
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Plant Sci., 29 May 2017
Sec. Plant Physiology
Volume 8 - 2017 | https://doi.org/10.3389/fpls.2017.00889
 Elizabeth Estrada-Johnson1†
Elizabeth Estrada-Johnson1† Fabiana Csukasi1†
Fabiana Csukasi1† Carmen M. Pizarro1
Carmen M. Pizarro1 José G. Vallarino1
José G. Vallarino1 Yulia Kiryakova2
Yulia Kiryakova2 Amalia Vioque1
Amalia Vioque1 Javier Brumos3
Javier Brumos3 Nieves Medina-Escobar1
Nieves Medina-Escobar1 Miguel A. Botella1José M. Alonso3
Miguel A. Botella1José M. Alonso3 Alisdair R. Fernie4
Alisdair R. Fernie4 José F. Sánchez-Sevilla5
José F. Sánchez-Sevilla5 Sonia Osorio1*
Sonia Osorio1* Victoriano Valpuesta1*
Victoriano Valpuesta1*The role of auxin in ripening strawberry (Fragaria ×ananassa) fruits has been restricted to the early stages of development where the growth of the receptacle is dependent on the delivery of auxin from the achenes. At later stages, during enlargement of the receptacle, other hormones have been demonstrated to participate to different degrees, from the general involvement of gibberellins and abscisic acid to the more specific of ethylene. Here we report the involvement of auxin at the late stages of receptacle ripening. The auxin content of the receptacle remains constant during ripening. Analysis of the transcriptome of ripening strawberry fruit revealed the changing expression pattern of the genes of auxin synthesis, perception, signaling and transport along with achene and receptacle development from the green to red stage. Specific members of the corresponding gene families show active transcription in the ripe receptacle. For the synthesis of auxin, two genes encoding tryptophan aminotransferases, FaTAA1 and FaTAR2, were expressed in the red receptacle, with FaTAR2 expression peaking at this stage. Transient silencing of this gene in ripening receptacle was accompanied by a diminished responsiveness to auxin. The auxin activity in the ripening receptacle is supported by the DR5-directed expression of a GUS reporter gene in the ripening receptacle of DR5-GUS transgenic strawberry plants. Clustering by co-expression of members of the FaAux/IAA and FaARF families identified five members whose transcriptional activity was increased with the onset of receptacle ripening. Among these, FaAux/IAA11 and FaARF6a appeared, by their expression level and fold-change, as the most likely candidates for their involvement in the auxin activity in the ripening receptacle. The association of the corresponding ARF6 gene in Arabidopsis to cell elongation constitutes a suggestive hypothesis for FaARF6a involvement in the same cellular process in the growing and ripening receptacle.
Fruit ripening is a complex and coordinated irreversible developmental process that leads to the production of a soft and edible ripe fruit. The hormone auxin is known to play a critical role in fruit growth, from flower formation to fruit ripening (Pattison et al., 2014). At early stage is believed to participate in the cell expansion associated to fruit growth, while at maturation a role has also been proposed (Trainotti et al., 2007; McAtee et al., 2013), although not definitive information exits on the cellular and molecular processes involved. Strawberry has been considered as an model to study the role played by the hormone auxin, at the molecular level, in fruit growth and ripening. In strawberry fruit, it was reported that growth of the receptacle ceased if achenes, the source of auxin, were removed after pollination, but growth of the receptacle was taken up again after the external application of 2-naphtoxyacetic acid (Nitsch, 1950). Later in development, the removal of the achenes accelerated some processes associated with fruit ripening (Given et al., 1988). Some studies have identified a number of genes that are down-regulated (Harpster et al., 1998; Aharoni et al., 2002) or up-regulated (Castillejo et al., 2004) after the external application of auxin to developing fruits. However, detailed studies on the content, synthesis, and signaling of this hormone in different fruit parts at different developmental stages are lacking.
Auxin sensing and signaling activity is relatively well established in plants (Weijers and Wagner, 2016). In this vein, some years ago, an F-box protein, TIR1, was identified as an auxin receptor (Dharmasiri et al., 2005; Kepinski and Leyser, 2005). The Arabidopsis genome encodes five other F-box proteins, AFB1-AFB5, that also function as auxin receptors (Dharmasiri et al., 2005; Parry et al., 2009). The auxin receptor TIR1 is part of the SCFTIR1 ubiquitin ligase complex and binds auxin directly (Dharmasiri et al., 2005; Kepinski and Leyser, 2005). Efficient auxin binding requires the assembly of an auxin co-receptor complex consisting of TIR1 and an Aux/indole-3-acetic acid (IAA) protein (Calderón Villalobos et al., 2012) that then induces poly-ubiquitination of the Aux/IAA protein and its targeting and degradation in the proteasome (Benjamins and Scheres, 2008). Most species produce a high number of Aux/IAA proteins, with some variability in the protein domains as well as in their expression pattern, that might support a variable range of auxin sensitivity within the plant (Dreher et al., 2006; Salehin et al., 2015; Xie et al., 2015; Weijers and Wagner, 2016). An alternative proteasome-independent auxin signaling pathway, located in the endoplasmic reticulum and the cell wall, has been proposed (Sauer and Kleine-Vehn, 2011). Its contribution to the auxin signaling has been reported in some processes such as cell wall loosening and cell expansion (Ljung, 2013).
Transcriptional regulators known as auxin response factors (ARF) are key elements in the transcriptional response of plants to auxin (Weijers and Wagner, 2016). The ARF proteins bind to cis-regulatory sequences (AuxREs) in the promoter of a set of auxin-dependent genes, controlling their expression and mediating the auxin-dependent growth and developmental processes (Guilfoyle and Hagen, 2007). Most species have a large family of ARF proteins, divided into subfamilies, that are responsible for the diverse roles played by auxin in different cellular processes (Finet et al., 2012). They typically contain a DNA-binding domain (B3) at the N-terminus flanked by a dimerization domain, a medium domain that mediates transcriptional regulation, and a C-terminus domain (PB1) for oligomerization and Aux/IAA heterodimerization (Guilfoyle, 2015; Weijers and Wagner, 2016). Variants of this domain architecture have been found between and within species (Finet et al., 2012), which contributes to the versatility of the transcriptional response to auxin in many different species and circumstances.
Two major pathways for IAA biosynthesis, the tryptophan-dependent and -independent pathways, have been reported for auxin biosynthesis in plants. However, it is accepted that a significant amount of auxin synthesis in plants is predominantly synthesized by the tryptophan-dependent two-step pathway that has as an intermediate indol-3-pyruvic acid (IPyA) (Brumos et al., 2014). The initial conversion of Trp to IPya is catalyzed by a tryptophane aminotransferase enzyme that includes multiple members in several species (TAA1 and TARs in Arabidopsis) (Stepanova et al., 2008). IPyA is then further converted to IAA by the YUCCA proteins, a family of flavin-dependent monooxygenases (Zhao et al., 2001; Stepanova et al., 2011). The TAA1, TAR, and YUCCA genes show specific expression patterns in several species, including the wild strawberry, associated with the involvement of auxin in different developmental processes (Kang et al., 2013; Brumos et al., 2014; Liu et al., 2014). Starting also from Trp a pathway with an intermediate indole-3-acetamide (IAM) has also been proposed to participate in the auxin biosynthesis plants, since both the IAM and the corresponding enzyme activities isolated in Arabidopsis and tobacco (Mano et al., 2010). In general, several mechanisms have been proposed to control auxin homeostasis, including IAA metabolism (biosynthesis, degradation, and conjugation), transport, and compartmentation (Ljung, 2013). Recently, an enzyme controlling IAA oxidation in Arabidopsis has been reported (Porco et al., 2016). In addition, a proportion of cellular auxin is conjugated to other molecules, being proposed that they might play a role of auxin storage molecules, or intermediates compounds in the auxin catabolic pathway (Ljung, 2013). Interestingly, some of these auxin-conjugates have been identified in fruits such as grape and tomato, with values of content changing along ripening (Böttcher et al., 2010).
The complexity of the auxin biosynthesis, sensing and signaling machinery, present in all plant species, makes it obligatory to have global information about their components, their variability, and their occurrence in relation to the process to be studied. RNA-seq analysis allows having this global information, at the transcript level, in a single experiment. A basic requirement for using this approach is the availability of the genomic sequence of the species. However, it has also been reported that mapping the reads in a closely related model species provides valid results (Conesa et al., 2016). Thus, in the case of Fragaria ananassa, with no sequenced genome, the sequenced F. vesca genome can be used to map the reads and obtain the transcription atlas of a process such as fruit ripening. The F. vesca map has been successfully used in the less-related species Rubus sp. (García-Seco et al., 2015). Transcriptomic studies in F. ananassa are complicated by the fact that it is an octoploid species (Tennessen et al., 2014). This ploidy might significantly increase the number of allelic variants that exist for the different components of the auxin sensing and signaling machinery. A partial solution to this challenge can be provided by the RNA-seq data as some methods have been reported that facilitate transcriptome assembly from RNA-seq data in species without a sequenced genome (Chevreux et al., 2004; Grabherr et al., 2011).
Our study is focused on expanding current knowledge about the role of auxin in strawberry fruit development and ripening. Our RNA-seq analysis of the ripening achene and receptacle identified the red stage-specific expression of selected members of the auxin synthesis, sensing and signaling pathways. This result was accompanied by the measurement of the auxin content in the receptacle at three developmental stages, and a functional test of the machinery by transient silencing of a main auxin synthesis gene. When taken together, our results support the involvement of auxin in the specific cellular processes taking place in the ripening receptacle.
Strawberry plants used for transient transformation were grown and maintained under glasshouse conditions (IFAPA-CIFA Churriana, Málaga, Spain). The transient silencing of FaTAR2 by agroinfiltration with the RNAi construct pBI-FaTAR2i, or pBI-intron as a control, was carried out in octoploid strawberry (Fragaria ×ananassa Duch.) cv Camarosa fruits, as described previously (Hoffmann et al., 2006), from March to May.
For the construction of pBI-FaTAR2i, a sequence of FaTAR2 (KY509034) was PCR-amplified from strawberry (Fragaria ×ananassa) cv Camarosa cDNA using gene- specific primers (forward, 5′ CTTGACCAACACCACTGAAA-3′; reverse, 5′ GTGTCTTCCTCCTCGGGTCA-3′). The forward and reverse primers contained an Nhel/SpeI and an XbaI/SacI restriction site, respectively. The fragment was ligated into the binary vector pSK that contained XbaI/NheI and SpeI/SacI restriction sites separated by an intron from strawberry (Hoffmann et al., 2006); the vector was thus cut with XbaI and NheI, and the fragment was ligated in the sense direction. Second, the vector was digested with SpeI and SacI, and the fragment was inserted in the antisense direction, obtaining the intron–hairpin construct pSK-TAR2i. This plasmid was cut with AscI, and the digested fragment was cloned into the AscI-cut pBI121, obtaining the construct pBI–FaTAR2i. The fruits remained attached to the plants after agroinfiltration. Seven to 10 days after injection, the fruits were harvested.
The strawberry cv Camarosa fruits used for RNAseq were harvested in four different developmental stages corresponding to green (G), white (W), turning (T), and red (R). These fruits were collected from plants that were grown under field conditions in Huelva, Spain. All fruits were frozen immediately in liquid nitrogen, and achenes were removed using a scalpel on frozen fruits. Transcriptome analysis was performed in three separate pools of 20 fruits each. Each pool was from four different plants. Libraries were sequenced on Illumina HiSeq2000 lanes using 2 × 100 bp reads. More than 30 million reads were generated for each sample. Analyses of transcript data were performed using FastQC, TopHat, Cufflinks, Blast2go, and IGV software as previously described (Trapnell et al., 2012; Sánchez-Sevilla et al., 2014). Normalized RNAseq fragment counts were used to measure the relative abundances of transcripts, expressed as fragments per kilobase of exon per million fragments mapped (FPKM).
Achenes of green strawberry fruits on the plant were carefully removed, using the tip of a scalpel blade. Fruits were harvested at 0, 24, 48, 72, and 96 h after treatment, immediately frozen in liquid nitrogen and stored at -80°C. The injection with NAA was performed in fruits at the turning stage with ca. 250 μl of a basic water solution (NaOH 50 μl in 500 ml of water) 2 mM in naphthalene acetic acid (NAA) and 2% DMSO. Mock solution contained the basic water with DMSO. Fruits were harvested after 24, 72, and 96 h, immediately frozen and stored at -80°C.
Indole-3-acetic acid (IAA) was identified by co-elution with an [2H5]-IAA standard (OlChemim Ltd, Olomouc, Czech Republic). Recovery experiments in which the amount of authentic IAA added at the start of the experiment was doubled yielded a result of 89.5 ± 2.1%, indicating a high stability of the metabolite and its derivative throughout the extraction, derivatization, and analytical processes.
Indole-3-acetic acid was extracted overnight from 6 g of receptacle fruits without achenes in 20 ml of 80% methanol. After extraction, each sample was reduced in vacuo and diluted with 20 ml of water. The aqueous phase was adjusted to pH 2.8 with 1 M HCl and partitioned four times with equal volumes of ethyl acetate. The ethyl acetate extracts were combined and evaporated to dryness. The residue was dissolved in 1 ml of 10% methanol and applied to a pre-equilibrated C18 cartridge1. The column was washed with aqueous acetic acid (pH 3.0), and then IAA was eluted with 80% methanol. After evaporation to dryness, the samples were derivatized and analyzed using internal [2H5]-IAA standard by GC-MS as in Osorio et al. (2011).
RNA extraction and transcriptome analysis by RNAseq were performed as previously described by Vallarino et al. (2015). A total of 10 independent samples corresponding to achene and receptacle, at four developmental stages (green, white, turning, and red), leaf and root, with three replications per sample, were analyzed. The total number of reads was over 990 million, and the average of reads per sample ranged from 26,4 to 40,9 millions (Supplementary Table S1). For gene expression analysis by qRT-PCR, first-strand cDNA synthesis of 1 mg of RNA in a final volume of 20 mL was performed using the iScript cDNA synthesis kit (Bio-Rad). Expression of the FaTAA1, FaTAR2, GUS, FaAux/IAA11, and FaTIR1 genes was analyzed by real-time qRT-PCR using the fluorescent intercalating dye SsoFast EvaGreen supermix in the MyiQ detection system (Bio-Rad). Relative quantification of the target expression level was performed using the comparative Ct method. The following primers were used: for analysis of FaTAA1 (forward, 5′-GGCCAGTGGATGAGCTATGT-3′; reverse, 5′-CCACCAGGAGAAGTGAGAGC-3′); FaTAR2 (forward, 5′-TGAGGAACTTGCTTGTGCTG-3′; reverse, 5′-TGGACCT CTCTGCTTCTGGT-3′); FaAUX/IAA11 (forward, 5′-TGGT GGTCAGGAGCATGATA-3′; reverse, 5′-TTAGCCTCTTCA CGGAACTAAGA-3′); FaTIR1 (forward, 5′-AGCCACTTG ATGAGCCACTTGATGTGGGTTTC-3′; reverse, 5′-AAAGC GCCTTATCACCAAAA-3′); FaARF6a (forward, 5′-AGTTT GTAAATAGTGTGTGGTGCAT-3′; reverse, 5′-CTGCATTGG GACAGACTTCAG-3′); and GUS (forward, 5′-GATCGCGA AAACTGTGGAAT-3′; reverse, 5′-AAAGACTTCGCGCT GATACC-3′). To normalize the gene expression levels for differences in the efficiency of cDNA synthesis, transcript levels of the constitutively expressed gene FaGAPH (Salvatierra et al., 2010) and/or FaCHP1 (Clancy et al., 2013) were measured.
For GUS analysis, tissues were incubated O/N at 37°C with GUS buffer (50 mM sodium phosphate buffer, pH 7.0, 10 mM Na2EDTA, 0.5 mM K4 [Fe(CN)6]⋅3H2O, 0.5 mM K3[Fe(CN)6], 0.1% Triton X-100, and 1 mg/mL X-Gluc) as previously described (Jefferson et al., 1987) and were then de-stained by incubating in ethanol/acetic acid 3:1 at RT for 24 h.
The FaTAA1 (KY509033) and FaTAR2 (KY509034) open reading frames were subcloned into pENTR/D-Topo and transferred into pDEST15 by Gateway LR recombination (Invitrogen) as previously described (Stepanova et al., 2008). GST-FaTAA1 was expressed in the BL21 Star (DE3) pLysS strain of Escherichia coli (Invitrogen) and induced by 0.5 mM isopropyl b-D-1-thiogalactopyranoside for 4 h. Equal volumes of protein extracts were loaded onto a native 10% polyacrylamide gel and run for 2 h at 100 V at 48°C. The in-gel aminotransferase activity was assayed as described (Pedraza et al., 2004) at 24°C over-night. Biochemical characterization of FaTAA1 and FaTAR2 was performed using recombinant GST-FaTAA1 and GST-TAR2 batch purified on glutathione-sepharose beads (Amersham Pharmacia) according to the manufacturer’s recommendations. Purified protein concentrations were estimated by SDS-PAGE followed by Coomassie Brilliant Blue staining. For a 100-mL reaction, 5 mg of FaTAA1 and FaTAR2 was used. Aminotransferase activity was assayed as previously described (Pedraza et al., 2004).
For HPLC-MS chromatography, reactions were carried out with the purified enzyme at 37°C for 1 h using previously described conditions (Koshiba and Matsuyama, 1993). To identify the products of the enzymatic reaction, indole-3-pyruvic acid and tryptophan (Sigma) were employed as standards. HPLC analysis was performed using a Shimadzu LC-MS 2010 EV module system with a UV photodiode array (190–800 nm) detection. For LC-MS analysis, the same system in combination with a single-stage quadrupole mass analyser coupled with electrospray ionization was utilized. The ion chromatograms were obtained using MS detection with negative ionization. Scans of peaks were stored from m/z of 50 to 2000 amu. The speed of the scan was 2000 amu/second. We used 0.1% acetic acid (HPLC grade) (solvent A) and acetonitrile (GC-MS grade) (solvent B) (VWR) as solvents for HPLC and LC-MS. The gradient program consisted of ratios of solvent A to solvent B as follows: 10:90 (0–5 min), 15:85 (5–10 min), 17:83 (10–20 min), 25:75 (20–30 min), 35:65 (30–40 min), 45:55 (40–50 min), 52:48 (50–55 min), 55:45 (55–58 min), 60:40 (58–58.5 min), 90:10 (58.5–59 min), and 10:90 (59–60 min). Data from LC chromatograms, UV spectrum, ion chromatograms, and MS spectrum were acquired from 0 to 60 min. After each run, the column was washed for 10 min using the solvents with the ratio of solvent A to B of 10:90. A ZORBRX Eclipse XDB-C18 reverse column (4.6 mm × 250 mm, 5-micron) (Agilent) was used to separate compounds from enzymatic reactions. The flow rate was 0.4 ml/min. The injection volume of samples was 30 μl for HLPC assay and 10 μl for LC-MS assay. We used 2 μl standards of freshly prepared IPA and Trp (0.1 μg/μl) to identify the product of the Trp aminotransferase reaction. Reaction products were identified by retention time, UV spectrum, ion chromatograms, and MS spectrum. Each experiment was performed in triplicate.
The unrooted phylogenetic tree shown in Supplementary Figures S1, S2 online was constructed using MEGA 5.052 with the neighbor-joining statistical method and bootstrap analysis (1000 replicates). Gene sequences were downloaded from Phytozome3 and GDR4; sequence alignment was performed using Clustal Omega5.
Previous studies of genes expressed in strawberry fruits identified some components of the auxin signaling pathway that were expressed not only in the green fruits but also in the red fruits (Bombarely et al., 2010). As this pattern could be an indication of auxin presence in the ripe receptacle, the content of this hormone was evaluated at three developmental stages. Thus, we measured the content of free IAA in the receptacle at green, white and red stages using GC-MS. The highest IAA content (5.14 ng/gFW) was observed in the green receptacle, and it was then diminished in the white and red receptacles (3.43 and 3.18 ng/gFW, respectively) (Figure 1A). This pattern of diminution in the transition from green to white and red stages was altered when the values were expressed on a dry weight basis (Figure 1B). In the transition from green to a red receptacle, the total number of cells remains constant with an increase in cell size, mostly by water uptake (Cheng and Breen, 1992). The maintenance of an equivalent amount of free auxin content per dry weight in the transition from green to red receptacle must be the result of either an active transport from the achene or the synthesis of auxin in the receptacle, concurrent with the development of this organ.
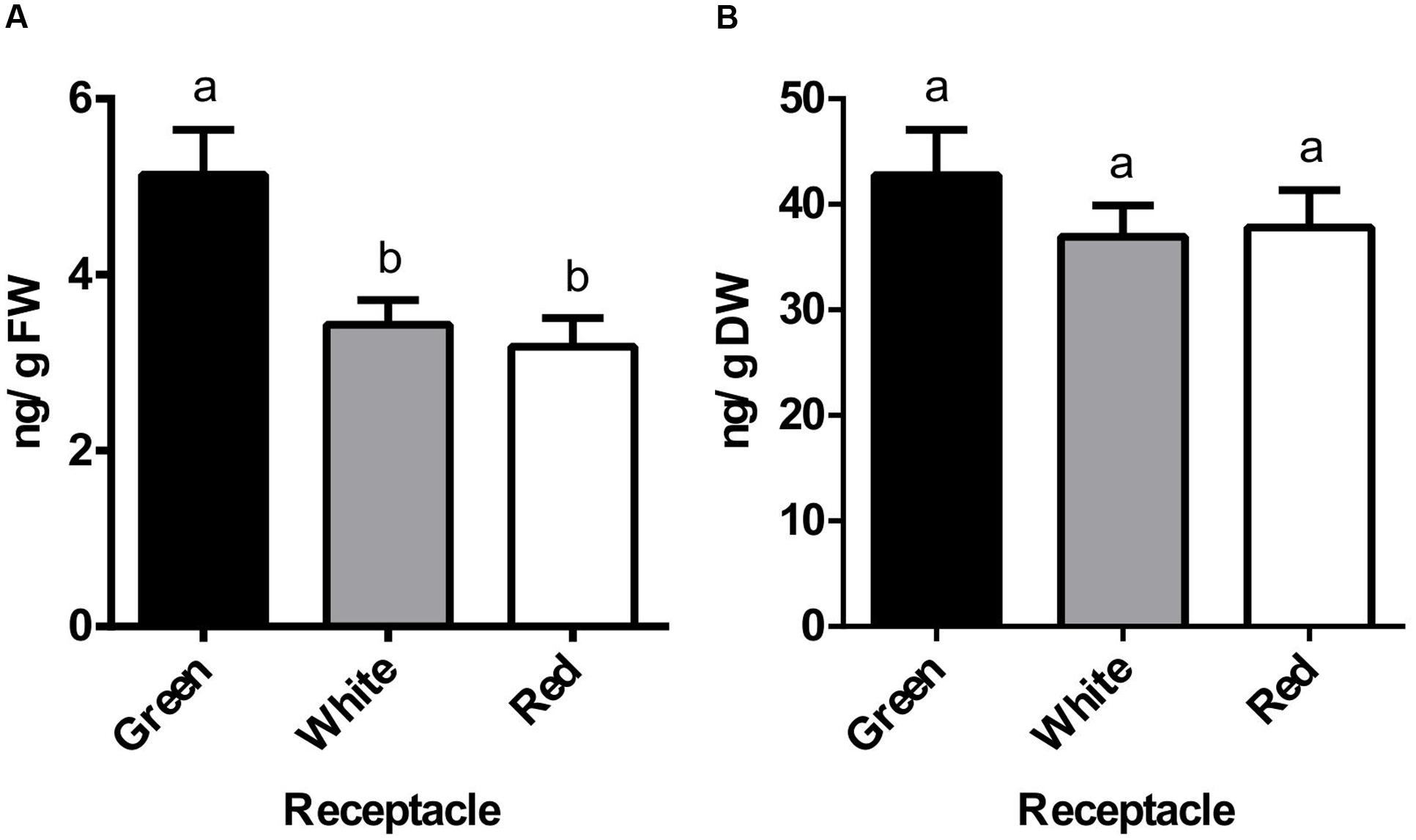
FIGURE 1. Changes in auxin endogenous levels during receptacle development and ripening. Endogenous levels of indole-3-acetic acid (IAA) in green, white and red receptacle as measured by GC-MS and expressed per gram of fresh weight (FW, A) or dry weight (DW, B). Bars represent the mean of four independent biological samples ± SE. Different letters indicate a significant difference between samples according to the corresponding ANOVA (P < 0.05).
In an effort to elucidate the role played by auxin in the ripening of strawberry fruit we performed RNA-seq in achenes and receptacle at different developmental stages (green, white, turning, and red) to identify differential expression of auxin related genes (Supplementary Table S2). Although the strawberry (Fragaria × ananassa) genome has not been sequenced, the genome of its wild relative F. vesca was sequenced (Shulaev et al., 2011), and more recently updated (Tennessen et al., 2014). Mapping the reads of an F. ananassa RNA-seq study to the F. vesca genome gives valid information about the gene expression of the octoploid species, as previously reported for another less closely related species such as Rubus sp. (García-Seco et al., 2015).
Auxin distribution by polar auxin transport is mediated by the PIN and AUX/LAX proteins (Vanneste and Friml, 2009). In the sequenced genome of the wild strawberry F. vesca, 10 FvPIN (Kang et al., 2013) and four FvAUX/LAX (Tennessen et al., 2014) (Supplementary Figure S1) genes have been identified. Only four FaPIN genes (FaPIN1, FaPIN4, FaPIN5, and FaPIN10) were expressed in the fruits of the cultivated strawberry (Fragaria ×ananassa), showing a developmental-specific pattern (Figures 2A,B). While FaPIN10 had higher expression in the achene than in the receptacle, the other three FaPIN genes displayed higher expression in the receptacle. In all cases, the pattern corresponded to decreased expression in the transition from the green to red stage, both in the achene and the receptacle. Regarding the F. ananassa AUX/LAX genes (FaAUX/LAX1, FaAUX/LAX2, FaAUX/LAX3, and FaAUX/LAX4), all were expressed in the receptacle and achenes (Figures 2C,D). In achenes, the highest expression corresponded to FaAUX/LAX4, while the other three members showed higher expression in the receptacle. In all cases, as occurred for FaPIN genes, the pattern was a continuous decrease from green to red stages in both the achene and receptacle. It is remarkable that most of the transport genes showed higher expression in the receptacle compared to the achene.
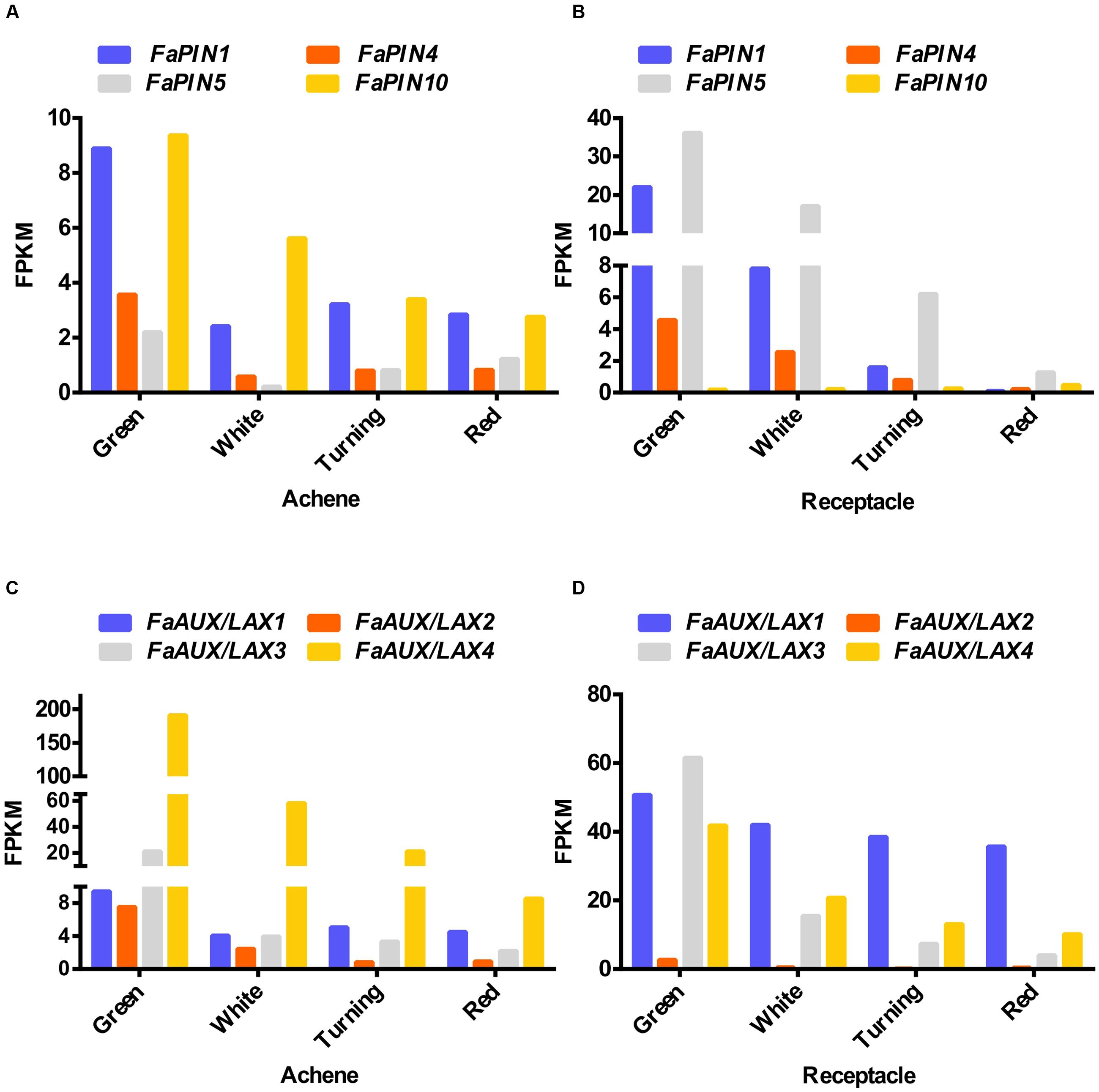
FIGURE 2. Expression of the four members of the FaPIN (A,B) and FaAUX/LAX (C,D) gene families in the achene (A,C) and the receptacle (B,D) at four developmental stages (green; white; turning; red), by RNAseq. FPKM, fragments per kilobase of exon per million fragments mapped.
The synthesis of auxin in the Trp-dependent pathway in Arabidopsis involves two steps that are catalysed by (i) the enzyme tryptophan aminotransferase (TAA and TAR), which converts tryptophan to indole-3-pyruvic acid (IPyA), and (ii) a family of flavin-dependent monooxygenases (YUCCA) that convert IPyA to indole-3-acetic acid (Stepanova et al., 2008, 2011). The TAA and YUCCA proteins jointly form a two-step biosynthetic route that constitutes the main auxin biosynthesis pathway in Arabidopsis. In addition, a number of studies in different species suggest that this pathway might be functional in fruits (Pattison et al., 2014). In the F. vesca genome, four tryptophan aminotransferases have been mapped (Supplementary Figure S2). The expression of the corresponding F. ananassa genes, analyzed by RNA-seq, shows that only three (FaTAA1, FaTAR1, and FaTAR2) are expressed in fruits (Figures 3A,B). In the achene, the three genes show the same pattern, i.e., a steep decrease in the transition from the green to the white stage (Figure 3A) and then decreasing to the red stage. The highest absolute value for expression is shown by FaTAR1. In the receptacle, the only gene showing expression over 1 FPKM is FaTAR2 (Figure 3B). Interestingly, the expression of this gene continuously increases from the green to the red stage. In relation to the YUCCA genes, only five of the nine genes mapped in F. vesca (Kang et al., 2013) are expressed in F. ananassa (Figures 3C,D). In the achene, the common pattern is a decrease from green to red, with the highest expression level being that of the FaYUC11 gene (Figure 3C). Even with low expression, some YUCCA genes were identified in the red receptacle (Figure 3D).
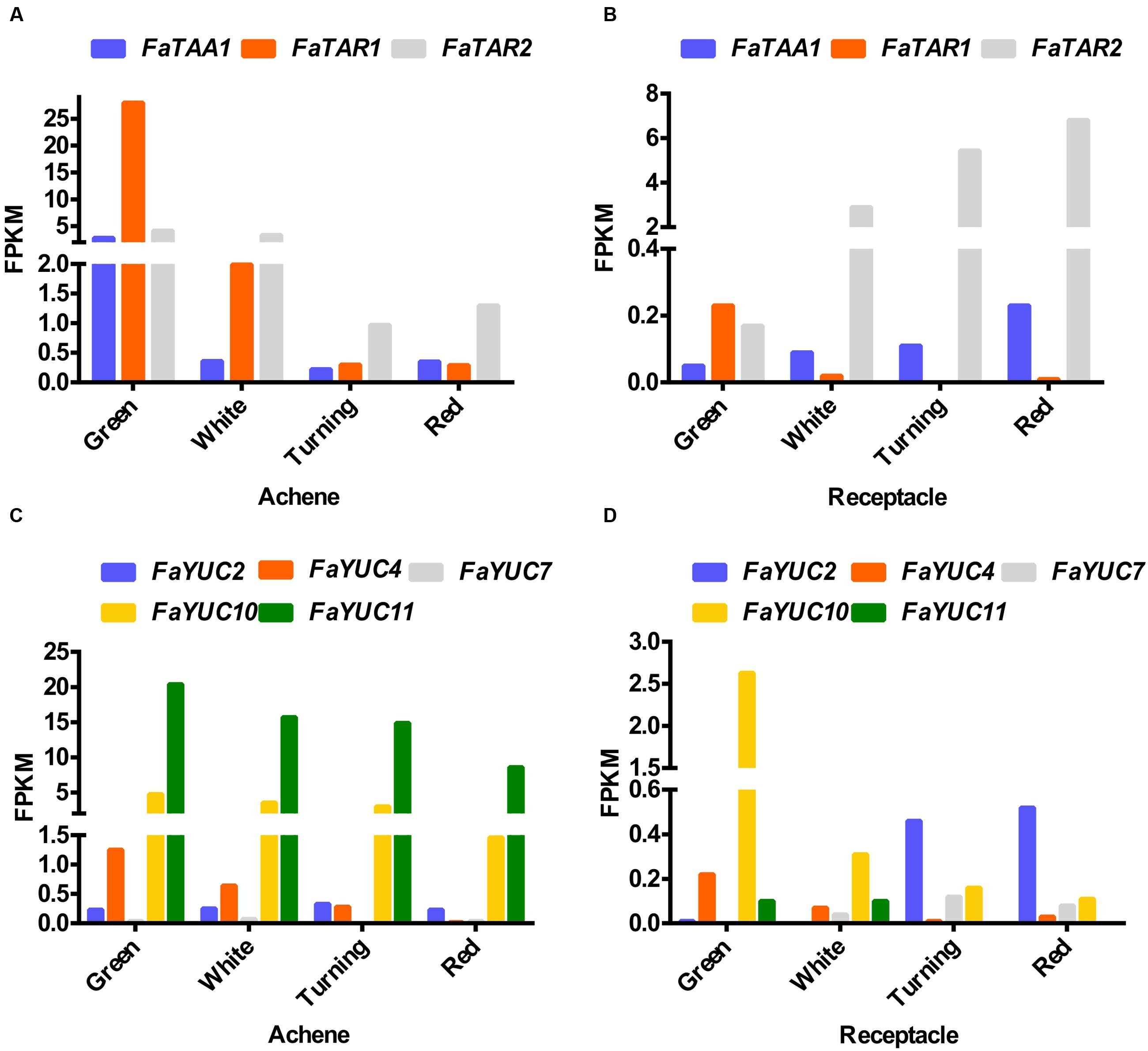
FIGURE 3. Expression of the three members of the tryptophan aminotransferase gene family (FaTAA1, FaTAR1, and FaTAR2) and four members of the flavin-dependent monooxygenases (FaYUC2, FaYUC4, FaYUC7, FaYUC10, and FaYUC11) in achene (A,C) and receptacle (B,D) at four developmental stages (green; white; turning; red), by RNAseq. FPKM, fragments per kilobase of exon per million fragments mapped.
Synthesis of auxin in plants from Trp has also been reported through the IAM pathway (Mano et al., 2010). A gene encoding a putative indole-3-acetamide hydrolase (AMI1) was identified in the F. vesca genome. The expression of the corresponding F. ananassa gene showed a decreased pattern from green to red stage in both achene and receptacle (Supplementary Table S2). Its contribution to the auxin content cannot be disregarded, mainly at early developmental stages. In addition to synthesis, homeostasis of auxin is maintained by the hormone conjugation and catabolism. The conjugation of auxin to amino acids is catalyzed by GH3 proteins (Staswick et al., 2005). Six members of the family were expressed in strawberry fruits showing a gene-specific pattern (Supplementary Table S2). In general, their expression is higher in achene. In receptacle, highest expression corresponds to FaGH3.1, that dramatically decreases from green to red stage. It is noteworthy than in grape berry the expression of the corresponding VvGH3.1 increased with ripening (Böttcher et al., 2010). This apparent discrepancy must be analyzed considering that the strawberry GH3 is a gene family, with gene-specific patterns, and the function of the auxin-conjugates is still under study (Ljung, 2013). There has also been reported the occurrence of amido-hydrolases in Arabidopsis (ILR1, ILL2, and IAR3) that convert back to free auxin some amino acid-conjugated forms of this hormone (LeClere et al., 2002). Two genes with high homology to these amido-hydrolases were identified in the F. vesca genome, and the corresponding F. ananassa genes showed a tissue- and developmental-specific pattern (Supplementary Table S2). The highest changes along fruit development were found for FaILR1 with a sharp decrease in expression from green to red achene, while expression in receptacle peaked at the turning stage. Interestingly, the presence of amide-conjugates of auxin in strawberry fruit has been reported long before, and changes in its content varied significantly only in achenes (Archbold and Dennis, 1984). Their function as a source of auxin must be considered, taking account the changes here reported on the expression of putative auxin-conjugating and de-conjugating enzymes.
Regarding auxin catabolism, a F. vesca gene with very high homology to the Arabidopsis AtDAO1 (Porco et al., 2016) was identified. The expression of the corresponding F. ananassa gene was higher in achene than in receptacle, increasing with maturation, and minor changes were observed in receptacle (Supplementary Table S2). Its contribution to auxin homeostasis in strawberry requires the confirmation of the enzyme activity and substrate specificity of the gene product.
Altogether, the expression of the auxin-synthesizing genes in strawberry fruits is directed to two organ/stage-specific points of active auxin biosynthesis, the green achene and, apparently at a lower level, the red receptacle. It is accepted that in the green fruit, the auxin synthesized in the achenes provides the hormone for the growth of the receptacle (Nitsch, 1955). The expression patterns of auxin transporters reported here (Figure 2) would support an active auxin transport in the receptacle. However, the synthesis of this hormone in red receptacle was unexpected. Therefore, focus was addressed to this organ, characterizing in more depth the synthesis and action of auxin in the ripening receptacle.
We first confirmed by qRT-PCR the expression of FaTAA1 and FaTAR2 in the ripening receptacle. The gene-specific primers designed from the assembled transcripts in the RNA-seq analysis (Grabherr et al., 2011) were used. The expression values obtained (Supplementary Figure S3) validated the previously found RNA-seq data (Figure 3B). In the transition from the green to the red receptacle, there is a significant increase in the expression of both FaTAA1 and FaTAR2. The increase was higher for FaTAR2, which also showed expression levels that were higher than those of FaTAA1 in the receptacle (Figure 3B).
The Trp aminotransferase activity of AtTAA1 has been studied in Arabidopsis to confirm its involvement in the production of IPyA from Trp (Stepanova et al., 2008). Thus, full-length cDNAs were cloned for FaTAA1 and FaTAR2 to produce the corresponding enzymes in E. coli, as described in the “Materials and Methods” section. The GST-purified proteins were assayed for aminotransferase activity in the presence of different concentrations of Trp, as previously described (Stepanova et al., 2008). The reaction products for FaTAA1 were analyzed by HPLC (Figures 4A,C,E). In the presence of Trp, two single peaks, corresponding to the enol and keto forms of IPyA, appeared at the end of the incubation period. Their identities were confirmed by HPLC analysis of the external standard. The controls, with boiled enzyme preparation and E. coli extract, are reported in Supplementary Figure S4. The transferase activity of the purified protein was also confirmed in gel, using a nitroblue tetrazolium dye (NBT) (Supplementary Figure S5).
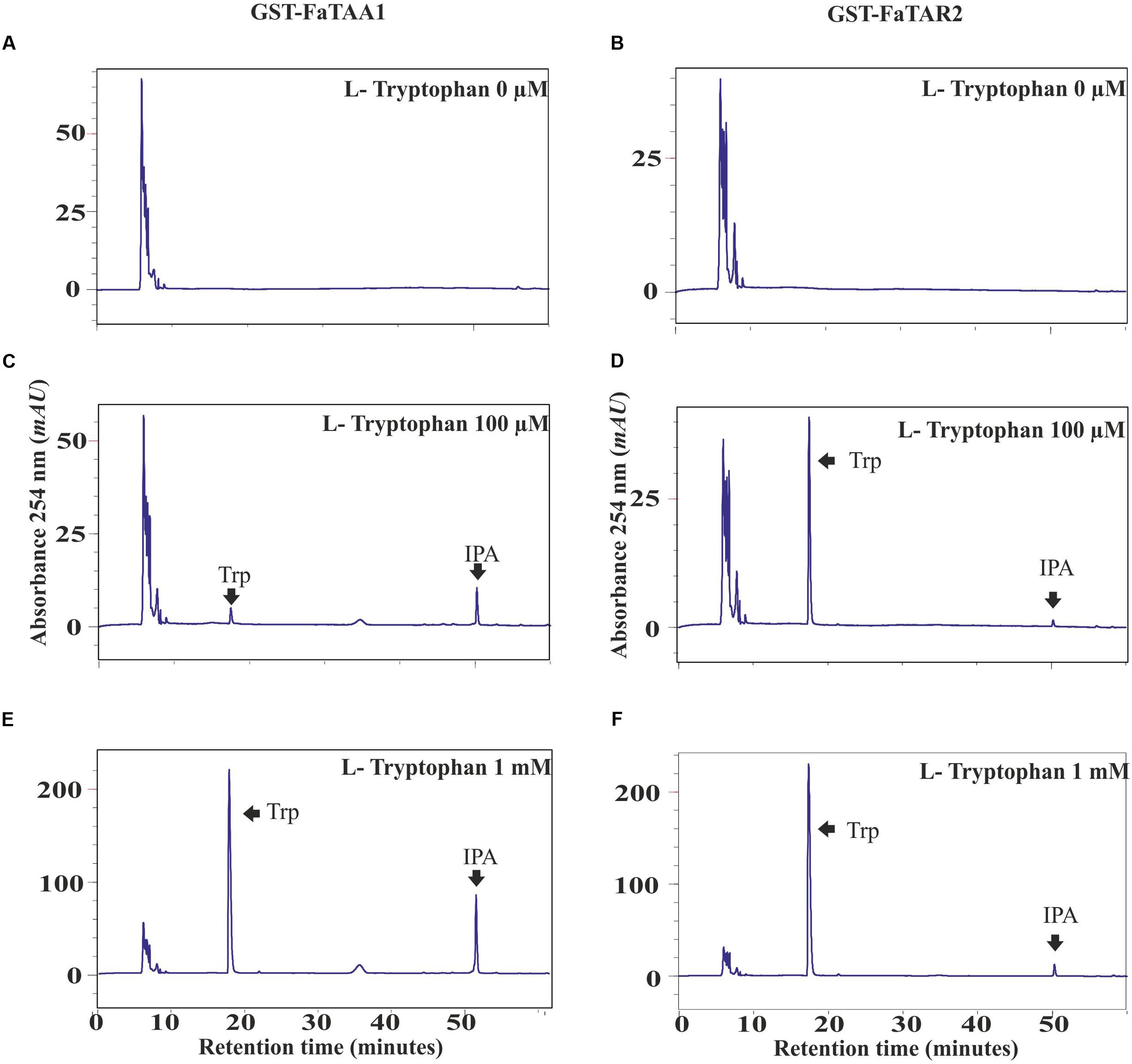
FIGURE 4. Trp aminotransferase (AT) activity of FaTAA1 and FaTAR2. HPLC chromatograms of the products of the in vitro AT reactions catalyzed by the purified GST-FaTAA1 (A–C) and GST-FaTAR2 (D–F) protein.
The same protocols were followed for the FaTAR2 protein. The chromatograms after incubation of the purified fusion protein GST-FaTAR2 with increasing concentrations of the substrate are shown in Figures 4B,D,F. At the highest concentration assayed, 1 mM Trp, a single peak corresponding to the more abundant enol form of IPyA was detected. The controls are shown in Supplementary Figure S4. The Trp aminotransferase activity of FaTAR2 was apparently lower than the activity of FaTAA1. However, this cannot be concluded from the present results as the protein concentration and purification degree of the two GST-fused enzymes, FaTAA1 and FaTAR2, were not evaluated. These results confirm that FaTAA1 and FaTAR2 work as Trp aminotransferases in strawberry as previously described in Arabidopsis, indicating that they are involved in auxin biosynthesis.
To investigate the role played by auxin in strawberry fruit development, the synthetic auxin response promoter DR5 (Ulmasov et al., 1997; Ottenschlager et al., 2003), fused to the β-glucuronidase (GUS) gene was used to generate transgenic strawberry plants. The roots of the transgenic strawberry plants showed a GUS expression pattern that was similar to that observed in the corresponding transgenic Arabidopsis (Ottenschlager et al., 2003) (Figure 5A, left), indicating that GUS expression in DR5-GUS transgenic strawberry plants is a bona fide indicator of auxin output signaling. Moreover, treatment with NAA increased the GUS staining in the root (Figure 5A), indicative of a functional response of the DR5 promoter to auxin in this organ. In the green fruit, DR5-driven GUS staining was observed in the receptacle around the achene attachment (Figure 5B). A closer view of a sectioned achene showed strong GUS staining in the embryo (Figure 5C). In the green receptacle surrounding the achenes, we observed a gradient of GUS staining that initiated at the achene (Figure 5D). Several attempts to visualize GUS activity in the red receptacle failed, as we always obtained a dispersed faint blue color that was not clearly visualized (Supplementary Figure S6). Either a dilution effect or the restriction of the activity to a limited set of cells could explain the weak stain obtained in the red receptacle. Then, GUS transcription was evaluated in the receptacle at three stages, as this process could overcome these problems. The result showed that in the red receptacle, the expression was maintained, and slightly higher, compared to the green and white stages (Figure 5E). Overall, the pattern of GUS activity and GUS expression in strawberry fruits confirms the previously reported auxin activity in the achene and green receptacle and the auxin responsiveness in red receptacle.
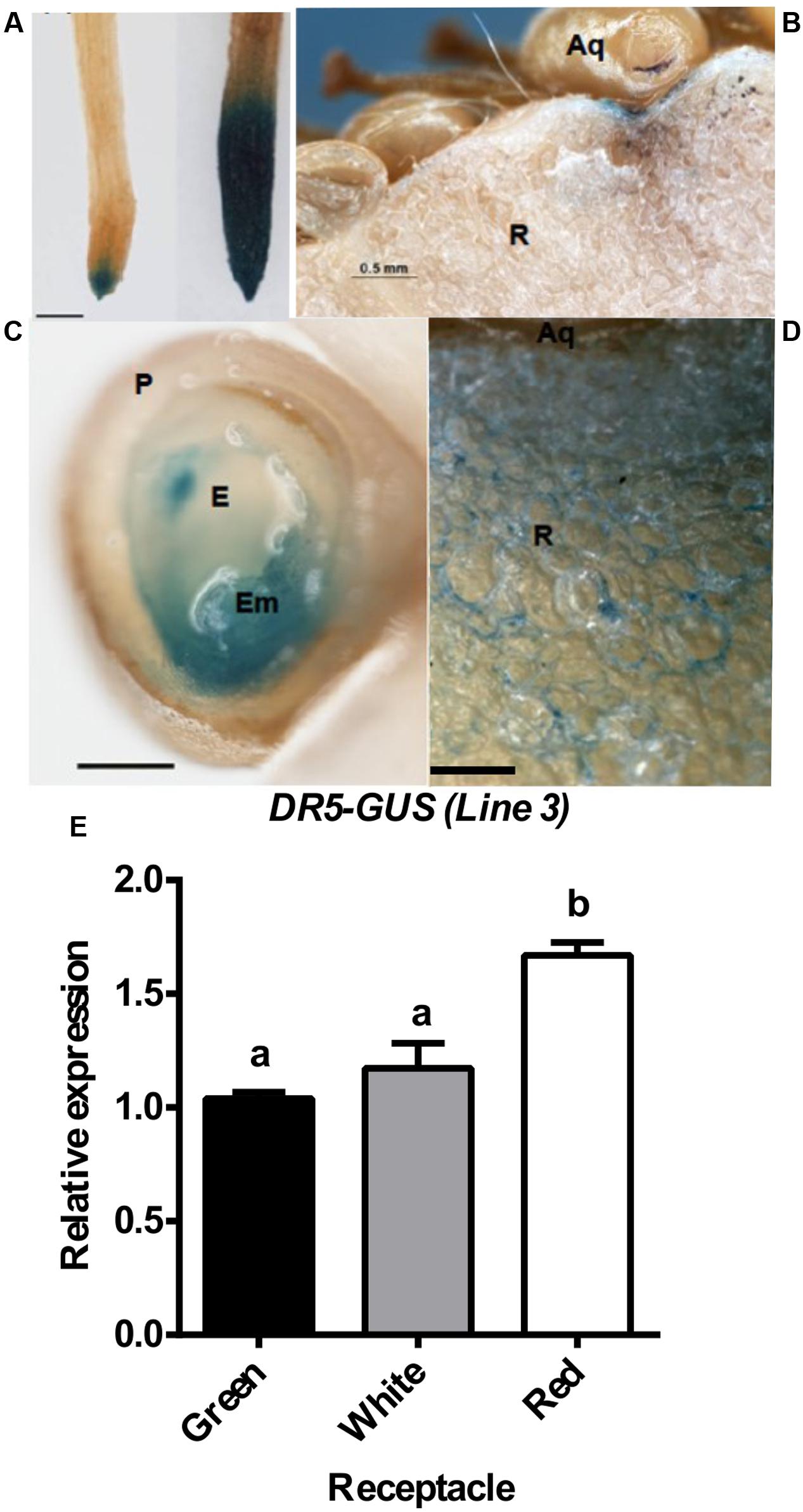
FIGURE 5. Auxin distribution in strawberry. DR5-GUS expression pattern in strawberry roots (A) and green fruits (B–D). For the NAA treatment, in (A), plants were grown in vitro and treated with a solution of 50 μM NAA in water for 24 h (right), or just with water for control plants (left). Scale bars (B) = 500 μm, (C,D) = 250 μm,. P, pericarp; E, endosperm; Em, embryo; Aq, achene; R, receptacle. (E) Relative expression by qRT-PCR of the GUS gene in receptacle at three developmental stages. Different letters indicate a significant difference between samples according to the corresponding ANOVA (P < 0.05).
Transcriptional responses to auxin are continuously evaluated and result from the interplay of a number of elements that conform to the signaling pathway (Weijers and Wagner, 2016). We restricted our analysis to the receptacle as the main focus of our research. The pathway starts with the auxin receptor genes TIR1 and AFB (Dharmasiri et al., 2005). In the F. vesca genome, four auxin receptors are present, while three are expressed in F. ananassa (Supplementary Figure S7). Their study in the receptacle shows that FaTIR1 slightly increases its expression from the green to red stage, while the expression of FaAFB2 decreases, and FaAFB5 remains constant (Figure 6A). The analysis was extended to the repressors Aux/IAA and the transcription factors ARFs, whose interaction is critical to trigger the transcriptional response to auxin. In most species, a high number of members have been found for these gene families. Thus, in F. vesca, 21 Aux/IAA and 19 ARF genes have been identified (Kang et al., 2013). The possible selectivity of the Aux/IAA-ARF interactions determined by their structural features (Piya et al., 2014) and their co-expression pattern would account for the specificity of the auxin involvement in various developmental processes (Weijers and Wagner, 2016). Therefore, the expression in the developing receptacle of the corresponding genes of F. ananassa (FaAux/IAA and FaARF) was analyzed in the RNA-seq expression data. A total of 19 FaAux/IAA genes were expressed in ripening receptacle of strawberry fruit with different patterns (Figure 6B). Most of them showed the highest expression at the green stage and then continuously decreased up to the red stage, in agreement with the occurrence of auxin in the green receptacle (Figure 6B). However, three genes showed a different pattern. The FaAux/IAA8a gene showed the highest expression at all stages, with a small increase from the green to red stage. Two other genes, FaAux/IAA14b and FaAux/IAA11, increased their expression up to a maximum at the turning stage and then decreased (Figure 6B). The expression of the FaARF genes is shown in Figure 6C. Most of the genes showed decreasing expression, with minor changes, from the green to red stage, but the FaARF6a gene, whose expression was the highest in the receptacle compared to the other FaARFs, showed a dramatic increase from 77 FPKM at the green to 236 FPKM at the red stage (Figure 6C).
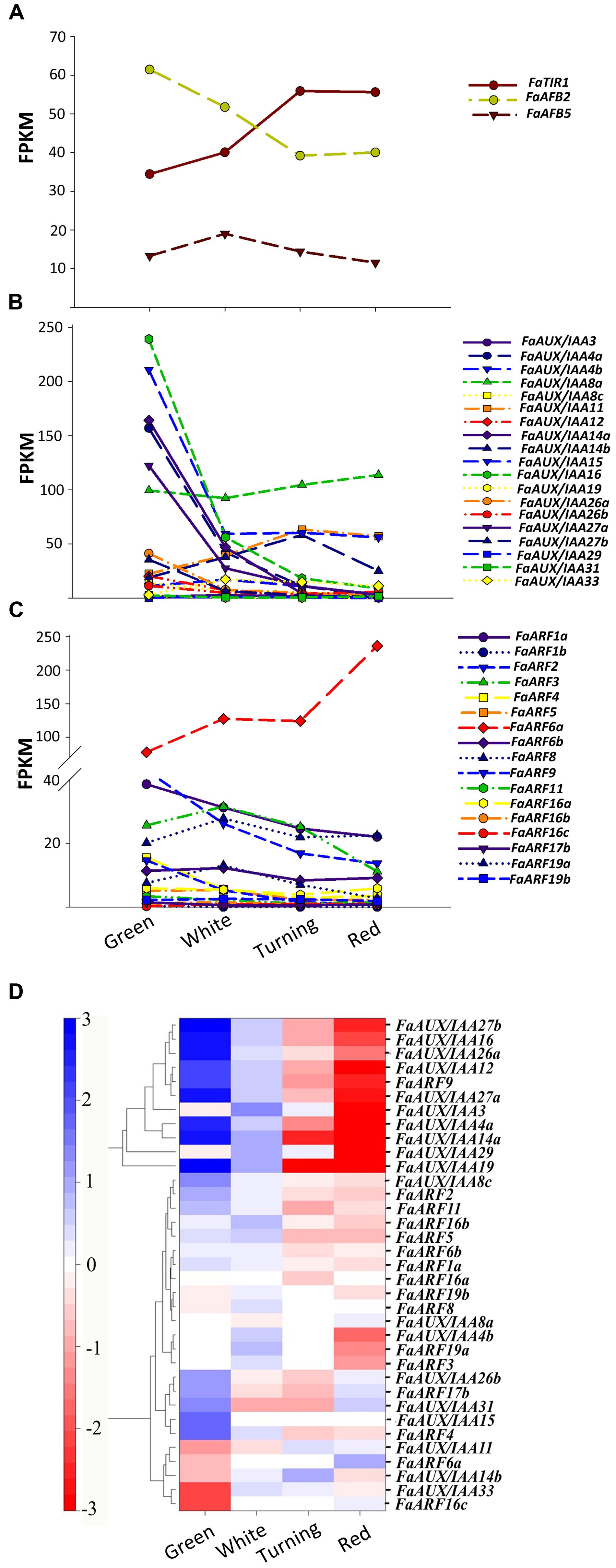
FIGURE 6. Expression of the gene family of auxin receptors (A), repressors (FaAUX/IAA; B), and transcription factors (FaARF; C) in the receptacle at four developmental stages (green; white; turning; red) by RNAseq. FPKM, fragments per kilobase of exon per million fragments mapped. (D) K-means clusters of 35 genes showing distinct stage- and tissue-specific expression patterns. The scale: averaged log2 “relative RPKM value” of all genes in each cluster.
Clustering by co-expression analysis of the FaAux/IAA and FaARF genes in the receptacle during ripening was performed to identify the possible positive interactions between members of the two gene families (Figure 6D). Two main clusters are formed. One of them (upper) includes those genes with the highest decrease in the transition from the green to red stage. The other cluster includes genes that showed variable and not very drastic changes in expression during the growth and ripening of the receptacle. In the lower part of this cluster are grouped, in a sub-cluster, those genes whose expression increased with the progression of ripening (FaAux/IAA11, FaARF6a, FaAux/IAA14b, FaAux/IAA33, and FaARF16c). In this group, the two genes showing the highest expression level and highest change in the transition from the green to red stage are FaAux/IAA11 and FaARF6a.
The values obtained for the expression of key genes of the auxin reception and signaling pathway in the ripening receptacle by RNA-seq were validated by qRT-PCR. The results, shown in Figure 7, confirm the RNA-seq data, i.e., there is an increase in the expression of the genes FaAux/IAA11 (Figure 7A) and FaARF6a, (Figure 7B), jointly with the auxin receptor FaTIR1 (Figure 7C), parallel to the ripening process. Altogether, these results point to the operation of the auxin responsive components of the strawberry receptacle at the ripe stage.
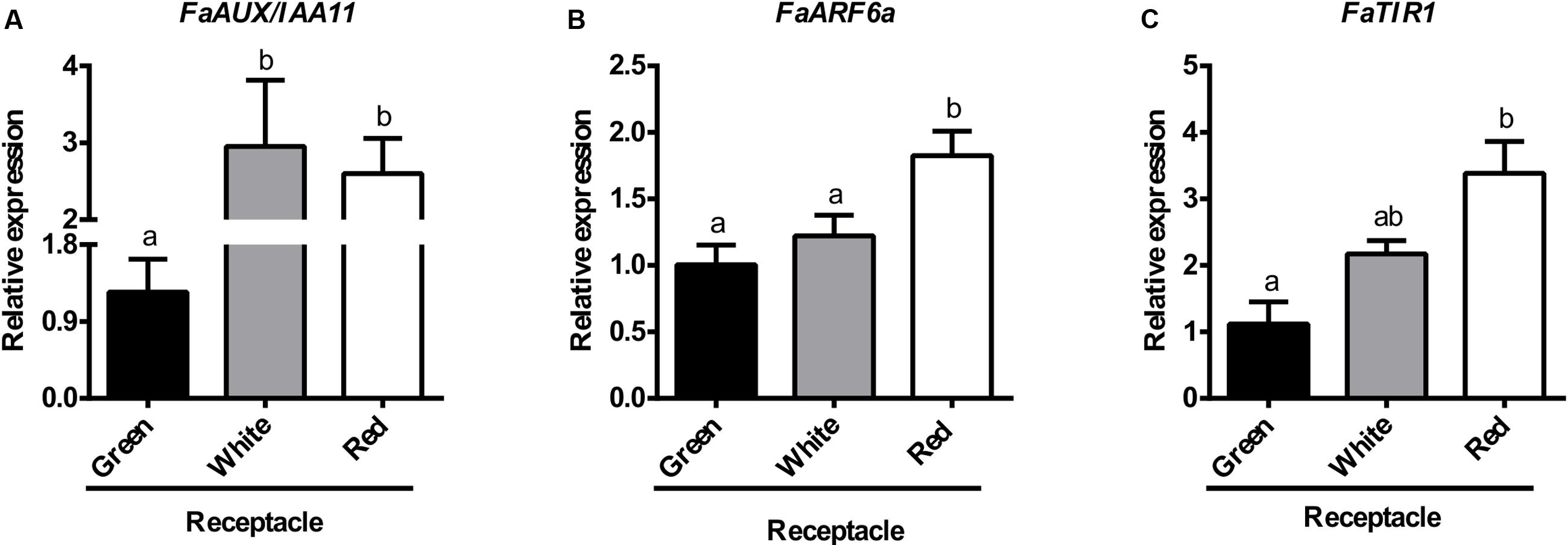
FIGURE 7. Relative transcript levels of FaAUX/IAA11 (A), FaARF6a (B), and FaTIR (C), as determined by qRT-PCR in different developmental stages of receptacle. Bars represent the mean of three independent biological samples ± SE. Different letters indicate a significant difference between samples according to the corresponding ANOVA (P < 0.05).
In relation to a proteasome-independent auxin signaling pathway, a gene with high homology to the Arabidopsis auxin binding protein ABP1 (Sauer and Kleine-Vehn, 2011) was identified in F. vesca, being its expression in F. ananassa higher in achene than in receptacle, where did not show significant differences along ripening (Supplementary Table S2).
Not only were the auxin reception and signaling genes expressed in ripe receptacle but so were the genes encoding for Trp aminotransferase (FaTAA1, FaTAR2) (Supplementary Figure S3). This result points to the synthesis of this hormone within this organ at this stage. Therefore, it was possible to interfere with the auxin biosynthetic pathway in this organ at this stage. In strawberry, transient silencing in fruits is a valid alternative to permanent transformation for those genes expressed in this organ (Hoffmann et al., 2006). Thus, RNAi silencing of the biosynthetic gene with the highest value of expression in the red receptacle, FaTAR2, was performed. Two of six fruits injected with the RNAi construct showed a significant reduction in the expression of FaTAR2 gene in the receptacle (Supplementary Figure S8). Silencing of chalcone synthase (CHS) was used as the positive control (Hoffmann et al., 2006). The phenotype of the silenced fruits did not show any apparent difference in relation to the control, but the CHS-silenced fruits showed a characteristic lack of color development (Figure 8A).
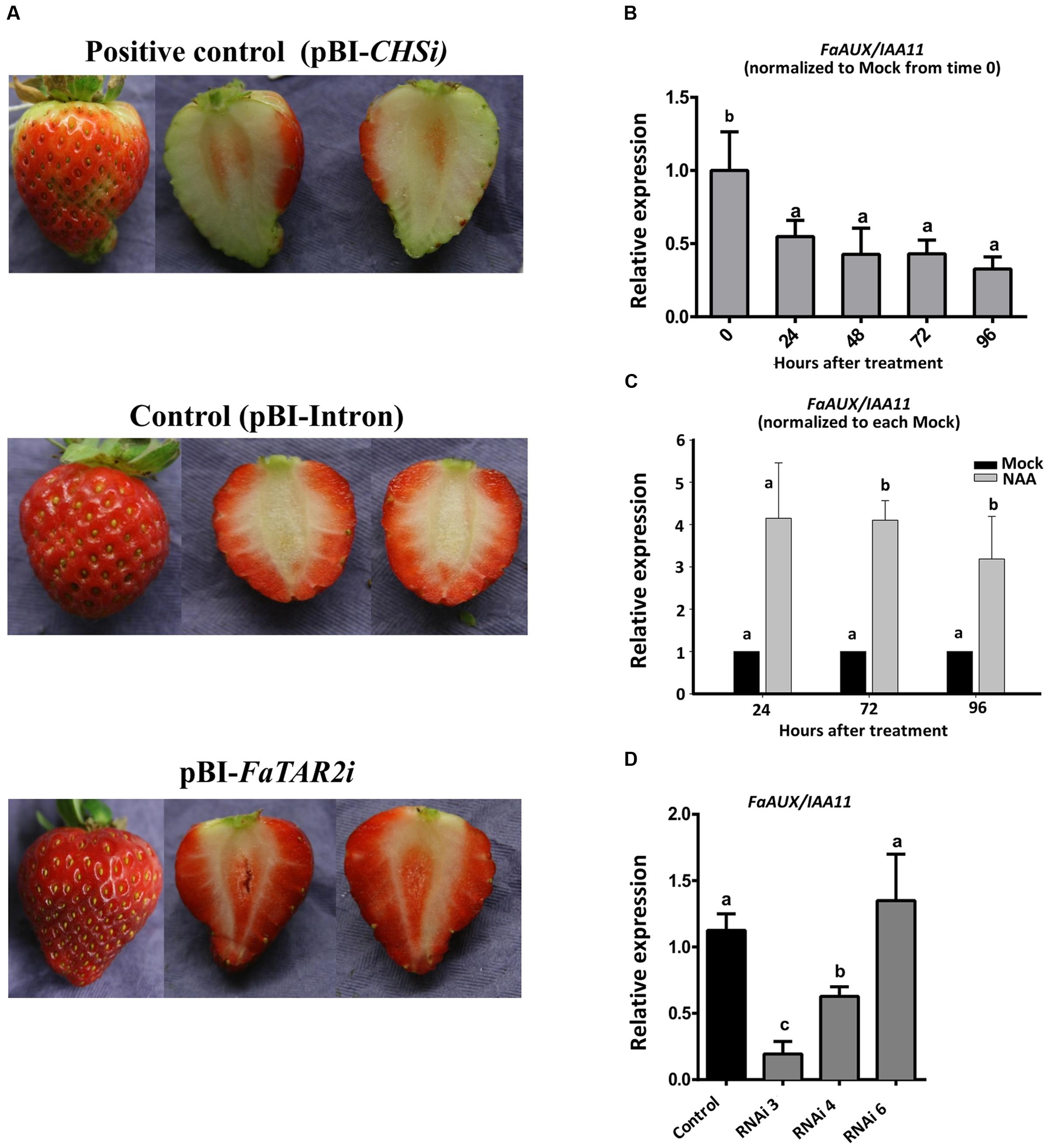
FIGURE 8. (A) Strawberry transgenic fruits agroinfiltrated with empty vector (control), with the FaTAR2-RNAi construct (RNAi) and with Chalcona synthase-RNAi, FaCHS-RNAi, as a positive control at 7 days after injection. (B) FaAUX/IAA11 expression by qRT-PCR of de-achened green strawberry fruits and covered with a lanoline paste. (C) Time course expression of FaAUX/IAA11, by qRT-PCR, of the receptacle of strawberry fruits injected, at the turning stage, with a water solution of NAA and the corresponding control (mock solution). (D) FaAUX/IAA11 expression in silenced fruit agroinfiltrated with empty vector (control) and with FaTAR2-RNAi construct. Error bars indicate +SE of three biological replicates. Different letters indicate a significant difference between samples according to the corresponding ANOVA (P < 0.05).
The silencing of FaTAR2 is expected to be accompanied by a decrease in the auxin content. The small amount of the sample made it unfeasible to quantify the hormone in the silenced fruits. However, transcript detection of auxin-regulated genes was a valid alternative. The Aux/IAA genes are transcriptionally up-regulated by auxin treatment, likely as part of a feedback control mechanism (Abel et al., 1994; Krogan and Berleth, 2015). Thus, we tested whether the FaAux/IAA11 gene, which is highly expressed in the ripe strawberry receptacle (Figure 6B), was sensitive to auxin changes. For this purpose, achenes were removed from green fruits and the expression of FaAux/IAA11 followed in the receptacle. The expression of this gene clearly decreased after 24 h removal of the achenes, and continued up to 96 h (Figure 8B). Another set of fruits, at the turning stage, were injected with a solution of NAA. The expression of FaAux/IAA11 in the receptacle was increased in comparison with fruits injected with the mock (Figure 8C). These results confirmed the effect of auxin on the transcription of this gene. Thus, the decreased expression of FaAux/IAA11 in the FaTAR2-silenced receptacle (Figure 8D) might be an indication of a reduced auxin concentration in this organ in comparison with control fruits and fruits where silencing was not effective. Altogether, these results support the synthesis of auxin in the red receptacle. This process is accompanied by a high expression of genes of the auxin reception and signaling pathway, such as FaTIR1, FaAux/IAA11, and FaARF6a, in this organ at this stage.
During recent years, there has been a continuous advancement in the comprehension of its synthesis, transport, perception, and signaling (Ljung, 2013; Adamowski and Friml, 2015; Kramer and Ackelsberg, 2015; Salehin et al., 2015; Weijers and Wagner, 2016). The study of these components in different organ/cell scenarios has been useful to reveal their involvement in cellular processes such as cell division, cell enlargement (Velasquez et al., 2016), cell differentiation (Yang and Wang, 2016), and subcellular processes such as remodeling of the cell wall (Pacheco-Villalobos et al., 2016) and lignin biosynthesis (Zhang et al., 2014). Most of them occur at some stage of strawberry fruit development.
The global transcriptional data in the two parts of the strawberry fruit, the achene and the receptacle, separately showed a specific pattern for gene expression of every member of the gene families FaPIN, FaAUX1/LAX, FaTIR1/AFB, FaTAA1/TAR, FaAux/IAA, and FaARF. Focusing on the receptacle, it is noteworthy the high expression of auxin transport genes at the green stage, and the enhanced transcription of some members of the auxin perception and signaling families at the red stage.
Our expression analysis of the genes encoding tryptophan amino transferases, whose in vitro activity of the enzymes has been demonstrated, supports that active auxin biosynthesis occurs in the green achenes and decreases thereafter. The pattern of expression of the auxin transporters supports that active transport is occurring to or from the receptacle. An important observation is the maintenance of auxin content in the receptacle of the ripe fruits, something that was previously reported many years ago but went relatively unnoticed. Analysis of the free auxin content separately in the achene and the receptacle revealed that the levels peaked at the same stage in both organs, corresponding to the transition from free nuclear to cellular endosperm in the achene and the initiation of the exponential growth of the receptacle. Afterward, the auxin content decreased in both tissues, but a late increase was found during ripening in the receptacle (Archbold and Dennis, 1984). The IAA values measured by us in the receptacle are in the range of those previously reported for the receptacle of wild F. vesca (Osorio et al., 2011) and for the white receptacle of the cultivated strawberry (Symons et al., 2012). They are approximately 10-fold lower than the values in other plant tissues, such as Arabidopsis roots (Basu et al., 2011) and tomato roots, leaves and fruits (Albacete et al., 2008; Pattison and Catalá, 2011), and more than 20-fold lower than in the achenes at the white stage (Symons et al., 2012). Thus, even at a low level, auxin is present in the ripe receptacle, and this is a novelty of the present work. Moreover, the increase in the expression of FaTAR2 with ripening, as well as in specific members of the FaAux/IAA (FaAux/IAA11, FaAux/IAA14b, FaAux/IAA33) and FaARF (FaARF6a, FaARF16c) families, points to a cell-autonomous auxin synthesis and action in the ripe receptacle.
The strawberry receptacle consists of a fleshy pith at the center surrounded by cortical tissue containing parenchymal and epidermal cells. Vascular bundles traverse the pith and the cortex to the achenes (Perkins-Veazie, 1995). During growth and ripening, the different cell types differ in their division rate, final cell size, cell wall composition and metabolic activity (Darrow, 1966; Fait et al., 2008; Nasopoulou et al., 2014). The processes associated with these changes involve cell wall remodeling, lignin biosynthesis and drastic changes in primary and secondary metabolism. Therefore, we hypothesize that the action of auxin in the ripening receptacle might be spatially limited to specific cell types, and associated with some of these processes.
The high number of Aux/IAA and ARF proteins in most of the species offers a wide range of combinatorial interactions that would account for the numerous specific plant processes in which auxin participates. This complex interaction network translates the local accumulation of auxin to gene expression by specific ARF transcription factors (Weijers and Wagner, 2016). In the sequenced genome of diploid F. vesca, 21 members of the Aux/IAA family and 19 members of the ARF family have been identified (Kang et al., 2013). Our analysis of the F. ananassa transcriptome in this species showed the expression of 19 Aux/IAA and 16 ARF members of these families in fruit, at different developmental stages, leaf and root. Their diverse expression pattern reveals the existence of a complex auxin signaling machinery that is prepared for the action of this hormone in different processes. Moreover, co-expression analysis might identify the possible Aux/IAA-ARF interacting partners involved in specific tissue/stage processes. Thus, in the ripening receptacle, where we here report the synthesis of auxin by an active FaTAR2 enzyme, the FaAux/IAA11 and FaARF6a clustered together. The nomenclature of the F. vesca genes of these families (Kang et al., 2013), as well as the F. ananassa genes, was determined by their sequence closeness to the Arabidopsis genes. In this species, it was found that AtARF6 interacted in a yeast two-hybrid assay, confirmed by bimolecular fluorescence complementation, with a number of Aux/IAA proteins, including AtAux/IAA11 (Piya et al., 2014). Moreover, co-expression analysis of the two gene families, Aux/IAA and ARF, showed that positive correlation in this species was high for AtAux/IAA11 and AtARF6 in flower buds and flowers (Piya et al., 2014). Altogether, our results point to FaARF6a as a main final transcriptional regulator, likely interacting with FaAux/IAA11, translating the activity of auxin in the ripening receptacle.
Sequence analysis of FaARF6a identified in the encoded protein the characteristic features of the class A ARFs (Supplementary Figure S9), which includes the N-terminal B3 DNA-binding domain, flanked by the dimerization domains, the Q-rich activation domain, and the C-terminal PB1 domain for oligomerization and Aux/IAA-ARF heterodimerization (Guilfoyle and Hagen, 2012; Weijers and Wagner, 2016). The class A ARFs are classified as transcriptional activators (Ulmasov et al., 1999). Thus, it is expected that FaARF6a targets genes with enhanced expression in the receptacle as ripening advances in this organ.
In Arabidopsis, AtARF6 has been demonstrated to play a central role in auxin’s regulation of cell elongation in the hypocotyl (Oh et al., 2014). The identification of AtARF6 targets was performed by chromatin-immunoprecipitation followed by sequencing (ChIP-Seq) analysis and further comparison with genes previously described as auxin-activated (Tiwari et al., 2003). A total of 255 AtARF6 targets were identified, many of them involved in cell elongation. Interestingly, in the ripening receptacle, cell enlargement is a process that increases rapidly when cell division ceases (Cheng and Breen, 1992). At the transition from the green to white stage, the highest increase in the expression of FaARF6a occurs. Moreover, in elongating hypocotyl, the DNA binding capacity of AtARF6 was blocked by the gibberellin-inactivated repressor RGA (Oh et al., 2014), which constitutes a good example of gibberellin involvement in the auxin response. In the ripening receptacle, we have previously reported the involvement of gibberellin in its development, with a peak at the white stage, and FaRGA expression dramatically decreasing from the green to white stage (Csukasi et al., 2011). Whether FaARF6a constitutes a link between auxin and gibberellin in the cell enlargement taking place in the ripening receptacle deserves to be investigated. Interestingly, in strawberry fruits where the FaGAMYB gene was silenced, the expression of FaARF6 was significantly down-regulated (Vallarino et al., 2015). FaGAMYB is a GA-responsive transcription factor that plays a key role in the changes occurring in receptacle development prior to its ripening. An analysis of the FaARF6a targets during this process would provide information on the specific molecular processes regulated by this gene in the strawberry receptacle. Moreover, cell elongation is not a uniform process in strawberry receptacle development. Thus, there is a distinct gradient in cell size in the central pith and in the cortex, with the smaller cells near the periphery and the larger ones toward the inside (Havis, 1943). In addition, the cortex develops more rapidly than the pith and at a higher relative rate. All of these points mean that the cell elongation in the developing receptacle might be circumscribed to specific groups of cells, depending on time and space. This result must be considered when investigating the molecular events associated with cell elongation and the involvement of specific gene products such as FaARF6a.
VV conceived the project. NM-E, MB, SO, and VV planned, designed and supervised the research. EE-J, FC, CP, JV, YK, AV, JB, JS-S, and SO performed the experiments. All authors contribute to data analysis and the writing of the manuscript.
The project was funded by Grants BIO2013-44199-R and AGR12-40066-CO2-02 (MINECO, Spain). EE-J and FC were awarded with a FPI fellowship (MINECO, Spain). SO was supported Ministerio de Ciencia e Innovación and University of Malaga (Spain) through the grant Ramón and Cajal program (Sonia Osorio, RYC-09170).
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The Supplementary Material for this article can be found online at: http://journal.frontiersin.org/article/10.3389/fpls.2017.00889/full#supplementary-material
Abel, S., Oeller, P. W., and Theologis, A. (1994). Early auxin-induced genes encode short lived nuclear proteins. Proc. Natl. Acad. Sci. 91, 326–330. doi: 10.1073/pnas.91.1.326
Adamowski, M. and Friml, J. (2015) PIN-dependent auxin transport: action, regulation, and evolution. The Plant Cell 27, 20–32. doi: 10.1105/tpc.114.134874
Aharoni, A., Keizer, L. C. P., Van Den Broeck, H. C., Blanco-Portales, R., Munoz-Blanco, J., Bois, G., et al. (2002). Novel insight into vascular, stress, and auxin-dependent and -independent gene expression programs in strawberry, a non-climacteric fruit. Plant Physiol. 129, 1019–1031. doi: 10.1104/pp.003558
Albacete, A., Ghanem, M. E., Martínez-Andújar, C., Acosta, M., Sánchez-Bravo, J., and Martínez, V. et al. (2008). Hormonal changes in relation to biomass partitioning and shoot growth impairment in salinized tomato (Solanum lycopersicum) plants. J. Exp. Bot. 59, 4119–4131. doi: 10.1093/jxb/ern251
Archbold, D. D., and Dennis, F. G. (1984). Quantification of fre aba and free and conjugated IAA in strawberry achene and receptacle tissue during fruit development. J. Amer. Soc. Hortic. Sci. 109, 330–335.
Basu, P., Brown, K. M., and Pal, A. (2011). Detailed quantitative analysis of architectural traits of basal roots of young seedlings of bean in response to auxin and ethylene. Plant Physiol. 155, 2056–2065. doi: 10.1104/pp.110.168229
Benjamins, R., and Scheres, B. (2008). Auxin: the looping star in plant development. Annu. Rev. Plant Biol. 59, 443–465. doi: 10.1146/annurev.arplant.58.032806.103805
Bombarely, A., Merchante, C., Csukasi, F., Cruz-Rus, E., Caballero, J. L., Medina-Escobar, N., et al. (2010). Generation and analysis of ESTs from strawberry (Fragaria x ananassa) fruits and evaluation of their utility in genetic and molecular studies. BMC Genomics 17:503. doi: 10.1186/1471-2164-11-503
Böttcher, C., Keyzers, R. A., Boss, P. K., and Davies, C. (2010). Sequestration of auxin by the indole-3-acetic acid-amido synthetase GH3-1 in grape berry (Vitis vinifera L.) and the proposed role of auxin conjugation during ripening. J. Exp. Bot. 61, 3615–3625. doi: 10.1093/jxb/erq174
Brumos, J., Alonso, J. M., and Stepanova, A. N. (2014). Genetic aspects of auxin biosynthesis and its regulation. Physiol. Plant. 151, 3–12. doi: 10.1111/ppl.12098
Calderón Villalobos, L. I., Lee, S., De Oliveira, C., Ivetac, A., Brandt, W., Armitage, L. et al. (2012). A combinatorial TIR1/AFB-Aux/IAA co-receptor system for differential sensing of auxin. Nat. Chem. Biol. 8, 477–485. doi: 10.1038/nchembio.926
Castillejo, C., de la Fuente, J., Iannetta, P., Botella, M., and Valpuesta, V. (2004). Pectin esterase gene family in strawberry fruit: study of fape1, a ripening-specific isoform. J. Exp. Bot. 55, 909–918. doi: 10.1093/jxb/erh102
Cheng, G. W., and Breen, P. J. (1992). Cell count and size in relation to fruit size among strawberry cultivars. J. Am. Soc. Hortic. Sci. 117, 946–950.
Chevreux, B., Pfisterer, T., Drescher, B., Driesel, A. J., Müller, W. E., Wetter, T., et al. (2004). Using the miraEST assembler for reliable and automated mRNA transcript assembly and SNP detection in sequenced ESTs. Genome Res. 14, 1147–1159. doi: 10.1101/gr.1917404
Clancy, M. A., Rosli, H. G., Chamala, S., Barbazuk, W. B., Civello, P. M., and Folta, K. M. (2013). Validation of reference transcripts in strawberry (Fragaria spp.). Mol. Genet. Genomics 288, 671–681. doi: 10.1007/s00438-013-0780-6
Conesa, A., Madrigal, P., Tarazona, S., Gomez-Cabrero, D., Cervera, A., McPherson, A., et al. (2016). A survey of best practices for RNA-seq data analysis. Genome Biol. 17:13. doi: 10.1186/s13059-016-0881-8.
Csukasi, F., Osorio, S., Gutierrez, J. R., Kitamura, J., Giavalisco, P., Nakajima, M., et al. (2011). Gibberellin biosynthesis and signalling during development of the strawberry receptacle. New Phytol. 19, 376–390. doi: 10.1111/j.1469-8137.2011.03700.x
Darrow, G. M. (1966). The Strawberry: History, Breeding and Physiology. New York, NY: Holt, Rinehart and Winston.
Dharmasiri, N., Dharmasiri, S., Weijers, D., Lechner, E., Yamada, M., Hobbie, L., et al. (2005). Plant development is regulated by a family of auxin receptor f box proteins. Dev. Cell 9, 109–119. doi: 10.1016/j.devcel.2005.05.014
Dreher, K. A., Brown, J., Saw, R. E., and Callis, J. (2006). The arabidopsis aux/iaa protein family has diversified in degradation and auxin responsiveness. Plant Cell 18, 699–714. doi: 10.1105/tpc.105.039172
Fait, A., Hanhineva, K., Beleggia, R., Dai, N., Rogachev, I., Nikiforova, V. J., et al. (2008). Reconfiguration of the achene and receptacle metabolic networks during strawberry fruit development. Plant Physiol. 148, 730–750. doi: 10.1104/pp.108.120691
Finet, C., Berne-Dedieu, A., Scutt, C. P., and Marlétaz, F. (2012). Evolution of the ARF gene family in land plants: old domains, new tricks. Mol. Biol. Evol. 30, 45–56. doi: 10.1093/molbev/mss220
García-Seco, D., Zhang, Y., Gutierrez-Mañero, F. J., Martin, C., and Ramos-Solano, B. (2015). RNA-Seq analysis and transcriptome assembly for blackberry (Rubus sp. Var. Lochness) fruit. BMC Genomics 16:5. doi: 10.1186/s12864-014-1198-1
Given, N. K., Venis, M. A., and Grierson, D. (1988). Hormonal regulation of ripening in the strawberry, a non-climacteric fruit. Planta 174, 402–406. doi: 10.1007/BF00959527
Grabherr, M. G., Haas, B. J., Yassour, M., Levin, J. Z., Thompson, D. A., and Amit, I. et al. (2011). Full-length transcriptome assembly from RNA-seq data without a reference genome. Nat. Biotechnol. 29, 644–652. doi: 10.1186/s12864-014-1198-1
Guilfoyle, T. J. (2015). The PB1 domain in auxin response factor and Aux/IAA Proteins: a versatile protein interaction module in the auxin response. Plant Cell 27, 33–43. doi: 10.1105/tpc.114.132753
Guilfoyle, T. J., and Hagen, G. (2007). Auxin response factors. Curr. Opin. Plant Biol. 10, 453–460. doi: 10.1016/j.pbi.2007.08.014
Guilfoyle, T. J., and Hagen, G. (2012). Getting a grasp on domain III/IV responsible for Auxin Response Factor-IAA protein interactions. Plant Sci. 190, 82–88. doi: 10.1016/j.plantsci.2012.04.003
Harpster, M. H., Brummell, D. A., and Dunsmuir, P. (1998). Expression analysis of a ripening-specific, auxin-repressed endo-1,4-beta -glucanase gene in strawberry. Plant Physiol. 118, 1307–1316. doi: 10.1104/pp.118.4.1307
Havis A. L. (1943). Developmental analysis of the strawberry fruit. Am. J. Bot. 30, 311–314. doi: 10.2307/2437460
Hoffmann, T., Kalinowsk, G., and Schwab W. (2006). RNAi-induced silencing of gene expression in strawberry fruit (Fragaria x ananassa) by agroinfiltration: a rapid assay for gene function analysis. Plant J. 48, 818–826. doi: 10.1111/j.1365-313X.2006.02913.x
Jefferson, R. A., Kavanagh, T. A., and Bevan, M. W. (1987). Gus fusion: B-glucoronidase as a sensitive and versatile gene fusion marker in higer plants. EMBO J. 6, 3901–3907.
Kang, C., Darwish, O., Geretz, A., Shahan, R., Alkharouf, N., and Liu, Z. (2013). Genome-scale transcriptomic insights into early-stage fruit development in woodland strawberry Fragaria vesca. Plant Cell 25, 1960–1978. doi: 10.1105/tpc.113.111732
Kepinski, S., and Leyser, O. (2005). The Arabidosis F-box protein TIR1 is an auxin receptor. Nature 435, 446–451. doi: 10.1038/nature03542
Koshiba, T., and Matsuyama, H. (1993) An in vitro system of indole-3- acetic acid formation from tryptophan in maize (Zea mays) coleoptile extracts. Plant Physiol. 102, 1319–1324. doi: 10.1104/pp.102.4.1319
Kramer, E. M., and Ackelsberg, E. M. (2015). Auxin metabolism rates and implications for plant development. Front. Plant Sci. 6:150. doi: 10.3389/fpls.2015.00150
Krogan, N. T., and Berleth, T. (2015). The identification and characterization of specific ARF-Aux/IAA regula- tory modules in plant growth and development. Plant Signal. Behav. 10:e992748. doi: 10.4161/15592324.2014.992748
LeClere, S., Tellez, R., Rampey, R. A., Matsuda, S. P., and Bartel, B. (2002). Charac- terization of a family of IAA-amino acid conjugate hydrolases from Arabidopsis. J. Biol. Chem. 277, 20446–20452. doi: 10.1074/jbc.M111955200
Liu, H., Xie, W. F., Zhang, L., Valpuesta, V., Ye, Z. W., and Gao, Q. H., et al. (2014). Auxin biosynthesis by the YUCCA6 flavin monooxygenase gene in woodland strawberry. J. Integr. Plant Biol. 56, 350–363. doi: 10.1111/jipb.12150
Ljung, K. (2013). Auxin metabolism and homeostasis during plant development. Development 140, 943–950. doi: 10.1242/dev.086363
Mano, Y., Nemoto, K., Suzuki, M., Seki, H., Fujii, I., and Muranaka, T. (2010). The AMI1 gene family: indole-3-acetamide hydrolase functions in auxin biosynthesis in plants. J. Exp. Bot. 81, 25–32. doi: 10.1093/jxb/erp292
McAtee, P., Karim, S., Schaffer, R., and David, K. (2013). A dynamic interplay between phytohormones is required for fruit development, maturation, and ripening. Front. Plant Sci. 4:79. doi: 10.3389/fpls.2013.00079
Nasopoulou, C., Pohjanen, J., Koskimäki, J. J., Zabetakis, I., and Pirttilä, A. M. (2014). Localization of strawberry (Fragaria x ananassa) and Methylobacterium extorquens genes of strawberry flavor biosynthesis in strawberry tissue by in situ hybridization. J. Plant Physiol. 171, 1099–1105. doi: 10.1016/j.jplph.2014.03.018
Nitsch, J. P. (1950). Growth and morphogenesis of the strawberry as related to auxin. Am. J. Bot. 37, 211–215. doi: 10.2307/2437903
Nitsch, J. P. (1955). Free auxins and free tryptophane in the strawberry. Plant Physiol. 30, 33–39. doi: 10.1104/pp.30.1.33
Oh, E., Zhu, J. Y., Bai, M. Y., Arenhart, R. A., Sun, Y., and Wang, Z.Y. (2014). Cell elongation is regulated through a central circuit of interacting transcription factors in the Arabidopsis hypocotyl. Elife 3:e03031. doi: 10.7554/eLife.03031
Osorio, S., Bombarely, A., Giavalisco, P., Usadel, B., Stephens, C., Aragüez, I., et al. (2011). Demethylation of oligogalacturonides by FaPE1 in the fruits of the wild strawberry fragaria vesca triggers metabolic and transcriptional changes associated with defence and development of the fruit. J. Exp. Bot. 62, 2855–2873. doi: 10.1093/jxb/erq465
Ottenschlager, I., Wolff, P., Wolverton, C., Bhalerao, R. P., Sandberg, G. R., Ishikawa, H., et al. (2003). Gravity-regulated differential auxin transport from columella to lateral root cap cells. Proc. Natl. Acad. Sci. U.S.A. 100, 2987–2991. doi: 10.1073/pnas.0437936100
Pacheco-Villalobos, D., Díaz-Moreno, S. M., van der Schuren, A., Tamaki, T., Kang, Y. H., Gujas, B., et al. (2016). The effects of high steady state auxin levels on root cell elongation in brachypodium. Plant Cell 28, 1009–1024. doi: 10.1105/tpc.15.01057
Parry, G., Calderón-Villalobos, L. I., Prigge, M., Peret, B., Dharmasiri, S., Itoh, H., et al. (2009). Complex regulation of the TIR1/AFB family of auxin receptors. Proc. Natl. Acad. Sci. U.S.A. 106, 22540–22545. doi: 10.1073/pnas.0911967106
Pattison, R. J., and Catalá, C. (2011). Evaluating auxin distribution in tomato (solanum lycopersicum) through an analysis of the pin and aux/lax gene families. Plant J. 70, 585–598. doi: 10.1111/j.1365-313X.2011.04895.x
Pattison, R. J., Csukasi, F., and Catalá, C. (2014). Mechanisms regulating auxin action during fruit development. Physiol. Plant. 151, 62–72. doi: 10.1111/ppl.12142
Pedraza, R. O., Ramírez-Mata, A., Xiqui, M. L., and Baca, B. E. (2004). Aromatic amino acid aminotransferase activity and indole-3-acetic acid production by associative nitrogen-fixing bacteria. FEMS Microbiol. Lett. 233, 15–21. doi: 10.1016/j.femsle.2004.01.047
Perkins-Veazie, P. (1995). Growth and ripening of strawberry fruit. Hortic. Rev. 17, 267–297. doi: 10.1002/9780470650585.ch8
Piya, S., Shrestha, S. K., Binder, B., Stewart, C. N. Jr., Hewezi, T. (2014) Protein-protein interaction and gene co-expression maps of ARFs and Aux/IAAs in Arabidopsis. Front. Plant Sci. 5:744. doi: 10.3389/fpls.2014.00744
Porco, S., Pencik, A., Rashed, A., Voß, U., Casanova-Sáez, R., Anthony Bishopp, A., Golebiowska, A., et al. (2016). Dioxygenase-encoding AtDAO1 gene controls IAA oxidation and homeostasis in Arabidopsis. Proc. Natl. Acad. Sci. U.S.A. 113, 11016–11021. doi: 10.1073/pnas.1604375113
Salehin, M., Bagchi, R., and Estelle, M. (2015) SCFTIR1/AFB-based auxin perception: mechanism and role in plant growth and development. Plant Cell 27, 9–19. doi: 10.1105/tpc.114.133744
Salvatierra, A., Pimentel, P., Moya-Leon, M. A., Caligari, P., and Herrera, R. (2010). Comparison of transcriptional profiles of flavonoid genes and anthocyanin contents during fruit development of two botanical forms of Fragaria chiloensis ssp. chiloensis. Phytochemistry 71, 1839–1847. doi: 10.1016/j.phytochem.2010.08.005
Sánchez-Sevilla, J., Cruz-Rus, E., Valpuesta, V., Botella, M., and Amaya, I. (2014). Deciphering gamma-decalactone biosynthesis in strawberry fruit using a combination of genetic mapping, RNA-Seq and eQTL analyses. BMC Genomics 15:218. doi: 10.1186/1471-2164-15-218
Sauer, M., and Kleine-Vehn, J. (2011). Auxin binding protein1: the outsider. Plant Cell 23, 2033–2043. doi: 10.1105/tpc.111.087064
Shulaev, V., Sargent, D. J., Crowhurst, R. N., Mockler, T. C., Folkerts, O., Delcher A. L. et al. (2011). The genome of woodland strawberry (Fragaria vesca). Nat. Genet. 43, 109–116. doi: 10.1038/ng.740
Staswick, P. E., Serban, B., Rowe, M., Tiryaki, I., Maldonado, M. T, Maldonado, M. C., et al. (2005). Characterization of an Arabidopsis enzyme family that conjugates amino acids to indole-3-acetic acid. Plant Cell 17, 616–627. doi: 10.1105/tpc.104.026690
Stepanova, A. N., Yun, J., Robles, L. M., Novak, O., He, W., Guo, H., et al. (2011). The Arabidopsis YUCCA1 avin monooxygenase functions in the indole-3-pyruvic acid branch of auxin biosynthesis. Plant Cell 23, 3961–3973. doi: 10.1105/tpc.111.088047
Stepanova, A. N., Robertson-Hoyt, J., Yun, J., Benavente, L. M., Xie, D. Y., Dolezal, K., et al. (2008). TAA1- mediated auxin biosynthesis is essential for hormone crosstalk and plant development. Cell 133, 177–191. doi: 10.1016/j.cell.2008.01.047
Symons, G. M., Chua, Y. J., Ross, J. J., Quittenden, L. J., Davies, N. W., and Reid, J. B. (2012). Hormonal changes during non-climacteric ripening in strawberry. J. Exp. Bot. 63, 4741–4750. doi: 10.1093/jxb/errs311437
Tennessen, J. A., Govindarajulu, R., Ashman, T. L., and Liston, A. (2014). Evolutionary origins and dynamics of octoploid strawberry subgenomes revealed by dense targeted capture linkage maps. Genome Biol. Evol. 6, 3295–3313. doi: 10.1093/gbe/evu261
Tiwari, S. B., Hagen, G., and Guilfoyle, T. (2003). The roles of auxin response factor domains in auxin-responsive transcription. Plant Cell 15, 533–543. doi: 10.1105/tpc.008417
Trainotti, L., Tadiello, A., and Casadoro, G. (2007). The involvement of auxin in the ripening of climacteric fruits comes of age: the hormone plays a role of its own and has an intense interplay with ethylene in ripening peaches. J. Exp. Bot. 58, 3299–3308. doi: 10.1093/jxb/erm178
Trapnell, C., Roberts, A., Goff, L., Pertea, G., Kim, D., Kelley, D., et al. (2012). Differential gene and transcript expression analysis of RNA-seq experiments with TopHat and Cufflinks. Nat. Protoc. 7, 562–578. doi: 10.1038/nprot.2012.016
Ulmasov, T., Hagen, G., and Guilfoyle, T. J. (1999). Activation and repression of transcription by auxin-response factors. Proc. Natl. Acad. Sci. U.S.A. 96, 5844–5849. doi: 10.1073/pnas.96.10.5844
Ulmasov, T., Murfett, J., Hagen, G., and Guilfoyle, T. J. (1997). Aux/lAA Proteins repress expression of reporter genes containing natural and highly active synthetic auxin response elements. Plant Cell 9, 1963–1971. doi: 10.1105/tpc.9.11.1963
Vallarino, J. G., Osorio, S., Bombarely, A., Casañal, A., Cruz-Rus, E., Sánchez-Sevilla, J. F. et al. (2015). Central role of a FaGAMYB in the transition of the strawberry receptacle from development to ripening. New Physiol. 208, 482–496. doi: 10.1111/nph.13463
Vanneste, S., and Friml, J. (2009). Auxin: a trigger for change in plant development. Cell 136, 1005–1016. doi: 10.1016/j.cell.2009.03.001
Velasquez, S. M., Barbez, E., Kleine-Vehn, J., and Estevez, J. M. (2016). Auxin and cellular elongation. Plant Physiol. 170, 1206–1215. doi: 10.1104/pp.15.01863
Weijers, D., and Wagner, D. (2016). Transcriptional Responses to the Auxin Hormone. Annu. Rev. Plant Biol. 67, 539–574. doi: 10.1146/annurev-arplant-043015-112122.
Xie, R., Pang, S., Ma, Y., Deng, L., He, S., Yi, S., et al. (2015). The ARF, AUX/IAA and GH3 gene families in citrus: genome-wide identification and expression analysis during fruitlet drop from abscission zone A. Mol. Genet. Genomics 290, 2089–2105. doi: 10.1007/s00438-015-1063-1
Yang, J. H., and Wang, H. (2016). Molecular mechanisms for vascular development and secondary cell wall formation. Front. Plant Sci. 7:356. doi: 10.3389/fpls.2016.00356
Zhang, Y., Paschold, A., Marcon, C., Liu, S., Tai, H., Nestler, J., et al. (2014). The Aux/IAA gene rum1 involved in seminal and lateral root formation controls vascular patterning in maize (Zea mays L.) primary roots. J. Exp. Bot. 65, 4919–4930. doi: 10.1093/jxb/eru249
Keywords: auxin, fruit, strawberry, transcriptome regulation, ripening
Citation: Estrada-Johnson E, Csukasi F, Pizarro CM, Vallarino JG, Kiryakova Y, Vioque A, Brumos J, Medina-Escobar N, Botella MA, Alonso JM, Fernie AR, Sánchez-Sevilla JF, Osorio S and Valpuesta V (2017) Transcriptomic Analysis in Strawberry Fruits Reveals Active Auxin Biosynthesis and Signaling in the Ripe Receptacle. Front. Plant Sci. 8:889. doi: 10.3389/fpls.2017.00889
Received: 17 February 2017; Accepted: 12 May 2017;
Published: 29 May 2017.
Edited by:
Shucai Wang, Northeast Normal University, ChinaReviewed by:
Ke Duan, Shanghai Academy of Agricultural Sciences, ChinaCopyright © 2017 Estrada-Johnson, Csukasi, Pizarro, Vallarino, Kiryakova, Vioque, Brumos, Medina-Escobar, Botella, Alonso, Fernie, Sánchez-Sevilla, Osorio and Valpuesta. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Victoriano Valpuesta, dmFscHVlc3RhQHVtYS5lcw== Sonia Osorio, c29zb3Jpb0B1bWEuZXM=
†These authors have contributed equally to this work.
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.