, W. T., Meldrum, R. A., Tran−Nguyen, L. T., et al. (2018). Investigation of the diversity of effector genes in the banana pathogen, Fusarium oxysporum f. sp. cubense, reveals evidence of horizontal gene transfer. Mol. Plant Pathol. 19, 1155–1171. doi: 10.1111/mpp.12594
Dangl, J. L., and Jones, J. D. G. (2001). Plant pathogens and integrated defence responses to infection. Nature 411, 826–833. doi: 10.1038/35081161
Davière, J. M., Langin, T., and Daboussi, M. J. (2001). Potential role of transposable elements in the rapid reorganization of the Fusarium oxysporum genome. Fungal Genet. Biol. 34, 177–192. doi: 10.1006/fgbi.2001.1296
de Lamo, F. J., and Takken, F. L. W. (2020). Biocontrol by Fusarium oxysporum Using Endophyte-Mediated Resistance. Front. Plant Sci. 11:37. doi: 10.3389/fpls.2020.00037
de Vega-Bartol, J. J., Martín-Dominguez, R., Ramos, B., García-Sánchez, M. A., and Díaz-Mínguez, J. M. (2011). New virulence groups in Fusarium oxysporum f. sp. phaseoli: the expression of the gene coding for the transcription factor ftf1 correlates with virulence. Phytopathology 101, 470–479. doi: 10.1094/PHYTO-09-10-0252
De Wit, P. J. (2016). Apoplastic fungal effectors in historic perspective; a personal view. N. Phytol. 212, 805–813. doi: 10.1111/nph.14144
Dean, R., Van Kan, J. A., Pretorius, Z. A., Hammond−Kosack, K. E., Di Pietro, A., Spanu, P. D., et al. (2012). The Top 10 fungal pathogens in molecular plant pathology. Mol. Plant Pathol. 13, 414–430. doi: 10.1111/j.1364-3703.2011.00783.x
DeIulio, G. A., Guo, L., Zhang, Y., Goldberg, J. M., Kistler, H. C., and Ma, L. J. (2018). Kinome expansion in the Fusarium oxysporum species complex driven by accessory chromosomes. Msphere. 3, 231–218e. doi: 10.1128/mSphere.00231-18
Di Pietro, A., García−Maceira, F. I., Méglecz, E., and Roncero, M. I. G. (2001). A MAP kinase of the vascular wilt fungus Fusarium oxysporum is essential for root penetration and pathogenesis. Mol. Microbiol. 39, 1140–1152. doi: 10.1111/j.1365-2958.2001.02307.x
Di, X., Cao, L., Hughes, R. K., Tintor, N., Banfield, M. J., and Takken, F. L. (2017). Structure–function analysis of the Fusarium oxysporum Avr2 effector allows uncoupling of its immune−suppressing activity from recognition. N. Phytol. 216, 897–914. doi: 10.1111/nph.14733
Dobbs, J. T., Kim, M. S., Dudley, N. S., Klopfenstein, N. B., Yeh, A., Hauff, R. D., et al. (2020). Whole genome analysis of the koa wilt pathogen (Fusarium oxysporum f. sp. koae) and the development of molecular tools for early detection and monitoring. BMC Genomics 21:1–15. doi: 10.1186/s12864-020-07156-y
Dong, S., Raffaele, S., and Kamoun, S. (2015). The two-speed genomes of filamentous pathogens: waltz with plants. Curr. Opin. Genet. Dev. 35, 57–65. doi: 10.1016/j.gde.2015.09.001
Duan, Y., Qu, W., Chang, S., Li, C., Xu, F., Ju, M., et al. (2020). Identification of Pathogenicity Groups and Pathogenic Molecular Characterization of Fusarium oxysporum f. sp. sesami in China. Phytopathology 110, 1093–1104. doi: 10.1094/PHYTO-09-19-0366-R
Edel-Hermann, V., and Lecomte, C. (2019). Current status of Fusarium oxysporum formae speciales and races. Phytopathology 109, 512–530. doi: 10.1094/PHYTO-08-18-0320-RVW
Epstein, L., Kaur, S., Chang, P. L., Carrasquilla-Garcia, N., Lyu, G., Cook, D. R., et al. (2017). Races of the celery pathogen Fusarium oxysporum f. sp. apii are polyphyletic. Phytopathology 107, 463–473. doi: 10.1094/PHYTO-04-16-0174-R
Flor, H. H. (1971). Current status of the gene-for-gene concept. Annu. Rev. Phytopathol. 9, 275–296. doi: 10.1146/annurev.py.09.090171.001423
Fokkens, L., Shahi, S., Connolly, L. R., Stam, R., Schmidt, S. M., Smith, K. M., et al. (2018). The multi-speed genome of Fusarium oxysporum reveals association of histone modifications with sequence divergence and footprints of past horizontal chromosome transfer events. BioRxiv 465070. [preprint]. doi: 10.1101/465070
Fourie, G., Steenkamp, E. T., Gordon, T. R., and Viljoen, A. (2009). Evolutionary relationships among the Fusarium oxysporum f. sp. cubense vegetative compatibility groups. Appl. Environ. Microbiol. 75, 4770–4781. doi: 10.1128/AEM.00370-09
Fraser-Smith, S., Czislowski, E., Meldrum, R. A., Zander, M., O’neill, W., Balali, G. R., et al. (2014). Sequence variation in the putative effector gene SIX 8 facilitates molecular differentiation of Fusarium oxysporum f. sp. cubense. Plant Pathol. 63, 1044–1052. doi: 10.1111/ppa.12184
Fujinaga, M., Ogiso, H., Shinohara, H., Tsushima, S., Nishimura, N., Togawa, M., et al. (2005). Phylogenetic relationships between the lettuce root rot pathogen Fusarium oxysporum f. sp. lactucae races 1, 2, and 3 based on the sequence of the intergenic spacer region of its ribosomal DNA. J. Gen. Plant Pathol. 71, 402–407. doi: 10.1007/s10327-005-0226-z
Gawehns, F., Houterman, P. M., Ichou, F. A., Michielse, C. B., Hijdra, M., Cornelissen, B. J. C., et al. (2014). The Fusarium oxysporum effector Six6 contributes to virulence and suppresses I-2-mediated cell death. Mol. Plant Microbe Interact. 27, 336–348. doi: 10.1094/MPMI-11-13-0330-R
Gonzalez-Cendales, Y., Catanzariti, A. M., Baker, B., Mcgrath, D. J., and Jones, D. A. (2016). Identification of I−7 expands the repertoire of genes for resistance to Fusarium wilt in tomato to three resistance gene classes. Mol. Plant Pathol. 17, 448–463. doi: 10.1111/mpp.12294
Gordon, T. R. (2017). Fusarium oxysporum and the Fusarium wilt syndrome. Annu. Rev. Phytopathol. 55, 23–39. doi: 10.1146/annurev-phyto-080615-095919
Gordon, T. R., and Martyn, R. D. (1997). The evolutionary biology of Fusarium oxysporum. Annu. Rev. Phytopathol. 35, 111–128. doi: 10.1146/annurev.phyto.35.1.111
Gordon, W. L. (1965). Pathogenic strains of Fusarium oxysporum. Can. J. Bot. 43, 1309–1318. doi: 10.1139/b65-138
Guo, L., Han, L., Yang, L., Zeng, H., Fan, D., Zhu, Y., et al. (2014). Genome and transcriptome analysis of the fungal pathogen Fusarium oxysporum f. sp. cubense causing banana vascular wilt disease. PLoS One 9:e95543. doi: 10.1371/journal.pone.0095543
Guo, L., Yang, L., Liang, C., Wang, J., Liu, L., and Huang, J. (2016). The G-protein subunits FGA2 and FGB1 play distinct roles in development and pathogenicity in the banana fungal pathogen Fusarium oxysporum f. sp. cubense. Physiol. Mol. Plant Pathol. 93, 29–38. doi: 10.1016/j.pmpp.2015.12.003
Henry, P., Kaur, S., Pham, Q. A. T., Barakat, R., Brinker, S., Haensel, H., et al. (2020). Genomic differences between the new Fusarium oxysporum f. sp. apii (Foa) race 4 on celery, the less virulent Foa races 2 and 3, and the avirulent on celery f. sp. coriandrii. BMC Genomics 21:1–23. doi: 10.1186/s12864-020-07141-5
Hill, A. L., Reeves, P. A., Larson, R. L., Fenwick, A. L., Hanson, L. E., and Panella, L. (2011). Genetic variability among isolates of Fusarium oxysporum from sugar beet. Plant Pathol. 60, 496–505. doi: 10.1111/j.1365-3059.2010.02394.x
Hogenhout, S. A., Van der Hoorn, R. A., Terauchi, R., and Kamoun, S. (2009). Emerging concepts in effector biology of plant-associated organisms. Mol. Plant Microbe Interact. 22, 115–122. doi: 10.1094/MPMI-22-2-0115
Houterman, P. M., Cornelissen, B. J., and Rep, M. (2008). Suppression of plant resistance gene-based immunity by a fungal effector. PLoS Pathog. 4:e1000061. doi: 10.1371/journal.ppat.1000061
Houterman, P. M., Ma, L., Van Ooijen, G., De Vroomen, M. J., Cornelissen, B. J., Takken, F. L., et al. (2009). The effector protein Avr2 of the xylem-colonizing fungus Fusarium oxysporum activates the tomato resistance protein I-2 intracellularly. Plant J. 58, 970–978. doi: 10.1111/j.1365-313X.2009.03838.x
Houterman, P. M., Speijer, D., Dekker, H. L., de Koster, C. G., Cornelissen, B. J., and Rep, M. (2007). The mixed xylem sap proteome of Fusarium oxysporum−infected tomato plants. Mol. Plant Pathol. 8, 215–221. doi: 10.1111/j.1364-3703.2007.00384.x
Hu, G., de Hart, A. K., Li, Y., Ustach, C., Handley, V., Navarre, R., et al. (2005). EDS1 in tomato is required for resistance mediated by TIR−class R genes and the receptor−like R gene Ve. Plant J. 42, 376–391. doi: 10.1111/j.1365-313X.2005.02380.x
Husaini, A. M., Sakina, A., and Cambay, S. R. (2018). Host–pathogen interaction in Fusarium oxysporum infections: Where do we stand? Mol. Plant Microbe Interact. 31, 889–898. doi: 10.1094/MPMI-12-17-0302-CR
Imazaki, I., Kurahashi, M., Iida, Y., and Tsuge, T. (2007). Fow2, a Zn (II) 2Cys6−type transcription regulator, controls plant infection of the vascular wilt fungus Fusarium oxysporum. Mol. Microbiol. 63, 737–753. doi: 10.1111/j.1365-2958.2006.05554.x
Jain, S., Akiyama, K., Kan, T., Ohguchi, T., and Takata, R. (2003). The Gprotein beta subunit FGB1 regulates development and pathogenicity in Fusarium oxysporum. Curr. Genet. 43, 79–86. doi: 10.1007/s00294-003-0372-9
Jain, S., Akiyama, K., Mae, K., Ohguchi, T., and Takata, R. (2002). Targeted disruption of a G protein alpha subunit gene results in reducedpathogenicity in Fusarium oxysporum. Curr. Genet. 41, 407–413. doi: 10.1007/s00294-002-0322-y
Jain, S., Akiyama, K., Takata, R., and Ohguchi, T. (2005). Signaling via the G protein α subunit FGA2 is necessary for pathogenesis in Fusarium oxysporum. FEMS Microbiol. Lett. 243, 165–172. doi: 10.1016/j.femsle.2004.12.009
Jiménez-Gasco, M. M., Milgroom, M. G., and Jiménez-Díaz, R. M. (2002). Gene genealogies support Fusarium oxysporum f. sp. ciceris as a monophyletic group. Plant Pathol. 51, 72–77. doi: 10.1046/j.0032-0862.2001.00610.x-i1
Joobeur, T., King, J. J., Nolin, S. J., Thomas, C. E., and Dean, R. A. (2004). The Fusarium wilt resistance locus FOM-2 of melon contains a single resistance gene with complex features. Plant J. 39, 283–297. doi: 10.1111/j.1365-313X.2004.02134.x
Joshi, R. (2018). A review of Fusarium oxysporum on its plant interaction and industrial use. J. Med. Plant. 6, 112–115. doi: 10.22271/plants.2018.v6.i3b.07
Kang, S., Demers, J., del Mar, Jimenez-Gasco, M., and Rep, M. (2014). “Fusarium oxysporum,” in Genomics of Plant-Associated Fungi and Oomycetes: Dicot Pathogens, eds A. Lichens-Park, C. Kole, and R. A. Dean (Berlin: Springer), 99–119.
Kashiwa, T., Inami, K., Fujinaga, M., Ogiso, H., Yoshida, T., Teraoka, T., et al. (2013). An avirulence gene homologue in the tomato wilt fungus Fusarium oxysporum f. sp. lycopersici race 1 functions as a virulence gene in the cabbage yellows fungus F. oxysporum f. sp. conglutinans. J. Gen. Plant Pathol. 79, 412–421. doi: 10.1007/s10327-013-0471-5
Kawabe, M., Kobayashi, Y., Okada, G., Yamaguchi, I., Teraoka, T., and Arie, T. (2005). Three evolutionary lineages of tomato wilt pathogen, Fusarium oxysporum f. sp. lycopersici, based on sequences of IGS, MAT1, and pg1, are each composed of isolates of a single mating type and a single or closely related vegetative compatibility group. J. Gen. Plant. Pathol. 71, 263–272. doi: 10.1007/s10327-005-0203-6
Kistler, H. C. (1997). Genetic diversity in the plant-pathogenic fungus Fusarium oxysporum. Phytopathology 87, 474–479. doi: 10.1094/PHYTO.1997.87.4.474
Kistler, H. C., Benny, U., Boehm, E. W. A., and Katan, T. (1995). Genetic duplication in Fusarium oxysporum. Curr. Genet. 28, 173–176. doi: 10.1007/BF00315784
Kuang, H., Padmanabhan, C., Li, F., Kamei, A., Bhaskar, P. B., Ouyang, S., et al. (2009). Identification of miniature inverted-repeat transposable elements (MITEs) and biogenesis of their siRNAs in the Solanaceae: new functional implications for MITEs. Genome Res. 19, 42–56. doi: 10.1101/gr.078196.108
Kuldau, G. A., and Yates, I. E. (2000). “Evidence for Fusarium endophytes,” in Microbial Endophytes, eds C. W. Bacon and J. F. White (New York, NY: Marcel Dekker, Inc.), 85–117.
Laurence, M. H., Summerell, B. A., and Liew, E. C. Y. (2015). Fusarium oxysporum f. sp. canariensis: evidence for horizontal gene transfer of putative pathogenicity genes. Plant Pathol. 64, 1068–1075. doi: 10.1111/ppa.12350
Leslie, J. F., and Summerell, B. A. (2008). The Fusarium laboratory manual. Hoboken, NJ: John Wiley & Sons.
Li, E., Wang, G., Xiao, J., Ling, J., Yang, Y., and Xie, B. (2016). A SIX1 homolog in Fusarium oxysporum f. sp. conglutinans is required for full virulence on cabbage. PLoS One 11:e0152273. doi: 10.1371/journal.pone.0152273
Li, J., Fokkens, L., and Rep, M. (2021). A single gene in Fusarium oxysporum limits host range. Mol. Plant Pathol. 22, 108–116. doi: 10.1111/mpp.13011
Lievens, B., Claes, L., Vakalounakis, D. J., Vanachter, A. C., and Thomma, B. P. (2007). A robust identification and detection assay to discriminate the cucumber pathogens Fusarium oxysporum f. sp. cucumerinum and f. sp. radicis−cucumerinum. Environ. Microbiol. 9, 2145–2161. doi: 10.1111/j.1462-2920.2007.01329.x
Lievens, B., Houterman, P. M., and Rep, M. (2009a). Effector gene screening allows unambiguous identification of Fusarium oxysporum f. sp. lycopersici races and discrimination from other formae speciales. FEMS Microbiol. Lett. 300, 201–215. doi: 10.11a11/j.1574-6968.2009.01783z.x
Lievens, B., Rep, M., and Thomma, B. P. (2008). Recent developments in the molecular discrimination of formae speciales of Fusarium oxysporum. Pest Manag. Sci. 64, 781–788. doi: 10.1002/ps.1564
Lievens, B., Van Baarlen, P., Verreth, C., Van Kerckhove, S., Rep, M., and Thomma, B. P. (2009b). Evolutionary relationships between Fusarium oxysporum f. sp. lycopersici and F. oxysporum f. sp. radicis-lycopersici isolates inferred from mating type, elongation factor-1α and exopolygalacturonase sequences. Mycol. Res. 113, 1181–1191. doi: 10.1016/j.mycres.2009.07.019
Lim, G. T. T., Wang, G. P., Hemming, M. N., Basuki, S., McGrath, D. J., Carroll, B. J., et al. (2006). Mapping the I-3 gene for resistance to Fusarium wilt in tomato: application of an I-3 marker in tomato improvement and progress towards the cloning of I-3. Australas. Plant Pathol. 35, 671–680. doi: 10.1071/AP06073
Lim, G. T. T., Wang, G. P., Hemming, M. N., McGrath, D. J., and Jones, D. A. (2008). High resolution genetic and physical mapping of the I-3 region of tomato chromosome 7 reveals almost continuous microsynteny with grape chromosome 12 but interspersed microsynteny with duplications on Arabidopsis chromosomes 1, 2 and 3. Theor. Appl. Genet. 118, 57–75. doi: 10.1007/s00122-008-0876-2
Liu, Z., Zhang, X., Liu, X., Fu, C., Han, X., Yin, Y., et al. (2016). The chitin synthase FgChs2 and other FgChss co-regulate vegetative development and virulence in F. graminearum. Sci. Rep. 6, 1–12. doi: 10.1038/srep34975
Lyons, R., Czislowski, E., Zeil-Rolfe, I., Kaur, S., Chen, A., et al. (2019). Unique Secreted in Xylem Genes in Banana-Infecting Endophytic Fusarium oxysporum. MDPI 36:180. doi: 10.3390/proceedings2019036180
Ma, L. J., Geiser, D. M., Proctor, R. H., Rooney, A. P., O’Donnell, K., Trail, F., et al. (2013). Fusarium pathogenomics. Annu. Rev. Microbiol. 67, 399–416. doi: 10.1146/annurev-micro-092412-155650
Ma, L., Cornelissen, B. J. C., and Takken, F. L. W. (2013). A nuclear localization for Avr2 from Fusarium oxysporum is required to activate the tomato resistance protein I-2. Front. Plant Sci. 4:94. doi: 10.3389/fpls.2013.00094
Ma, L. J., Shea, T., Young, S., Zeng, Q., and Kistler, H. C. (2014). Genome sequence of Fusarium oxysporum f. sp. melonis strain NRRL 26406, a fungus causing wilt disease on melone. Genome Announc. 2, 730–714e. doi: 10.1128/genomeA.00730-14
Ma, L. J., Van Der Does, H. C., Borkovich, K. A., Coleman, J. J., Daboussi, M. J., Di Pietro, A., et al. (2010). Comparative genomics reveals mobile pathogenicity chromosomes in Fusarium. Nature 464, 367–373. doi: 10.1038/nature08850
Ma, L., Houterman, P. M., Gawehns, F., Cao, L., Sillo, F., Richter, H., et al. (2015). The AVR2–SIX5 gene pair is required to activate I−2−mediated immunity in tomato. N. Phytol. 208, 507–518. doi: 10.1111/nph.13455
Maldonado, B. L. D., Villarruel, O. J. L., Calderón, O. M. A., and Sánchez, E. A. C. (2018). Secreted in Xylem (Six) Genes in Fusarium oxysporum f. sp. cubense and Their Potential Acquisition by Horizontal Transfer. Adv. Biotech. Micro. 10:555779. doi: 10.19080/AIBM.2018.10.555778
Mandeel, Q., and Baker, R. (1991). Mechanisms involved in biological control of Fusarium wilt of cucumber with strains of nonpathogenic Fusarium oxysporum. Phytopathology 81, 462–469. doi: 10.1094/phyto-81-462
Mandeel, Q., Ayub, N., and Gul, J. (2005). Survey of Fusarium species in an environment of Bahrain. VI. Biodiversity of the genus Fusarium in root-soil ecosystem of halophytic date palm (Phoenix dactylifera) community. Cryptogamie Mycol. 26, 365–404.
McGrath, D., Gillespie, D., and Vawdrey, L. (1987). Inheritance of resistance to Fusarium oxysporum f. sp. lycopersici races 2 and 3 in Lycopersicon pennellii. Aust. J. Agric. Res. 38, 729–733. doi: 10.1071/AR9870729
Meldrum, R. A., Fraser-Smith, S., Tran-Nguyen, L. T. T., Daly, A. M., and Aitken, E. A. B. (2012). Presence of putative pathogenicity genes in isolates of Fusarium oxysporum f. sp. cubense from Australia. Australas. Plant Pathol. 41, 551–557. doi: 10.1007/s13313-012-0122-x
Mes, J. J., van Doorn, A. A., Wijbrandi, J., Simons, G., Cornelissen, B. J., and Haring, M. A. (2000). Expression of the Fusarium resistance gene I-2 colocalizes with the site of fungal containment. Plant J. 23, 183–193. doi: 10.1046/j.1365-313x.2000.00765.x
Michielse, C. B., and Rep, M. (2009). Pathogen profile update: Fusarium oxysporum. Mol. Plant Pathol. 10:311. doi: 10.1111/j.1364-3703.2009.00538.x
Michielse, C. B., van Wijk, R., Reijnen, L., Cornelissen, B. J., and Rep, M. (2009a). Insight into the molecular requirements for pathogenicity of Fusarium oxysporum f. sp. lycopersici through large-scale insertional mutagenesis. Genome Biol. 10:R4. doi: 10.1186/gb-2009-10-1-r4
Michielse, C. B., van Wijk, R., Reijnen, L., Manders, E. M., Boas, S., Olivain, C., et al. (2009b). The nuclear protein Sge1 of Fusarium oxysporum is required for parasitic growth. PLoS Pathog. 5:e1000637. doi: 10.1371/journal.ppat.1000637
Moore, N. Y., Pegg, K. G., Allen, R. N., and Irwin, J. A. G. (1993). Vegetative compatibility and distribution of Fusarium oxysporum f. sp. cubense in Australia. Aus. J. Exp. Agric. 33, 797–802. doi: 10.1071/EA9930797
Namiki, F., Shiomi, T., Kayamura, T., and Tsuge, T. (1994). Characterization of the formae speciales of Fusarium oxysporum causing wilts of cucurbits by DNA fingerprinting with nuclear repetitive DNA sequences. Appl. Environ. Microbiol. 60, 2684–2691. doi: 10.1128/aem.60.8.2684-2691.1994
Niño-Sánchez, J., Casado-Del Castillo, V., Tello, V., De Vega-Bartol, J. J., Ramos, B., Sukno, S. A., et al. (2016). The FTF gene family regulates virulence and expression of SIX effectors in Fusarium oxysporum. Mol. Plant Pathol. 17, 1124–1139. doi: 10.1111/mpp.12373
Niño-Sánchez, J., Tello, V., Casado-del Castillo, V., Thon, M. R., Benito, E. P., and Díaz-Mínguez, J. M. (2015). Gene expression patterns and dynamics of the colonization of common bean (Phaseolus vulgaris L.) by highly virulent and weakly virulent strains of Fusarium oxysporum. Front. Microbiol. 6:234. doi: 10.3389/fmicb.2015.00234
Nirmaladevi, D., Venkataramana, M., Srivastava, R. K., Uppalapati, S. R., Gupta, V. K., Yli-Mattila, T., et al. (2016). Molecular phylogeny, pathogenicity and toxigenicity of Fusarium oxysporum f. sp. lycopersici. Sci. Rep. 6, 1–14. doi: 10.1038/srep21367
Niu, X., Zhao, X., Ling, K. S., Levi, A., Sun, Y., and Fan, M. (2016). The FonSIX6 gene acts as an avirulence effector in the Fusarium oxysporum f. sp. niveum-watermelon pathosystem. Sci. Rep. 6, 1–7. doi: 10.1038/srep28146
O’Donnell, K., and Cigelnik, E. (1997). Two divergent intragenomic rDNA ITS2 types within a monophyletic lineage of the fungus Fusarium are nonorthologous. Mol. Phylogenet. Evol. 7, 103–116. doi: 10.1006/mpev.1996.0376
O’Donnell, K., Kistler, H. C., Cigelnik, E., and Ploetz, R. C. (1998). Multiple evolutionary origins of the fungus causing Panama disease of banana: concordant evidence from nuclear and mitochondrial gene genealogies. Proc. Natl. Acad. Sci. U S A. 95, 2044–2049. doi: 10.1073/pnas.95.5.2044
O’Donnell, K., Rooney, A. P., Proctor, R. H., Brown, D. W., McCormick, S. P., Ward, T. J., et al. (2013). Phylogenetic analyses of RPB1 and RPB2 support a middle Cretaceous origin for a clade comprising all agriculturally and medically important fusaria. Fungal Genet. Biol. 52, 20–31. doi: 10.1016/j.fgb.2012.12.004
O’Donnell, K., Gherbawy, Y., Schweigkofler, W., Adler, A., and Prillinger, H. (1999). Phylogenetic analyses of DNA sequence and RAPD data compared in Fusarium oxysporum and related species from maize. J. Phytopathol. 147, 445–452. doi: 10.1111/j.1439-0434.1999.tb03849.x
Ogiso, H., Fujinaga, M., Saito, H., Takehara, T., and Yamanaka, S. (2002). Physiological races and vegetative compatibility groups of Fusarium oxysporum f. sp. lactucae isolated from crisphead lettuce in Japan. J. Gen. Plant Pathol. 68, 292–299. doi: 10.1007/PL00013094
Olivain, C., and Alabouvette, C. (1999). Process of tomato root colonization by a pathogenic strain of Fusarium oxysporum f. sp. lycopersici in comparison with a non−pathogenic strain. N. Phytol. 141, 497–510. doi: 10.1046/j.1469-8137.1999.00365.x
Palmero, D., Iglesias, C., de Cara, M., Lomas, T., Santos, M., and Tello, J. C. (2009). Species of Fusarium isolated from river and sea water of southeastern Spain and pathogenicity on four plant species. Plant Dis. 93, 377–385. doi: 10.1094/PDIS-93-4-0377
Pasquali, M., Dematheis, F., Gilardi, G., Gullino, M. L., and Garibaldi, A. (2005). Vegetative compatibility groups of Fusarium oxysporum f. sp. lactucae from lettuce. Plant Dis. 89, 237–240. doi: 10.1094/PD-89-0237
Pietro, A. D., Madrid, M. P., Caracuel, Z., Delgado-Jarana, J., and Roncero, M. I. G. (2003). Fusarium oxysporum: exploring the molecular arsenal of a vascular wilt fungus. Mol. Plant Pathol. 4, 315–325. doi: 10.1046/j.1364-3703.2003.00180.x
Pinaria, A. G., Laurence, M. H., Burgess, L. W., and Liew, E. C. Y. (2015). Phylogeny and origin of Fusarium oxysporum f. sp. vanillae in Indonesia. Plant Pathol. 64, 1358–1365. doi: 10.1111/ppa.12365
Ploetz, R. C., and Correll, J. C. (1988). Vegetative compatibility among races of Fusarium oxysporum f.sp. cubense. Plant Dis. 72, 325–328. doi: 10.1094/pd-72-0325
Ponukumati, S. V., Elliott, M. L., and Des Jardin, E. A. (2019). Comparison of Secreted in Xylem (SIX) genes in two fusarium wilt pathogens of ornamental palms. Plant Pathol. 68, 1663–1681. doi: 10.1111/ppa.13090
Pradhan, A., Ghosh, S., Sahoo, D., and Jha, G. (2020). Fungal effectors, the double edge sword of phytopathogens. Curr. Genet. 67, 27–40. doi: 10.1007/s00294-020-01118-3
Puhalla, J. E. (1985). Classification of strains of Fusarium oxysporum on the basis of vegetative compatibility. Can. J. Bot. 63, 179–183. doi: 10.1139/b85-020
Punja, Z. K., and Parker, M. (2000). Development of fusarium root and stem rot, a new disease on greenhouse cucumber in British Columbia, caused by Fusarium oxysporum f. sp. radicis-cucumerinum. Can. J. Plant Pathol. 22, 349–363. doi: 10.1080/07060660009500453
Raffaele, S., and Kamoun, S. (2012). Genome evolution in filamentous plant pathogens: why bigger can be better. Nat. Rev. Microbiol. 10, 417–430. doi: 10.1038/nrmicro2790
Rana, A., Sahgal, M., and Johri, B. N. (2017). “Fusarium oxysporum: genomics, diversity and plant–host interaction,” in Developments in Fungal Biology and Applied Mycology, eds S. K. Deshmukh, T. Satyanarayana, and B. N. Johri (Singapore: Springer), 159–199. doi: 10.1007/978-981-10-4768-8_10
Rao, V. S., Srinivas, K., Sujini, G. N., and Kumar, G. N. (2014). Protein-protein interaction detection: methods and analysis. Int. J. Proteom. 2014:147648. doi: 10.1155/2014/147648
Recorbet, G., Steinberg, C., Olivain, C., Edel, V., Trouvelot, S., Dumas-Gaudot, E., et al. (2003). Wanted: pathogenesis−related marker molecules for Fusarium oxysporum. N. Phytol. 159, 73–92. doi: 10.1046/j.1469-8137.2003.00795.x
Rep, M. (2005). Small proteins of plant-pathogenic fungi secreted during host colonization. FEMS Microbiol. Lett. 253, 19–27. doi: 10.1016/j.femsle.2005.09.014
Rep, M., and Kistler, H. C. (2010). The genomic organization of plant pathogenicity in Fusarium species. Curr. Opin. Plant Biol. 13, 420–426. doi: 10.1016/j.pbi.2010.04.004
Rep, M., Dekker, H. L., Vossen, J. H., de Boer, A. D., Houterman, P. M., Speijer, D., et al. (2002). Mass spectrometric identification of isoforms of PR proteins in xylem sap of fungus-infected tomato. Plant Physiol. 130, 904–917. doi: 10.1104/pp.007427
Rep, M., Meijer, M., Houterman, P. M., van der Does, H. C., and Cornelissen, B. J. (2005). Fusarium oxysporum evades I-3-mediated resistance without altering the matching avirulence gene. Mol. Plant Microbe Interact. 18, 15–23. doi: 10.1094/MPMI-18-0015
Rep, M., Van Der Does, H. C., Meijer, M., Van Wijk, R., Houterman, P. M., Dekker, H. L., et al. (2004). A small, cysteine−rich protein secreted by Fusarium oxysporum during colonization of xylem vessels is required for I−3−mediated resistance in tomato. Mol. Microbiol. 53, 1373–1383. doi: 10.1111/j.1365-2958.2004.04177.x
Rocha, L. O., Laurence, M. H., Ludowici, V. A., Puno, V. I., Lim, C. C., Tesoriero, L. A., et al. (2016). Putative effector genes detected in Fusarium oxysporum from natural ecosystems of Australia. Plant Pathol. 65, 914–929. doi: 10.1111/ppa.12472
Sasaki, K., Nakahara, K., Tanaka, S., Shigyo, M., and Ito, S. I. (2015). Genetic and pathogenic variability of Fusarium oxysporum f. sp. cepae isolated from onion and Welsh onion in Japan. Phytopathology 105, 525–532. doi: 10.1094/PHYTO-06-14-0164-R
Schlechtendal, D. F. L. (1824). Flora Berolinensis: Pars secunda: cryptogamia 2. Berolini: Sumtibus Ferdinandi Dümmler.
Schmidt, S. M., Houterman, P. M., Schreiver, I., Ma, L., Amyotte, S., Chellappan, B., et al. (2013). MITEs in the promoters of effector genes allow prediction of novel virulence genes in Fusarium oxysporum. BMC Genomics 14:119. doi: 10.1186/1471-2164-14-119
Schmidt, S. M., Lukasiewicz, J., Farrer, R., van Dam, P., Bertoldo, C., and Rep, M. (2016). Comparative genomics of Fusarium oxysporum f. sp. melonis reveals the secreted protein recognized by the Fom−2 resistance gene in melon. N. Phytol. 209, 307–318. doi: 10.1111/nph.13584
Scott, J. W., and Jones, J. P. (1989). Monogenic resistance in tomato to Fusarium oxysporum f. sp. lycopersici race 3. Euphytica 40, 49–53. doi: 10.1007/BF00023296
Simbaqueba, J., Catanzariti, A. M., González, C., and Jones, D. A. (2018). Evidence for horizontal gene transfer and separation of effector recognition from effector function revealed by analysis of effector genes shared between cape gooseberry and tomato−infecting formae speciales of Fusarium oxysporum. Mol. Plant Pathol. 19, 2302–2318. doi: 10.1111/mpp.12700
Simons, G., Groenendijk, J., Wijbrandi, J., Reijans, M., Groenen, J., Diergaarde, P., et al. (1998). Dissection of the Fusarium I-2 gene cluster in tomato reveals six homologs and one active gene copy. Plant Cell 10, 1055–1068. doi: 10.1105/tpc.10.6.1055
Skovgaard, K., Nirenberg, H. I., O’Donnell, K., and Rosendahl, S. (2001). Evolution of Fusarium oxysporum f. sp. vasinfectum races inferred from multigene genealogies. Phytopathology 91, 1231–1237. doi: 10.1094/PHYTO.2001.91.12.1231
Snyder, W. C., and Hansen, H. N. (1940). The species concept in Fusarium. Am. J. Bot. 27, 64–67. doi: 10.2307/2436688
Stall, R. E., and Walter, J. M. (1965). Selection and inheritance of resistance in tomato to isolates of races 1 and 2 of the Fusarium wilt organism. Phytopathology 55, 1213–1215.
Takken, F., and Rep, M. (2010). The arms race between tomato and Fusarium oxysporum. Mol. Plant Pathol. 11, 309–314. doi: 10.1111/j.1364-3703.2009.00605.x
Taylor, A., Armitage, A. D., Handy, C., Jackson, A. C., Hulin, M. T., Harrison, R. J., et al. (2019). Basal Rot of Narcissus: Understanding Pathogenicity in Fusarium oxysporum f. sp. naricissi. Front. Microbiol. 10:2905. doi: 10.3389/fmicb.2019.02905
Taylor, A., Vágány, V., Jackson, A. C., Harrison, R. J., Rainoni, A., and Clarkson, J. P. (2016). Identification of pathogenicity−related genes in Fusarium oxysporum f. sp. cepae. Mol. Plant Pathol. 17, 1032–1047. doi: 10.1111/mpp.12346
Thatcher, L. F., Gardiner, D. M., Kazan, K., and Manners, J. M. (2012). A highly conserved effector in Fusarium oxysporum is required for full virulence on Arabidopsis. Mol. Plant Microbe Interact. 25, 180–190. doi: 10.1094/MPMI-08-11-0212
Vakalounakis, D. J. (1996). Root and Stem Rot of Cucumber Caused by Fusarium oxysporum f. sp. radicis-cucumerinum f. sp. nov. Plant Dis. 80, 313–316.
van Dam, P., and Rep, M. (2017). The Distribution of Miniature Impala Elements and SIX Genes in the Fusarium Genus is Suggestive of Horizontal Gene Transfer. J. Mol. Evol. 85, 14–25. doi: 10.1007/s00239-017-9801-0
van Dam, P., Fokkens, L., Ayukawa, Y., van der Gragt, M., Ter Horst, A., Brankovics, B., et al. (2017). A mobile pathogenicity chromosome in Fusarium oxysporum for infection of multiple cucurbit species. Sci. Rep. 7:9042. doi: 10.1038/s41598-017-07995-y
van Dam, P., Fokkens, L., Schmidt, S. M., Linmans, J. H., Kistler, H. C., Ma, L. J., et al. (2016). Effector profiles distinguish formae speciales of Fusarium oxysporum. Environ. Microbiol. 18, 4087–4102. doi: 10.1111/1462-2920.13445
Van der Biezen, E. A., and Jones, J. D. G. (1998). Plant disease resistance proteins and the gene-for-gene concept. Trends Plant Sci. 23, 454–456. doi: 10.1016/s0968-0004(98)01311-5
van der Does, H. C., and Rep, M. (2007). Virulence genes and the evolution of host specificity in plant-pathogenic fungi. Mol. Plant Microbe Interact. 20, 1175–1182. doi: 10.1094/mpmi-20-10-1175
Van Der Does, H. C., Fokkens, L., Yang, A., Schmidt, S. M., Langereis, L., Lukasiewicz, J. M., et al. (2016). Transcription Factors Encoded on Core and Accessory Chromosomes of Fusarium oxysporum Induce Expression of Effector Genes. PLoS Genet. 12:e1006401. doi: 10.1371/journal.pgen.1006401
Van Der Does, H. C., Lievens, B., Claes, L., Houterman, P. M., Cornelissen, B. J., and Rep, M. (2008). The presence of a virulence locus discriminates Fusarium oxysporum isolates causing tomato wilt from other isolates. Environ. Microbiol. 10, 1475–1485. doi: 10.1111/j.1462-2920.2007.01561.x
van der Hoorn, R. A., and Kamoun, S. (2008). From guard to decoy: a new model for perception of plant pathogen effectors. Plant Cell 20, 2009–2017. doi: 10.1105/tpc.108.060194
VanderMolen, G. E., Beckman, C. H., and Rodehorst, E. (1987). The ultrastructure of tylose formation in resistant banana following inoculation with Fusarium oxysporum f. sp. cubense. Physiol. Mol. Plant Pathol. 31, 185–200. doi: 10.1016/0885-5765(87)90063-4
Vlaardingerbroek, I., Beerens, B., Rose, L., Fokkens, L., Cornelissen, B. J., and Rep, M. (2016a). Exchange of core chromosomes and horizontal transfer of lineage-specific chromosomes in Fusarium oxysporum. Environ. Microbiol. 18, 3702–3713. doi: 10.1111/1462-2920.13281
Vlaardingerbroek, I., Beerens, B., Schmidt, S. M., Cornelissen, B. J., and Rep, M. (2016b). Dispensable chromosomes in Fusarium oxysporum f. sp. lycopersici. Mol. Plant Pathol. 17, 1455–1466. doi: 10.1111/mpp.12440
Weimer, J. L. (1944). Some root rots and a foot rot of lupines in the Southeastern part of the Unites States. J. Agric. Res. 68:441.
Widinugraheni, S., Niño-Sánchez, J., Van Der Does, H. C., Van Dam, P., García-Bastidas, F. A., Subandiyah, S., et al. (2018). A SIX1 homolog in Fusarium oxysporum f. sp. cubense tropical race 4 contributes to virulence towards Cavendish banana. PLoS One 13:e0205896. doi: 10.1371/journal.pone.0205896
Williams, A. H., Sharma, M., Thatcher, L. F., Azam, S., Hane, J. K., Sperschneider, J., et al. (2016). Comparative genomics and prediction of conditionally dispensable sequences in legume-infecting Fusarium oxysporum formae speciales facilitates identification of candidate effectors. BMC Genomics 17:191. doi: 10.1186/s12864-016-2486-8
Yadeta, K. A., and Thomma, B. P. J. (2013). The xylem as battleground for plant hosts and vascular wiltpathogens. Front. Plant Sci. 4:97. doi: 10.3389/fpls.2013.00097
Zhang, J. X., Mace, M. E., Stipanovic, R. D., and Bell, A. A. (1993). Production and fungitoxicity of the terpenoid phytoalexins in cotton inoculated with Fusarium oxysporum f. sp. vasinfectum. J. Phytopathol. 139, 247–252. doi: 10.1111/j.1439-0434.1993.tb01423.x
Zhou, E., Jia, Y., Singh, P., Correll, J. C., and Lee, F. N. (2007). Instability of the Magnaporthe oryzae avirulence gene AVR-Pita alters virulence. Fungal Genet. Biol. 44, 1024–1034. doi: 10.1016/j.fgb.2007.02.003
Zipfel, C., and Rathjen, J. P. (2008). Plant Immunity: AvrPto targets the frontline. Curr. Biol. 18, R218–R220. doi: 10.1016/j.cub.2008.01.016
Keywords: Fusarium oxysporum, Fusarium oxysporum species complex, Secreted in Xylem, effectors, pathogenicity, host specificity
Citation: Jangir P, Mehra N, Sharma K, Singh N, Rani M and Kapoor R (2021) Secreted in Xylem Genes: Drivers of Host Adaptation in Fusarium oxysporum. Front. Plant Sci. 12:628611. doi: 10.3389/fpls.2021.628611
Received: 12 November 2020; Accepted: 01 March 2021;
Published: 22 April 2021.
Edited by:
Paloma Melgarejo, Ministerio de Agricultura, Alimentación y Medio Ambiente (Spain), SpainReviewed by:
Antonio Di Pietro, University of Córdoba, SpainFrank L. W. Takken, University of Amsterdam, Netherlands
Copyright © 2021 Jangir, Mehra, Sharma, Singh, Rani and Kapoor. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Rupam Kapoor, kapoor_rupam@yahoo.com