
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Plant Sci. , 16 June 2017
Sec. Technical Advances in Plant Science
Volume 8 - 2017 | https://doi.org/10.3389/fpls.2017.01033
 María L. Giachero1
María L. Giachero1 Nathalie Marquez1,2
Nathalie Marquez1,2 Adrien Gallou3
Adrien Gallou3 Celina M. Luna4
Celina M. Luna4 Stéphane Declerck5
Stéphane Declerck5 Daniel A. Ducasse1*
Daniel A. Ducasse1*In this work, we described an in vitro system adequate for investigating the pathosystem soybean/arbuscular mycorrhizal fungi (AMF)/Fusarium virguliforme. Pre-mycorrhized plantlets with Rhizophagus irregularis were infected by F. virguliforme either locally via a plug of gel supporting mycelium (Method 1) or via a macroconidia suspension applied to the medium surface (Method 2). Root colonization by the AMF and infection by the pathogen were similar to the usual observations in pot experiments. Within a period of 18 days, more than 20% of the roots were colonized by the AMF and infection by the pathogen was observed in all the plants. In presence of AMF, a decrease in symptoms and in the level of root tissue infection was noticed. With Method 1, smaller necrotic lesions were observed in the pre-mycorrhized plantlets. In Method 2, pathogen infection was slower but more homogenous. These results demonstrated the suitability of the in vitro cultivation system to study the pathosystem soybean/AMF/F. virguliforme. We propose this in vitro cultivation system for studying the mechanisms involved in the biocontrol conferred by AMF against F. virguliforme in soybean.
Studies on the belowground plant–microbes interaction have increased tremendously in number and complexity in recent years (Brink, 2016). During the last decade, this increasing interest resulted in the development of several innovative techniques to investigate the intricate rhizosphere community (Oburger and Schmidt, 2016). Soil is now viewed as a complex universe where plants interact with a wide range of microbes and minerals, resulting in a true system where nothing can be modified without affecting everything else (Young and Crawford, 2004). Many of the microorganisms that are normally present in the soil, actually inhabit the rhizosphere and interact with plants. Those plant–microorganisms interactions may be beneficial or harmful. Among the first are the arbuscular mycorrhizal fungi (AMF). These soil fungi have been reported to improve plant resistance/tolerance to pests and diseases (Gutjahr and Parniske, 2013). AMF are obligate root symbionts that rely on their host for their carbohydrates and as a consequence, it is impossible to grow them in axenic conditions, i.e., independently from a suitable host plant. Nevertheless, in the last decades, a number of cultivation systems have been developed to grow them in vitro with root organs (Gutjahr and Parniske, 2013) or whole plants (Voets et al., 2005, 2009; Dupré de Boulois et al., 2006; Koffi et al., 2009). These in vitro systems have allowed the in-deep study of the plant-AMF symbiotic association (Koffi et al., 2013).
Soil borne pathogens represent a threat to agriculture generating important yield losses, depending upon the pathogen and the crop (Oerke, 2006). One example is the “Sudden Death Syndrome” (SDS), a severe disease in soybean caused by a complex of at least four species of Fusarium sp., among which Fusarium virguliforme and F. tuccumaniae are the most prevalent in Argentina (O’Donnell et al., 2010).
Fusarium virguliforme causes root rot as well as vascular discoloration of roots and stems. Root infection is often accompanied by foliar symptoms characterized, in the early stages of infection, by interveinal chlorosis followed at the later stages by necrosis and, in the most severe cases, by flower and pod abscission (Rupe, 1989) and defoliation (Genre et al., 2005). In the worst situations, SDS can cause up to 90% yield losses has observed in some areas of Argentina (Genre et al., 2005). Symptoms may be more severe on leaves, but reduction in yield is mostly attributed to both root infection and foliar symptoms (Rupe et al., 1989; Genre et al., 2008).
Recently, Bressano et al. (2010) developed an in vitro system to infect soybean plantlets with Macrophomina phaseolina, a fungal soil-borne pathogen. This system allowed the study of the pre-penetration and penetration process of the pathogen within the roots as well as the early responses of the host plant to infection. Similarly, the tripartite interactions, involving a mycorrhizal plant and a leaf pathogen (potato/AMF with the oomycete Phytophthora infestans) (Gallou et al., 2011) or nematodes (banana/AMF with the nematode Radopholus similis) (Koffi et al., 2013) and even with beneficial microorganisms [e.g., barrel medic/AMF with Trichoderma harzianum (De Jaeger et al., 2011)] have been developed. Gallou (2011) also succeeded in the tripartite association between mycorrhizal potato plantlets and the root pathogen Rhizoctonia solani. Using this model, the author was able to investigate the molecular mechanisms involved in the increased resistance of the pre-mycorrhized potato plantlets to this soilborne pathogen.
In vitro cultivation systems offers a number of advantages over pot culture experiments which are the absence of unwanted contaminants and the possibility for highly controlled and non-destructive dynamical observations of the interaction between AMF and pathogens (Declerck et al., 2005).
At present, there is no described methodology for the in vitro three-way interaction between soybean plantlets, an AMF and a soil-borne fungus. The development of such a system would facilitate comprehension of the mechanisms involved in the increased resistance to F. virguliforme in AMF-colonized soybean plantlets.
In the present study we reported and described, for the first time, an in vitro cultivation system associating pre-mycorrhized soybean plantlets infected by F. virguliforme. We also monitored and compared the early steps of F. virguliforme infection progress in presence/absence of the AMF and proposed some reasons to the differences observed.
A strain of Rhizophagus irregularis (Błaszk., Wubet, Renker & Buscot) C. Walker & A. Schüßler as [‘irregulare’] MUCL 41833 was provided by the Glomeromycota in vitro collection1. The AMF was grown in association with Ri T-DNA transformed carrot (Daucus carota L.) roots clone DC1 on Petri plates (90 mm diameter) containing the modified Strullu-Romand (MSR) medium (Declerck et al., 1998) solidified with 3 g L-1 Phytagel (Sigma–Aldrich, St. Louis, United States), following the method detailed in (Cranenbrouck et al., 2005). The Petri plates were incubated in the dark in an inverted position at 27°C during several months until thousands of spores were obtained.
A strain of F. virguliforme O’Donnell & T. Aoki MUCL 53605, originally isolated from soybean (Glycine max) in Argentina (Buenos Aires, San Pedro) (Aoki et al., 2005) was supplied by the Mycothèque de l’Université catholique de Louvain (MUCL2). A plug of gel containing several macroconidia and mycelium was placed on 50 ml Potato Dextrose Agar (PDA) (Scharlau Chemie S.A., Barcelona, Spain) in Petri plates (90 mm diameter). The Petri plates were incubated at 25°C in the dark for 7 days.
Seeds of Medicago truncatula Gaertn. cv. Jemalong A 17 (SARDI, Australia) were surface-disinfected by immersion in calcium hypochlorite (3.5% active calcium) for 12 min, rinsed three time in deionized sterilized (121°C for 15 min) water and germinated in groups of 15 on Petri plates (90 mm diameter) filled with 35 ml MSR medium without sucrose and vitamins, and solidified with 3 g L-1 Phytagel. The Petri plates were incubated at 27°C in the dark for 4 days and subsequently exposed to light for 1 day, before use.
Seeds of soybean [G. max (L.) Merr.] cv DON MARIO 4800 were surface-disinfected by immersion in ethanol 70% for 1 min, followed by calcium hypochlorite (10% active calcium) for 2 min, rinsed three time in deionized sterilized (121°C for 15 min) water and germinated in groups of 6 on Petri plates (145 mm diameter) filled with 60 ml MSR medium without sucrose and vitamins, and solidified with 3 g L-1 Phytagel. The Petri plates were incubated at 27°C in the dark for 4 days, after exposition to light [average photosynthetic photon flux (PPF) of 225 μmol m-2 s-1] for 24 h.
The Mycelium Donor Plant (MDP) in vitro culture system developed by Voets et al. (2009) and adapted by Anene et al. (2013) for banana was used. Briefly, the cover of a 55 mm diameter Petri plate (named root compartment, RC) was introduced in the base of a 145 mm diameter Petri plate (named hyphal compartment, HC) (Figure 1A). The RC and HC were filled with 20 and 100 ml MSR medium lacking vitamins and sucrose and solidified with 3 g L-1 Phytagel, respectively. The RC was leaned to the border of the HC and a small opening (±2 mm diameter) was made in the base and the lid of the 145 mm diameter Petri plate. One 4-day-old M. truncatula seedling was transferred in the RC with the roots plated on the MSR medium and shoot extending outside the Petri plate via the hole. A plug (9 mm × 5 mm) of MSR medium containing ± 100 spores of R. irregularis MUCL 41833 was placed in the vicinity of the roots. The system was then sealed with Parafilm (Pechiney, Plastic Packaging, Chicago, IL, United States) and the hole plastered with sterilized (121°C for 15 min) silicon grease (VWR International, Belgium) to avoid contaminations (Figure 1A). The systems were transferred in a growth chamber set at 20/18°C (day/night), 70% relative humidity, with a photoperiod of 16 h d-1 and a PPF of 300 μmol m-2 s-1. Except from the aerial part of M. truncatula plants, the rest of the system was covered with an opaque plastic bag to keep the AMF and M. truncatula roots in the dark.

FIGURE 1. Mycorrhizal Donor Plant (MDP) in vitro culture system for the mycorrhization of Medicago truncatula (Mt) and subsequent infection by Fusarium virguliforme [adapted from Voets et al. (2009) and Anene et al., 2013]. (A) Bi-compartmented in vitro culture system with a root compartment (RC) and a hyphal compartment (HC). A plant of M. truncatula (15-days-old) is inserted in the RC and associated to the arbuscular mycorrhizal fungus (AMF) Rhizophagus irregularis MUCL 41833. Bar = 1 cm. (B) Soybean plantlet (7-days-old) inserted in the HC, with the roots in direct contact with the extraradical mycelium of R. irregularis – Bar = 1 cm. (C) Root (R) of soybean plants (15-days-old) infected with a plug of Potato Dextrose Agar (PDA) (25 mm2) covered with F. virguliforme (method 1). Bar = 1 cm. (D) Roots of soybean plantlets (15-days-old) infected with a 2 ml suspension of 1.52 × 106 macroconidia/ml of F. virguliforme (method 2) – Bar = 1 cm.
After 6 weeks of culture, the mycelium crossed the plastic barrier separating the RC from the HC and a profuse extra-radical mycelium (ERM) network bearing numerous spores developed in the HC. Two weeks later, a new hole was made in the base and the lid of the systems, at the opposite side of M. truncatula, to allow the insertion of a 5-days old soybean plantlet with the roots in direct contact with the ERM and shoot extending outside the Petri plate (Figure 1B). Simultaneously soybean plantlets were inserted in the MDP in vitro culture systems as above but in the absence of AMF (i.e., the control). The systems were sealed carefully and incubated horizontally in a growth chamber under the same conditions as above (with an opaque plastic bag to keep the roots in the dark).
After 2 weeks of contact between the soybean roots and the ERM, the RC (i.e., the cover of the 55 mm diameter Petri plate containing the M. truncatula plantlet) was removed and replaced by 25 ml MSR medium without sucrose and vitamins and solidified with 3 g L-1 Phytagel. At that moment, the systems were inoculated with either a plug of PDA (25 mm2) covered by the pathogen (referred as Method 1 – Figure 1C) or with 2 ml suspension of 1.52 × 106 macroconidia ml-1 in sterile water (referred as Method 2 – Figure 1D). In both inoculation methods, the systems were divided randomly into four treatments, each with three replicates: pre-mycorrhized soybean plantlets with/without F. virguliforme (+AMF+Fv, +AMF-Fv) and non-mycorrhizal soybean plantlets with/without F. virguliforme (-AMF+Fv, -AMF-Fv).
Roots were harvested 48 h after contact of the first hyphae of the pathogen with the root surface (i.e., 96 h after inoculation) for Method 1 or after 72 h post-inoculation for Method 2. Colonization by R. irregularis and damage caused by the pathogen were subsequently assessed.
Symptoms on roots and leaves were monitored just before plantlets were harvested (i.e., 96 h and 72 h after inoculation by the pathogen for Method 1 and 2, respectively).
The evolution of root necrosis differed according to the pathogen inoculation method. In Method 1, the size of the necrotic area at the point where the hyphae of the pathogen contacted the root surface was monitored and compared between mycorrhizal and not mycorrhizal plantlets. In Method 2, the entire root system was observed at 24, 48, and 72 h. Independently of the method of inoculation with the pathogen, the symptoms on leaves were also visually monitored.
Root colonization by AMF was estimated 96 h (Method 1) and 72 h (Method 2) after inoculation by the pathogen. Within such short period it was estimated that the pathogen did not impact the root colonization parameters of the AMF and consequently no colonization analysis was conducted prior to infection by the pathogen.
Roots were cleared in 10% KOH at room temperature for 3 h, rinsed with distilled water, bleached and acidified with HCl 1% and stained with Trypan blue 0.2% at room temperature for 15 min. The percentage of root colonization was subsequently estimated according to McGonigle et al. (1990). Two hundred intersections were observed under a compound microscope (Olympus SZ40, Olympus Optical GmbH, Hamburg, Germany) at 10–40× magnification. Total root colonization (%RC), abundance of arbuscules (%A), and intraradical spores/vesicles (%V) were determined. At each intersection, a mark was made if the vertical crosshair crosses an arbuscule, vesicle or hyphae. Intersections were counted in the following categories: “negative” (i.e., no AM fungus observed in root), “hyphae only,” “arbuscules” and “vesicles.” If the vertical crosshair crossed one or more arbuscules or vesicles, the category was incremented by one, and similarly for intersections where hyphae only were observed. When both arbuscules and vesicles were visualized at an intersection, the total number of intersections was only added by one. The arbuscular colonization and vesicles colonization were calculated by dividing the count for the “arbuscules” and “vesicles” categories, respectively, by the total number of intersections examined. Hyphal colonization was calculated as the proportion of non-negative intersections.
Infection by F. virguliforme was confirmed under the microscope (Nikon labophot-2, Japan) at 10–40× magnification and the estimation of root infection was done following staining with Trypan blue 0.2% as above. The estimation of the level of roots infection by F. virguliforme was done following the methodology described by McGonigle et al. (1990) slightly modified. For each treatment, 10 mm long root pieces were selected randomly, mounted in glycerine on microscope slides and covered with 40 × 22 mm coverslips. Between 2 and 4 slides, each covered with 10 pieces of roots, were used per replicate. Roots were aligned parallel to the long axis of the slide and observed as follows under a magnification of 40×: to examine each intersection, the plane of focus was moved completely through the root and whether the vertical crosshair actually cuts any hyphae, were considered like positive. Intersections were counted in the following categories; ‘absence of hyphae’ (no fungal material in root), and ‘presence of hyphae.’ Hyphal colonization was calculated as the proportion of non-negative intersections.
When pre-mycorrhized F. virguliforme infected roots were examined, both mycelia were clearly distinguished by their morphology and growing pattern. While R. irregularis grew displaying an intercellular net-like pattern with well-stained cell walls, the pathogen showed a thinner and more transparent hyphae and straighter and linear growing pattern. The percentage of infected roots was estimated as the ratio between infected roots and total number of roots examined.
To determine whether the cells of a brownish tissue was dead or not, an FDA-PI staining was performed on detached roots (see Supplementary Material).
The experiment was repeated twice. Percentages of root colonization by AMF and root infection by F. virguliforme were tested for normal distribution. Data were subsequently subjected to the LSD Fisher’s honest significant difference (HSD) test in order to identify the significant differences (P ≤ 0.05) between treatments. Data analysis was performed with the statistical package INFOSTAT (Di Rienzo et al., 2011).
Root colonization by the AMF was estimated 96 h (Method 1) and 72 h (Method 2) after inoculation by F. virguliforme. This corresponded to 19 (Method 1) and 18 (Method 2) days post-plating of the soybean plantlets in the HC containing the ERM of the AMF. The non-mycorrhizal controls were similarly harvested and observed.
Intraradical hyphae, arbuscules and vesicles/spores were observed in the +AMF-Fv and +AMF+Fv treatments. In general, no significant differences were observed in %A or %V between the +AMF-Fv and +AMF+Fv treatments (Table 1) unless in one of the %RC. No root colonization was noticed in the non-mycorrhizal controls (-AMF+Fv and -AMF-Fv).
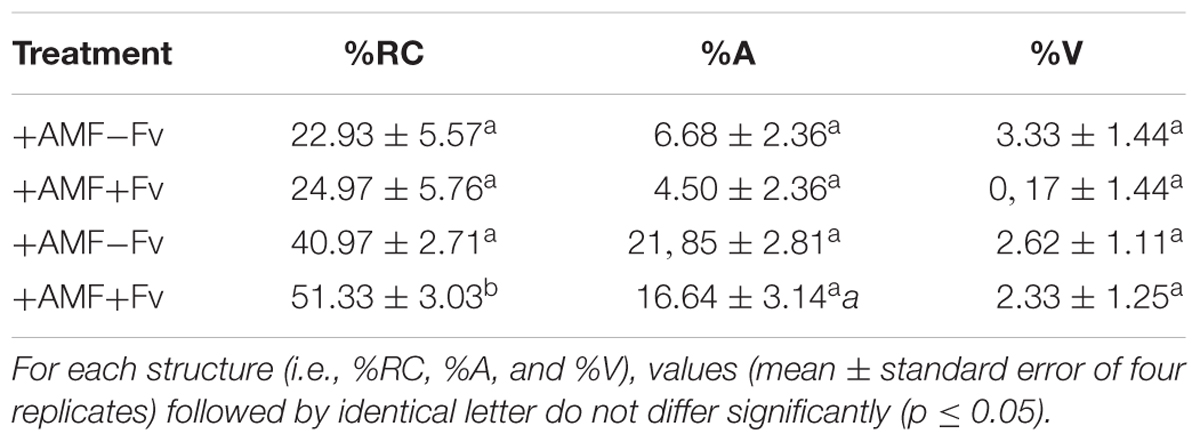
TABLE 1. Root colonization of soybean plants plated on actively growing extraradical mycelium networks of Rhizophagus irregularis in presence (+AMF+Fv) or absence (+AMF-Fv) of the pathogen Fusarium virguliforme.
In Method 1 (inoculation with a plug of PDA supporting mycelium), hyphae of F. virguliforme reached the surface of the roots 2 days after inoculation, independently of the presence/absence of AMF associated to the soybean plantlets. Following contact, the pathogen developed profusely from a few hyphae (Figure 2A) to a dense network of hyphae during the following 48 h (Figure 2B). Pathogen roots penetration and subsequent root tissues invasion was confirmed by microscopic observation. Careful examination of F. virguliforme within the roots revealed hyphae growing inter and intracellularly. Intercellular growth was predominantly parallel to the root axis (Figure 2C), whereas intracellular hyphae was perpendicular to it (Figure 2D). Many hyphae were also observed in the root tip zone (Figure 2E). Fungal structures with a swollen and pigmented cell wall were evidenced (Figures 2C,D).

FIGURE 2. Roots from soybean plantlets cultivated in vitro in the MDP in vitro culture system showing early infection stages by F. virguliforme observed under the stereo microscope (A,B) and under the microscope (C–E). (A) Fungal hyphae contacting the host root surface 2 days after inoculation of the pathogen (soybean plantlet of 17-days-old). Bar = 2 mm. (B) Hyphal development 4 days after inoculation of the pathogen (soybean plantlet of 19-days-old). Bar = 2 mm. (C–E) Soybean roots stained with Trypan blue 0.2%. (C) Intercellular growth of F. virguliforme hyphae. Bar = 5 μm. (D) Swollen and pigmented invasive structures (black arrow), intracellular growth of hyphae (arrowhead). Bar = 5 μm. (E) Hyphae in the tip roots zone. Bar = 20 μm.
In Method 2 (inoculation with macroconidia) a faster and denser development of hyphae was noticed on the surface of the MSR medium. Similarly to the inoculation via Method 1, hyphae and infective fungal structures were observed in the roots extending inter and intracellularly. The whole root system was infected by the pathogen irrespective of the treatment. Nevertheless, the percentage of root infection was significantly lower (p = 0.0129) in the +AMF+Fv treatment as compared to the -AMF+Fv treatment (Table 2).

TABLE 2. Percentage of root infection in presence (+AMF+Fv) or absence (-AMF+Fv) of the mycorrhizal fungi.
Necrotic areas were observed on the surface of the roots of the plantlets in both treatments, -AMF+Fv and +AMF+Fv, and with both inoculation method. In Method 1, small necrotic areas of about 1.3 mm diameter were noticed near the penetration point of the pathogen, in the soybean plantlets of the +AMF+Fv treatment (Figure 3A). To the contrary, in the plantlets of the -AMF+Fv treatment, F. virguliforme developed a necrotic area on either side of the infection point with a diameter of about 20 mm (˜95% higher) (Figure 3B). No symptoms were detected in roots of pre-mycorrhized soybean plantlets without F. virguliforme (+AMF-Fv) (Figure 3C) and non-mycorrhizal soybean plantlets without F. virguliforme (-AMF-Fv) (Figure 3D). State of cells viability was assessed by FDA-PI staining. (Supplementary Material).
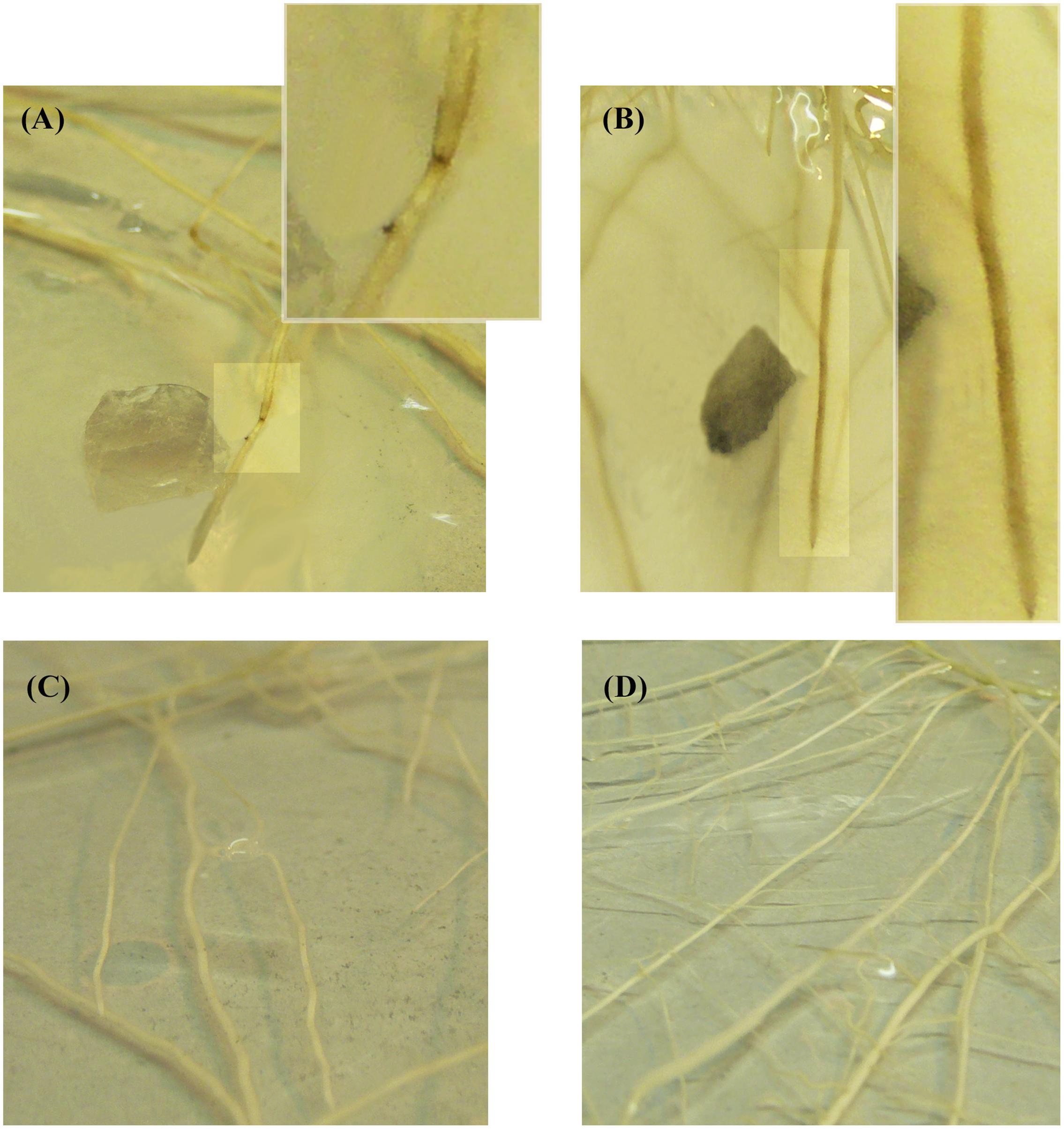
FIGURE 3. Roots from soybean plantlets cultivated in vitro in modified Strullu-Romand (MSR) medium observed by naked eyes. (A) pre-mycorrhized soybean plantlets inoculated with a plug of PDA (25 mm2) covered by F. virguliforme (+AMF+Fv). The white box shows an enlarged picture of root necrotic symptoms from A, 96 h following inoculation. (B) Non-mycorrhizal soybean plantlets inoculated with a plug of PDA (25 mm2) covered by F. virguliforme. The white box shows an enlarged picture of root necrotic symptoms from B, 96 h following inoculation. (C) pre-mycorrhized soybean plantlets without F. virguliforme (+AMF-Fv). (D) Non-mycorrhizal soybean plantlets without F. virguliforme (–AMF-Fv).
The evaluation of leaf symptoms was performed 96 and 72 h after inoculation by F. virguliforme. No symptoms were observed on the leaves of any soybean plantlets independently of the presence/absence of AMF and of the pathogen inoculation method.
In this study we reported for the first time on the tripartite interaction between soybean, a root symbiont (the AMF R. irregularis MUCL 41833) and a major root pathogen (F. virguliforme MUCL 53605) under in vitro culture conditions. Both microorganisms colonized the plantlet roots and the triple interaction was monitored by direct observation and under the microscope. The pathogen was inoculated following two methodologies: using a plug of gel covered by the pathogen (Method 1) and using a suspension of macroconidia (Method 2). In both cases, the pathogen was able to establish infection in the root tissues. This was evaluated under the microscope by determining the percentages of root infection by the pathogen. The same approach was followed to evaluate the level of root colonization by the AMF.
Root organ cultures colonized by AMF have been used to study mycorrhizal interaction for many years (Fortin et al., 2002; Elsen et al., 2003), while the interaction with whole plants is more recent (Voets et al., 2005; Dupré de Boulois et al., 2006; Koffi et al., 2009; Voets et al., 2009). For instance, Gallou et al. (2011) investigated the tripartite interaction between pre-mycorrhized potato plantlets and a leaf pathogen, P. infestans. Finally, Anene et al. (2013) investigated the interaction between pre-colonized banana plantlets and the nematode R. similis. Bressano et al. (2010) monitored the development of M. phaseolina in soybean roots under in vitro conditions. These authors demonstrated not only that the pathogen under study was able of infecting the host roots but also that the infecting structures observed in vitro and in vivo were similar. Therefore these systems paved the way for the tripartite interaction study between soybean, AMF and F. virguliforme.
In the present study, two inoculation methods were tested and compared. Both methods were effective, and 100% of the plantlets were infected within 3 days. However, the infection process differed slightly between the two methods. With Method 1, the number of infection sites was limited and almost restricted to the area where the plug supporting the pathogen was placed. Hyphae extending form the plug contacted the neighboring roots and developed localized symptoms. Conversely, with Method 2, infection was almost homogenous with multiple infection units distributed on the whole root system. Method 1 seems thus, particularly adapted for short-term time-course infection studies since the development of the pathogen can be easily and accurately monitored. This inoculation method is suitable for performing studies on dynamic of defense genes expression. Following the same rationale, studies on induced systemic resistance (ISR) or mycorrhizal induced resistance (MIR), generally conducted in tri-partite systems separating roots infected by a pathogen from non-infected roots (Khaosaad et al., 2007), can also be performed with this method. Besides, our methodology is feasible to be applied with a high-throughput system analysis like RNA sequencing (RNA-seq), DNA microarrays or even a chemical library.
Method 2, on the other hand, is more adapted when a high level of infection in whole root system is requested representing an infection shock for the plant.
Root symptoms were observed in the mycorrhizal as well as non-mycorrhizal plantlets. However, in the pre-mycorrhized plantlets infected by Method 1, the size of the necrotic lesion on the roots was approximately 95% smaller as compared to the non-mycorrhizal plantlets. Similarly, the level of necrosis on the whole root system was less intense in the pre-mycorrhized plantlets infected by Method 2. No symptoms were evidenced on the leaves. This was probably related to the short-term infection study (96 and 72 h for inoculation Method 1 and 2, respectively).
Interestingly, when the presence of the pathogen inside the root tissue was quantified, the amount of intraradical hyphae of the pathogen was significantly lower in the pre-mycorrhized plantlets as compared to the non-mycorrhizal ones. These observations support the hypothesis that early AMF root colonization, i.e., prior to any interaction with the pathogen, increases the ability of the colonized plantlet to withstand the competition with the pathogen. This hypothesis was supported by numerous studies reporting reduction of severity in diseases caused by soil-borne and above-ground pathogens in pre-mycorrhized plants (Whipps, 2004; Pozo et al., 2009).
These results, demonstrated the suitability of the in vitro cultivation system to study the tripartite interaction between soybean, an AMF (R. irregularis) and a major root pathogen (F. virguliforme). It offers large perspectives to investigate the mechanisms involved in early and transient protection conferred to plants in presence of the symbiotic fungus. In particular, molecular studies [e.g., defense gene expression analysis (Gallou et al., 2011)] and microarray approaches (Gallou et al., 2012) would benefit from such system.
The main innovations of the system here presented compared to other similar systems developed previously by other authors are referred to the type of pathogen, a double colonization of the same plant organ by two microorganism and the verification of this fact and to corroborate symptoms attenuation in an in vitro system. In other words, we worked with a soil-borne pathogen that infects roots. This implies not only the use of a different pathogen not previously reported, but also that the pathogen develops in a different environment such as the soil, which is not a minor fact and has not been reported previously. On the other hand, infection by the pathogen and colonization by the mycorrhizal fungus in the roots was verified, this is to say, a complex interaction system was established in the same organ of the plant, the roots. And finally, it is the first time that symptoms attenuation is reported in an in vitro mycorrhizal system and although it is true that this symptoms mitigation was estimated qualitatively, it opens the possibilities to use this system to perform more exhaustive studies.
However, the system also have some limitations which most important is the limited space for the protagonists to develop. Indeed the in vitro cultivation system is not adapted for long-term interaction studies due to the fast growth of the soybean plantlet as well as the pathogen on the synthetic growth medium. The use of the autotrophic in vitro cultivation system represents a substantial improvement with regard to root organ cultures often used in interaction studies of AMF with pathogens, mainly because of the presence of photosynthetic active tissues and the absence of sugar and vitamins in the growth medium. This model system offers a highly controlled option to investigate the molecular (e.g., defense gene expression) as well as other mechanisms involved in the bio-protection conferred by AMF to a major soil-borne pathogen of soybean.
MG and NM evenly contributed to the manuscript in experiment developing and writing. AG contributed with experiment developing and critical review, CL contributed with critical views, SD and DD contributed in experimental planning and writing.
This work was supported by INTA research funds (PNPV-1135023) and Twinning Program (MINCyT N° 740/09).
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
MG acknowledges the support of the INTA’s program for Ph.D. in the context of Biological Sciences Doctoral Career of the National University of Córdoba-Argentine. The authors would like to thank María Mercedes Scandiani for providing the strain of Fusarium virguliforme and Dr. Leandro Ortega for assistance in microscopy.
The Supplementary Material for this article can be found online at: http://journal.frontiersin.org/article/10.3389/fpls.2017.01033/full#supplementary-material
Anene, A., Koffi, M. C., Vos, C., and Declerck, S. (2013). Rhizophagus irregularis MUCL 41833 decreases the reproduction ratio of Radopholus similis in the banana cultivar Yangambi km5. Nematology 15, 629–632. doi: 10.1163/15685411-00002742
Aoki, T., ODonnell, K., and Scandiani, M. (2005). Sudden death syndrome of soybean in South America is caused by four species of Fusarium: Fusarium brasiliense sp. nov., F. cuneirostrum sp. nov., F. tucumaniae, and F. virguliforme. Mycoscience 46, 162–183. doi: 10.1007/s10267-005-0235-y
Bressano, M., Lorena Giachero, M., Luna, C. M., and Ducasse, D. A. (2010). An in vitro method for examining infection of soybean roots by Macrophomina phaseolina. Physiol. Mol. Plant Pathol. 74, 201–204. doi: 10.1016/j.pmpp.2009.12.003
Brink, S. C. (2016). Unlocking the secrets of the rhizosphere. Trends Plant Sci. 21, 169–170. doi: 10.1016/j.tplants.2016.01.020
Cranenbrouck, S., Voets, L., Bivort, C., Renard, L., Stullu, D., and Declerck, S. (2005). “Methodologies for in vitro cultivation of arbuscular mycorrhizal fungi with root organs,” in In Vitro Culture of Mycorrhizas, Vol. 527, ed. S. Declerck, D. G. Strullu, J. A. Fortin (Heidelberg: Springer-Verlag), 341–348. doi: 10.1007/3-540-27331-x_18
De Jaeger, N., Ivan, E., Dupré de Boulois, H., and Declerck, S. (2011). Trichoderma harzianum might impact phosphorus transport by arbuscular mycorrhizal fungi. FEMS Microbiol. Ecol. 77, 558–567. doi: 10.1111/j.1574-6941.2011.01135.x
Declerck, S., Strullu, D., and Plenchette, C. (1998). Monoxenic culture of the intraradical forms of Glomus sp. isolated from a tropical ecosystem: a proposed methodology for germplasm collection. Mycologia 90, 579–585. doi: 10.2307/3761216
Declerck, S., Strullu, D. G., and Fortin, A. (eds). (2005). In Vitro Culture of Mycorrhizas. Berlin: Springer Science & Business Media. doi: 10.1007/b138925
Di Rienzo, J. A., Casanoves, F., Balzarini, M. G., Gonzalez, L., Tablada, M., and Robledo, C. W. (2011). InfoStat versión 2011. Argentina: Grupo InfoStat, FCA, Universidad Nacional de Córdoba.
Dupré de Boulois, H., Voets, L., Delvaux, B., Jakobsen, I., and Declerck, S. (2006). Transport of radiocaesium by arbuscular mycorrhizal fungi to Medicago truncatula under in vitro conditions. Environ. Microbiol. 8, 1926–1934. doi: 10.1111/j.1462-2920.2006.01070.x
Elsen, A., Declerck, S., and De Waele, D. (2003). Use of root organ cultures to investigate the interaction between Glomus intraradices and Pratylenchus coffeae. Appl. Environ. Microbiol. 69, 4308–4311. doi: 10.1128/AEM.69.7.4308-4311.2003
Fortin, J. A., Bécard, G., Declerck, S., Dalpé, Y., St-Arnaud, M., Coughlan, A. P., et al. (2002). Arbuscular mycorrhiza on root-organ cultures. Can. J. Bot. 80, 1–20. doi: 10.1139/b01-139
Gallou, A. (2011). Impact of Rhizophagus sp. (syn. Glomus sp.) and Trichoderma harzianum on the Potato Resistance against Rhizoctonia solani and Phytophthora infestans, Two Major Potato Pathogens. Ph.D.thesis, University Catholic of Louvain, Louvain.
Gallou, A., Declerck, S., and Cranenbrouck, S. (2012). Transcriptional regulation of defence genes and involvement of the WRKY transcription factor in arbuscular mycorrhizal potato root colonization. Funct. Integr. Genomics 12, 183–198. doi: 10.1007/s10142-011-0241-4
Gallou, A., Lucero Mosquera, H. P., Cranenbrouck, S., Suárez, J. P., and Declerck, S. (2011). Mycorrhiza induced resistance in potato plantlets challenged by Phytophthora infestans. Physiol. Mol. Plant Pathol. 76, 20–26. doi: 10.1016/j.pmpp.2011.06.005
Genre, A., Chabaud, M., Faccio, A., Barker, D. G., and Bonfante, P. (2008). Prepenetration apparatus assembly precedes and predicts the colonization patterns of arbuscular mycorrhizal fungi within the root cortex of both Medicago truncatula and Daucus carota. Plant Cell 20, 1407–1420 doi: 10.1105/tpc.108.059014
Genre, A., Chabaud, M., Timmers, T., Bonfante, P., and Barker, D. G. (2005). Arbuscular mycorrhizal fungi elicit a novel intracellular apparatus in Medicago truncatula root epidermal cells before infection. Plant Cell 17, 3489–3499. doi: 10.1105/tpc.105.035410
Gutjahr, C., and Parniske, M. (2013). Cell and developmental biology of arbuscular mycorrhiza symbiosis. Annu. Rev. Cell Dev. Biol. 29, 593–617. doi: 10.1146/annurev-cellbio-101512-122413
Khaosaad, T., Garcia-Garrido, J., Steinkellner, S., and Vierheilig, H. (2007). Take-all disease is systemically reduced in roots of mycorrhizal barley plants. Soil Biol. Biochemi. 39, 727–734. doi: 10.1016/j.soilbio.2006.09.014
Koffi, M. C., de la Providencia, I. E., Elsen, A., and Declerck, S. (2009). Development of an in vitro culture system adapted to banana mycorrhization. Afr. J. Biotechnol. 8, 2750–2756 doi: 10.1071/RD12044
Koffi, M. C., Vos, C., Draye, X., and Declerck, S. (2013). Effects of Rhizophagus irregularis MUCL 41833 on the reproduction of Radopholus similis in banana plantlets grown under in vitro culture conditions. Mycorrhiza 23, 279–288. doi: 10.1007/s00572-012-0467-6
McGonigle, T., Miller, M., Evans, D., Fairchild, G., and Swan, J. (1990). A new method which gives an objective measure of colonization of roots by vesicular-arbuscular mycorrhizal fungi. New Phytol. 115, 495–501. doi: 10.1111/j.1469-8137.1990.tb00476.x
Oburger, E., and Schmidt, H. (2016). New methods to unravel rhizosphere processes. Trends Plant Sci. 21, 243–255. doi: 10.1016/j.tplants.2015.12.005
O’Donnell, K., Sink, S., Scandiani, M. M., Luque, A., Colletto, A., Biasoli, M., et al. (2010). Soybean sudden death syndrome species diversity within North and South America revealed by multilocus genotyping. Phytopathology 100, 58–71. doi: 10.1094/PHYTO-100-1-0058
Oerke, E. C. (2006). Crop losses to pests. J. Agric. Sci. 144, 31–43. doi: 10.1017/s0021859605005708
Pozo, M. J., Verhage, A., García-Andrade, J., García, J. M., and Azcón-Aguilar, C. (2009). “Priming plant defence against pathogens by arbuscular mycorrhizal fungi,” in Mycorrhizas-Functional Processes and Ecological Impact, eds C. Azcón-Aguilar, J. M. Barea, S. Gianinazzi, and V. Gianinazzi-Pearson (Berlin: Springer), 123–135. doi: 10.1007/978-3-540-87978-7_9
Rupe, J. (1989). Frequency and pathogenicity of Fusarium solani recovered from soybean with sudden death syndrome. Plant Dis. 73, 581–584. doi: 10.1094/PD-73-0581
Rupe, J. C., Gbur, E. E., and Marx, D. (1989). Spatial and temporal distribution of sudden death syndrome of soybeans. Phytopatology 79, 1219.
Voets, L., Dupré de Boulois, H., Renard, L., Strullu, D.-G., and Declerck, S. (2005). Development of an autotrophic culture system for the in vitro mycorrhization of potato plantlets. FEMS Microbiol. Lett. 248, 111–118. doi: 10.1016/j.femsle.2005.05.025
Voets, L., de la Providencia, I. E., Fernandez, K., IJdo, M., Cranenbrouck, S., and Declerck, S. (2009). Extraradical mycelium network of arbuscular mycorrhizal fungi allows fast colonization of seedlings under in vitro conditions. Mycorrhiza 19, 347–356. doi: 10.1007/s00572-009-0233-6
Whipps, J. (2004). Prospects and limitations for mycorrhizas in biocontrol of root pathogens. Can. J. Bot. 82, 1198–1227. doi: 10.1139/b04-082
Keywords: arbuscular mycorrhizal fungi, Fusarium virguliforme, in vitro system, soybean, plant–microbe interaction
Citation: Giachero ML, Marquez N, Gallou A, Luna CM, Declerck S and Ducasse DA (2017) An In Vitro Method for Studying the Three-Way Interaction between Soybean, Rhizophagus irregularis and the Soil-Borne Pathogen Fusarium virguliforme. Front. Plant Sci. 8:1033. doi: 10.3389/fpls.2017.01033
Received: 22 December 2016; Accepted: 29 May 2017;
Published: 16 June 2017.
Edited by:
Roger Deal, Emory University, United StatesReviewed by:
Jozef Samaj, Palacký University, Olomouc, CzechiaCopyright © 2017 Giachero, Marquez, Gallou, Luna, Declerck and Ducasse. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Daniel A. Ducasse, ZHVjYXNzZS5kYW5pZWxAaW50YS5nb2IuYXI=
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.