 Kashif Mahmood
Kashif Mahmood Zhenhua Xu
Zhenhua Xu Ashraf El-Kereamy
Ashraf El-Kereamy José A. Casaretto
José A. Casaretto Steven J. Rothstein
Steven J. Rothstein- Department of Molecular and Cellular Biology, University of Guelph, Guelph, ON, Canada
Production of anthocyanins is one of the adaptive responses employed by plants during stress conditions. During stress, anthocyanin biosynthesis is mainly regulated at the transcriptional level via a complex interplay between activators and repressors of anthocyanin biosynthesis genes. In this study, we investigated the role of a NAC transcription factor, ANAC032, in the regulation of anthocyanin biosynthesis during stress conditions. ANAC032 expression was found to be induced by exogenous sucrose as well as high light (HL) stress. Using biochemical, molecular and transgenic approaches, we show that ANAC032 represses anthocyanin biosynthesis in response to sucrose treatment, HL and oxidative stress. ANAC032 was found to negatively affect anthocyanin accumulation and the expression of anthocyanin biosynthesis (DFR, ANS/LDOX) and positive regulatory (TT8) genes as demonstrated in overexpression line (35S:ANAC032) compared to wild-type under HL stress. The chimeric repressor line (35S:ANAC032-SRDX) exhibited the opposite expression patterns for these genes. The negative impact of ANAC032 on the expression of DFR, ANS/LDOX and TT8 was found to be correlated with the altered expression of negative regulators of anthocyanin biosynthesis, AtMYBL2 and SPL9. In addition to this, ANAC032 also repressed the MeJA- and ABA-induced anthocyanin biosynthesis. As a result, transgenic lines overexpressing ANAC032 (35S:ANAC032) produced drastically reduced levels of anthocyanin pigment compared to wild-type when challenged with salinity stress. However, transgenic chimeric repressor lines (35S:ANAC032-SRDX) exhibited the opposite phenotype. Our results suggest that ANAC032 functions as a negative regulator of anthocyanin biosynthesis in Arabidopsis thaliana during stress conditions.
Introduction
Anthocyanin pigments belong to the flavonoid-class of secondary metabolites originating from the phenylpropanoid pathway. Production of anthocyanins is spatio-temporally regulated in different plant species (Jackson et al., 1992; Procissi et al., 1997; Gou et al., 2011). In addition, anthocyanin biosynthesis is also induced in response to a variety of stress conditions such as UV light, HL (Vanderauwera et al., 2005; Cominelli et al., 2008; Morishita et al., 2009; Lotkowska et al., 2015; Viola et al., 2016), nutrient deficiency (Jiang et al., 2007; Peng et al., 2008), drought (Castellarin et al., 2007), salinity (Eryilmaz, 2006; Lotkowska et al., 2015) and extreme temperature fluctuations (Christie et al., 1994; Yamane et al., 2006). Under HL and UV irradiation, anthocyanins act as a sun screen and function in protecting photosynthetic organs from photoinhibition (Hughes et al., 2005; Albert et al., 2009). Other abiotic stresses such as drought, salinity and extreme temperature fluctuations also result in ROS production, and anthocyanin accumulation during these conditions is likely to confer tolerance through ROS scavenging (Nakabayashi et al., 2014).
The enzymatic pathway involved in the production of anthocyanin pigments has been well elucidated with the identification of a number of mutants in different plant species, consisting of early and late biosynthesis genes (EBGs and LBGs). EBGs such as chalcone synthase (CHS), chalcone isomerase (CHI), flavanone 3-hydroxylase (F3H), and flavonoid 3′-hydroxylase (F3′H) are involved in the production of different types of flavonoids including anthocyanin. LBGs such as dihydroflavonol 4-reductase (DFR), leucoanthocyanidin oxygenase (LDOX), anthocyanidin reductase (ANR) and UDP-glucose:flavonoid 3-O-glucosyltransferase (UF3GT) are specific for anthocyanin biosynthesis only. The regulation of anthocyanin production is mainly achieved through a concerted action of a number of transcription factors (TFs). Both positive and negative transcriptional regulatory mechanisms for anthocyanin production have been identified. Positive transcriptional regulation in Arabidopsis thaliana involves the formation of a MBW protein complex, comprising members of the MYBs (TT2, PAP1; MYB75, PAP2, MYB113 or MYB114), bHLHs (TT8, GL3 and EGL3) and WD40-repeat proteins (TTG1) (Walker et al., 1999; Carey et al., 2004; Teng et al., 2005; Baudry et al., 2006; Quattrocchio et al., 2006; Gonzalez et al., 2008). In the context of negative regulation, two members of the small R3-MYB TF family have been identified in Arabidopsis to repress anthocyanin biosynthesis. One is AtMYBL2 that interacts with TT8 to form the L2BW complex which represses anthocyanin biosynthesis (Dubos et al., 2008; Matsui et al., 2008). Interplay between MBW and L2BW complex has thus been proposed to regulate anthocyanin biosynthesis in A. thaliana (Dubos et al., 2008; Matsui et al., 2008). Another small R3-MYb protein, CPC, has been implicated to repress anthocyanin biosynthesis by interacting with PAP1/PAP2 and interfering with the formation of the MBW complex (Zhu et al., 2009; Nemie-feyissa et al., 2014). In addition to R3-MYBs, a SQUAMOSA PROMOTER BINDING PROTEIN-LIKE TF, SPL9, was recently identified as a negative regulator of anthocyanin synthesis (Gou et al., 2011). Furthermore, three members of the Lateral Boundary Domain TF family, LBD37, LBD38, and LBD39, have been identified to directly repress expression of PAP1 and PAP2 which encode key regulators of anthocyanin biosynthesis in A. thaliana (Rubin et al., 2009).
Production of anthocyanin pigments during adverse growth conditions is considered one of the adaptive responses employed by plants (Chalker-Scott, 1999; Stewart et al., 2001; Peng et al., 2008; Cui et al., 2014) and again, interplay between positive and negative transcriptional regulators governs anthocyanin accumulation under unfavorable conditions. For example, under HL stress, expression of genes encoding the TFs that form the MBW complex, such as PAP1, PAP2, TT8, EGL3, TTG1 are strongly induced (Cominelli et al., 2008). Similar changes in the expression pattern of PAP1, PAP2 and TT8 were observed in Arabidopsis plants grown under different abiotic stress conditions (Kilian et al., 2007; Peng et al., 2008). In contrast, expression of negative regulators of anthocyanin biosynthesis such as AtMYBL2, LBD37 and LBD38 is reduced during stress conditions (Kilian et al., 2007; Dubos et al., 2008; Rubin et al., 2009). Sugars have also been implicated in the induction of anthocyanin pigments (Weiss, 2000; Teng et al., 2005; Solfanelli et al., 2006; Kilian et al., 2007; Dubos et al., 2008; Rubin et al., 2009), especially in the hormonal regulation of anthocyanin biosynthesis. For example, treatment of Arabidopsis with ABA and jasmonic acid (JA) induced anthocyanin production only in the presence of sucrose (Loreti et al., 2008). The involvement of sugars in the hormone-induced anthocyanin production seems relevant since the ABA and JA biosynthesis is triggered by salinity, drought and HL stresses (Wang et al., 2001; Walia et al., 2006; Galvez-Valdivieso et al., 2009; Ramel et al., 2013), and these adverse conditions also result in the increased accumulation of soluble sugar in different plant species (Lichtenthaler et al., 1981; Dubey and Singh, 1999; Kempa et al., 2008; Krasensky and Jonak, 2012; Schmitz et al., 2014).
In Arabidopsis thaliana, the NAC (No Apical Meristem/NAM, Arabidopsis ATAF1/2, and Cup-shaped Cotyledon2/CUC2) TF family has been shown to regulate diverse biological processes ranging from plant development to responses to stress conditions (Duval et al., 2002; Guo and Gan, 2006; Ko et al., 2007; Balazadeh et al., 2010; Yang et al., 2011; Lee et al., 2012; Mahmood et al., 2016). Although, some members of the NAC TF family have been implicated in modulating the phenylpropanoid pathway specific to lignin biosynthesis (Kubo et al., 2005; Mitsuda et al., 2005, 2007; Zhong et al., 2008; Ohashi-Ito et al., 2010), only a few have been identified to regulate anthocyanin biosynthesis. For instance, ANAC078 has been shown to positively regulate anthocyanin production under HL stress (Morishita et al., 2009) whereas JUB1/ANAC042 was shown to negatively affect the biosynthesis of anthocyanins in Arabidopsis (Wu et al., 2012). Here, we analyzed the role of ANAC032 in the regulation of anthocyanin biosynthesis. By employing biochemical, molecular and transgenic approaches, we show that ANAC032 negatively regulates anthocyanin biosynthesis in response to abiotic stresses including HL, salinity and oxidative stress. ANAC032 expression was found to be induced by a range of stress conditions, suggesting an important regulatory role of ANAC032 in the biosynthesis of anthocyanin during stress.
Materials and Methods
Growth Conditions and Treatments
For sucrose-induced anthocyanin accumulation, surface sterilized seeds of wild-type and ANAC032 transgenic lines were germinated and grown on half-strength MS ager plates supplemented with 0, 1.5, 3, and 6% sucrose for 5 days under long-day (16 h/8 h light/dark) as well as under continuous light conditions. For JA-induced anthocyanin accumulation, surface sterilized seeds were germinated and grown on half-strength MS agar plates (1% sucrose) supplemented with 0, 25, and 50 μM MeJA for 12 days under long-day conditions (∼100 μmol m-2 s-1). For ABA treatment, 8-day-old in vitro grown seedlings were transferred to half-strength agar plates (1% sucrose) supplemented with 0, 10, and 20 μM ABA, and grown for 4 days under long-day conditions.
For HL stress, transgenic and wild-type plants were grown on LA4 Sunshine mix for 17 days under long-day growth conditions at 150 μmol m-2 s-1. Plants were then transferred to a separate growth chamber set at 450 μmol m-2 s-1 for 4 days. Plants grown at 150 μmol m-2 s-1 were treated as control (normal light, NL). For salinity stress, plants were irrigated with saline water (200 mM NaCl) three times at 4 days interval. Plants were harvested 2 weeks after the first irrigation.
Generation of Transgenic Plants
Generation of transgenic plants, ANAC032 overexpression (35S:ANAC032), chimeric repressors (35S:ANAC032-SRDX) and PANAC032:GUS, have been described in Mahmood et al. (2016).
Histochemical Detection of B-glucuronidase (GUS) Activity
To analyze the GUS activity in response to HL stress, 20-day-old plants of the PANAC032:GUS transgenic line were treated with optimal (control, 150 μmol m-2 s-1) or HL stress (HL, ∼1000 μmol m-2 s-1) for 24 h. The whole rosettes were first collected in chilled 90% acetone and were then immersed in GUS staining solution [100 mM sodium phosphate buffer pH 7.2, 10 mM EDTA, 0.5 mM potassium ferricyanide, 0.5 mM potassium ferrocyanide, 2 mM X-Gluc (5-bromo-4-chloro-3-indolyl-β-D-glucuronic acid), 0.1% triton X-100, 20% methanol]. The samples were incubated overnight in the dark at 37°C after a brief vacuum infiltration. Rosettes were photographed after removing chlorophyll using 70% ethanol. For GUS activity in response to sucrose treatment, seeds of the PANAC032:GUS transgenic line were germinated and grown on half-strength MS agar plates supplemented with or without 1% sucrose for 5 days. The seedlings were subsequently transferred to fresh half-strength MS agar plates containing 1 and 6% sucrose for 24 h. After the treatment, seedlings were transferred to GUS staining solution and staining was performed as described above done in the same way as described for HL stress.
Determination of Anthocyanin Content
Anthocyanin content was determined using the acidified methanol method with some modifications. For salinity and HL stress, ∼ 50 mg leaf tissues were homogenized in 500 μL of methanol-HCl (1% v/v) and kept at 4°C overnight. Samples were centrifuged at 14,000 rpm for 10 min. Supernatants were transferred to 2 mL centrifuge tubes. Anthocyanins were extracted using the extraction solvent chloroform:H2O 1.75:1 (v/v). Briefly, to each supernatant sample, chloroform and deionized H2O were added in 1.75:1 ratio (v/v) and vortexed vigorously for 30 s. Each sample was again centrifuged at 14,000 rpm for 10 min and the supernatant layer was transferred to a new 1.5 ml centrifuge tube. Samples were analyzed by spectrophotometric method at 530 and 657 nm wavelengths. Anthocyanin content was calculated by subtracting A530 values from those for A657. For sucrose- and MeJA-induced anthocyanin accumulation, twenty seedlings from each genotype were pooled in a single replicate after 5 and 12 days of growth, respectively. For ABA-induced anthocyanin, eight seedlings were pooled together in a single replicate after 4 days of treatment. For these three treatments, samples were homogenized in 250 μL of methanol-HCl (1% v/v) and anthocyanins were analyzed as described.
Determination of Total Soluble Sugars
Total soluble sugars were determined through colorimetric quantification method as described in (Buysse and Merckx, 1993), with slight modifications. Briefly, 50 mg of the leaf tissues were homogenized in 0.5 mL ethanol (80%) and then centrifuged at 14,000 rpm for 5 min at 4°C. Supernatants were transferred to fresh 2 mL tubes. Homogenization and centrifugation steps were repeated 2–3 times, and supernatants were pooled. The final reaction mixture contained 0.5 mL of supernatants, 0.5 mL phenol (28% w/w) and 2.5 mL conc. sulphuric acid. Reaction was allowed to take place for 15 min and then the absorbance of the samples was recorded at 490 nm using plate reader. Glucose was used as a standard to determine the total soluble sugars in the samples.
RNA Isolation, cDNA Synthesis and Quantitative RT-PCR Analysis
Total RNA from leaf samples was isolated using the Total RNA Purification Kit (Norgen Biotek Corp). One microgram of the total RNA from each sample was reverse transcribed into cDNA using qScriptTM cDNA SuperMix (Quanta Biosciences) according to the manufacturer’s protocol. Real-time RT-PCR was performed using the PerfeCTa® SYBR® Green SuperMix (Quanta Biosciences) following the manufacturer’s protocol. Expression values from two technical and three biological replicates were analyzed using the comparative CT method. ACT7 and UBC21 were used as internal reference genes for normalization. The primer sequences used in this study are provided in Supplementary Table S1.
Histochemical Staining for Lignin Analysis (Wiesner Staining)
For lignin analysis in roots, 1-week-old seedlings were incubated in Wiesner solution (Phloroglucinol-HCl) (Mitra and Loqué, 2014) for 5 min and then visualized under a light microscope and photographed.
Results
ANAC032 Represses Sucrose-Induced Anthocyanin Biosynthesis
In a previous study, by generating and analyzing ANAC032 overexpressors (35S:ANAC032) and chimeric repressors (35S:ANAC032-SRDX), we have shown that ANAC032 promotes age-dependent and stress-induced senescence in A. thaliana (Mahmood et al., 2016). In addition to regulating the senescence processes, ANAC032 transgenic lines were also found to accumulate differential levels of anthocyanin pigments particularly during early growth phases (Supplementary Figure S1). Since production of anthocyanins is considered an important adaptive response particularly under stress conditions, we decided to investigate in more detail the mechanism by which ANAC032 affects anthocyanin accumulation during stress conditions.
Because sucrose is known to have a positive effect on the biosynthesis of anthocyanins, high sucrose treatment has been used as a reliable assay to analyze mutants involved in anthocyanin biosynthesis and regulation (Teng et al., 2005; Solfanelli et al., 2006; Rubin et al., 2009). To determine the role of ANAC032 in regulating anthocyanin biosynthesis, we first examined if the expression of ANAC032 is affected by sucrose. qRT-PCR results showed that treatment of Arabidopsis seedlings with higher sucrose levels (6% sucrose) induced the expression of ANAC032 compared to control conditions (1% sucrose) (Figure 1A). The effect of sucrose on the expression of ANAC032 was further investigated in detail using a PANAC032:GUS transgenic line. Histochemical analysis showed that the ANAC032 promoter drove expression of GUS only in the roots when seedlings were grown on plates lacking sucrose. However, upon transfer to plates with 6% sucrose, an increased GUS activity was also detected in cotyledons (Figure 1B). A similar pattern of GUS activity was observed when PANAC032:GUS seedlings were transferred from plates containing 1% sucrose to 6% sucrose (Figure 1C). A weak GUS activity was detected in the cotyledons of the PANAC032:GUS seedlings grown on 1% sucrose, however, an obvious increase in the GUS activity was observed in the cotyledons upon transfer to 6% sucrose (Figure 1C). These results show that ANAC032 expression is induced by sucrose treatment, most likely in a concentration dependent manner.
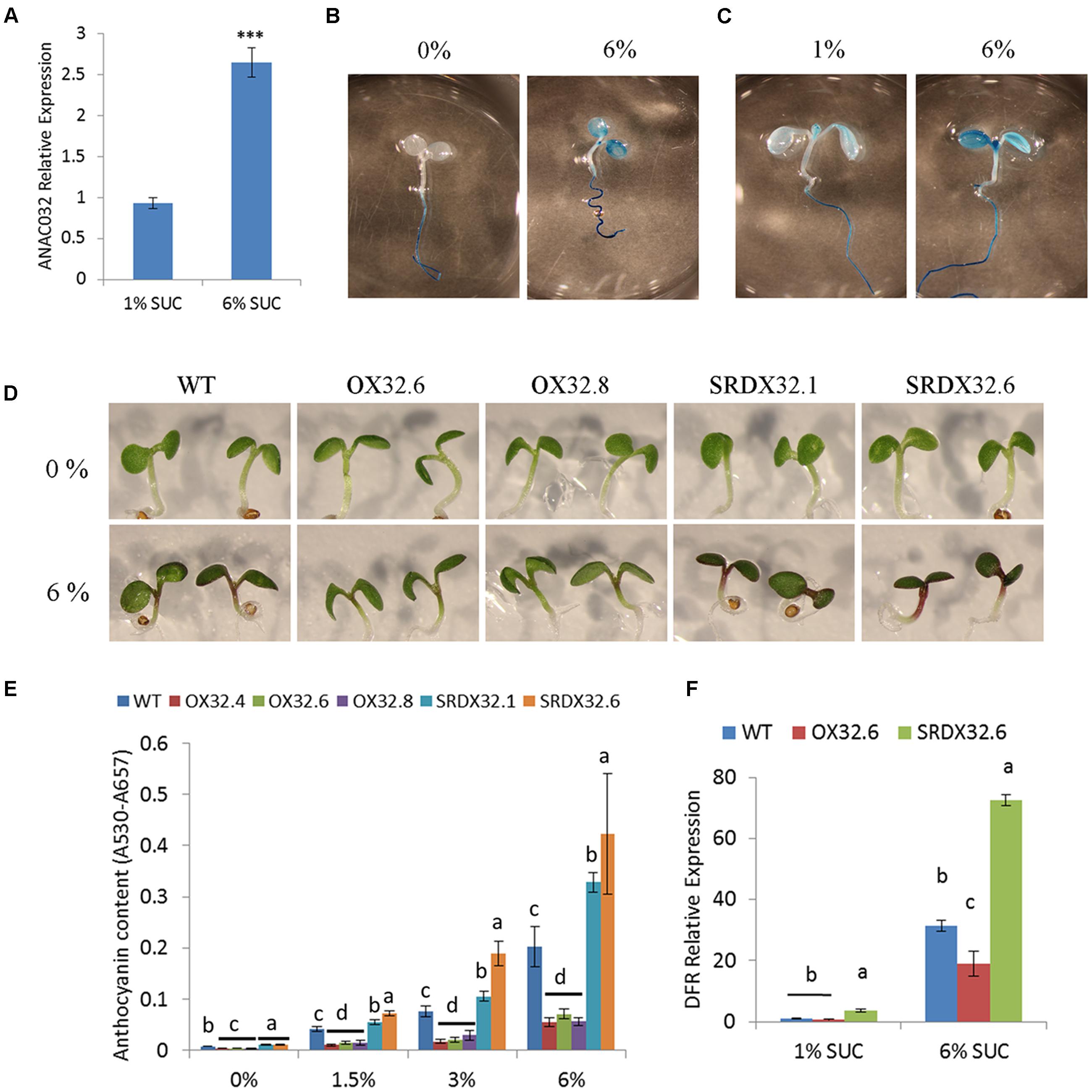
FIGURE 1. Regulation of sucrose–induced anthocyanin by ANAC032 in Arabidopsis. (A) qRT-PCR analysis of ANAC032 expression in response to sucrose treatment. Five-day-old seedlings were transferred to fresh half-strength MS plates supplemented with 1% or 6% sucrose for 24 h under continuous light conditions. Data represent mean values (±SD). UBC21 was used as internal control. Data were analyzed using Student’s t-test (∗∗∗P< 0.01). (B,C) β-glucuronidase activity of PANAC032: GUS line in response to high sucrose treatment. Five-day-old seedlings of PANAC032: GUS line, grown on half-strength MS plates with 0% (B) and 1% (C) sucrose were transferred to MS plates supplemented with 6% sucrose for 24 h, followed by incubation in GUS-staining solution overnight in dark at 37°C. (D) Phenotypic response of wild-type (WT) and ANAC032 transgenic lines (OX32; overexpressors and SRDX32; chimeric repressors) after 5 days of growth on half-strength agar plates containing 0 and 6% sucrose. (E) Anthocyanin content in the seedlings of ANAC032 transgenic lines, grown on half-strength MS agar plates containing 0, 1.5, 3, and 6% sucrose, after 5 days of growth. Data represent mean values (±SD; n = 3). (F) qRT-PCR analysis of DFR (dihydroflavanol reductase) expression in response to high sucrose treatment. Five-day-old seedlings of WT and ANAC032 transgenic lines were transferred to fresh half strength MS plates containing 1 and 6% sucrose for 24 h. UBC21 was used as internal control. Within each treatment, bars with different letters in (E,F) are statistically not similar to each other according to one way ANOVA LSD test (P < 0.05).
Induction of ANAC032 by sucrose prompted us to examine if anthocyanin biosynthesis may be altered in ANAC032 overexpression and SRDX lines upon treatment with sucrose. Seeds of wild-type and ANAC032 transgenic lines were grown on half-strength MS plates containing 0, 1.5, 3, and 6% sucrose under long-day growth conditions. After 5 days, anthocyanin levels increased in wild-type seedlings in a concentration dependent manner upon treatment with sucrose (Figures 1C,D). However, the seedlings of ANAC032 overexpression lines accumulated significantly lower anthocyanin pigments compared to wild-type seedlings (Figures 1C,D). In contrast, seedlings of both SRDX lines accumulated higher levels of anthocyanin pigments, suggesting that ANAC032 negatively regulates anthocyanin biosynthesis in A. thaliana in response to high sucrose (Figures 1C,D). The effect of high sucrose treatment on anthocyanin biosynthesis was further investigated by analyzing the expression of dihydroflavonol 4-reductase (DFR; responsible for the conversion of dihydroquercetin to leucocyanidin) in wild-type and transgenic lines. qRT-PCR results show that although DFR expression was induced by high sucrose in wild-type seedlings, its expression was significantly lower in an ANAC032 overexpression line (Figure 1F). On the contrary, an ANAC032 SRDX line had significantly higher transcript levels of DFR (Figure 1F).
ANAC032 Represses Anthocyanin Production in Response to High Light Stress
In our previous study, ANAC032 was shown to promote leaf senescence in response to HL stress (Mahmood et al., 2016). Here, using qRT-PCR and PANAC032:GUS line, we show that the expression of ANAC032 is induced by HL stress (Figures 2A,B). Analysis of ANAC032 transgenic lines exposed to a moderate HL stress (∼450 μmol m-2 s-1) showed that ANAC032 overexpression lines accumulated considerably lower levels of anthocyanin pigment compared to wild-type plants after 4 days of HL treatment (Figures 2C,D). In contrast, ANAC032 SRDX lines exhibited the opposite phenotype (Figures 2C,D). These results indicate that ANAC032 represses anthocyanin biosynthesis in response to HL stress. Since HL induces the accumulation of soluble sugars (Schmitz et al., 2014) and because ANAC032 transgenic lines accumulated altered anthocyanin content, we predicted that total soluble sugars accumulation might have been altered as well in the transgenic lines during HL stress. Indeed, ANAC032 SRDX lines accumulated significantly higher soluble sugars compared to wild-type under HL stress and at least two of the overexpression lines accumulated less (Figure 2E), suggesting a relationship between sugar metabolism and anthocyanin production.
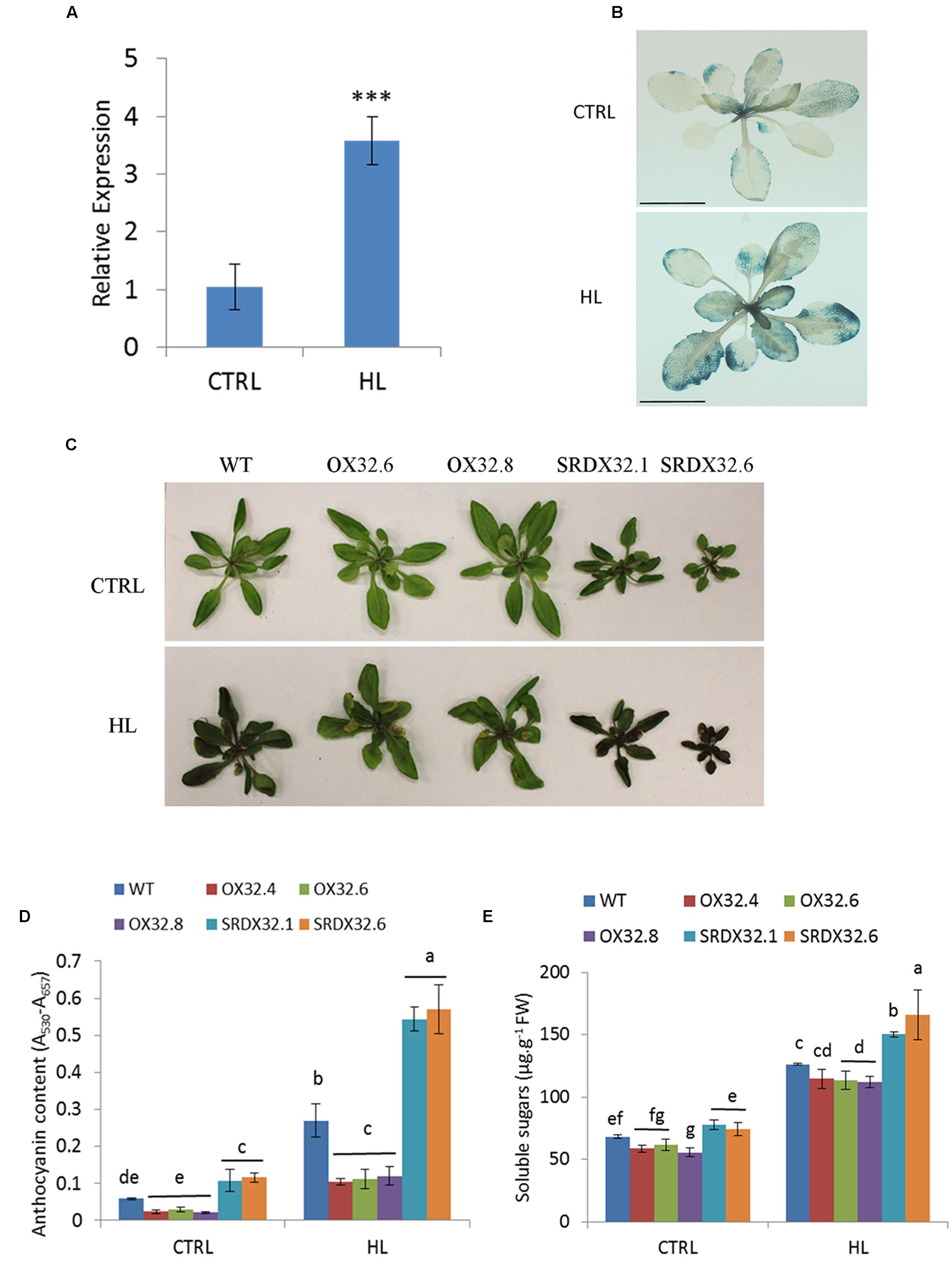
FIGURE 2. Regulation of high light-induced anthocyanin biosynthesis. (A) Expression analysis of ANAC032 in response to high light stress using qRT-PCR assay. Data represent mean values (±SD). ACT7 was used as internal control. Data were analyzed statistically using Student’s t-test (∗∗∗P < 0.01). (B) β-glucuronidase activity of PANAC032: GUS line in response to high light treatment. Twenty-day-old plants of PANAC032 : GUS line were transferred to control/optimal (150 μmol m-2 s-1) and high light (∼1000 μmol m-2 s-1) for 24 h and then incubated in GUS-staining solution overnight (scale bar = 1 cm). (C) Phenotype of WT and ANAC032 transgenic lines grown under optimal (control) and high light. (D) Biochemical analysis of anthocyanin content in WT and ANAC032 transgenic lines in response to high light stress. (E) Analysis of total soluble sugars in response to HL stress. 17-day-old plants of WT and ANAC032 overexpression and SRDX lines were grown under high light (∼450 μmol m-2 s-1) for 4 days. Plants grown under optimal light conditions (∼150 μmol m-2 s-1) were considered as control. Data represent values from three biological replicates. Bars with different letters are not statistically similar to each other according to one way ANOVA LSD test (P < 0.05).
Since HL stress leads to an oxidative burst associated with an increased proliferation of reaction oxygen species (ROS), we recapitulated the similar trend for anthocyanin biosynthesis in ANAC032 transgenic lines by exposing them to 3-AT. 3-AT is known to inhibit catalase activity, thereby resulting in an enhanced accumulation of H2O2. After 17 days of growth on half-strength MS agar plates supplemented with 0 and 7.5 μM 3-AT, ANAC032 overexpression lines accumulated considerably less anthocyanins compared to wild-type at 7.5 μM 3-AT (Figures 3A,B). On the contrary, ANAC032 SRDX lines accumulated significantly more anthocyanin pigments than wild-type (Figures 3A,B).
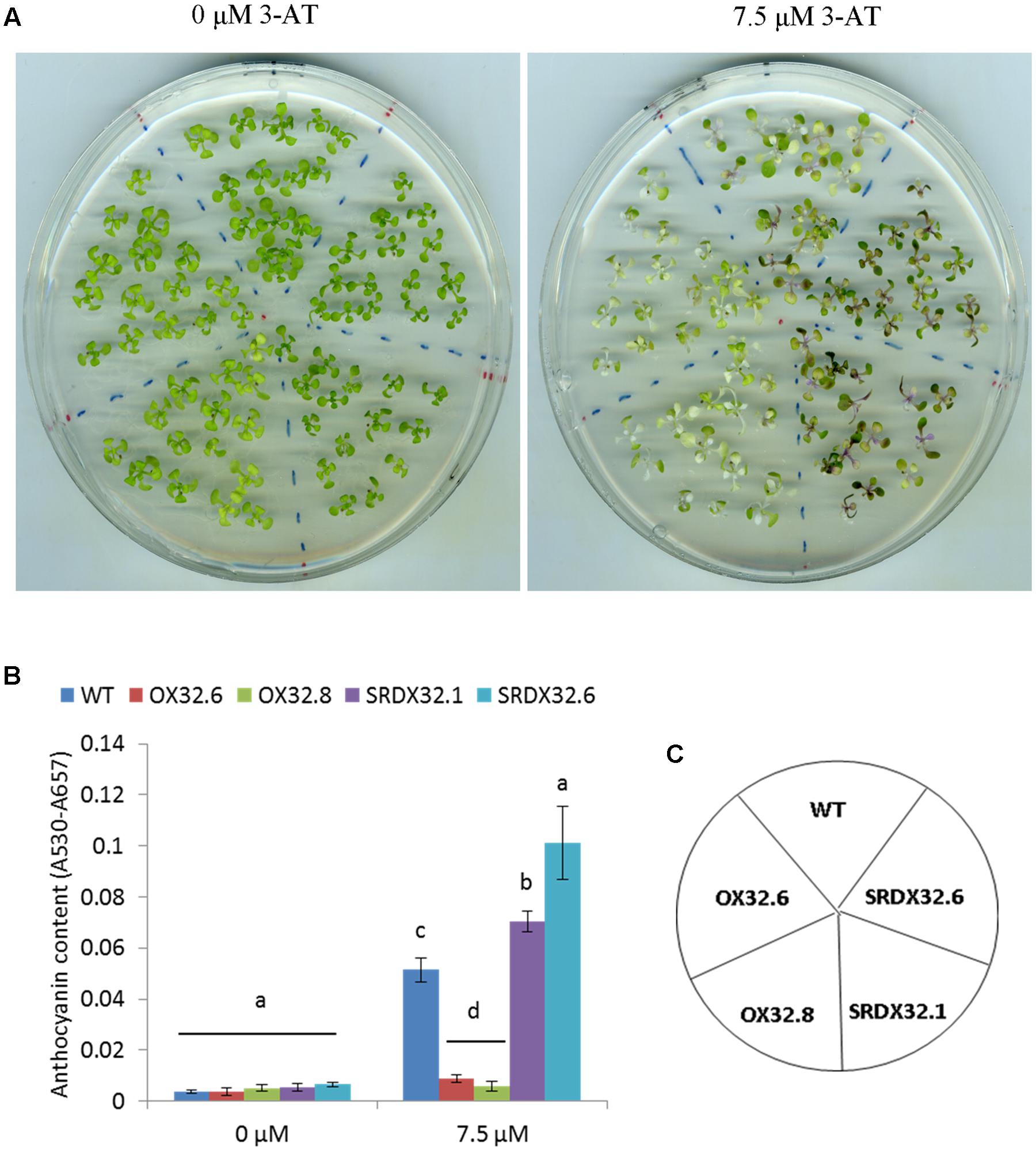
FIGURE 3. Effect of 3-AT on anthocyanin biosynthesis in WT and ANAC032 transgenic lines. (A) Phenotypic response of WT and ANAC032 overexpression and SRDX lines for anthocyanin biosynthesis in response to 3-AT treatment after 17 days of growth under long-day conditions. (B) Biochemical analysis of anthocyanin content in WT and ANAC032 transgenic lines after 17 days on 0 and 7.5 μM of 3-AT. (C) Template showing the position of the genotypes shown in (A). Data represent mean values from three biological replicates. Within each treatment, bars with different letters are not statistically similar to each other according to one way ANOVA LSD test (P < 0.05).
Expression of Anthocyanin Biosynthesis and Regulatory Genes Is Affected by ANAC032
Given the altered HL-induced anthocyanin accumulation in ANAC032 transgenic lines, we analyzed the molecular basis of these differences by investigating the transcript levels of genes involved in anthocyanin biosynthesis and its regulation. In the context of anthocyanin biosynthesis genes (ABGs), the expression levels of PAL1, CHS, DFR and ANS were determined in rosette leaves of wild-type and ANAC032 transgenic lines treated with HL stress. Consistent with the biochemical phenotype, qRT-PCR results showed that expression of particularly CHS, DFR and ANS, were drastically induced in wild-type leaves upon exposure to HL (Figure 4A). Interestingly, the expressions levels of both DFR and ANS, two late biosynthesis genes (LBGs), were significantly lower in the overexpression line compared to wild-type (Figure 4A) while transcript levels of both genes were significantly higher in the ANAC032 SRDX line (Figure 4A). Although expression of CHS was significantly induced in both transgenic lines, only the SRDX line presented significantly higher transcript levels compared to wild-type (Figure 4A). Next, we examined the transcript levels of regulatory genes known to activate or repress ABGs in A. thaliana. Amongst the activators, expression of PAP1 (a MYB TF) and TT8 (bHLH TF) was largely affected by HL in wild-type plants (Figure 4B). Unexpectedly, despite overexpression lines having significantly lower anthocyanin content (Figure 3C), PAP1 transcript levels were higher in the overexpression line compared to wild-type under HL (Figure 4B). Although, the PAP1 expression was higher in the SRDX line under control conditions, its expression was similar to wild-type under HL (Figure 4B). In the case of TT8, its transcript levels were significantly lower in the overexpression line compared to wild-type plants whereas they were higher in the SRDX line (Figure 4B). The expression of GL3 (a bHLH homolog of TT8) and TTG1 (WD40 repeat) were either slightly altered or remained unchanged in ANAC032 transgenic lines under both control and HL conditions (Figure 4B). In addition, we also examined the transcript abundance of known repressors of anthocyanin biosynthesis. Results show that expression of MYBL2 and LBD37 were downregulated by HL in wild-type plants whereas that of CPC and SPL9 remained unchanged (Figure 4C). Under control conditions, the SRDX line displayed lower transcript levels of MYBL2 while the overexpression line had transcript levels comparable to wild-type. However, under HL stress, the overexpression line and the SRDX line presented, respectively, higher and lower transcript levels of MYBL2 than wild-type (Figure 4C). Transcript levels of SPL9 were significantly reduced in the SRDX line but were similar in the overexpression line and wild-type under both control and HL conditions (Figure 4C). Although expression of LBD37 were reduced in wild-type plants by HL treatment, its expression was further reduced in the overexpression line and induced in the SRDX line compared to the control condition (Figure 4C). The expression of CPC was only elevated in the overexpression line while remained unaltered in the SRDX line compared to wild-type plants under HL (Figure 4C).
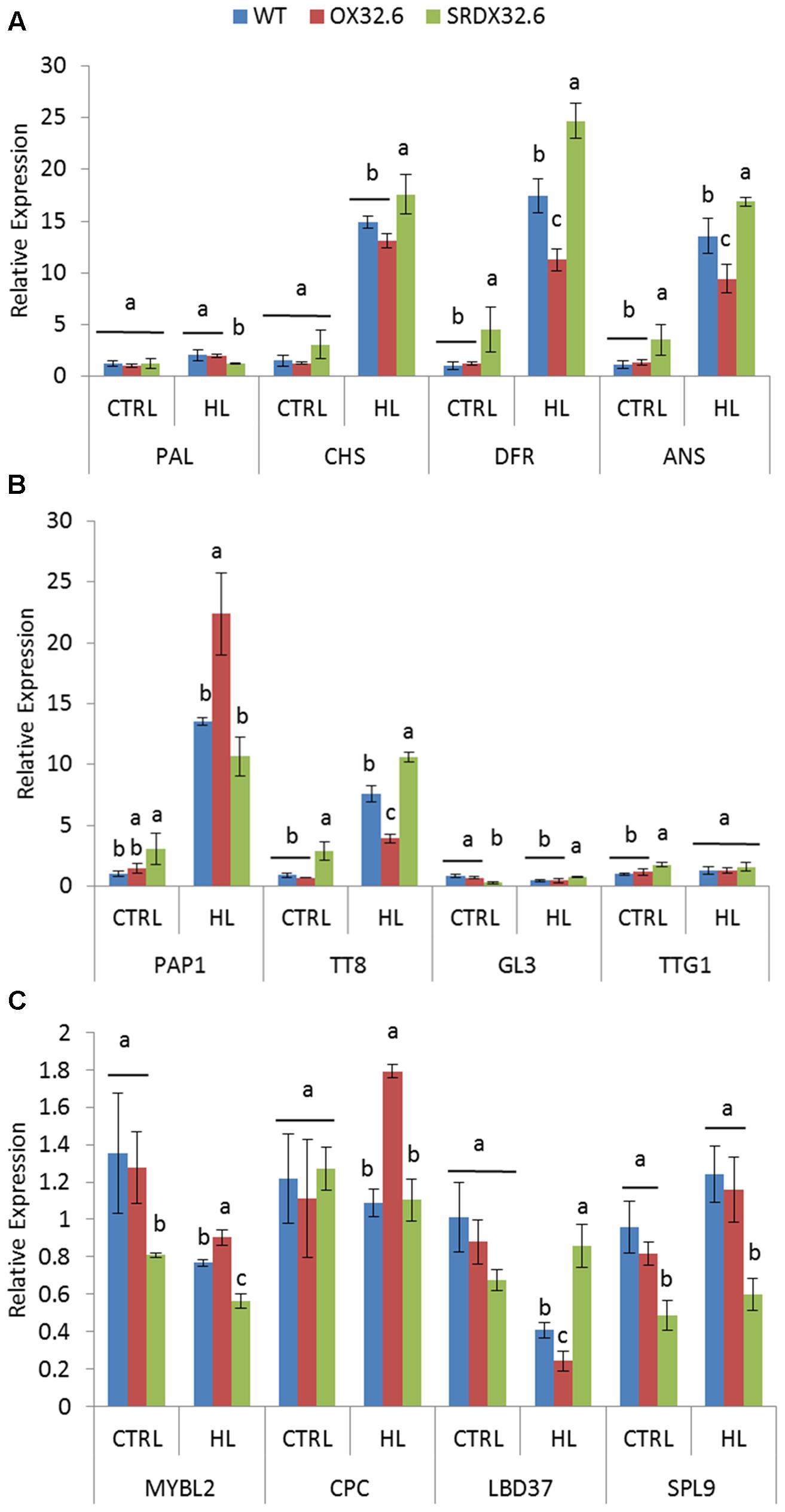
FIGURE 4. Expression analysis of anthocyanin biosynthesis and regulatory genes in response to high light stress. (A) qRT-PCR analysis of anthocyanin biosynthesis genes (ABGs). (B) qRT-PCR analysis of positive regulators (transcriptional activators) of anthocyanin biosynthesis. (C) qRT-PCR analysis of negative regulators (transcriptional repressors) of anthocyanin biosynthesis. Seventeen-day-old WT and ANAC032 transgenic lines were treated with optimal/control (150 μmol m-2 s-1) and high light (∼450 μmol m-2 s-1) for 4 days. Data represent mean relative expression values (±SD). ACT7 was used as internal reference gene. Within each treatment, bars with different letters are not statistically similar to each other according one way ANOVA LSD test (P < 0.05).
ANAC032 Negatively Regulates MeJA and ABA Signaling Required for Stress-Induced Anthocyanin Accumulation
Stress-regulated plant hormones such as ABA and JA are also known to induce anthocyanin biosynthesis in plants, suggesting that stresses can induce anthocyanin accumulation through signals integrated by these stress hormones (Loreti et al., 2008; Shan et al., 2009). Since expression of ANAC032 is induced by MeJA and ABA (Mahmood et al., 2016), we hypothesized that ANAC032 may inhibit JA- or ABA-inductive effects on anthocyanin production. First, we studied the effect of MeJA on anthocyanin biosynthesis. Results show that after 12 days of growth on plates supplemented with 0, 25, and 50 μM MeJA, wild-type seedlings accumulated anthocyanins in a concentration dependent manner (Figures 5A,B). The effect of MeJA on the accumulation of anthocyanins was reduced in ANAC032 overexpression lines compared to wild-type (Figures 5A,B). In contrast, ANAC032 SRDX lines accumulated significantly higher levels of anthocyanin pigments compared to wild-type seedlings (Figures 5A,B). Similar findings were observed when ANAC032 transgenic lines were treated with ABA (Figures 5C,D). Taken together these results suggest that ANAC032 negatively regulates ABA and JA signals that lead to anthocyanin accumulation.
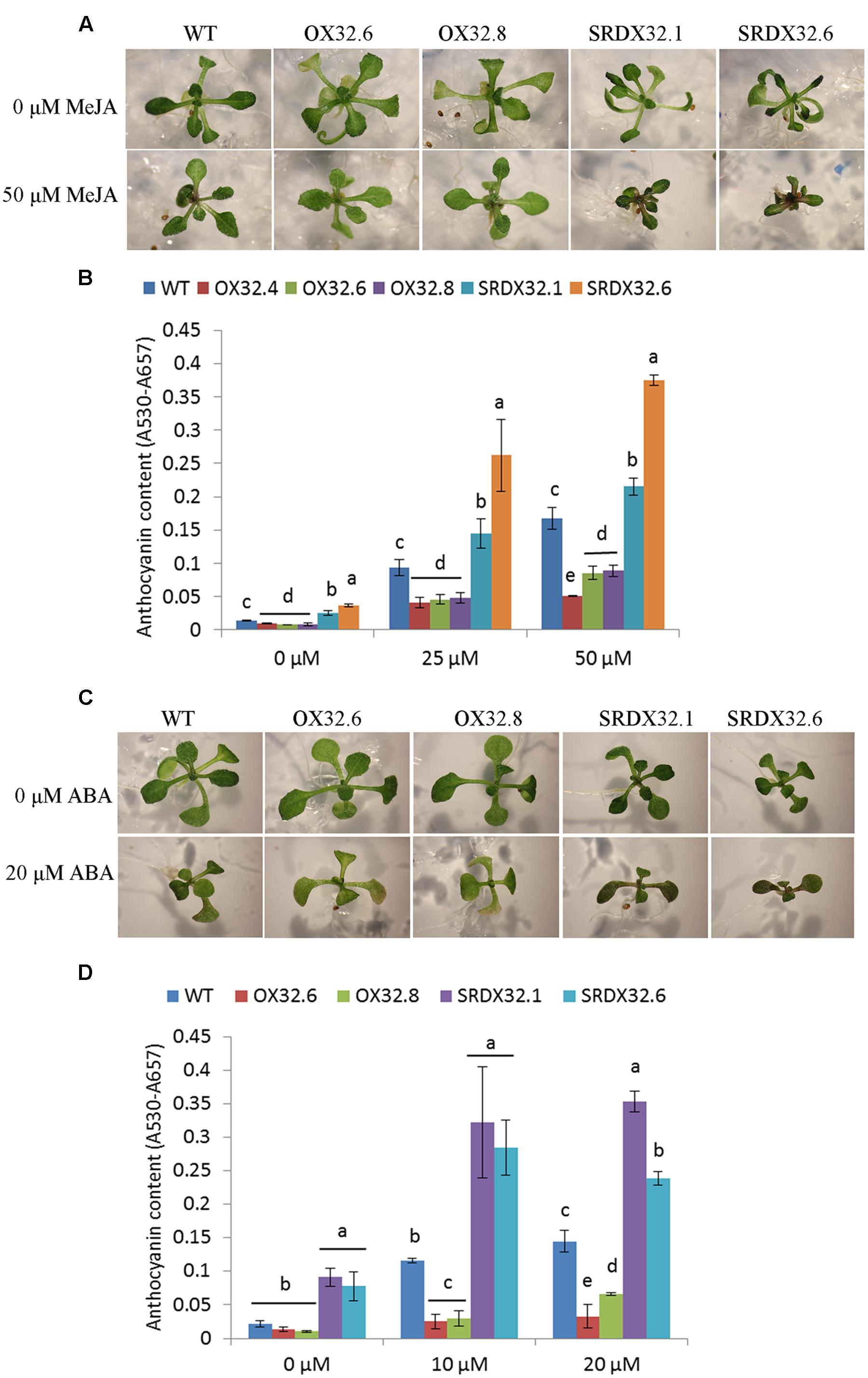
FIGURE 5. Hormonal regulation of anthocyanin accumulation. (A) Phenotypic response of WT and ANAC032 transgenic lines for anthocyanin biosynthesis in response to MeJA treatment after 15 days of growth under long-day conditions. (B) Anthocyanin content in response to MeJA treatment. Seeds were germinated and grown on half-strength MS agar plates (1% sucrose) supplemented with 0, 25, and 50 μM MeJA for 12 days under long-day conditions. Twenty seedlings were pooled in each replicate for each genotype to analyze anthocyanin content. (C) Phenotypic response of WT and ANAC032 transgenic lines for anthocyanin biosynthesis in response to ABA treatment. (D) Anthocyanin content in response to ABA treatment. Eight-day-old seedlings of wild-type and ANAC032 transgenic lines were exposed to 0, 10, and 20 μM ABA and were grown under long-day condition for 4 days. Eight seedlings were pooled in each replicate for each genotype. Data represent mean values (±SD, n = 3). Within each treatment, bars with different letters in (B,D) are not similar to each other statistically according to one way ANOVA LSD test (P < 0.05).
ANAC032 Negatively Regulates Anthocyanin Biosynthesis in Response to Salinity Stress
Given the inhibitory effect of ANAC032 on the ABA- and MeJA-induced anthocyanin accumulation, we speculated that ANAC032 would also negatively regulate anthocyanin production in response to stresses that trigger ABA and JA biosynthesis, such as salinity stress (Zhu, 2002; Walia et al., 2006). Consistent with this, challenging WT and ANAC032 transgenic lines with 200 mM saline solutions (NaCl) showed that ANAC032 overexpression lines accumulated less anthocyanin contents whereas the SRDX lines accumulate more compared to wild-type, suggesting that ANAC032 can also reduce anthocyanin production in response to salinity stress (Figure 6A). The altered accumulation of anthocyanin pigment in ANAC032 transgenic lines was further confirmed by analyzing the expression of anthocyanin biosynthesis and regulatory genes. Since no significant differences in anthocyanin content were found between genotypes under control conditions, the expression analysis was only performed on plants exposed to salinity stress. Consistent with these biochemical changes, transcript levels of all the ABGs analyzed (PAL, CHS, DFR and ANS) were significantly reduced in the overexpression line compared to wild-type (Figure 6B) and the opposite expression pattern in the SRDX line, particularly for DFR and ANS (Figure 6B). In accordance with the reduced transcript levels of these biosynthesis genes, expression of PAP1, TT8 and GL3, which constitute the transcriptional complex to activate the expression of ABGs, was also drastically reduced in the overexpression line compared to wild-type (Figure 6C). The expression of these genes, including TTG1, in contrast, was significantly induced in the SRDX line (Figure 6C). Similar to what was observed in the case of HL stress; the most drastic differences were observed for TT8 in the ANAC032 transgenic lines (Figure 6C).
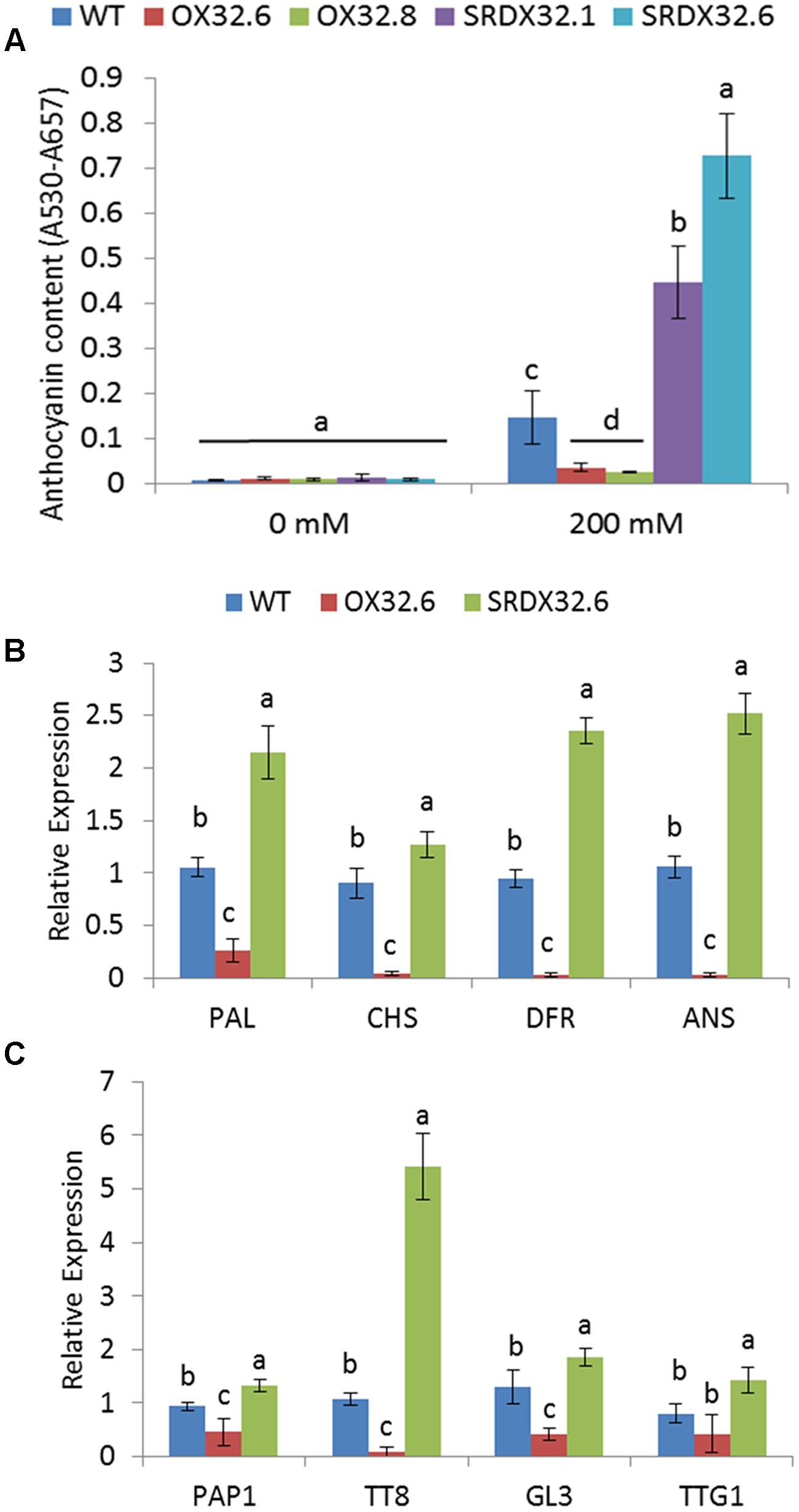
FIGURE 6. Salinity-induced anthocyanin biosynthesis in ANAC032 transgenic lines. (A) Anthocyanin content in response to salinity stress. Three-week-old plants of WT and ANAC032 transgenic lines were treated with 200 mM NaCl for 2 weeks. Data represent values from three biological replicates (±SD). Bars with same letters are not significantly different from each other according to one way ANOVA LSD test (P < 0.05). (B) qRT-PCR analysis of ABGs in leaves of plants subjected to salinity stress. (C) qRT-PCR analysis of transcription factors that positively regulate the expression of ABGs plants subjected to salinity stress. ACT7 was used as internal control. For each gene, bars with same letters are not significantly different from each other according to one way ANOVA LSD test (P < 0.05).
Lignin Biosynthesis Is Not Regulated by ANAC032
ANAC032 is predominantly expressed in roots under normal growth conditions (Figures 1B,C), suggesting that its expression may influence root development. Analysis of primary root growth showed that although both wild-type and ANAC032 overexpression lines had comparable primary root growth, the chimeric repressors produced significantly shorter roots under normal growth conditions (Figures 7A,B). Since lignin and anthocyanin are produced from same phenylpropanoid pathway and many mutants exhibiting ectopic lignification produce shorter roots (Abdulrazzak et al., 2006; Lucas et al., 2011) we anticipated that ANAC032 may affect lignin biosynthesis in the roots. Phloroglucinol-HCL staining of roots showed that both ANAC032 overexpression and SRDX lines had lignification patterns similar to wild-type as no ectopic lignification was observed in any of the transgenic line (Figure 7C). These results show that ANAC032 is unlikely involved in the regulation of lignin biosynthesis, and that the retarded root growth in SRDX032 lines could be due to reduced cell division and/or expansion.
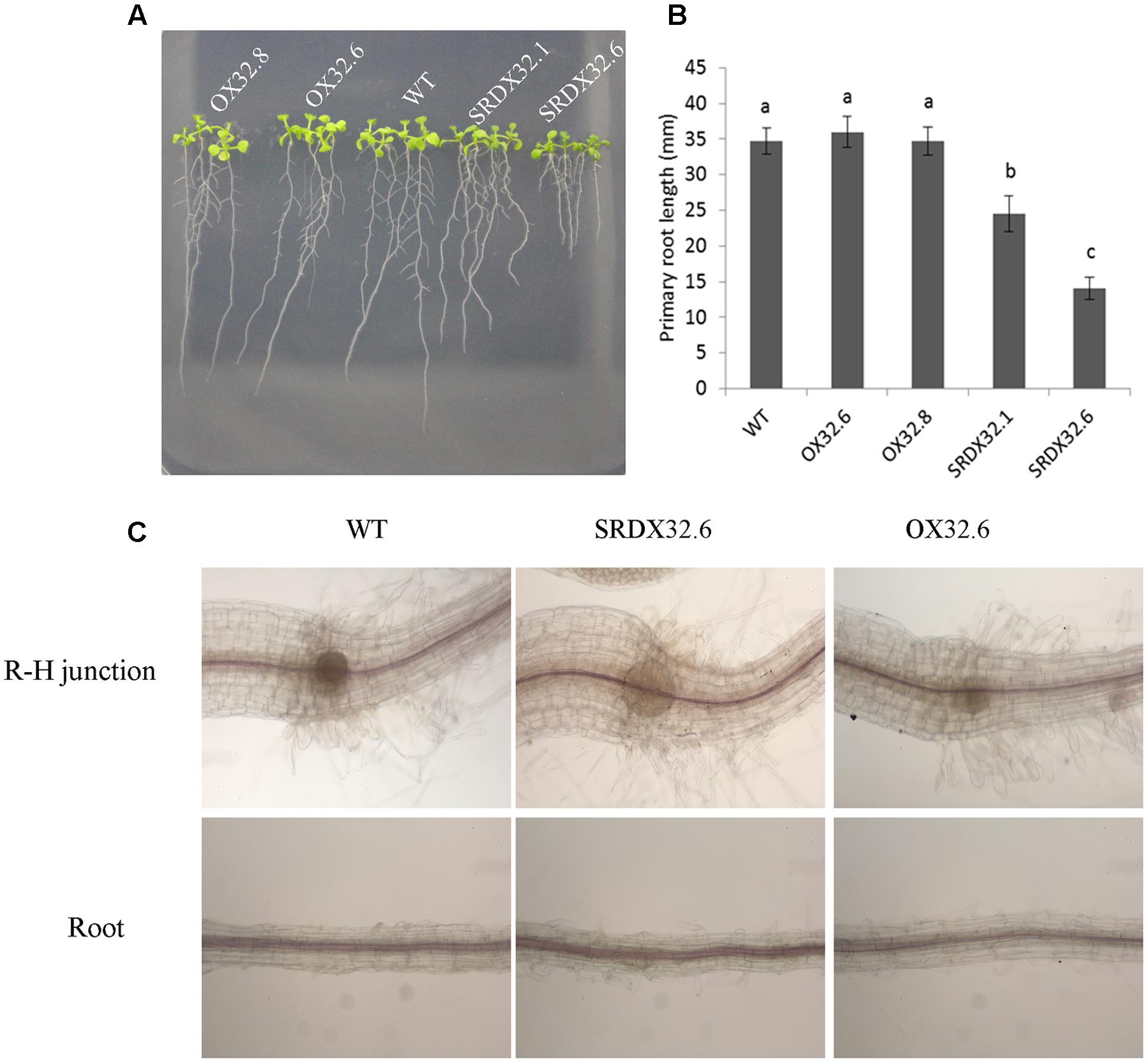
FIGURE 7. (A) Phenotype of 10-day-old seedlings grown on half-strength MS agar plates supplemented with 1% sucrose under long-day condition. (B) Primary root lengths of 7-day-old seedlings of wild-type and ANAC032 transgenic lines. Data represent values from three biological replicates. Each replicate included 20 roots. Bars with same letters are not statistically different from each other according to one way ANOVA-LSD test (P < 0.05). (C) Analysis of lignification pattern in WT and ANAC032 transgenic lines. Phloroglucinol-HCl staining of lignin in roots and hypocotyl regions of 7-day-old seedlings. Roots were analyzed under a light microscope (10 X magnification). (R-H, root-hypocotyl junction).
Discussion
In plants, spatio-temporal regulation of anthocyanin biosynthesis takes place in a highly ordered manner (Martins et al., 2013; Xu et al., 2013). A marked increase in the accumulation of anthocyanin pigment has been observed during a variety of biotic and abiotic stresses and they have been proposed to have a protective role during stress conditions (Chalker-Scott, 1999). While the enzymatic pathway involved in the production of anthocyanins is largely known, the transcriptional regulation of their biosynthetic pathway is not fully understood. In this study, we investigated the role of ANAC032 in the regulation of anthocyanin production in A. thaliana.
A number of NAC TFs have been implicated in mediating stress responses in A. thaliana, yet only JUB1 and ANAC078 have been implicated in the regulation of anthocyanin production (Morishita et al., 2009; Wu et al., 2012). An increase in the expression of late ABGs (e.g., DFR and LDOX/ANS) was observed in transgenic plants overexpressing ANAC078. Similarly, TT2, TT8 and TTG1 that form the MBW complex were also induced in the ANAC078 lines (Morishita et al., 2009). JUB1 expression, however, was shown to have a negative impact on the expression of ABGS and MBWs in Arabidopsis (Wu et al., 2012). Although, JUB1 was shown to be important for survival under stress conditions, such as salinity and oxidative stress (Wu et al., 2012), the regulation of anthocyanin biosynthesis has not been studied in detail under these stress conditions. We show that in early growth stages, ANAC032 had a negative impact on production of anthocyanin pigment which was lost in the later stage of plant growth (Supplementary Figure S1). However, under different stress conditions such as HL, oxidative stress and salinity stress, there is a clear negative effect on the production of anthocyanin pigments in plants overexpressing ANAC032 (Figures 2, 3, and 6). At the molecular level, we also observed a reduction in the transcript levels of late ABGs (such as DFR and LDOX/ANS) in an ANAC032 overexpression line and induction of those genes in the ANAC032-SRDX line compared to wild-type (Figure 4A). In the context of TFs forming the MBW complex and that regulate the expression of ABGs, the expression of TT8 was significantly reduced in the overexpression line and induced in the SRDX line compared to wild-type, suggesting that a negative impact on anthocyanin biosynthesis may be due to a reduction in TT8 expression which eventually affects the assembly of the MBW complex (Figure 4B). TT8 activates the expression of DFR by directly binding with its promoter region (Baudry et al., 2006). Despite reduced accumulation of anthocyanin pigments, the ANAC032 overexpression line had significantly higher transcript levels of PAP1 compared to wild-type, yet they accumulate less anthocyanin. Given the reduced transcript levels of TT8 in overexpression plants under HL, it is likely that increased PAP1 expression is a response to lower levels of TT8 to compensate for its deficiency. This proposition is in agreement with previous findings proposing that PAP1 and its MYB homolog, TT2, can activate the expression of TT8 (Baudry et al., 2006).
Studies have shown that TT8 is mainly regulated at the transcriptional level, and that TT8 can regulate its own expression (Nesi et al., 2000; Baudry et al., 2006). Furthermore, new mechanisms for the negative transcriptional regulation of TT8 and DFR have also been elucidated with the identification of repressors of anthocyanin biosynthesis. For example, AtMYBL2 has been shown to interact with TT8 (Matsui et al., 2008) along with GL3 and EGL3 (Dubos et al., 2008) resulting in the formation of an inactive complex, L2BW, which competes with the active MBW complex to regulate ABGs (Dubos et al., 2008; Matsui et al., 2008). Similarly, SPL9, another repressor of anthocyanin biosynthesis, interferes with the formation of the MBW complex by competing with TT8 for its interaction with PAP1 (Gou et al., 2011). Since ANAC032 overexpressors and chimeric repressors exhibited contrasting phenotypes with regard to anthocyanin accumulation, it is highly unlikely that ANAC032 directly represses the expression of DFR and TT8. The repressive effect on the expression of these genes, therefore, appears to be indirect which could be due to another factor that represses the activation of these genes. Consistent with this is that ANAC032-SRDX plants had reduced expression of AtMYBL2 under both control and HL whereas the overexpression line presents increased transcription of AtMYBL2 compared to wild-type, particularly under HL stress (Figure 4C). The altered transcript levels of AtMYBL2 may, therefore, be responsible for the differential expressions of TT8 and DFR in the ANAC032 transgenic lines under HL stress. Moreover, the expression of SPL9 was only reduced in the ANAC032-SRDX line under both control and HL conditions. The three members of the Lateral Organ Boundary Domain family of TF, LBD37, LBD38 and LBD39, negatively regulate anthocyanin biosynthesis in response to HL, high sucrose and nitrogen stress conditions in A. thaliana (Rubin et al., 2009). In our study, the expression of LBD37 was repressed by HL in wild-type plants (Figure 4C), however, it was more drastically reduced in the overexpression line. Since these TFs directly repress the expression of PAP1/2 (Rubin et al., 2009), a significant increase in the expression of PAP1 was observed in the overexpression line under HL stress compared to wild-type. Despite the obvious induction of PAP1 in overexpression line, it was unable to overcome the negative impact on the expression of TT8 and DFR in ANAC032 overexpression line under HL.
Most abiotic stresses have been shown to cause the accumulation of soluble sugars (glucose, fructose, and sucrose) in several plant species (Kempa et al., 2008; Krasensky and Jonak, 2012). In grape skin, production of anthocyanin has also been correlated with sugar accumulation (Boss et al., 1996). Moreover, exogenous sucrose treatment triggers the induction of anthocyanin production in Arabidopsis (Teng et al., 2005; Solfanelli et al., 2006). Hyper-accumulation of soluble sugars during stress conditions, therefore, may be not only important for acclimation responses but may also trigger anthocyanin biosynthesis. Consistent with this hypothesis is that the suc2 mutant which lacks a functional phloem-loading Suc-proton symporter-, not only accumulates increased anthocyanin pigments but also contains higher levels of soluble sugars (Lloyd and Zakhleniuk, 2004). Similarly, another Arabidopsis mutant, phosphoglucomutase (pgm), which accumulates soluble sugars, had higher levels of anthocyanin pigments compared to wild-type and had a drastic increase in the expression of PAP1, 4CL, CHS, CHI, F3H, DFR, and LDOX (Solfanelli et al., 2006). We show that ANAC032 is also induced by sucrose treatment and altering ANAC032 expression negatively regulated anthocyanin production upon treatment with exogenous sucrose in a concentration dependent manner (Figure 1). Based on these findings, it is likely that ANAC032 represses anthocyanin production by altering sugar metabolism. In agreement with this, is that the ANAC032 overexpression lines, which accumulate less anthocyanin, had reduced levels of soluble sugars and that the SRDX032 lines accumulated more pigments compared to wild-type, particularly under HL.
A number of studies have shown that plants produce hormones like ABA and JA when exposed to different abiotic stress conditions such as HL, drought and salinity (Wang et al., 2001; Walia et al., 2006; Galvez-Valdivieso et al., 2009; Ramel et al., 2013). Production of these stress hormones evokes various adaptive responses in plants, including anthocyanin production. We have shown that ANAC032 expression is also induced by ABA and MeJA treatment. Similar to what was observed under HL and salinity stress, ANAC032 overexpression lines accumulated significantly less anthocyanins whereas SRDX lines accumulated more pigments upon ABA and MeJA treatment compared to wild-type (Figure 5). These findings suggest that ANAC032 represses anthocyanin biosynthesis through a negative regulation of ABA and JA signaling. The altered anthocyanin production in response to ABA and MeJA may also involve differential accumulation of soluble sugars as was observed in the case of HL stress. Several lines of evidence support this notion; (i) the ABA and MeJA-induced anthocyanin biosynthesis in A. thaliana was shown to be dependent upon the presence of sucrose in the nutrient medium (Loreti et al., 2008); (ii) many ABA-, JA- and stress-inducible genes are also co-regulated by sugars (Reinbothe et al., 1994; Sadka et al., 1994); and (iii) exogenous treatment with ABA results in the accumulation of soluble sugars, thus mimicking a stress response (Hiratsuka et al., 2001; Nagao et al., 2005).
In summary, we have investigated the role of ANAC032 in the regulation of anthocyanin biosynthesis in A. thaliana. Induced expression of ANAC032 represses anthocyanin accumulation and alters the expression of anthocyanin biosynthesis (DFR, ANS/LDOX) and regulatory genes (TT8) under stresses, more likely through modulated expression of the negative regulators of anthocyanin biosynthesis, AtMYBL2 and SPL9. However, given that the identification of the complete network of negative regulators of anthocyanin biosynthesis in Arabidopsis is not fully known, modulation of yet other negative regulators by ANAC032 can not be ruled out. Our data also suggest that the differential accumulation of anthocyanin is due to altered ABA, MeJA and sugar signaling.
Author Contributions
KM and SR conceived the project. KM, AE-K, JC, and SR designed the experiments. KM and ZX performed the experiments. KM analyzed the data. KM, JC, and SR wrote the manuscript. All authors read and approved the final version of the manuscript.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgment
This research was supported by funding from Natural Sciences and Engineering Research Council of Canada (NSERC) to SR.
Supplementary Material
The Supplementary Material for this article can be found onlineat: http://journal.frontiersin.org/article/10.3389/fpls.2016.01548
Abbreviations
3-AT, 3-aminotriazole; ABA, abscisic acid; ATAF1,2 and CUC2 (cup-shaped cotyledon); GUS, β-glucuronidase; HL, high light; MeJA, methyl jasmonate; NAC, NAM (no apical meristem).
References
Abdulrazzak, N., Pollet, B., Ehlting, J., Larsen, K., Asnaghi, C., Ronseau, S., et al. (2006). A coumaroyl-ester-3-hydroxylase insertion mutant reveals the existence of nonredundant meta-hydroxylation pathways and essential roles for phenolic precursors in cell expansion and plant growth. Plant Physiol. 140, 30–48. doi: 10.1104/pp.105.069690
Albert, N. W., Lewis, D. H., Zhang, H., Irving, L. J., Jameson, P. E., and Davies, K. M. (2009). Light-induced vegetative anthocyanin pigmentation in Petunia. J. Exp. Bot. 60, 2191–2202. doi: 10.1093/jxb/erp097
Balazadeh, S., Siddiqui, H., Allu, A. D., Matallana-Ramirez, L. P., Caldana, C., Mehrnia, M., et al. (2010). A gene regulatory network controlled by the NAC transcription factor ANAC092/AtNAC2/ORE1 during salt-promoted senescence. Plant J. 62, 250–264. doi: 10.1111/j.1365-313X.2010.04151.x
Baudry, A., Caboche, M., and Lepiniec, L. (2006). TT8 controls its own expression in a feedback regulation involving TTG1 and homologous MYB and bHLH factors, allowing a strong and cell-specific accumulation of flavonoids in Arabidopsis thaliana. Plant J. 46, 768–779. doi: 10.1111/j.1365-313X.2006.02733.x
Boss, P. K., Davies, C., and Robinson, S. P. (1996). Anthocyanin composition and anthocyanin pathway gene expression in grapevine sports differing in berry skin colour. Aust. J. Grape Wine Res. 2, 163–170. doi: 10.1111/j.1755-0238.1996.tb00104.x
Buysse, J. A. N., and Merckx, R. (1993). An improved colorimetric method to quantify sugar content of plant tissue. J. Exp. Bot. 44, 1627–1629. doi: 10.1093/jxb/44.10.1627
Carey, C. C., Strahle, J. T., Selinger, D. A., and Chandler, V. L. (2004). Mutations in the pale aleurone color1 regulatory gene of the Zea mays anthocyanin pathway have distinct phenotypes relative to the functionally similar TRANSPARENT TESTA GLABRA1 gene in Arabidopsis thaliana. Plant Cell 16, 450–464. doi: 10.1105/tpc.018796.1
Castellarin, S. D., Pfeiffer, A., Sivilotti, P., Degan, M., Peterlunger, E., and Di Gaspero, G. (2007). Transcriptional regulation of anthocyanin biosynthesis in ripening fruits of grapevine under seasonal water deficit. Plant Cell Environ. 30, 1381–1399. doi: 10.1111/j.1365-3040.2007.01716.x
Chalker-Scott, L. (1999). Environmental significance of anthocyanins in plant stress responses. Photochem. Photobiol. 70, 1–9. doi: 10.1111/j.1751-1097.1999.tb01944.x
Christie, P. J., Alfenito, M. R., and Walbot, V. (1994). Impact of low-temperature stress on general phenylpropanoid and anthocyanin pathways: enhancement of transcript abundance and anthocyanin pigmentation in maize seedlings. Planta 194, 541–549. doi: 10.1007/BF00714468
Cominelli, E., Gusmaroli, G., Allegra, D., Galbiati, M., Wade, H. K., Jenkins, G. I., et al. (2008). Expression analysis of anthocyanin regulatory genes in response to different light qualities in Arabidopsis thaliana. J. Plant Physiol. 165, 886–894. doi: 10.1016/j.jplph.2007.06.010
Cui, L. G., Shan, J. X., Shi, M., Gao, J. P., and Lin, H. X. (2014). The miR156-SPL9-DFR pathway coordinates the relationship between development and abiotic stress tolerance in plants. Plant J. 80, 1108–1117. doi: 10.1111/tpj.12712
Dubey, R. S., and Singh, A. K. (1999). Salinity induces accumulation of soluble sugars and alters the activity of sugar metabolising enzymes in rice plants. Biol. Plant. 42, 233–239. doi: 10.1023/A:1002160618700
Dubos, C., Le Gourrierec, J., Baudry, A., Huep, G., Lanet, E., Debeaujon, I., et al. (2008). MYBL2 is a new regulator of flavonoid biosynthesis in Arabidopsis thaliana. Plant J. 55, 940–953. doi: 10.1111/j.1365-313X.2008.03564.x
Duval, M., Hsieh, T. F., Kim, S. Y., and Thomas, T. L. (2002). Molecular characterization of AtNAM: a member of the Arabidopsis NAC domain superfamily. Plant Mol. Biol. 50, 237–248. doi: 10.1023/A:1016028530943
Eryilmaz, F. (2006). The relationships between salt stress and anthocyanin content in higher plants. Biotechnol. Biotechnol. Equip. 20, 47–52. doi: 10.1080/13102818.2006.10817303
Galvez-Valdivieso, G., Fryer, M. J., Lawson, T., Slattery, K., Truman, W., Smirnoff, N., et al. (2009). The high light response in Arabidopsis involves ABA signaling between vascular and bundle sheath cells. Plant Cell 21, 2143–2162. doi: 10.1105/tpc.108.061507
Gonzalez, A., Zhao, M., Leavitt, J. M., and Lloyd, A. M. (2008). Regulation of the anthocyanin biosynthetic pathway by the TTG1/bHLH/Myb transcriptional complex in Arabidopsis seedlings. Plant J. 53, 814–827. doi: 10.1111/j.1365-313X.2007.03373.x
Gou, J., Felippes, F. F., Liu, C., Weigel, D., and Wang, J. (2011). Negative regulation of anthocyanin biosynthesis in Arabidopsis by a miR156-targeted SPL transcription factor. Plant Cell 23, 1512–1522. doi: 10.1105/tpc.111.084525
Guo, Y., and Gan, S. (2006). AtNAP, a NAC family transcription factor, has an important role in leaf senescence. Plant J. 46, 601–612. doi: 10.1111/j.1365-313X.2006.02723.x
Hiratsuka, S., Onodera, H., Kawai, Y., Kubo, T., Itoh, H., and Wada, R. (2001). ABA and sugar effects on anthocyanin formation in grape berry cultured in vitro. Sci. Hortic. 90, 121–130. doi: 10.1016/S0304-4238(00)00264-8
Hughes, N. M., Neufeld, H. S., and Burkey, K. O. (2005). Functional role of anthocyanins in high-light winter leaves of the evergreen herb Galax urceolata. New Phytol. 168, 575–587. doi: 10.1111/j.1469-8137.2005.01546.x
Jackson, D., Roberts, K., and Martin, C. (1992). Temporal and spatial control of expression of anthocyanin biosynthetic genes in developing flowers of Antirrhinum majus. Plant J. 2, 425–434. doi: 10.1111/j.1365-313X.1992.00425.x
Jiang, C., Gao, X., Liao, L., Harberd, N. P., and Fu, X. (2007). Phosphate starvation root architecture and anthocyanin accumulation responses are modulated by the gibberellin-DELLA signaling pathway in Arabidopsis. Plant Physiol. 145, 1460–1470. doi: 10.1104/pp.107.103788
Kempa, S., Krasensky, J., Dal Santo, S., Kopka, J., and Jonak, C. (2008). A central role of abscisic acid in stress-regulated carbohydrate metabolism. PLoS ONE 3:e3935. doi: 10.1371/journal.pone.0003935
Kilian, J., Whitehead, D., Horak, J., Wanke, D., Weinl, S., Batistic, O., et al. (2007). The AtGenExpress global stress expression data set: protocols, evaluation and model data analysis of UV-B light, drought and cold stress responses. Plant J. 50, 347–363. doi: 10.1111/j.1365-313X.2007.03052.x
Ko, J.-H., Yang, S. H., Park, A. H., Lerouxel, O., and Han, K.-H. (2007). ANAC012, a member of the plant-specific NAC transcription factor family, negatively regulates xylary fiber development in Arabidopsis thaliana. Plant J. 50, 1035–1048. doi: 10.1111/j.1365-313X.2007.03109.x
Krasensky, J., and Jonak, C. (2012). Drought, salt, and temperature stress-induced metabolic rearrangements and regulatory networks. J. Exp. Bot. 63, 1593–1608. doi: 10.1093/jxb/err460
Kubo, M., Udagawa, M., Nishikubo, N., Horiguchi, G., Yamaguchi, M., Ito, J., et al. (2005). Transcription switches for protoxylem and metaxylem vessel formation. Genes Dev. 19, 1855–1860. doi: 10.1101/gad.1331305
Lee, S., Seo, P. J., Lee, H.-J., and Park, C.-M. (2012). A NAC transcription factor NTL4 promotes reactive oxygen species production during drought-induced leaf senescence in Arabidopsis. Plant J. 70, 831–844. doi: 10.1111/j.1365-313X.2012.04932.x
Lichtenthaler, H. K., Buschmann, C., Doll, M., Fietz, H. J., Bach, T., Kozel, U., et al. (1981). Photosynthetic activity, chloroplast ultrastructure, and leaf characteristics of high-light and low-light plants and of sun and shade leaves. Photosynth. Res. 2, 115–141. doi: 10.1007/BF00028752
Lloyd, J. C., and Zakhleniuk, O. V. (2004). Responses of primary and secondary metabolism to sugar accumulation revealed by microarray expression analysis of the Arabidopsis mutant, pho3. J. Exp. Bot. 55, 1221–1230. doi: 10.1093/jxb/erh143
Loreti, E., Povero, G., Novi, G., Solfanelli, C., Alpi, A., and Perata, P. (2008). Gibberellins, jasmonate and abscisic acid modulate the sucrose-induced expression of anthocyanin biosynthetic genes in Arabidopsis. New Phytol. 179, 1004–1016. doi: 10.1111/j.1469-8137.2008.02511.x
Lotkowska, M. E., Tohge, T., Fernie, A. R., Xue, G.-P., Balazadeh, S., and Mueller-Roeber, B. (2015). The Arabidopsis transcription factor MYB112 promotes anthocyanin formation during salinity and under high light stress. Plant Physiol. 169, 1862–1880. doi: 10.1104/pp.15.00605
Lucas, M., Swarup, R., Paponov, I. A., Swarup, K., Casimiro, I., Lake, D., et al. (2011). SHORT-ROOT regulates primary, lateral, and adventitious root development in Arabidopsis. Plant Physiol. 155, 384–398. doi: 10.1104/pp.110.165126
Mahmood, K., El-Kereamy, A., Kim, S.-H., Nambara, E., and Rothstein, S. J. (2016). ANAC032 positively regulates age-dependent and stress-induced senescence in Arabidopsis thaliana. Plant Cell Physiol. doi: 10.1093/pcp/pcw120 [Epub ahead of print].
Martins, T. R., Berg, J. J., Blinka, S., Rausher, M. D., and Baum, D. A. (2013). Precise spatio-temporal regulation of the anthocyanin biosynthetic pathway leads to petal spot formation in Clarkia gracilis (Onagraceae). New Phytol. 197, 958–969. doi: 10.1111/nph.12062
Matsui, K., Umemura, Y., and Ohme-Takagi, M. (2008). AtMYBL 2, a protein with a single MYB domain, acts as a negative regulator of anthocyanin biosynthesis in Arabidopsis. Plant J. 55, 954–967. doi: 10.1111/j.1365-313X.2008.03565.x
Mitra, P. P., and Loqué, D. (2014). Histochemical staining of Arabidopsis thaliana secondary cell wall elements. J. Vis. Exp. 87:e51381. doi: 10.3791/51381
Mitsuda, N., Iwase, A., Yamamoto, H., Yoshida, M., Seki, M., Shinozaki, K., et al. (2007). NAC transcription factors, NST1 and NST 3, are key regulators of the formation of secondary walls in woody tissues of Arabidopsis. Plant Cell 19, 270–280. doi: 10.1105/tpc.106.047043
Mitsuda, N., Seki, M., Shinozaki, K., and Ohme-takagi, M. (2005). The NAC transcription factors NST1 and NST2 of Arabidopsis regulate secondary wall thickenings and are required for anther dehiscence. Plant Cell 17, 2993–3006. doi: 10.1105/tpc.105.036004.1
Morishita, T., Kojima, Y., Maruta, T., Nishizawa-Yokoi, A., Yabuta, Y., and Shigeoka, S. (2009). Arabidopsis NAC transcription factor, ANAC 078, regulates flavonoid biosynthesis under high-light. Plant Cell Physiol. 50, 2210–2222. doi: 10.1093/pcp/pcp159
Nagao, M., Minami, A., Arakawa, K., Fujikawa, S., and Takezawa, D. (2005). Rapid degradation of starch in chloroplasts and concomitant accumulation of soluble sugars associated with ABA-induced freezing tolerance in the moss Physcomitrella patens. J. Plant Physiol. 162, 169–180. doi: 10.1016/j.jplph.2004.06.012
Nakabayashi, R., Yonekura-Sakakibara, K., Urano, K., Suzuki, M., Yamada, Y., Nishizawa, T., et al. (2014). Enhancement of oxidative and drought tolerance in Arabidopsis by overaccumulation of antioxidant flavonoids. Plant J. 7, 367–379. doi: 10.1111/tpj.12388
Nemie-feyissa, D., Olafsdottir, S. M., Heidari, B., and Lillo, C. (2014). Phytochemistry nitrogen depletion and small R3-MYB transcription factors affecting anthocyanin accumulation in Arabidopsis leaves. Phytochemistry 98, 34–40. doi: 10.1016/j.phytochem.2013.12.006
Nesi, N., Debeaujon, I., Jond, C., Pelletier, G., Caboche, M., and Lepiniec, L. (2000). The TT8 gene encodes a basic helix-loop-helix domain protein required for expression of DFR and BAN genes in Arabidopsis siliques. Plant Cell 12, 1863–1878. doi: 10.1105/tpc.12.10.1863
Ohashi-Ito, K., Oda, Y., and Fukuda, H. (2010). Arabidopsis VASCULAR-RELATED NAC-DOMAIN6 directly regulates the genes that govern programmed cell death and secondary wall formation during xylem differentiation. Plant Cell 22, 3461–3473. doi: 10.1105/tpc.110.075036
Peng, M., Hudson, D., Schofield, A., Tsao, R., Yang, R., Gu, H., et al. (2008). Adaptation of Arabidopsis to nitrogen limitation involves induction of anthocyanin synthesis which is controlled by the NLA gene. J. Exp. Bot. 59, 2933–2944. doi: 10.1093/jxb/ern148
Procissi, A., Dolfini, S., Ronchi, A., and Tonelli, C. (1997). Light-dependent spatial and temporal expression of pigment regulatory genes in developing maize seeds. Plant Cell 9, 1547–1557. doi: 10.1105/tpc.9.9.1547
Quattrocchio, F., Verweij, W., Kroon, A., Spelt, C., Mol, J., and Koes, R. (2006). PH4 of Petunia is an R2R3 MYB protein that activates vacuolar acidification through interactions with basic-helix-loop-helix transcription factors of the anthocyanin pathway. Plant Cell 18, 1274–1291. doi: 10.1105/tpc.105.034041
Ramel, F., Ksas, B., and Havaux, M. (2013). Jasmonate: a decision maker between cell death and acclimation in the response of plants to singlet oxygen. Plant Signal. Behav. 8:e26655. doi: 10.4161/psb.26655
Reinbothe, S., Mollenhauer, B., and Reinbothe, C. (1994). JIPs and RIPs: the regulation of plant gene expression by jasmonates in response to environmental cues and pathogens. Plant Cell 6, 1197–1209. doi: 10.1105/tpc.6.9.1197
Rubin, G., Tohge, T., Matsuda, F., Saito, K., and Scheible, W.-R. (2009). Members of the LBD family of transcription factors repress anthocyanin synthesis and affect additional nitrogen responses in Arabidopsis. Plant Cell 21, 3567–3584. doi: 10.1105/tpc.109.067041
Sadka, A., DeWald, D. B., May, G. D., Park, W. D., and Mullet, J. E. (1994). Phosphate modulates transcription of soybean VspB and other sugar-inducible genes. Plant Cell 6, 737–749. doi: 10.2307/3869876
Schmitz, J., Heinrichs, L., Scossa, F., Fernie, A. R., Oelze, M. L., Dietz, K. J., et al. (2014). The essential role of sugar metabolism in the acclimation response of Arabidopsis thaliana to high light intensities. J. Exp. Bot. 65, 1619–1636. doi: 10.1093/jxb/eru027
Shan, X., Zhang, Y., Peng, W., Wang, Z., and Xie, D. (2009). Molecular mechanism for jasmonate-induction of anthocyanin accumulation in Arabidopsis. J. Exp. Bot. 60, 3849–3860. doi: 10.1093/jxb/erp223
Solfanelli, C., Solfanelli, C., Poggi, A., Poggi, A., Loreti, E., Loreti, E., et al. (2006). Sucrose-specic induction of the anthocyanin biosynthetic pathway in Arabidopsis. Plant Physiol. 140, 637–646. doi: 10.1104/pp.105.072579
Stewart, A. J., Chapman, W., Jenkins, G. I., Graham, I., Martin, T., and Crozier, A. (2001). The effect of nitrogen and phosphorus deficiency on flavonol accumulation in plant tissues. Plant, Cell Environ. 24, 1189–1197. doi: 10.1046/j.1365-3040.2001.00768.x
Teng, S., Keurentjes, J., Bentsink, L., Koornneef, M., and Smeekens, S. (2005). Sucrose-specific induction of anthocyanin biosynthesis in Arabidopsis requires the MYB75/PAP1 gene. Plant Physiol. 139, 1840–1852. doi: 10.1104/pp.105.066688.1840
Vanderauwera, S., Zimmermann, P., Rombauts, S., Vandenabeele, S., Langebartels, C., Gruissem, W., et al. (2005). Genome-wide analysis of hydrogen peroxide-regulated gene expression in Arabidopsis reveals a high light-induced transcriptional cluster involved in anthocyanin biosynthesis. Plant Physiol. 139, 806–821. doi: 10.1104/pp.105.065896
Viola, I. L., Camoirano, A., and Gonzalez, D. H. (2016). Redox-dependent modulation of anthocyanin biosynthesis by the TCP transcription factor TCP15 during exposure to high light intensity conditions in Arabidopsis. Plant Physiol. 170, 74–85. doi: 10.1104/pp.15.01016
Walia, H., Wilson, C., Wahid, A., Condamine, P., Cui, X., and Close, T. J. (2006). Expression analysis of barley (Hordeum vulgare L.) during salinity stress. Funct. Integr. Genomics 6, 143–156. doi: 10.1007/s10142-005-0013-0
Walker, A. R., Davison, P. A., Bolognesi-Winfield, A. C., James, C. M., Srinivasan, N., Blundell, T. L., et al. (1999). The TRANSPARENT TESTA GLABRA1 locus, which regulates trichome differentiation and anthocyanin biosynthesis in Arabidopsis, encodes a WD40 repeat protein. Plant Cell 11, 1337–1349. doi: 10.1105/tpc.11.7.1337
Wang, X. Q., Ullah, H., Jones, A. M., and Assmann, S. M. (2001). G protein regulation of ion channels and abscisic acid signaling in Arabidopsis guard cells. Science 292, 2070–2072. doi: 10.1126/science.1059046
Weiss, D. (2000). Regulation of flower pigmentation and growth: multiple signaling pathways control anthocyanin synthesis in expanding petals. Physiol. Plant 110, 152–157. doi: 10.1034/j.1399-3054.2000.110202.x
Wu, A., Allu, A. D., Garapati, P., Siddiqui, H., Dortay, H., Zanor, M.-I., et al. (2012). JUNGBRUNNEN 1, a reactive oxygen species-responsive NAC transcription factor, regulates longevity in Arabidopsis. Plant Cell 24, 482–506. doi: 10.1105/tpc.111.090894
Xu, W., Grain, D., Le Gourrierec, J., Harscoët, E., Berger, A., Jauvion, V., et al. (2013). Regulation of flavonoid biosynthesis involves an unexpected complex transcriptional regulation of TT8 expression, in Arabidopsis. New Phytol. 198, 59–70. doi: 10.1111/nph.12142
Yamane, T., Seok, T. J., Goto-Yamamoto, N., Koshita, Y., and Kobayashi, S. (2006). Effects of temperature on anthocyanin biosynthesis in grape berry skins. Am. J. Enol. Vitic. 57, 54–59.
Yang, S.-D., Seo, P. J., Yoon, H.-K., and Park, C.-M. (2011). The Arabidopsis NAC transcription factor VNI2 integrates abscisic acid signals into leaf senescence via the COR/RD genes. Plant Cell 23, 2155–2168. doi: 10.1105/tpc.111.084913
Zhong, R., Lee, C., Zhou, J., McCarthy, R. L., and Ye, Z. H. (2008). A battery of transcription factors involved in the regulation of secondary cell wall biosynthesis in Arabidopsis. Plant Cell 20, 2763–2782. doi: 10.1105/tpc.108.061325
Zhu, H., Fitzsimmons, K., Khandelwal, A., and Kranz, R. G. (2009). CPC, a single-repeat R3 MYB, is a negative regulator of anthocyanin biosynthesis in Arabidopsis. Mol. Plant 2, 790–802. doi: 10.1093/mp/ssp030
Keywords: ANAC032, anthocyanin biosynthesis, Arabidopsis thaliana, high light, salinity, oxidative stress
Citation: Mahmood K, Xu Z, El-Kereamy A, Casaretto JA and Rothstein SJ (2016) The Arabidopsis Transcription Factor ANAC032 Represses Anthocyanin Biosynthesis in Response to High Sucrose and Oxidative and Abiotic Stresses. Front. Plant Sci. 7:1548. doi: 10.3389/fpls.2016.01548
Received: 19 July 2016; Accepted: 03 October 2016;
Published: 14 October 2016.
Edited by:
Jirong Huang, Shanghai Institutes for Biological Sciences (CAS), ChinaReviewed by:
Yongzhen Pang, Institute of Botany (CAS), ChinaAi-Xia Cheng, Shandong University, China
Copyright © 2016 Mahmood, Xu, El-Kereamy, Casaretto and Rothstein. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Steven J. Rothstein, cm90aHN0ZWlAdW9ndWVscGguY2E=
†Present address: Ashraf El-Kereamy, Division of Agriculture and Natural Resources, University of California, Cooperative Extension Kern County, Bakersfield, CA, USA