
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Plant Sci., 24 October 2018
Sec. Plant Breeding
Volume 9 - 2018 | https://doi.org/10.3389/fpls.2018.01547
This article is part of the Research TopicGenetics and Genomics of Plant Reproduction for Crop BreedingView all 25 articles
 Micaela Mancini1
Micaela Mancini1 Hugo Permingeat1
Hugo Permingeat1 Carolina Colono1
Carolina Colono1 Lorena Siena1
Lorena Siena1 Fulvio Pupilli2
Fulvio Pupilli2 Celeste Azzaro1
Celeste Azzaro1 Diva Maria de Alencar Dusi3
Diva Maria de Alencar Dusi3 Vera Tavares de Campos Carneiro3
Vera Tavares de Campos Carneiro3 Maricel Podio1
Maricel Podio1 José Guillermo Seijo4,5
José Guillermo Seijo4,5 Ana María González4,6
Ana María González4,6 Silvina A. Felitti1
Silvina A. Felitti1 Juan Pablo A. Ortiz1
Juan Pablo A. Ortiz1 Olivier Leblanc7
Olivier Leblanc7 Silvina C. Pessino1*
Silvina C. Pessino1*Apomixis is a clonal mode of reproduction via seeds, which results from the failure of meiosis and fertilization in the sexual female reproductive pathway. In previous transcriptomic surveys, we identified a mitogen-activated protein kinase kinase kinase (N46) displaying differential representation in florets of sexual and apomictic Paspalum notatum genotypes. Here, we retrieved and characterized the N46 full cDNA sequence from sexual and apomictic floral transcriptomes. Phylogenetic analyses showed that N46 was a member of the YODA family, which was re-named QUI-GON JINN (QGJ). Differential expression in florets of sexual and apomictic plants was confirmed by qPCR. In situ hybridization experiments revealed expression in the nucellus of aposporous plants’ ovules, which was absent in sexual plants. RNAi inhibition of QGJ expression in two apomictic genotypes resulted in significantly reduced rates of aposporous embryo sac formation, with respect to the level detected in wild type aposporous plants and transformation controls. The QGJ locus segregated independently of apospory. However, a probe derived from a related long non-coding RNA sequence (PN_LNC_QGJ) revealed RFLP bands cosegregating with the Paspalum apospory-controlling region (ACR). PN_LNC_QGJ is expressed in florets of apomictic plants only. Our results indicate that the activity of QGJ in the nucellus of apomictic plants is necessary to form non-reduced embryo sacs and that a long non-coding sequence with regulatory potential is similar to sequences located within the ACR.
Asexual reproduction can naturally occur in ovules of several flowering plant taxa through apomixis, an alternative route to sexuality, which allows the formation of maternal embryos within seeds (Nogler, 1984; Carman, 1997). This atypical trait relies on developmental alterations which cause unreduced cells within the ovule to acquire a reproductive fate. Although mechanistically diverse, apomictic pathways are usually classified into two major classes (i.e., sporophytic and gametophytic), depending on the origin of maternal embryos (Hand and Koltunow, 2014). During sporophytic apomixis, embryogenesis occurs spontaneously in somatic cells of the ovule, leading to the formation of seeds that harbor supernumerary maternal embryos. In contrast, gametophytic apomixis involves the differentiation of functional, unreduced embryo sacs (2n-ES) within the ovule, followed by egg cell parthenogenetic development into embryos (Bicknell and Koltunow, 2004). Depending on the origin of 2n-ESs, gametophytic apomixis can be further subcategorized into: (1) diplospory, when the megaspore mother cell (MMC) fails meiosis and enters into gametogenesis; or (2) apospory, when one or several nucellar or integumental cells, which are usually somatic companions of the MMC, acquire a gametic fate. In all gametophytic apomicts the embryo develops autonomously, while the formation of the endosperm can be either autonomous or fertilization-dependent (pseudogamous) (Bicknell and Koltunow, 2004).
In the last decade, transcriptomic surveys have allowed the identification of hundreds of candidate genes allegedly associated with apomixis in di- and monocotyledonous plants. However, predictions for most of these candidates revealed they belong to a few functional categories, including signal transduction, cell-cycle control, protein turnover, intercellular signaling, transposon activity and transcriptional regulation (Pessino et al., 2001; Rodrigues et al., 2003; Albertini et al., 2004; Cervigni et al., 2008; Laspina et al., 2008; Sharbel et al., 2009, 2010; Yamada-Akiyama et al., 2009; Garcia-Aguilar et al., 2010; Polegri et al., 2010; Okada et al., 2013). Particularly, a transcript fragment (N46) displaying homology with mitogen-activated protein kinase kinase kinases (MAP3K/MAPKKK/MEKK) and similarity to the Arabidopsis gene At1g53570 (MAPKKK3) was identified in Paspalum notatum, an aposporous sub-tropical grass (Laspina et al., 2008). Interestingly, another transcript showing homology with the same Arabidopsis gene (A-148-3) mapped to the apospory controlling region (ACR) of Paspalum simplex, a single, non-recombinant, dominant superlocus, which confers nearly 100% apospory (Polegri et al., 2010), epigenetically controlled parthenogenesis (Podio et al., 2014a) and the capacity to form endosperm with unbalanced parental genome contributions (Ortiz et al., 2013).
Based on these results and considering the essential roles of MAPKs in plant development (Musielak and Bayer, 2014; Xu and Zhang, 2015), we rationalized that the P. notatum At1g53570 homolog N46 might be involved in the switch from sexuality to aposporous apomixis in this species. The central biological question of our work was the following: is the Paspalum At1g53570 ortholog (N46) involved in the developmental molecular cascade controlling apospory, either as trigger or participant? To test this hypothesis, we first mined transcriptomic resources available for P. notatum to complete the characterization of N46 sequences. We also conducted spatio-temporal expression analyses in sexual and apomictic genotypes of P. notatum, and used Brachiaria brizantha, a related aposporous species, as a validation control. Finally, we made functional analyses in P. notatum by producing RNA interference (RNAi) lines. Moreover, we mapped N46 onto the P. notatum genome to explore the occurrence of genetic linkage with apomixis. Finally, we determined that the At1g53570-like transcript previously identified by Polegri et al. (2010) (see above) was not a protein-coding ortholog of At1g53570 and N46, but a lncRNA showing only partial similarity with these genes.
The P. notatum genotypes used in this work belong to the IBONE’s germplasm collection (Instituto de Botánica del Nordeste, UNNE-CONICET, Corrientes, Argentina) and are listed below: (i) natural apomictic tetraploid genotype Q4117 (2n = 4x = 40) (Ortiz et al., 1997); (ii) experimentally obtained sexual tetraploid genotype Q4188 (2n = 4x = 40) (Quarin et al., 2003); (iii) colchicine-treated sexual double-diploid genotype C4-4x (2n = 4x = 40) (Quarin et al., 2001); and (iv) 55 F1 hybrids derived from a cross between Q4188 and Q4117, 29 of them sexual and 26 apomictic (Stein et al., 2007). The Brachiaria brizantha genotypes used here were: (i) sexual diploid genotype BRA 002747 (B105) (2n = 2x = 18); and (ii) cultivar Marandu BRA 00591 (B30), a facultative tetraploid apomictic (2n = 4x = 36). Both genotypes belong to the Embrapa’s germplasm collection and are maintained at Embrapa Genetic Resources and Biotechnology, Brasilia Federal District, Brazil.
The N46 full-length sequences were retrieved from 454/Roche FLX + floral transcriptome databases generated in prior work (Ortiz et al., 2017) and available at DDBJ/ENA/GenBank under the accessions GFMI00000000 and GFNR00000000, versions GFMI02000000 and GFNR01000000, respectively. Analysis of DNA similarity was done by using the BLASTN and BLASTX packages at the NCBI1, the Arabidopsis Information Resource2 and the Gramene3 websites, as well as exploring the Oryza Repeats Database4. For open reading frame (ORF) detection, the NCBI ORF Finder tool was used5. Gene schemes were constructed with the WormWeb Exon-Intron graphic maker6. Alignments and phylogenetic analyses were done with ClustalW2 (Larkin et al., 2007) and MEGA6 (Tamura et al., 2013) software packages, respectively. The evolutionary history was inferred using the UPGMA method (Sneath and Sokal, 1973). Evolutionary distances were computed using the Poisson correction method (Zuckerkandl and Pauling, 1965) (units: number of amino acid substitutions per site). The lncRNA similarity survey was done onto the plant lncRNA GreeNC database (Paytuví Gallart et al., 2016).
Genomic DNA was extracted from 200 mg of leaves by using CTAB (Paterson et al., 1993). To reveal the presence of the 76-nt intron in the genome, amplification reactions were carried out with a primer pair complementary to the intron flanking regions: FIP upper (5′-ATTTGCAAGGACCAACATCC-3′, Tm: 59.80°C) and FIP lower (5′-ATGGCAAGCAACTTCGATTC-3′, Tm: 60.22°C). To amplify the entire QGJ sequence we used the following primers: N46full upper: 5′GCGTGTACGCCTCTCTCTCT3′, Tm: 59.78°C; N46full lower: 5′CTGCATCCTGGGTGAAAAAT3′, Tm: 59.93°C. Reactions (final volume 25 μL) included 1X Real mix qPCR (BIODYNAMICS), 2 mM MgCl2, 200 μM dNTPs, 200 nM gene-specific primers and 60 ng genomic DNA. Amplifications were performed in a BIO-RAD thermocycler, programmed as follows: 1 min at 94°C, 35 cycles of 1 min at 94°C, 2 min at 57°C and 2 min at 72°C, and a final elongation step of 5 min at 72°C.
To evaluate the representation of different splice variants by semiquantitative RT-PCR, total RNA was extracted from leaves and/or spikelets at premeiosis/meiosis and reverse transcribed using Superscript II (INVITROGEN). Amplifications were conducted with: (1) the same primer pair flanking the 76-nt intron described above; or (2) a primer pair located inside the intron: IP upper (5′-AAACAGCATGGTGCAGTCAA-3′, Tm: 60.31°C) and IP lower (5′-TCAGGTGGACAATTGATGAGA-3′, Tm: 59.07°C). Each reaction (final volume 25 μL) included the same components used for genomic amplifications and were run in the same thermocycler, but 20 ng of cDNA were used as template, and the cycling was 5 min at 94°C, 25 cycles of 30 s at 94°C, 30 s at 57°C and 45 s at 72°C and a final elongation step of 5 min at 72°C.
To quantitate QGJ expression in reproductive organs, the following samples were collected: (1) spikelets of apomictic and sexual P. notatum and B. brizantha plants (P. notatum: Q4117 and C4-4x genotypes; B. brizantha: B30 and B105 genotypes) at premeiosis, meiosis and postmeiosis; (2) ovaries of apomictic and sexual B. brizantha plants (B30 and B105 genotypes) at the same above-mentioned stages; (3) spikelets at meiosis/young leaves of wild type (Q4117 genotype, apomictic) and transformant (RNAi1, RNAi2, TC1, TC2) P. notatum plants. Total RNA was extracted with the SV Total RNA Isolation Kit (PROMEGA), which includes a DNAse treatment step. cDNAs were synthetized with Superscript II (INVITROGEN). All qPCR reactions (final volume: 25 μL) included 200 nM gene-specific primers, 1X Real mix qPCR (BIODYNAMICS) and 20 ng of cDNA. Three biological replicates were processed each into three technical replicates. Replicates with templates produced in the absence of Superscript II (INVITROGEN) and without templates were included (negative controls). Amplifications were performed in a Rotor-Gene Q thermocycler (QIAGEN), programmed as follows: 2 min at 94°C, 45 cycles of 15 s at 94°C, 30 s at 62°C and 40 s at 72°C and a final elongation step of 5 min at 72°C. QGJ-specific primers were: (1) N46N upper (5′GGCCCTGCATCTCCTACTTCAT3′, Tm: 68°C) and N46N lower (5′’TGCCCAAACGTCCCACTGC3′, Tm: 62°C), which amplified QGJ in all allelic contexts (used for chronological expression analysis); (2) N46S upper (5′AATCGAAGTTGCTTGCCATC3′, Tm: 60°C) and N46S lower (5′GCTCTGTTAGACCGCTGCTT3′, Tm: 59°C), which were located outside the N46 segment cloned into the pBS86-N46 vector (used for analysis of expression in transgenic plants). Non-template reactions were included as controls. β-tubulin was used as an internal reference gene, as recommended by Felitti et al. (2011), Ochogavía et al. (2011), and Podio et al. (2014b), who worked in the same plant model. Relative quantitative expression levels were calculated by using REST-RG (Relative Expression Software Tool V 2.0.7 for Rotor Gene, Corbett Life Sciences) considering take-off and amplification efficiency values for each particular reaction.
Spikelets of P. notatum (genotypes Q4117 and C4-4x) and B. brizantha (genotypes B30 and B105) were collected at premeiosis/meiosis. Flowers were dissected, fixed in 4% paraformaldehyde/0.25% glutaraldehyde/0.01 M phosphate buffer pH 7.2, dehydrated in an ethanol series and embedded in paraffin (for P. notatum) or butyl-methyl-methacrylate (BMM) (for B. brizantha). Specimens were cut into sections of 10 μm (Paspalum) or 3.5 μm (Brachiaria) and placed onto slides treated with poly-L-lysine 100 μg/mL. The paraffin or BMM were removed with xylene or acetone series, respectively. Prior to hybridization, control sections were stained with acridine orange and examined under UV light to verify RNA integrity. A plasmid including the original N46 fragment isolated by Laspina et al. (2008) was linearized using restriction enzymes NcoI or SalI (Promega). Sense and anti-sense probes were labeled with the Roche Dig RNA Labeling kit (SP6/T7), following the manufacturers’ instructions, and hydrolyzed to 150–200 bp fragments. Prehybridization was carried out in 0.05 M Tris–HCl pH 7.5 buffer containing 1 μg/mL proteinase K in a humid chamber at 37°C for 10 min. Hybridization was carried out overnight in a humid chamber at 42°C, in 10 mM Tris–HCl pH 7.5 buffer containing 300 mM NaCl, 50% formamide (deionized), 1 mM EDTA pH 8, 1 X Denhardt’s solution, 10% dextran sulfate, 600 ng/mL tRNA and 600 ng/mL of probe. Detection was performed following the instructions of the Roche Dig Detection kit, using anti DIG AP and NBT/BCIP as substrates. Sections were mounted in glycerol 50% and observed under Leica DMRX (Paspalum experiments) or Zeiss-Axiophot (Brachiaria experiments) light microscopes.
A vector containing an N46 hairpin (pBS86-N46) (Pact1D:rfa-n46-s:rga2i:rfa-n46-as:T35s/Pubi:bar:Tnos) was constructed from cloning the complete N46 fragment (451 bp) reported by Laspina et al. (2008) into the selector, bar-containing plasmid pBS86 (Thompson et al., 1987), which includes two insertion sites in opposite orientation (cgf-s and cgf-as). The rice act1 promoter was considered suitable, because it drives expression in male and female reproductive tissues in rice (Zhang et al., 1991) and P. notatum (Mancini et al., 2014). Briefly, attB1 and attB2 Gateway sequences were included in the 5′ and 3′ ends of N46-specific PCR primers (Forward primer: GGGGACAAGTTTGTACAAAAAA GCAGGCTTCCCCTCCTCCCCTGTGCCGAC; Reverse primer: GGGGAC CACTTTGTACAAGAAAGCTGGGTTAAGCCTCCCCAAACGGACCAT). An amplicon was generated from a pGemTeasy N46 clone (Laspina et al., 2008), purified by using a Qiagen column and mixed with a Gateway donor vector and BP Clonase enzymes. The recombination mix was used to transform DH5α competent cells (INVITROGEN). The entry clone was then transferred into the Gateway Destination vector pBS86 using LR clonase. The insertion was validated by sequencing at the Plant Biotechnology Centre, Melbourne, VIC, Australia. The pBS86-N46 vector, together with the reporter plasmid pact1-gfbsd2 (Ochiai-Fukuda et al., 2006) carrying the eGFP gene (encoding an enhanced green fluorescent protein) were used to co-transform P. notatum plants (Q4117 genotype) with a protocol previously developed in our laboratory (Mancini et al., 2014). Transformation events were identified by PCR amplification of the transgenes from genomic DNA using the following primers: eGFPF 5′GGGGACAGCTTTCTTGTACAAAGTGGGGATGGTGAGCAAGGGCGAGGAGCT3′ (Tm: 65.4°C)/eGFPR 5′-GGGGACAACTTTGTATAAAGTTGGTTACTTGTACAGCTCGTCCATGCC-3′ (Tm: 66.1°C) (used to detect eGFP within pact1-gfbsd2) and BARXLF 5′-CCGGCGGTCTGCACCATCGT-3′ (Tm: 66°C)/BARXR 5′-ATCTCGGTGACGGGCAGGAC-3′ (Tm: 66°C) (used to detect BAR within pBS86-N46). Reactions of 25 μl final volume included 1x Taq polymerase buffer (PROMEGA), 0.2 mM forward and reverse primers, 2 mM MgCl2, 0.2 μM dNTPs, 50 ng genomic DNA and 1U Taq Polymerase (PROMEGA). Positive (with 20 ng of pact1-gfbsd2 or pBS86-N46) and negative (non-template) controls were run in parallel. Cycling consisted of 5 min at 94°C, 35 cycles of 30 s at 94°C, 1 min at the annealing temperature (Ta) and 30 s at 72°C, and a final 10 min extension at 72°C. The Ta was set at 2°C less than the lower predicted Tm. Calli transient transformation and eGFP Pact1-directed expression in reproductive tissues was followed by using an Eclipse E200 fluorescence microscopy (Nikon, Tokyo, Japan) with an standard filter cube for excitation 470/40 nm; emission 535/50 nm. Transgenic plants were grown in controlled chambers at IICAR, CONICET-Facultad de Ciencias Agrarias, Universidad Nacional de Rosario, Argentina, under a 14 h photoperiod (150–200 μE.m-2.s-1) at 26 ± 2°C.
Spikelets at anthesis were fixed in FAA (70% ethanol:formaldehyde:acetic acid 18:1:1) for 24–48 h. Ovaries were dissected and placed in 70% ethanol for at least 24 h, treated with 3% H2O2 during 2 h and dehydrated in an ethanol series (50%, 70%, 95% and twice 100%; 30 min each step). Next, they were cleared using a series of methyl salicylate/ethanol (v:v) solutions (1:1, 3:1, 5.6:1; 30 min for each step). Finally, ovaries were incubated in methyl salicylate for at least 12 h and examined using a Leica DM2500 microscope equipped with DIC optics. Pollen viability was estimated by staining with Alexander’s reagent (Alexander, 1980). Purple-stained grains were considered to be viable whereas lack of staining (i.e., pale-green/non-colored grains) indicated sterility. Observations were carried out in a Nikon Eclipse E200 microscope.
The average number of aposporous and meiotic embryo sacs per ovule was compared among four independent transgenic events and the control genotype Q4117. A modified Shapiro–Wilk test was used to test the normal distribution of the variables (Shapiro and Wilk, 1965). Due to the non-normal distribution detected, the variables were compared using the non-parametric tests of Wilcoxon (Wilcoxon, 1945) and Kruskal–Wallis (Kruskal and Wallis, 1952). Confidence intervals for observed proportions were calculated following the method described by Newcombe (1998), derived from a procedure outlined by Wilson (1927) with a correction for continuity7. Chi2 tests for homogeneity were calculated with the R software8.
An F1 population of 55 individuals, derived from a cross between sexual Q4188 as pistillate parent and apomictic Q4117 as male progenitor and characterized for reproductive modes (29 sex: 26 apo) (Stein et al., 2007) was used for linkage analyses. For N46 bulked segregant analysis (BSA), 30 μg of genomic DNA from the two parental lines and two equitable bulks of 10 sexual and 10 apomictic F1 hybrid progenies were digested with EcoRI, HindIII, and PstI. Samples were loaded in 1% agarose gel (1xTAE), electrophoresed at 40 mA and blotted onto nylon membranes (Hybond N, Amersham) using 10x SSC buffer. DNA was fixed at 80°C for 2 h. DIG probe labeling (the same N46 fragment used for ISH analyses), hybridization and detection were performed as described by Ortiz et al. (1997). For A-148-3 linkage analysis, 5 μg of genomic DNA from the two parental lines and the 55 F1s were digested using EcoRI, electrophoresed and finally blotted onto Nylon membranes. A-148-3 was converted into an RFLP probe according to Polegri et al. (2010). Probe 32P labeling, blot hybridization and exposition to X-ray films was performed according to Pupilli et al. (2001).
PCR amplifications were conducted from 50 ng of cDNA produced from total RNA extracted from leaves or flowers, with upper primer LNCU: 5′-AATTGTGCGAAATCCAATCA-3′ and lower primer LNCL: 5′-TTCACCATTACTGCCCACAA-3′. The cycling program included 1 cycle of 1 min at 94°C, 30 cycles of 1 min at 94°C, 2 min at 57°C and 2 min at 72°C and a final elongation cycle of 5 min at 72°C.
Laspina et al. (2008) reported similarity between a mRNA fragment differentially expressed in florets of sexual and apomictic P. notatum plants (N46) and a full-length cDNA transcribed from the maize gene GRMZM6G513881 (see footnote 3; NCBI Reference Sequence NM_001137220.1), which encodes a MAP3K protein. Here, we took advantage of Paspalum 454/Roche FLX + floral transcriptomes recently developed in our laboratory to recover the N46 full cDNA sequences from apomictic (Q4117) and sexual (C4-4x) genotypes and carry out molecular phylogenetic analysis. In the apomictic floral transcriptome library, we detected one isogroup (apoisogroup 00379) represented by four homologous isotigs, namely apoisotig 03083 (GFMI02003139.1), apoisotig 03084 (GFMI02003140.1), apoisotig 03085 (GFMI02003141.1) and apoisotig 03086 (GFMI02003142.1). In the sexual floral transcriptome library, we also found one isogroup (sexisogroup 02509), but it contained a single isotig (sexisotig 08547; GFNR01008571.1). ClustalW nucleotide (nt) sequence alignments revealed that apoisotigs 03085 and 03086 and sexisotig 08547 were highly similar, differing only by a few polymorphisms (SNPs and INDELs) (Supplementary Figure S1). Apoisotigs 03083 and 03084, were respectively, identical to apoisotigs 03085 and 03086, except for a 76-nt insertion with the canonical donor-receptor sites of GU-AG-type introns, which corresponds to a partially conserved intron in maize GRMZM6G513881. These results suggest that P. notatum N46-like floral sequences are genetic and splice variants of a single locus with at least two different alleles in the apomictic genotype (apoisotigs 03085/03083 and 03086/03084) and a third allele detected in the sexual genotype (sexisotig 08547). The intron-like insertion (located between positions 1857–1934 and 1833–1908 in apoisotigs 03083 and 03084, respectively) modifies the reading frame to produce a protein with a variable C-terminal end (Figure 1 and Supplementary Figure S2). Genomic amplification with flanking primers showed that the intron-like region is present in both apomictic and sexual plants (Supplementary Figure S3). Although the non-processed form had been sequenced only from the apomictic samples in the 454/Roche FLX + transcriptome libraries, semi-quantitative RT-PCR experiments showed that both variants (processed and non-processed) were represented in flowers of apomictic and sexual plants (Supplementary Figure S3). Moreover, qPCR of cDNAs originated from a mix of flowers at different developmental stages (from premeiosis to anthesis) with primers located inside the intron revealed no significant differential representation between reproductive modes (not shown). BLASTX searches using N46 full-length sequences as queries identified homology to MAP3K genes belonging to the YODA family (best annotated match: Oryza sativa mitogen-activated protein kinase kinase kinase YODA isoform X1 XP_015617106.1; 79% identity; E-val: 0.0; query coverage: 74%; alignment length: 1,688). A phylogenetic tree inferred using 22 homologous protein sequences from different species showed that all Paspalum sequences grouped into a single cluster within the Poaceae clade, supporting the conclusion that they are allelic isoforms or, alternatively, expressed from gene copies that diverged recently (Supplementary Figure S4). Finally, the whole QGJ sequence was amplified by using primers located at the borders (see “Materials and Methods”), to confirm its existence without the need of computational assembly. Based on its identity as a member of the YODA family, we named the N46 locus QUI-GON JINN (QGJ), after another character of the Star Wars saga.
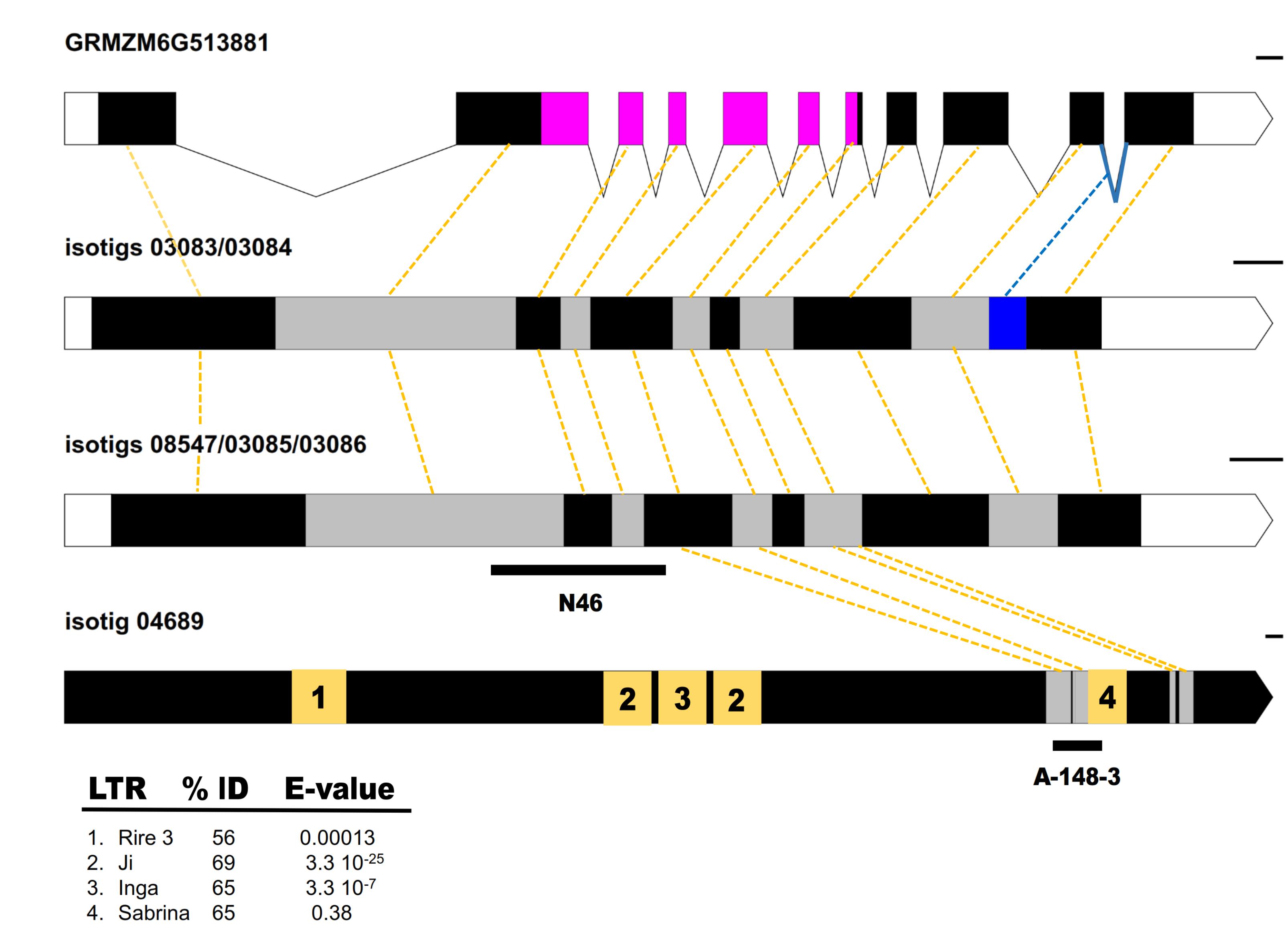
FIGURE 1. Structural and functional annotations of Paspalum QUI-GON JINN and PN_LNC_QGJ sequences. (A) Annotated genomic sequence of the QGJ maize ortholog GRMZM6G513881. White and black boxes denote UTRs and exons, respectively. Lines denote introns. The STK domain is indicated in magenta. The intron involved in the Paspalum splice variants in indicated in blue. (B) Transcript splice variant corresponding to isotigs 03083/03084, with the last intron represented in the sequence (blue). Gray and black boxes denote different subsequent exons. White boxes denote 5′ and 3′ UTRs. (C) Transcript splice variant corresponding to isotigs 08547/03085/03086. The position of the original N46 fragment (Laspina et al., 2008) was indicated below. Gray and black boxes denote different subsequent exons. White boxes denote 5′ and 3′ UTRs. (D) PN_LNC_QGJ long non-coding RNA. Sequences similar to repetitive elements were numbered on the yellow boxes according to (E) and unknown sequences are indicated in black. Sequences similar to QGJ are indicated in gray. The position of the original A-148-3 fragment (Polegri et al., 2010) was indicated below. (E) Summary table for PN_LNC_QGJ BLAST analysis (Gramineae Repeats Database). Yellow dashed lines join corresponding exons. Bars at the right indicate the drawing scale: 100 bases.
The QGJ expression was quantified in spikelets of sexual and apomictic P. notatum genotypes at different developmental stages (1: premeiosis; 2: late premeiosis/meiosis; 3: post-meiosis) by using real-time PCR. The primer pair used for amplification was complementary to all known QGJ variants (see “Materials and Methods,” qPCR experiments). At stage 1 (premeiosis), QGJ transcripts were equally represented in both P. notatum reproductive types (Figure 2A). Later, during late premeiosis/meiosis, the expression in the sexual plant was significantly higher (Figure 2A). In contrast, an opposite pattern was observed at post-meiosis (Figure 2A). Besides, we took advantage of the EMBRAPA (Brasilia, Brazil) collection of Brachiaria brizantha plants, another well-characterized aposporous pseudogamous system (Pagliarini et al., 2012), to validate the results. Brachiaria brizantha (syn. Urochloa brizantha) is, like P. notatum, a rhizomatous perennial grass (Poaceae), which reproduces through aposporous pseudogamous apomixis. As P. notatum plants, aposporous Brachiaria genotypes form supernumerary non-reduced embryo sacs lacking antipodals from nucellar somatic cells surrounding the MMC. The B. brizantha genome also includes a single ACR lacking recombination, which may be evolutionary related to the Paspalum one, since it is located in a chromosomal background displaying partial synteny to rice chromosome 2 (Pessino et al., 1997, 1998). In Brachiaria spikelets a similar expression profile was detected, but at premeiosis overexpression was detected in the apomictic genotype (Figure 2B). However, a more detailed quantification of QGJ in RNA samples extracted in Brachiaria isolated ovaries revealed overexpression in apomictic plants at late premeiosis/meiosis (Figure 2C), which suggest the occurrence of contrasting representation patterns in different tissues.
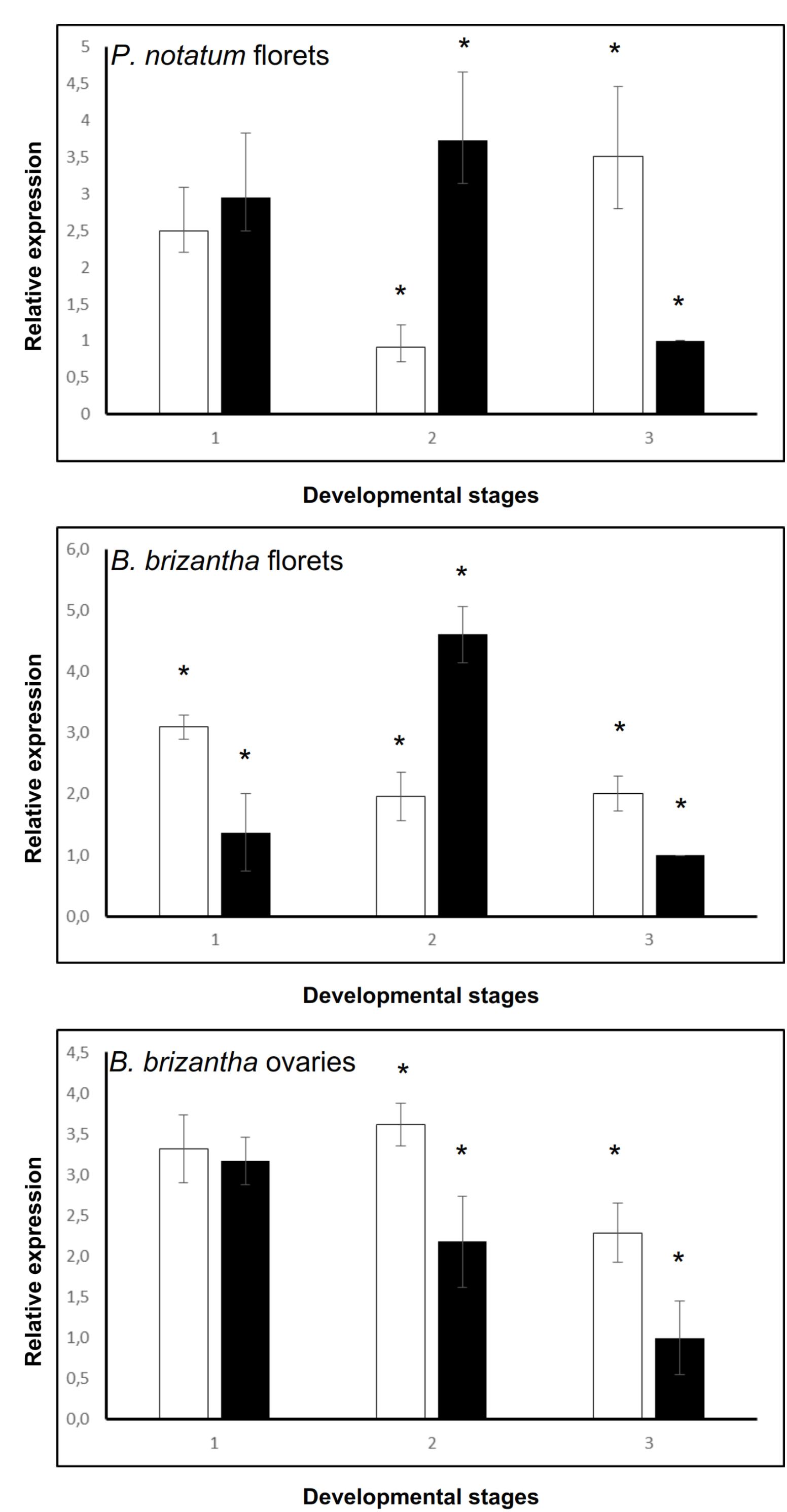
FIGURE 2. Relative quantitative expression of QUI-GON JINN across reproductive stages. (A) P. notatum florets. (B) B. brizantha florets. (C) B. brizantha ovaries. White bars: apomictic plant (genotype Q4117). Black bars: sexual plant (genotype C4-4x). 1: premeiosis; 2: late premeiosis/meiosis; 3: postmeiosis. Control: sexual plant, postmeiosis stage (relative expression = 1). Samples displaying statistically significant differential expression between apomictic and sexual plants were marked with an asterisk at the top.
The site of expression was then examined through in situ hybridization in developing ovaries and anthers of P. notatum, using the original N46 clone to produce the sense and antisense probes (Figure 3). N46 is complementary to a region conserved among all QGJ variants, so the experiment has no potential to differentiate them. In premeiotic flowers of sexual plants, the antisense probe showed weak signal in the ovule nucellus and integuments and moderate to strong signal in the MMC, the anther tapetum and pollen mother cells (Figure 3A). Meanwhile, in apomictic plants the same probe revealed strong signal in the ovule nucellus and MMCs, and a diminished signal in anthers (Figure 3B). The sense probe showed weal signal in ovules of both sexual and apomictic plants (Figures 3C,D). We validated the observed in situ differential expression of QGJ genes in B. brizantha aposporous ovaries, by using the same N46 probe (Figure 4). In the Brachiaria experiments, thinner microtome slice cuts, together with a microscope with a higher resolving power were used (see “Materials and Methods”), allowing a more accurate detection of the hybridization pattern. In premeiotic ovules of sexual plants, a weak to moderate signal was detected in the ovule nucellus, while a moderate to strong signal appeared in the MMCs (Figure 4A). After meiosis I, the signal became mainly restricted to the micropylar cell of female dyads, yet some signal could also be observed in the nucellus (Figure 4B and Supplementary Figure S5). In tetrads, a strong signal was detected in the non-functional (micropylar) megaspores, while the functional one (located close to the chalazal end of the ovule) showed low signal (Figure 4C). In premeiotic ovules of apomictic plants, a weak to moderate signal was detected in the ovule nucellus, while a moderate to strong signal appeared in the MMCs (Figure 4D). During aposporous initials (AI) differentiation, moderate to strong signal was detected in the ovule nucellus, except for the cell layer surrounding the MMC (the AI onset site) (Figures 4E,F). Note that at this stage the MMC has enlarged and formed a meiocyte instead of entering meiosis I (meiosis frequently fails in obligate aposporous plants). Apospory initials originate from this proximal cell layer lacking signal (Figure 4F). While strong signal was detected in pollen mother cells of the sexual plant (Figure 4G), a moderate to weak signal was observed in pollen mother cells of the aposporous genotype (Figure 4H). Finally, hybridizations using sense probes detected weak signals in the nucellus of both plant types (Figures 4I,J). Our results indicate that, in sexual plants, QGJ is weakly expressed in nucellar tissues during meiosis. At this stage, its expression is restricted to the non-functional (micropylar) megaspores, which are adjacent to the functional (chalazal) megaspore. In contrast, and in agreement with our previous results from Paspalum, a strong expression of QGJ was observed in nucellar cells of apomictic plants. However, the proximal layer of cells originating the AI lacked signal, suggesting that QGJ is expressed in cells adjacent to the functional germ cells, i.e., non-functional reduced megaspores or nucellar cells located aside the non-reduced megaspores in sexual and apomictic plants, respectively.
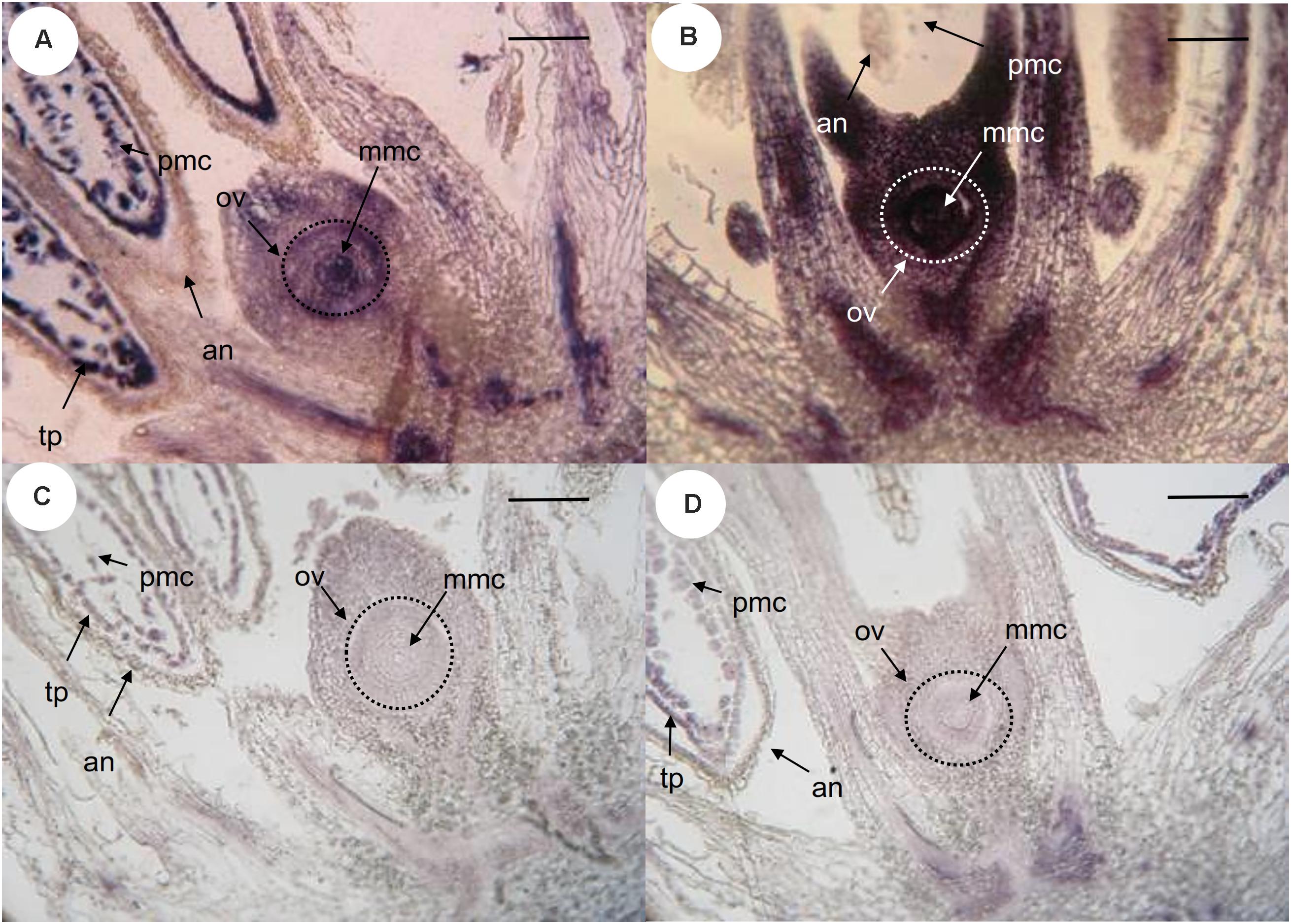
FIGURE 3. In situ hybridization of QGJ on Paspalum notatum reproductive organs at late premeiosis stage. Homologous in situ hybridization was carried out with the original N46 fragment. (A,C) Ovule and anthers of sexual plant (C4-4x), late premeiotic stage. (B,D) Ovule and anthers of apomictic plant (Q4117), late premeiotic stage. (A,B) Antisense probe. (C,D) Sense probe. Bars: 20 μm. References: an, anther; mmc, megaspore mother cell; ov, ovule; tp, tapetum; pmc, pollen mother cells.
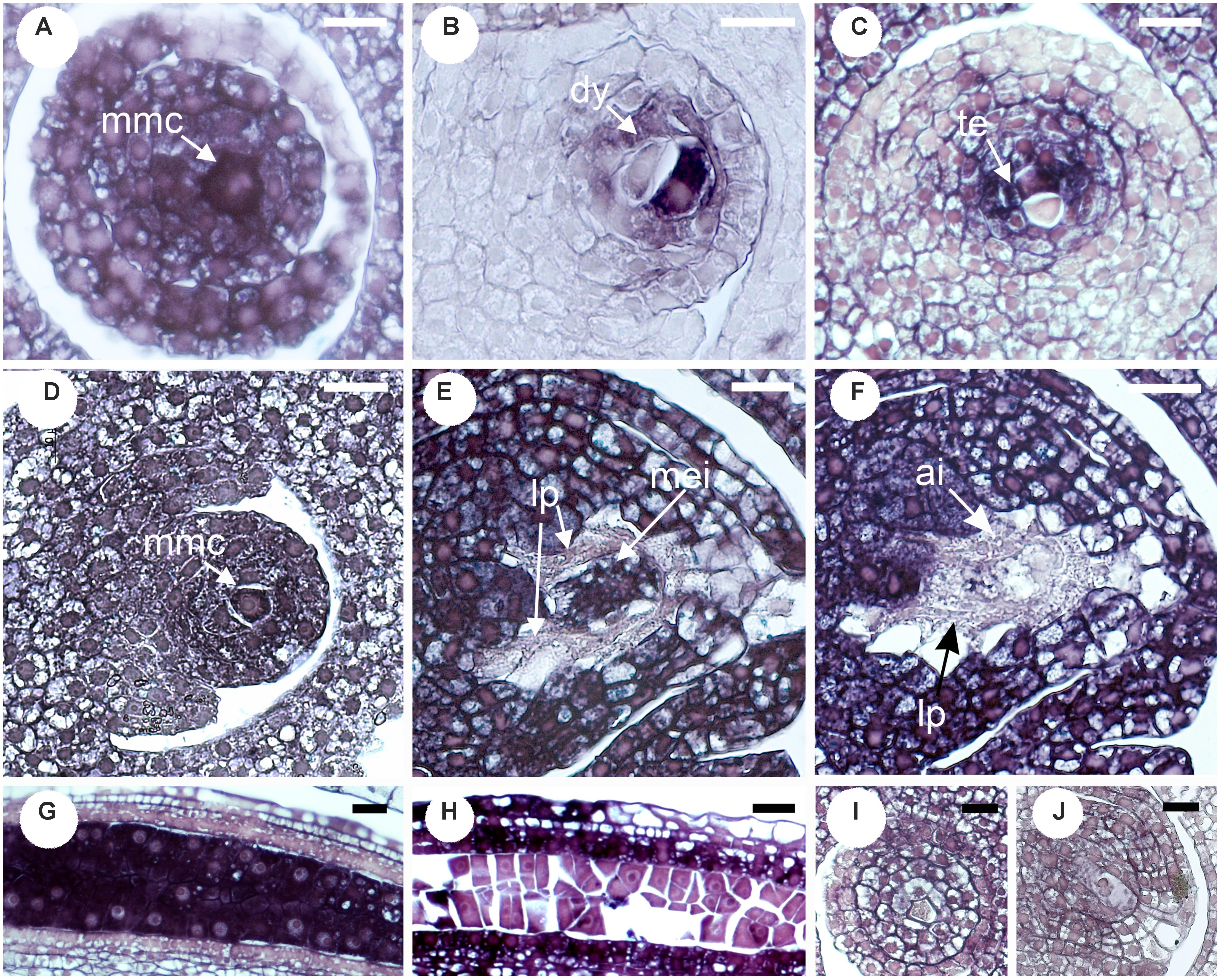
FIGURE 4. Validation of QGJ in situ expression in reproductive organs of aposporous Brachiaria brizantha. Heterologous in situ hybridization was carried out with the original N46 fragment. Ovules (A–C,I) and anther (G) of sexual plant B105; ovules (D–F,J) and anther (H) of apomictic plant B30. (A–H) Antisense probe. (I,J) Sense probe. (A) Megaspore mother cell (mmc). (B) Meiosis (dyad stage). (C) Meiosis (tetrad stage showing two megaspores). (D) Archesporial cell stage. (E,F) Meiosis (mmc entering meiosis = meiocyte) and AI formation stage. (G,H) Pollen mother cell stage. (I,J) Meiosis stage. Bars: 10 μm. References: mmc, megaspore mother cell; dy, dyad; te, tetrads; lp, layer of proximal cells surrounding the MMC; ai, apospory initial.
Next, we decided to investigate if a diminished expression of QGJ in an apomictic background gives rise to altered reproductive phenotypes. Firstly, a plant transformation vector including an N46 hairpin (pBS86-N46) was constructed by cloning the complete N46 fragment in sense and antisense orientation within plasmid pBS86 (see “Materials and Methods”). Then, QGJ RNAi lines were obtained by Q4117 biolistic co-transformation with plasmids pact1-gfbsd2 (which expresses an enhanced green fluorescent protein gene eGFP under the rice ACT1 promoter) and pBS86-N46 (see “Materials and Methods”). From 41 positive transgenic events (for pact1-gfbsd2, pBS86-N46 or both), two groups of lines were selected, which had, respectively, been transformed with: (1) the reporter plasmid pact1-gfbsd2; and (2) the RNAi plasmid pBS86-N46 and the reporter plasmid pact1-gfbsd2 (Figures 5A–H,K,L). Plants belonging to the first group were classified as transformation control lines (since they allow evaluation of reproductive phenotypes in plants subjected to in vitro culture and transformation procedures), while those corresponding to the second group were labeled as RNAi lines. These plants were classified as T0, since they were regenerated from bombarded calli. Prior to the molecular and cytoembryological analysis, two rounds of small rhizomes subculture were conducted (P. notatum is perennial and reproduces by forming rhizomes). Only four plants flowered in the isolated GMO chamber under controlled conditions (transformation control lines #TC1/#TC2 and RNAi lines #RNAi1/#RNAi2), together with a wild type control. Fluorescence analysis in transgenic lines carrying the pact1-gfbsd2 vector (both RNAi and TC) confirmed that the rice Act1 promoter drives expression in male and female reproductive tissues of P. notatum (Figures 5I,J).

FIGURE 5. Generation of RNAi lines from apomictic genotype (Q4117). (A,B) Schematic representation of transformation vectors pact1-gfbsd2 and pBS86-N46. (C) BIOMIX biolistic device used for plant transformation as disposed previous to bombardment. (D) Calli in osmotic medium. (E) GFP expressing cells (marked by white arrows) indicating transient transformation of calli. (F) Regenerating plants subjected to selective agent glufosinate-ammonium 1 mg/L. (G,H) Rusticated plant in the growth chamber. (I,J) Ovary (I) and pollen grains (J) expressing eGFP. (K) PCR amplification of eGFP in T0 plants. (L) PCR amplification of BAR in T0 plants.
Quantitative RT-PCR analyses revealed that the QGJ expression was significantly attenuated in floral tissues of both RNAi lines compared to the apomictic wild type ecotype (relative expression levels ranging from 0.478 to 0.656; Supplementary Figure S6A). Lower expression levels were detected in leaves in comparison to flowers of Q4117 and no significant reduction in expression was detected in leaves of RNAi lines (Supplementary Figure S6B). Rates of viable pollen were slightly reduced in the two RNAi lines (#RNAi1 and #RNAi2) with respect to both the control lines (#TC1 and #TC2) and the wild type (Q4117) (Table 1). However, even if statistically significant, this minor alteration might not imply physiological consequences. Female reproductive development was examined by observing cleared ovules at anthesis and determining the type and number of ES per ovule. Relatively high proportions of aborted ovaries, i.e., containing no ES, were detected for lines #RNAi1 and #RNAi2 compared to both wild type and control plants (20–30% vs. <10%; Table 2). Defects in both initiation and completion of AES formation affected female reproductive development in RNAi lines (Table 2 and Figure 6). A lower number of AES per ovule was detected for both #RNAi1 and #RNAi2 (Table 2) and most of them exhibited weak/abortive phenotypes such as small size, ragged borders, and no detectable polar nuclei (Figure 6). Conversely, in control lines (#TC1 and #TC2), the average number of AES per ovule was similar to that of Q4117 (Table 2). Finally, the proportion of ovules containing meiotic ES (MES) showed no statistical difference among Q4117, control lines and RNAi lines (Table 2). We concluded that the significant reduction of QGJ transcripts after introducing a QGJ hairpin construction in the apomictic genotype Q4117 impaired the formation of AESs. Our data suggest that expression of QGJ in nucellar cells is necessary for aposporous development in P. notatum.
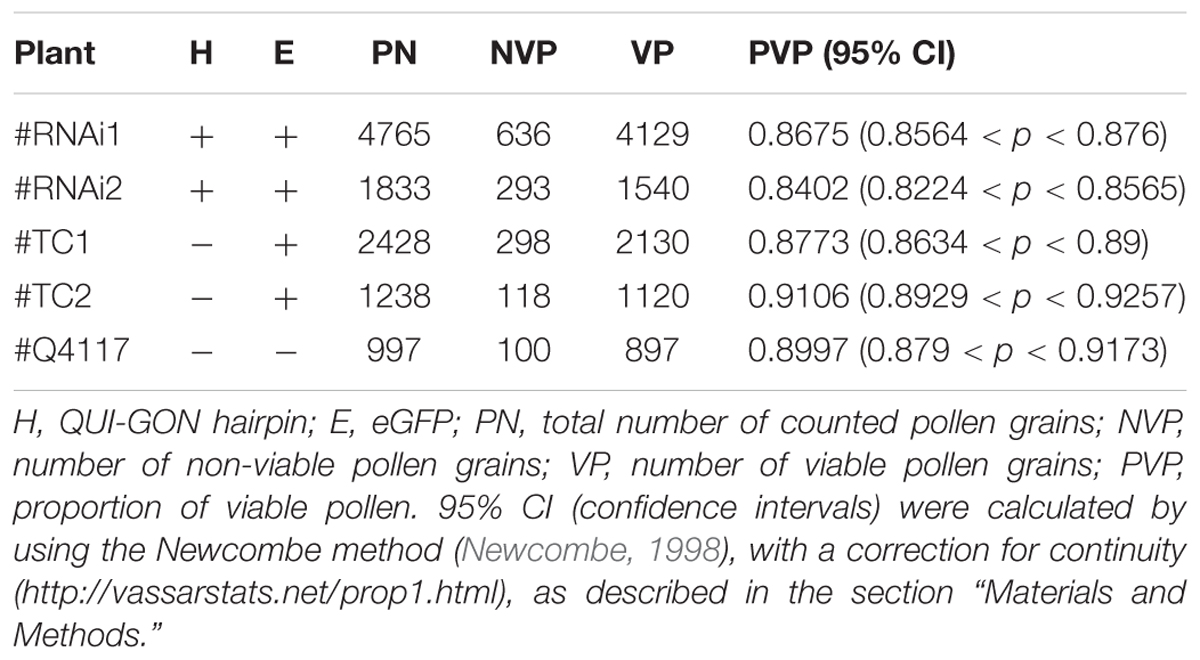
TABLE 1. Pollen viability of transformed and control plants.
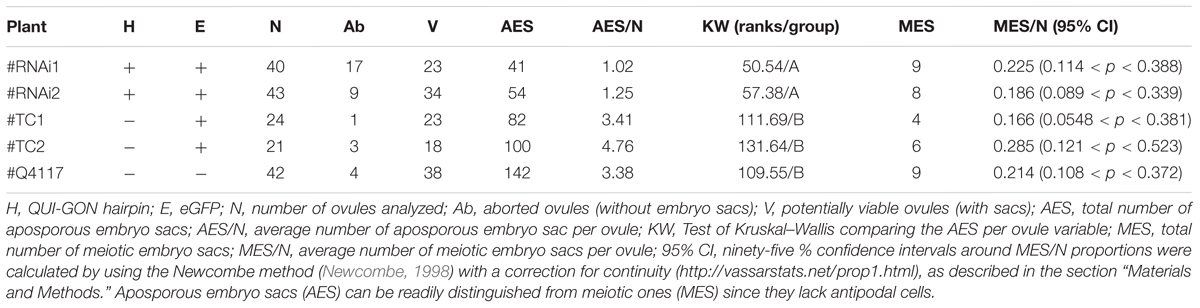
TABLE 2. Cytoembryological analysis of transformed and control plants.
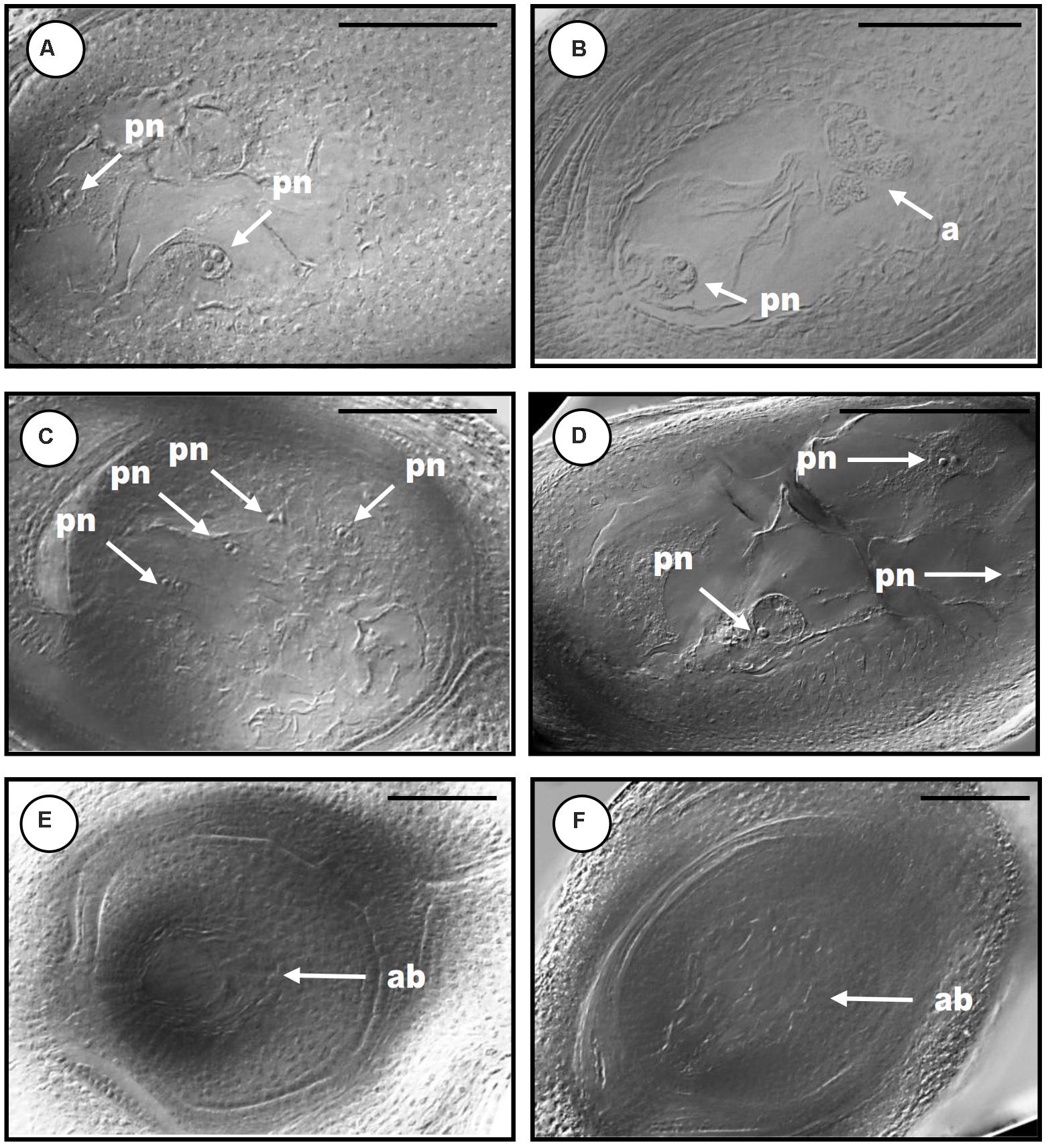
FIGURE 6. Cytoembryological analysis of the reproductive behavior in QGJ RNAi lines. Mature ovules of wild type genotype Q4117 (A,B), transformation control lines #TC1 (C) and #TC2 (D) and QGJ RNAi lines #RNAi1 (E) and #RNAi2 (F). Aposporous embryo sacs (A,C,D). Meiotic embryo sac (with a proliferating mass of antipodals) (B). Aborted embryo sacs (E,F). References: pn, polar nuclei; a, antipodals; ab, aborted. Bars: 100 μm.
Possible co-segregation of the QGJ locus with apospory was examined in P. notatum by using bulked-segregant analysis (Figure 7). The complete N46 original fragment was hybridized onto genomic DNA samples originated from two parental plants (Q4188, sexual female parent and Q4117, apomictic pollen donor) and genomic DNA bulks made from 10 sexual and 10 apomictic F1 plants derived from the Q4188 × Q4117 cross. Segregation in F1 is expected due to ACR hemizygosity and the heterozygous nature of a high number of parental genomic loci. Although genomic DNA digestion with three different restriction enzymes (EcoRI, HindIII, and PstI) produced several polymorphic bands between parental plants, none of them resulted polymorphic between F1 bulks. The bulked results show the sexual polymorphisms are not specific to sexual F1s. However, the apomictic band present in the parental PstI digest disappeared in the apomictic F1s. Since PstI is sensitive to certain contexts (CpNpG sites), this observation might suggest a methylation change occurring during hybridization. In silico mapping onto the Gramene website revealed that the putative ortholog to QGJ is located in chromosome 11 (Os11g0207200, E-value 0.0), in a genomic region showing no synteny with the ACR of P. notatum. However, other high score hits (Os02g0555900, Os02t0666300-01, Os12g0577700, E-values 5.7E-86, 3E-58, and 7.6E-15) are protein kinase genes located in a rice genome region (chromosome 2 long arm, positions: 21,002,439–21,008,209, 27,048,920–27,061,955 and chromosome 12 long arm, positions: 23,885,845–23,888,835, respectively) syntenic to the P. notatum ACR. Our results showed no evidence of a genetic link between QUI-GON JINN and the ACR, but suggested that sequences showing significant similarity to this gene might be located within the genomic region controlling apospory.
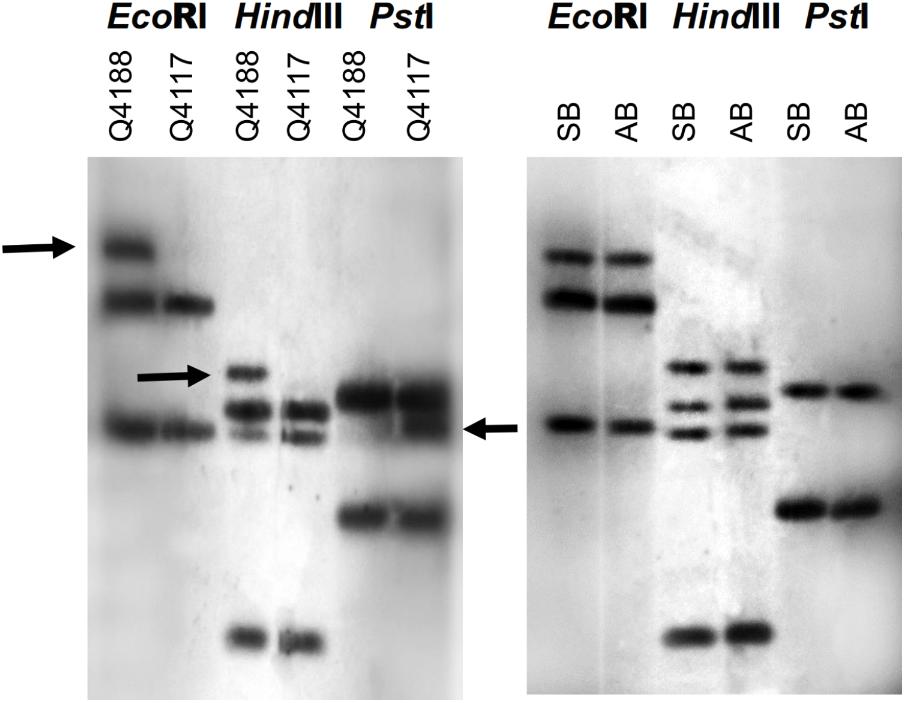
FIGURE 7. Bulked segregant analysis of the QGJ locus on the P. notatum genome. Three different restriction enzymes (EcoRI, HindIII, and PstI) were used to digest the genomic DNA samples. The complete original N46 fragment was used as a probe. Left panel: Polymorphisms (marked with arrows) were detected between Q4188 (sexual genotype, mapping population female parent) and Q4117 (apomictic genotype, mapping population male parent). Right panel: Linkage analysis was performed by bulking genomic DNA extracted from 10 sexual (SB) and 10 apomictic (AB) F1 individuals obtained from crossing Q4188 (female) and Q4117 (male).
In a previous work, Polegri et al. (2010) reported full linkage between the Paspalum simplex transcriptome fragment A-148-3 and apospory, and determined that this candidate was also homologous to At1g53570. A search for the complete sequence of A-148-3 in the Paspalum Roche 454 floral libraries revealed that it was not a PN_QGJ ortholog, since it showed the best hit of similarity with apoisotig 04689 (GFMI02004742.1) (query cover 100%; E-value: 6 e-143, identities 90%; position on apoisotig 04689: 5588–5955). Interestingly, apoisotig 04689 is a 6835-nt sequence with no coding potential and specific to the apomictic P. notatum floral transcriptome libraries (reads in the apomictic library: 133; reads in the sexual library: 0). A search in the GreeNC database revealed similarity with six predicted plant lncRNAs at E-values ≤ 1 e-10 (best match: Zmays_GRMZM2G024551_T01; E-value: 1.00411e-27; alignment length: 226 nt, involving the segment flanked by positions 4304–4530 within apoisotig 04689; positive: 177). Given its partial similarity with QGJ, its lack of protein-coding potential, its similarity to sequences included in the GreeNC database and its detected expression in the apomictic floral transcriptome, we inferred that isotig 04689 is a long non-coding RNA (lncRNA) related to QGJ, and was renamed accordingly as PN_LNC_QGJ (after P. notatum long non-coding QGJ). The complete QGJ functional sequence (apoisotig 03085, 2377 nt) has similarity to the PN_LNC_QGJ transcript in positions ranging 1033–1326/1352–1475 (matching positions 5493–5800/6255–6387 in the PN_LNC_QGJ sequence) (Figure 1). Meanwhile, the original N46 fragment spanned positions 733–1174 in the QGJ sequence (apoisotig 03085) (Figure 1).
Mapping of the A-148-3 original transcript in a P. notatum population (55 F1 plants) revealed one polymorphic band strictly cosegregating with apomixis (Figure 8). Another band showed partial linkage, confirming association of the sequence with proximal regions. Besides, the two additional monomorphic/non-segregating bands were detected, which could be related with the same locus or, alternatively, with other genomic regions located elsewhere (Figure 8). Furthermore, reverse-transcribed PCR experiments with PN_LNC_QGJ specific primers conducted in several apomictic and sexual P. notatum individuals showed that PN_LNC_QGJ is expressed only in apomictic plants (Supplementary Figure S7).
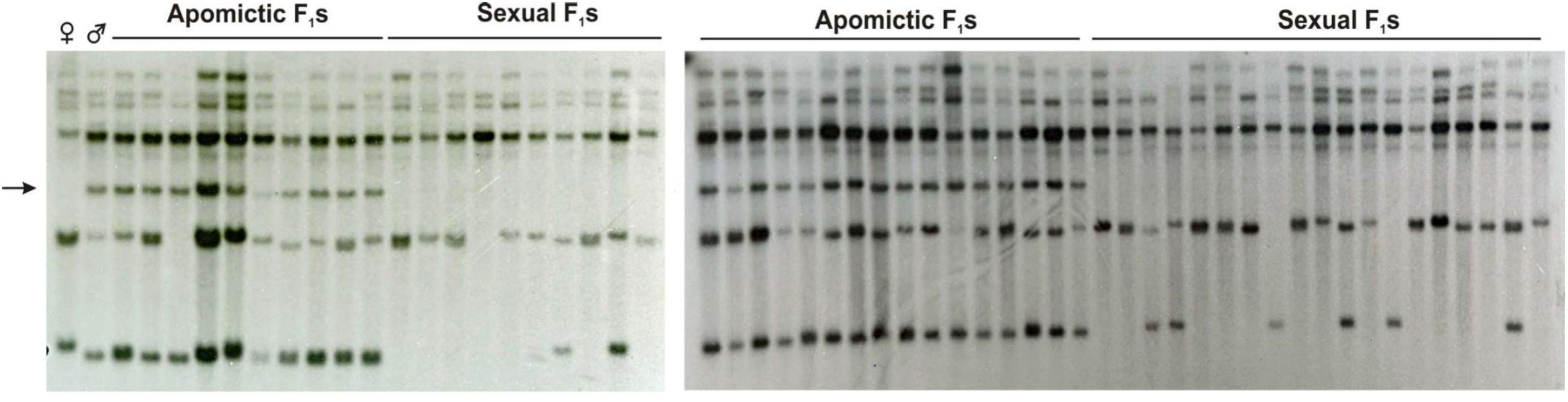
FIGURE 8. A-148-3 mapping onto the P. notatum genome by RFLP analysis. Hybridizing banding pattern of genomic DNA EcoRI digests of a P. notatum segregating population, composed of a female sexual parent (Q4188,♀), a male aposporous parent (Q4177, ♂) together with 29 sexual and 26 aposporous F1 plants. The arrow indicates a band inherited from the aposporous parent, which strictly co-segregates with apospory.
Based on these observations, we concluded that original transcript fragment A-148-3 is part of a long non-coding RNA (namely PN_LNC_QGJ) differentially expressed in apomictic and sexual plants. Besides, a sequence similar to the A-148-3 probe is located in the P. notatum ACR. However, further experiments should be conducted to determine if the copy of the gene located at the ACR is producing the differentially expressed lncRNA (considering the existence of monomorphic bands, which could represent copies located in other parts of the genome). Moreover, the existence of a functional link between the altered QGJ expression and a possible regulator activity of PN_LNC_QGJ has yet to be investigated through functional analysis. The alignments between the Z. mays reference genomic sequence GRMZM6G513881 and the sequences of all PN_QGJ and PN_LNC_QGJ isotigs are displayed in Supplementary Figure S8.
The extracellular signal-regulated kinase 1/2 (ERK1/2) cascade is a central signaling pathway that modulates a wide variety of cellular processes, including proliferation, differentiation, survival, apoptosis, and stress response (Wortzel and Seger, 2011). The intracellular communication between membrane receptors and their nuclear or cytoplasmic targets upon stimulation is mediated by a limited number of signaling pathways, including a group of mitogen-activated protein kinase (MAPK) cascades (Wortzel and Seger, 2011). MAPK signal transduction cascades consist of three sequentially activated kinases. Upstream signals activate MAPK kinase kinases (MAPKKKs), which in turn phosphorylate MAPK kinases (MKKs); subsequently, MKKs activate specific MAPKs. The downstream targets of MAPKs can be either transcription factors or cytoskeletal proteins. Phosphorylation and activation of a MAPK can lead to changes in its subcellular localization and its activity on transcriptional effectors, thereby reprogramming gene expression (Fiil et al., 2009). In particular, the Arabidopsis genome encodes 60 putative MAP3Ks (including the MEKK, RAF, and ZIK subfamilies), 10 MAPKKs, and 20 MAPKs, involved in a plethora of different responses to specific ligands (Ichimura et al., 2002). Here, we characterized the expression and function of QUI-GON JINN (QGJ), the putative ortholog to Arabidopsis At1g53570/AtMEKK3, in reproductive organs of sexual and apomictic P. notatum plants. AtMEKK3 belongs to the MEKK subfamily related to budding yeast Ste11p (Lukowitz et al., 2004). It comprises 12 members (Ichimura et al., 2002), including critical regulators of plant cell division and differentiation during reproduction, i.e., YODA/AtMAPKKK4 (early embryogenesis) (Lukowitz et al., 2004), AtMEKK20 (male gamete differentiation) (Borg et al., 2011) and ScFRK/AtMEKK19/20 (female gametophyte development) (Daigle and Matton, 2015).
Our results suggests that QGJ plays a role in promoting the acquisition of a gametophytic cell fate by AIs, a critical step in the establishment of the aposporous pathway, or alternatively affects the development of the embryo sacs. The fact that QGJ is not expressed in the cell layer originating AIs, but in the adjacent ones, suggests that a non-cell-autonomous signaling mechanism might be operating. Such mechanisms are commonly used for intercellular communication during plant development (Van Norman et al., 2011). In contrast, pollen viability and meiotic embryo sac formation seem poorly affected in RNAi plants, showing that the QGJ partial silencing is not influencing male/female meiosis in an obligate apomictic background. However, since the meiotic pathway is per se diminished in obligate apomictic plants (the Q4117 apomictic genotype naturally shows a rate of meiotic embryo sac development of around 3–4%) (Ortiz et al., 1997), it is difficult to evaluate the QGJ post-transcriptional attenuation consequences on the sexual developmental pathway when using the genetic background transformed here. Such analyses should be conducted in a mutant/transformant line derived from a sexual plant, an experiment that we plan to complete in the near future. Moreover, the slight decrease in pollen viability detected in the Paspalum transgenic lines might reflect a mild response to the introduction of the hairpin in a tissue where the gene is naturally down-regulated with respect to sexual plants, as revealed by in situ hybridization. Podio et al. (2012) had analyzed anaphase I configurations and pollen viability in aposporous and sexual tetraploid cytotypes of P. notatum and found reduced pollen viability in the aposporous genotypes, including Q4117. A reduced activity of QGJ in pollen mother cells of aposporous plants (Figures 3, 4) could also explain the diminished expression detected in apo plants in comparative qPCR experiments conducted on florets (Figure 2), since anthers represent a high proportion of the floret tissue at this stage. Based on all these observations, we hypothesized that, similarly to YODA (Lukowitz et al., 2004), QGJ mediates a signaling pathway acting as a key regulator to define cell lineage during plant reproduction. While YODA is required for normal development of the zygote and the cells of the basal lineage originating the suspensor, QGJ might play a role during the sporophytic-to-gametophytic transition phase. However, although our results indicate that QGJ activity is essential to non-reduced embryo sacs formation, overexpression experiments under ovule specific promoters will be necessary to assess if expression in the nucellus of sexual plants is fully responsible for apomictic development. An alternative hypothesis is that this candidate is involved later, after the fate decision has been made, in either sexual or aposporous embryo formation.
Polegri et al. (2010) reported the isolation of A-148-3, a P. simplex transcript homologous to the predicted Arabidopsis QGJ ortholog (At1g53570), showing constitutive expression in apomictic genotypes during all reproductive developmental stages and linkage to the P. simplex ACR. However, full sequence analysis revealed that A-148-3 is not a QGJ ortholog, but an lncRNA with partial similarity to QGJ. Genetic linkage analyses in a Q4188 (sexual) × Q4117 (apomictic) F1 population confirmed that a sequence showing similarity with A-148-3 is located within the P. notatum ACR. On the contrary, we found no evidence that N46 and the ACR are genetically linked. Note that the A-148.3 probe has some potential to hybridize QGJ (the original 354 bp-long A-148-3 sequence includes a 147-nt insertion with 78 % similarity to P. notatum QGJ). Moreover, from the 451 nt covered by the original fragment N46, a 112-nucleotide segment keeps partial similarity (72%) with PN_LNC_QGJ. Although these similarities are limited, involve only a portion of the probes (40% of A-148.3 and 25% of N46), and the experimental conditions were strict enough to ensure specific detection, the possibility of some cross hybridization cannot be fully discarded, especially for A-148.3. However, the detection of contrasting genomic hybridization patterns when using N46 and A-148.3 as alternative probes (Figures 7, 8), suggests that each of them has capacity for specific detection. The ACR is a genomic region specific to apomictic genotypes, which is highly heterochromatic and harbors almost intact exonic sequences interlaced within highly repetitive sequences (Podio et al., 2014a). PN_LNC_QGJ is expressed in floral tissues of aposporous plants only, and it includes large, non-coding stretches similar to retrotransposons and two short exonic QGJ regions (439 nt in total out of 6835). Long non-coding RNAs have recently emerged as critical regulators of gene expression in many eucaryotes, including plants (Ariel et al., 2015; Chekanova, 2015; Liu et al., 2015). Therefore, considering the sequence relationship between QGJ and PN_LNC_QGJ, it is tempting to speculate that PN_LNC-QGJ could mediate QGJ modulation in reproductive tissues of Paspalum apomicts. Among many putative mechanisms, our results point out at least two of them: (1) a change in splicing leading to the formation of variants; and (2) the induction of nucellar expression via miRNA hijacking. However, no claim of a functional relationship between QGJ and PN_LNC_QGJ can be currently made, since it is not supported by functional analysis data. Moreover, expression of PN_LNC_QGJ from the ACR should be further confirmed. Shortly, we will focus in determining if the miss-expression of QGJ is caused by a transcriptional regulatory event or is alternatively influenced by PN_LNC-QGJ, and to determine if PN_LNC-QGJ is expressed from the ACR. Interestingly, non-coding transcripts carrying exonic sequences were proposed to regulate PN_SERK and PS_ORC3, two genes putatively involved in apomixis in P. notatum and P. simplex, respectively (Podio et al., 2014b; Siena et al., 2016).
In the past decade, numerous candidate genes for apomixis were identified (Hand and Koltunow, 2014; Ronceret and Vielle-Calzada, 2015) but how the underlying networks integrate into sexual reproduction and alter expression patterns remain largely unknown. Our work posits that a MAP3K signaling pathway of an ERK1/2 cascade is pivotal to aposporous embryo sac differentiation. However, the rest of the members of the ERK cascade and their interactions with this kinase remain unknown. Interestingly, besides N46 (QGJ), Laspina et al. (2008) reported the differential expression of several other genes involved in ERK cascades in comparisons between sexual and apomictic plants: an LRR family protein (N79), a GPI anchored protein (N20), phosphatidylinositol 4K (N23), a Ser/Thr phosphatase (N102), the PRIP-interacting protein (N69) and a kinesin (N114). From them, only N20 and N69 have been further characterized (Felitti et al., 2011; Siena et al., 2014). N20 (later renamed N20GAP-1) is ortholog to genes At4g26466 (LORELEI, encoding a GPI-anchored protein) and/or At5g56170 (LORELEI-like), shows partial cosegregation with apospory and is increasingly overexpressed in apomictic plants from premeiosis to antesis (Felitti et al., 2011). N69 is ortholog to gene AT1G45231 (TGS1, encoding a trimethylguanosine synthase which has a dual role in splicing and transcription), and, contrarily, is increasingly overexpressed in sexual plants from premeiosis to antesis (Siena et al., 2014). Moreover, evidence was shown that TGS1 Ser298 phosphorylation is promoted by an ERK cascade to activate transcriptional activity at some promoters (Kapadia et al., 2013). The availability of Paspalum RNAi lines for N20 (LORELEI), N69 (TGS1) and N46 (QGJ MAP3K, reported here) would allow to investigate in detail a possible biological link among these molecules. Though functional approaches are challenging in polyploid, highly heterozygous apomictic species like P. notatum, the development of reference genomes, transformation protocols and advanced microscopy tools will likely accelerate the discovery of the central mechanisms underlying the switch from sexuality to apomixis.
MM: transformation experiments and the genotypic/phenotypic analysis. HP: transformation experiments supervision. CC and CA: RNA extractions, real time experiments, and bioinformatic analysis. LS: developmental stage classification and RNA extraction. FP: PN_LNC_QGJ mapping. DdAD, VdCC, and MP: Brachiaria in situ hybridization. MP: PN_QGJ genomic hybridization. JS and AG: Paspalum in situ hybridization. SF: transformation vector construction. JO: phylogenetic analysis and experimental design. OL: experimental design, sequence analysis, and manuscript writing. SP: experimental design, in silico analysis, qPCR analysis, and manuscript writing.
Thanks are due to the European Union’s Horizon 2020 Research and Innovation Programme under the Marie Skłodowska-Curie Grant Agreement No. 645674; Agencia Nacional de Promoción Científica y Tecnológica (ANPCyT), Argentina, Projects PICT 2007-00476, PICT 2011-1269, PICT-2014-1080; Consejo Nacional de Investigaciones Científicas y Técnicas (CONICET), Argentina, Project: PIP 11220090100613; CNPq/PROSUL project number 490749/2008-9, Brazil and the Ufficio Relazioni Europee e Internazionali del Consiglio Nazionale delle Ricerche, Italy (Laboratori Congiunti Bilaterali Internazionali CNR, Prot. 0005651).
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The handling Editor is currently organizing a Research Topic with one of the authors FP, and confirms the absence of any other collaboration.
MM, CC, CA, and MP received fellowships from CONICET. MP, SF, LS, JS, AG, JO, and SP are research staff members of CONICET. Thanks are due to Dr. María Sartor for cytoembryology/ISH experiments technical support and discussion, Ana Cristina M. M. Gomes for help with the Brachiaria ovules sectioning and Dr. Celina Beltrán for collaborating with statistical analyses.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fpls.2018.01547/full#supplementary-material
Albertini, E., Marconi, G., Barcaccia, G., Raggi, L., and Falcinelli, M. (2004). Isolation of candidate genes for apomixis in Poa pratensis. Plant Mol. Biol. 56, 879–894. doi: 10.1007/s11103-004-5211-y
Alexander, M. P. (1980). Differential staining of aborted and non-aborted pollen. Stain Technol. 44, 117–122. doi: 10.3109/10520296909063335
Ariel, F., Romero-Barrios, N., Jégu, T., Benhamed, M., and Crespi, M. (2015). Battles and hijacks: noncoding transcription in plants. Trends Plant Sci. 20, 362–371. doi: 10.1016/j.tplants.2015.03.003
Bicknell, R. A., and Koltunow, A. M. (2004). Understanding apomixis: recent advances and remaining conundrums. Plant Cell 16, 228–245. doi: 10.1105/tpc.017921
Borg, M., Brownfield, L., Khatab, H., Sidorova, A., Lingaya, M., and Twell, D. (2011). The R2R3 MYB transcription factor DUO1 activates a male germline-specific regulon essential for sperm cell differentiation in Arabidopsis. Plant Cell 23, 534–549. doi: 10.1105/tpc.110.081059
Carman, J. G. (1997). Asynchronous expression of duplicate genes in angiosperms may cause apomixis, bispory, tetraspory, and polyembryony. Biol. J. Linn. Soc. 61, 51–94. doi: 10.1111/j.1095-8312.1997.tb01778.x
Cervigni, G. D., Paniego, N., Díaz, M., Selva, J. P., Zappacosta, D., Zanazzi, D., et al. (2008). Expressed sequence tag analysis and development of associated markers in a near-isogenic plant system of Eragrostis curvula. Plant Mol. Biol. 67, 7–10. doi: 10.1007/s11103-007-9282-4
Chekanova, J. A. (2015). Long non-coding RNA and their functions in plants. Curr. Opin. Plant Biol. 27, 207–216. doi: 10.1016/j.pbi.2015.08.003
Daigle, C., and Matton, D. P. (2015). Genome-wide analysis of MAPKKKs shows expansion and evolution of a new MEKK class involved in solanaceous species sexual reproduction. BMC Genomics 16:1037. doi: 10.1186/s12864-015-2228-3
Felitti, S. A., Seijo, J. G., González, A. M., Podio, M., Laspina, N. V., Siena, L., et al. (2011). Expression of lorelei-like genes in aposporous and sexual Paspalum notatum plants. Plant Mol. Biol. 77, 337–354. doi: 10.1007/s11103-011-9814-9
Fiil, B. K., Petersen, K., Petersen, M., and Mundy, J. (2009). Gene regulation by MAP kinase cascades. Curr. Opin. Plant Biol. 12, 615–621. doi: 10.1016/j.pbi.2009.07.017
Garcia-Aguilar, M., Michaud, C., Leblanc, O., and Grimanelli, D. (2010). Inactivation of a DNA methylation pathway in maize reproductive organs results in apomixis-like phenotypes. Plant Cell 22, 3249–3267. doi: 10.1105/tpc.109.072181
Hand, M., and Koltunow, A. (2014). The genetic control of apomixis: asexual seed formation. Genetics 197, 441–450. doi: 10.1534/genetics.114.163105
Ichimura, K., Shinozaki, K., Tena, G., Sheen, J., Henry, Y., Champion, A., et al. (2002). Mitogen-activated protein kinase cascades in plants: a new nomenclature. Trends Plant Sci. 7, 301–308. doi: 10.1016/S1360-1385(02)02302-6
Kapadia, B., Viswakarma, N., Parsa, K. V. L., Kain, V., Behera, S., Suraj, S. K., et al. (2013). ERK2-mediated phosphorylation of transcriptional coactivator binding protein PIMT/NcoA6IP at Ser298 augments hepatic gluconeogenesis. PLoS One 8:e83787. doi: 10.1371/journal.pone.0083787
Kruskal, W. H., and Wallis, W. A. (1952). Use of ranks in one-criterion variance analysis. J. Am. Stat. Assoc. 47, 583–621. doi: 10.1080/01621459.1952.10483441
Larkin, M. A., Blackshields, G., Brown, N. P., Chenna, R., MCGettigan, P. A., McWilliam, H., et al. (2007). Clustal W and Clustal X version 2.0. Bioinformatics 23, 2947–2948. doi: 10.1093/bioinformatics/btm404
Laspina, N. V., Vega, T., Martelotto, L., Stein, J., Podio, M., Ortiz, J. P., et al. (2008). Gene expression analysis at the onset of aposporous apomixis in Paspalum notatum. Plant Mol. Biol. 67, 615–628. doi: 10.1007/s11103-008-9341-5
Liu, X., Hao, L., Li, D., Zhu, L., and Hu, S. (2015). Long non-coding RNAs and their biological roles in plants. Genomics Proteomics Bioinformatics 13, 137–147. doi: 10.1016/j.gpb.2015.02.003
Lukowitz, W., Roeder, A., Parmenter, D., and Somerville, C. (2004). A MAPKK kinase gene regulates extra-embryonic cell fate in Arabidopsis. Cell 116, 109–119. doi: 10.1016/S0092-8674(03)01067-5
Mancini, M., Woitovich, N., Permingeat, H., Podio, M., Siena, L. A., Ortiz, J. P. A., et al. (2014). Development of a modified transformation platform for apomixis candidate genes research in Paspalum notatum (bahiagrass). In Vitro Cell. Dev. Biol. Plant 50, 412–424. doi: 10.1007/s11627-014-9596-2
Musielak, T. J., and Bayer, M. (2014). YODA signalling in the early Arabidopsis embryo. Biochem. Soc. Trans. 42, 408–412. doi: 10.1042/BST20130230
Newcombe, R. G. (1998). Two-sided confidence intervals for the single proportion: comparison of seven methods. Stat. Med. 17, 857–872. doi: 10.1002/(SICI)1097-0258(19980430)17:8<857::AID-SIM777>3.0.CO;2-E
Nogler, G. A. (1984). “Gametophytic apomixis,” in Embryology of Angiosperms, ed. B. M. Johri (Berlin, Germany: Springer-Verlag), 475–518.
Ochiai-Fukuda, T., Takahashi-Ando, N., Ohsato, S., Igawa, T., Kadokura, K., Hamamoto, H., et al. (2006). A fluorescent antibiotic resistance marker for rapid production of transgenic rice plants. J. Biotechnol. 122, 521–527. doi: 10.1016/j.jbiotec.2005.09.015
Ochogavía, A. C., Seijo, J. G., González, A. M., Podio, M., Laspina, N. V., Duarte Silveira, E., et al. (2011). Characterization of retrotransposon sequences expressed in inflorescences of apomictic and sexual Paspalum notatum plants. Sex. Plant Reprod. 24, 231–246. doi: 10.1007/s00497-011-0165-0
Okada, T., Hu, Y., Tucker, M. R., Taylor, J. M., Johnson, S. D., Spriggs, A., et al. (2013). Enlarging cells initiating apomixis in Hieracium praealtum transition to an embryo sac program prior to entering mitosis. Plant Physiol. 163, 216–231. doi: 10.1104/pp.113.219485
Ortiz, J. P., Pessino, S. C., Leblanc, O., Hayward, M. D., and Quarin, C. L. (1997). Genetic fingerprint for determinig the mode of reproduction in Paspalum notatum, a subtropical apomictic forage grass. Theoret. Appl. Genet. 95, 850–856. doi: 10.1007/s001220050635
Ortiz, J. P. A., Quarin, C. L., Pessino, S. C., Acuña, C., Martínez, E. J., Espinoza, F., et al. (2013). Harnessing apomictic reproduction in grasses: what we have learned from Paspalum. Ann. Bot. 112, 767–787. doi: 10.1093/aob/mct152
Ortiz, J. P. A., Revale, S., Siena, L. A., Podio, M., Delgado, L., Stein, J., et al. (2017). A reference floral transcriptome of sexual and apomictic Paspalum notatum. BMC Genomics 18:318. doi: 10.1186/s12864-017-3700-z
Pagliarini, M. S., Carneiro Vieira, M. L., and Borges do Valle, C. (2012). “Meiotic behavior in intra- and interspecific sexual and somatic polyploid hybrids of some tropical species,” in Meiosis Molecular Mechanisms and Cytogenetic Diversity, ed. A. Swan (London: Intech Open). doi: 10.5772/30518
Paterson, A. H., Brubaker, C. L., and Wendel, J. F. (1993). A rapid method for extraction of cotton (Gossypium spp.) genomic DNA suitable for RFLP or PCR analysis. Plant Mol. Biol. Rep. 11, 122–127. doi: 10.1007/BF02670470
Paytuví Gallart, A., Hermoso Pulido, A., Martínez de Lagrán, I. A., Sanseverino, W., and Aiese Cigliano, R. (2016). GREENC: a Wiki-based database of plant lncRNAs. Nucleic Acids Res. 44, 1161–1166. doi: 10.1093/nar/gkv1215
Pessino, S. C., Espinoza, F., Martínez, E. J., Ortiz, J. P. A., Valle, E. M., and Quarin, C. L. (2001). Isolation of cDNA clones differentially expressed in flowers of apomictic and sexual Paspalum notatum. Hereditas 134, 35–42. doi: 10.1111/j.1601-5223.2001.00035.x
Pessino, S. C., Evans, C., Ortiz, J. P. A., Armstead, I., do Valle, C. B., and Hayward, M. D. (1998). A genetic map of the apospory region in Brachiaria hybrids: identification of two markers closely associated with the trait. Hereditas 128, 153–158. doi: 10.1111/j.1601-5223.1998.00153.x
Pessino, S. C., Ortiz, J. P. A., Leblanc, O., do Valle, C. B., Evans, C., and Hayward, M. D. (1997). Identification of a maize linkage group related to apomixis in Brachiaria. Theoret. Appl. Genet. 94, 439–444. doi: 10.1007/s001220050434
Podio, M., Cáceres, M. E., Samoluk, S. S., Seijo, J. G., Pessino, S. C., Ortiz, J. P. A., et al. (2014a). A methylation status analysis of the apomixis specific region in Paspalum spp. suggests an epigenetic control of parthenogenesis. J. Exp. Bot. 65, 6411–6424. doi: 10.1093/jxb/eru354
Podio, M., Felitti, S. A., Siena, L. A., Delgado, L., Mancini, M., Seijo, G., et al. (2014b). Characterization and expression analysis of SOMATIC EMBRYOGENESIS RECEPTOR KINASE (SERK) genes in sexual and apomictic Paspalum notatum. Plant Mol. Biol. 84, 479–495. doi: 10.1007/s11103-013-0146-9
Podio, M., Siena, L. A., Hojsgaard, D., Stein, J., Quarin, C. L., and Ortiz, J. P. A. (2012). Evaluation of meiotic abnormalities and pollen viability in aposporous and sexual tetraploid Paspalum notatum (Poaceae). Plant Syst. Evol. 298, 1625–1633. doi: 10.1007/s00606-012-0664-y
Polegri, L., Calderini, O., Arcioni, S., and Pupilli, F. (2010). Specific expression of apomixis-linked alleles revealed by comparative transcriptomic analysis of sexual and apomictic Paspalum simplex Morong flowers. J. Exp. Bot. 61, 1869–1883. doi: 10.1093/jxb/erq054
Pupilli, F., Labombarda, P., Cáceres, M. E., Quarin, C. L., and Arcioni, S. (2001). The chromosome segment related to apomixis in Paspalum simplex is homoeologous to the telomeric region of the long arm of rice chromosome 12. Mol. Breed. 8, 53–61. doi: 10.1023/A:1011966922301
Quarin, C. L., Espinoza, F., Martínez, E. J., Pessino, S. C., and Bovo, O. A. (2001). A rise of ploidy level induces the expression of apomixis in Paspalum notatum. Sex. Plant Reprod. 13, 243–249. doi: 10.1007/s004970100070
Quarin, C. L., Urbani, M. H., Blount, A. R., Martínez, E. J., Hack, C. M., Burton, G. W., et al. (2003). Registration of Q4188 and Q4205, sexual tetraploid germplasm lines of Bahiagrass. Crop Sci. 43, 745–746. doi: 10.2135/cropsci2003.0745
Rodrigues, J. C., Cabral, G. B., Dusi, D. M. A., Mello, L. V., Rinden, D., and Carneiro, V. T. C. (2003). Identification of differentially expressed cDNA sequences in ovaries of sexual and apomictic plants of Brachiaria brizantha. Plant Mol. Biol. 53, 745–757. doi: 10.1023/B:PLAN.0000023664.21910.bd
Ronceret, A., and Vielle-Calzada, J. P. (2015). Meiosis, unreduced gametes, and parthenogenesis: implications for engineering clonal seed formation in crops. Plant Reprod. 28, 91–102. doi: 10.1007/s00497-015-0262-6
Siena, L. A., Ortiz, J. P. A., Leblanc, O., and Pessino, S. (2014). PnTgs1-like expression during reproductive development supports a role for RNA methyltransferases in the aposporous pathway. BMC Plant Biol. 14:297. doi: 10.1186/s12870-014-0297-0
Shapiro, S. S., and Wilk, M. B. (1965). An analysis of variance test for normality (complete samples). Biometrika 52, 591–611. doi: 10.1093/biomet/52.3-4.591
Sharbel, T. F., Voigt, M. L., Corral, J. M., Galla, G., Kumlehn, J., Klukas, C., et al. (2010). Apomictic and sexual ovules of Boechera display heterochronic global gene expression patterns. Plant Cell 22, 655–671. doi: 10.1105/tpc.109.072223
Sharbel, T. F., Voigt, M. L., Corral, J. M., Thiel, T., Varshney, A., Kumlehn, J., et al. (2009). Molecular signatures of apomictic and sexual ovules in the Boechera holboellii complex. Plant J. 58, 870–882. doi: 10.1111/j.1365-313X.2009.03826.x
Siena, L. A., Ortiz, J. P. A., Calderini, O., Paolocci, F., Cáceres, M. E., Kaushal, P., et al. (2016). An apomixis-linked ORC3-like pseudogene is associated with silencing of its functional homolog in apomictic Paspalum simplex. J. Exp. Bot. 67, 1965–1978. doi: 10.1093/jxb/erw018
Sneath, P. H. A., and Sokal, R. R. (1973). “The estimation of taxonomic resemblance,” in Numerical Taxonomy. The Principles and Practice of Numerical Classification, eds D. Kennedy and R. B. Park (San Francisco, CA: Freeman), 129–132.
Stein, J., Pessino, S. C., Martínez, E. J., Rodríguez, M. P., Siena, L. A., Quarin, C. L., et al. (2007). A genetic map of tetraploid Paspalum notatum Flügge (bahiagrass) based on single-dose molecular markers. Mol. Breed. 20, 153–166. doi: 10.1007/s11032-007-9083-0
Tamura, K., Stecher, G., Peterson, D., Filipski, A., and Kumar, S. (2013). MEGA6: molecular evolutionary genetics analysis Version 6.0. Mol. Biol. Evol. 30, 2725–2729. doi: 10.1093/molbev/mst197
Thompson, C. J., Movva, N. R., Tizard, R., Crameri, R., Davies, J. E., and Lauwereys, M. (1987). Characterization of the herbicide-resistance gene bar from Streptomyces hygroscopicus. EMBO J. 6, 2519–2523. doi: 10.1002/j.1460-2075.1987.tb02538.x
Van Norman, J. M., Breakfield, N. W., and Benfey, P. N. (2011). Intercellular communication during plant development. Plant Cell 23, 855–864. doi: 10.1105/tpc.111.082982
Wilcoxon, F. (1945). Individual comparisons by ranking methods. Biometrics Bull. 1, 80–83. doi: 10.2307/3001968
Wilson, E. B. (1927). Probable inference, the law of succession, and statistical inference. J. Am. Stat. Assoc. 22, 209–212. doi: 10.1080/01621459.1927.10502953
Wortzel, I., and Seger, R. (2011). The ERK cascade: distinct functions within various subcellular organelles. Genes Cancer 2, 195–209. doi: 10.1177/1947601911407328
Xu, J., and Zhang, S. (2015). Mitogen-activated protein kinase cascades in signalling plant growth and development. Trends Plant Sci. 20, 56–64. doi: 10.1016/j.tplants.2014.10.001
Yamada-Akiyama, H., Akiyama, Y., Ebina, M., Xu, Q., Tsuruta, S., Yazaki, J., et al. (2009). Analysis of expressed sequence tags in apomictic Guinea grass (Panicum maximum). J. Plant Physiol. 166, 750–761. doi: 10.1016/j.jplph.2008.10.001
Zhang, W., McElroy, D., and Wu, R. (1991). Analysis of rice actl 5′ region activity in transgenic rice plants. Plant Cell 3, 1155–1165.
Keywords: apomixis, apospory, LNC-QGJ, MAP3K, Paspalum notatum, plant reproduction, QGJ, QUI-GON
Citation: Mancini M, Permingeat H, Colono C, Siena L, Pupilli F, Azzaro C, de Alencar Dusi DM, de Campos Carneiro VT, Podio M, Seijo JG, González AM, Felitti SA, Ortiz JPA, Leblanc O and Pessino SC (2018) The MAP3K-Coding QUI-GON JINN (QGJ) Gene Is Essential to the Formation of Unreduced Embryo Sacs in Paspalum. Front. Plant Sci. 9:1547. doi: 10.3389/fpls.2018.01547
Received: 02 July 2018; Accepted: 03 October 2018;
Published: 24 October 2018.
Edited by:
Andrea Mazzucato, Università degli Studi della Tuscia, ItalyReviewed by:
Takashi Okada, University of Adelaide, AustraliaCopyright © 2018 Mancini, Permingeat, Colono, Siena, Pupilli, Azzaro, de Alencar Dusi, de Campos Carneiro, Podio, Seijo, González, Felitti, Ortiz, Leblanc and Pessino. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Silvina C. Pessino, cGVzc2lub0Bhcm5ldC5jb20uYXI=
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.