
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Plant Sci. , 31 May 2019
Sec. Plant Pathogen Interactions
Volume 10 - 2019 | https://doi.org/10.3389/fpls.2019.00547
This article is part of the Research Topic Fusarium wilt of Banana, a Recurring Threat to Global Banana Production View all 13 articles
 Lilia C. Carvalhais1*
Lilia C. Carvalhais1* Juliane Henderson1
Juliane Henderson1 Vivian A. Rincon-Florez1
Vivian A. Rincon-Florez1 Cecilia O’Dwyer1
Cecilia O’Dwyer1 Elizabeth Czislowski2
Elizabeth Czislowski2 Elizabeth A. B. Aitken2
Elizabeth A. B. Aitken2 André Drenth1
André Drenth1Fusarium wilt is currently spreading in banana growing regions around the world leading to substantial losses. The disease is caused by the fungus Fusarium oxysporum f. sp. cubense (Foc), which is further classified into distinct races according to the banana varieties that they infect. Cavendish banana is resistant to Foc race 1, to which the popular Gros Michel subgroup succumbed last century. Cavendish effectively saved the banana industry, and became the most cultivated commercial subgroup worldwide. However, Foc tropical race 4 (TR4) subsequently emerged in Southeast Asia, causing significant yield losses due to its high level of aggressiveness to cultivars of Cavendish, and other commonly grown cultivars. Preventing further spread is crucially important in the absence of effective control methods or resistant market-acceptable banana cultivars. Implementation of quarantine and containment measures depends on early detection of the pathogen through reliable diagnostics. In this study, we tested the hypothesis that secreted in xylem (SIX) genes, which currently comprise the only known family of effectors in F. oxysporum, contain polymorphisms to allow the design of molecular diagnostic assays that distinguish races and relevant VCGs of Foc. We present specific and reproducible diagnostic assays based on conventional PCR targeting SIX genes, using as templates DNA extracted from pure Foc cultures. Sets of primers specifically amplify regions of: SIX6 in Foc race 1, SIX1 gene in TR4, SIX8 in subtropical race 4, SIX9/SIX10 in Foc VCG 0121, and SIX13 in Foc VCG 0122. These assays include simplex and duplex PCRs, with additional restriction digestion steps applied to amplification products of genes SIX1 and SIX13. Assay validations were conducted to a high international standard including the use of 250 Fusarium spp. isolates representing 16 distinct Fusarium species, 59 isolates of F. oxysporum, and 21 different vegetative compatibility groups (VCGs). Tested parameters included inter and intraspecific analytical specificity, sensitivity, robustness, repeatability, and reproducibility. The resulting suite of assays is able to reliably and accurately detect R1, STR4, and TR4 as well as two VCGs (0121 and 0122) causing Fusarium wilt in bananas.
Commercial banana production is under serious threat worldwide. A destructive disease caused by the fungus Fusarium oxysporum f. sp. cubense tropical race 4 (TR4) is spreading rapidly throughout the banana growing regions around the globe (Ordonez et al., 2015; Zheng et al., 2018). The soilborne nature of this pathogen is one of the main reasons why eradication and containment of TR4 are very challenging. In the absence of effective disease resistance in market-accepted banana varieties, one of the remaining measures to avoid further losses is to prevent spread through early detection with reliable diagnostics, and subsequent containment of new incursions.
Tropical race 4 was first identified in samples obtained from Sumatra, Indonesia, in 1992 (Ploetz, 1994; Ploetz, 2004). However, reports of infected Cavendish banana (Musa spp. AAA genome group) date from nearly 30 years before in Taiwan (Su et al., 1977; Ploetz, 1994). Since its emergence, TR4 has not only decimated the Cavendish banana industry in Taiwan, but has been making its way through Southeast Asia in China, Indonesia, Malaysia, Philippines, Laos, Myanmar, Vietnam, and Pakistan (Ordonez et al., 2015, 2016; Chittarath et al., 2018; Hung et al., 2018; Zheng et al., 2018). Intercontinental spread has also occurred with reports of the pathogen in Australia, Jordan, Lebanon, Oman, and Mozambique (Butler, 2013; García-Bastidas et al., 2014; Ploetz, 2015b; Dita et al., 2018).
The present situation of Fusarium wilt is familiar: another race of the causal agent (race 1 or R1) triggered a devastating epidemic in the banana growing regions of Central America in the first half of last century (Stover, 1990; Ploetz, 2005). It was “widely regarded as one of the most destructive plant diseases in recorded history” (Moore et al., 1995). A major commercial subgroup at the time, “Gros Michel” (Musa acuminata AAA genome group), was highly susceptible to R1 (Stover, 1962). The banana industry could only recover because cultivars belonging to the Cavendish subgroup were found to be resistant to R1 (Ploetz, 1994).
Resistance to Fusarium wilt R1, high productivity, the use of temperature-controlled transportation in boxes, and consumer acceptance turned Cavendish into the main subgroup used worldwide for export (Ploetz, 2005). Other varieties of banana are fundamental to the subsistence of households of many developing countries with low incomes, either as a staple or as a cash crop (FAOSTAT, 2018). With predictions of many of these countries being the main contributors to population growth (Gerland et al., 2014), the protection of banana production is essential to safeguard not only one of the most popularly consumed and exported fruits globally, but also livelihoods of millions of people (FAOSTAT, 2018).
To facilitate reference to groups that infect specific plant species, F. oxysporum has been sub-divided into formae speciales (Armstrong, 1981). However, given that not all varieties of the same plant species are necessarily susceptible to a particular forma specialis, these are further classified into races, e.g., F. oxysporum f. sp. lycopersici race 1 infects a specific group of tomato varieties (Alexander, 1945). The term forma specialis cubense is utilized to delineate those populations that can cause disease on banana. Nevertheless, the race classification can be inconsistent as surprisingly little work focused on identifying the host specificity of this pathogen (Ploetz, 2015a). In general, the cultivars within the subgroup Gros Michel (AAA), Pisang Awak (ABB) and others that belong to the AAB genomic group (e.g., Maqueno, Silk, and Pome) are susceptible to R1 (Stover and Buddenhagen, 1986; Stover and Simmonds, 1987; Stover, 1990). The susceptibility of banana varieties to races 2 and 4 is still unclear as insufficient variety trials have been systematically conducted for these two races. Race 4 was further divided into two groups, TR4 and subtropical race 4 (STR4). Lower temperatures are typically associated with STR4 infection and disease progression in cultivars of the Cavendish subgroup in subtropical regions, while TR4 is virulent to these varieties in all environments (Ploetz, 2005). Reclassification of TR4 into the new species Fusarium odoratissimum was proposed based on the genetic diversity of Foc isolates in the Indonesian centre of origin (Maryani et al., 2019).
An additional classification commonly used for the fungi causing Fusarium wilt is based on vegetative compatibility. Strains are classified into the same vegetative compatibility group (VCG) when they are able to anastomose and form a stable heterokaryon with each other (Leslie, 1993). This system reflects well the similarity of strains based on phenotypical traits (Caten and Jinks, 1966); however, genetic relatedness between different groupings cannot be inferred as mutations in the vic loci can lead to vegetative incompatibility even in closely related isolates (Bentley et al., 1998). Although different races can be associated with most VCGs, there are several limitations in the use of VCGs to support race classifications. Various VCGs can be associated with the same race, and evolutionary relationships cannot be taken into consideration (Ordonez et al., 2015). For example, VCG 0126 exhibits a closer phylogenetic affiliation to race 4 and, similarly to isolates classified into this race, have the ability to produce odorous aldehydes on media (Moore et al., 1991; Czislowski et al., 2017). Nonetheless, they are considered race 1 due to the host range on which it is able to cause disease (Pegg et al., 1994).
Breeding of new banana cultivars has been attempted, but until now has only had very limited success. Progress in banana breeding has been hampered by many factors, which include lack of germplasm evaluation, lack of fundamental genetic studies of diploid plants to identify characteristics, as well as lack of a long-term commitment to fund breeding. Besides, the occurrence of varying ploidy levels in cultivated varieties (which are polyploid) and the need for diploids for crosses, parthenocarpy, sterility, poor seed set, germination, and survival have hampered progress in banana breeding programs (Pillay and Tenkouano, 2011). Nonetheless, the growing impact of TR4 and other diseases such as black leaf streak (black Sigatoka, caused by the fungus Pseudocercospora fijiensis) and several improvements in breeding processes have yielded a renewed interest in this area (Pillay and Tenkouano, 2011; Li et al., 2015), while transgenic approaches to obtain TR4-resistant Cavendish are undergoing field-testing (Dale et al., 2017).
Once banana plants are infected, there is no effective treatment for Fusarium wilt. Fungicides or soil fumigation are ineffective to control or eradicate this disease (Ploetz, 2015b). Thus, in the absence of effective resistance or control methods, one of the remaining ways to manage this disease is preventing further spread and ensuring rapid containment of new infected plants. For this, early identification of infected plants combined with sensitive, accurate and robust diagnostic methods to detect incursions at an early stage are paramount. Diagnosing the specific VCGs and races is required as similar early symptoms caused by distinct strains are observed in various locally grown non-Cavendish and commercialized banana varieties (Karangwa et al., 2016). Diagnostics based solely on morphology is not reliable to differentiate distinct Foc and non-pathogenic strains of F. oxysporum (Pérez Vicente et al., 2014). Several molecular diagnostics methods have been developed, which mostly rely on core genomic regions and hence are unlikely to be closely linked or associated with pathogenic characteristics of different races. A molecular diagnostic method based on the intergenic spacer region (IGS) of the nuclear ribosomal gene cluster of Foc is widely used and is efficient for detecting TR4 (Dita et al., 2010). However, this assay was not designed to detect other strains that are closely related to TR4 and are also able to infect Cavendish, such as those affiliated to VCG 0121 and VCG 0122 (Ordonez et al., 2015; Czislowski et al., 2017; Mostert et al., 2017). Another study conducted by Lin et al. (2009) reported primers that are able to detect all race 4 strains, and hence can detect strains assigned to VCG 0121 and VCG 0122. However, these primers are unable to distinguish TR4 and STR4. Despite being closely related to TR4 VCGs 01213/16 and being able to attack Cavendish varieties in the tropics, the classification of R4 VCGs 0121 and 0122 as TR4 or STR4 is still a matter of debate (Moore et al., 1993; Bentley et al., 1998; Buddenhagen, 2009). Some of the isolates belonging to VCGs 0121 and 0122 have been reported to infect Cavendish causing symptoms that were less severe than those caused by TR4 isolates (Buddenhagen, 2009), which suggests that predisposing conditions may need to be in place for symptoms to be expressed. Therefore, we opted for classifying here VCGs 0121 and 0122 as R4, without further sub-classifications as tropical or subtropical. Foc strains affiliated to these VCGs can cause disease in Cavendish (Mostert et al., 2017), thus having molecular tools available to detect VCG 0121 and VCG 0122 is especially beneficial for countries reliant on banana cultivation under subtropical conditions.
A common target for diagnostics is genes that encode proteins that are strongly correlated with virulence (de Sain and Rep, 2015). In F. oxysporum-infected tomato, some of these proteins have been detected in the xylem sap and named Secreted-in-Xylem (SIX) (Rep et al., 2004; Houterman et al., 2007; Gawehns et al., 2014). When these proteins or other molecules (e.g., secondary metabolites and small RNAs) are associated with processes in the host in favor of colonization by the pathogen and disease progression, they are known as effectors (Weiberg et al., 2013; de Sain and Rep, 2015). Homologs of the SIX genes are found across different formae speciales and their profile can be used to distinguish distinct formae speciales, races and isolates (Lievens et al., 2009; Chakrabarti et al., 2011; van Dam et al., 2016; Czislowski et al., 2017). SIX gene profiling has been successfully used to distinguish the three races in F. oxysporum f. sp. lycopersici (Lievens et al., 2009). The mode of action of these proteins in the pathogenicity process remains to be elucidated, but some studies have suggested an effect on plant defense signaling (Thatcher et al., 2012; Kazan and Lyons, 2014).
Diagnostic tools to identify races associated with banana Fusarium wilt are needed to allow early detection of new incursions and distinguish isolates infecting varieties that are generally more susceptible to a wider range of Fusarium (e.g., Lady Finger). However, poor assay validation often leads to a huge gap between development and implementation of diagnostic methods (Chilvers, 2012). Evidence for the specificity and reliability of diagnostic assays is required and therefore validation needs to be conducted according to rigorous standards.
The gene SIX8 has been previously shown to contain enough variation to differentiate Foc race 4 from the races 1 and 2 as well as STR4 from TR4 through a conventional PCR (Fraser-Smith et al., 2014). However, only three non-pathogenic isolates of F. oxysporum were included in the screening, and the detection of TR4 was based on the absence of an amplification product. Diagnosing a disease based on the absence of an amplification product is unreliable because the absence of a pathogen would provide the same outcome. In addition, basic validation parameters such as sensitivity and robustness cannot be assessed if there is no amplicon for the assessment of positive diagnostics. Therefore, the overall objective of our study was to assess whether sequence variation in SIX genes of Fusarium races could be used to develop a series of assays that can detect relevant VCGs and races of Fusarium wilt of banana. We specifically sought to address whether: (1) SIX genes exhibit specific sequences that are conserved within each race; (2) there are sufficient single nucleotide polymorphisms (SNPs) across SIX gene homologs to enable the design of primers which target unique sequences within the above races; (3) the primers targeting specific SIX genes homologs can be used in PCR assays to reach a level of specificity, sensitivity, robustness and repeatability that meets international standards for diagnostic assays; and (4) our proposed assays can detect all races and VCGs mentioned above and outperform popularly used molecular diagnostic tests previously designed to detect R4 (STR4 and TR4).
Reliable detection of different races and VCGs of Fusarium will enable decision-making by the banana industry stakeholders.
Regions in the SIX genes conserved within races of Foc were identified by analyzing sequences generated by Czislowski et al. (2017), which are available in the National Center for Biotechnology Information database1 (NCBI Resource Coordinators, 2017), under accessions KX434886-KX435052. These sequences had been generated by whole-genome sequencing (WGS) analysis followed by Foc-SIX specific PCR to characterize the diversity and evolution of the SIX genes in a collection of 89 isolates representing 23 genetic lineages of Foc (Czislowski et al., 2017). Based on the data of Czislowski et al. (2017), it was possible to identify in this current study an exclusive SIX gene homolog for each race of Foc. Each of these exclusive homologs was present in a different SIX gene. Consensus sequences of these SIX gene homologs were aligned for the identification of conserved regions within races of Foc. Alignments were performed either in Geneious (v10.0.9) (Kearse et al., 2012) or Clustal Omega2 (Sievers and Higgins, 2014). SNP-rich regions within the distinct SIX genes were then targeted for race-specific primer design, as described in the following sections.
Sets of primers were designed either to anneal to SNP-rich regions within the targeted SIX gene or to anneal to regions flanking a specific SNP that contained a sequence which included a restriction enzyme site. Details of races, VCGs, target SIX gene homologs, primer sequences, and annealing temperatures are listed in Table 1. The strategy for identifying TR4 isolates was based on the exclusive presence of the homolog “a” of SIX1 in TR4 (Czislowski et al., 2017). For the design of specific primers to detect TR4 isolates, full-length sequences from all SIX1 gene homologs (“a” to “i”) were aligned using the software Clustal Omega2. The primer set SIX1_266 (Table 1) was designed to anneal to regions which were rich in SNPs that were exclusive to the SIX1 gene homologs “a,” “b,” or “c.” These gene homologs are unique to TR4, R4 VCG 0121 and R4 VCG 0122, respectively (Czislowski et al., 2017). The 266 bp-product amplified by these primers contains a unique recognition site for the restriction enzyme HpyAV (New England BioLabs), which is present in SIX1 homolog “a” and absent in homologs “b” and “c.” After restriction digestion, two DNA fragments are predicted to be generated, one with 124 bp and the other with 142 bp.
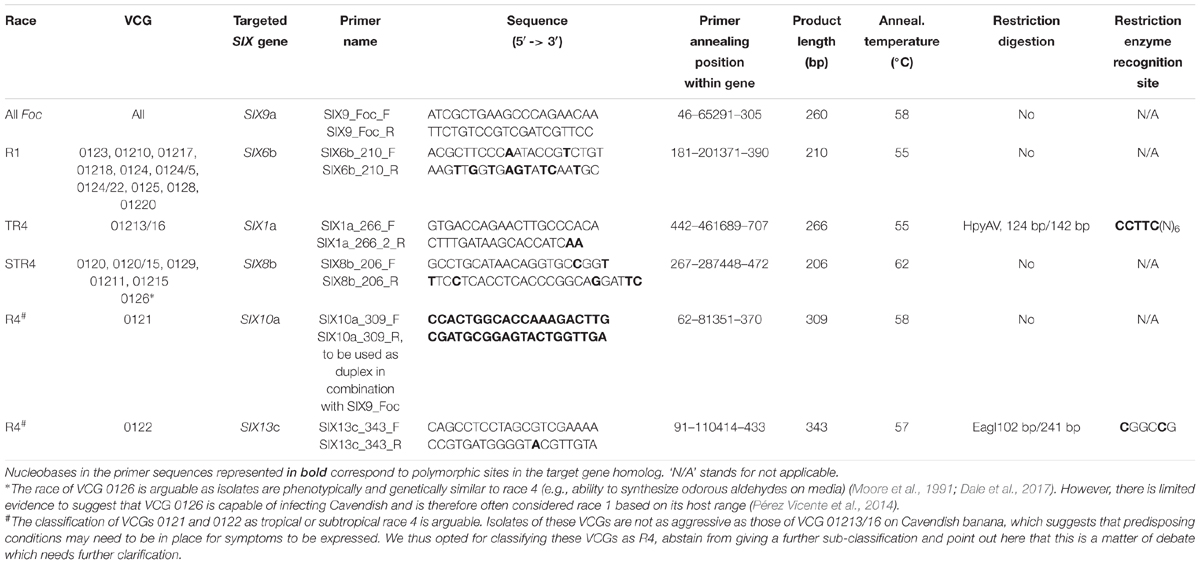
Table 1. Primer sequences designed in this study based on SIX gene sequences generated in Czislowski et al. (2017).
A different approach was adopted to detect R4 VCG 0121 and R4 VCG 0122. To specifically detect R4 VCG 0121, primers were designed to target the SIX10 gene which is present exclusively in this VCG (Czislowski et al., 2017) using Primer-BLAST3. A 309 bp-product is predicted to be amplified with this primer set. However, we obtained the same amplicon for several F. oxysporum strains isolated from crops other than banana (data not shown). We thus opted for the use of a duplex PCR combining the primer set that targets SIX10 in combination with the SIX9 gene that is conserved in all isolates of Foc (Czislowski et al., 2017). The primer set SIX9_Foc was designed to amplify a 260-bp product targeting the homolog “a” of SIX9 using the software Primer3 (Untergasser et al., 2012).
To detect VCG 0122, primers were designed to amplify a 343-bp region of the gene SIX13 that contains a restriction site exclusive to this VCG recognized by the restriction enzyme EagI (New England Biolabs). A restriction digestion step after amplification was necessary as there was insufficient sequence variation to design primers that would amplify only the homolog “c” of SIX13 (Czislowski et al., 2017). Two fragments of this PCR product were predicted to be generated after the restriction digestion, one with 102 bp and the other with 241.
To identify STR4 isolates, primers were designed to anneal to SNP-rich regions of the SIX8 gene which are unique to the homolog “b,” only present in STR4 (Czislowski et al., 2017). Likewise, to detect R1 isolates, primers were designed to anneal to the polymorphic regions of the homolog “b” of the SIX6 gene, found to be present only in R1 by Czislowski et al. (2017).
DNA material extracted from isolates of Fusarium was used as templates for the amplification tests in this study. DNA was extracted from 5 to 7-day-old monoconidial cultures, grown on half-strength potato dextrose agar (PDA, Difco Laboratories) at 25°C, using the DNeasy Plant Mini Kt (Qiagen) on the QIACube (Qiagen) following the manufacturer’s recommendations. Approximately 50–100 mg of mycelium were used as initial material in the extractions (amount recovered by scraping the medium surface of a 100-mm diameter standard Petri dish fully covered with mycelium).
For all amplifications, MyTaqTM Red DNA Polymerase (Bioline) was used in 20 uL reactions according to the manufacturer’s recommendations. The primer and DNA template concentration in the final PCR reaction were 0.4 μM and 5 pg. uL-1, respectively. The following cycling conditions were used for all PCR: 95°C for 3 min, 30 cycles of 95°C for 15 s, and annealing temperature were primer-set dependant (see Table 1) for 15 s and 72°C for 10 s. In the cases of follow-up restriction digestions (for SIX1 and SIX13 products), no purification step of the PCR products is needed before enzymatic digestions. Restriction digestions for the enzymes HpyAV and EagI (New England Biolabs) were performed using 4 μL of the amplification product and 0.4 units (U) of enzyme in a total volume of 10 μL. The enzyme EagI is also known as Eco521. Star activity is a property exhibited by some restriction enzymes in which they show relaxed or inaccurate sequence recognition (Wei et al., 2008). HpyAV and EagI-HF have not been predicted to have star activity in the buffers recommended by the manufacturer4. If the recommended conditions are used as reported here, no restriction enzyme star activity is expected to occur. The restriction enzymes digestions were conducted at 37°C for 1 h and subsequently inactivated for 20 min at 65°C. A volume of 5 μL of amplification products and 10 μL restriction digestions were run on a 1.5% agarose gel and post-stained with ethidium bromide (1 μg mL-1).
The primer set that partially amplifies the translation elongation factor 1α gene (TEF-1α) was used in an additional PCR for all samples to confirm that each fungal DNA sample was amenable to amplification and control for false negatives (O’Donnell et al., 1998).
To validate the diagnostic assays, the international standards as proposed in the “guidelines for the validation and verification of quantitative and qualitative test methods” were followed (National Association of Testing Authorities, 2018). These guidelines delineate particular requirements that a proposed method should meet to fulfill the purpose for its intended use (i.e., “fit-for-purpose”). As validation parameters, we included inter and intraspecific analytical specificity, sensitivity, robustness, repeatability, and reproducibility.
A total of 250 isolates of Fusarium spp. were screened in the validation tests. These included 16 different plant-associated Fusarium species in addition to 21 VCGs commonly associated with Foc (VCGs 120 to 01223).
For validation of assay specificity, we tested each primer set with 16 Fusarium species (Table 2). These Fusarium isolates were obtained from infected plant material in Australia. We also included in the validations nine different formae speciales of F. oxysporum and 21 different VCGs of Foc. Sensitivity was tested in a 10-fold serially diluted positive control within the range of 10 ng to 0.1 fg.μL-1 (Supplementary Table S1). Before the preparation of dilutions, DNA concentrations were measured in a Qubit fluorometer with the Quant-iT High-Sensitivity dsDNA Assay Kit (Thermo Fisher Scientific).

Table 2. Strains used for inter-specific validation of the diagnostic assays using the primer sets SIX1a_266 (TR4), SIX9_Foc/SIX10a_309 duplex (R4 VCG 0121), SIX13c_343 (R4 VCG 0121), SIX8b_206 (STR4) and SIX6b_210 (R1) and DNA extracted from Fusarium isolates obtained from infected plant material in Australia.
To test for assay robustness, two different Taq polymerases were used for the amplification of eight positive controls, in duplicate, in two different thermal cyclers. The DNA polymerases used were MyTaqTM HS DNA polymerase (Bioline, MyTaq reaction buffer with 1 mM dNTPs, 3 mM MgCl2) and ThermoFisher Scientific Taq Polymerase [Taq Buffer with (NH4)2SO4], using reagent concentrations and cycling conditions as previously described. The robustness of the assays was tested by changing the manufacturer of the DNA polymerases. Different manufacturers of restriction enzymes were not used for the assays that included an additional restriction digestion step (SIX1 and SIX13). The reason for that is based on reportedly different outcomes observed according to the type of polymerase (Schierwater and Ender, 1993), while the results obtained with restriction enzymes from different manufacturers are more consistent due to the intrinsic specificity of these enzymes.
For repeatability, six to eight positive controls from a variety of VCGs in six separate occasions were tested by the same operator for each primer set. For all assays, two negative controls were also included: one that contained a DNA sample that was not targeted by the primers and another that lacked DNA template. Reproducibility was tested by conducting separate assays with two different operators, on three different occasions, with the same set of samples. All tests included two technical replicates. Two previously reported conventional PCR methods were also compared with our assays: one utilized primers which amplify a region of the intergenic spacer (IGS) region of the nuclear ribosomal operon to detect TR4 (Dita et al., 2010); the other utilized primers developed from a Random Amplified Polymorphic DNA marker specific to Foc Race 4 (Lin et al., 2009).
Sequences from previously reported homologs of each SIX gene (Czislowski et al., 2017) were aligned and conserved regions within each race were identified as potential primer annealing sites. The alignment of different homologs of the genes SIX1, SIX13, SIX8, SIX6 allowed the identification of exclusive sequences for TR4, R4 (VCG 0122), STR4 and R1, respectively. As SIX10 was reported to be only present in R4 VCG 0121, and only one homolog has been reported (Czislowski et al., 2017), no alignment was needed prior to primer design targeting this gene.
Details on the presence of sufficient sequence polymorphisms that enabled race-specific primer design are described in the subsections dedicated to each Foc race below. All diagnostic assays were validated using DNA extracted from 38 isolates of Fusarium affiliated to 16 different species and 59 isolates of F. oxysporum obtained from healthy plant tissues of a range of plant species including, amongst others, Asparagus officinalis, Citrullus lanatus, Cucumis melo, Euphorbia dallachyna, Musa spp., Phoenix sp., Solanum lycopersicum, Solanum tuberosum, Trianthema portulacastrum, and Zingiber officinale. This included endophytes that were asymptomatic on their hosts and F. oxysporum classified into different formae speciales. A total of 150 Foc isolates affiliated to different races were used in our tests, including 32 TR4 isolates obtained in various countries (Tables 2–4).

Table 3. Intra-specific validation of the diagnostic assays using the primer sets SIX1a_266 (TR4), SIX9_Foc/SIX10a_309 duplex (R4 VCG 0121), SIX13c_343 (R4 VCG 0121), SIX8b_206 (STR4) and SIX6b_210 (R1) and DNA extracted from Fusarium isolates obtained from infected plant material in Australia.
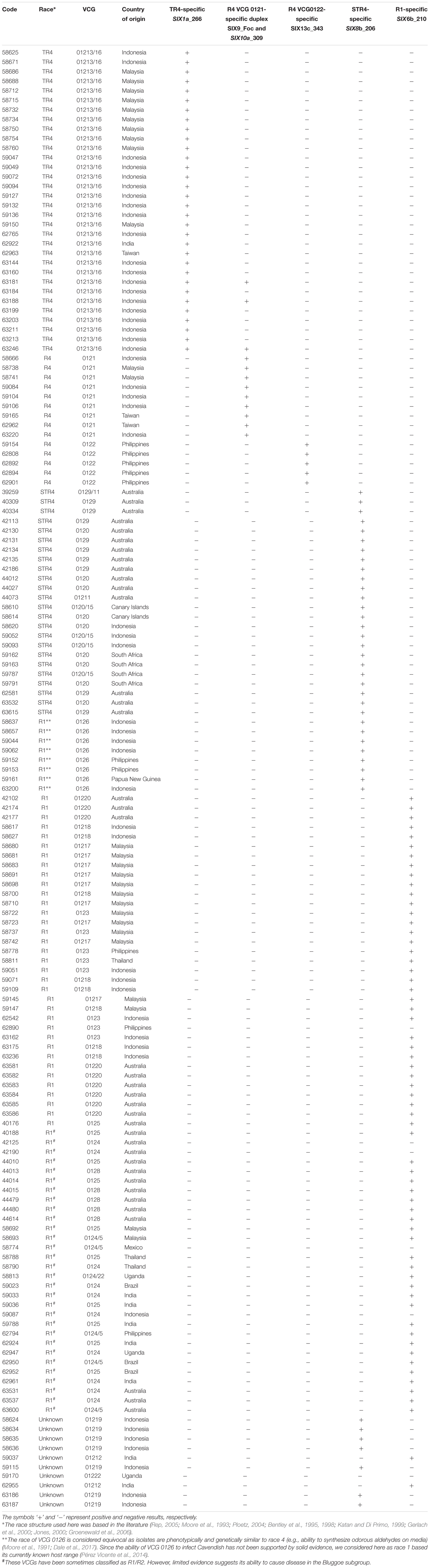
Table 4. Validation of the diagnostic assays using the primer sets SIX1a_266 (TR4), SIX9_Foc/SIX10a_309 duplex (R4 VCG 0121), SIX13c_343 (R4 VCG 0121), SIX8b_206 (STR4) and SIX6b_210 (R1) and DNA extracted from isolates from the Fusarium oxysporum f. sp. cubense (Foc) species complex infecting Musa spp.
Sequences of the SIX1 homologs “a” to “i” were aligned and the primer set SIX1a_266 was designed to anneal to regions that were conserved within homologs “a,” “b,” and “c,” and were exclusive to TR4, R4-VCG 0121, and R4-VCG 0122, respectively (Table 1). These primers flanked a recognition site of the enzyme HpyAV, which is only present in the SIX1 gene homolog “a,” which is unique to TR4 (Czislowski et al., 2017). Figure 1 shows amplification products for TR4 isolates and HpyAV-digested fragments. The amplified product of 266 bp was digested into two fragments with the enzyme HpyAV, one with 124 bp and the other with 142 bp (Figure 1). The 266-bp amplification product is also obtained for R4-VCG 0121 and R4-VCG 0122; however, this product is not digested by HpyAV in strains associated with these VCGs (Figure 1). No false positives or false negatives were obtained for other Fusarium species, F. oxysporum strains, formae speciales of other F. oxysporum, or races tested (Tables 2–4). This assay was validated and proven to be repeatable, robust, and specific. The limit of detection is 1 pg.μL-1 (Supplementary Table S1).
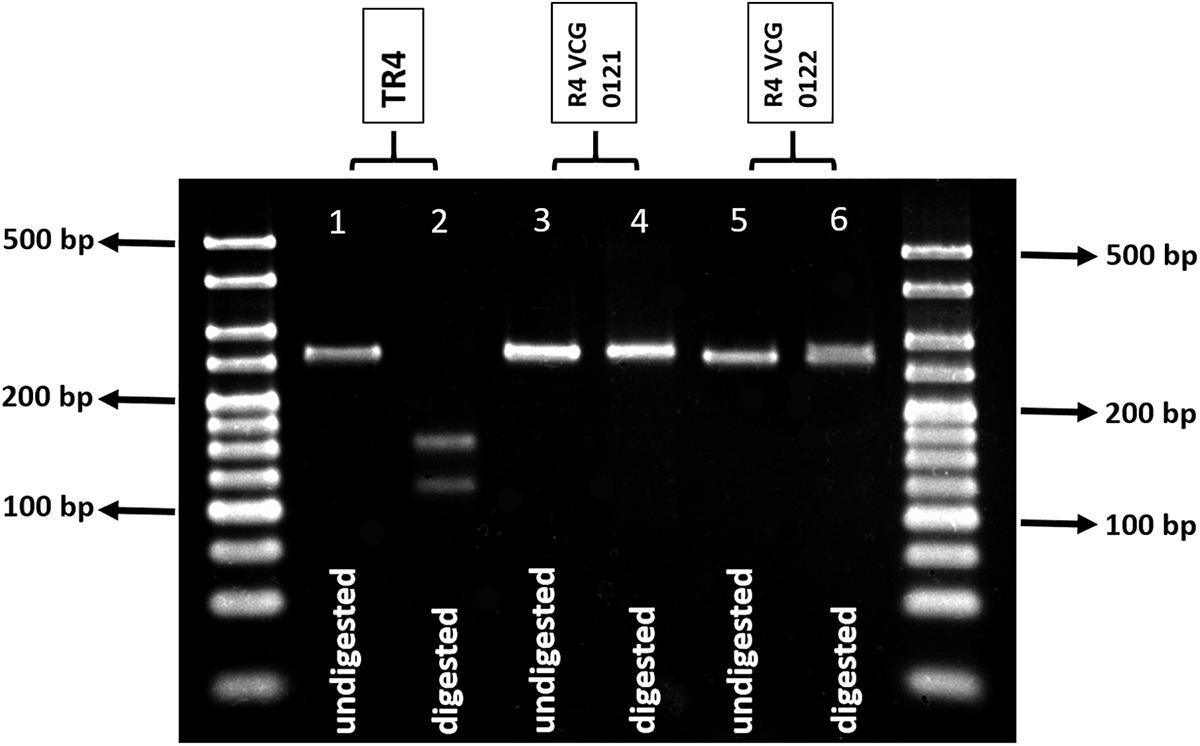
Figure 1. Agarose gel (3%) showing undigested and digested 266-bp amplification products obtained with the primer set (SIX1a_266) designed to target the SIX gene homolog present exclusively in TR4 (SIX1a). Ladder used: 25 bp Bioline Hyperladder. Lane 1, undigested amplicon from TR4 isolate (5 μL); lane 2, HpyAV-treated amplicon from TR4 isolate (10 μL); lane 3, undigested amplicon from R4 VCG 0121 isolate (5 μL); lane 4, HpyAV-treated amplicon from R4 VCG 0121 isolate (10 μL); lane 5, undigested amplicon from R4 VCG 0122 isolate (5 μL); and lane 6, HpyAV-treated amplicon from R4 VCG 0122 isolate (10 μL).
The primer set which targets the gene SIX10 in VCG 0121 (SIX10a_309) generated the expected amplification product in nine isolates of F. oxysporum that were asymptomatic endophytes in plant genera distinct to Musa (data not shown). For this reason, we developed a duplex PCR as the diagnostic assay for the detection of VCG 0121, which then included primers that target the homolog “a” of the gene SIX9. According to currently available data, this gene homolog seems to be present in Foc but absent in other formae speciales of F. oxysporum (Czislowski et al., 2017; Tables 3, 4). Two amplicons with the expected lengths were obtained using the primer sets SIX9_Foc and SIX10a_309, with 260 and 309 bp, respectively (Figure 2). The resultant duplex PCR has a limit of detection of 0.1 ng.μL-1 and met most validation criteria (Tables 2–4 and Supplementary Table S1). Four of 32 TR4 isolates from Indonesia tested positive for this duplex-PCR assay (Table 4), inferring the presence of SIX10 in these isolates. The limit of detection was 0.01 ng.μL-1 (Supplementary Table S1).
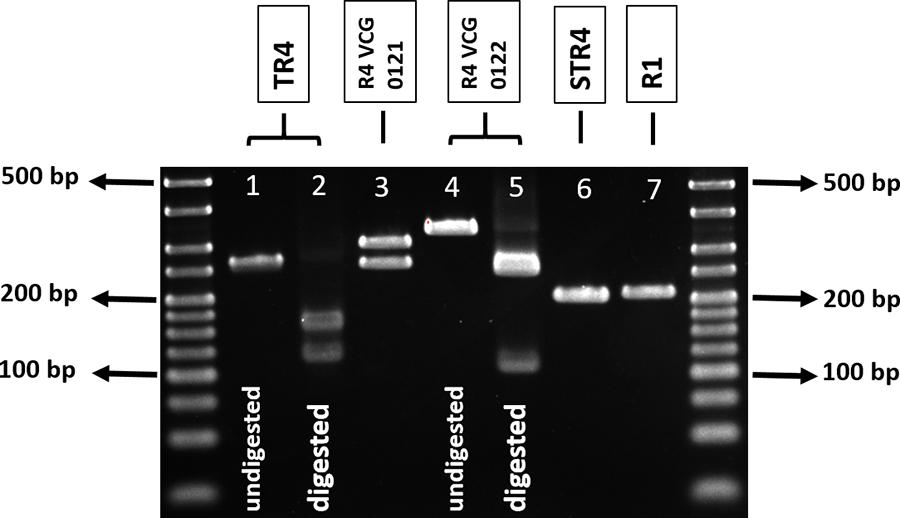
Figure 2. Agarose gel (3%) showing amplification products obtained with primers designed to target different races and VCGs of Fusarium oxysporum f. sp. cubense. Ladder used: 25 bp Bioline Hyperladder. Lane 1, undigested 266 bp SIX1 gene amplicon (5 μL); lane 2, TR4-specific HpyAV-treated 266 bp SIX1 gene amplicon (10 μL), showing 124 and 142 bp digests; lane 3, R4 VCG 0121-specific 309 and 260 bp amplicons from duplex PCR of the genes SIX10 and SIX9 (5 μL), respectively; lane 4, undigested 343 bp SIX13 gene amplicon (5 μL); lane 5, R4 VCG 0122-specific EagI-treated 343 bp SIX13 gene amplicon, showing 102 and 241 digests (10 μL); lane 6, STR4-specific 206 bp SIX8 gene amplicon (5 μL); and lane 7, R1-specific SIX6 210 bp gene amplicon (5 μL).
This assay was designed to target the gene SIX13 and flanked a R4-VCG 0122-specific recognition site for the restriction enzyme Eagl (Table 1). The expected 343 bp product was amplified and, after digestion with the enzyme EagI, expected fragments of 102 and 241 bp were obtained (Figure 2). The assay using the primer set SIX13c_343 followed by an EagI-restriction digestion was specific, with no false negatives or false positives detected (Tables 3–5). The limit of detection of this assay was 0.01 g.μL-1 (Supplementary Table S1). An example of a gel showing amplification products of SIX13 obtained with a range of DNA template concentrations is shown is Supplementary Figure S1. All validation criteria of repeatability and robustness were met.
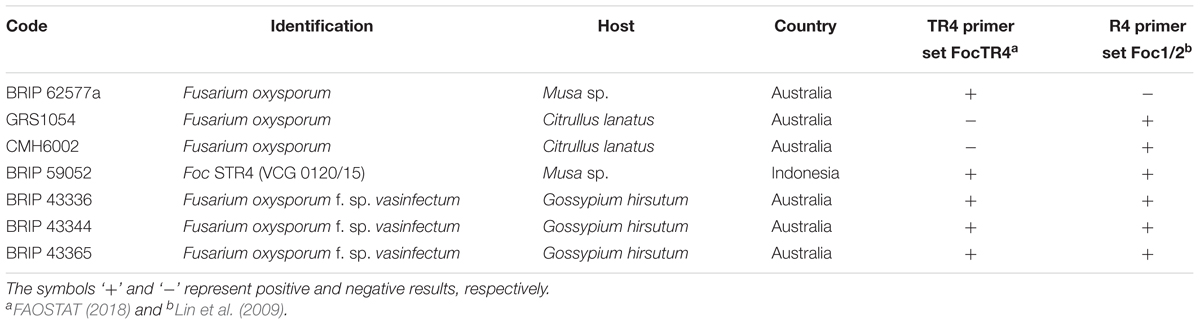
Table 5. List of strains tested with primer sets reported in previous studies, which target TR4 and/or R4 strains and produced false positive results.
The primer set SIX8b_206 was designed to amplify exclusively STR4 isolates (Table 1). A 206 bp product was amplified for all tested STR4 isolates (Figure 2) and no false positives were obtained (Tables 2–4). Amplification products with the expected sizes were also obtained for isolates belonging to the VCGs 01219 (whose race is still undetermined) and VCG 0126 (classified as R1) (Table 4). This assay was proven to be repeatable, reproducible, robust, sensitive and specific. The limit of detection was 0.01 ng.μL-1 (Supplementary Table S1).
The primer set SIX6b_210 was designed to detect exclusively R1 isolates (Table 1). The primers generated the expected 210 bp amplicon for most R1-associated VCGs (60 isolates out of 65, Figure 1 and Table 4). Three strains classified as R1 did not generate the expected PCR product, with one of them belonging to the VCG 0123 (BRIP 62890), three to the VCG 0124 (BRIPs 42125, 42190, and 59087), and the other to the VCG 0124/5 (BRIP 58774) (Table 4). The test was shown to be repeatable, robust, reproducible, and the limit of detection was 0.1 ng. uL-1 (Supplementary Table S1).
Although all primer sets generated amplicons using DNA templates at a concentration of 0.5 ng. μL-1 in the PCR reaction for most isolates, some false negatives were observed at this particular concentration. Thus, we recommend using DNA concentrations between 50 pg. μL-1 and 5 pg. μL-1 in the PCR reaction as these concentrations worked consistently well in the assays conducted to determine the limit of detection (Supplementary Table S1).
We confirmed unpublished results obtained by the Department of Agriculture and Fisheries (Henderson J., unpublished), which revealed that the primers FocTR4, previously reported to detect TR4 (Dita et al., 2010), tested positive for one endophytic strain isolated from an asymptomatic banana plant grown in Northern Queensland, Australia (BRIP62577, Table 5). Furthermore, the same primer set and the Foc1/2 primer set reported by Lin et al., 2009 tested positive for one STR4 and three F. oxysporum f. sp. vasinfectum isolates (BRIP 59052, 43336, 43344, and 43365, Table 5). The Foc1/2 primer set also produced amplification products for two endophytic strains of F. oxysporum colonizing watermelon plants (Citrullus lanatus) (GRS1054 and CMH6002, Table 5). As the Foc1/2 primer set cannot distinguish between TR4 and STR4, it is expected that amplicons for STR4 strains are produced, such as for BRIP 59052 (Table 5).
This study supports the utilization of SIX genes as targets for molecular diagnostics for races of Foc. The results demonstrate that for each race, specific conserved SIX gene sequences can be identified especially for TR4, R4-VCG 0122, and STR4. Sufficient SNPs across SIX gene homologs enabled the differentiation of TR4, R4-VCG 0121, R4-VCG 0122, STR4, and R1 strains through conventional PCR using primers targeting polymorphic regions of the genes SIX1, SIX9/SIX10, SIX13, SIX8, and SIX6, respectively. However, only a small number of SNPs was present to distinguish TR4 and R4-VCG 0122 from other races and VCGs. Therefore, following the PCRs targeting SIX1 and SIX13, a restriction digestion step with the enzymes HpyAV and EagI was required to detect the specific homologs present in TR4 and R4 VCG 0122, respectively. Overall, our validation experiments revealed that our assays are specific, sensitive, robust and repeatable.
Strains with VCG 0122 have been recently reported to infect the Cavendish cultivar Grande Naine (AAA) in the Davao del Norte area in the Philippines between 2006 and 2007 (Mostert et al., 2017), but Maryani et al. (2019) reported that these strains were not able to infect Cavendish in glasshouse trials. These contradictory results may stem from the low aggressiveness of these strains, which may not infect plants under artificial conditions. Some factors influencing the success of the infections in the field may not have been known and taken into account. The primer set SIX1a_266 generates amplicons for not only TR4, but also the R4 VCGs 0121 and 0122. Therefore, the primer sets SIX10a_309 and SIX13c_343 were additionally designed to detect exclusively R4 VCGs 0121 and 0122, respectively. Other PCR primers that can detect VCG 0121 and TR4 have been reported by Lin et al. (2009), but they cannot discriminate between TR4 and STR4 isolates. In the study by Czislowski et al. (2017) within Foc, only one homolog of the gene SIX10 was identified and it was present exclusively in VCG 0121; consequently SIX10 appeared to be a suitable specific target for VCG 0121. The SIX10 gene is however, present in other F. oxysporum strains, as we observed from amplification products using isolates other than Foc. This led to the use of the additional primer set SIX9_Foc, which targets a gene homolog of SIX9 previously shown to be present in all Foc (Czislowski et al., 2017). Despite the reported monophyletic nature of TR4 isolates (Ordonez et al., 2015; Maryani et al., 2019), our results indicate that there is some level of variability associated with the SIX gene profiles of these isolates given that amplicons of SIX10 in the duplex PCR were observed in 12.5% of the TR4 isolates tested (4 out of 32). However, this finding does not affect the use of our diagnostic toolkit. For example, if Fusarium wilt symptoms are reported in a Cavendish plantation in a region where there is no record of TR4, the first molecular diagnostic method that should be conducted is a PCR using the SIX1a_266 primer set, followed by restriction digestion with the HpyAV enzyme. This is important given the increased risks associated with a TR4 incursion compared to the other races. If the expected 124 and 142 bp digestion products are obtained after the treatment with HpyAV, there is strong evidence to suggest that the isolate is TR4. An amplification reaction with primers targeting the gene SIX9 should be run simultaneously with the primer set SIX1a_266 to verify whether the isolate is Foc. If this is confirmed, the isolate may be considered TR4. DNA from isolates with VCGs 0121 and 0122 will also generate an amplification product with this primer set but, because they lack the HpyAV restriction site, the 266-bp product will not be digested into two fragments. If this is the case, then the next step would be either to use the duplex PCR to verify whether it detects R4 VCG 0121, or the SIX13c_343 primer set to identify R4 VCG 0122. We advise that a PCR targeting the SIX9 gene should be conducted in parallel with the race-specific PCR assay to confirm that the tested isolate is a Foc and not an endophytic Fusarium strain that shares the same SIX gene homolog with a pathogenic strain. In Figure 3 we suggest a workflow for the use of the different primer sets designed in this study.
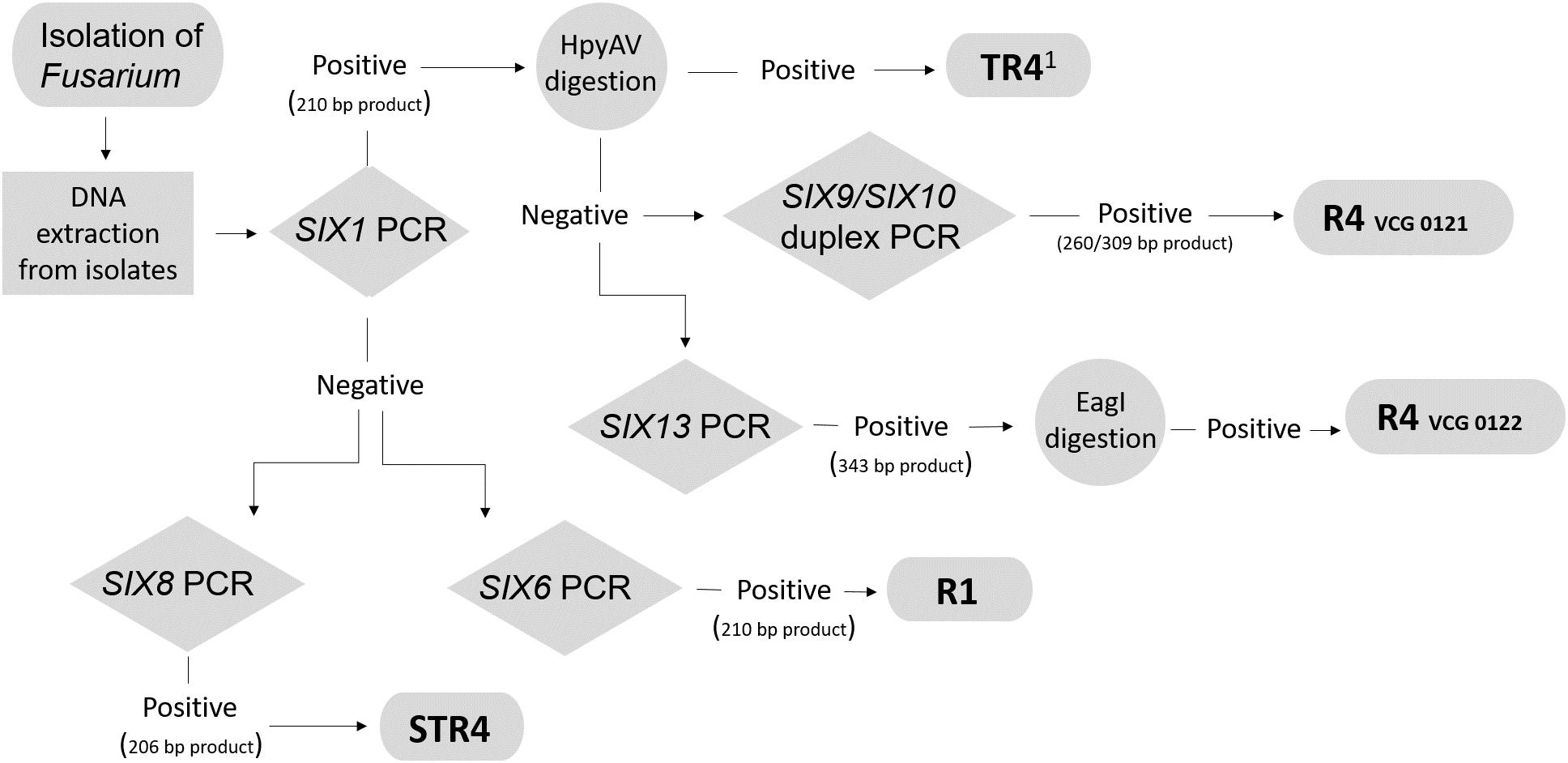
Figure 3. Proposed molecular diagnostic workflow to distinguish races and vegetative compatibility groups (VCGs) of F. oxysporum f. sp. cubense. based on a conventional PCR targeting the SIX genes. TR41 has been proposed to be reclassified as Fusarium odoratissimum (Maryani et al., 2019).
The primer set SIX8b_206 was designed to amplify exclusively STR4 isolates. Amongst a wide range of strains tested, 24 STR4 isolates from the eight different VCGs considered to be STR4 were included in the assays (Table 4). A 206 bp product was amplified for all tested STR4 isolates and no false positives were obtained (Table 2). Amplification of products with the expected size were also obtained for isolates belonging to the VCGs 01219 and 0126, whose races are either not determined or still controversial. The race associated with the VCG 01219 has not been determined yet. However, our findings corroborate those of other studies that documented strains which belong to the VCG 01219 have the same SIX gene profile as all the other VCGs commonly associated with STR4 (Czislowski et al., 2017). In addition, despite VCG 0126 having been associated with R1 given its host range, our results are in agreement with reports suggesting closer genetic relatedness of VGC 0126 to isolates belonging to VCGs associated with STR4 than those associated with R1 (Bentley et al., 1998; Groenewald et al., 2006; Czislowski et al., 2017).
Although the expected results were obtained for most strains tested with the SIX6 primers designed to amplify R1, five R1 isolates did not show the expected amplification product. Three of them belong to VCG 0124 and one to VCG 0123. A possible reason is the polyphyletic nature of R1 strains (Ordonez et al., 2016), which complicates the design of specific primers for this race. A recent phylogenetic analysis of Foc using strains isolated in the Indonesian center of origin revealed that diversity among genetic lineages of Foc is higher than previously anticipated (Maryani et al., 2019). Identification and phylogeny of these isolates were based on the genes encoding the translation elongation factor-1alpha (tef1), the RNA polymerase II largest submunit (rpb1), and the RNA polymerase II second largest subunit (rpb2), leading to the formal description of nine independent lineages (Maryani et al., 2019). TR4 isolates belong to the same lineage and were proposed to be reclassified as F. odoratissimum (Maryani et al., 2019). A previous attempt to design a real-time PCR specific for Foc R1 isolates has been reported but only four isolates belonging to this race were included and no VCG information was given (Yang et al., 2015). The distribution of putative effectors in natural and agro-ecosystems suggests that SIX6, which was chosen in this study to detect R1, may have been horizontally transferred across F. oxysporum strains (Rocha et al., 2016).
Pathogenicity genes have been common targets for molecular diagnostic assays of plant diseases (Loreti and Gallelli, 2002; Zaccardelli et al., 2007; Serdani et al., 2013). In fungi, the most typical avirulence factors are secreted small proteins (less than 200 amino acids) (Rep, 2005). Disease resistance is often triggered when the host’s innate immune system identifies these proteins (Kim et al., 2016). However, it is not unusual that fungi adopting endophytic lifestyles also harbor genes that encode these proteins (Kim et al., 2016). The occurrence of putative effector genes has been investigated in strains of the F. oxysporum species complex isolated from asymptomatic plants occurring in natural environments (Rocha et al., 2016; Jelinski et al., 2017). The genes PDA1, PELD, SGE1, and SIX were evaluated because of their involvement in pathogenicity and functional diversity (Rocha et al., 2016). From these putative effector genes, SIX genes were reported to be generally less prevalent, which suggests that they may not have a crucial function in natural populations of F. oxysporum (Rocha et al., 2016). SIX genes have already been used as targets for molecular diagnostics, as in the case of a loop-mediated isothermal amplification assay targeting the SIX3 gene of F. oxysporum f. sp. lycopersici (Ayukawa et al., 2017). The proposed method claims to detect point mutations and distinguish race 3 strains from other races of F. oxysporum f. sp. lycopersici (Ayukawa et al., 2017). Furthermore, a previous study detected enough sequence variation in the SIX8 gene to design a primer set which distinguished R4 strains from races 1 and 2 and another set which differentiates STR4 from TR4 by the presence of an amplification product only in STR4 (Fraser-Smith et al., 2014). The main issue with this approach is its reliance on the absence of a band to test positive for TR4, which is not acceptable as a robust diagnostic test.
A range of molecular diagnostics methods has been previously proposed to distinguish R4 from other races of Foc. Lin et al. (2009) proposed primers to detect R4 isolates based on a random amplification of polymorphic DNA (RAPD) product that was unique to R4 strains. However, their validation tests included only seven reference Foc isolates from other races (two R1 isolates, two R2, and three STR4 isolates). This primer set was further tested in at least two other studies. Dita et al., 2010 found that these primers amplified nine different Foc VCGs, including several that included strains which are not classified as R4, such as the ones belonging to the VCG 01210 that typically includes R1 isolates (Dita et al., 2010). Another study reported that the primers developed by Lin et al. (2009) tested positive for endophytic strains isolated from healthy banana plants and R1 isolates (Magdama, 2017). This primer set is also unable to distinguish STR4 from TR4 isolates, which is a major drawback for regions where STR4 is endemic and that still have no record of TR4. Alternative primers targeting a ribosomal IGS have been developed and found to be quite reliable (Dita et al., 2010). This region was chosen as it was deemed more polymorphic than others and more suitable as a sensitive diagnostic test due to the multi-copy nature of this region (Fourie et al., 2009; Dita et al., 2010), which was confirmed by our tests (Supplementary Table S1). Nonetheless, the primers FocTR4 which were proposed by Dita et al., 2010 were tested in another study which included endophytic strains isolated from healthy banana plants (Magdama, 2017). An endophytic F. oxysporum strain which was obtained from Gros Michel roots tested positive although it was isolated from an asymptomatic plant grown in a region where TR4 is absent (Magdama, 2017). In our study, we also obtained false positives for one endophytic strain isolated from an asymptomatic banana plant, one STR4 and three F. oxysporum f. sp. vasinfectum isolates with primers FocTR4 (Dita et al., 2010; Table 5). It is possible that there is a higher risk of obtaining false positives when targeting core genomic regions, as genetically related strains may contain the same sequence and still differ in pathogenicity. Primers designed to amplify genes or sequences associated with pathogenicity may be more specific in a diagnostic assay; however, their sensitivity may be lower compared to other regions that are often present as multiple copies in the genome, such as ribosomal regions (Black et al., 2013).
It is important to point out that our molecular diagnostics toolkit was designed and validated to be used only with DNA extracted from pure cultures of F. oxysporum as templates. Thus, we strongly discourage the use of our assays beyond the parameters validated here. Further tests with the use of DNA extracted from infected plant material are needed and require extensive validation. The ubiquitous nature of F. oxysporum existing in plant tissue as saprophytic and/or endophytic strains with apparent diverse and fickle genotypes renders the development of molecular based diagnostic assays direct from plant tissue, challenging. This would be the main reason why we would be hesitant to convert our conventional PCR assays into loop-mediated isothermal amplification (LAMP) assays, which are mainly used either in crude samples extracts or DNA extracted directly from tissues rather than cultures (Zhang et al., 2014). LAMP is a technique that is used for amplifying a specific segment of DNA under isothermal conditions through the strand-displacing Bst DNA polymerase (Zhang et al., 2014). This tool is commonly advocated as being sensitive, low cost and mobile; however, the combined price of the needed equipment, kits and reagents is considerable. In addition, some expertise is needed for the interpretation of the results in the field, suitable controls are also necessary, and results need to be confirmed in a centralized laboratory through an alternative effective diagnostic assay. Our results also suggest that there is not enough variability in the SIX gene sequences across Foc races to allow the design of four primers that recognize six distinct regions. For example, our assay for TR4 relies on the presence of two SNPs in the SIX1 homolog “a,” which is exclusive to TR4 strains and is part of the recognition site of the HpyAV enzyme.
The availability of specific, sensitive, and robust diagnostic assays to identify plant pathogens is vital for the early detection and further containment or eradication of plant diseases. Accurate identification of the race or VCG of Foc using a sensitive, robust, user-friendly, and accessible assay by laboratories in any part of the world can provide reliable diagnostics to growers. This would also assist local governments to take suitable control measures to prevent threats to food security or economic losses to the banana industry.
JH, EA, EC, and AD conceived the study. JH, AD, and LC designed the experiments. EA and EC provided the sequences for primer design. CO, VR-F, and LC carried out the experiments. LC and AD drafted the manuscript. All authors contributed to the final manuscript.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
We would like to acknowledge Hort Innovation Australia for the funding (Project BA16005). We thank Dean Beasley and Tuan Nguyen from the Department of Agriculture and Fisheries of the Queensland Government in Australia for their subculturing and supplying the fungal cultures, and Anna Smyth for technical assistance.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fpls.2019.00547/full#supplementary-material
FIGURE S1 | Example of agarose gel showing the limit of detection of amplification products obtained with the primer set SIX13c_343 within a range of DNA template concentrations.
TABLE S1 | Limit of detection of primer sets SIX1a_266, SIX6b_210, SIX8b_206, SIX9_Foc, SIX10a_309, SIX13c_343, and the previously published primers Foc-TR4 (Dita et al., 2010), and EF (O’Donnell et al., 1998) within the DNA template concentration range of 10 ng⋅μL-1 to 0.1 fg⋅μL-1.
Alexander, L. J. (1945). Physiologic specialization in the tomato wilt fungus Fusarium oxysporum f. sp. lycopersici. J. Agric. Res. 70, 303–313.
Armstrong, G. (1981). “Formae speciales and races of Fusarium oxysporum causing wilt diseases,” in Fusarium: Diseases, Biology, and Taxonomy, eds P. Nelson, T. Toussoun, and R. Cook (University Park, PA: Pennsylvania State University Press), 391–399.
Ayukawa, Y., Hanyuda, S., Fujita, N., Komatsu, K., and Arie, T. (2017). Novel loop-mediated isothermal amplification (LAMP) assay with a universal QProbe can detect SNPs determining races in plant pathogenic fungi. Sci. Rep. 7:4253. doi: 10.1038/s41598-017-04084-y
Bentley, S., Pegg, K. G., and Dale, J. L. (1995). Genetic variation among a world-wide collection of isolates of Fusarium oxysporum f. sp. cubense analysed by RAPD-PCR fingerprinting. Mycol. Res. 99, 1378–1384. doi: 10.1016/s0953-7562(09)81225-2
Bentley, S., Pegg, K. G., Moore, N. Y., Davis, R. D., and Buddenhagen, I. W. (1998). Genetic variation among vegetative compatibility groups of Fusarium oxysporum f. sp. cubense analyzed by DNA fingerprinting. Phytopathology 88, 1283–1293. doi: 10.1094/PHYTO.1998.88.12.1283
Black, J., Dean, T., Byfield, G., Foarde, K., and Menetrez, M. (2013). Determining fungi rRNA copy number by PCR. J. Biomol. Tech. 24, 32–38. doi: 10.7171/jbt.13-2401-004
Buddenhagen, I. (2009). “Understanding strain diversity in Fusarium oxysporum f. sp. cubense and history of introduction of’ tropical race 4 to better manage banana production,” in Prodeedings of the 3rd International Symposium on Banana: ISHS-ProMusa Symposium on Recent Advances in Banana Crop Protection for Sustainable, (Belgium: ISHS), 193–204. doi: 10.17660/ActaHortic.2009.828.19
Caten, C. E., and Jinks, J. L. (1966). Heterokaryosis - its significance in wild homothallic Ascomycetes and Fungi Imperfecti. Br. Mycol. Soc. Trans. 49, 81–93. doi: 10.1016/s0007-1536(66)80038-4
Chakrabarti, A., Rep, M., Wang, B., Ashton, A., Dodds, P., and Ellis, J. (2011). Variation in potential effector genes distinguishing Australian and non-Australian isolates of the cotton wilt pathogen Fusarium oxysporum f.sp. vasinfectum. Plant Pathol. 60, 232–243. doi: 10.1111/j.1365-3059.2010.02363.x
Chilvers, M. I. (2012). Molecular diagnostics in plant disease diagnostic clinics. What’s the status? Fungal Genom. Biol. 2:e102. doi: 10.4172/2165-8056.1000e102
Chittarath, K., Mostert, D., Crew, K. S., Viljoen, A., Kong, G., Molina, A. B., et al. (2018). First report of Fusarium oxysporum f. sp. cubense Tropical Race 4 (VCG 01213/16) associated with Cavendish Bananas in Laos. Plant Dis. 102, 449–450. doi: 10.1094/PDIS-09-13-0954-PDN
Czislowski, E., Fraser-Smith, S., Zander, M., O’Neill, W. T., Meldrum, R. A., Tran-Nguyen, L. T. T., et al. (2017). Investigation of the diversity of effector genes in the banana pathogen, Fusarium oxysporum f. sp. cubense, reveals evidence of horizontal gene transfer. Mol. Plant Pathol. 19, 1155–1171. doi: 10.1111/mpp.12594
Dale, J., James, A., Paul, J. Y., Khanna, H., Smith, M., Peraza-Echeverria, S., et al. (2017). Transgenic Cavendish bananas with resistance to Fusarium wilt tropical race 4. Nat. Commun. 8:1496. doi: 10.1038/s41467-017-01670-6
de Sain, M., and Rep, M. (2015). The role of pathogen-secreted proteins in fungal vascular wilt diseases. Int. J. Mol. Sci. 16, 23970–23993. doi: 10.3390/ijms161023970
Dita, M., Barquero, M., Heck, D., Mizubuti, E. S. G., and Staver, C. P. (2018). Fusarium wilt of banana: current knowledge on epidemiology and research needs toward sustainable disease management. Front. Plant Sci. 9:1468. doi: 10.3389/fpls.2018.01468
Dita, M. A., Waalwijk, C., Buddenhagen, I. W., Souza, M. T., and Kema, G. H. J. (2010). A molecular diagnostic for tropical race 4 of the banana Fusarium wilt pathogen. Plant Pathol. 59, 348–357. doi: 10.1111/j.1365-3059.2009.02221.x
FAOSTAT (2018). FAOSTAT Online Database. Available at http://faostat.fao.org/ (accessed December 2018).
Fourie, G., Steenkamp, E. T., Gordon, T. R., and Viljoen, A. (2009). Evolutionary relationships among the Fusarium oxysporum f. sp. cubense vegetative compatibility groups. Appl. Environ. Microbiol. 75, 4770–4781. doi: 10.1128/AEM.00370-09
Fraser-Smith, S., Czislowski, E., Meldrum, R. A., Zander, M., O’Neill, W., Balali, G. R., et al. (2014). Sequence variation in the putative effector gene SIX8 facilitates molecular differentiation of Fusarium oxysporum f. sp. cubense. Plant Pathol. 63, 1044–1052.
García-Bastidas, F., Ordóñez, N., Konkol, J., Al-Qasim, M., Naser, Z., Abdelwali, M., et al. (2014). First report of Fusarium oxysporum f. sp. cubense tropical race 4 associated with Panama disease of banana outside Southeast Asia. Plant Dis. 98:694. doi: 10.1094/PDIS-09-13-0954-PDN
Gawehns, F., Houterman, P. M., Ichou, F. A., Michielse, C. B., Hijdra, M., Cornelissen, B. J. C., et al. (2014). The Fusarium oxysporum effector SIX6 contributes to virulence and suppresses I-2-mediated cell death. Mol. Plant Microbe Interact. 27, 336–348. doi: 10.1094/MPMI-11-13-0330-R
Gerlach, K. S., Bentley, S., Moore, N. Y., Pegg, K. G., and Aitken, E. A. B. (2000). Characterisation of Australian isolates of Fusarium oxysporum f. sp. cubense by DNA fingerprinting. Aust. J. Agric. Res. 51, 945–953.
Gerland, P., Raftery, A. E., Sevcikova, H., Li, N., Gu, D. A., Spoorenberg, T., et al. (2014). World population stabilization unlikely this century. Science 346, 234–237. doi: 10.1126/science.1257469
Groenewald, S., Van den Berg, N., Marasas, W. F. O., and Viljoen, A. (2006). The application of high-throughput AFLP’s in assessing genetic diversity in Fusarium oxysporum f. sp. cubense. Mycol. Res. 110, 297–305. doi: 10.1016/j.mycres.2005.10.004
Houterman, P. M., Speijer, D., Dekker, H. L., de Koster, C. G., Cornelissen, B. J. C., and Rep, M. (2007). The mixed xylem sap proteome of Fusarium oxysporum-infected tomato plants. Mol. Plant Pathol. 8, 215–221. doi: 10.1111/j.1364-3703.2007.00384.x
Hung, T. N., Hung, N. Q., Mostert, D., Viljoen, A., Chao, C. P., and Molina, A. B. (2018). First report of Fusarium wilt on Cavendish bananas, caused by Fusarium oxysporum f. sp. cubense Tropical Race 4 (VCG 01213/16), in Vietnam. Plant Dis. 102, 448–448. doi: 10.1094/pdis-08-17-1140-pdn
Jelinski, N. A., Broz, K., Jonkers, W., Ma, L. J., and Kistler, H. C. (2017). Effector gene suites in some soil isolates of Fusarium oxysporum are not sufficient predictors of vascular wilt in tomato. Phytopathology 107, 842–851. doi: 10.1094/PHYTO-12-16-0437-R
Karangwa, P., Blomme, G., Beed, F., Niyongere, C., and Viljoen, A. (2016). The distribution and incidence of banana Fusarium wilt in subsistence farming systems in east and central Africa. Crop Prot. 84, 132–140. doi: 10.1016/j.cropro.2016.03.003
Katan, T., and Di Primo, P. (1999). Current status of vegetative compatibility groups in Fusarium oxysporum. Phytoparasitica 27, 273–277. doi: 10.1016/j.meegid.2011.01.012
Kazan, K., and Lyons, R. (2014). Intervention of phytohormone pathways by pathogen effectors. Plant Cell 26, 2285–2309. doi: 10.1105/tpc.114.125419
Kearse, M., Moir, R., Wilson, A., Stones-Havas, S., Cheung, M., Sturrock, S., et al. (2012). Geneious Basic: an integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics 28, 1647–1649. doi: 10.1093/bioinformatics/bts199
Kim, K. T., Jeon, J., Choi, J., Cheong, K., Song, H., Choi, G., et al. (2016). Kingdom-wide analysis of fungal small secreted proteins (SSPs) reveals their potential role in host association. Front. Plant Sci. 7:186. doi: 10.3389/fpfs.2016.00186
Leslie, J. F. (1993). Fungal vegetative compatibility. Annu. Rev. Phytopathol. 31, 127–150. doi: 10.1146/annurev.py.31.090193.001015
Li, W. M., Dita, M., Wu, W., Hu, G. B., Xie, J. H., and Ge, X. J. (2015). Resistance sources to Fusarium oxysporum f. sp. cubense tropical race 4 in banana wild relatives. Plant Pathol. 64, 1061–1067.
Lievens, B., Houterman, P. M., and Rep, M. (2009). Effector gene screening allows unambiguous identification of Fusarium oxysporum f.sp. lycopersici races and discrimination from other formae speciales. FEMS Microbiol. Lett. 300, 201–215. doi: 10.1111/j.1574-6968.2009.01783.x
Lin, Y. H., Chang, J. Y., Liu, E. T., Chao, C. P., Huang, J. W., and Chang, P. F. L. (2009). Development of a molecular marker for specific detection of Fusarium oxysporum f. sp. cubense race 4. Eur. J. Plant Pathol. 123, 353–365. doi: 10.1007/s10658-008-9372-4
Loreti, S., and Gallelli, A. (2002). Rapid and specific detection of virulent Pseudomonas avellanae strains by PCR amplification. Eur. J. Plant Pathol. 108, 237–244.
Magdama, F. (2017). Population Biology of Fusarium oxysporum Associated with Banana in Ecuador. doctor of philosophy dissertation. University Park, PA: Pennsylvania State University.
Maryani, N., Lombard, L., Poerba, Y. S., Subandiyah, S., Crous, P. W., and Kema, G. H. J. (2019). Phylogeny and genetic diversity of the banana Fusarium wilt pathogen Fusarium oxysporum f. sp. cubense in the Indonesian centre of origin. Stud. Mycol. 92, 155–194. doi: 10.1016/j.simyco.2018.06.003
Moore, N., Bentley, S., Pegg, K., and Jones, D. (1995). Musa Disease Fact Sheet No 5. Fusarium wilt of Banana. Montpellier: INIBAP.
Moore, N. Y., Hargreaves, P. A., Pegg, K. G., and Irwin, J. A. G. (1991). Characterization of strains of Fusarium oxysporum f. sp. cubense by production of volatiles. Aust. J. Bot. 39, 161–166.
Moore, N. Y., Pegg, K. G., Allen, R. N., and Irwin, J. A. G. (1993). Vegetative compatibility and distribution of Fusarium oxysporum f. sp. cubense in Australia. Aust. J. Exp. Agric. 33, 797–802. doi: 10.1371/journal.pone.0181630
Mostert, D., Molina, A. B., Daniells, J., Fourie, G., Hermanto, C., Chao, C. P., et al. (2017). The distribution and host range of the banana Fusarium wilt fungus, Fusarium oxysporum f. sp. cubense, in Asia. PLoS One 12:e0181630. doi: 10.1371/journal.pone.0181630
National Association of Testing Authorities (2018). “Guidelines for the Validation and Verification of Quantitative and Qualitative Test Methods”, in: Technical Note 17. Canberra, ACT: National Association of Testing Authorities.
NCBI Resource Coordinators (2017). Database resources of the national center for biotechnology information. Nucleic Acids Res. 41, D8–D20. doi: 10.1093/har/gkw1071
O’Donnell, K., Kistler, H. C., Cigelnik, E., and Ploetz, R. C. (1998). Multiple evolutionary origins of the fungus causing Panama disease of banana: concordant evidence from nuclear and mitochondrial gene genealogies. Proc. Natl. Acad. Sci. U.S.A. 95, 2044–2049. doi: 10.1073/pnas.95.5.2044
Ordonez, L. N., Garcia-Bastidas, F., Laghari, H. B., Akkary, M. Y., Harfouche, E. N., al Awar, B. N., et al. (2016). First report of Fusarium oxysporum f. sp. cubense tropical race 4 causing Panama disease in Cavendish bananas in Pakistan and Lebanon. Plant Dis. 100, 209–210.
Ordonez, N., Seidl, M. F., Waalwijk, C., Drenth, A., Kilian, A., Thomma, B. P. H. J., et al. (2015). Worse comes to worst: bananas and Panama disease - when plant and pathogen clones meet. PLoS Pathog. 11:e1005197. doi: 10.1371/journal.ppat.1005197
Pegg, K., Moore, N., and Sorenson, S. (1994). “Variability in populations of Fusarium oxysporum f. sp. cubense from the Asia/Pacific region,” in The improvement and testing of Musa: a global partnership, ed. D. R. Jones (Honduras: FHIA), 70–82.
Pérez Vicente, L., Dita, M., and Martinez De La Parte, E. (2014). Technical Manual: Prevention and Diagnostic of Fusarium wilt (Panama disease) of Banana Caused by Fusarium oxysporum f. sp. cubense Tropical Race 4 (TR4). Rome: FAO.
Pillay, M., and Tenkouano, A. (2011). Banana Breeding: Progress and Challenges. Boca Raton, FL: CRC Press.
Ploetz, R. C. (1994). Panama disease: return of the first banana menace. Int. J. Pest Manag. 40, 326–336. doi: 10.1080/09670879409371908
Ploetz, R. C. (2004). Diseases and pests: a review of their importance and management. InfoMusa 13, 11–16.
Ploetz, R. C. (2005). Panama disease, an old nemesis rears its ugly head: part 1, the beginnings of the banana export trades. Plant Health Prog. 1, 1–10. doi: 10.1094/PHP-2005-1221-01-RV
Ploetz, R. C. (2015a). Fusarium wilt of banana. Phytopathology 105, 1512–1521. doi: 10.1094/phyto-04-15-0101-rvw
Ploetz, R. C. (2015b). Management of Fusarium wilt of banana: a review with special reference to tropical race 4. Crop Prot. 73, 7–15. doi: 10.1016/j.cropro.2015.01.007
Ploetz, R. C., and Correll, J. C. (1988). Vegetative compatibility among races of Fusarium oxysporum f. sp. cubense. Plant Dis. 72, 325–328.
Rep, M. (2005). Small proteins of plant-pathogenic fungi secreted during host colonization. FEMS Microbiol. Lett. 253, 19–27. doi: 10.1016/j.femsle.2005.09.014
Rep, M., van der Does, H. C., Meijer, M., van Wijk, R., Houterman, P. M., Dekker, H. L., et al. (2004). A small, cysteine-rich protein secreted by Fusarium oxysporum during colonization of xylem vessels is required for I-3-mediated resistance in tomato. Mol. Microbiol. 53, 1373–1383. doi: 10.1111/j.1365-2958.2004.04177.x
Rocha, L. O., Laurence, M. H., Ludowici, V. A., Puno, V. I., Lim, C. C., Tesoriero, L. A., et al. (2016). Putative effector genes detected in Fusarium oxysporum from natural ecosystems of Australia. Plant Pathol. 65, 914–929. doi: 10.1111/ppa.12472
Schierwater, B., and Ender, A. (1993). Different thermostable DNA polymerases may amplify differentRAPD products. Nucleic Acids Res. 21, 4647–4648. doi: 10.1093/nar/21.19.4647
Serdani, M., Curtis, M., Miller, M. L., Kraus, J., and Putnam, M. L. (2013). Loop-mediated isothermal amplification and polymerase chain reaction methods for specific and rapid detection of Rhodococcus fascians. Plant Dis. 97, 517–529. doi: 10.1094/PDIS-02-12-0214-RE
Sievers, F., and Higgins, D. G. (2014). Clustal Omega, accurate alignment of very large numbers of sequences. Methods Mol. Biol. 1079, 105–116. doi: 10.1007/978-1-62703-646-7_6
Stover, R. (1990). “Fusarium wilt of banana: some history and current status of the disease,” in Fusarium wilt of Banana, ed. R. C. Ploetz (St. Paul, MN: APS Press), 1–7.
Stover, R., and Simmonds, N. (1987). Bananas. Tropical agricultural series. New York, NY: John Wiley and Sons, Inc, 158–468.
Stover, R. H. (1962). Fusarial wilt (Panama Disease) of bananas and other Musa species. Kew: Commonwealth Mycological Institute.
Stover, R. H., and Buddenhagen, I. W. (1986). Banana breeding - polyploidy, disease resistance and productivity. Fruits 41, 175–191.
Su, H. J., Chuang, T. Y., and Kong, W. S. (1977). Physiological race of fusarial wilt fungus attacking Cavendish banana of Taiwan. Taiwan Banana Res. Inst. Spec. Publ. 2, 1–21.
Thatcher, L. F., Gardiner, D. M., Kazan, K., and Manners, J. M. (2012). A highly conserved effector in Fusarium oxysporum is required for full virulence on Arabidopsis. Mol. Plant Microbe Interact. 25, 180–190. doi: 10.1094/MPMI-08-11-0212
Untergasser, A., Cutcutache, I., Koressaar, T., Ye, J., Faircloth, B. C., Remm, M., et al. (2012). Primer3 - new capabilities and interfaces. Nucleic Acids Res. 40, e115. doi: 10.1093/nar/gks596
van Dam, P., Fokkens, L., Schmidt, S. M., Linmans, J. H. J., Kistler, H. C., Ma, L. J., et al. (2016). Effector profiles distinguish formae speciales of Fusarium oxysporum. Environ. Microbiol. 18, 4087–4102. doi: 10.1111/1462-2920.13445
Wei, H., Therrien, C., Blanchard, A., Guan, S., and Zhu, Z. (2008). The fidelity index provides a systematic quantitation of star activity of DNA restriction endonucleases. Nucleic Acids Res. 36:e50. doi: 10.1093/nar/gkn182
Weiberg, A., Wang, M., Lin, F. M., Zhao, H. W., Zhang, Z. H., Kaloshian, I., et al. (2013). Fungal small RNAs suppress plant immunity by hijacking host RNA interference pathways. Science 342, 118–123. doi: 10.1126/science.1239705
Yang, L. L., Sun, L. X., Ruan, X. L., Qiu, D. Y., Chen, D. H., Cai, X. Q., et al. (2015). Development of a single-tube duplex real-time fluorescence method for the rapid quantitative detection of Fusarium oxysporum f. sp. cubense race 1 (FOC1) and race 4 (FOC4) using TaqMan probes. Crop Prot. 68, 27–35. doi: 10.1016/j.cropro.2014.11.004
Zaccardelli, M., Campanile, F., Spasiano, A., and Merighi, M. (2007). Detection and identification of the crucifer pathogen, Xanthomonas campestris pv. campestris, by PCR amplification of the conserved Hrp/type III secretion system gene hrcC. Eur. J. Plant Pathol. 118, 299–306. doi: 10.1007/s10658-007-9115-y
Zhang, X., Lowe, S. B., and Gooding, J. J. (2014). Brief review of monitoring methods for loop-mediated isothermal amplification (LAMP). Biosens. Bioelectron. 61, 491–499. doi: 10.1016/j.bios.2014.05.039
Keywords: tropical race 4, molecular diagnostics, Panama disease, SIX genes, plant pathogens, Fusarium oxysporum
Citation: Carvalhais LC, Henderson J, Rincon-Florez VA, O’Dwyer C, Czislowski E, Aitken EAB and Drenth A (2019) Molecular Diagnostics of Banana Fusarium Wilt Targeting Secreted-in-Xylem Genes. Front. Plant Sci. 10:547. doi: 10.3389/fpls.2019.00547
Received: 21 December 2018; Accepted: 10 April 2019;
Published: 31 May 2019.
Edited by:
Jesús Mercado-Blanco, Instituto de Agricultura Sostenible (IAS), SpainReviewed by:
David J. Studholme, University of Exeter, United KingdomCopyright © 2019 Carvalhais, Henderson, Rincon-Florez, O’Dwyer, Czislowski, Aitken and Drenth. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Lilia C. Carvalhais, bC5jYXJ2YWxoYWlzQHVxLmVkdS5hdQ==
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.