 Suravoot Yooyongwech1
Suravoot Yooyongwech1 Rujira Tisarum
Rujira Tisarum Suriyan Cha-um
Suriyan Cha-um
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Plant Sci. , 08 August 2017
Sec. Plant Breeding
Volume 8 - 2017 | https://doi.org/10.3389/fpls.2017.01400
This article is part of the Research Topic Tailoring crops for higher water productivity View all 12 articles
The objective of this study was to elevate water deficit tolerance by improving soluble sugar and free proline accumulation, photosynthetic pigment stabilization, photosynthetic abilities, growth performance and storage root yield in sweet potato cv. ‘Tainung 57’ using a foliar application of paclobutrazol (PBZ). The experiment followed a Completely Randomized Block Design with four concentrations of PBZ: 0 (control), 17, 34, and 51 μM before exposure to 47.5% (well irrigation), 32.3% (mild water deficit) or 17.5% (severe water deficit) soil water content. A sweet potato cultivar, ‘Japanese Yellow’, with water deficit tolerance attributes was the positive check in this study. Total soluble sugar content (sucrose, glucose, and fructose) increased by 3.96-folds in ‘Tainung 57’ plants treated with 34 μM PBZ grown under 32.3% soil water content (SWC) compared to the untreated plants, adjusting osmotic potential in the leaves and controlling stomatal closure (represented by stomatal conductance and transpiration rate). In addition, under the same treatment, free proline content (2.15 μmol g-1 FW) increased by 3.84-folds when exposed to 17.5% SWC. PBZ had an improved effect on leaf size, vine length, photosynthetic pigment stability, chlorophyll fluorescence, and net photosynthetic rate; hence, delaying wilting symptoms and maintaining storage root yield (26.93 g plant-1) at the harvesting stage. A positive relationship between photon yield of PSII (ΦPSII) and net photosynthetic rate was demonstrated (r2 = 0.73). The study concludes that soluble sugar and free proline enrichment in PBZ-pretreated plants may play a critical role as major osmoprotectant to control leaf osmotic potential and stomatal closure when plants were subjected to low soil water content, therefore, maintaining the physiological and morphological characters as well as storage root yield.
Sweet potato is classified as a candidate rain-fed crop for the semi-arid and arid regions (Gomes and Carr, 2001, 2003; Gomes et al., 2005). However, climate change has resulted in drought or water shortage or water deficit (WD) conditions (Rockstrom, 2003; Blum, 2005; Neumann, 2008; Falkenmark, 2013). Under these conditions, crop water use efficiency, crop water content are directly influenced, leading to low net photosynthetic rate, growth reduction, and storage root yield loss (van Heeden and Laurie, 2008; Gajanayake et al., 2014; Yooyongwech et al., 2016). Biochemical, physiological, and morphological performances of sweet potato plants under WD conditions have been well assessed using multivariate cluster analysis and ‘Japanese Yellow’ and ‘PROC 65-3’ have been identified as WD tolerance cultivars (Yooyongwech et al., 2013). Soluble sugar enrichment and free proline accumulation at the cellular level play a key role in osmotic adjustment, which promotes drought defense mechanisms (Wang et al., 2000; Yooyongwech et al., 2016).
Triazole compounds such as paclobutrazol (PBZ) (Sankar et al., 2007), triadimefon (Manivannan et al., 2008), hexaconazole (Chehelpar et al., 2016), and uniconazole (Zhang et al., 2007) promote osmoregulatory effects on plants. Paclobutrazol [(2RS, 3RS)-1-4(-chlorophenyl)-4, dimethyl-2-1,2,4-triszol-1-yl-penten-3-ol] is a known gibberellic acid inhibitor (anti-GA) and has been applied as plant growth retardant. In sweet potato, flood tolerance (cv. ‘Taoyuan 2’; 5 days flooding) and chilling tolerance traits (cvs. ‘TN71’ and ‘TN65’; ambient temperature at 7°C for 5 days) have been reported using PBZ-pretreatment at 0.5 mg plant-1 and 300 μg plant-1, respectively. Under flooding and chilling stress, the regulation of antioxidative system, i.e., enzymatic and non-enzymatic defense mechanisms, in PBZ-pretreated plants were reported (Lin et al., 2006a,b). PBZ also induces drought tolerance in Stevia rebaudiana (Hajihashemi and Ehsanpour, 2013, 2014), Aesculus hippocastanum (Percival and Noviss, 2008), and Arachis hypogaea (Sankar et al., 2007). Moreover, the possible hypotheses on drought tolerance regulation by PBZ have been proposed which state that it maintains the endogenous cytokinin levels (zeatin and zeatin riboside) and stabilize leaf water potential (Zhu et al., 2004) causing increased leaf and epidermal thickness (Sankar et al., 2013). Alternatively, regulation of free proline and glycine betaine as major osmoprotectants (Hajihashemi and Ehsanpour, 2013) and promotion of enzymatic and non-enzymatic antioxidant activities, reduce the toxicity derived from drought stress (Sankar et al., 2007; Hajihashemi and Ehsanpour, 2014; Jungklang et al., 2016). However, knowledge on PBZ-regulated sugar and free proline enrichment in sweet potato grown under WD condition is limited. The aim of this investigation was to: (a) improve the WD tolerant abilities in sweet potato subjected to varying degrees of water stress using foliar PBZ concentrations, and (b) evaluate biochemical, physiological, and morphological characteristics and storage root traits in each treatment.
Two sweet potato genotypes, namely ‘Japanese Yellow’ (WD tolerance; positive check) and ‘Tainung 57’ (WD sensitive) obtained from Agricultural Extension Group, Phichit province, Thailand, were used as master stock materials (Yooyongwech et al., 2013). Single vine cuttings (15 ± 1 cm in length) without leaf blades were propagated and planted into plastic pots (∅ = 20 cm) containing 2 kg garden soil (EC = 2.687 dS m-1; pH = 5.5; organic matter = 10.36%; total nitrogen = 0.17%; total phosphorus = 0.07%; and total potassium = 1.19%). The cuttings planted in the pot culture were incubated in a greenhouse under 500-1,000 μmol m-2 s-1 photosynthetic photon flux density with a 10 h d-1 photoperiod, 28 ± 2°C ambient temperature and 80 ± 5% relative humidity, for 4 weeks. The ‘Tainung 57’ plants were treated with exogenous foliar applications of PBZ at 0, 17, 34, and 51 μM (25 mL plant-1). Each treatment consisted of well-watered (WW; 47.5% soil water content), mild water deficit (MWD; 32.3% soil water content, by withholding water for 7 days), and severe water deficit (SWD; 17.5% soil water content, by withholding water for 14 days) conditions (Supplementary Figure S1). Growth characteristics (vine length and number of leaves), soluble sugar, free proline content, leaf osmotic potential, photosynthetic pigments, chlorophyll fluorescence, net photosynthetic rate (Pn), stomatal conductance (gs), and transpiration rate (E) in each treatment was measured. In addition, number of storage roots per plant, storage root yield, vine fresh weight, root fresh weight, vine dry weight, and root dry weight in the harvesting period (150 days after cutting) were recorded.
Sucrose, glucose, and fructose content in the second fully expanded leaf from shoot tip were measured following the method of Karkacier et al. (2003). In brief, sweet potato leaves were collected and freeze-dried using a freeze-dryer. Fifty-milligram sample was ground in a mortar with liquid nitrogen. Following this, 1 mL of nanopure water was added and centrifuged at 12,000 rpm for 15 min. The supernatant was collected and filtered through a 0.45 μm membrane filter (VertiPureTM, Vertical®). Twenty microliters of the filtrate was injected into Waters HPLC equipped with a MetaCarb 87C column and a guard column. Deionized water was used as the mobile phase at a flow rate of 0.5 mL min-1. The online detection was performed using a Waters 410 differential refractometer detector and the data was analyzed by Empower® software. Sucrose, glucose, and fructose (Fluka, United States) were used as the standards.
Free proline in the second fully expanded leaf from shoot tip was extracted and analyzed according to the method of Bates et al. (1973). In brief, 50 mg of fresh material was ground with liquid nitrogen in a mortar. The homogenate powder was mixed with 1 mL aqueous sulfosalicylic acid (3%, w/v) and filtered through Whatman #1 filter paper (Whatman, England). The extracted solution was reacted with an equal volume of glacial acetic acid and ninhydrin reagent (1.25 mg ninhydrin in 30 mL glacial acetic acid and 20 mL 6 M H3PO4) and incubated at 95°C for 1 h. The reaction was terminated by placing the container in an ice bath. The reaction mixture was mixed vigorously with 2 mL of toluene. After cooling to 25°C, the chromophore was measured at 520 nm by spectrophotometer (HACH DR/4000; Model 48000, HACH Company, Loveland, CO, United States) using L-proline as a calibration standard.
Osmotic potential in the leaves of sweet potato was measured, according to Lanfermeijer et al. (1991). In brief, 100 mg of fresh leaf tissue was chopped into small pieces, transferred to 1.5 mL micro tube, and then crushed using a glass rod. The 20 μL of extracted solution was dropped directly onto a filter paper in an osmometer chamber (5520 Vapro®, Wescor, Logan, UT, United States) and subsequently the data were collected. The osmolarity (mmol kg-1) was converted to osmotic potential (MPa) using conversion factor of osmotic potential measurement.
Chlorophyll a (Chl a), chlorophyll b (Chl b), total chlorophyll (TC), and total carotenoid content in the second fully expanded leaf from shoot tip were analyzed according to the method of Shabala et al. (1998), whereas total carotenoid (Cx+c) content was assayed following Lichtenthaler (1987) method. One hundred milligram leaf tissue was homogenized in glass vials using 10 mL of 99.5% acetone, and blended using a homogenizer. The glass vials were sealed with Parafilm® to prevent evaporation, and then stored at 4°C for 48 h. Chl a and Chl b concentrations were measured at 662 and 644 nm, whereas Cx+c concentration measured at 470 nm using UV-VIS spectrophotometer against acetone (99.5%) as a blank.
Chlorophyll fluorescence emission was measured from the adaxial surface of the second fully expanded leaf from the shoot tip using a fluorescence monitoring system (model FMS 2; Hansatech Instruments Ltd., Norfolk, United Kingdom) in the pulse amplitude modulation mode (Loggini et al., 1999). A leaf, kept in dark for 30 min was initially exposed to the modulated measuring beam of far-red light (LED source) with typical peak at 735 nm. Original (F0) and maximum (Fm) fluorescence yields were measured under weak modulated red light (<0.5 μmol m-2 s-1) with 1.6 s pulses of saturating light (>6.8 μmol m-2 s-1 PAR) and calculated using FMS software for Windows®. The variable fluorescence yield (Fv) was calculated using the equation: Fv = Fm - F0. The ratio of variable to maximum fluorescence (Fv/Fm) was calculated as the maximum quantum yield of PSII photochemistry. The photon yield of PSII (ΦPSII) in the light was calculated as: ΦPSII = (Fm′ -F)/Fm′ after 45 s of illumination, when steady state was achieved (Maxwell and Johnson, 2000).
Net photosynthetic rate (Pn; μmol m-2 s-1), stomatal conductance (gs; mmol H2O m-2 s-1), and transpiration rate (E; mmol m-2 s-1) of the second fully expanded leaf from shoot tip were measured using a Portable Photosynthesis System with an Infra-red Gas Analyzer (Model LI 6400, LI-COR® Inc., Lincoln, NE, United States). Transpiration rate was measured continuously by monitoring the content of air entering and exiting the IRGA headspace chamber, according to Cha-um et al. (2007). Flow rate of IRGA headspace chamber was set as 500 μmol s-1 with ambient CO2 concentration (350 ± 10 μmol CO2 mol-1) and 1,000 μmol m-2 s-1 photosynthetic photon flux density provided by 6400-02 B LED Red/Blue light source.
Vine length, leaf length, leaf width, number of leaves, storage root yield, vine fresh weight, root fresh weight, vine dry weight, and root dry weight of sweet potato were measured. Vine and roots were dried at 80°C in a hot-air oven for 2 days, and then incubated in desiccator before the measurement of dry weight. In addition, the leaf toxic symptoms, i.e., leaf wilting and leaf chlorosis, were observed (Supplementary Figure S2).
The experiment was arranged as 4 × 2 factorial in a Completely Randomized Block Design (CRBD) with eight replicates (n = 8). Two-way analysis of variance (ANOVA) in each parameter was validated using SPSS software (Supplementary Tables S1–S3). The mean values obtained were compared using Tukey’s HSD and analyzed with SPSS software. Pearson correlation between Chl a content and Fv/Fm, TC and ΦPSII, ΦPSII and Pn, and Pn reduction and plant dry weight was analyzed.
Morphological characteristics of the storage roots of sweet potato at the harvest stage are shown in Figure 1. There was no significant difference observed in the storage root yield of ‘Japanese Yellow’, the positive check, under WW and WD conditions (only 5.4% reduction). ‘Tainung 57’ plants grown under WW conditions and pretreated with PBZ had a higher storage root yield than the non-pretreated plants (Figure 1). In WD treatment, storage root yield was significantly improved especially in 37 μM PBZ pretreated plants (Figure 1) Storage root yield of untreated ‘Tainung 57’ was significantly decreased by 47.55% when exposed to 17.5% SWC (SWD) compared to WW conditions. However, when the plants were treated with 17, 34, and 51 μM of PBZ, the yield decreased up to 37.68, 35.77, and 34.95%, respectively (Table 1).

FIGURE 1. Storage root characteristics of sweet potato pretreated with paclobutrazol (PBZ) grown under well watering [WW; 47.5% soil water content (SWC)] or severe water deficit (SWD) (17.5% SWC; 14 days water withholding) subsequently recovery prior to storage root harvesting process (150 days after planting).
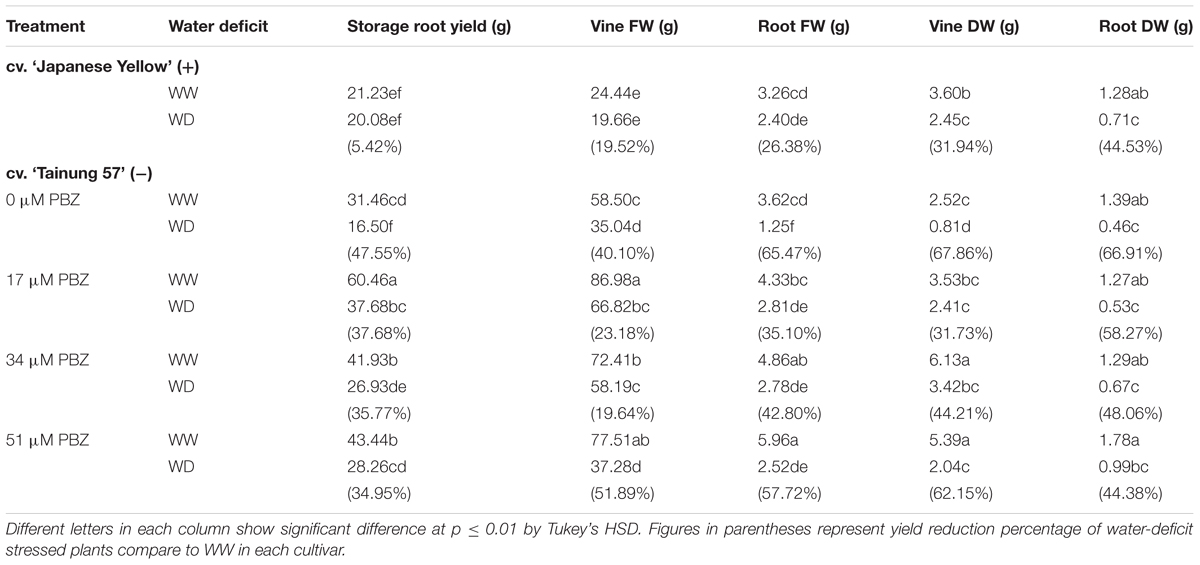
TABLE 1. Storage root yield, vine fresh weight, root fresh weight, vine dry weight, and root dry weight of sweet potato pretreated with paclobutrazol (PBZ) subsequently subjected to well watering [WW; 47.5% soil water content (SWC)] or severe water deficit (SWD; 17.5% SWC) and then recovered until storage root harvesting stage (150 days after cutting).
Vine fresh weight, root fresh weight, vine dry weight, and root dry weight in PBZ-pretreated plants raised under WW conditions were greater than those in water stress. In WD stress conditions, vine fresh weight, root fresh weight, vine dry weight, and root dry weight in non-pretreated plants declined by 40.10, 65.47, 67.86, and 66.91%, respectively; whereas, these parameters improved with the 34 μM PBZ treatment (19.64, 42.80, 44.21, and 48.06%, respectively). In addition, under WD conditions the root fresh weight (35.10% reduction) and vine dry weight (31.73% reduction) in 17 μM PBZ-pretreated plants were maintained. In the positive check cv. ‘Japanese Yellow,’ the above traits in WD stressed plants showed higher performance than the ‘Tainung 57’ plants (Table 1).
Leaf wilting and chlorosis were evidently observed in the older leaves when sweet potato plants were exposed to SWD, especially in non-treated plants (Supplementary Figure S2). Vine length was significantly reduced by PBZ treatment, whereas leaf length, leaf width, and number of leaves remained same. In WW conditions, plant height or vine length of sweet potato pretreated with 34 μM PBZ was reduced by 18.78% when compared with control. Under MWD conditions (32.3% SWC), vine length, leaf area, leaf width, and number of leaves of sweet potato cv. ‘Japanese Yellow’ were maintained (with only 6.96, 2.54, 4.25, and 6.04% reduction, respectively) with a lower reduction rate as compared to the cv. ‘Tainung 57’ (with 31.62, 10.01, 13.78, and 19.36% reduction, respectively). The degree of reduction in each parameter was amplified with the extent of water deficiency and genetic variations, especially in cv. ‘Tainung 57’ (Table 2). In PBZ-pretreated plants, overall growth performances under WD conditions were alleviated. Interestingly, vine length, leaf area, leaf width, and number of leaves in 34 μM PBZ pretreated plants of cv. ‘Tainung 57’ were maintained in both MWD (10.52, 3.09, 0.79, and 3.13% reduction) and SWD (23.02%, 13.00%, 20.10% and 34.92% reduction) (Table 2).
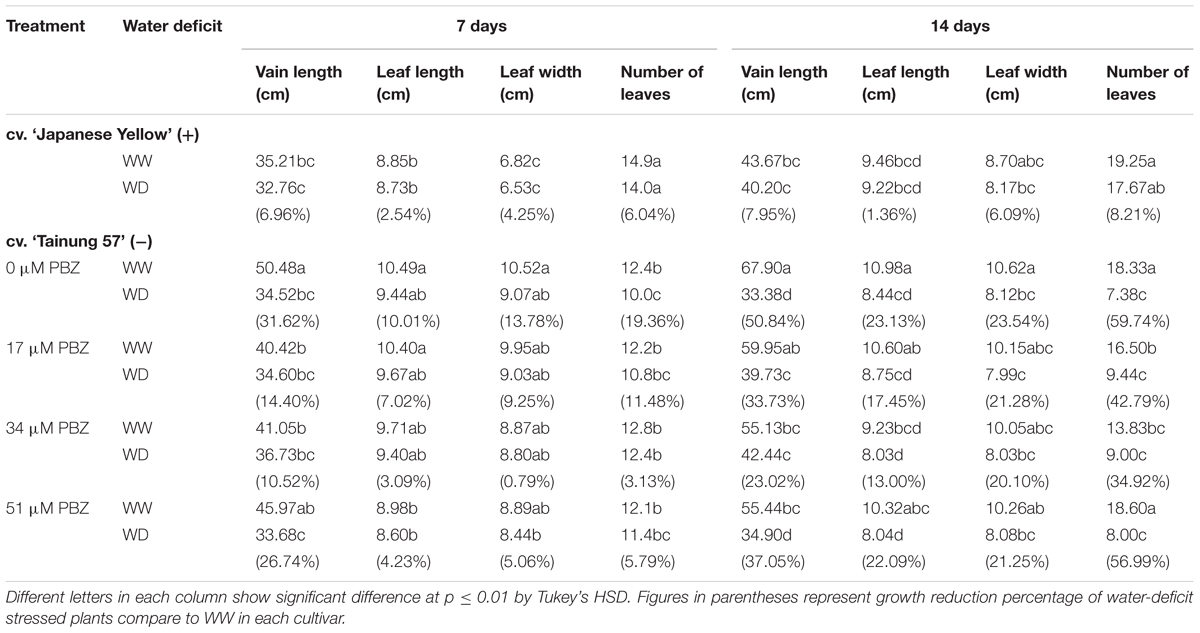
TABLE 2. Vine length, leaf length, leaf width, and number of leaves of sweet potato pretreated with PBZ subsequently subjected to WW (47.5% SWC) or mild water deficit (32.3% SWC) for 7 days or SWD (17.5% SWC) for 14 days.
Free proline in sweet potato cv. ‘Japanese Yellow’ grown under MWD was enriched by 1.87-fold (2.78 μmol g-1 FW) over the WW control. However, the proline levels of PBZ pretreated and untreated cv. ‘Tainung 57’ were not significantly different when exposure to MWD (Figure 2A). In SWD, free proline peaked (2.81 μmol g-1 FW) in the positive check cv. ‘Japanese Yellow’, while it was maintained in cv. ‘Tainung 57’ without PBZ treatment (Figure 2B). In addition, free proline content in PBZ-pretreated cv. ‘Tainung 57’ grown under SWD was significantly enriched by 2.94- (2.03 μmol g-1 FW), 3.84- (2.15 μmol g-1 FW), and 2.28-fold (2.14 μmol g-1 FW), when pretreated with 17, 34, and 51 μM PBZ, respectively (Figure 2B).

FIGURE 2. Free proline and total soluble sugar in leaf tissues of sweet potato pretreated with PBZ subsequently subjected to mild water deficit (32.3% SWC; 7 days water withholding) (A,C) or SWD (17.5% SWC; 14 days water withholding) (B,D). Different letters in each bar show significant difference at p ≤ 0.01 by Tukey’s HSD. Data presented as mean ± SE.
Total soluble sugar in MWD stressed plants increased by 2.87- (26.85 mg g-1 DW) and 1.85-folds (27.61 mg g-1 DW) over the control in cvs. ‘Japanese Yellow’ and ‘Tainung 57,’ respectively. The increase was most prominent at 34 μM PBZ where the total soluble sugar concentration was 49.33 mg g-1 DW (3.69-fold over control) (Figure 2C). Under SWD conditions, total soluble sugar was enriched in both the positive check cv. ‘Japanese Yellow’ (3.63-folds over control) and the treated cv. ‘Tainung 57’ (1.86-folds over control). Under SWD conditions, there was a significant increase in total soluble sugar content in ‘Tainung 57’, by 3.91-, 2.39-, and 1.53-folds over well irrigated conditions when sprayed by 17, 34 and 51 μM PBZ, respectively (Figure 2D). In WD conditions, the enhancement in sugar classes followed a fructose > glucose > sucrose trend in the leaf tissues and this may play a key role in osmotic adjustment in sweet potato. Sucrose content (6.93 mg g-1 DW) in 34 μM PBZ pretreated plants grown under MWD was enriched by 6.08-folds over control. Glucose and fructose in cvs. ‘Japanese Yellow’ and ‘Tainung 57’ increased in relation to the degree of WD stress. Glucose and fructose contents peaked at 11.32 mg g-1 DW (2.87-folds over control) and 31.08 mg g-1 DW (4.21-folds over control) (Table 3). In SWD, sucrose, glucose, and fructose in positive check cv. ‘Japanese Yellow’ were enriched by 3.97-folds (3.02 mg g-1 DW), 1.99-folds (13.44 mg g-1 DW), and 5.19-folds (35.71 mg g-1 DW), respectively, over the control. In contrast, sucrose content in PBZ-pretreated plants cv. ‘Tainung 57’ in both well irrigated and SWD stress was unchanged (Table 3). However, glucose and fructose in the leaf tissues of 34 μM PBZ-pretreated plants of cv. ‘Tainung 57’ peaked at 13.35 mg g-1 DW (2.29-folds over control) and 26.94 mg g-1 DW (2.49-folds over control), respectively (Table 3).
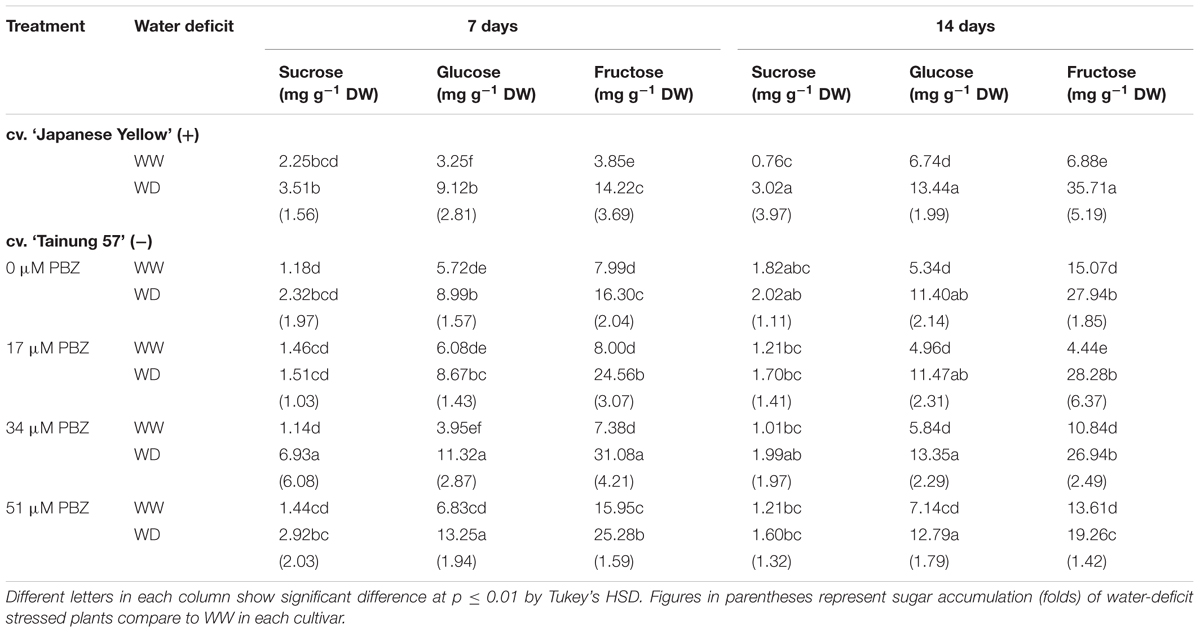
TABLE 3. Sucrose, glucose and fructose content in leaf tissues of sweet potato pretreated with PBZ subsequently subjected to WW (47.5% SWC), mild water deficit (32.3% SWC; 7 days water withholding) or SWD (17.5% SWC; 14 days water withholding).
Osmotic potential (Ψs) in WD condition was decreased depending on the degree of stress. Leaf Ψs in 34 μM PBZ-pretreated plants of cv. “Tainung 57” was similar to the positive check (cv. “Japanese Yellow”) when subjected to MWD (Figure 3A). In SWD, leaf Ψs in all the treatments significantly declined (Figure 3B). It was confirmed that free proline and soluble sugar enrichment in the PBZ-pretreated sweet potato had a potential to control the osmotic potential in the leaf tissues, preventing the water loss via stomatal closure. In WD conditions, gs in 34 μM PBZ-pretreated plants of cv. “Tainung 57” was stabilized in the same pattern as the positive check (Figures 3C,D).
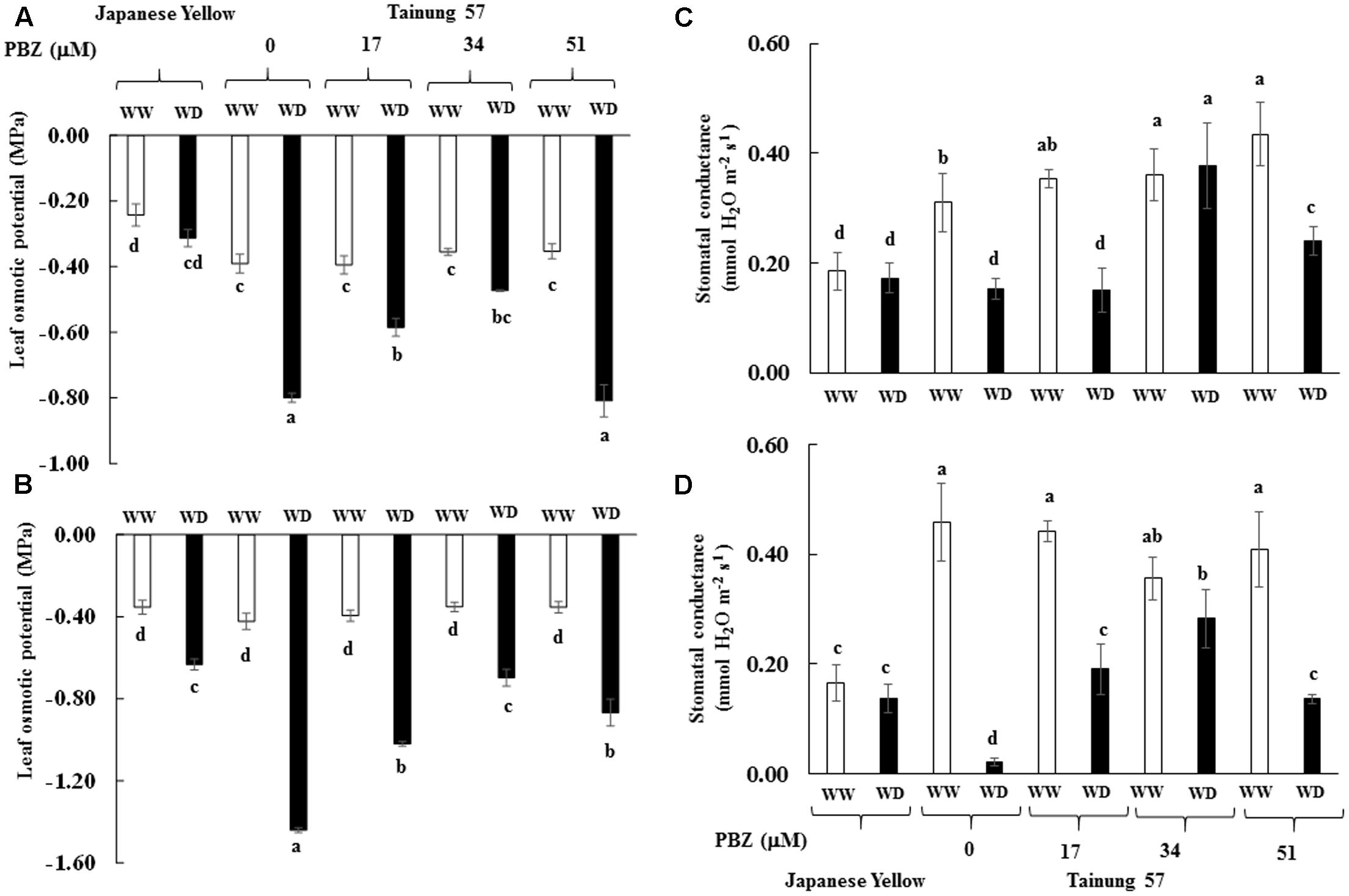
FIGURE 3. Leaf osmotic potential (Ψs) and stomatal conductance (gs) of sweet potato pretreated with PBZ subsequently subjected to mild water deficit (32.3% SWC; 7 days water withholding) (A,C) or SWD (17.5% SWC; 14 days water withholding) (B,D). Different letters in each bar show significant difference at p ≤ 0.01 by Tukey’s HSD. Data presented as mean ± SE.
Photosynthetic pigments, i.e., Chl a, Chl b, and Cx+c, in the positive check cv. ‘Japanese Yellow’ under WD conditions were maintained, whereas those in cv. ‘Tainung 57’ were significantly degraded, especially in SWD. Chl a and Chl b in WD stressed plants of cv. ‘Japanese Yellow’ were maintained (with only 3.84 and 3.65% degradation, respectively) under MWD and declined by 5.25 and 2.27%, respectively, under SWD. Total carotenoid (Cx+c) content in leaf tissues of plants under SWD conditions significantly dropped by 14.32%, but was unaffected under MWD conditions (Table 4). In cv. ‘Tainung 57’, Chl b and Cx+c contents significantly declined by 8.30 and 27.41%, respectively, whereas Chl a was stable. Chl a, Chl b, and Cx+c in plants grown under SWD conditions were significantly reduced by 13.97, 6.64 and 34.63, respectively. The Chl b and Cx+c in 34 μM PBZ (5.32 and 7.98%) and 51 μM PBZ-pretreated plants (4.61 and 9.48%) were alleviated when subjected to MWD. In SWD, Chl a and Chl b in 34 μM PBZ (11.71 and 2.82%, respectively) and 51 μM PBZ-pretreated plants (9.59 and 4.90%, respectively) were maintained, whereas Cx+c, a sensitive indicator, was significantly reduced (Table 3).
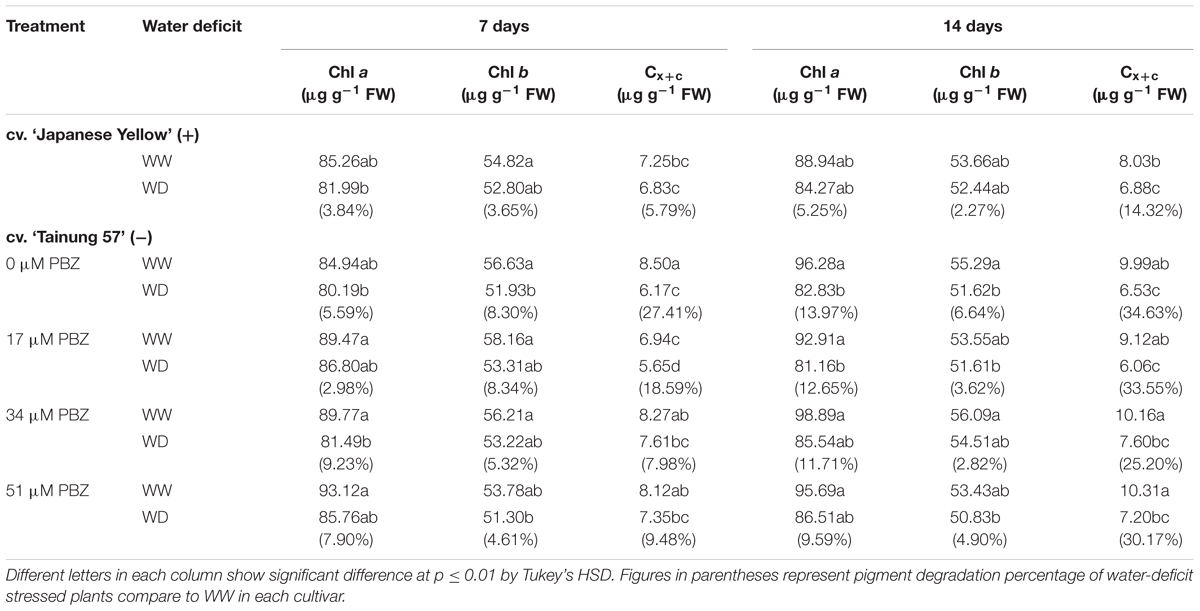
TABLE 4. Chlorophyll a (Chl a), chlorophyll b (Chl b), and total carotenoids (Cx+c) content in leaf tissues of sweet potato pretreated with PBZ subsequently subjected to WW (47.5% SWC), mild water deficit (32.3% SWC; 7 days water withholding) or SWD (17.5% SWC; 14 days water withholding).
Maximum quantum yield of PSII (Fv/Fm) in cv. ‘Tainung 57’ grown under MWD was significantly dropped (13.40% diminution), whereas it was maintained in cv. ‘Japanese Yellow’ (only 2.01% diminution) (Figure 4A). In MWD, Fv/Fm in PBZ-pretreated plants was retained. Similarly, Fv/Fm in 34 and 51 μM PBZ-pretreated plants grown under SWD was maintained (only 2.95 and 1.34% diminution, respectively) (Figure 4B). Photon yield of PSII (ΦPSII) in MWD was maintained, whereas it was declined by 6.8% in 51 mM PBZ-pretreated plants (Figure 4C). In contrast, ΦPSII in 51 mM PBZ-pretreated plants was maintained (1.58% diminution) similar to cv. ‘Japanese Yellow’. A significant diminution of ΦPSII (13.08% reduction) was revealed in cv. ‘Tainung 57’ plants without PBZ-pretreatment (Figure 4D). Pn in cv. ‘Tainung 57’ was lower than that of cv. ‘Japanese Yellow’ and significantly dropped when plants exposed to MWD and SWD (Figures 5A,B). In MWD, the Pn in cv. ‘Japanese Yellow’ was maintained (12.6% reduction), whereas it was decreased by 72.3% in cv. ‘Tainung 57’ plants without PBZ-pretreatment (Figure 5A). Moreover, Pn in cv. ‘Japanese Yellow’ grown under SWD was reduced by 37.09% and it was sharply declined by 93.36% in cv. ‘Tainung 57’ non-treated plants. Interestingly, Pn in 34 mM PBZ-pretreated plants under MWD and SWD was alleviated (with only 41 and 47.5% reduction, respectively) when compared with other treatments (Figures 5A,B). In addition, transpiration rate (E) in cv. ‘Japanese Yellow’ and cv. ‘Tainung 57’ pretreated with 34 mM PBZ subsequently grown under MWD was sustained (with only 6.46% reduction) (Figure 5C). In SWD, E was a sensitive parameter that has significantly declined (Figure 5D). A positive correlation between Chl a content and Fv/Fm (R2 = 0.60), TC content and ΦPSII (R2 = 0.58), ΦPSII and Pn (R2 = 0.73), Pn reduction and plant dry weight (R2 = 0.45) was demonstrated (Figure 6).
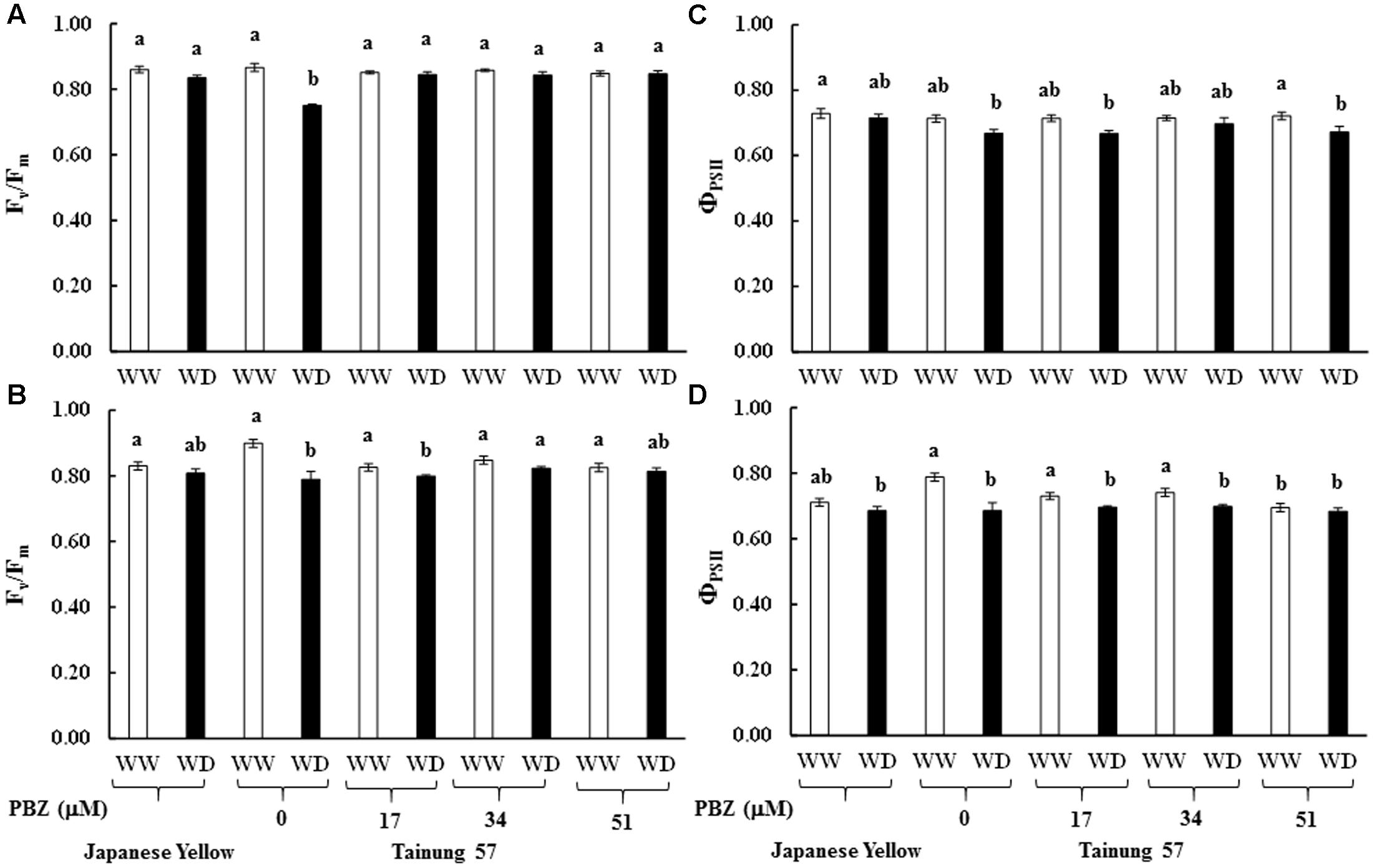
FIGURE 4. Maximum quantum yield of PSII (Fv/Fm) and proton yield of PSII (ΦPSII) of sweet potato pretreated with PBZ subsequently subjected to mild water deficit (32.3% SWC; 7 days water withholding) (A,C) or SWD (17.5% SWC; 14 days water withholding) (B,D). Different letters in each bar show significant difference at p ≤ 0.01 by Tukey’s HSD. Data presented as mean ± SE.
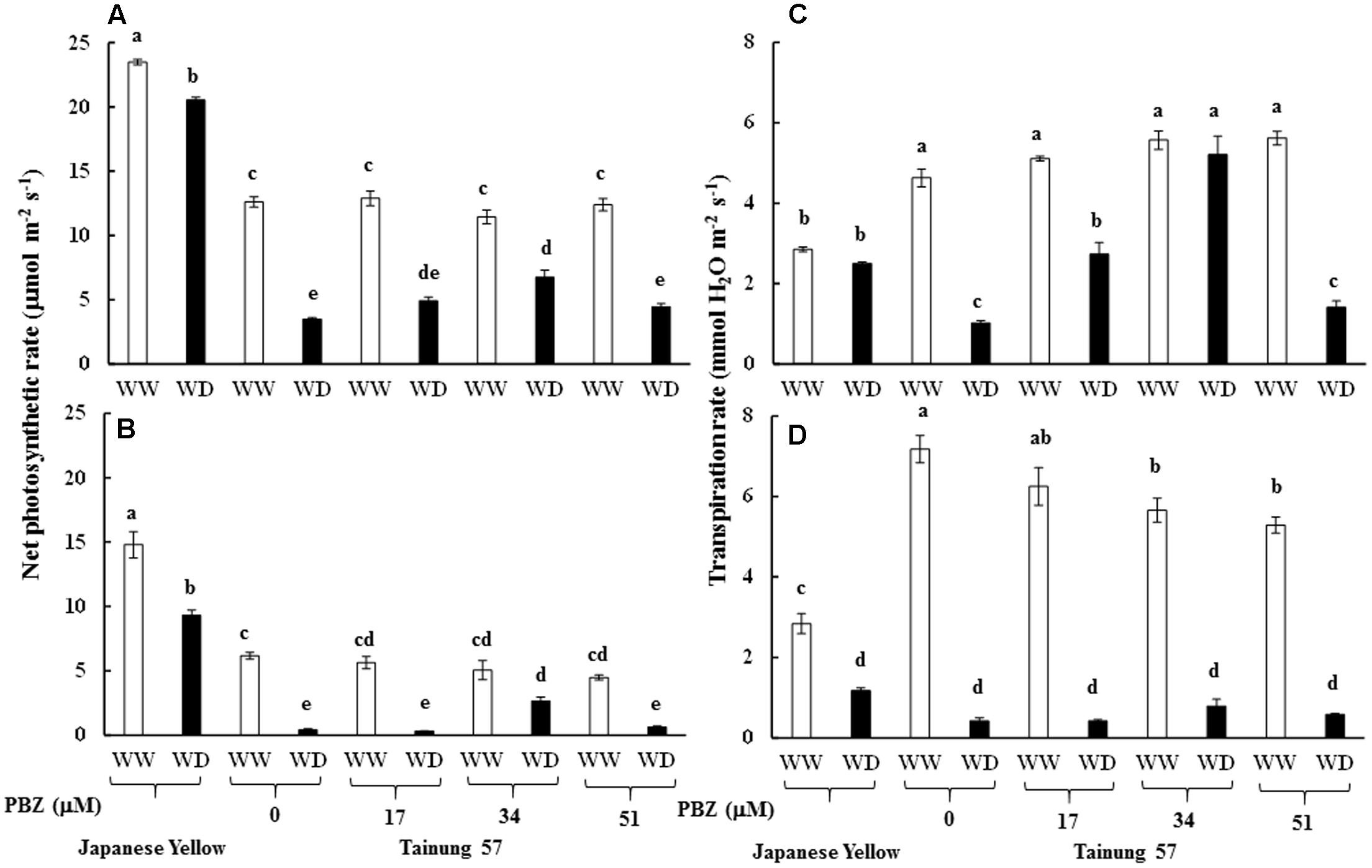
FIGURE 5. Net photosynthetic rate (Pn) and transpiration rate (E) of sweet potato pretreated with PBZ subsequently subjected to mild water deficit (32.3% SWC; 7 days water withholding) (A,C) or SWD (17.5% SWC; 14 days water withholding) (B,D). Different letters in each bar show significant difference at p ≤ 0.01 by Tukey’s HSD. Data presented as mean ± SE.
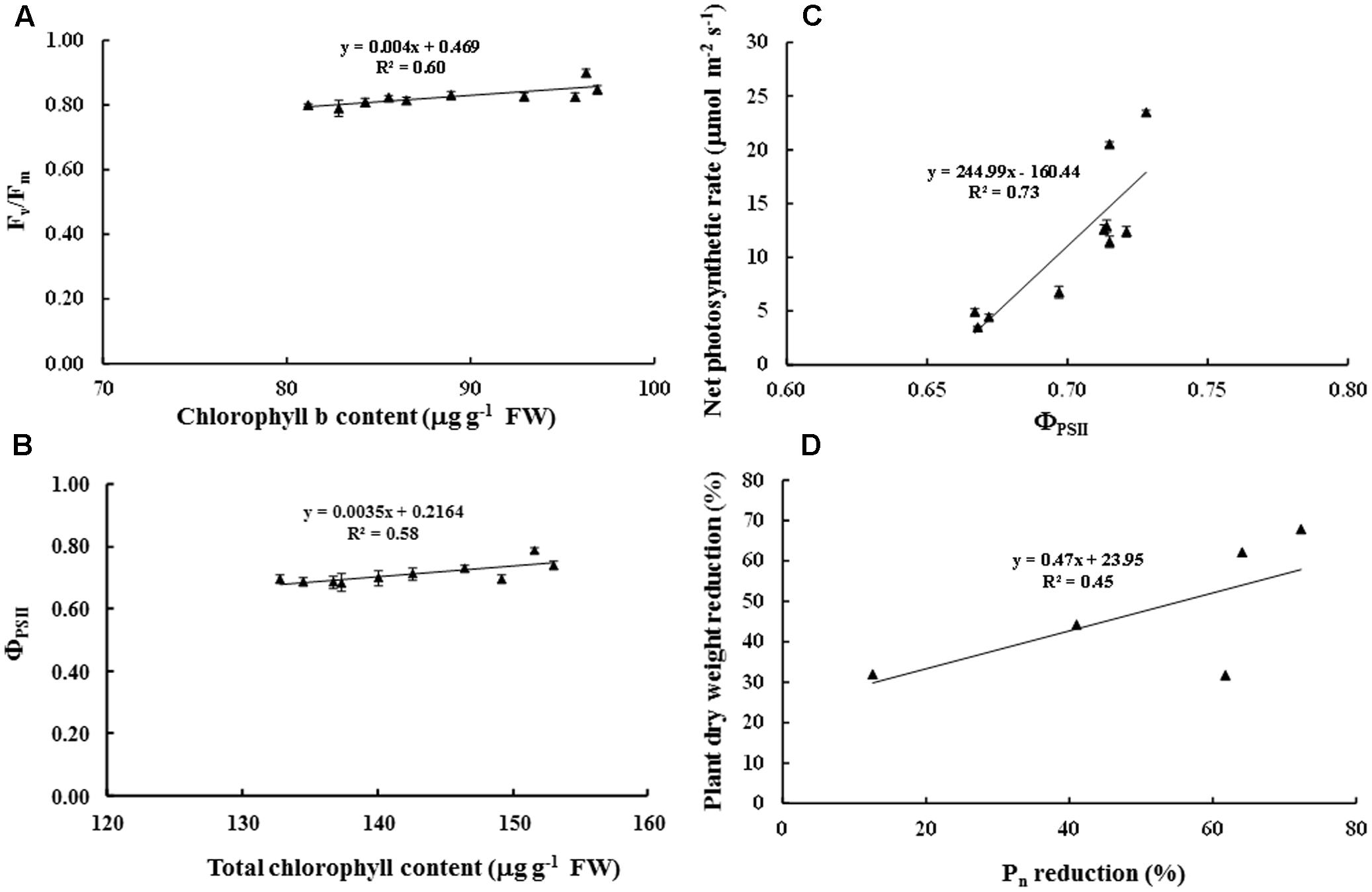
FIGURE 6. Relationship between chlorophyll a content and Fv/Fm (A), total chlorophyll content and ΦPSII (B), ΦPSII and net photosynthetic rate (Pn) (C), and Pn reduction and plant dry weight reduction of sweet potato pretreated with PBZ subsequently subjected to water deficit conditions (D). Data presented as mean ± SE.
Storage root yield of sweet potato was improved by 37 μM PBZ pretreatment, especially in the water shortage at the early growth stage. Drought stress delay the time of flowering in potted red firespike PBZ pretreatment (0.24 mg plant-1 soil drench) improved it and maintained the number of flowers (6 flowers plant-1) equal to control (Rezazadeh et al., 2016). In other studies, fruit yield of tomato (Lycopersicon esculentum Mill. cv. Hybrid Alex 63) pretreated with 50 mg L-1 PBZ was 1.37-folds higher than non-treated plants. This yield declined by 4.79% when the pretreated plants were subjected to WD conditions at 60% field capacity (FC) (Mohamed et al., 2011). In addition, tomato fruit yield (3.89 kg plant-1) and number of fruits (31 fruits plant-1) in 30 mg L-1 PBZ pretreated tomato plants were maintained when exposed to drought condition (Latimer, 1992). Overall, there were enhanced yield performances, in PBZ-pretreated plants than non-pretreated plants under both WW and WD conditions (Table 1).
Root fresh- and dry weight traits in sweet potato were identified as very sensitive to the drought stress, which was improved by PBZ-pretreatment in the present study. Studies on English oak, European beech, and Lombardy poplar tree species also indicate a 4.56-, 3.50-, and 3.67-fold increase in root dry weight, respectively, when treated with 0.5 g PBZ plant-1 (Percival and Sacre, 2014). In WD stress (water withholding for 3 weeks), root dry weight of horse chestnut was recovered (with only 18.4% reduction) by 500 mg L-1 PBZ treatment (Percival and Noviss, 2008). Shoot biomass (mg plant-1) in irrigated finger millets with or without 100 μM PBZ was unchanged. Shoot dry weight (37.17% reduction) and root dry weight (13.04% reduction) of 60 mg L-1 PBZ-treated tomato grown under drought condition were maintained (Latimer, 1992). Moreover, dry weight of 50 mg L-1 PBZ pretreated plants declined by 20.45% as compared to 36.77% reduction in non-treated plants (Bayat and Sepehri, 2012). Shoot dry weight in turf grass, Festuca arundinacea L. cv. Master and Lolium perenne L. cv. Barrage, was very sensitive to the WD conditions (25% FC), resulting in a 95.45 and 97.68% decrease, respectively, whereas this was improved by using 30 mg L-1 PBZ treated seeds (with up to 3.14% reduction) (Shahrokhi et al., 2011).
Paclobutrazol, an anti-GA, has been reported as plant growth retardant. Plant height, a good indicator of PBZ function, in Phillyrea angustifolia was reduced depending on the degree of PBZ concentration in the soil (Fernández et al., 2006). In Pathumma (Curcuma alismatifolia Gagnep. cv. Chiang Mai Pink), shoot height of 3.75 g L-1 PBZ-treated plants was inhibited by 48.93% compared to the untreated plants. In addition, when these plants were treated with 1,500 mg L-1 PBZ (soil drenching) under water withholding conditions for 20 days (5% SWC) and 30 days (4% SWC), the shoot length was maintained when compared to non-treated plants (Jungklang et al., 2016). Likewise, 2.0 mg pot-1 PBZ soil drenching resulted in reduced shoot height in sunflower and zinnia by 26.3 and 42.1%, respectively (Ahmad et al., 2014). Syzygium myrtifolium (Roxb.) Walp. treated with 3.75 g L-1 PBZ had decreased plant height and leaf area by 19.93 and 60.02%, respectively (Roseli et al., 2012). Leaf area of 30 mL L-1 PBZ-pretreated P. angustifolia plants grown under WW conditions was reduced by 83.25%. Therefore, the growth performances of PBZ-pretreated plants were improved when exposed to MWD conditions (I60) and then declined under SWD conditions (I40) (Fernández et al., 2006). Correspondingly, shoot height, root length, and leaf area of 10 mg L-1 PBZ-pretreated peanut plants were improved compared to the plants without PBZ treatment when exposed to drought conditions (Sankar et al., 2014a).
Free proline enrichment in sweet potato cv. ‘Japanese Yellow’ under WD conditions has been well established as a WD tolerance indicator (Yooyongwech et al., 2013). In the WD condition, free proline biosynthesis via pyrroline-5-carboxylate synthetase (P5CS) and pyrroline-5-carboxylate reductase (P5CR) was upregulated to promote proline as major osmoprotectant at the cellular level (Delauney and Verma, 1993; Kishor et al., 2005). In two cultivars of sweet potato (“Untacip” and “Huambachero”), free proline was regulated by WD conditions in both leaf and storage root tissues (Rodríguez-Delfin et al., 2012). In cell suspension culture, free proline in sweet potato cv. ‘Tainung 57’ grown under 0.6 M sorbitol-induced WD conditions was enriched by 5.25-folds over control (Wang et al., 1999). In the present study, free proline content in the leaf tissues of sweet potato cv. ‘Tainung 57’ under MWD conditions were unchanged. In contrast, free proline content in PBZ-pretreated plants under SWD conditions was significantly increased when compared with cv. ‘Japanese Yellow’ without PBZ. This is the first report of proline regulation by a combination of PBZ and water deficiency in sweet potato. It is possible that PBZ regulates the free proline accumulation, especially in combination with WD stress (Hajihashemi and Ehsanpour, 2013). Similarly, free proline content in 50 mg L-1 PBZ treated tomato plants grown under 60% FC peaked at 54.56 mg g-1 FW (1.52-fold over control) (Mohamed et al., 2011). In contrast, free proline content in 10 mg L-1 PBZ-pretreated peanut under WD conditions (1.04-folds over control) was lower than non-treated plants (1.49-folds over control) (Sankar et al., 2014b).
Total soluble sugar enrichment in sweet potato cell culture under 0.6 M sorbitol-induced water deficiency has been well established (Wang et al., 1999). It may function as major osmoprotectant at the cellular level when plants were exposed to WD conditions, leading to control the osmotic potential in the leaf and stomata closure to prevent the water loss (Sánchez et al., 1998; Iannucci et al., 2002). Recent studies indicate that glucose and fructose content in WD stressed sweet potato cultivars “‘Tainung 57”’ and “PROC65-3” increased, whereas sucrose content was unaffected (Yooyongwech et al., 2016). It was confirmed that the activity of acid soluble invertase enzyme was reached threefolds over control to convert sucrose into fructose and glucose when plants were subjected to WD stress (Pelleschi et al., 1997). In addition, fructose, glucose, sucrose, and total soluble sugar in English oak, Lombardy poplar, and European beach treated by 0.5 g PBZ plant-1 soil drench application increased by 1.3- to 2.0-folds over control (Percival and Sacre, 2014). In untreated and PBZ treated (50 mg L-1) WD stressed tomato (60% FC), there was a 1.16- and 1.52-fold increase in total soluble sugars (Mohamed et al., 2011). In 6% PEG-induced WD stress, water soluble carbohydrate and reducing sugars were significantly declined in untreated S. rebaudiana Bertoni, whereas the sugar content was elevated by 2 mg L-1 upon PBZ foliar application (Hajihashemi and Ehsanpour, 2013). Total soluble sugar enrichment in PBZ-pretreated sweet potato may play a vital role in the osmotic adjustment at cellular level of plant under WD conditions. In PBZ-pretreated plants, sugar, a major soluble carbohydrate derived from transitory starch degradation, was evidently observed (Hare et al., 1998; Gupta and Kaur, 2005) that maintains the leaf water potential under drought conditions (Zhu et al., 2004). In strawberry tree, water consumption was reduced by 10% (60 mg PBZ plant-1) and 20% (100 mg PBZ plant-1) compared to the control, causing lower gs and CO2 assimilation after treatment with PBZ (Navarro et al., 2007). It seems to help the plants in acclimatization before the exposure to WD conditions (Cha-um et al., 2009). Under drought conditions, PBZ elevates the gs and water use efficiency in WW conditions as validated in potted red firespike (Rezazadeh et al., 2016), tomato (Pal et al., 2016), and wheat (Dwivedi et al., 2017). It is possible that PBZ improves the stomatal function to prevent the water loss from transpiration when exposed to drought stress (Berova and Zlatev, 2003).
Recent studies indicate that, photosynthetic pigments in sweet potato cultivars, ‘PROC 65-3,’ ‘Japanese Yellow,’ and “Tainung 57” under low SWC were degraded, in relation to the degree of WD stress (Yooyongwech et al., 2014). Similarly, TC in sweet potato cv. Beauregard was reduced depending on the decrease in soil moisture content, while it was stable in cv. Evangeline (Gajanayake et al., 2014). In tomato, TC content in the leaf tissues of 50 mg L-1 PBZ-pretreated plants under 60% FC was maintained (Mohamed et al., 2011). In horse chestnut, TC and Cx+c in plants under WD stress was declined by 69.39 and 65.61%, respectively, and then improved to 55.03 and 55.63%, respectively, on 500 mg L-1 PBZ treatment (Percival and Noviss, 2008). In addition, the improvement of TC and Cx+c in 1-2 mg L-1 PBZ pretreated plants was confirmed in S. rebaudiana grown under 6% polyethylene glycol (PEG)-induced drought condition (Hajihashemi and Ehsanpour, 2013).
Overall photosynthetic abilities, i.e., chlorophyll fluorescence, Pn and E, in PBZ-pretreated sweet potato were improved than those without PBZ. Fv/Fm and Pn in horse chestnut plants under WD conditions declined by 58.02 and 62.95%, respectively, and was recovered by 33.33 and 57.01%, respectively, in the presence of 500 mg L-1 PBZ (Percival and Noviss, 2008). Apart from these parameters, PBZ has been studied to show positive effects on cuticular wax biosynthesis (Jenks et al., 2001), leaf water potential (Zhu et al., 2004; Bañon et al., 2009), water content (Jungklang and Saengnil, 2012; Hajihashemi and Ehsanpour, 2013), and water consumption (Navarro et al., 2007). Hence, PBZ has a cumulative effect in enhancing WD tolerance in plant species (Gilley and Fletcher, 1997; Berova and Zlatev, 2003; Navarro et al., 2009). In present study, a positive relation in physiological (photosynthetic abilities) and growth (plant dry weight) parameters was demonstrated (Figure 6). Similarly, TC concentration in the leaf tissues of Quercus robur and Q. ilex treated with 2 and 4 ppm PBZ foliar spray alleviates in relation to maintain chlorophyll fluorescence (performance index or PI) and Pn, leading to improve survival percentage (Percival and Albalushi, 2007).
Total soluble sugar and free proline in 34 μM PBZ pretreated sweet potato cv. ‘Tainung 57’ may play a critical role in the osmotic adjustment to stabilize photosynthetic pigments, enhance photosynthetic abilities and control water transpiration, leading to sustained growth and storage root yield against WD stress.
This study was designed, directed and coordinated by SC and SY as principal investigator, provided conceptual and technical guidance for all aspects. CT, RT, and TS performed and analyzed the biochemical, physiological, morphological, and yield traits.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
We would like to sincerely thank National Science and Technology Development Agency (NSTDA, Thailand) for funding support and Agricultural Extension Group, Phichit province, Thailand for sweet potato master stock.
The Supplementary Material for this article can be found online at: http://journal.frontiersin.org/article/10.3389/fpls.2017.01400/full#supplementary-material
Ahmad, I., Whipker, B. E., Dole, J. M., and McCall, I. (2014). Paclobutrazol and ancymidol lower water use of potted ornamental plants and plugs. Eur. J. Hortic. Sci. 79, 318–326.
Bañon, S., Miralles, J., Navarro, A., and Sánchez-Blanco, M. J. (2009). Influence of paclobutrazol and substrate on daily evapotranspiration of potted geranium. Sci. Hortic. 122, 572–578.
Bates, L. S., Waldren, R. P., and Teare, I. D. (1973). Rapid determination of free proline for water-stress studies. Plant Soil 39, 205–207. doi: 10.1016/j.dental.2010.07.006
Bayat, S., and Sepehri, A. (2012). Paclobutrazol and salicylic acid application ameliorates the negative effect of water stress on growth and yield of maize plants. J. Res. Agric. Sci. 8, 127–139.
Berova, M., and Zlatev, Z. (2003). Physiological response of paclobutrazol-treated triticale plants to water stress. Biol. Plant. 46, 133–136.
Blum, A. (2005). Drought resistance, water use efficiency, and yield potential are they compatible, dissonant, or mutually exclusive? Aust. J. Agric. Res. 56, 1159–1168.
Cha-um, S., Puthea, O., and Kirdmanee, C. (2009). An effective in-vitro acclimatization using uniconazole treatments and ex-vitro adaptation of Phalaenopsis orchid. Sci. Hortic. 121, 468–473. doi: 10.1016/j.scienta.2009.02.027
Cha-um, S., Supaibulwatana, K., and Kirdmanee, C. (2007). Glycinebetaine accumulation, physiological characterizations and growth efficiency in salt-tolerant and salt-sensitive lines of indica rice (Oryza sativa L. ssp. indica) in response to salt stress. J. Agron. Crop Sci. 193, 157–166.
Chehelpar, N., Tohidi-Moghadam, H. R., and Ghoushchi, F. (2016). Hexaconazole foliar application alleviates water deficit effects in common bean. Pesq. Agropec. Trop. 46, 301–310. doi: 10.1590/1983-40632016v4641432
Delauney, A. J., and Verma, D. P. S. (1993). Proline biosynthesis and osmoregulation in plants. Plant J. 4, 215–223. doi: 10.1046/j.1365-313X.1993.04020215.x
Dwivedi, S. K., Arora, A., and Kumar, S. (2017). Paclobutrazol-induced alleviation of water-deficit damage in relation to photosynthetic characteristics and expression of stress markers in contrasting wheat genotypes. Photosynthetica 55, 351–359. doi: 10.1007/s11099-016-0652-5
Falkenmark, M. (2013). Growing water scarcity in agriculture: future challenge to global water security. Philos. Trans. R. Soc. A 371:20120410. doi: 10.1098/rsta.2012.0410
Fernández, J. A., Belenzategui, L., Bañón, S., and Franco, J. A. (2006). Induction of drought tolerance by paclobutrazol and irrigation deficit in Phillyrea angustifolia during the nursery period. Sci. Hortic. 107, 277–283. doi: 10.1016/j.scienta.2005.07.008
Gajanayake, B., Reddy, K. R., Shankle, M. W., and Arancibia, R. A. (2014). Growth, developmental, and physiological responses of two sweet potato (Ipomoea batatas L. [Lam.]) cultivars to early season soil moisture deficit. Sci. Hortic. 168, 218–228.
Gilley, A., and Fletcher, R. A. (1997). Relative efficacy of pacloburtrazol, propiconazole and tetraconazole as stress protectants in wheat seedlings. Plant Growth Regul. 21, 169–175. doi: 10.1023/A:1005804717016
Gomes, F., and Carr, M. K. V. (2001). Effects of water availability and vine harvesting frequency on the productivity of sweet potato in Southern Mozambique. I. Storage root and vine yields. Exp. Agric. 37, 523–537.
Gomes, F., and Carr, M. K. V. (2003). Effects of water availability and vine harvesting frequency on the productivity of sweet potato in Southern Mozambique. III. Crop yield and water use response functions. Exp. Agric. 39, 409–421.
Gomes, F., Carr, M. K. V., and Squire, G. R. (2005). Effects of water availability and vine harvesting frequency on the productivity of sweet potato in Southern Mozambique. IV. Radiation interception, dry matter production and partitioning. Exp. Agric. 41, 93–108.
Gupta, A. K., and Kaur, N. (2005). Sugar signalling and gene expression in relation to carbohydrate metabolism under abiotic stresses in plants. J. Biosci. 30, 761–776. doi: 10.1007/BF02703574
Hajihashemi, S., and Ehsanpour, A. A. (2013). Influence of exogenously applied paclobutrazol on some physiological traits and growth of Stevia rebaudiana under in vitro drought stress. Biologia 68, 414–420. doi: 10.2478/s11756-013-0165-7
Hajihashemi, S., and Ehsanpour, A. A. (2014). Antioxidant response of Stevia rebaudiana B. to polyethylene glycol and paclobutrazol treatments under in vitro culture. Appl. Biochem. Biotechnol. 172, 4038–4052. doi: 10.1007/s12010-014-0791-8
Hare, P. D., Cress, W. A., and van Staden, J. (1998). Dissecting the roles of osmolyte accumulation during stress. Plant Cell Environ. 21, 535–553. doi: 10.1046/j.1365-3040.1998.00309.x
Iannucci, A., Russo, M., Arena, L., di Fonzo, N., and Martiniello, P. (2002). Water deficit effects on osmotic adjustment and solute accumulation in leaves of annual clovers. Eur. J. Agron. 16, 111–122. doi: 10.1016/S1161-0301(01)00121-6
Jenks, M. A., Andersen, L., Teusink, R. S., and Williams, M. H. (2001). Leaf cuticular waxes of potted rose cultivars as affects by plan development, drought and paclobutrazol treatments. Physiol. Plant. 112, 62–70.
Jungklang, J., and Saengnil, K. (2012). Effect of paclobutrazol on pathumma cv. Chiang Mai Pink under water stress. Songklanakarin J. Sci. Technol. 34, 361–366.
Jungklang, J., Saengnil, K., and Uthaibutra, J. (2016). Effects of water-deficit stress and paclobutrazol on growth, relative water content, electrolyte leakage, proline content and some antioxidant changes in Curcuma alismatifolia Gagnep. cv. Chiang Mai Pink. Saudi J. Biol. Sci. (in press). doi: 10.a016/j.sjbs.2015.09.017
Karkacier, M., Ebras, M., Uslu, M. K., and Aksu, M. (2003). Comparison of different extraction and detection methods for sugars using amino-bonded phase HPLC. J. Chromatogr. Sci. 41, 331–333. doi: 10.1093/chromsci/41.6.331
Kishor, P. B. K., Sangam, S., Amrutha, R. N., Laxmi, P. S., Naidu, K. R., Rao, R. R. S. S., et al. (2005). Regulation of proline biosynthesis, degradation, uptake and transport in higher plants: its implications in plant growth and abiotic stress tolerance. Curr. Sci. 88, 424–438.
Lanfermeijer, F. C., Koerselman-Kooij, J. W., and Borstlap, A. C. (1991). Osmosensitivity of sucrose uptake by immature pea cotyledons disappears during development. Plant Physiol. 95, 832–838. doi: 10.1104/pp.95.3.832
Latimer, J. G. (1992). Drought, paclobutrazol, abscisic acid, and gibberellic acid as alternatives to daminozide in tomato transplant production. J. Am. Soc. Hortic. Sci. 117, 243–247.
Lichtenthaler, H. K. (1987). Chlorophylls and carotenoids: pigments of photosynthetic biomembranes. Method Enzymol. 148, 350–380. doi: 10.1016/0076-6879(87)48036-1
Lin, K. H., Pai, F. H., Hwang, S. Y., and Lo, H. F. (2006a). Pre-treating paclobutrazol enhanced chilling tolerance of sweet potato. Plant Growth Regul. 49, 249–262.
Lin, K. H., Tsou, C. C., Hwang, S. Y., Chen, L. F. O., and Lo, H. F. (2006b). Paclobutrazol pre-treatment enhanced flooding tolerance of sweet potato. J. Plant Physiol. 163, 750–760.
Loggini, B., Scartazza, A., Brugnoli, E., and Navari-Izzo, F. (1999). Antioxidant defense system, pigment composition, and photosynthetic efficiency in two wheat cultivars subjected to drought. Plant Physiol. 119, 1091–1099. doi: 10.1104/pp.119.3.1091
Manivannan, P., Jaleel, C. A., Somasundaram, R., and Panneerselvam, R. (2008). Osmoregulation and antioxidant metabolism in drought-stressed Helianthus annuus under triadimefon drenching. C R Biol. 331, 418–425. doi: 10.1016/j.crvi.2008.03.003
Maxwell, K., and Johnson, G. N. (2000). Chlorophyll fluorescence-a practical guide. J. Exp. Bot. 51, 659–668.
Mohamed, G. F., Agamy, R. A., and Rady, M. M. (2011). Ameliorative effects of some antioxidants on water-stressed tomato (Lycopersicon esculentum Mill.) plants. J. Appl. Sci. Res. 7, 2470–2478.
Navarro, A., Sánchez-Blanco, M. J., and Bañon, S. (2007). Influence of paclobutrazol on water consumption and plant performance of Arbutus unedo seedlings. Sci. Hortic. 111, 133–139. doi: 10.1016/j.scienta.2006.10.014
Navarro, A., Sánchez-Blanco, M. J., Morte, A., and Bañon, S. (2009). The influence of mycorrhizal inoculation and paclobutrazol on water and nutritional status of Arbutus unedo L. Environ. Exp. Bot. 66, 362–371. doi: 10.1016/j.envexpbot.2009.04.005
Neumann, P. M. (2008). Coping mechanisms for crop plants in drought-prone environments. Ann. Bot. 101, 901–907. doi: 10.1093/aob/mcn018
Pal, S., Zhao, J., Khan, A., Yadav, N. S., Batushansky, A., Barak, S., et al. (2016). Paclobutrazol induces tolerance in tomato to deficit irrigation through diversified effects on plant morphology, physiology and metabolism. Sci. Rep. 6:39321. doi: 10.1038/srep39321
Pelleschi, S., Rocher, J. P., and Prioul, J. L. (1997). Effect of water restriction on carbohydrate metabolism and photosynthesis in mature maize leaves. Plant Cell Environ. 20, 493–503. doi: 10.1046/j.1365-3040.1997.d01-89.x
Percival, G. C., and Albalushi, A. M. S. (2007). Paclobutrazol-induced drought tolerance in contaminerized English and Evergreen oak. Aboricul. Urban For. 33, 397–409.
Percival, G. C., and Noviss, K. (2008). Triazole induced drought tolerance in horse chesnut (Aesculus hippocastanum). Tree Physiol. 28, 1685–1692. doi: 10.1093/treephys/28.11.1685
Percival, G. C., and Sacre, K. (2014). The influence of soluble carbohydrates, slow release nitrogen and a plant growth regulator on transplant survival of trees. Aboricult. J. 36, 140–160. doi: 10.1080/03071375.2014.943559
Rezazadeh, A., Harkess, R. L., and Bi, G. (2016). Effects of paclobutrazol and flurprimidol on water stress amelioration in potted red firespike. HortTechnology 26, 26–29.
Rockstrom, J. (2003). Water for food and nature in drought-prone tropics: vapour shift in rain-fed agriculture. Philos. Trans R. Soc. B 358, 1997–2009. doi: 10.1098/rstb.2003.1400
Rodríguez-Delfin, A., Posadas, A., León-Velarde, C., Mares, V., and Quiroz, R. (2012). Effect of salt and water stress on the proline and total chlorophyll content and nutrients uptake on two sweet potato cultivars grown on soilless culture. Acta Hortic. 947, 55–62. doi: 10.17660/ActaHortic.2012.947.4
Roseli, A. N. M., Ying, T. F., and Ramlan, M. F. (2012). Morphological response of Syzygium myrtifolium (Roxb.) Walp. to paclobutrazol. Sans Malays. 41, 1187–1192.
Sánchez, F. J., Manzanares, M., de Andres, E. F., Tenorio, J. L., and Ayerbe, L. (1998). Turgor maintenance, osmotic adjustment and soluble sugar and proline accumulation in 49 pea cultivars in response to water stress. Field Crop Res. 59, 225–235. doi: 10.1016/S0378-4290(98)00125-7
Sankar, B., Gopinathan, P., Karthishwaran, K., and Somasundaram, R. (2014a). Variation in growth of peanut plants under drought stress condition and in combination with paclobutrazol and abscisic acid. Curr. Bot. 5, 14–21.
Sankar, B., Gopinathan, P., Karthishwaran, K., and Somasundaram, R. (2014b). Biochemical content variation in Arachis hypogaea under drought stress with or without paclobutrazol and abscisic acid. J. Ecobiotechnol. 6, 9–14.
Sankar, B., Jaleel, C. A., Manivannan, P., Kishorekumar, A., Somasundaram, R., and Panneerselvam, R. (2007). Effect of paclobutrazol on water stress amelioration through antioxidants and free radical scavenging enzymes in Arachis hypogaea L. Colloids Surf. B Biointerfaces 60, 229–235. doi: 10.1016/j.colsurfb.2007.06.016
Sankar, B., Karthishwaran, K., and Somasundaram, R. (2013). Leaf anatomy changes in peanut plants in relation to drought stress with or without paclobutrazol and abscisic acid. J. Phytol. 5, 25–29.
Shabala, S. N., Shabala, S. I., Martynenko, A. I., Babourina, O., and Newman, I. A. (1998). Salinity effect on bioelectric activity, growth, Na+ accumulation and chlorophyll fluorescence of maize leaves: a comparative survey and prospects for screening. Aust. J. Plant Physiol. 25, 609–616. doi: 10.1071/PP97146
Shahrokhi, M., Tehranifar, A., Hadizadeh, H., and Selahvarzi, Y. (2011). Effect of drought stress and paclobutrazol-treated seeds on physiological response of Festuca arundinacea L. Master and Lolium perenne L. Barrage. J. Biol. Environ. Sci. 5, 77–85.
van Heeden, P. D. R., and Laurie, R. (2008). Effects of prolonged restriction in water supply on photosynthesis, shoot development and storage root yield in sweet potato. Physiol. Plant. 134, 99–109. doi: 10.1111/j.1399-3054.2008.01111.x
Wang, H. L., Lee, P. D., Chen, W. L., Huang, D. J., and Su, J. C. (2000). Osmotic stress-induced changes of sucrose metabolism in cultured sweet potato cells. J. Exp. Bot. 51, 1991–1999. doi: 10.1093/jexbot/51.353.1991
Wang, H. L., Lee, P. D., Liu, L. F., and Su, J. C. (1999). Effect of sorbitol induced osmotic stress on the changes of carbohydrate and free amino acid pools in sweet potato cell suspension cultures. Bot. Bull. Acad. Sin. 40, 219–225.
Yooyongwech, S., Samphumphuang, T., Theerawitaya, C., and Cha-um, S. (2014). Physio-morphological responses of sweet potato [Ipomoea batatas L. (Lam.)] genotypes to water-deficit stress. Plant Omics J. 7, 361–368.
Yooyongwech, S., Samphumphuang, T., Tisarum, R., Theerawitaya, C., and Cha-um, S. (2016). Arbuscular mycorrhizal fungi (AMF) improved water deficit tolerance in two different sweet potato genotypes involves osmotic adjustment via soluble sugar and free proline. Sci. Hortic. 198, 107–117. doi: 10.1016/j.scienta.2015.11.002
Yooyongwech, S., Theerawitaya, C., Samphumphuang, T., and Cha-um, S. (2013). Water-deficit tolerant identification in sweet potato genotypes [Ipomoea batatas (L.) Lam.] in vegetative developmental stage using multivariate physiological indices. Sci. Hortic. 162, 242–251.
Zhang, M., Duan, L., Tian, X., He, Z., Li, J., Wang, B., et al. (2007). Uniconazole-induced tolerance of soybean to water deficit stress in relation to changes in photosynthesis, hormones and antioxidant system. J. Plant Physiol. 164, 709–717. doi: 10.1016/j.jplph.2006.04.008
Keywords: sweet potato, paclobutrazol, water deficit, pigment, proline, total soluble sugar
Citation: Yooyongwech S, Samphumphuang T, Tisarum R, Theerawitaya C and Cha-um S (2017) Water-Deficit Tolerance in Sweet Potato [Ipomoea batatas (L.) Lam.] by Foliar Application of Paclobutrazol: Role of Soluble Sugar and Free Proline. Front. Plant Sci. 8:1400. doi: 10.3389/fpls.2017.01400
Received: 25 October 2016; Accepted: 27 July 2017;
Published: 08 August 2017.
Edited by:
Partha Sarathi Basu, Indian Institute of Pulses Research, IndiaReviewed by:
Koushik Chakraborty, Indian Council of Agricultural Research (ICAR), IndiaCopyright © 2017 Yooyongwech, Samphumphuang, Tisarum, Theerawitaya and Cha-um. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Suriyan Cha-um, c3VyaXlhbmNAYmlvdGVjLm9yLnRo
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.