 Irene Serrano
Irene Serrano Laura Campos
Laura Campos
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
MINI REVIEW article
Front. Plant Sci. , 08 February 2018
Sec. Plant Physiology
Volume 9 - 2018 | https://doi.org/10.3389/fpls.2018.00139
This article is part of the Research Topic Post-Translational Modifications in Plant Nuclear Signaling: Novel Insights into Responses to Environmental Changes View all 9 articles
Ubiquitination, the reversible protein conjugation with ubiquitin (Ub), is a post-translational modification that enables rapid and specific cellular responses to stimuli without requirement of de novo protein synthesis. Although ubiquitination also displays non-proteolytic functions, it often acts as a signal for selective protein degradation through the ubiquitin-proteasome system (UPS). In plants, it has become increasingly apparent that the UPS is a central regulator of many key cellular and physiological processes, including responses to biotic and abiotic stresses. In the nucleus, protein regulation via the UPS orchestrates gene expression, genome maintenance, and signal transduction. Here, we focus on E3 Ub-ligase proteins as major components of the ubiquitination cascade that confer specificity of substrate recognition. We provide an overview on how they contribute to nuclear proteome plasticity during plant responses to environmental stress signals.
Modulation of protein activity by post-translational modifications (PTMs) enables rapid and specific cellular responses to stimuli without the requirement of energy-consuming de novo protein synthesis. Ubiquitination regulates a plethora of cellular functions, from growth and development to responses to biotic and abiotic stimuli (Moon, 2004; Vierstra, 2009). This PTM involves the covalent attachment of one or more ubiquitin (Ub) proteins to a Lys (K) residue within specific target proteins through a stepwise cascade involving three enzymes: E1 (Ub-activating), E2 (Ub-conjugating), and E3 (Ub-ligase) (Vierstra, 2009). Ubiquitination confers different fates to the ubiquitinated protein depending on the number and location of the attached Ub molecules (Sadowski et al., 2012). Polyubiquitination via K48 Ub linkage is the best characterized form of ubiquitination and results in selective protein degradation through the ubiquitin-proteasome system (UPS) (Vierstra, 2009; Sadanandom et al., 2012).
The importance of the UPS in plants is exemplified by the fact that approximately 6% of the anticipated gene loci of the Arabidopsis thaliana genome (about 1600 genes) is involved in UPS-related functions (Mazzucotelli et al., 2006; Vierstra, 2009). Most of them encode putative E3 Ub-ligases that determine substrate specificity and thus are the most studied components of the ubiquitination cascade (Callis, 2014). E3 Ub-ligases can be classified into four main subfamilies [Homologous to E6-associated protein Carboxyl Terminus (HECT), Really Interesting New Gene (RING), U-box, and Cullin-RING ligases (CRLs)], depending on their subunit composition and mode of action (Vierstra, 2009). RING and U-box proteins are able to directly transfer Ub from Ub-E2 intermediates to the target (Stone, 2005; Yee and Goring, 2009). RING-type E3 Ub-ligases can also act as part of a multi-subunit CRL complex such as the SCF (Skp1, Cullin, F-box) complex in which the Cullin (CUL) protein acts as a molecular scaffold bringing together the RING protein (that binds to the E2) and the F-box protein (that directs substrate recognition) (Hua and Vierstra, 2011). In other CRL complexes, the CUL protein binds to “Bric a brac, Tramtrack and Broad Complex” (BTB) or UV-Damaged DNA-Binding Protein1 (DDB1) domain proteins that act as substrate-specific adaptors or scaffold proteins, respectively (Mazzucotelli et al., 2006). The UPS and, more particularly, E3 Ub-ligases have been shown to play a crucial role in the regulation of many steps of plant immune signaling (Marino et al., 2012). Indeed, ubiquitination modulates perception of pathogen-associated molecular patterns (PAMPs) by plasma membrane-associated receptors during PAMP-triggered immunity (PTI) and regulates the accumulation of intracellular immune receptors to prevent the constitutive activation of effector-triggered immunity (ETI), which is often associated with hypersensitive cell death (HR) at the sites of infection (Furlan et al., 2012; Li et al., 2014). In addition, E3 Ub-ligases are also involved in the regulation of signaling responses downstream of pathogen perception through, for instance, targeting vesicle trafficking components or nuclear proteins, such as transcription factors (TFs) (Zhou and Zeng, 2016).
Plant ability to survive abiotic stresses, including salinity, drought, cold, heat, radiation, and nutrient availability also relies on proteome plasticity. The UPS plays a central role in the efficient perception of these environmental stresses, as well as in the downstream responses, by suppressing stress signaling pathways during favorable growth conditions, eliminating negative regulators of signaling responses to stimulus or attenuating the signaling pathway once the stress has ceased (Lyzenga and Stone, 2012; Stone, 2014).
Beyond the importance of ubiquitination in controlling cytosolic proteins, the regulation of the nuclear machinery via the UPS is emerging as a crucial component of plant responses to biotic and abiotic stresses. Here we review the control of the homeostasis of nuclear proteins, especially the TFs, by the UPS, and its impact on plant adaptation to environmental signals. Due to space limitations, we focus on E3 Ub-ligases for which the nuclear target protein has been identified (Table 1). Finally, although hormones play a major role in the regulation of plant responses to environmental signals, we refer the reader to various reviews covering the extensively characterized role of SCF complexes in the transcriptional control of hormone signaling (Trujillo and Shirasu, 2010; Robert-Seilaniantz et al., 2011; Nagels Durand et al., 2016; Nguyen et al., 2016; Yang et al., 2016) and discuss here examples of non SCF-related hormone responses.
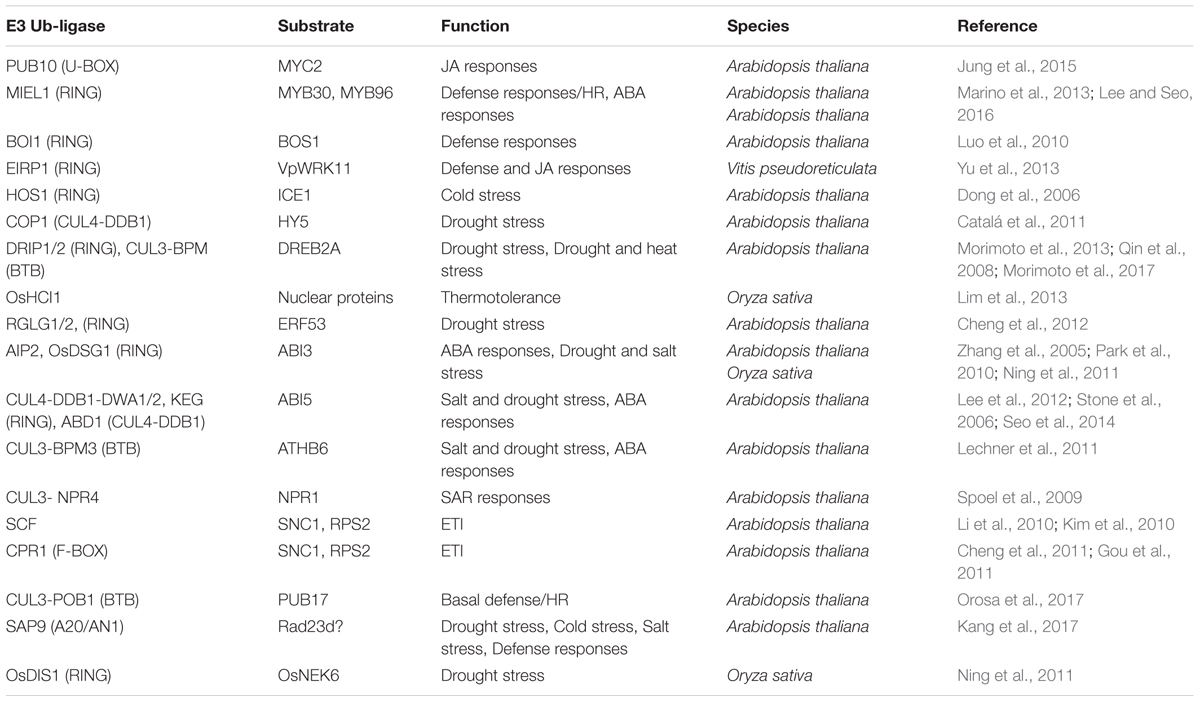
TABLE 1. Plant E3-Ub ligases with known nuclear targets and their function.
Extensive research has uncovered TFs as major nuclear proteins targeted by UPS activity during plant–pathogen interactions. The TF MYC2 acts as a master regulator that integrates multiple signals to coordinate plant defense and development through repression and activation of jasmonic acid (JA)/ethylene (ET)- and JA/wound-responsive gene expression, respectively (Abe, 2002; Boter, 2004; Lorenzo, 2004). The E3 Ub-ligase PUB10 interacts with and ubiquitinates MYC2 in the nucleus, targeting MYC2 for proteasomal degradation (Jung et al., 2015). The importance of this regulatory mode has been underlined in a recent report showing that polyubiquitinated MYC2 is deubiquitinated by nuclear UBIQUITIN PROTEASE12 (UBP12) and UBP13, thus counteracting PUB10 activity (Jeong et al., 2017). In agreement, roots of pub10 mutant seedlings and those of seedlings overexpressing MYC2, UBP12, or UBP13 displayed a similar phenotype and were hypersensitive to methyl-jasmonate, whereas seedlings overexpressing PUB10 and myc2, ubp12, or ubp13 mutant seedlings were hyposensitive (Jung et al., 2015). The role of PUB10, UBP12, or UBP13 in the MYC2-mediated plant response to pathogens was not tested in these reports. However, given that (i) MYC2 represses defense-related JA/ET-responsive genes and that (ii) UBP12 and UBP13 negatively regulate plant immunity (Ewan et al., 2011), it is tempting to speculate that these UPS-related proteins additionally modulate MYC2-dependent defense responses.
We previously identified the RING-type E3 Ub-ligase MIEL1 as being able to interact with the Arabidopsis defense-activating TF MYB30. This interaction leads to MYB30 proteasomal degradation and thus to the attenuation of Arabidopsis defense and HR responses (Marino et al., 2013). MYB30 positively regulates plant resistance by promoting the biosynthesis of very long chain fatty acids (VLCFAs). Coherently, miel1 mutant plants displayed enhanced VLCFA-related gene expression and enhanced resistance responses after inoculation with bacteria (Marino et al., 2013). MIEL1 expression is rapidly downregulated after bacterial infection, which allows MYB30 accumulation thus promoting defense and HR (Marino et al., 2013).
The RING-type E3 Ub-ligase BOI1 interacts with the Arabidopsis defense-related MYB TF BOS1 in yeast and in planta (Luo et al., 2010). BOI1 ubiquitinates BOS1 in vitro and the proteasome-dependent degradation of BOS1 was shown in Arabidopsis (Luo et al., 2010). In agreement, co-regulation of the expression of BOI1 and BOS1 in response to fungal treatment or cell death-promoting conditions suggests a functional link between the two proteins. However, no evidence that BOI1 targets BOS1 for proteasomal degradation in vivo has been reported and both bos1 mutant and BOI1 RNAi Arabidopsis plants (in which the BOS1 protein should accumulate) display enhanced susceptibility to fungal infection (Mengiste, 2003; Luo et al., 2010). Thus, the molecular details behind the BOS1-BOI1 association remain unclear.
The RING-type E3 Ub-ligase EIRP1 promotes defense in Chinese wild grapevine (Vitis pseudoreticulata) by interacting with the nuclear TF VpWRKY11. VpWRKY11 activates the expression of JA-responsive genes that negatively regulate resistance to fungal infection (Yu et al., 2013). Thus, EIRP1-mediated proteasomal degradation of VpWRKY11 results in reduced JA-responsive signaling and enhanced resistance to the fungus (Yu et al., 2013).
In addition to the regulatory roles during plant–pathogen interactions, the UPS, and more particularly E3 Ub-ligases, play a prevalent role in the control of TF levels during plant responses to adverse environmental conditions. Plant responses to cold stress rely on the activation of CBF proteins and their downstream genes by the TF ICE1. The RING-type E3 Ub-ligase HOS1 interacts with ICE1 in the nucleus and triggers its ubiquitination and degradation. In agreement with HOS1 being a negative regulator of freezing tolerance, HOS1 overexpression in Arabidopsis increases sensitivity to freezing stress (Dong et al., 2006).
The Arabidopsis TF HY5 regulates crucial processes including photomorphogenesis, responses to hormones and freezing tolerance (Lau and Deng, 2010; Catalá et al., 2011). Low temperatures induce HY5 expression independently of CBF TFs, and the HY5 protein is stabilized through nuclear depletion of the RING E3 Ub-ligase COP1 within a CUL4-DDB1 complex, which results in anthocyanin accumulation and cold acclimation (Catalá et al., 2011). Interestingly, the UPS additionally controls HY5 accumulation through its interaction with the small prefolding protein PFD4 that accumulates in the nucleus in response to low temperature in a process that requires DELLA proteins (Perea-Resa et al., 2017). PFD4 attenuates anthocyanin biosynthesis during cold acclimation by promoting COP1-independent HY5 ubiquitination and proteasomal degradation (Perea-Resa et al., 2017).
The rice RING-type E3 Ub-ligase-encoding OsHCI1 gene is induced by heat and cold treatments. Overexpression of OsHCI1 in Arabidopsis confers enhanced heat tolerance (Lim et al., 2013). Under high temperatures, OsHCI1 moves along the cytoskeleton from Golgi vesicles to the nucleus where it induces translocation of substrate nuclear proteins to the cytoplasm by monoubiquitination (Lim et al., 2013). This work highlights the importance of nucleo-cytoplasmic partitioning of E3 Ub-ligases in the regulation of plant stress responses. The TF DREB2A has been characterized as a positive regulator of drought-responsive genes by controlling the expression of water deficit-inducible genes (Sakuma, 2006; Sakuma et al., 2006). The RING E3 Ub-ligases DRIP1 and DRIP2 interact with DREB2A in the nucleus and mediate its ubiquitination and degradation via the UPS. In response to dehydration, Arabidopsis drip1drip2 double mutant plants present significantly higher survival rates, compared to wild-type plants under drought conditions (Qin et al., 2008; Morimoto et al., 2013). It was thus proposed that DRIP1/DRIP2-mediated DREB2A ubiquitination and degradation occurs under normal growth conditions. Under stress, proteasomal degradation of DREB2A would be blocked leading to its accumulation and the activation of the expression of drought-responsive genes (Qin et al., 2008). A recent report identified a 30-amino acid domain in DREB2A that mediates its nuclear interaction with BTB-MATH proteins (BPMs). BPMs are substrate-recruiting components of CUL3-based E3 Ub-ligases that induce DREB2A degradation and modulate drought and heat stress responses through regulation of DREB2A stability (Morimoto et al., 2017).
An additional Arabidopsis TF involved in the regulation of drought responses is ERF53, whose expression is induced by drought. ERF53 interacts with the RING E3 Ub-ligases RGLG1 and RGLG2. RGLG2 localizes at the plasma membrane but under drought stress translocates to the nucleus, where it interacts with ERF53 to mediate its degradation. In agreement with RGLG2 being a negative regulator of drought stress responses, the rglg1rglg2 double mutant exhibits drought tolerance (Cheng et al., 2012). These results show that, as DRIP1/2, RGLG1/2 negatively regulate drought stress responses in Arabidopsis.
The hormone abscisic acid (ABA) plays a key role in plant adaptation to abiotic stress by modulating physiological processes, such as seed dormancy and germination, seedling growth and stomatal aperture (Lopez-Molina et al., 2001; Finkelstein et al., 2005). Ubiquitination through CRLs, RING-type and U-box E3 Ub-ligases has been shown to regulate ABA signaling (Yu et al., 2015; Yang et al., 2016). The RING E3 Ub-ligase AIP2 negatively regulates the TF ABI3, a central regulator of ABA signaling (Zhang et al., 2005). In rice, OsDSG1 has been characterized as an AIP2 ortholog. Mutations in Osdsg1 increase rice tolerance to drought and high salinity (Park et al., 2010; Ning et al., 2011). AFP, an ABI5-interacting protein, attenuates ABA signaling by facilitating ABI5 proteasomal degradation in nuclear bodies (Lopez-Molina, 2003) and the 26S proteasome subunit RPN10 targets ABI5 for degradation (Smalle et al., 2003). In addition, the RING E3 Ub-ligase KEEP ON GOING (KEG) negatively regulates ABI5 accumulation, confirming the direct regulation of ABI5 by the UPS (Stone et al., 2006). In the presence of ABA, induction of ABI5 expression and KEG degradation result in increased ABI5 levels. Besides RING proteins, CRL-type E3 Ub-ligases also contribute to ABA signaling during abiotic stress responses. For example, CUL4-DDB1-based E3 Ub-ligases utilize DWA1 and DWA2 as substrate receptors and act as negative regulators of ABI5 abundance in the nucleus (Lee et al., 2012). In agreement, dwa1, dwa2, and cul4 mutants are ABA- and salt-hypersensitive and display enhanced salt-, drought-, and ABA-responsive gene expression after ABA treatment (Lee et al., 2012). ABD1 is another CUL4-DDB1-associated protein able to interact with ABI5 in the nucleus and leading to its degradation (Seo et al., 2014). Thus, abd1 mutants display ABA hypersensitivity, reduced stomatal aperture and water loss and, thus, improved tolerance to drought stress. In addition, the DDA1 protein is part of the substrate adaptor module of CUL4-DDB1-based E3 Ub-ligases and interacts with the ABA receptor PYL8 in the nucleus leading to its degradation (Irigoyen et al., 2014). Finally, the BTB-MATH protein BPM3 downregulates the abundance of the TF ATHB6 that negatively regulates drought responses in Arabidopsis (Söderman et al., 1999; Himmelbach et al., 2002; Lechner et al., 2011). As previously shown for ABI5 (Lopez-Molina et al., 2001), ABA negatively regulates ATHB6 protein turnover. ATHB6 overexpression and BPM silencing results in ABA insensitivity, enhanced water loss, increased transpiration and larger stomatal aperture, which correlates with attenuated ABA-dependent induction of salt-, drought-, and ABA-responsive genes (Lechner et al., 2011).
Besides its role in the control of plant defense, the RING-type E3 Ub-ligase MIEL1 regulates abiotic stresses in a ABA-dependent manner by promoting the degradation of the TF MYB96 (Lee and Seo, 2016). In the absence of ABA, MIEL1 accumulates and targets MYB96 to attenuate ABA signal transduction whereas, in the presence of the hormone, MIEL1 is degraded by the UPS, promoting MYB96 accumulation. These finding suggest that MIEL1 may facilitate the ABA-mediated crosstalk between biotic and abiotic stresses (Lee and Luan, 2012).
Besides TFs, the UPS can regulate the proteolysis of additional crucial nuclear regulatory components. One example is the transcriptional co-activator NPR1, a central node for basal and salicylic acid (SA)-mediated systemic acquired resistance (SAR) through induction of PR1 gene expression (Kinkema, 2000). NPR1 accumulation is tightly regulated by both its subcellular localization and nuclear PTMs (Kinkema, 2000; Withers and Dong, 2016). Under resting conditions, NPR1 forms multimers in the cytoplasm and is constantly degraded through its interaction with CUL3-NPR4 (Spoel et al., 2009). Pathogen-induced accumulation of SA induces a redox change that promotes dissociation of the NPR1 complex and translocation of NPR1 monomers to the nucleus, where they promote gene transcription by binding to TGA and NIMIN TFs (Zhou et al., 2000; Weigel, 2005) and by directly activating the expression of WRKY TFs (Wang et al., 2010). A recent report uncovered that a dynamic balance among different PTMs not only facilitates NPR1 degradation but also switches NPR1 association with WRKY transcriptional repressors to TGA transcriptional activators (Saleh et al., 2016). In resting conditions, NPR1 is phosphorylated at Ser55/Ser59, inhibiting its SUMOylation and promoting its interaction with WRKY70, which results in repressed PR1 transcription (Saleh et al., 2016). After pathogen challenge, SA accumulation promotes Ser55/Ser59 dephosphorylation, NPR1 SUMOylation and dissociation from WRKY70. SUMOylation of NPR1 is required for (i) phosphorylation at Ser11/Ser15, triggering a signal amplification loop that generates more activated NPR1 able to interact with TGA TFs and induce PR1 gene expression, and for (ii) ubiquitination by the CUL3-NPR4 E3 Ub-ligase, ensuring a strong but transient SAR response (Spoel et al., 2009; Saleh et al., 2016). Since NPR1 not only activates SAR but also suppresses ETI responses, NPR1 degradation may promote ETI by releasing its repression of HR in cells with an elevated SA concentration (Withers and Dong, 2016). Interestingly, the presence of a BTB domain in NPR1 suggests that it may act as an adaptor in a CUL3-based complex, although no NPR1 substrate has been described to date. In agreement with its role as a master regulator of SAR, the Pseudomonas syringae type III effector AvrPtoB has been recently shown to target NPR1 and mediate its proteasomal degradation following SA accumulation (Chen et al., 2017). This interaction, which is likely to occur in the cytoplasm prior to NPR1 nuclear translocation, results in suppressed SA-mediated defense signaling. The U-Box E3 Ub-ligase PUB17 has been described as a positive regulator of plant immunity (Yang et al., 2006; He et al., 2015), which represents a rare example of physical interaction among E3 Ub-ligases in plants. The BTB domain E3 Ub-ligase POB1 interacts with PUB17 in the nucleus promoting PUB17 proteasomal degradation (Orosa et al., 2017). As a result, POB1 acts as a suppressor of basal defense and HR responses and this requires POB1 dimerization, nuclear localization and interaction with PUB17. In addition of targeting proteins for degradation, E3 Ub-ligases can control protein turnover by modification of UPS-related proteins. SAP9 is an A20/AN1-type zinc finger protein involved in development and in the regulation of multiple stress responses in Arabidopsis. SAP9 expression is induced in response to drought, cold, and salt stress as well as after ABA and pathogen challenge (Kang et al., 2017). SAP9 displays E3 Ub-ligase activity and interacts with Rad23d, a shuttle factor for the transport of ubiquitinated substrates to the proteasome (Kang et al., 2017). SAP9 may thus facilitate the degradation of ubiquitinated targets by promoting their interaction with Rad23. Alternatively, monoubiquitination of Ub-binding domain proteins by SAP9 could impair their ability to bind polyubiquitinated targets (Kang et al., 2017), in which case SAP9 would promote protein stability rather than degradation via the UPS.
In rice, RING E3 Ub-ligase OsDIS1 is mostly localized in the nucleus and negatively regulates drought stress responses both transcriptionally and post-translationally (Ning et al., 2011). OsDIS1, whose expression is induced under drought conditions, suppresses the expression of a wide range of drought-responsive genes. This could be due to OsDIS1-mediated ubiquitination of drought-related TFs, which would result in their degradation or structural modification (Ning et al., 2011). Additionally, OsDIS1 interacts with OsNEK6, a tubulin complex-related serine/threonine kinase, leading to its degradation via the UPS. Since OsNEK6 overexpression leads to increased tolerance to drought stress, it is possible that OSNEK6 promotes OSDIS1 translocation from the nucleus to the tubulin complex, providing an elegant example of regulation of the regulator.
Regulation of gene expression plays a central role during plant adaptation to environmental stimuli. The UPS and more particularly E3 Ub-ligase proteins have emerged as crucial components in the control of gene expression during plant responses to biotic and abiotic signals. Here, we have mainly focused on the proteolytic activities of ubiquitination during the modulation of nuclear protein levels in response to stress. Although less studied, ubiquitination also displays non-proteolytic functions during transcription, for example in regulating chromatin dynamics by H2B histone monoubiquitination. Indeed, the RING E3 Ub-ligase HISTONE MONOUBIQUITINATION1 (HUB1) monoubiquitinates histone H2B, positively regulates resistance against necrotrophic fungi in an ET- and SA-dependent manner (Dhawan et al., 2009) and, together with HUB2, confers tolerance to salt stress (Zhou et al., 2017). Further work on non-proteolytic roles of ubiquitination should provide a more complete picture of the different functions of this important PTM.
Fine-tuning of gene expression is essential to establish an optimal response while preventing the detrimental effects caused by stress conditions. Temporal and spatial control of gene expression is achieved by the orchestrated action of diverse regulatory mechanisms that modulate the intensity and duration of signaling and ensure cellular homeostasis. Since ubiquitination is a reversible PTM, it is crucial to gain a deeper knowledge on the dynamics of ubiquitination and on its coordination with other events that control gene expression during plant adaptation to environmental stresses.
All authors listed have made a substantial, direct and intellectual contribution to the work, and approved it for publication.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
IS was supported by an AgreenSkills fellowship within the EU Marie-Curie FP7 COFUND People Program (grant agreement n° 267196). LC was funded by the Alfonso Martín Escudero Foundation. Our work is supported by the French Laboratory of Excellence project “TULIP” (ANR-10-LABX-41; ANR-11-IDEX-0002-02). We apologize to all colleagues whose work could not be discussed because of space limitations.
Abe, H. (2002). Arabidopsis AtMYC2 (bHLH) and AtMYB2 (MYB) function as transcriptional activators in abscisic acid signaling. Plant Cell 15, 63–78. doi: 10.1105/tpc.006130
Boter, M. (2004). Conserved MYC transcription factors play a key role in jasmonate signaling both in tomato and Arabidopsis. Genes Dev. 18, 1577–1591. doi: 10.1101/gad.297704
Callis, J. (2014). The ubiquitination machinery of the ubiquitin system. Arabidopsis Book 12:e0174. doi: 10.1199/tab.0174
Catalá, R., Medina, J., and Salinas, J. (2011). Integration of low temperature and light signaling during cold acclimation response in Arabidopsis. Proc. Natl. Acad. Sci. U.S.A. 108, 16475–16480. doi: 10.1073/pnas.1107161108
Chen, H., Chen, J., Li, M., Chang, M., Xu, K., Shang, Z., et al. (2017). A bacterial type III effector targets the master regulator of salicylic acid signaling, NPR1, to subvert plant immunity. Cell Host Microbe 22, 777.e7–788.e7. doi: 10.1016/j.chom.2017.10.019
Cheng, M.-C., Hsieh, E.-J., Chen, J.-H., Chen, H.-Y., and Lin, T.-P. (2012). Arabidopsis RGLG2, functioning as a RING E3 ligase, interacts with AtERF53 and negatively regulates the plant drought stress response. Plant Physiol. 158, 363–375. doi: 10.1104/pp.111.189738
Cheng, Y. T., Li, Y., Huang, S., Huang, Y., Dong, X., Zhang, Y., et al. (2011). Stability of plant immune-receptor resistance proteins is controlled by SKP1-CULLIN1-F-box (SCF)-mediated protein degradation. Proc. Natl. Acad. Sci. U.S.A. 108, 14694–14699. doi: 10.1073/pnas.1105685108
Dhawan, R., Luo, H., Foerster, A. M., AbuQamar, S., Du, H.-N., Briggs, S. D., et al. (2009). HISTONE MONOUBIQUITINATION1 interacts with a subunit of the mediator complex and regulates defense against necrotrophic fungal pathogens in Arabidopsis. Plant Cell 21, 1000–1019. doi: 10.1105/tpc.108.062364
Dong, C. H., Agarwal, M., Zhang, Y., Xie, Q., and Zhu, J. K. (2006). The negative regulator of plant cold responses, HOS1, is a RING E3 ligase that mediates the ubiquitination and degradation of ICE1. Proc. Natl. Acad. Sci. U.S.A. 103, 8281–8286. doi: 10.1073/pnas.0602874103
Ewan, R., Pangestuti, R., Thornber, S., Craig, A., Carr, C., O’Donnell, L., et al. (2011). Deubiquitinating enzymes AtUBP12 and AtUBP13 and their tobacco homologue NtUBP12 are negative regulators of plant immunity. New Phytol. 191, 92–106. doi: 10.1111/j.1469-8137.2011.03672.x
Finkelstein, R., Gampala, S. S. L., Lynch, T. J., Thomas, T. L., and Rock, C. D. (2005). Redundant and distinct functions of the ABA response loci ABA-INSENSITIVE(ABI)5 and ABRE-BINDING FACTOR (ABF)3. Plant Mol. Biol. 59, 253–267. doi: 10.1007/s11103-005-8767-2
Furlan, G., Klinkenberg, J., and Trujillo, M. (2012). Regulation of plant immune receptors by ubiquitination. Front. Plant Sci. 3:238. doi: 10.3389/fpls.2012.00238
Gou, M., Shi, Z., Zhu, Y., Bao, Z., Wang, G., and Hua, J. (2011). The F-box protein CPR1/CPR30 negatively regulates R protein SNC1 accumulation. Plant J. 69, 411–420. doi: 10.1111/j.1365-313X.2011.04799.x
He, Q., McLellan, H., Boevink, P. C., Sadanandom, A., Xie, C., Birch, P. R. J., et al. (2015). U-box E3 ubiquitin ligase PUB17 acts in the nucleus to promote specific immune pathways triggered by Phytophthora infestans. J. Exp. Bot. 66, 3189–3199. doi: 10.1093/jxb/erv128
Himmelbach, A., Hoffmann, T., Leube, M., Hohener, B., and Grill, E. (2002). Homeodomain protein ATHB6 is a target of the protein phosphatase ABI1 and regulates hormone responses in Arabidopsis. EMBO J. 21, 3029–3038. doi: 10.1093/emboj/cdf316
Hua, Z., and Vierstra, R. D. (2011). The cullin-RING ubiquitin-protein ligases. Annu. Rev. Plant Biol. 62, 299–334. doi: 10.1146/annurev-arplant-042809-112256
Irigoyen, M. L., Iniesto, E., Rodriguez, L., Puga, M. I., Yanagawa, Y., Pick, E., et al. (2014). Targeted degradation of abscisic acid receptors is mediated by the ubiquitin ligase substrate adaptor DDA1 in Arabidopsis. Plant Cell 26, 712–728. doi: 10.1105/tpc.113.122234
Jeong, J. S., Jung, C., Seo, J. S., Kim, J. K., and Chua, N.-H. (2017). The deubiquitinating enzymes UBP12 and UBP13 positively regulate MYC2 levels in jasmonate responses. Plant Cell 29, 1406–1424. doi: 10.1105/tpc.17.00216
Jung, C., Zhao, P., Seo, J. S., Mitsuda, N., Deng, S., and Chua, N.-H. (2015). PLANT U-BOX PROTEIN10 regulates MYC2 stability in Arabidopsis. Plant Cell 27, 2016–2031. doi: 10.1105/tpc.15.00385
Kang, M., Lee, S., Abdelmageed, H., Reichert, A., Lee, H.-K., Fokar, M., et al. (2017). Arabidopsis stress associated protein 9 mediates biotic and abiotic stress responsive ABA signaling via the proteasome pathway. Plant Cell Environ. 40, 702–716. doi: 10.1111/pce.12892
Kim, S. H., Gao, F., Bhattacharjee, S., Adiasor, J. A., Nam, J. C., and Gassmann, W. (2010). The Arabidopsis resistance-like gene SNC1 is activated by mutations in SRFR1 and contributes to resistance to the bacterial effector AvrRps4. PLOS Pathog. 6:e1001172. doi: 10.1371/journal.ppat.1001172
Kinkema, M. (2000). Nuclear localization of NPR1 is required for activation of PR gene expression. Plant Cell 12, 2339–2350. doi: 10.1105/tpc.12.12.2339
Lau, O. S., and Deng, X. W. (2010). Plant hormone signaling lightens up: integrators of light and hormones. Curr. Opin. Plant Biol. 13, 571–577. doi: 10.1016/j.pbi.2010.07.001
Lechner, E., Leonhardt, N., Eisler, H., Parmentier, Y., Alioua, M., Jacquet, H., et al. (2011). MATH/BTB CRL3 receptors target the homeodomain-leucine zipper ATHB6 to modulate abscisic acid signaling. Dev. Cell 21, 1116–1128. doi: 10.1016/j.devcel.2011.10.018
Lee, H. G., and Seo, P. J. (2016). The Arabidopsis MIEL1 E3 ligase negatively regulates ABA signalling by promoting protein turnover of MYB96. Nat. Commun. 7:12525. doi: 10.1038/ncomms12525
Lee, J.-H., Yoon, H.-J., Terzaghi, W., Martínez, C., Dai, M., Li, J., et al. (2012). DWA1 and DWA2, two Arabidopsis DWD protein components of CUL4-based E3 ligases, act together as negative regulators in ABA signal transduction. Plant Cell 22, 1716–1732. doi: 10.1105/tpc.109.073783
Lee, S. C., and Luan, S. (2012). ABA signal transduction at the crossroad of biotic and abiotic stress responses. Plant Cell Environ. 35, 53–60. doi: 10.1111/j.1365-3040.2011.02426.x
Li, B., Lu, D., and Shan, L. (2014). Ubiquitination of pattern recognition receptors in plant innate immunity. Mol. Plant Pathol. 15, 737–746. doi: 10.1111/mpp.12128
Li, Y., Li, S., Bi, D., Cheng, Y. T., Li, X., and Zhang, Y. (2010). SRFR1 negatively regulates plant NB-LRR resistance protein accumulation to prevent autoimmunity. PLOS Pathog. 6:e1001111. doi: 10.1371/journal.ppat.1001111
Lim, S. D., Cho, H. Y., Park, Y. C., Ham, D. J., Lee, J. K., and Jang, C. S. (2013). The rice RING finger E3 ligase, OsHCI1, drives nuclear export of multiple substrate proteins and its heterogeneous overexpression enhances acquired thermotolerance. J. Exp. Bot. 64, 2899–2914. doi: 10.1093/jxb/ert143
Lopez-Molina, L. (2003). AFP is a novel negative regulator of ABA signaling that promotes ABI5 protein degradation. Genes Dev. 17, 410–418. doi: 10.1101/gad.1055803
Lopez-Molina, L., Mongrand, S., and Chua, N. H. (2001). A postgermination developmental arrest checkpoint is mediated by abscisic acid and requires the ABI5 transcription factor in Arabidopsis. Proc. Natl. Acad. Sci. U.S.A. 98, 4782–4787. doi: 10.1073/pnas.081594298
Lorenzo, O. (2004). JASMONATE-INSENSITIVE1 encodes a MYC transcription factor essential to discriminate between different jasmonate-regulated defense responses in Arabidopsis. Plant Cell 16, 1938–1950. doi: 10.1105/tpc.022319
Luo, H., Laluk, K., Lai, Z., Veronese, P., Song, F., and Mengiste, T. (2010). The Arabidopsis botrytis susceptible1 interactor defines a subclass of RING E3 ligases that regulate pathogen and stress responses. Plant Physiol. 154, 1766–1782. doi: 10.1104/pp.110.163915
Lyzenga, W. J., and Stone, S. L. (2012). Abiotic stress tolerance mediated by protein ubiquitination. J. Exp. Bot. 63, 599–616. doi: 10.1093/jxb/err310
Marino, D., Froidure, S., Canonne, J., Ben Khaled, S., Khafif, M., Pouzet, C., et al. (2013). Arabidopsis ubiquitin ligase MIEL1 mediates degradation of the transcription factor MYB30 weakening plant defence. Nat. Commun. 4:1476. doi: 10.1038/ncomms2479
Marino, D., Peeters, N., and Rivas, S. (2012). Ubiquitination during plant immune signaling. Plant Physiol. 160, 15–27. doi: 10.1104/pp.112.199281
Mazzucotelli, E., Belloni, S., Marone, D., De Leonardis, A., Guerra, D., Di Fonzo, N., et al. (2006). The E3 ubiquitin ligase gene family in plants: regulation by degradation. Curr. Genomics 7, 509–522. doi: 10.2174/138920206779315728
Mengiste, T. (2003). The BOTRYTIS SUSCEPTIBLE1 gene encodes an R2R3MYB transcription factor protein that is required for biotic and abiotic stress responses in Arabidopsis. Plant Cell 15, 2551–2565. doi: 10.1105/tpc.014167
Moon, J. (2004). the ubiquitin-proteasome pathway and plant development. Plant Cell 16, 3181–3195. doi: 10.1105/tpc.104.161220
Morimoto, K., Mizoi, J., Qin, F., Kim, J.-S., Sato, H., Osakabe, Y., et al. (2013). Stabilization of Arabidopsis DREB2A is required but not sufficient for the induction of target genes under conditions of stress. PLOS ONE 8:e80457. doi: 10.1371/journal.pone.0080457
Morimoto, K., Ohama, N., Kidokoro, S., Mizoi, J., Takahashi, F., Todaka, D., et al. (2017). BPM-CUL3 E3 ligase modulates thermotolerance by facilitating negative regulatory domain-mediated degradation of DREB2A in Arabidopsis. Proc. Natl. Acad. Sci. U.S.A. 114, E8528–E8536. doi: 10.1073/pnas.1704189114
Nagels Durand, A., Pauwels, L., and Goossens, A. (2016). The ubiquitin system and jasmonate signaling. Plants 5:6. doi: 10.3390/plants5010006
Nguyen, D., Rieu, I., Mariani, C., and van Dam, N. M. (2016). How plants handle multiple stresses: hormonal interactions underlying responses to abiotic stress and insect herbivory. Plant Mol. Biol 91, 727–740. doi: 10.1007/s11103-016-0481-8
Ning, Y., Jantasuriyarat, C., Zhao, Q., Zhang, H., Chen, S., Liu, J., et al. (2011). The SINA E3 ligase OsDIS1 negatively regulates drought response in rice. Plant Physiol. 157, 242–255. doi: 10.1104/pp.111.180893
Orosa, B., He, Q., Mesmar, J., Gilroy, E. M., McLellan, H., Yang, C., et al. (2017). BTB-BACK domain protein POB1 suppresses immune cell death by targeting ubiquitin E3 ligase PUB17 for degradation. PLOS Genet. 13:e1006540. doi: 10.1371/journal.pgen.1006540
Park, G.-G., Park, J.-J., Yoon, J., Yu, S.-N., and An, G. (2010). A RING finger E3 ligase gene, Oryza sativa delayed seed germination 1 (OsDSG1), controls seed germination and stress responses in rice. Plant Mol. Biol. 74, 467–478. doi: 10.1007/s11103-010-9687-3
Perea-Resa, C., Rodríguez-Milla, M. A., Iniesto, E., Rubio, V., and Salinas, J. (2017). Prefoldins negatively regulate cold acclimation in Arabidopsis thaliana by promoting nuclear proteasome-mediated HY5 degradation. Mol. Plant 10, 791–804. doi: 10.1016/j.molp.2017.03.012
Qin, F., Sakuma, Y., Tran, L. S. P., Maruyama, K., Kidokoro, S., Fujita, Y., et al. (2008). Arabidopsis DREB2A-interacting proteins function as RING E3 ligases and negatively regulate plant drought stress-responsive gene expression. Plant Cell 20, 1693–1707. doi: 10.1105/tpc.107.057380
Robert-Seilaniantz, A., Grant, M., and Jones, J. D. G. (2011). Hormone crosstalk in plant disease and defense: more than just jasmonate-salicylate antagonism. Annu. Rev. Phytopathol. 49, 317–343. doi: 10.1146/annurev-phyto-073009-114447
Sadanandom, A., Bailey, M., Ewan, R., Lee, J., and Nelis, S. (2012). The ubiquitin-proteasome system: central modifier of plant signalling. New Phytol. 196, 13–28. doi: 10.1111/j.1469-8137.2012.04266.x
Sadowski, M., Suryadinata, R., Tan, A. R., Roesley, S. N., and Sarcevic, B. (2012). Protein monoubiquitination and polyubiquitination generate structural diversity to control distinct biological processes. IUBMB Life 64, 136–142. doi: 10.1002/iub.589
Sakuma, Y. (2006). Functional analysis of an Arabidopsis transcription factor, DREB2A, involved in drought-responsive gene expression. Plant Cell 18, 1292–1309. doi: 10.1105/tpc.105.035881
Sakuma, Y., Maruyama, K., Qin, F., Osakabe, Y., Shinozaki, K., and Yamaguchi-Shinozaki, K. (2006). Dual function of an Arabidopsis transcription factor DREB2A in water-stress-responsive and heat-stress-responsive gene expression. Proc. Natl. Acad. Sci. U.S.A. 103, 18822–18827. doi: 10.1073/pnas.0605639103
Saleh, A., Withers, J., Mohan, R., Marqués, J., Gu, Y., Yan, S., et al. (2016). Posttranslational modifications of the master transcriptional regulator NPR1 enable dynamic but tight control of plant immune responses. Cell Host Microbe 19, 127–130. doi: 10.1016/j.chom.2015.12.008
Seo, K. I., Lee, J. H., Nezames, C. D., Zhong, S., Song, E., Byun, M. O., et al. (2014). ABD1 is an Arabidopsis DCAF substrate receptor for CUL4-DDB1-based E3 ligases that acts as a negative regulator of abscisic acid signaling. Plant Cell 26, 695–711. doi: 10.1105/tpc.113.119974
Smalle, J., Kurepa, J., Yang, P., Emborg, T. J., Babiychuk, E., Kushnir, S., et al. (2003). The pleiotropic role of the 26S proteasome subunit RPN10 in Arabidopsis growth and development supports a substrate-specific function in abscisic acid signaling. Plant Cell 15, 965–980.
Söderman, E., Hjellström, M., Fahleson, J., and Engström, P. (1999). The HD-Zip gene ATHB6 in Arabidopsis is expressed in developing leaves, roots and carpels and up-regulated by water deficit conditions. Plant Mol. Biol. 40, 1073–1083.
Spoel, S. H., Mou, Z., Tada, Y., Spivey, N. W., Genschik, P., and Dong, X. (2009). Proteasome-mediated turnover of the transcription coactivator NPR1 plays dual roles in regulating plant immunity. Cell 137, 860–872. doi: 10.1016/j.cell.2009.03.038
Stone, S. L. (2005). Functional analysis of the RING-type ubiquitin ligase family of Arabidopsis. Plant Physiol. 137, 13–30. doi: 10.1104/pp.104.052423
Stone, S. L. (2014). The role of ubiquitin and the 26S proteasome in plant abiotic stress signaling. Front. Plant Sci. 5:135. doi: 10.3389/fpls.2014.00135
Stone, S. L., Williams, L. A., Farmer, L. M., Vierstra, R. D., and Callis, J. (2006). KEEP ON GOING, a RING E3 ligase essential for Arabidopsis growth and development, is involved in abscisic acid signaling. Plant Cell 18, 3415–3428. doi: 10.1105/tpc.106.046532
Trujillo, M., and Shirasu, K. (2010). Ubiquitination in plant immunity. Curr. Opin. Plant Biol. 13, 402–408. doi: 10.1016/j.pbi.2010.04.002
Vierstra, R. D. (2009). The ubiquitin–26S proteasome system at the nexus of plant biology. Nat. Rev. Mol. Cell Biol. 10, 385–397. doi: 10.1038/nrm2688
Wang, H., Avci, U., Nakashima, J., Hahn, M. G., Chen, F., and Dixon, R. A. (2010). Mutation of WRKY transcription factors initiates pith secondary wall formation and increases stem biomass in dicotyledonous plants. Proc. Natl. Acad. Sci. U.S.A. 107, 22338–22343. doi: 10.1073/pnas.1016436107
Weigel, R. R. (2005). Interaction of NIMIN1 with NPR1 modulates PR gene expression in Arabidopsis. Plant Cell 17, 1279–1291. doi: 10.1105/tpc.104.027441
Withers, J., and Dong, X. (2016). Posttranslational modifications of npr1: a single protein playing multiple roles in plant immunity and physiology. PLOS Pathog. 12:e1005707. doi: 10.1371/journal.ppat.1005707
Yang, C.-W., Gonzalez-Lamothe, R., Ewan, R. A., Rowland, O., Yoshioka, H., Shenton, M., et al. (2006). The E3 ubiquitin ligase activity of Arabidopsis PLANT U-BOX17 and its functional tobacco homolog ACRE276 are required for cell death and defense. Plant Cell 18, 1084–1098. doi: 10.1105/tpc.105.039198
Yang, W., Zhang, W., and Wang, X. (2016). Post-translational control of ABA signalling: the roles of protein phosphorylation and ubiquitination. Plant Biotechnol. J. 15, 4–14. doi: 10.1111/pbi.12652
Yee, D., and Goring, D. R. (2009). The diversity of plant U-box E3 ubiquitin ligases: from upstream activators to downstream target substrates. J. Exp. Bot. 60, 1109–1121. doi: 10.1093/jxb/ern369
Yu, F., Wu, Y., and Xie, Q. (2015). Precise protein post-translational modifications modulate ABI5 activity. Trends Plant Sci. 20, 569–575. doi: 10.1016/j.tplants.2015.05.004
Yu, Y., Xu, W., Wang, J., Wang, L., Yao, W., Yang, Y., et al. (2013). The Chinese wild grapevine (Vitis pseudoreticulata) E3 ubiquitin ligase Erysiphe necator-induced RING finger protein 1 (EIRP1) activates plant defense responses by inducing proteolysis of the VpWRKY11 transcription factor. New Phytol. 200, 834–846. doi: 10.1111/nph.12418
Zhang, X., Garreton, V., and Chua, N.-H. (2005). The AIP2 E3 ligase acts as a novel negative regulator of ABA signaling by promoting ABI3 degradation. Genes Dev. 19, 1532–1543. doi: 10.1101/gad.1318705
Zhou, B., and Zeng, L. (2016). Conventional and unconventional ubiquitination in plant immunity. Mol. Plant Pathol. 18, 1313–1330. doi: 10.1111/mpp.12521
Zhou, J.-M., Trifa, Y., Silva, H., Pontier, D., Lam, E., Shah, J., et al. (2000). NPR1 differentially interacts with members of the TGA/OBF family of Transcription factors that bind an element of the PR-1Gene required for induction by salicylic acid. Mol. Plant Microbe Interact. 13, 191–202. doi: 10.1094/MPMI.2000.13.2.191
Keywords: E3 ubiquitin-ligase, plant cell nucleus, post-translational modification, 26S proteasome, transcription factor
Citation: Serrano I, Campos L and Rivas S (2018) Roles of E3 Ubiquitin-Ligases in Nuclear Protein Homeostasis during Plant Stress Responses. Front. Plant Sci. 9:139. doi: 10.3389/fpls.2018.00139
Received: 18 November 2017; Accepted: 24 January 2018;
Published: 08 February 2018.
Edited by:
Christian Mazars, UMR5546 Laboratoire de Recherche en Sciences Vegetales (LRSV), FranceReviewed by:
Lirong Zeng, University of Nebraska System, United StatesCopyright © 2018 Serrano, Campos and Rivas. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Irene Serrano, aXJlc2VycmFAZ21haWwuY29t Susana Rivas, c3VzYW5hLnJpdmFzQGlucmEuZnI=
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.