
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Plant Sci. , 21 February 2018
Sec. Plant Breeding
Volume 9 - 2018 | https://doi.org/10.3389/fpls.2018.00216
This article is part of the Research Topic Generating Useful Genetic Variation in Crops by Induced Mutation View all 8 articles
 Miriam E. Szurman-Zubrzycka1
Miriam E. Szurman-Zubrzycka1 Justyna Zbieszczyk1
Justyna Zbieszczyk1 Marek Marzec1Janusz Jelonek1Beata Chmielewska1
Marek Marzec1Janusz Jelonek1Beata Chmielewska1 Marzena M. Kurowska1Milena Krok1
Marzena M. Kurowska1Milena Krok1 Agata Daszkowska-Golec1
Agata Daszkowska-Golec1 Justyna Guzy-Wrobelska1
Justyna Guzy-Wrobelska1 Damian Gruszka1Monika Gajecka1Patrycja Gajewska1
Damian Gruszka1Monika Gajecka1Patrycja Gajewska1 Magdalena Stolarek1Piotr Tylec1
Magdalena Stolarek1Piotr Tylec1 Paweł Sega1Sabina Lip1Monika Kudełko1Magdalena Lorek1Małgorzata Gorniak-Walas1
Paweł Sega1Sabina Lip1Monika Kudełko1Magdalena Lorek1Małgorzata Gorniak-Walas1 Anna Malolepszy1†Nina Podsiadlo1
Anna Malolepszy1†Nina Podsiadlo1 Katarzyna P. Szyrajew1Anete Keisa2Zodwa Mbambo3
Katarzyna P. Szyrajew1Anete Keisa2Zodwa Mbambo3 Elena Todorowska4Marek Gaj1Zygmunt Nita5Wanda Orlowska-Job5Miroslaw Maluszynski1
Elena Todorowska4Marek Gaj1Zygmunt Nita5Wanda Orlowska-Job5Miroslaw Maluszynski1 Iwona Szarejko1*
Iwona Szarejko1*TILLING (Targeting Induced Local Lesions IN Genomes) is a strategy used for functional analysis of genes that combines the classical mutagenesis and a rapid, high-throughput identification of mutations within a gene of interest. TILLING has been initially developed as a discovery platform for functional genomics, but soon it has become a valuable tool in development of desired alleles for crop breeding, alternative to transgenic approach. Here we present the HorTILLUS (Hordeum—TILLING—University of Silesia) population created for spring barley cultivar “Sebastian” after double-treatment of seeds with two chemical mutagens: sodium azide (NaN3) and N-methyl-N-nitrosourea (MNU). The population comprises more than 9,600 M2 plants from which DNA was isolated, seeds harvested, vacuum-packed, and deposited in seed bank. M3 progeny of 3,481 M2 individuals was grown in the field and phenotyped. The screening for mutations was performed for 32 genes related to different aspects of plant growth and development. For each gene fragment, 3,072–6,912 M2 plants were used for mutation identification using LI-COR sequencer. In total, 382 mutations were found in 182.2 Mb screened. The average mutation density in the HorTILLUS, estimated as 1 mutation per 477 kb, is among the highest mutation densities reported for barley. The majority of mutations were G/C to A/T transitions, however about 8% transversions were also detected. Sixty-one percent of mutations found in coding regions were missense, 37.5% silent and 1.1% nonsense. In each gene, the missense mutations with a potential effect on protein function were identified. The HorTILLUS platform is the largest of the TILLING populations reported for barley and best characterized. The population proved to be a useful tool, both in functional genomic studies and in forward selection of barley mutants with required phenotypic changes. We are constantly renewing the HorTILLUS population, which makes it a permanent source of new mutations. We offer the usage of this valuable resource to the interested barley researchers on cooperative basis.
Mutants are essential for functional analysis of genes. There are many techniques, available today, of creating mutants that can be used to study gene function. Among these techniques there are standard chemical or physical mutagenic treatments causing mutations that are spread throughout the genome, as well as modern techniques of gene editing, such as CRISPR/Cas9-based system that can be used for creation of mutations within a specific target gene (Cong et al., 2013). The latter one, however, is still not available for many plant species, for which the protocols of transformation and/or in vitro regeneration are not available or are difficult to optimize for a wider range of genotypes. In the case of barley, such protocols are well established only for one variety “Golden Promise” (a Scottish cultivar developed in 1967; Lawrenson et al., 2015), while transformation efficiency of modern barley cultivars is still insufficient for a routine use. Therefore, the methods, such as TILLING (Targeting Induced Local Lesions IN Genomes), are still relevant for functional analysis of genes in many species, including barley.
TILLING is a strategy used for functional analysis of genes that combines the classical mutagenesis and a rapid, high-throughput identification of mutations within a gene of interest. TILLING has been initially developed as a discovery platform for functional genomics in Arabidopsis thaliana (McCallum et al., 2000). In a few years, besides Arabidopsis (Till et al., 2003), TILLING populations have been created for other model plants, e.g., Lotus japonicus (Perry et al., 2009) and Medicago truncatula (Le Signor et al., 2009), and model animals: fruit fly (Winkler et al., 2005), zebra fish (Wienholds et al., 2003), and rat (Smits et al., 2004). The main advantage of TILLING as a reverse genetics strategy is that it can be applied to any species, regardless of its genome size and ploidy level.
Soon TILLING has become a valuable tool in development of desired alleles for crop breeding (reviewed in Kurowska et al., 2011; Tadele, 2016). The utility of TILLING has been demonstrated in many crop species, including cereals: maize (Till et al., 2004), rice (Till et al., 2007; Suzuki et al., 2008), sorghum (Xin et al., 2008), durum (Uauy et al., 2009), bread (Slade et al., 2005; Uauy et al., 2009; Fitzgerald et al., 2010; Chen et al., 2012), and einkorn wheat (Rawat et al., 2012); legumes and oil crops: pea (Dalmais et al., 2008), common bean (Porch et al., 2009), soybean (Cooper et al., 2008), sunflower (Kumar et al., 2013), rapeseed (Gilchrist et al., 2013); vegetables: tomato (Gady et al., 2009; Minoia et al., 2010; Piron et al., 2010; Okabe et al., 2011), pumpkin (Vicente-Dólera et al., 2014), cucumber (Fraenkel et al., 2014), radish (Kohzuma et al., 2017); industrial species: cotton (Aslam et al., 2016), flax (Chantreau et al., 2013), tobacco (Reddy et al., 2012); and staple tropical crops, such as banana (Jankowicz-Cieslak et al., 2012).
Barley is among the most important cereal species grown worldwide. In the statistics showing the world grain production in 2016/2017, barley is at the fourth position in terms of the harvested acreage and quantity of produced grain, following corn, wheat, and rice (www.statista.com/statistics/263977/world-grain-production-by-type/). Barley grain is used mainly as animal fodder, as a source of malt in brewery industry and as a component of various health foods. Barley exhibits a high genetic adaptability to a wide range of environments and serves as a model cereal for studying mechanisms of adaptation to abiotic stresses (Dawson et al., 2015). The role of barley as an important crop species is reflected in development of molecular tools, among them TILLING, that assist its functional analysis and breeding. Over years several TILLING populations have been created for this crop (Caldwell et al., 2004; Talamè et al., 2008; Gottwald et al., 2009; Lababidi et al., 2009), and the release and assembling of barley genome sequence have facilitated the use of TILLING platforms for functional genomics studies (International Barley Genome Sequencing Consortium et al., 2012; Beier et al., 2017; Mascher et al., 2017).
In this paper we describe a new TILLING platform in barley created at the University of Silesia in Katowice, Poland. The population called HorTILLUS (Hordeum vulgare—TILLING—University of Silesia)—was developed for spring barley variety “Sebastian” after double treatment with two chemical mutagens: sodium azide (NaN3) and N-Methyl-N-nitrosourea (MNU). It can serve as a platform for functional genomics and pre-breeding programs of barley. The HorTILLUS population has proved its utility as a reverse genetics tool in many barley studies concerning e.g., brassinosteroid metabolism (Gruszka et al., 2016), DNA repair (Stolarek et al., 2015a,b), strigolactone signaling (Marzec et al., 2016), waterlogging tolerance (Mendiondo et al., 2016), or drought and ABA response (Daszkowska-Golec et al., 2017). This population is being constantly renewed to make it functional for many years. We present the HorTILLUS platform as a renewable source of new mutations and mutants which we want to share within barley community in a cooperative manner.
Spring barley cultivar “Sebastian” (Supplementary Figure 1) was used as a parent variety to create the HorTILLUS population. This cultivar was chosen because of its high yield potential, good malting quality, resistance to lodging and high resistance to stem rust (Puccinia graminis) and leaf rust (Puccinia hordei). It is also characterized by an average resistance to powdery mildew (Blumeria graminis f.sp. hordei), net blotch (Pyrenophora teres), and scald (Rynchosporium secalis).
Seeds of “Sebastian” were mutagenized with the use of two mutagens: sodium azide (NaN3) and N-Methyl-N-nitrosourea (MNU), with inter-incubation germination period (iig) (Szarejko and Maluszynski, 1999; Szarejko et al., 2017). Two treatments were applied, both with the same dose of NaN3, but two different doses of MNU: a higher dose (1.5 mM NaN3/3 h–6 h iig – 0.75 mM MNU/3 h) and a lower dose (1.5 mM NaN3/3 h–6 h iig – 0.5 mM MNU/3 h).
The scheme of creation of HorTILLUS population is shown in Figure 1. The M1 population was grown at experimental field and harvested individually. All M1 spikes were stored but only one M2 plant was developed from each M1 plant in order to avoid repetitions of the same mutations in M2 progenies. After 2 weeks of seedlings hardening at 4°C, M2 plants were grown under controlled conditions in a greenhouse (photoperiod 16/8 h, light intensity 400 μM/m2/s and temperature 22/20°C during day and night, respectively) until harvest.
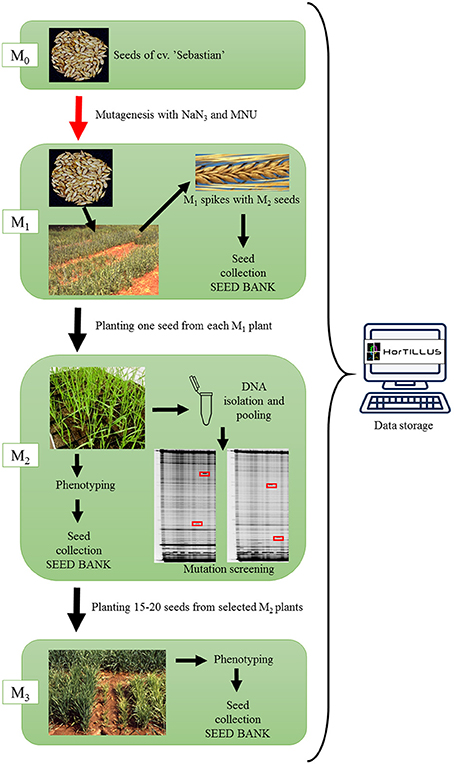
Figure 1. The scheme of HorTILLUS population creation.
M2 generation of initial HorTILLUS population has been created using almost 15,000 M1 plants developed after double treatment with sodium azide and MNU. Among those M1 plants, 4,716 were developed after treatment with a higher dose of MNU (1.5 mM NaN3/3 h–6 h iig – 0.75 mM MNU/3 h) and 10,116 after treatment with a lower dose of MNU (1.5 mM NaN3/3 h–6 h iig – 0.5 mM MNU/3 h). One seed from each M1 plant was planted and in total 13,181 M2 seeds germinated. DNA isolation and phenotypic analysis were performed on over 11,000 M2 plants.
All M2 plants were morphologically characterized at different developmental stages—from a seedling stage to maturity. During post-harvest analysis plants were measured and described for several characters, such as: number of tillers, fertility, number, and weight of seeds and any visible morphological changes. Mutants with visible changes were photographed using Fujifilm S9600 digital camera and the images were stored in a database.
Fifteen to twenty seeds from 3,481 individual M2 plants producing a high number of seeds were planted in the field belonging to Plant Breeding and Acclimatization Institute in Strzelce. M3 seeds from the same M2 plant were sown in two neighboring rows in plots and all M3 seedlings were carefully labeled. M3 progenies were phenotyped at different developmental stages, for different traits, such as presence of chlorophyll seedlings, shape and color of rosettes, morphology of leaves and spikes, time of flowering, height of plants, etc. M3 plants with morphological changes were also photographed. All data collected for individual plants and lines of M3 generation were stored in the database. M4 seeds were stored in a seed bank.
The proper storage of material in a seed bank is crucial for maintaining the TILLING population. Therefore, we vacuum-packed the seeds of individual plants from M1-M4 generations (using vacuum packer Tepro pp 5.4) and stored them at 4°C.
It should be stressed that after a few years of intensive usage, some seed samples of M2 plants were almost expended and/or the germination capacity of others decreased. To keep the HorTILLUS population suitable for a long-term use we have decided to gradually renew our population by constantly growing new M2 plants and isolating DNA from them. These M2 plants derived from the same mutagenic treatments as those used for creation of the initial TILLING platform.
DNA was isolated from M2 HorTILLUS plants individually using a modified micro-CTAB method (Doyle and Doyle, 1987) that ensures a large amount of high quality DNA. Leave samples (~3 cm2 segments) of 2–4-weeks-old plants were collected in silica gel in order to remove water from the tissue. Dried samples were ground in single eppendorf tubes with 3 mm glassballs (Sigma-Aldrich) using an electric mill (Retsch, MM200 and MM301). The exact protocol for DNA isolation from barley used for this study was described elsewhere (Szarejko et al., 2017). DNA was isolated from over 11,000 M2 plants.
DNA quantity and quality was measured using NanoDrop™ ND-1000 UV-Vis Spectrophotometer. Stocks were diluted to 100 ng/μl. DNA stocks were stored at −80°C, whereas the dilutions were stored in −20°C. The dilutions were used to prepare eight-fold pools in 96-well plates (Supplementary Figure 2). One well of the pool plate contains DNA from 8 different M2 plants, so one pool plate contains DNA from 768 M2 plants. Eight-fold pools served as a templates for PCR reactions.
Candidates for TILLING analysis were chosen based on literature data as genes potentially related to biological processes of our interest, such as brassinosteroid metabolism, response to abiotic stresses, root development or DNA repair (Table 1). GenBank (http://www.ncbi.nlm.nih.gov/genbank/) database was searched for sequences of candidates (mainly from Arabidopsis and rice). The selection of some genes was performed before the release of barley genome sequence (International Barley Genome Sequencing Consortium et al., 2012; Beier et al., 2017), therefore barley genomic sequences of candidates were unavailable. The homologous sequences of gene of interest from rice were retrieved in order to clone barley homologs. Rice mRNA or amino acid sequence were used as a query in barley Expressed Sequence Tags (EST) database (http://compbio.dfci.harvard.edu/tgi/). For the best of obtained hits primers were designed using i.e., Primer3 (http://bioinfo.ut.ee/primer3-0.4.0/) with the purpose of amplifying, sequencing and assembling a full coding sequences of barley homologs. After that, the barley coding sequence was aligned with the genomic sequence of the rice homolog using BLASTN algorithm or Splign (http://www.ncbi.nlm.nih.gov/sutils/splign/splign.cgi) what allowed to predict exon/intron putative borders in the barley gene and design primers for amplification of intron regions. Assembling of amplified sequences resulted in full barley genomic sequence. When the barley genome sequence became publicly available, it was possible to search directly for barley genes in databases, such as EnsemblPlants database (http://plants.ensembl.org/Hordeum_vulgare/Info/Index) or PLAZA 3.0 Monocots database (https://bioinformatics.psb.ugent.be/plaza/versions/plaza_v3_monocots/organism/view/Hordeum+vulgare).
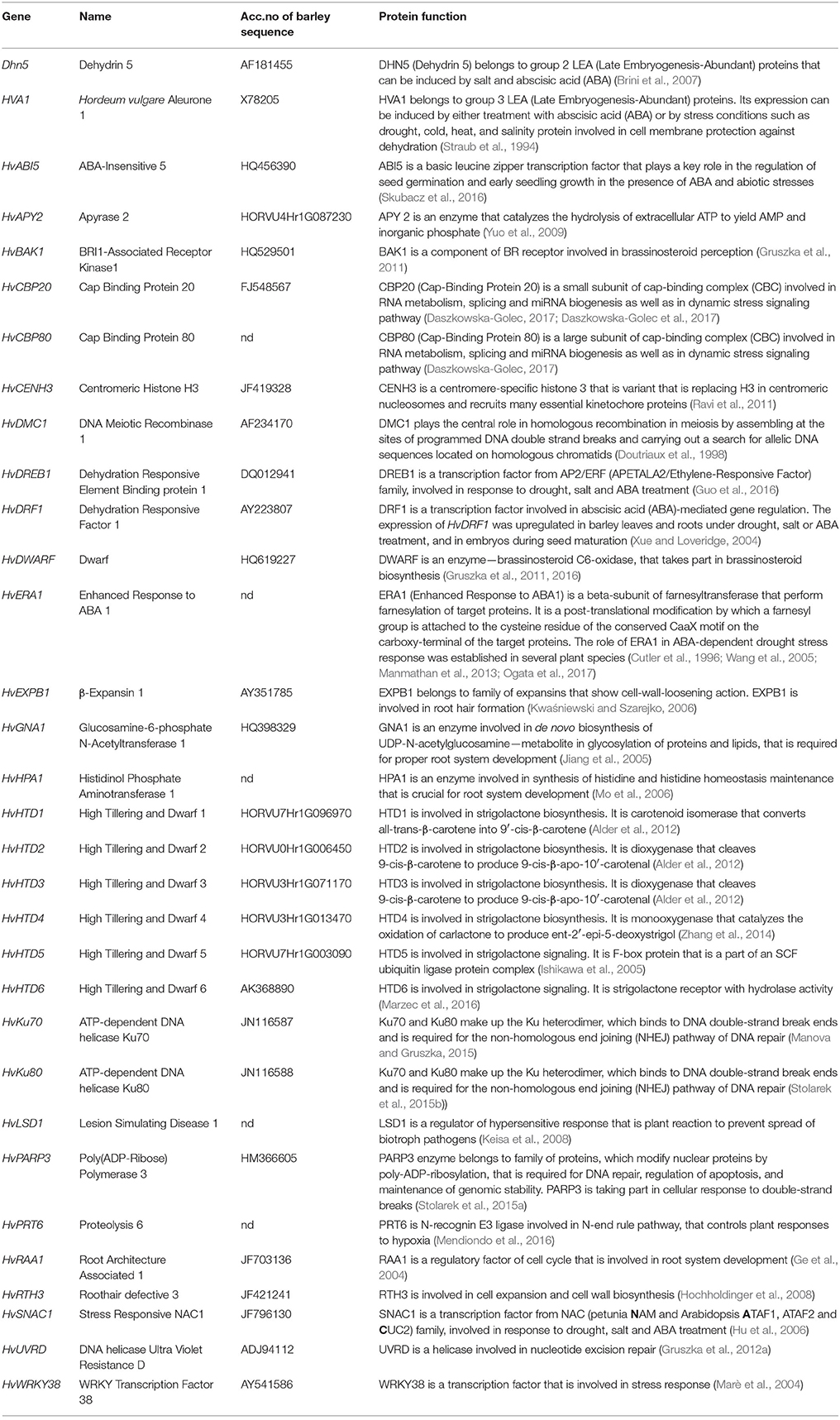
Table 1. Candidate genes selected for TILLING analysis.
In order to analyze a gene with the use of TILLING strategy it is necessary to determine a fragment of a gene where a point mutation has the highest probability to occur and change a protein function. Each of the studied genes was analyzed using CODDLE software (Choosing codons to Optimize Discovery of Deleterious Lesions; http://blocks.fhcrc.org/proweb/coddle/). As an input, the genomic and coding sequences of barley genes were used. CODDLE performs a BLAST search of known proteins to find conserved regions. CODDLE is based on two algorithms—SIFT (Sorting Intolerant From Tolerant) and PSSM (Position Specific Scoring Matrix). Both these algorithms weigh the identity and frequency of the amino acids found at each position in the protein. CODDLE separately handles the prediction of changes that could truncate the protein and destabilize the RNA (nonsense changes and splice junction changes), and the prediction of missense changes which should alter function of the gene product (located in conserved amino acid blocks in the CDS). In an addition to CODDLE analysis, to verify the conserved regions within the protein of interest we performed alignments of homologous proteins using ClustalW (http://www.ebi.ac.uk/Tools/msa/clustalw2/). Because both agents used as mutagens for HorTILLUS create mostly G/C to A/T transitions, also the GC content was analyzed within fragments chosen for analysis. Fragments for TILLING analysis were 700–1,200 bp in length and included a regions determined by the above described methods. Primers that allow amplification of chosen region(s) (Supplementary Table 1) were designed using i.e. Primer3 (http://primer3.ut.ee/).
The method of mutation detection used in described study consists of the following steps:
(1) PCR on eight-fold DNA pools using differentially labeled primers (forward primer labeled with IRDye700, reverse primer labeled with IRDye800)
(2) Heteroduplex formation by denaturation and slow renaturation of PCR products (heteroduplex with a single nucleotide mismatch appear only in pools that contain DNA from plant carrying a mutation within amplified fragment)
(3) Enzymatic digestion using endonuclease (Cel-1) that specifically recognize and cuts heteroduplex in a mismatch position
(4) Purification of products using ethanol
(5) Electrophoresis in polyacrylamide gel in LI-COR Sequencer
(6) Analysis of gel image and identification of positive pools containing DNA with potential mutation within amplified fragment—PCR products should be visible in both channels in LI-COR sequencer (700 and 800 nm). When heteroduplex is cut by Cel-1, one fragment contains only IRDye700 label, the other only IRDye800 label, that is why the bands indicating mutation are visible in different channels. The sum of length of this two corresponding bands should be equal to the length of PCR product
(7) Identification of particular plants from positive pools carrying potential mutations by performing steps 1–6 on two-fold pools containing DNA of parental variety (non-mutated) mixed with DNA of each individual from positive pool separately
(8) Sequencing of analyzed fragments from selected plants in order to confirm mutations and check their type and zygosity state.
IRDye700 and IRDye800 labeled primers as well as unlabeled ones were diluted to 20 μM and used to prepare primer “cocktail” (in a 3*: 2: 4*: 1 proportion for IRDye700 labeled forward, unlabeled forward, IRDye800 labeled reverse, unlabeled reverse primers, respectively). PCR reactions were performed in 20 μl volume and the concentration of PCR substrates, as well as the conditions of amplification were optimized for each pair of primers individually. After amplification, the heteroduplex formation was performed at 95°C per 3 min for initial denaturation, and then at 70°C per 20 s (x 70 cycles) −0.1°C per cycle for slow renaturation. After heteroduplex formation 10 μl of samples were treated with 20 μl of 0.1 × Celery Juice Extract (CJE) containing Cel I enzyme, which was kindly provided by B. Till from IAEA, Seibersdorf Laboratory, where it was isolated according to the protocol described previously (Till et al., 2006). The enzymatic cleavage was performed at 45°C for 15 min. The next step was purification of products with ethanol. Sixty microliters of 96% ethanol with 1% of sodium acetate were added to each sample and centrifuged at 4°C, 18,000 × g for 20 min in order to precipitate DNA fragments. Afterwards, the ethanol was removed and DNA pellets were washed in 30 μl of 70% ethanol and centrifuged in the same conditions as previously for 15 min. After drying the samples from the rest of ethanol at 80°C, 3 μl of STOP buffer, containing 5% of bromophenol blue-xylene, 40% of formamide, and 1% of EDTA, was added.
For the purpose of visualization of products, samples were denatured and the polyacrylamide electrophoresis was carried out in LI-COR sequencers (LI-COR 4300). Electrophoresis was performed in denaturating 6% polyacrylamide gel in 1xTBE (Tris—Boric Acid—EDTA) running buffer, at settings of 3,000 V, 30 mA, and 30 W.
The phpMyAdmin program and the MySQL system were used for creating HorTILLUS database. phpMyAdmin is one of the most popular tools with freeware license for administrating the databases. MySQL is a relational database management system (RDBMS) that runs as a server providing multi-user access to a number of databases.
HorTILLUS database includes a detailed description of individual plants from M2 generation and their progenies. The data about plants from various generations are collected in individual tables due to differential analyses subjected to generations. Several tables describing inter alia: phenotype and molecular analysis were created for each of generation. All the tables are linked by dedicated key entries that enable the identification of a single M2 plant and all next-generation (M3, M4, M5) individuals derived from this plant. The information on the preceding generation (M1) is also provided. This resolution enables a flexible scanning of database contents and a simultaneous browsing of the data in different generations.
HorTILLUS database includes description of changes in plant phenotypes and the information about mutations found in the analyzed genes (such as type of mutation, its precise position in TILLed fragment and zygosity state in M2 plant). Furthermore, there is information about fertility, number and weight of grains for each plant of M1, M2 generations and their progenies. The information about the number of grains used for phenotypic analyses and the number of grains left in the seed bank is also attached. All this options enable instant and simple supervision of seed bank status and, if necessary, multiplication of mutants which grains are running out.
The HorTILLUS database contains detailed information about quality and concentration of DNA isolated from M2 plants and about its place of storage. Hence, it is possible to prepare new dilutions of DNA and re-stock existing DNA pools. One of the tables of the database includes the information about created DNA pools used in mutation searching. After finding a potential mutation in a pool, all eight M2 plants whose DNA were pooled in this single bulk can be easily identified.
HorTILLUS database is supported by forms created in PHP and HTML languages. Therefore, it is possible to edit collected data and to enter new information using any Web browser. Created search scripts allow to ask a query about phenotype description and/or information about molecular analysis. Additionally, creation of database consisting of many individual tables allows their free modification and addition of extra tables for collecting additional information from any new studies.
Among 13,181 germinated M2 plants, 10% were lethal. In half of the cases (5.1%) lethality was caused by chlorophyll defects that led to development of albina, albinoviridis, striata, viridis, or xantha seedlings. A higher dose of MNU (0.7 mM) led to the higher frequency of lethal seedlings (12.1%) than the 0.5 mM MNU mM used in a combination with the same dose of 1.5 mM NaN3 (9.1%). Eleven thousand one hundred and ninety-nine M2 plants have been grown till maturity, among them 87.4% were fertile and produced seeds (Table 2). All M2 plants were characterized for obvious morphological changes. Reduction of plant height was the most common alteration of phenotype—almost 8% of analyzed plants were dwarf (>50% shorter than the parental variety) or semi-dwarfs (50–75% height of the parental variety “Sebastian”). Other phenotypic changes observed in M2 plants include alterations in plant architecture, time of flowering, and changes in the development of rosette, leaves, spikes, and awns (Table 3), including alterations in shape, color, and sizes of these organs.
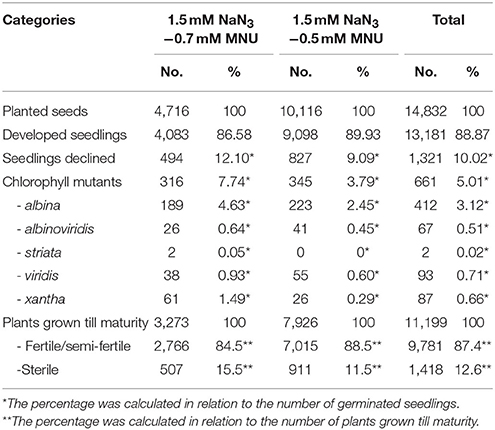
Table 2. Status of initial HorTILLUS M2 generation.
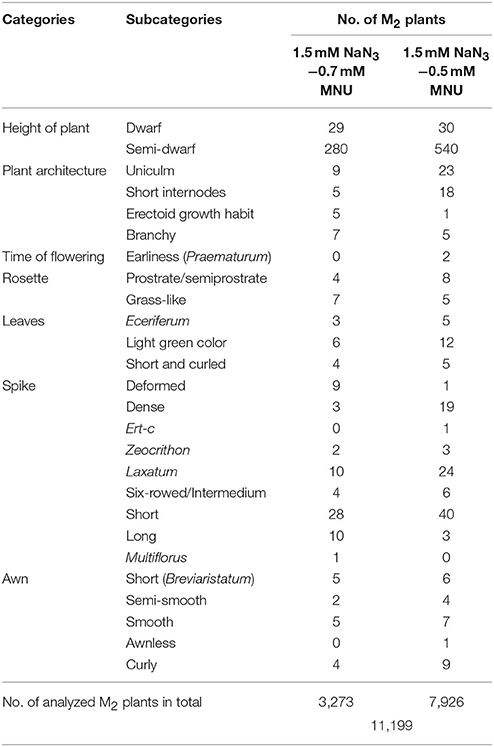
Table 3. Phenotypic changes observed in M2 generation.
We performed field observations of M3 progenies derived from 3,481 M2 plants that produced the highest amount of seeds, enough to provide a sample for M3 seed bank and M3 evaluation. Analysis of M3 families revealed that 30% of them carried morphological mutations. About one third of M3 families with morphological changes were homozygous for the observed phenotype but majority of M3 progenies segregated for one or more morphological characters (Table 4). Among these characters, alterations in plant height and growth habit were most often detected. We identified 138 homozygous M3 lines with dwarf or semi-dwarf plant height and 277 M3 families that segregated for this trait. Among changes of plant architecture, the erectoid growth type was most common (33 homozygous lines and 118 M3 families segregating for this character (Figure 2). Furthermore, forms exhibiting changes in time of flowering, color and morphology of leaves, color and shape of kernels and morphology of spikes and awns were observed in M3 generation (Figure 3). The frequency of M3 progenies with changes in these categories varied from ca.1% for early mutants to 15% for mutants with changes in plant growth type (Figure 2). To ensure a pedigree-based material for further forward mutant screens, we have harvested individually ca. 30,000 M3 plants and stored the vacuum-packed M4 seeds in refrigerator.
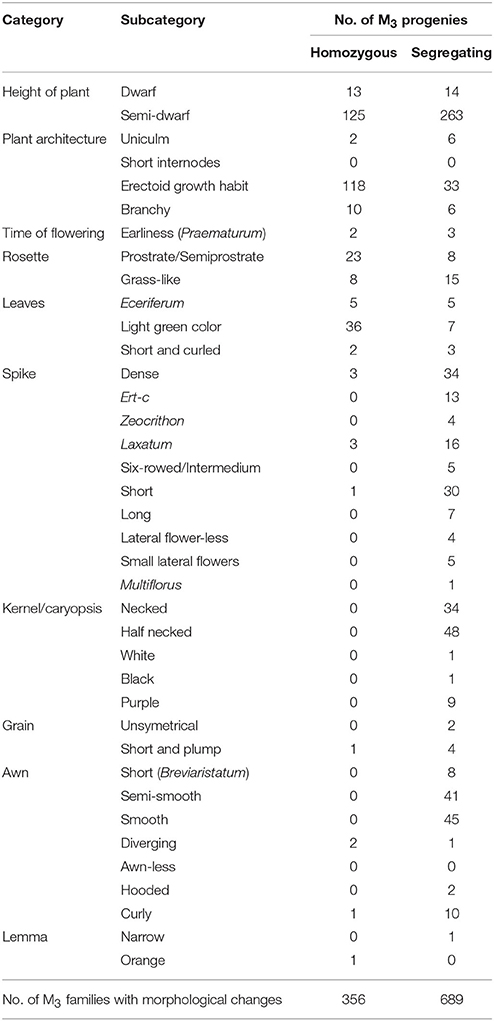
Table 4. Morphological changes analyzed in M3 progenies of 3,481 M2 plants.
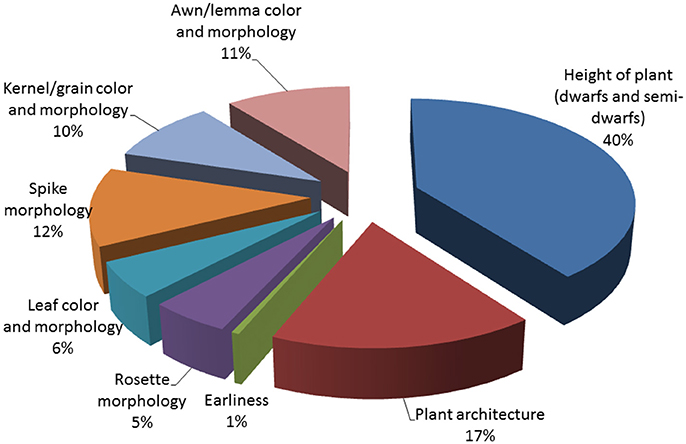
Figure 2. Types of morhpological alterations observed in M3 progenies (in %).

Figure 3. Examples of morphological changes observed in M3 HorTILLUS plants: (A) curly prostrate rosette, (B) yellow stripped rosette, (C) earliness, (D) dense spike, (E) multiflorus, (F) curly awns, (G) white husk, (H) Calcaroides, (I) awnless.
We have evaluated the usefulness of the HorTILLUS population for mutation identification based on 32 gene TILLed. The genes which were chosen for this analysis were related to different aspects of plant growth and development, such as tolerance to abiotic stresses, growth regulators metabolism, or DNA repair (Table 1). Based on the CODDLE analysis, one to three gene fragments, for which there was the highest probability that mutation would affect an encoded protein function, were used for mutation detection. For each gene fragment, 3,072–6,912 M2 plants were used for mutation identification. In total, 182.16 Mb were screened in 40 gene fragments and 382 mutations were found (Table 5). Based on these results, we estimated the average mutation density in the HorTILLUS population as 1 mutation per 477 kb. The mutation density varied from gene to gene—the highest (1/86 kb) was observed in HvRAA1 and the lowest (1/7,066 kb) in one of the HvKu70 fragments. Only in one fragment of HvPARP3 gene we have not found any mutation. On average, we have identified 11.9 mutations per gene, with a range of 4–34 mutations. The density of mutations was higher in the HorTILLUS subpopulation developed after treatment with a higher concentration of MNU (1 mutation per 414 kb) than in the subpopulation derived from treatment with a lower concentration of MNU, where 1 mutation was found per 608 kbp (Supplementary Table 2). This result is consistent with the frequency of chlorophyll mutants among M2 plants derived from these two treatments (7.7 and 3.8% after treatment with the higher and lower dose of mutagens, respectively).
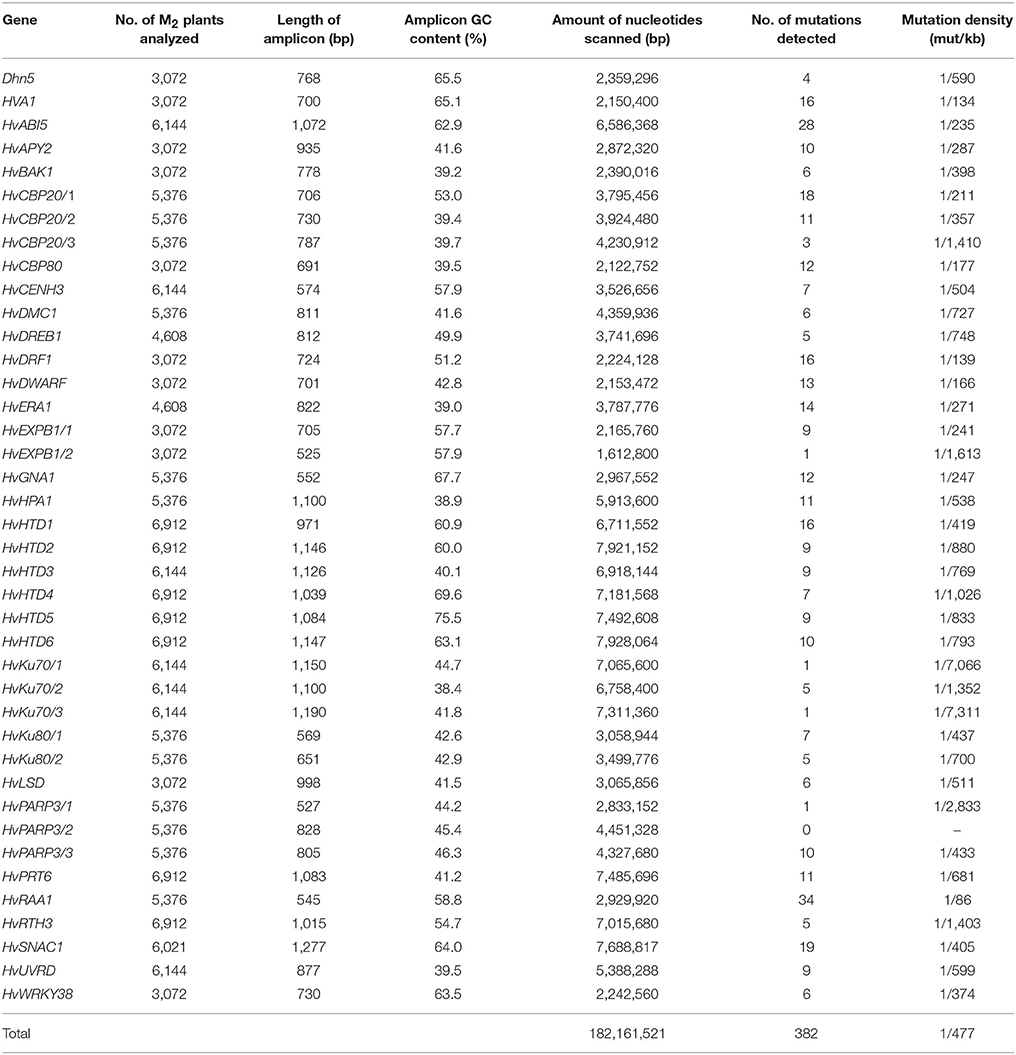
Table 5. Mutation density in HorTILLUS population based on 32 genes TILLed.
Mutagens used for creation of HorTILLUS population caused mainly G/C to A/T transitions (88% of mutations; Table 6). Among them, 60% were G>A transitions and 40% C>T transition. Among nucleotide substitutions we have also found other types of transitions—T>C and A>G (4.5% of all mutations), as well as some transversions (C>A, G>T, A>C, C>G, T>G, and G>C). The transversions accounted for 7.5% of all mutations found in HorTILLUS.
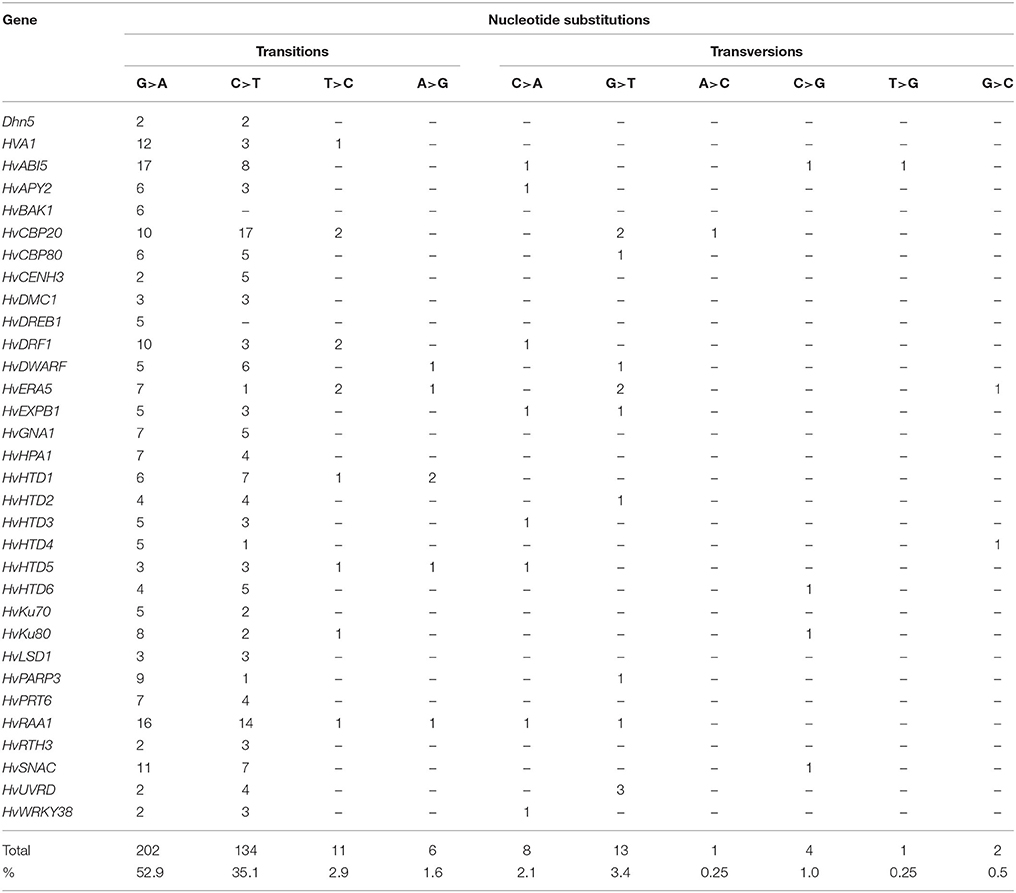
Table 6. Types of mutations found in HorTILLUS population based on 32 genes TILLed.
The available DNA sequences in which mutations were detected allowed for an investigation of whether the G/C to A/T transitions (the predominant type of mutations) are distributed randomly in barley genes or whether the local bias in the nucleotide composition surrounding the methylated guanine (O6-metG) exists. Therefore, we have examined 5′-NGN-3′ sequence context for G/C to A/T transitions in a subgroup of 12 TILLed genes (Table 7). Purines formed more than 90% of nucleotides observed at the−1 site, with predominant guanine (55.4%), whereas at position +1 there was no such a bias. The predominant local sequence context bias for G/C to A/T transitions observed in our population was 5′-RmetGN-3′.
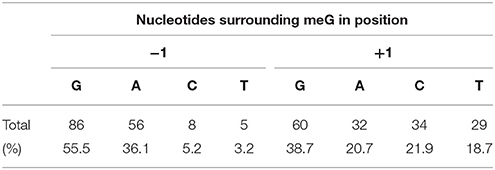
Table 7. The nucleotides flanking the mutated guanine (O6-metG) in HorTILLUS population, based on 12 genes TILLed.
Among 382 identified mutations in HorTILLUS population, 261 (68.3%) occurred in coding gene regions and 121 (31.7%) in noncoding regions, mostly introns (Supplementary Table 3). Converting these values to density of mutations gives 1 mutation per 444 kb and 1 mutation per 548 kb in the coding and noncoding sequences, respectively. Among mutations in the coding regions, 61.5% were missense, 37.5% were silent and 1% were nonsense (Table 8). Over 57% of all mutations found in the HorTILLUS population were heterozygous, while 42% were in homozygous state in M2 generation.
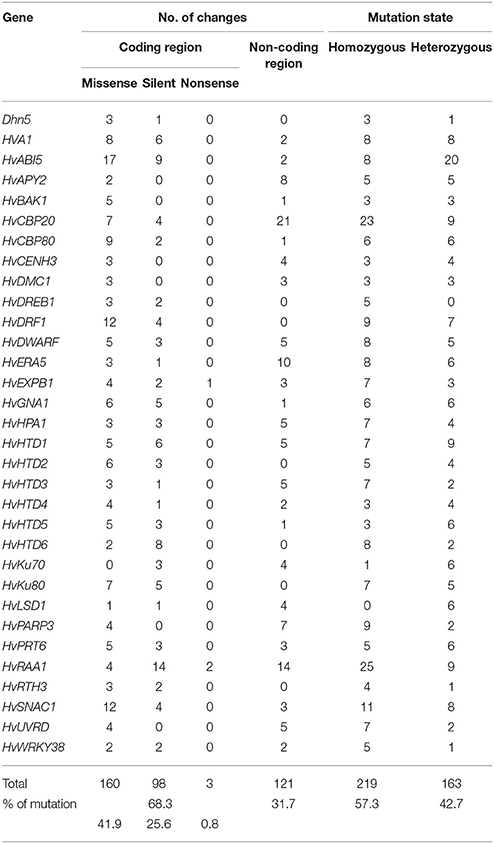
Table 8. Spectrum of mutations identified in HorTILLUS population, based on 32 genes TILLed.
The mutation density in HorTILLUS population, calculated as 1 mutation per 477 kbp, is among the highest mutation densities reported for barley. In the other barley TILLING populations created till now after chemical mutagenesis, this value ranges from 1 mutation/2,500 kb to 1 mutation/374 kb (Caldwell et al., 2004; Talamè et al., 2008, 2009; Gottwald et al., 2009; Lababidi et al., 2009; Kurowska et al., 2012; Supplementary Table 4). It should be noted, however, that the mutation density estimated in these populations was based on the analysis of only 2–11 genes that included in total 4.5–52.3 Mb of barley sequences, while we have examined 32 genes and more than 182 Mb. The mutation density in three barley populations for which more than 2 genes were analyzed (HorTILLUS, Barke, and TILLMore) does not differ significantly according to the non-parametric Anova test (Kruskal–Wallis; Supplementary Figure 3). The relatively high mutation density in HorTILLUS confirms a high efficiency of mutagenesis using a combined treatment with sodium azide and MNU (Till et al., 2007).
The mutation density in HorTILLUS varied between different gene fragments from 1/86 to 1/7,066 kbp. The reason for this phenomenon may be the local sequence context of nucleotides surrounding the methylated guanine. Guanine is the main target of methylating agents, such as EMS and MNU. Most of mutations induced by these agents arise from alkylation of guanine at the O6 position, which leads to the formation of O6-metG, the DNA lesion with the strongest mutagenic property (Kleibl, 2002). We have investigated the nucleotide composition in the relation to G/C to A/T transitions and we clearly demonstrated the strong specificity of the nucleotide surrounding the methylated guanine at the −1 position. In 90% of cases it was purine. A nonrandom distribution of mutated guanine was also shown in other studies (Richardson et al., 1987; Mironov et al., 1993; Kurowska et al., 2012), where the motif 5′-RmetGN-3′ was detected for 68–82% G/C to A/T transitions. It is thought that the base close to the lesion can strongly influence the mutagenic efficiency by the local repair deficiencies. Another reason for a wide range of mutation densities in different genes may be related to the differentiated robustness of the PCR reaction that depends on the specificity of primers or the efficiency of their association with template. Dyes attached to primers may change their 3D structure, what in some cases, may influence the efficiency of primer-DNA association. For this reason, when using labeled primers, the PCR efficiency, and thus mutation detection efficiency, could be lower for some fragments. To overcome this problem there is a possibility to use other mutation detection methods that do not require labeled primers. We have optimized the method of mutation identification with the use of Fragment Analyzer™ (Advanced Analytical Technologies) and we are applying it currently for TILLING analyses.
N-Methyl-N-nitrosourea (MNU), one of the mutagens used for development of HorTILLUS platform, was applied also in several TILLING experiments, for example in G. max (Cooper et al., 2008) and O. sativa (Suzuki et al., 2008), where it caused only G/C to A/T transitions. Treatment of barley with sodium azide (the second mutagen used in our study) also leads mostly to this kind of transitions (Talamè et al., 2008). As expected, in HorTILLUS population we found mainly G/C to A/T mutations (88%). Most of them (60%) were G to A transition, and 40% were C to T transitions. G to A transitions arise from O6 alkylation of guanine in the nontranscribed (sense) DNA strand, due to mispairing of O6-metG with thymine in the first replication cycle, which leads to its replacement by adenine in the subsequent replication round. Alkylation of guanine in the transcribed (antisense) strand results in the C to T transition. The higher proportion of G>A transitions observed in this study may indicate a slight bias in the repair of O6-metG lesions between DNA strands, with a more efficient repair of the transcribed strand. Similar bias was observed after MNU treatment in the hprt (hypoxanthine-guanine phosphoribosyltransferase) gene in rat fibroblasts (Jansen et al., 1994). In HorTILLUS population we also found other types of transitions—T to C and A to G. The T/A to C/G mutations may arise from methylation of O4 position of thymine which appears with a much lower frequency compared to O6−meG (Kleibl, 2002). In the first replication cycle, O4-metT mispairs with guanine and in the subsequent cycle it leads to the T/A to C/G transition. In our TILLING population this type of nucleotide substitutions occurred with a low frequency (4.5%). Besides transitions we found also some transversions (7.5% of all mutations). Sodium azide, used as one of the mutagens in our experiments, is known to be highly mutagenic in barley (Nilan et al., 1973) and other crop species, whereas it is marginally mutagenic in A. thaliana and mammals (Gruszka et al., 2012b). It leads mainly to transitions, but also to transversions (Olsen et al., 1993). Thus, the transversions observed in HorTILLUS population might be caused by mutagenic activity of NaN3, although MNU is known to induce a relatively high level of transversions as well (Kurowska et al., 2012).
Interestingly, the comparison of alterations in coding and noncoding regions of TILLed DNA fragments showed a slightly higher mutation density in the coding sequences (1/444 vs. 1/548 kb, respectively). The noncoding regions represented introns, and in some cases the 5′ and 3′UTR. The slightly higher mutation density in the coding regions shows a lack of preference in mutagenic action and/or repair system in relation to the type of sequences. The most useful mutations in terms of functional genetics are changes that affect protein function, mainly missense, nonsense and splice junction mutations. Based on 32 genes TILLed, the majority of mutations occurring in coding sequences in HorTILLUS population were missense (61%). We have also found three independent nonsense mutations leading to the premature STOP codon, while 98 mutations (37% of changes occurring in coding regions) were silent. These results are in agreement with the spectrum of mutations observed after EMS or sodium azide treatment in barley (Caldwell et al., 2004; Talamè et al., 2008; Gottwald et al., 2009) and clearly show that HorTILLUS population is an useful reverse genetics tool.
Most of the mutations found in M2 plants were in heterozygous state (57%). The ratio of homozygous to heterozygous mutations in M2 generation should be 1:2 (Till et al., 2006). The lower level of heterozygotes among analyzed plants with mutations could be connected with a method of mutation detection applied in the study. When using the described LI-COR method of mutation identification, signals of heterozygous mutations are by half weaker than the signals of homozygous ones in the eight-fold DNA pools. Therefore, some of heterozygous mutations could be missed during analysis of LI-COR gel images.
One of the problems often raised in regards to application of TILLING platforms as a reverse genetics tool is a high density of background mutations that may affect the plant phenotype. Based on the analysis of HorTILLUS population presented above, we estimate the average mutation density as ca. 1 mutation per 500 kb. This density gives about 10,000 mutations (nucleotide substitutions) in barley genome whose size is around 5 Gb. Taking into consideration only the coding part of barley genome, which size is estimated at 65.3 Mb (1.4%; Mascher et al., 2017), the number of mutations in coding sequences may account for ca. 130 mutations. Therefore, for functional genomics studies we recommend to overcome the problem of off target mutations by backcrossing the identified mutant to the parent variety. After two backcrosses this number will be reduced by 75%, giving about 32 background mutations left. As our analysis showed that about 40% of identified mutations are silent, this further reduces the number of off target mutations with potential effect, to about 19 in barley genome. In addition, when we work with TILLING as a reverse genetics strategy, we usually know the biological process in which a candidate gene is involved. Therefore, we analyze the identified mutant according to the phenotypic characters related to this process. To confirm that the identified mutation is really responsible for a phenotypic change in the mutant, we always perform a co-segregation analysis in F2 progeny of mutant x parent variety, and/or we evaluate a set of allelic mutants with different changes in the gene. We are aware that TILLING approach may be time and labor consuming but it is the only strategy available for plants with large genomes and/or poorly studied to produce mutants in genes of interest. The new strategy of genome editing (CRISPR-Cas9), which was reported also for barley (Lawrenson et al., 2015), is up to now restricted to one barley variety “Golden Promise,” due to difficulties in transformation of other barley cultivars. Additionally, TILLING is not a GMO strategy, which may be considered as an advantage in some countries.
HorTILLUS is one of few TILLING platforms created for barley. In Supplementary Table 4 we have summarized the characteristics of all barley TILLING populations developed till now. The HorTILLUS platform is the largest of those populations and is well characterized. It has been evaluated in terms of induced mutations in a wide range of genes located across barley genome and characterized for phenotypic changes in a large M3 population. The HorTILLUS platform has already proven its usefulness in functional genomics studies. The platform has been utilized to reveal function of genes involved in brassinosteroid and strigolactone metabolism (Gruszka et al., 2016; Marzec et al., 2016), DNA repair (Stolarek et al., 2015a,b), tolerance to abiotic stresses (Mendiondo et al., 2016; Daszkowska-Golec et al., 2017). In addition, the individual collection of ca. 30,000 M3 progenies gives a unique basis for forward screening for mutants of interest. We would like to emphasize that our HorTILLUS population is available to the barley community on a cooperative basis.
We have created HorTILLUS—the TILLING platform for barley with an average mutation density 1/477 kb, calculated on the basis of 32 gene TILLed. This platform proved to be a useful tool, both in functional genomic studies and in forward selection of barley mutants with required phenotypic changes, including agronomically important traits. A great advantage of the HorTILLUS platform, compared to the other TILLING populations in barley, is that we are constantly renewing it by replacing the M2 plants, whose DNA and/or seeds are depleted, with the new ones. This makes HorTILLUS a permanent source of mutations that can be used for many years. We offer the usage of this valuable resource to the interested barley researchers on a cooperative basis.
IS and MMal: Conceived and designed the study; JZ: Performed mutagenesis; JZ, JJ: Handled M1 and M2 generations; JJ, MS-Z: Isolated DNA and prepared pools; JZ, MS-Z, BC, MMar: Phenotyped M2 plants; MS-Z, MMar, JJ, BC, MKur, MKr, AD-G, JG-W, DG, MGaje, PG, MS, PT, PS, SL, MKud, ML, MG-W, AM, NP, KS, AK, ZM, ET: Performed mutation screening of different fragments; JZ, MGaj, MS-Z, MMar, DG, WO-J, ZN: Handled and phenotyped M3 population; JZ: Maintained seed bank; MMar: Created the database; MS-Z and IS: Wrote the manuscript.
This work was supported by Polish Ministry of Science and Higher Education PBZ-MNiSW-2/3/2006/8, European Regional Development Fund through the Innovative Economy for Poland 2007–2013, project WND-POIG.01.03.01-00-101/08 POLAPGEN-BD “Biotechnological tools for breeding cereals with increased resistance to drought,” task 22 and International Atomic Energy Agency Research Contract RC 15419 “Biotechnology Package for Enhancing Induced Mutagenesis in Barley”.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
We would like to thank Bradley Till from International Atomic Energy Agency, Vienna, for providing us a Celery Juice Extract containing Cel-1 enzyme and for many advice regarding mutation detection.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fpls.2018.00216/full#supplementary-material
Alder, A., Jamil, M., Marzorati, M., Bruno, M., Vermathen, M., Bigler, P., et al. (2012). The path from β-carotene to carlactone, a strigolactone-like plant hormone. Science 335, 1348–1351 doi: 10.1126/science.1218094
Aslam, U., Cheema, H. M., Ahmad, S., Khan, I. A., Malik, W., and Khan, A. A. (2016). COTIP: cotton TILLING platform, a resource for plant improvement and reverse genetic studies. Front. Plant. Sci. 7:1863. doi: 10.3389/fpls.2016.01863
Beier, S., Himmelbach, A., Colmsee, C., Zhang, X., Barrero, R., Zhang, Q., et al. (2017). Construction of a map-based reference genome sequence for barley, Hordeum vulgare L. Sci. Data 4:170044. doi: 10.1038/sdata.2017.44
Brini, F., Hanin, M., Lumbreras, V., Amara, I., Khoudi, H., Hassairi, A., et al. (2007). Overexpression of wheat dehydrin DHN-5 enhances tolerance to salt and osmotic stress in Arabidopsis thaliana. Plant Cell Rep. 26, 2017–2026. doi: 10.1007/s00299-007-0412-x
Caldwell, D., McCallum, N., Shaw, P., Muehlbauer, G., Marshall, D., and Waugh, R. (2004). A structured mutant population for forward and reverse genetics in barley (Hordeum vulgare L.). Plant J. 40, 143–150. doi: 10.1111/j.1365-313X.2004.02190.x
Chantreau, M., Grec, S., Gutierrez, L., Dalmais, M., Pineau, C., Demailly, H., et al. (2013). PT-flax (phenotyping and TILLinG of flax): development of a flax (Linum usitatissimum L.) mutant population and TILLinG platform for forward and reverse genetics. BMC Plant Biol. 13:159. doi: 10.1186/1471-2229-13-159
Chen, L., Huang, L., Min, D., Phillips, A., Wang, S., Madqwick, P., et al. (2012). Development and characterization of a new TILLING population of common bread wheat (Triticum aestivum L.). PLoS ONE 7:e41570. doi: 10.1371/journal.pone.0041570
Cong, L., Ran, F., Cox, D., Lin, S., Barretto, R., Habib, N., et al. (2013). Multiplex genome engineering using CRISPR/Cas systems. Science 339, 819–823. doi: 10.1126/science.1231143
Cooper, J. L., Till, B. J., Laport, R. G., Darlow, M. C., Kleffner, J. M., Jamai, A., et al. (2008). TILLING to detect induced mutations in soybean. BMC Plant Biol. 8:9. doi: 10.1186/1471-2229-8-9
Cutler, S., Ghassemian, M., Bonetta, D., Cooney, S., and McCourt, P. (1996). A protein farnesyl transferase involved in ABA signal transduction in Arabidopsis. Science 273, 1239–1241. doi: 10.1126/science.273.5279.1239
Dalmais, M., Schmidt, J., Le Signor, C., Moussy, F., Burstin, J., Savois, V., et al. (2008). UTILLdb, a Pisum sativum in silico forward and reverse genetics tool. Genome Biol. 9:R43. doi: 10.1186/gb-2008-9-2-r43
Daszkowska-Golec, A. (2017). Emerging roles of the nuclear cap-binding complex in abiotic stress responses. Plant Physiol. 176, 242–253. doi: 10.1104/pp.17.01017
Daszkowska-Golec, A., Skubacz, A., Marzec, M., Słota, M., Kurowska, M., Gajecka, M., et al. (2017). Mutation in HvCBP20 (Cap Binding Protein 20) adapts barley to drought stress at phenotypic and transcriptomic levels. Front. Plant Sci. 8:942. doi: 10.3389/fpls.2017.00942
Dawson, I., Russell, J., Powell, W., Steffenson, B., Thomas, W., and Waugh, R. (2015). Barley: a translational model for adaptation to climate change. New Phytol. 206, 913–931. doi: 10.1111/nph.13266
Doutriaux, M. P., Couteau, F., Bergounioux, C., and White, C. (1998). Isolation and characterisation of the RAD51 and DMC1 homologs from Arabidopsis thaliana. Mol. Gen. Genet. 257, 283–291. doi: 10.1007/s004380050649
Doyle, J. J., and Doyle, J. L. (1987). A rapid DNA isolation procedure for small quantities of fresh leaf tissue. Phytochem. Bull. 19, 11–15.
Fitzgerald, T. L., Kazan, K., Li, Z., Morell, M. K., and Manners, J. M. (2010). A high-throughput method for the detection of homologous gene deletions in hexaploid wheat. BMC Plant Biol. 10:264. doi: 10.1186/1471-2229-10-264
Fraenkel, R., Kovalski, I., Troadec, C., Bendahmane, A., and Perl-Treves, R. (2014). Development and evaluation of a cucumber TILLING population. BMC Res. Notes 7:846. doi: 10.1186/1756-0500-7-846
Gady, A. L., Hermans, F. W., Van de Wal, M. H., van Loo, E. N., Visser, R. G., and Bachem, C. W. (2009). Implementation of two high throughput techniques in a novel application: detecting point mutations in large EMS mutated plant populations. Plant Methods 5:13. doi: 10.1186/1746-4811-5-13
Ge, L., Chen, H., Jiang, J., Zhao, Y., Xu, M., Xu, Y., et al. (2004). Overexpression of OsRAA1 causes pleiotropic phenotypes in transgenic rice plants, including altered leaf, flower, and root development and root response to gravity. Plant Physiol. 135, 1502–1513. doi: 10.1104/pp.104.041996
Gilchrist, E., Sidebottom, C., Koh, C., MacInnes, T., Sharpe, A., and Haughn, W. (2013). A mutant Brassica napus (Canola) population for the identification of new genetic diversity via TILLING and next generation sequencing. PLoS ONE 8:e84303. doi: 10.1371/journal.pone.0084303
Gottwald, S., Bauer, P., Komatsuda, T., Lundqvist, U., and Stein, N. (2009). TILLING in the two-rowed barley cultivar “Barke” reveals preferred sites of functional diversity in the gene HvHox1. BMC Res. Notes 2:258. doi: 10.1186/1756-0500-2-258
Gruszka, D., Gorniak, M., Glodowska, E., Wierus, E., Oklestkova, J., Janeczko, A., et al. (2016). A Reverse-genetics mutational analysis of the barley HvDWARF gene results in identification of a series of alleles and mutants with short stature of various degree and disturbance in BR biosynthesis allowing a new insight into the process. Int. J. Mol. Sci. 17:600. doi: 10.3390/ijms17040600
Gruszka, D., Marzec, M., and Szarejko, I. (2012a). The barley EST DNA Replication and Repair Database (bEST-DRRD) as a tool for the identification of the genes involved in DNA replication and repair. BMC Plant Biol. 12:88. doi: 10.1186/1471-2229-12-88
Gruszka, D., Szarejko, I., and Maluszynski, M. (2011). Identification of barley DWARF gene involved in brassinosteroid synthesis. Plant Growth Regul. 65, 343–358. doi: 10.1007/s10725-011-9607-9
Gruszka, D., Szarejko, I., and Maluszynski, M. (2012b). “Sodium azide as a mutagen,” in Plant Mutation Breeding and Biotechnology, eds Q. Shu, B. P. Forster, and H. Nakagawa (Wallingford, UK: CABI Publishing House), 159–166.
Guo, B., Wei, Y., Xu, R., Lin, S., Luan, H., Lv, C., et al. (2016). Genome-wide analysis of APETALA2/Ethylene-Responsive factor (AP2/ERF) gene family in barley (Hordeum vulgare L.). PLoS ONE. doi: 10.1371/journal.pone.0161322
Hochholdinger, F., Wen, T., Zimmerman, R., Chimot-Marolle, P., da Costa e Silva, O., Bruce, W., et al. (2008). The maize (Zea mays L.) roothairless3 encodes a putative GPIanchored, monocot-specific, COBRA-like protein that significantly affects grain yield. Plant J. 54, 888–898. doi: 10.1111/j.1365-313X.2008.03459.x
Hu, H., Dai, M., Yao, J., Xiao, B., Li, X., Zhang, Q., et al. (2006). Overexpressing a NAM, ATAF, and CUC (NAC) transcription factor enhances drought resistance and salt tolerance in rice. Proc. Natl. Acad. Sci. 103, 12987–12992. doi: 10.1073/pnas.0604882103
International Barley Genome Sequencing Consortium, Mayer, K. F., Waugh, R., Brown, J. W., Schulman, A., Langridge, P., et al. (2012). A physical, genetic and functional sequence assembly of the barley genome. Nature 491, 711–716. doi: 10.1038/nature11543
Ishikawa, S., Maekawa, M., Arite, T., Onishi, K., Takamure, I., and Kyozuka, J. (2005). Suppression of tiller bud activity in tillering dwarf mutants of rice. Plant Cell Physiol. 46, 79–86. doi: 10.1093/pcp/pci022
Jankowicz-Cieslak, J., Huynh, O., Brozynska, M., Nakitandwe, J., and Till, B. (2012). Induction, rapid fixation and retention of mutations in vegetatively propagated banana. Plant Biotechnol. J. 10, 1056–1066. doi: 10.1111/j.1467-7652.2012.00733.x
Jansen, J., Mohn, G., Vrieling, H., van Taijlingen, C., Lohman, P., and van Zeeland, A. (1994). Molecular analysis of hprt gene mutations in skin fibroblast of rats exposed in vivo to N-methyl-N-nitrosourea or N-ethyl-N-nitrosourea. Cancer Res. 54, 2478–2485.
Jiang, H., Wang, S., Dang, L., Wang, S., Chen, H., Wu, Y., et al. (2005). A novel short-root gene encodes a glucosamine-6-phosphate acetyltransferase required for maintaining normal root cell shape in rice. Plant Physiol. 138, 232–242. doi: 10.1104/pp.104.058248
Keisa, A., Kanberga, K., Gill, U., Kleinhofs, A., and Rostoks, N. (2008). Cloning and characterization of barley homologues of the Arabidopsis LSD1 gene: putative regulators of hypersensitive response. Acta Univ. Latviensis 745, 87–101.
Kleibl, K. (2002). Molecular mechanisms of adaptive response to alkylating agents in Escherichia coli and some remarks on O6−methylguanine DNA-methyltransferase in other organisms. Mutat. Res. 512, 67–84. doi: 10.1016/S1383-5742(02)00025-X
Kohzuma, K., Chiba, M., Nagano, S., Anai, T., Ueda, M., Oquchi, R., et al. (2017). Mutant selection in the self-incompatible plant radish (Raphanus sativus L. var. sativus) using two-step TILLING. Breed Sci. 67, 268–276. doi: 10.1270/jsbbs.16200
Kumar, A., Boualem, A., Bhattacharya, A., Parikh, S., Desai, N., Zambelli, A., et al. (2013). SMART – Sunflower Mutant population and Reverse genetic Tool for crop improvement. BMC Plant Biol. 13:38. doi: 10.1186/1471-2229-13-38
Kurowska, M., Daszkowska-Golec, A., Gruszka, D., Marzec, M., Szurman, M., Szarejko, I., et al. (2011). TILLING - a shortcut in functional genomics. J. Appl. Genet. 52:317. doi: 10.1007/s13353-011-0061-1
Kurowska, M., Labocha-Pawlowska, A., Gnizda, D., Maluszynski, M., and Szarejko, I. (2012). Molecular analysis of point mutations in a barley genome exposed to MNU and gamma rays. Mutat. Res. 738–739, 52–70. doi: 10.1016/j.mrfmmm.2012.08.008
Kwaśniewski, M., and Szarejko, I. (2006). Molecular cloning and characterization of β-expansin gene related to root hair formation in barley. Plant Physiol. 141, 1149–1158. doi: 10.1104/pp.106.078626
Lababidi, S., Mejlhede, N., Rasmussen, S., Backes, G., Al-Said, W., Baum, M., et al. (2009). Identification of barley mutants in the cultivar “Lux” at the Dhn loci through TILLING. Plant Breed. 128, 332–336. doi: 10.1111/j.1439-0523.2009.01640.x
Lawrenson, T., Shorinola, O., Stacey, N., Li, C., Østergaard, L., Patron, N., et al. (2015). Induction of targeted, heritable mutations in barley and Brassica oleracea using RNA-guided Cas9 nuclease. Genome Biol. 16:258. doi: 10.1186/s13059-015-0826-7
Le Signor, C., Savois, V., Aubert, G., Verdier, J., Nicolas, M., Pagny, G., et al. (2009). Optimizing TILLING populations for reverse genetics in Medicago truncatula. Plant Biotechnol. J. 7, 430–441. doi: 10.1111/j.1467-7652.2009.00410.x
Manmathan, H., Shaner, D., Snelling, J., Tisserat, N., and Lapitan, N. (2013). Virus-induced gene silencing of Arabidopsis thaliana gene homologues in wheat identifies genes conferring improved drought tolerance. J. Exp. Bot. 64, 1381–1392. doi: 10.1093/jxb/ert003
Manova, V., and Gruszka, D. (2015). DNA damage and repair in plants – from models to crops. Front. Plant Sci. 6:885. doi: 10.3389/fpls.2015.00885
Marè, C., Mazzucotelli, E., Crosatti, C., Francia, E., Stanca, A., and Cattivelli, L. (2004). Hv-WRKY38: a new transcription factor involved in cold- and drought-response in barley. Plant Mol. Biol. 55, 399–416. doi: 10.1007/s11103-004-0906-7
Marzec, M., Gruszka, D., Tylec, P., and Szarejko, I. (2016). Identification and functional analysis of the HvD14 gene involved in strigolactone signaling in Hordeum vulgare. Physiol. Plant. 158, 341–355. doi: 10.1111/ppl.12460
Mascher, M., Gundlach, H., Himmelbach, A., Beier, A., Twardziok, S., Wicker, T., et al. (2017). A chromosome conformation capture ordered sequence of the barley genome. Nature 544, 427–433. doi: 10.1038/nature22043
McCallum, C., Comai, L., Greene, E., and Henikoff, S. (2000). Targeted screening for induced mutations. Nat. Biotechnol. 18, 455–457. doi: 10.1038/74542
Mendiondo, G., Gibbs, D., Szurman-Zubrzycka, M., Korn, A., Marquez, J., Szarejko, I., et al. (2016). Enhanced waterlogging tolerance in barley by manipulation of expression of the N-end rule pathway E3 ligase PROTEOLYSIS6. Plant Biotechnol. J. 14, 40–50. doi: 10.1111/pbi.12334
Minoia, S., Petrozza, A., D'Onofrio, O., Piron, F., Mosca, G., Sozio, G., et al. (2010). A new mutant genetic resource for tomato crop improvement by TILLING technology. BMC Res. Notes 3:69. doi: 10.1186/1756-0500-3-69
Mironov, N. M., Bleicher, F., Martel-Planche, G., and Montesano, R. (1993). Nonrandom distribution of O6-methylguanine in H-ras gene sequence from DNA modified with N-methyl-N-nirosourea. Mutat. Res. 288, 197–205. doi: 10.1016/0027-5107(93)90085-T
Mo, X., Zhu, Q., Li, X., Li, J., Zeng, Q., Rong, H., et al. (2006). The hpa1 mutant of Arabidopsis reveals a crucial role of histidine homeostasis in root meristem maintenance. Plant Physiol. 141, 1425–1435. doi: 10.1104/pp.106.084178
Nilan, R., Sideris, E., Kleinhofs, A., Sander, C., and Konzak, C. (1973). Azide—a potent mutagen. Mut. Res. 17, 142–144.
Ogata, T., Nagatoshi, Y., Yamagishi, N., Yoshikawa, N., and Fujita, Y. (2017). Virus-induced down-regulation of GmERA1A and GmERA1B genes enhances the stomatal response to abscisic acid and drought resistance in soybean. PLoS ONE 12:e0175650. doi: 10.1371/journal.pone.0175650
Okabe, Y., Asamizu, E., Saito, T., Matsukura, C., Ariizumi, T., Brès, C., et al. (2011). Tomato TILLING technology: development of a reverse genetics tool for the efficient isolation of mutants from Micro-Tom mutant libraries. Plant Cell Physiol. 52, 1994–2005. doi: 10.1093/pcp/pcr134
Olsen, O., Wang, X., and von Wettstein, D. (1993). Sodium azide mutagenesis: preferential generation of A-T G-C transitions in the barley Ant18 gene. Proc. Nat. Acad. Sci. U.S.A. 90, 8043–8047.
Perry, J., Brachmann, A., Welham, T., Binder, A., Charpentier, M., Groth, M., et al. (2009). TILLING in Lotus japonicus identified large allelic series for symbiosis genes and revealed a bias in functionally defective ethyl methanesulfonate alleles toward glycine replacements. Plant Physiol. 151, 1281–1291. doi: 10.1104/pp.109.142190
Piron, F., Nicolai, M., Minoia, S., Piednoir, E., Moretti, A., Salgues, A., et al. (2010). An induced mutation in tomato eIF4E leads to immunity to two potyviruses. PLoS ONE 5:e11313. doi: 10.1371/journal.pone.0011313
Porch, T., Blair, M., Lariguet, P., Galeano, C., Pankhurst, C., and Broughton, W. (2009). Generation of a mutant population for TILLING common bean genotype Bat 93. J. Am. Soc. Hortic. Sci. 134, 348–355.
Ravi, M., Shibata, F., Ramahi, J. S., Nagaki, K., Chen, C., Murata, M., et al. (2011). Meiosis-specific loading of the centromere-specific histone CENH3 in Arabidopsis thaliana. PLoS Genet. 7:e1002121. doi: 10.1371/journal.pgen.1002121
Rawat, N., Sehgal, S., Joshi, A., Rothe, N., Wilson, D., McGraw, N., et al. (2012). A diploid wheat TILLING resource for wheat functional genomics. BMC Plant Biol. 12:205. doi: 10.1186/1471-2229-12-205
Reddy, T., Dwivedi, S., and Sharma, N. (2012). Development of TILLING by sequencing platform towards enhanced leaf yield in tobacco. Ind. Crops Prod. 40, 324–335. doi: 10.1016/j.indcrop.2012.03.031
Richardson, K. K., Richardson, F. C., Croby, R. M., Swenberg, J. A., and Skopek, T. R. (1987). DNA base changes and alkilation following in vivo exposure of Escherichia coli to N-methyl-N-nitrosourea or N-ethyl-N-nitrosourea. Proc. Natl. Acad. Sci. U.S.A. 84, 344–348. doi: 10.1073/pnas.84.2.344
Skubacz, A., Daszkowska-Golec, A., and Szarejko, I. (2016). The role and regulation of ABI5 (ABA-Insensitive 5) in plant development, abiotic stress responses and Phytohormone Crosstalk. Front. Plant Sci. 7:1884. doi: 10.3389/fpls.2016.01884
Slade, A. J., Fuerstenberg, S. I., Loeffler, D., Steine, M. N., and Facciotti, D. (2005). A reverse genetic, nontransgenic approach to wheat crop improvement by TILLING. Nat. Biotechnol. 23, 75–81. doi: 10.1038/nbt1043
Smits, B. M., Mudde, J., Plasterk, R. H., and Cuppen, E. (2004). Target-selected mutagenesis of the rat. Genomics 83, 332–334. doi: 10.1016/j.ygeno.2003.08.010
Stolarek, M., Gruszka, D., Braszewska-Zalewska, A., and Maluszynski, M. (2015a). Alleles of newly identified barley geneHvPARP3 exhibit changes inefficiency of DNA repair. DNA Repair 28, 116–130. doi: 10.1016/j.dnarep.2015.02.018
Stolarek, M., Gruszka, D., Braszewska-Zalewska, A., and Maluszynski, M. (2015b). Functional analysis of the new barley gene HvKu80 indicates that it plays a key role in double-strand DNA break repair and telomere length regulation. Mutagenesis 30, 785–797. doi: 10.1093/mutage/gev033
Straub, P., Shen, Q., and Ho, T. D. (1994). Structure and promoter analysis of an ABA- and stress-regulated barley gene, HVA1. Plant Mol. Biol. 26, 617–630. doi: 10.1007/BF00013748
Suzuki, T., Eiguchi, M., Kumamaru, T., Satoh, H., Matsusaka, H., Moriguchi, K., et al. (2008). MNU-induced mutant pools and high performance TILLING enable finding of any gene mutation in rice. Mol. Genet. Genomics 279, 213–223. doi: 10.1007/s00438-007-0293-2
Szarejko, I., and Maluszynski, M. (1999). High frequency of mutations after mutagenic treatment of barley seeds with NaN3 and MNH with application of inter-incubation germination period. Mutat. Breed. Newslett. 44, 28–30.
Szarejko, I., Szurman-Zubrzycka, M., Nawrot, M., Marzec, M., Gruszka, D., Kurowska, M., et al. (2017). “Creation of a TILLING population in barley after chemical mutagenesis with sodium azide and MNU,” in Biotechnologies for Plant Mutation Breeding, eds J. Jankowicz-Cieslak, T. H. Tai, J. Kumlehn, and B. J. Till (Springer International Publishing AG), 91–111.
Tadele, Z. (2016). Mutagenesis and TILLING to dissect gene function in plants. Curr. Genomics 17, 499–508. doi: 10.2174/1389202917666160520104158
Talamè, V., Bovina, R., Salvi, S., Sanguineti, M., Piffanelli, P., Lundquist, U., et al. (2009). “TILLING with TILLMore”, in Induced Plant Mutations in the Genomics Era, ed Q. Y. Shu (Rome: FAO), 240–242.
Talamè, V., Bovina, R., Sanguineti, M., Tuberosa, R., Lundqvist, U., and Salvi, S. (2008). TILLMore, a resource for the discovery of chemically induced mutations in barley. Plant Biotechnol. J. 6, 477–485. doi: 10.1111/j.1467-7652.2008.00341.x
Till, B. J., Cooper, J., Tai, T. H., Colowit, P., Greene, E. A., Henikoff, S., et al. (2007). Discovery of chemically induced mutations in rice by TILLING. BMC Plant Biol. 7:19. doi: 10.1186/1471-2229-7-19
Till, B. J., Reynolds, S. H., Weil, C., Springer, N., Burtner, C., Young, K., et al. (2004). Discovery of induced point mutations in maize genes by TILLING. BMC Plant Biol. 4:12. doi: 10.1186/1471-2229-4-12
Till, B. J., Zerr, T., Comai, L., and Henikoff, S. (2006). A protocol for TILLING and Ecotilling in plants and animals. Nat. Protoc. 1, 2465–2477. doi: 10.1038/nprot.2006.329
Till, B., Reynolds, S., Greene, E., Codomo, C., Enns, L., Johnson, J., et al. (2003). Large-scale discovery of induced point mutations with high-throughput TILLING. Genome Res. 13, 524–530. doi: 10.1101/gr.977903
Uauy, C., Paraiso, F., Colasuonno, P., Tran, R. K., Tsai, H., Berardi, S., et al. (2009). A modified TILLING approach to detect induced mutations in tetraploid and hexaploid wheat. BMC Plant Biol. 9:115. doi: 10.1186/1471-2229-9-115
Vicente-Dólera, N., Troadec, C., Moya, M., del Río-Celestino, M., Pomares-Viciana, T., Bendahmane, A., et al. (2014). First TILLING Platform in Cucurbita pepo: a new mutant resource for gene function and crop improvement. PLoS ONE 9:e112743. doi: 10.1371/journal.pone.0112743
Wang, Y., Ying, J., Kuzma, M., Chalifoux, M., Sample, A., McArthur, C., et al. (2005). Molecular tailoring of farnesylation for plant drought tolerance and yield protection. Plant J. 43, 413–424. doi: 10.1111/j.1365-313X.2005.02463.x
Wienholds, E., van Eeden, F., Kosters, M., Mudde, J., Plasterk, R. H. A., and Cuppen, E. (2003). Efficient target-selected mutagenesis in zebrafish genome. Genome Res. 13, 2700–2707. doi: 10.1101/gr.1725103
Winkler, S., Schwabedissen, A., Backasch, D., Bökel, C., Seidel, C., Bönisch, S., et al. (2005). Target-selected mutant screen by TILLING in Drosophila. Genome Res. 15, 718–723. doi: 10.1101/gr.3721805
Xin, Z., Wang, M. L., Barkley, N. A., Burow, G., Franks, C., Pederson, G., et al. (2008). Applying genotyping (TILLING) and phenotyping analyses to elucidate gene function in a chemically induced sorghum mutant population. BMC Plant Biol. 8:103. doi: 10.1186/1471-2229-8-103
Xue, G., and Loveridge, C. (2004). HvDRF1 is involved in abscisic acid-mediated gene regulation in barley and produces two forms of AP2 transcriptional activators, interacting preferably with a CT-rich element. Plant J. 37, 326–339. doi: 10.1046/j.1365-313x.2003.01963.x
Yuo, T., Toyota, M., Ichii, M., and Taketa, S. (2009). Molecular cloning of a root hairless gene rth1 in rice. Breed. Sci. 59, 13–20. doi: 10.1270/jsbbs.59.13
Keywords: TILLING, reverse genetics, mutation, barley, MNU, sodium azide
Citation: Szurman-Zubrzycka ME, Zbieszczyk J, Marzec M, Jelonek J, Chmielewska B, Kurowska MM, Krok M, Daszkowska-Golec A, Guzy-Wrobelska J, Gruszka D, Gajecka M, Gajewska P, Stolarek M, Tylec P, Sega P, Lip S, Kudełko M, Lorek M, Gorniak-Walas M, Malolepszy A, Podsiadlo N, Szyrajew KP, Keisa A, Mbambo Z, Todorowska E, Gaj M, Nita Z, Orlowska-Job W, Maluszynski M and Szarejko I (2018) HorTILLUS—A Rich and Renewable Source of Induced Mutations for Forward/Reverse Genetics and Pre-breeding Programs in Barley (Hordeum vulgare L.). Front. Plant Sci. 9:216. doi: 10.3389/fpls.2018.00216
Received: 31 December 2017; Accepted: 05 February 2018;
Published: 21 February 2018.
Edited by:
Apichart Vanavichit, Kasetsart University, ThailandReviewed by:
Guoxiong Chen, Chinese Academy of Sciences, ChinaCopyright © 2018 Szurman-Zubrzycka, Zbieszczyk, Marzec, Jelonek, Chmielewska, Kurowska, Krok, Daszkowska-Golec, Guzy-Wrobelska, Gruszka, Gajecka, Gajewska, Stolarek, Tylec, Sega, Lip, Kudełko, Lorek, Gorniak-Walas, Malolepszy, Podsiadlo, Szyrajew, Keisa, Mbambo, Todorowska, Gaj, Nita, Orlowska-Job, Maluszynski and Szarejko. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Iwona Szarejko, aXdvbmEuc3phcmVqa29AdXMuZWR1LnBs
†Present Address: Anna Malolepszy, Plant Molecular and Cellular Biology Laboratory, Salk Institute for Biological Studies, La Jolla, CA, United States
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.