 Martha V. T. Cely1
Martha V. T. Cely1 Marco A. Siviero2Janaina Emiliano1
Marco A. Siviero2Janaina Emiliano1 Flávia R. Spago1Vanessa F. Freitas1
Flávia R. Spago1Vanessa F. Freitas1 André R. Barazetti1
André R. Barazetti1 Erika T. Goya1
Erika T. Goya1 Gustavo de Souza Lamberti1Igor M. O. dos Santos1
Gustavo de Souza Lamberti1Igor M. O. dos Santos1 Admilton G. De Oliveira1
Admilton G. De Oliveira1 Galdino Andrade1*
Galdino Andrade1*- 1Laboratório de Ecologia Microbiana, Departamento de Microbiologia, Universidade Estadual de Londrina, Londrina, Brazil
- 2Grupo Arboris-Manejo Florestal, Dom Eliseu, Brazil
Schizolobium parahyba var. amazonicum (Huber ex Ducke) occurs naturally in the Brazilian Amazon. Currently, it is being planted extensively because of its fast growth and excellent use in forestry. Consequently, there is great interest in new strategies to increase wood production. The interaction between soil microorganisms and plants, specifically in the roots, provides essential nutrients for plant growth. These interactions can have growth-promoting effects. In this way, this study assessed the effect of the inoculation with arbuscular mycorrhizal fungi (AMF) and plant growth-promoting rhizobacteria (PGPR) on growth of S. parahyba var. amazonicum under field conditions. We used two native species of arbuscular mycorrhizal fungi, Claroideoglomus etunicatum (Ce), and Acaulospora sp. (Ac); two native strains of Rhizobium sp. (Rh1 and Rh2); and a non-native strain of Burkholderia sp. Different combinations of microorganisms were supplemented with chemical fertilizers (doses D1 and D2) in two planting methods, seed sowing and seedling planting. In seed sowing, the results showed that treatments with Ce/Rh1/Fertilizer D2 and Ac/No PGPR/Fertilizer D2 increased wood yield. In seedling planting, two combinations (Ac/Rh2/Fertilizer D1 and Ac/Rh1/Fertilizer D1) were more effective in increasing seedling growth. In these experiments, inoculation with AMF and PGPR increased wood yield by about 20% compared to the application of fertilizer alone.
Introduction
The negative impacts of agro-industrial development and wood exploitation in native forest areas have encouraged the development of projects focused on reforestation with homogeneous stands or intercropped species of rapid growth and high commercial value. These strategies are directed at degraded areas with the objective of forest restoration or wood production. The family Leguminosae is one of the most representative in terms of number and frequency of plant species in the Amazon region of Brazil (Silva et al., 1988). Some tree species (nodulating and non-nodulating) of this family are used or have high potential uses for timber production (Sprent and Parsons, 2000) and land restoration (Faria et al., 2010).
Schizolobium parahyba var. amazonicum (Huber ex Ducke), belonging to the family Leguminosae and subfamily Caesalpinioideae, is a non-nodulating species native to the Amazon. It is considered an ecologically and economically important species due to its significant wood potential; its commercial potential has been exploited since the 1970s. Today, it is the native species most planted in the Brazilian states of Amazonia, Pará, Maranhão, and Rondonia, covering 87,901 ha (ABRAF, 2012). Due to its fast growth, S. parahyba var. amazonicum can reach an annual wood yield of 30 m3 ha−1 year−1 with 6 years of age (Carvalho, 2007). Moreover, it is considered an important species for carbon sequestration because it produces high levels of biomass in a short period of time (Siviero et al., 2008). The quality of its wood is suitable for furniture and plywood production.
The choice of plant species that are used for restoration and wood production in degraded lands represents a great challenge, because these species need to be able to survive under conditions of low soil fertility. These restrictive factors for plant growth can be attenuated by the action of efficient soil microorganisms such as plant growth-promoting rhizobacteria (PGPR) and arbuscular mycorrhizal fungi (AMF; Chaer et al., 2011). The microbial community in the soil plays an important role in the sustainability of plant communities (Andrade, 2004). The interaction between microorganisms and plants, specifically in the roots, provides for important nutritional requirements of plants and also the microorganisms associated with them. Thus, as the roots directly affect the surrounding microbial populations, the microorganisms present in the rhizosphere can also influence plant growth (Giri et al., 2005).
PGPR are microorganisms that colonize the rhizosphere and promote plant growth. Among them, the N-fixing bacteria (NFB) such as Rhizobium species can establish symbiosis with leguminous plant species, resulting in a beneficial interaction for plant growth. Some diazotrophic bacteria can help plant nutrition through biological fixation of N2 or production of phytohormones (Vessey, 2003). AMF, associated with plant roots, increase the uptake of soil inorganic nutrients, mainly P (Neumann and George, 2010). In addition, other benefits related to AMF are the stabilization of soil aggregates (Rillig, 2004), increasing resistance to water stress (Garg and Chandel, 2010) and protection against pathogens (Jung et al., 2012). In the mycorrhizosphere, the soil surrounding the roots and fungal hyphae (Artursson et al., 2006), AMF can interact with PGPR bacterial species, as well as with endophytic bacteria. Some belong to the genus Burkholderia (Bianciotto and Bonfante, 2002). These interactions can provide potential benefits for plant development. The inoculation of compatible combinations of PGPR and AMF in forest and agricultural systems may result in a significant increase in plant growth (Biró et al., 2000; Nadeem et al., 2014; Hashem et al., 2016). Many studies (Marques et al., 2001; Valdenegro et al., 2001; Patreze and Cordeiro, 2004) have demonstrated the synergistic effect of the inoculation of Rhizobium and AMF in promoting nodulated legume tree species. However, little research has been carried out on this subject with legume trees of the subfamily Caesalpinoideae (Bryan et al., 1996) with the formation of nodules observed in a few cases (Sprent, 1983).
S. parahyba var. amazonicum is a non-nodulating legume, and Rhizobium bacteria may promote plant growth in this species in two ways. Some authors suggest that non-nodulating species of the family Leguminoseae can profit from N fixed by root-associated bacteria (rhizosphere bacteria or endophytes) like nodulating species (Bryan et al., 1996; Van Sambeek et al., 2007). On the other hand, it can be assumed that the Rhizobium act as plant growth-promoting bacteria in the rhizosphere and release phytohormones (Mehboob et al., 2012).
The use of growth-promoting microorganisms in S. parahyba var. amazonicum was assessed by Siviero et al. (2008), who showed that this species displays a positive response to inoculation with AMF in combination with N-fixing bacteria isolated from another plant species (exogenous, i.e., non-native bacteria). The authors observed differences between planting methods (seeds or seedlings) in inoculated plants. In the planting method with seeds, only AMF (Glomus intrarradices) inoculation increased biomass and wood production. In the planting method with seedlings, the dual inoculation of AMF (Glomus clarum) and PGPR (LEM6 or Rhi1 Rhizobium strains) was more effective. In this work, the authors suggested that the selection of native microorganisms is very important to obtain the best results in the field.
Our hypothesis was that the inoculation with indigenous microorganisms is more effective in promoting plant growth of S. parahyba var. amazonicum, and that the presence of inoculum would help plant roots to be more effective in using the chemical fertilizer applied. Therefore, this study assessed the effect on wood production, comparing inoculation with two indigenous AMF (Claroideoglomus etunicatum and Acaulospora sp.) isolated from S. parahyba var. amazonicum roots in interaction with three bacterial strains (two indigenous Rhizobium spp. and one exogenous Burkholderia sp.). The inoculation with different combinations of microorganisms and the addition of chemical fertilizer was investigated using a completely randomized block experiment. The effect of these factors on S. parahyba var. amazonicum growth was determined in situ over 2 years.
Materials and Methods
Experimental Field
The experiments were conducted in the municipality of Dom Eliseu – Pará State (Brazil) [4°17′36″ S and 47°33′15″ W]. Its climate is classified as humid mesothermic, with an average annual temperature of 25°C and annual rainfall of 2500 mm. The region in the wet season shows extensive rain from January to June, and a relative humidity of around 85%. The vegetation is a terra firme type with dense forest (da Silva et al., 2011). However, continuous deforestation had destroyed the original vegetation, leading to the emergence of large areas of savannas and secondary forest (SEICOM, 2012).
The soil in the experimental area was a Xanthic Ferralsol according to the FAO classification (FAO, 1994). Prior to experimentation, the soil was chemically analyzed using a composite sample collected from a depth of 0–20 cm and the physical–chemical analysis showed the following results: pH (CaCl2) 4.8, H+Al 2.9 cmolc dm−3, Al+3 0.2 cmolc dm−3; Ca+2 3.3 cmolc dm−3, Mg+2 1.0 cmolc dm−3, K+ 0.24 cmolc dm−3; P (Mehlich I) 10.0 cmolc dm−3, C 19.0 g dm−3; S- 4.2 cmolc dm−3, Na+ 4.0 cmolc dm−3, B 0.3 cmolc dm−3, Fe2+ 99.0 cmolc dm−3, Mn 7.3 cmolc dm−3, Cu 0.2 cmolc dm−3, and Zn 3.0 cmolc dm−3. Samples of soil collected in the experimental area showed a low number of AMF spores (3 spores/g of soil) when compared with other soils.
Experimental Design
Two experiments were conducted, each using different planting methods, seeds, and seedlings. The inoculation of each planting method occurred by using different combinations of two species of AMF (C. etunicatum and Acaulospora sp.), and three PGPR strains (Rhizobium sp1, Rhizobium sp2, and Burkholderia sp.). Additionally, two doses of chemical fertilizer (NPK formulation 10:20:20-N: Urea; P: P2O5; K: K2O) were applied. Dose 1 (D1) was 75 g of fertilizer per plant and dose 2 (D2) was 150 g of fertilizer per plant. The resulting 36 treatments of combinations of these three factors (AMF, PGPR, and Fertilizer) are described in Table 1. The treatments were arranged as a completely randomized block design with three repetitions. In the block, each treatment was represented by a row with 10 plants. The spacing was 3 × 2 m between plants and 6 m between blocks. The buffer area in the experiment was composed of three rows with non-inoculated and non-fertilized plants.

Table 1. Description of treatments.
Plant Inoculation
The seeds of S.parahyba var. amazonicum were collected from native forest in Pará state, where the tree occurs naturally. Before sowing, the seeds were scarified mechanically at one end. In the seed system, two seeds were sown in each pit and in the seedling system, one 30 day-old seedling (cultivated in a nursery in plastic bags of 1000 mL with non-sterile soil) was planted before being taken out of the plastic bag.
Microorganism Strains and Growth Conditions
Spores of AMF (C. etunicatum and Acaulospora sp.) were isolated from the rhizosphere of S. parahyba var. amazonicum in the Amazon Forest in Dom Eliseu, Pará, and propagated in pots with Urochloa decumbens as plant host. Ten grams of inoculum extracted from pots containing 50 spores/g of soil, colonized roots, and mycelia were added before seed sowing or seedling planting in the field.
The bacterial strains used as inoculum were two native ones [Rhizobium sp1 (Rh1) and Rhizobium sp2 (Rh2)] isolated from roots of S. parahyba var. amazonicum in the Amazon Forest in Dom Eliseu, Pará. In addition, an exogenous strain of Burkholderia sp. was used (Raimam et al., 2007). The Rhizobium strains were grown in Petri dishes with TY medium (Beringer, 1974) and the Burkholderia sp. strain in Nfb medium (Döbereiner and Day, 1976). For inoculation in the field, the bacteria were re-suspended in sterile saline (0.85% NaCl) plus carboxymethyl cellulose (0.1%) and adjusted by visual comparison with a McFarland standard scale to obtain a final cell concentration of ~109 cells mL−1. Before sowing, the seeds were inoculated by immersion in a bacterial suspension. Seedlings were inoculated with 10 mL of bacterial suspension around the plant.
Data Collection, Biomass, and Wood Yield Determination
Plant growth was determined by shoot diameter (at soil surface), shoot total height (TH), and biomass. Data were collected at 180, 280, 480, and 720 days after planting. At 720 days, we evaluated the diameter at breast height (DBH), TH, and height up to the first leaf (HFL). Biomass (BIO) was determined as described below and was determined for each plant based on the volume of the stem (Brow, 1997) and multiplied by the correction factor for S. parahyba var. amazonicum as suggested by Colpini et al. (2009).
Wood yield was determined by BIO-value multiplied by the wood specific density of S. parahyba var. amazonicum and the number of plants per hectare.
Statistical Analysis
Statistical analysis was performed using Statistica 7.0 (Statsoft Inc Statistica, 2004). Data were tested for normality using the Shapiro-Wilk test. ANOVA on the data sets (DBH, TH, and biomass) was carried out to determine the interactions of the factors AMF, PGPR, and Fertilizer. Differences between treatments were determined by Tukey's means test (HSD) at p ≤ 0.05 significance level. A principal component analyses (PCA) was carried out with all data. Time was considered a cofactor, and the treatments were grouped according to AMF inoculation to facilitate the interpretation.
Results
Seed Experiment
In the seeds planting experiment, AMF showed significant effects on DBH and TH at 280 days and BIO at 480 days. PGPR increased DBH and TH at 480 days. Fertilizer addition showed a significant effect on plant growth at all sampling times. The interaction between PGPR and fertilizer effect resulted in increased DBH (Table 2). BIO was significantly enhanced by both AMF C. etunicatum and Acaulospora sp. at 480 days (Figure 1A). Rhizobium strain Rh1 increased BIO by around 30% when compared with Burkholderia (Figure 1B). Both doses of fertilizer increased plant growth during the whole experiment (Figure 1C).
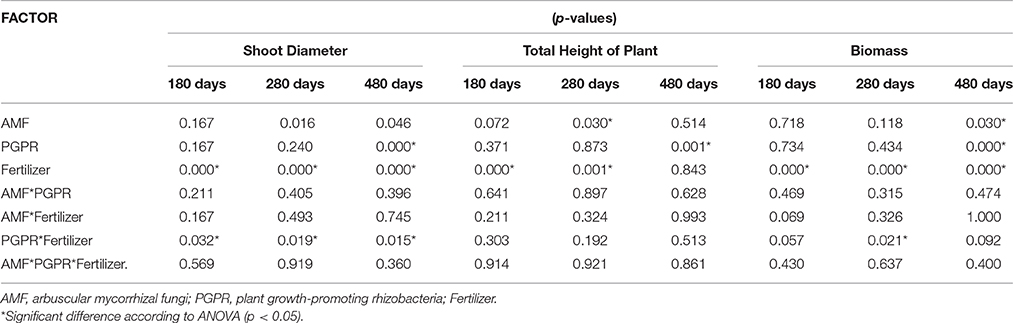
Table 2. Analysis of variance of plant growth of S. parahyba var. amazoicum at 180, 280, and 480 days after sowing seeds.
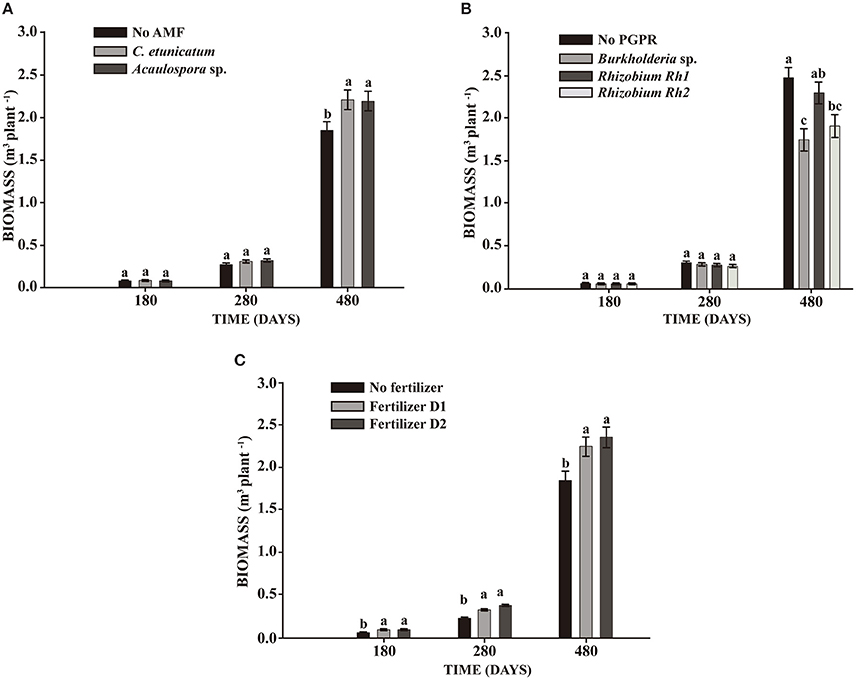
Figure 1. Effect on biomass production of S. parahyba var. amazonicum after 180, 280, and 480 days of sowing seeds. (A) Arbuscular mycorrhiza fungi (AMF) C. etunicatum and Acaulospora sp.; (B) PGPR strains Burkholderia sp., Rhizobium Rh1, and Rhizobium Rh2; (C) two doses of fertilizer (D1: 75 g plant−1 and D2: 150 g plant−1). Bars sharing the same letter are not statistically significantly different according to Tukey test (p < 0.05).
After 720 days, there was a significant effect on DBH and BIO in AMF plants from sown seeds and in fertilized plants. Bacterial inoculation, especially with Rhizobium Rh1, increased DBH, TH, and BIO. The interaction between AMF and bacteria also increased DBH, TH, and BIO. The interaction between Acaulospora sp. and Rhizobium Rh1 resulted in greater diameter and height of S. parahyba in fertilized and non-fertilized plants. On the other hand, the same treatment increased BIO but only in non-fertilized plants (Table 3). The interaction between Rhizobium Rh1 and C. etunicatum also increased DBH and BIO in non-fertilized and D2 plants, and TH was increased only in D2 plants in the interaction of C. etunicatum and Rhizobium Rh1 (Table 3).
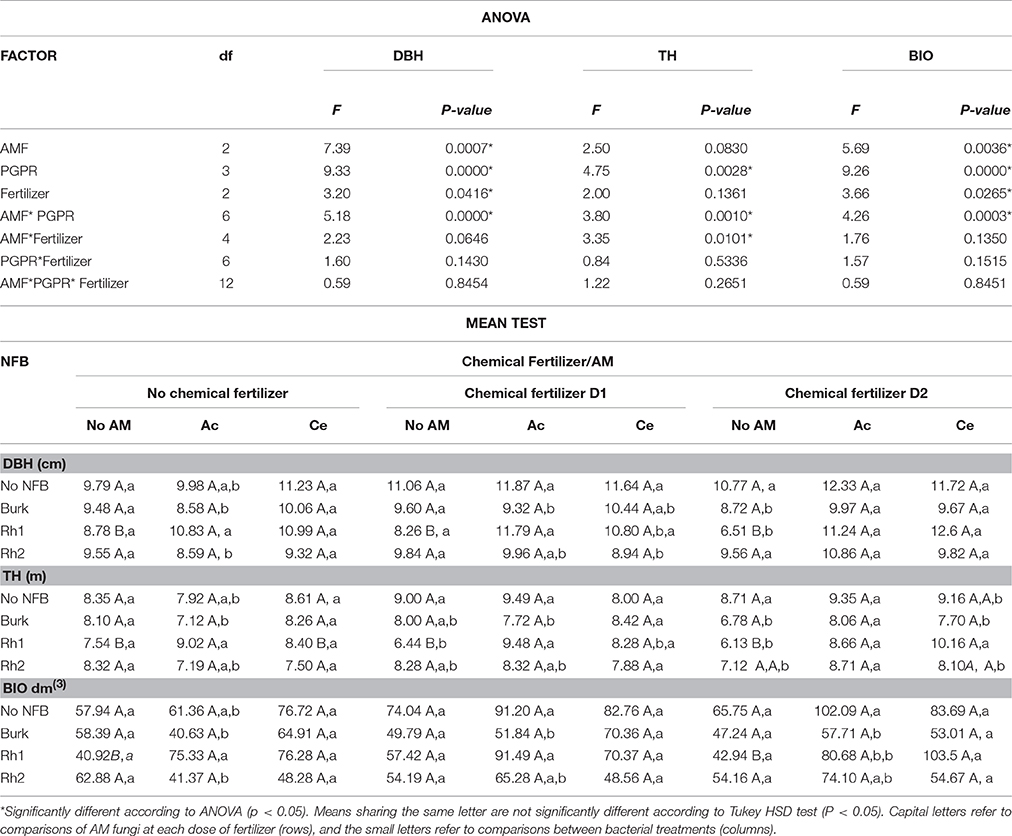
Table 3. Effect of arbuscular mycorrhizal fungi (AMF) Acaulospora sp. (Ac), Claroideoglomus etunicatum (Ce), and PGPR strains Burkholderia sp. (Burk), Rhizobium Rh1 and Rh2 on diameter at breast height (DBH), total height (TH), and biomass (BIO) after 2 years of sowing seeds.
PCA of No AMF plants (Figure 2A) revealed that principal component 1 (PC1) and principal component 2 (PC2) accounted for 60.2 and 21.3% of the data variation, respectively. PC1 comprised treatments with Rhizobium Rh1 and fertilizer (D1 and D2), and it showed a strong relation with TH, when compared with plants that were only fertilized. In PC2, D2 showed more influence on HFL and BIO. For plants inoculated with C. etunicatum (Figure 2B), PC1 accounted for 65.3% of data variation and PC2 for 13%. PC1 allowed comparison of two treatments: Rhizobium Rh1/No Fertilizer and No PGPR/Fertilizer D2. Both treatments showed more influence on HFL and BIO. PC2 showed that the combination of Rhizobium Rh1 and fertilizer D2 had a significant influence on BIO. With regard to inoculation with Acaulospora sp., PC1 accounted for 63.8% of data variation and PC2 for 20.4 % (Figure 2C). The treatments that showed the highest impact on TH and DBH were fertilizer D1 plus Rhizobium Rh1, and the application of D1 or D2 as well. This analysis showed that the inoculation with microorganisms was compatible with the application of fertilizer and that it had a significant effect on plant growth. Therefore, the more effective treatments were: No AMF/Rhizobium Rh1/Fertilizer D2; C. etunicatum/Rhizobium Rh1/No fertilizer; C. etunicatum/No PGPR/Fertilizer D2; Acaulospora sp./No PGPR/Fertilizer D1 and Acaulospora sp./Rhizobium Rh1/Fertilizer D2 compared to control plants.
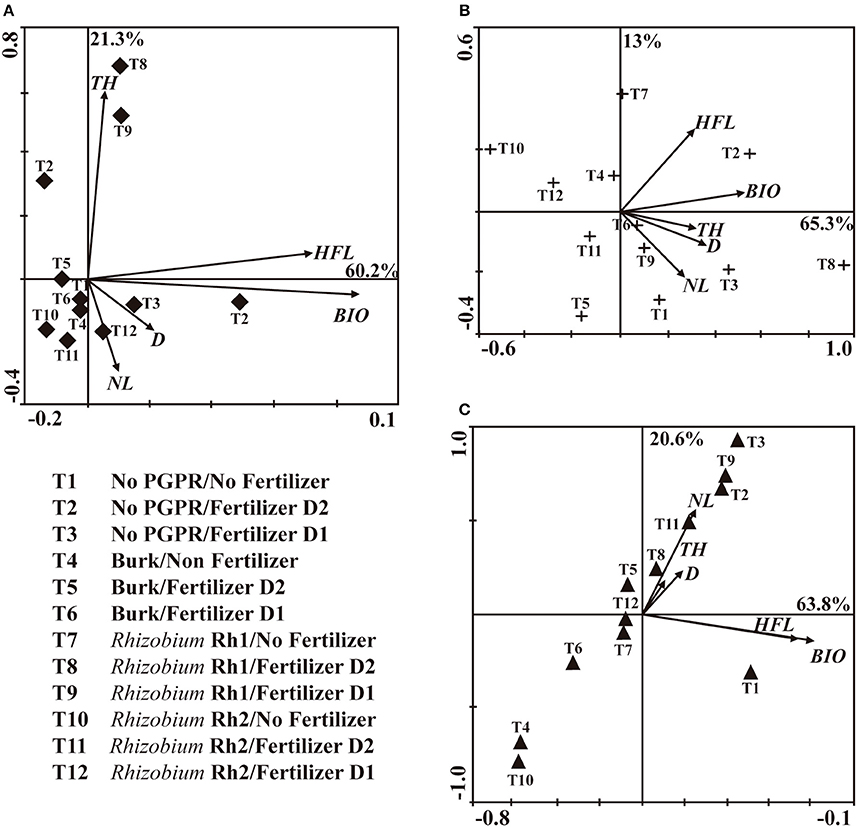
Figure 2. Principal component analysis (PCA) among AM fungi and PGPR bacteria (Burkholderia sp, Rhizobium (Rh1 and Rh2) on the shoot diameter (D), total height (TH), height at the first leave (HFL), number of leaves (NL) and biomass (BIO) two years after seeds sowing. (A) No AMF; (B) Claroideoglomus etunicatum; (C) Acaulospora sp.
In terms of wood yield in plants from sown seeds, only Ac/No PGPR/No fertilizer was more effective (up to 20% increase in wood production) compared control plants (No AMF, No PGPR, No Fertilizer). In addition, four treatments increased wood yield by 30% (No AM/Rh2/No fertilizer; Ce/Burkholderia (Burk)/No fertilizer; Ac/Rh2/Fertilizer D1; and No AMF/No PGPR/Fertilizer D2), and in seven treatments, there was a more than 40% increase (Ce/Burk/Fertilizer D1; Ce/Rh1/Fertilizer D1; Ce/Rh1/No fertilizer; No AM/No PGPR/Fertilizer D1; Ac/Rh2/Fertilizer D2; Ac/Rh1/No fertilizer; and Ce/No PGPR/No fertilizer). Four treatments increased wood yield by more than 50% (No AM/Rh1/Fertilizer D2; Ac/Rh1/Fertilizer D2; Ce/mon-PGPR/Fertilizer D1; Ce/No PGPR/Fertilizer D2), and three treatments by more than 60% (Ac/No PGPR/Fertilizer D1); Ac/Rh1/Fertilizer D1; and No AMF/Rh1/Fertilizer D1). Two treatments increased wood yield by 100% (Ac/No PGPR/Fertilizer D2 and Ce/Rh1/Fertilizer D2) (Figure 3), when compared to the control.
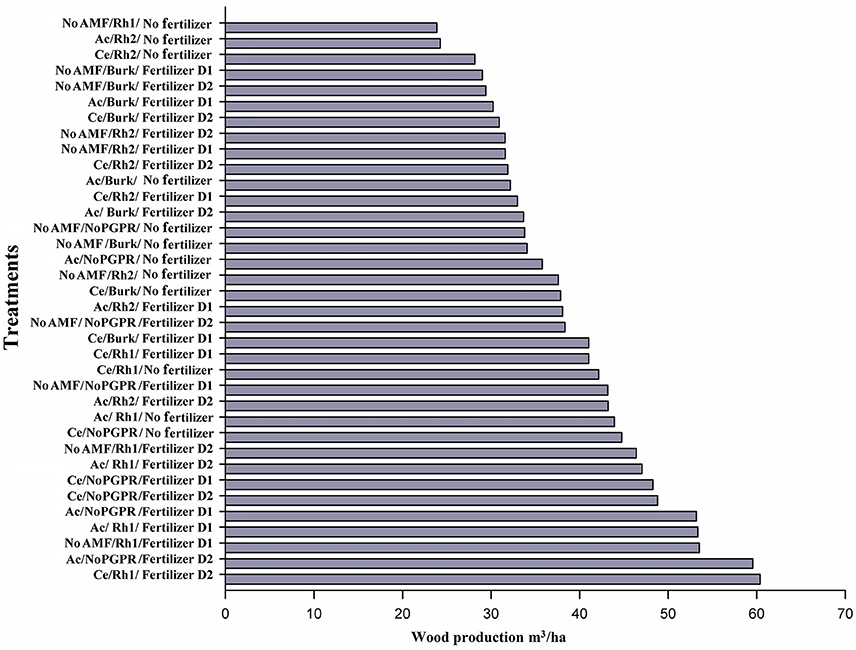
Figure 3. Wood production by different combinations of AMF [Acaulospora sp. (Ac) and Claroideoglomus etunicatum (Ce)], PGPR [Burkholderia sp. (Burk), Rhizobium Rh1, and Rhizobium Rh2], and chemical fertilizer [D1: 75 g plant−1, D2: 150 g plant−1] 2 years after sowing seeds.
The presence of AM fungi was more effective by up to 40% with C. etunicatum (Ce/non PGPR/No Fertilizer). On the other hand, when PGPR alone were used for inoculation, wood yield increased only by 30% in the presence of Rhizobium Rh2, and with other PGPR no effect was observed. When yield assessed, with increasing more than 50%, the addition of fertilizer (D1 or D2) was needed as well as AMF (Acaulospora or C. etunicatum).
Seedling Experiment
In the experiment with seedlings, mycorrhizal inoculation showed a significant effect on TH after 280 days. The inoculation of PGPR showed significant differences in all parameters assessed, increasing plant growth at 180, 280, and 480 days. The interaction between AMF and fertilizer showed significant differences in DBH during the whole experiment and in TH and BIO at 180 and 280 days (Table 4). No difference in BIO was observed between AMF and control plants (Figure 4A). On the other hand, inoculation of Rhizobium Rh1 increased BIO (Figure 4B). The addition of fertilizer showed a positive effect on BIO at all times evaluated (Figure 4C).
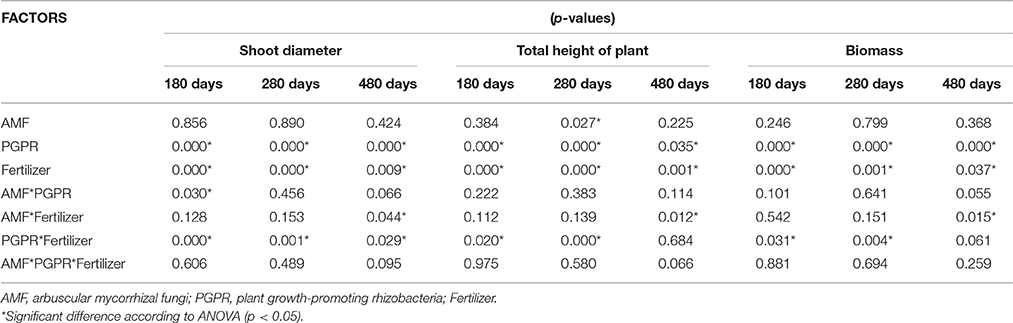
Table 4. Analysis of variance of plant growth of S. parahyba var. amazoicum at 180, 280, and 480 days after seedling planting.
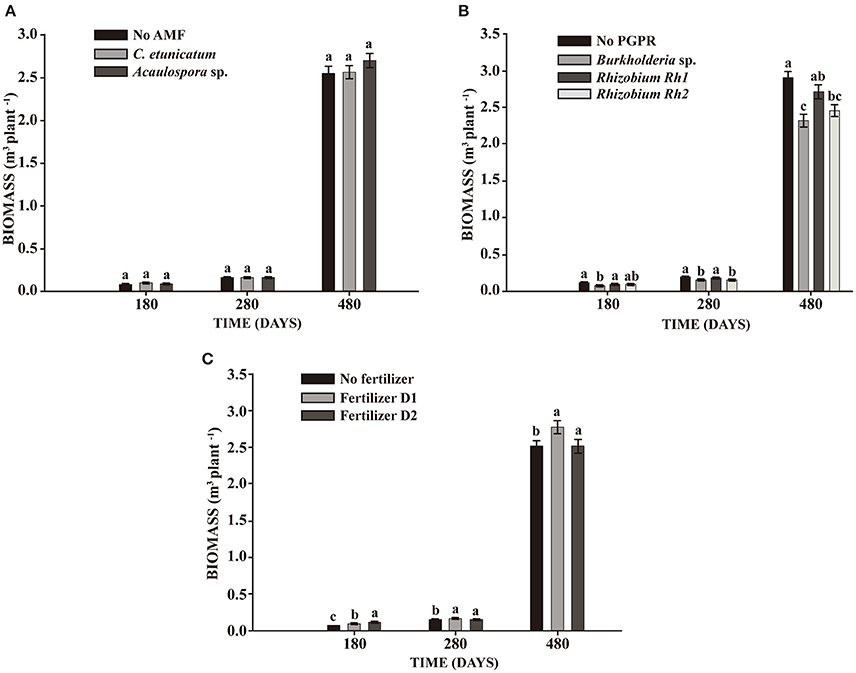
Figure 4. Effect on biomass production of S. parahyba var. amazonicum 180, 280, and 480 days after seedling planting. (A) Arbuscular mycorrhiza fungi (AMF) C. etunicatum and Acaulospora sp.; (B) PGPR strains Burkholderia sp., Rhizobium Rh1, and Rhizobium Rh2; (C) two doses of fertilizer (D1: 75 g plant−1 and D2: 150 g plant−1). Bars sharing the same letter are not statistically significantly different according to Tukey test (p < 0.05).
Two years (720 days) after planting and inoculated with AMF and PGPR and/or addition of fertilizer, trees in this experiment showed elevated DBH, TH, and BIO values. Plant growth increased in non-fertilized plant inoculated with Rhizobium Rh1 and C. etunicatum when compared with No AMF plants. D1-fertilized plants plus Acaulospora sp. and Burkholderia sp. or Rhizobium Rh1 increased TH. BIO was increased in non-fertilized plants when inoculated with C. etunicatum and Rhizobium Rh1, and in plants fertilized with D1 plus inoculated with Acaulospora sp. and Rhizobium Rh1 (Table 5).
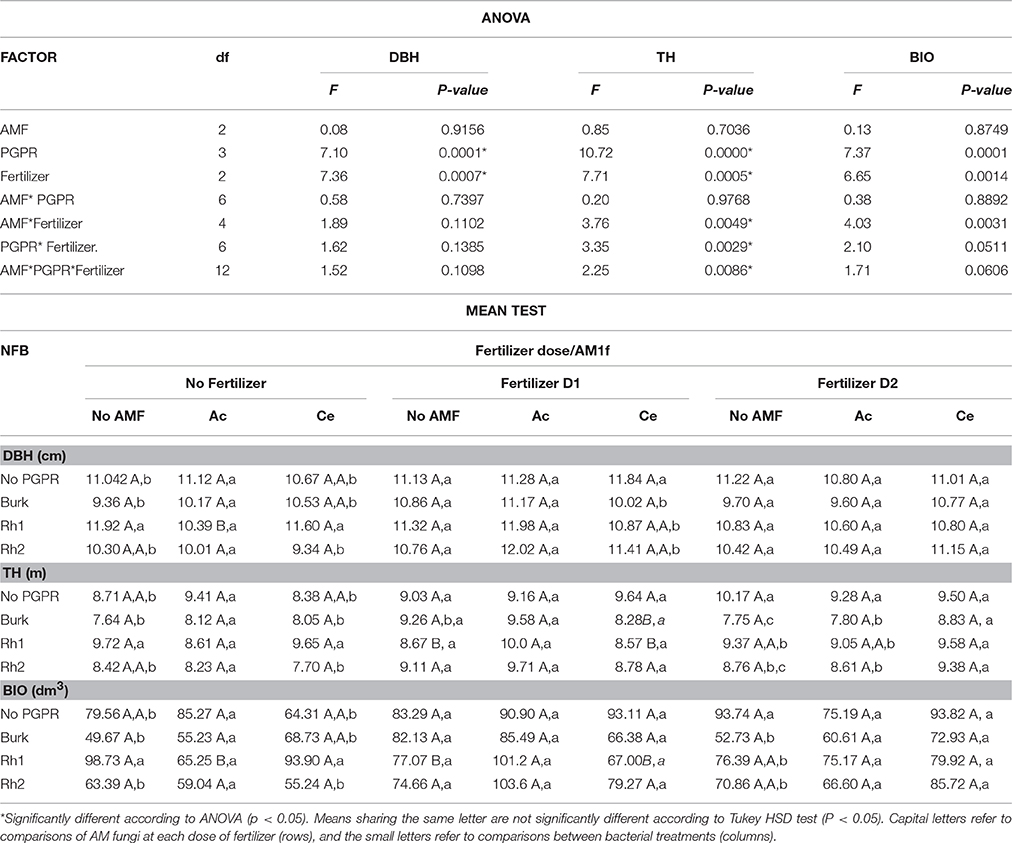
Table 5. Effect of AMF Acaulospora sp. (Ac), Claroideoglomus etunicatum (Ce), and PGPR strains Burkholderia sp. (Burk), Rhizobium Rh1 and Rh2 on diameter at breast height (DBH), total height (TH), and biomass (BIO) 2 years after seedling planting.
PCA of No AMF treatments allowed us to determine the first principal component (PC1) as accounting for 65.8% of data variation and the second principal component (PC2) for 17.2% of data variation. With PC1 comprising BIO, the treatments did not show significant influence on other variables. On the other hand, in the PC2, comprising No PGPR and Fertilizer D2 (T2), showed more relation with plant height. Inoculation with Rhizobium Rh1 without fertilizer (T7) also showed relation with TH (Figure 5A). For inoculation with C. etunicatum, PC1 explained 66.2 % of data variation and PC2 19.3 %. PC1 did not show significant relation with other variables. For PC2, the combination of Rhizobium Rh1 and No fertilizer (T7) showed more influence on D, TH, and BIO than did other treatments (Figure 5B). When Acaulospora sp. was used as inoculum, PC1 grouped 58.9% of data variation and PC2 19.2 %. The treatment with highest impact on plant growth was the combination between Rhizobium Rh1 and fertilizer D1 (T9). As for PC2, Rhizobium Rh2 plus fertilizer D1 (T12) showed a significant influence on BIO production (Figure 5C).
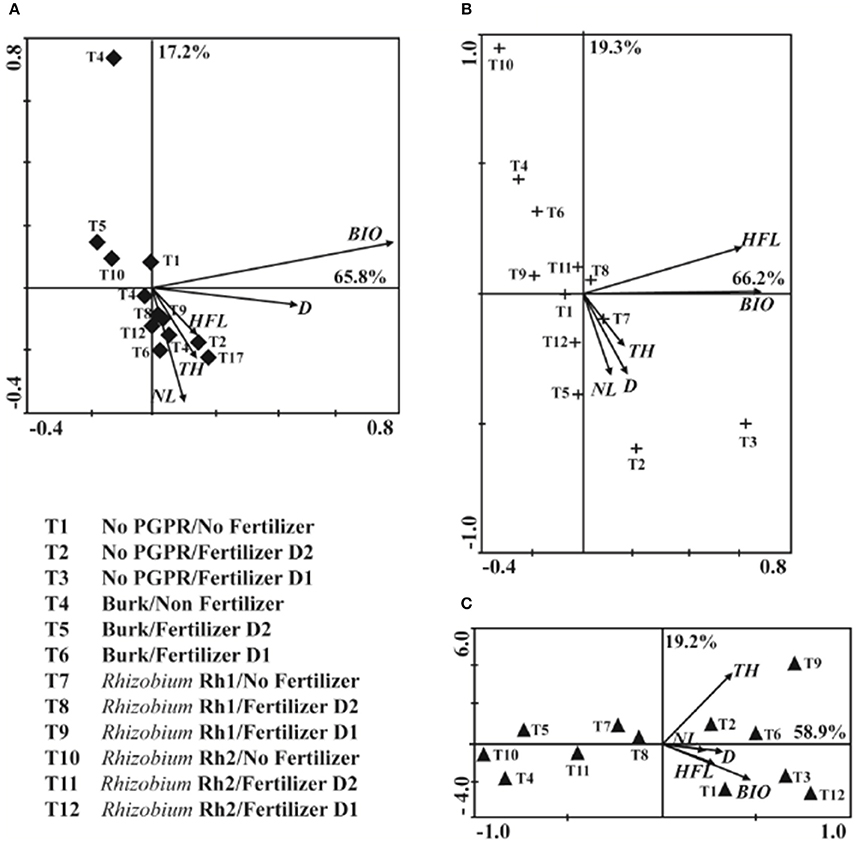
Figure 5. Principal component analysis (PCA) among AM fungi and PGPR bacteria (Burkholderia sp, Rhizobium Rh1 and Rh2) on the shoot diameter (D), total height (TH), height at the first leave (HFL), number of leaves (NL) and biomass (BIO) two years after seedling planting. (A) No AMF; (B) Claroideoglomus etunicatum; (C) Acaulospora sp.
Wood yield (720 days after seedling planting) increased with Acaulospora sp. in combination with Rhizobium Rh1 or Rhizobium Rh2 strains plus D1. Wood production was around 60 m3 ha−1 (Figure 6). Single inoculation with Rhizobium Rh1 and No AMF/No Fertilizer produced 57 m3 ha−1 (Figure 6). In this experiment, only two treatments (No AMF/Rh1/No Fertilizer and Ac/Rh1/Fertilizer D1) increased wood production by more than 20% (Figure 6). The Ac/Rh2/Fertilizer D1 treatment resulted in a 30% increase in wood production (Figure 6) when compared to controls. The results showed that wood production varied with inoculation with growth-promoting microorganisms. No PGPR/No Fertilizer produced 46.3 m3 ha−1, and values for only fertilization with D1 or D2 were 48.5 and 54 m3 ha−1, respectively (Figure 6).
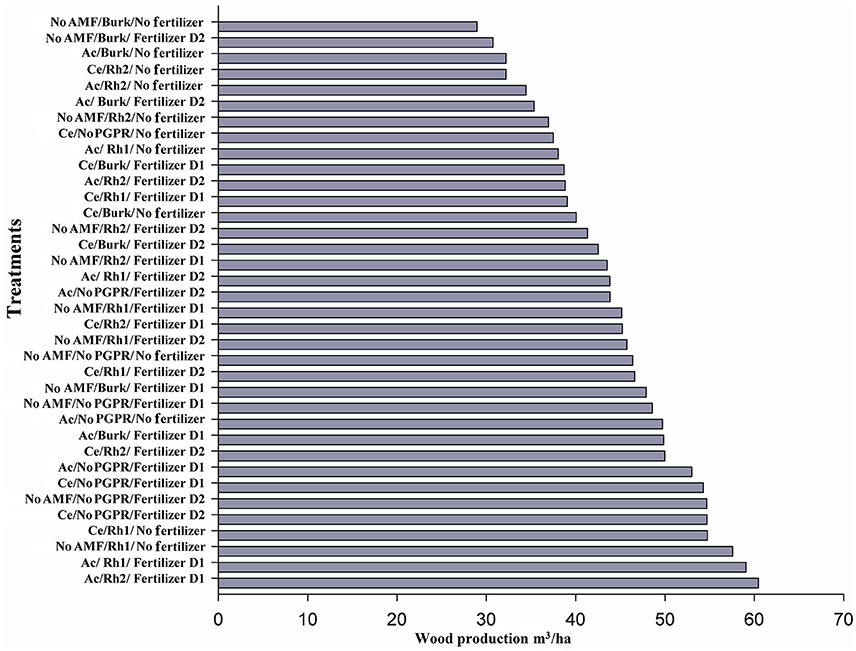
Figure 6. Wood production by different combinations of AMF [Acaulospora sp. (Ac) and Claroideoglomus etunicatum (Ce)], PGPR [Burkholderia sp. (Burk), Rhizobium Rh1, and Rh2], and chemical fertilizer [D1: 75 g plant−1, D2: 150 g plant−1] 2 years after seedling planting.
Discussion
Several biotic and abiotic factors influence the structural and functional diversity of bacterial communities (Berg and Smalla, 2009). In the relationship between plant and microbial rhizosphere communities, root exudates play an important role in selecting specific microbial populations (Bais et al., 2006). Hence, different plant species are associated with microorganisms that exhibit different responses in terms of survival and activity. In this way, it is necessary to evaluate and select microorganisms from site-specific plant associations, to optimize the inoculant for applications in plant production. The physiological characteristics of the inoculant organism determine to a great extent its fate and activity in soil (Van Veen et al., 1997). In the present study, Rhizobium strains promoted the growth of S. parahyba var. amazonicum when used alone or in combination with C. etunicatum or Acaulospora sp. in two planting methods. The indigenous isolates of Rhizobium were more effective than the exogenous strain of Burkholderia sp.
The interaction between AMF and Rhizobium improved the development of S. parahyba var. amazonicum trees from seeds and seedlings. It is believed that the mycorrhiza increase the effectiveness of Rhizobium as a result of the general increase in nutritional supply of the host plant (Barea et al., 2002; Bhowmik and Singh, 2004). The ability of Rhizobium bacteria to act as endophytes (Spencer et al., 1994; Lupwayi et al., 2004) and PGPR in non-nodulated plants has been confirmed by several studies in other plant species (Yanni et al., 2001; Hossain and Martensson, 2008). Acting as PGPR, rhizobia can support plant growth by solubilizing organic and inorganic phosphates and releasing phytohormones, enzymes, siderophores, exopolysaccharides, and riboflavin (Deshwal, 2013). They can also promote growth by inhibiting the growth of pathogens by the release of antibiotic compounds and/or iron immobilization by siderophore production (Mehboob et al., 2012). The double inoculation of Rhizobium and AM fungi has been shown to improve plant growth by increasing the nitrogen and phosphorus contents in plant biomass, resulting in improved soil nutrient availability (Matias et al., 2009).
Mycorrhizal inoculation can be integrated into nursery propagation of forestry species, thereby improving planting performance (Herrera et al., 1993). A more appropriate management of mycorrhizal symbiosis in poor soils would allow substantial reduction in the amount of minerals resulting in minimizing losses in productivity, while at the same time permitting a more sustainable production management (Soka and Ritchie, 2014). Due to the low fertility of the soil in the experimental area (Table 2), application of chemical fertilizers significantly promotes tree growth. This is a common practice in forestry systems in the area, even though this increases the cost of wood production. The main objective of this work was to reduce or improve efficiency of chemical fertilizer application by in situ microorganism inoculation of tropical legume trees.
This was demonstrated by the positive effect on plant growth and wood production with the application of combinations of AMF and Rhizobium, which were complemented with the addition of low doses of chemical fertilizer, especially in the seed system. The doses applied in the experiment were lower than those reported by Viégas et al. (2007) who used 255–272 g/plant for S. parahyba var. amazonicum cultivation in the Amazon area. Diameter, height and biomass of S. parahyba var. amazonicum after 480 days were equal to or greater than values obtained in plants fertilized with the recommended amount of fertilizer, suggesting a favorable and synergistic action between low fertilization and inoculation with Rhizobium and/or AMF.
S. parahyba var. amazonicum shows fast growth, reaching a volumetric production of up to 30 m3 ha−1 year−1 after 6 years of growth (Carvalho, 2007). In this study, the estimation of wood yield with inoculation of microorganisms reached more than 60 m3 ha−1 in 2 years with the best treatments (Ce/Rh1/Fertilizer D2 in seed sowing and Ac/Rh2/Fertilizer D1 in seedling planting) reaching the maximum yield in 2 years. The promotion of growth of S. parahyba var. amazonicum by microorganism inoculation has a secondary benefit: carbon sequestration. S. parahyba var. amazonicum has a low to moderate wood specific density (0.40 g cm−3), with carbon representing around 50% of dry matter. The amount of fixed carbon increases when wood production increases.
Conclusion
The use of microorganisms combined or not with fertilizer was more effective in plant growth and wood production in the seeds experiment as compared to the seedling experiment. Wood yield was almost the same in the two systems. However, when using seeds, many treatments increased plant growth and wood yield, and in the seedling system, only three treatments were more effective compared to control plants.
The use of native microorganisms as an inoculant for S. parahyba var. amazonicum was very effective, especially when combined with low doses of fertilizer, resulting in increased plant growth and wood yield under field conditions. In addition, the inoculation of Acaulospora sp. and bacteria improved the absorption of chemical fertilizer, enhancing wood yield. When compared with non-fertilized trees, the best treatments increased wood production by more than 50%. The inoculation of Acaulospora sp. and Rhizobium Rh1 with D1 fertilizer was the most effective treatment in both systems.
Author Contributions
MC: Experimental design and collect of data in the field. MS: Plant culture in the field. JE, FS, VF, AB, EG, GL: Field team control pest, average of plants in the field. IS, AO: Statistical analysis. GA: General coordinator, Head leader team.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We thank the National Council of Scientific and Technological Development (CNPq) who made this study possible by awarding PIBIC, MSc., PhD and Productivity in Research grants; as well as, the PUBLIC-FAEPE UEL/2016 fund. Dr. A. Leyva that helped with editing of the manuscript.
References
ABRAF (2012). Data from: Brazilian Association of Forest Plantation Producers. Forest plantations in Brazil. Available online at: http://www.abraflor.org.br/estatisticas.asp
Andrade, G. (2004). “Role of functional groups of microorganisms on the rhizosphere microcosm dynamics,” in Plant Surface Microbiology, eds A. Varma, L. Abbott, D. Werne, and R. Hampp (New York, NY: Springer), 51–68.
Artursson, V., Finlay, R. D., and Jansson, J. K. (2006). Interactions between arbuscular mycorrhizal fungi and bacteria and their potential for stimulating plant growth. Environ. Microbiol. 8, 1–10. doi: 10.1111/j.1462-2920.2005.00942.x
Bais, H. P., Weir, T. L., Perry, L. G., Gilroy, S., and Vivanco, J. M. (2006). The role of root exudates in rhizosphere interactions with plants and other organisms. Annu. Rev. Plant Biol. 57, 233–266. doi: 10.1146/annurev.arplant.57.032905.105159
Barea, J. M., Azcon, R., and Azcón-Aguilar, C. (2002). Mycorrhizosphere interactions to improve plant fitness and soil quality. A. Van Leeuw. J. Microb. 81, 343–351. doi: 10.1023/A:1020588701325
Berg, G., and Smalla, K. (2009). Plant species and soil type cooperatively shape the structure and function of microbial communities in the rhizosphere. FEMS Microbiol. Ecol. 68, 1. doi: 10.1111/j.1574-6941.2009.00654.x
Beringer, J. E. R. (1974). Factor transfer in Rhizobium leguminosarum. J. Gen. Microbiol. 84, 88–198. doi: 10.1099/00221287-84-1-188
Bhowmik, S. N., and Singh, C. S. (2004). Mass multiplication of AM inoculums, effect of plant growth-promoting rhizobacteria and yeast in rapid culturing of Glomus mosseae. Curr. Sci. 86, 705–709.
Bianciotto, V., and Bonfante, P. (2002). Arbuscular mycorrhizal fungi: a specialized niche for rhizospheric and endocellular bacteria. A. Van Leeuw. J. Microb. 81, 365–371. doi: 10.1023/A:1020544919072
Biró, B., Köves-Péchy, K., Vörös, I., Takács, T., Eggenberge, P., and Strasser, R. J. (2000). Interrelations between Azospirillum and Rhizobium nitrogen-fixers and arbuscular mycorrhizal fungi in the rhizosphere of alfalfa in sterile, AMF-free or normal soil conditions. Agric. Ecosyst. Environ. Appl. Soil Ecol. 15, 159–168. doi: 10.1016/S0929-1393(00)00092-5
Bryan, J. A., Berlym, G. P., and Gordon, J. C. (1996). Toward a new concept of the evolution of symbiotic nitrogen fixation in the Leguminosae. Plant Soil. 86, 151–159. doi: 10.1007/BF00035069
Chaer, G. M., Resende, A. S., Campello, E. F. C., De Faria, S. M., and Boddey, R. M. (2011). Nitrogen-fixing legume tree species for the reclamation of severely degraded lands in Brazil. Tree Physiol. 31, 139–149. doi: 10.1093/treephys/tpq116
Colpini, C., Travagin, D. P., Soares, T. S., and Silva, V. S. M. (2009). Determinação do volume, do fator de forma e da porcentagem de casca de árvores individuais em uma floresta ombrófila aberta na região noroeste de Mato Grosso. Acta Amaz. 39, 97–104. doi: 10.1590/S0044-59672009000100010
da Silva, K. E., Martins, S. V., Ribeiro, C. A., Santos, N. T., de Azevedo, C. P., Matos, F. D., et al. (2011). Floristic composition and similarity of 15 hectares in central Amazon. Rev. Biol. Trop. 59, 1927–1938.
Deshwal, V. K. (2013). Rhizobia unique plant growth promoting rhizobacteria: a review. Int. J. L. Sci. 2, 74–86.
Döbereiner, J., and Day, J. M. (1976). “Associative symbiosis in subtropical grasses: characterization of microorganisms and dinitrogen fixing sites,” in Proceedings of the International Symposium on Nitrogen Fixation II, eds W. E. Newton and C. J. Nyman (Washington, DC: University Press), 518–538.
Faria, S. M., Diedhiou, A. G., De Lima, H. C., Ribeiro, R. D., Galiana, A., Castilho, A. F., et al. (2010). Evaluating the nodulation status of leguminous species from the Amazonian forest of Brazil. J. Exp. Bot. 61, 3119–3127. doi: 10.1093/jxb/erq142
Garg, N., and Chandel, S. (2010). Arbuscular mycorrhizal networks: process and functions, a review. Agron. Sustain. Dev. 30, 581–599. doi: 10.1051/agro/2009054
Giri, B., Giang, P. H., Kumari, R., Prasad, R., and Varma, A. (2005). “Microorganisms in soils: roles in genesis and functions” in Soil Biology, eds F. Buscotand and A. Varma (New York, NY: Springer-Verlag), 19–43.
Hashem, A., Abd Allah, E. F., Alqarawi, A. A., Al-Huqail, A. A., Wirth, S., and Egamberdieva, D. (2016). The interaction between arbuscular mycorrhizal fungi and endophytic bacteria enhances plant growth of Acacia gerrardii under salt stress. Front. Microbiol. 7:1089. doi: 10.3389/fmicb.2016.01089
Herrera, M. A., Salamanca, C. P., and Barea, J. M. (1993). Inoculation of woody legumes with select arbuscular mycorrhizal fungi and rizobia to recover desertified Mediterranean ecosystems. Appl. Environ. Microb. 59, 129–133.
Hossain, M. D. S., and Martensson, A. (2008). Potential use of Rhizobium spp. to improve fitness of non-nitrogen-fixing plants. Acta Agr. Scand. B Soil Plant Sci. 58, 352–358. doi: 10.1080/09064710701788810
Jung, S. C., Martinez-Medina, A., Lopez-Raez, J. Á., and Pozo, M. J. (2012). Mycorrhiza-induced resistance and priming of plant defenses. J. Chem. Ecol. 38, 651–664. doi: 10.1007/s10886-012-0134-6
Lupwayi, N. Z., Clayton, G. W., Hanson, K. G., Rice, W. A., and Biederbeck, V. O. (2004). Endophytic rhizobia in barley, wheat and canola roots. Can. J. Plant Sci. 84, 37–45. doi: 10.4141/P03-087
Marques, M. S., Pagano, M., and Scotti, M. R. M. L. (2001). Dual inoculation of a woody legume (Centrolobium tomentosum) with rhizobia and mycorrhizal fungi in South-Eastern Brazil. Agroforest Syst. 52, 107–117. doi: 10.1023/A:1010637401475
Matias, S. R., Pagano, M. C., Muzzi, F. C., Oliveira, C. A., Carneiro, A. A., Horta, S. N., et al. (2009). Effect of rhizobia, mycorrhizal fungi and phosphate-solubilizing microorganisms in the rhizosphere of native plants used to recover an iron ore area in Brazil. Soil. Biol. 45, 259–266. doi: 10.1016/j.ejsobi.2009.02.003
Mehboob, I., Naveed, M., Zahir, Z. A., and Ashraf, M. (2012). “Potential of rizobia for sustainable production of non-legumes,” in Crop Production for Agricultural Improvement, eds M. Ashraf, M. Öztürk, M. S. A. Ahmad, and A. Aksoy (Rotterdam: Springer), 659–704.
Nadeem, S. M., Ahmad, M., Zahir, Z. A., Javaid, A., and Ashraf, M. (2014). The role of mycorrhizae and plant growth promoting rhizobacteria (PGPR) in improving crop productivity under stressful environments. Biotechnol. Adv. 32, 429–448. doi: 10.1016/j.biotechadv.2013.12.005
Neumann, E., and George, E. (2010). “Nutrient uptake: the arbuscular mycorrhiza fungal symbiosis as a plant nutrient acquisition strategy,” in Arbuscular Mycorrhizas: Physiology and Function, eds H. Koltai and Y. Kapulnik (New York, NY: Springer), 137–167.
Patreze, M. C., and Cordeiro, L. (2004). Nitrogen-fixing and vesicular arbuscular mycorrhizal symbioses in some tropical legume trees of tribe Mimoseae. Forest Ecol. Manag. 196, 275–285. doi: 10.1016/j.foreco.2004.03.034
Raimam, M. P., Albino, U., Cruz, M. F., Lovato, G. M., Spago, F., Ferracin, T. P., et al. (2007). Interaction among free-living N-fixing bacteria isolated from Droseravillosa var. villosa and AM fungi (Glomus clarum) in rice (Oryzasativa). Appl. Soil. Ecol. 35, 25–34. doi: 10.1016/j.apsoil.2006.05.013
Rillig, M. C. (2004). Arbuscular mycorrhizae, glomalin, and soil aggregation. Can. J. Soil Sci. 84, 355–363. doi: 10.4141/S04-003
Secretaria de Estado de Indústria Comércio e Mineração (SEICOM). (2012). Estatística Municipal. Dom Eliseu, Governo do Estado do Pará. Available online at: http://seicom.pa.gov.br/kitmineracao/estatistica-municipal/regiao-do-rio-capim/DomEliseu.pdf
Silva, M. F., Carreira, L. M. M., Távares, A. S., Ribeiro, I. C., Jardim, M. A. G., Lobo, M. G. A., et al. (1988). As leguminosas da Amazônia brasileira: lista prévia. Acta Bot. Bras. 2, 193–237.
Siviero, M. A., Marega, A., Lima Dos Santos, D., Rosselli, B. R., Huh, Y. S., Santinoni, I., et al. (2008). Interaction among N-fixing bacteria and AM fungi in Amazonian legume tree (Schizolobium amazonicum) in field conditions. Appl. Soil Ecol. 39, 144–152. doi: 10.1016/j.apsoil.2007.12.004
Soka, G., and Ritchie, M. (2014). Arbuscular mycorrhizal symbiosis and ecosystem processes: prospects for future research tropical soils. OJE 4, 11–22. doi: 10.4236/oje.2014.41002
Spencer, D., James, E. K., Ellis, G. J., Shaw, J. E., and Sprent, J. I. (1994). Interaction between rhizobia and potato tissues. J. Exp. Bot. 45, 1475–1482. doi: 10.1093/jxb/45.10.1475
Sprent, J. I. (1983). “Agricultural and horticultural systems: implications for forestry,” in Biological Nitrogen Fixation in Forest Ecosystems: Foundations and Applications, eds J. C. Gordon and C.T. Wheeler (Boston, MA: Martinus Nijhoff/Dr. W. Junk Publishers), 213–232.
Sprent, J. I., and Parsons, R. (2000). Nitrogen fixation in legume and non-legume trees. Field Crop. Res. 65, 183–196. doi: 10.1016/S0378-4290(99)00086-6
Valdenegro, M., Barea, J. M., and Azcón, R. (2001). Influence of arbuscular-mycorrhizal fungi, Rhizobium meliloti strains and PGPR inoculation on the growth of Medicago arborea used as model legume for revegetation and biological reactivation in a semiarid Mediterranean area. Plant Growth Regul. 34, 233–240. doi: 10.1023/A:1013323529603
Van Sambeek, J. W., Navarrete-Tindall, N. E., and Hunt, K. L. (2007). “Growth and foliar nitrogen concentrations of interplanted native woody legumes and pecan,” in Proceedings of the 16th Central Hardwoods Forest Conference (Newtown Square, PA: U.S. Department of Agriculture, Forest Service, Northern Research Station), 580–588.
Van Veen, J. A., Van Overbeek, L. S., and Van Elsas, J. D. (1997). Fate and activity of microorganisms introduced into soil. Microbiol. Mol. Biol. R. 6, 121–135.
Vessey, J. K. (2003). Plant growth promoting rhizobacteria as biofertilizers. Plant Soil. 255, 571–586. doi: 10.1023/A:1026037216893
Viégas, M. I., Azevedo, R. E. J., Alvez, T. M. A., and Sato, T. (2007). Efeito da Adubação NPK em Plantas Jovens de Paricá (Schizolobium amazonicum, Huber Ducke). Colombo: Embrapa Florestas.
Keywords: microorganism interaction, reforestation, Amazon forest, Schizolobium parahyba, mycorrhizal inoculant
Citation: Cely MVT, Siviero MA, Emiliano J, Spago FR, Freitas VF, Barazetti AR, Goya ET, Lamberti GdS, dos Santos IMO, De Oliveira AG and Andrade G (2016) Inoculation of Schizolobium parahyba with Mycorrhizal Fungi and Plant Growth-Promoting Rhizobacteria Increases Wood Yield under Field Conditions. Front. Plant Sci. 7:1708. doi: 10.3389/fpls.2016.01708
Received: 27 June 2016; Accepted: 31 October 2016;
Published: 22 November 2016.
Edited by:
Mohamed Hijri, Université de Montréal, CanadaReviewed by:
Raffaella Balestrini, National Research Council, ItalyChristian Staehelin, Sun Yat-sen University, China
Copyright © 2016 Cely, Siviero, Emiliano, Spago, Freitas, Barazetti, Goya, Lamberti, dos Santos, De Oliveira and Andrade. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Galdino Andrade, Z2FsLmFuZHJhZGU3QGdtYWlsLmNvbQ==