 Kiyomi Abe
Kiyomi Abe Hiroaki Ichikawa
Hiroaki Ichikawa
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
REVIEW article
Front. Plant Sci. , 21 September 2016
Sec. Plant Biotechnology
Volume 7 - 2016 | https://doi.org/10.3389/fpls.2016.01359
This article is part of the Research Topic Progress of cereal genome engineering mediated by Agrobacterium tumefaciens View all 10 articles
Identification and elucidation of functions of plant genes is valuable for both basic and applied research. In addition to natural variation in model plants, numerous loss-of-function resources have been produced by mutagenesis with chemicals, irradiation, or insertions of transposable elements or T-DNA. However, we may be unable to observe loss-of-function phenotypes for genes with functionally redundant homologs and for those essential for growth and development. To offset such disadvantages, gain-of-function transgenic resources have been exploited. Activation-tagged lines have been generated using obligatory overexpression of endogenous genes by random insertion of an enhancer. Recent progress in DNA sequencing technology and bioinformatics has enabled the preparation of genomewide collections of full-length cDNAs (fl-cDNAs) in some model species. Using the fl-cDNA clones, a novel gain-of-function strategy, Fl-cDNA OvereXpressor gene (FOX)-hunting system, has been developed. A mutant phenotype in a FOX line can be directly attributed to the overexpressed fl-cDNA. Investigating a large population of FOX lines could reveal important genes conferring favorable phenotypes for crop breeding. Alternatively, a unique loss-of-function approach Chimeric REpressor gene Silencing Technology (CRES-T) has been developed. In CRES-T, overexpression of a chimeric repressor, composed of the coding sequence of a transcription factor (TF) and short peptide designated as the repression domain, could interfere with the action of endogenous TF in plants. Although plant TFs usually consist of gene families, CRES-T is effective, in principle, even for the TFs with functional redundancy. In this review, we focus on the current status of the gene-overexpression strategies and resources for identifying and elucidating novel functions of cereal genes. We discuss the potential of these research tools for identifying useful genes and phenotypes for application in crop breeding.
Modern plant breeding has developed several key technologies in crop breeding that have contributed to increasing and sustaining food production. Among the key technologies, the most noteworthy is the development of high-yielding and semi-dwarf varieties in rice and wheat between the 1940s and late 1960s. This technology led to increased agricultural production and is known as the Green Revolution (Khush, 1999; Basu et al., 2010). Because the improved varieties were resistant to lodging and were grown with appropriate chemical fertilization, they showed increased productivity per unit area. A second important key technology is the development of F1 hybrid varieties. Particularly, in the United States, production of hybrid maize spread widely and rapidly, and hybrid varieties became substitutes for open-pollinated varieties in 1940s. The productivity per unit area of the hybrid maize has dramatically increased: five times greater than that of pre-hybrid varieties (Crow, 1998).
Plant breeding by genetic engineering was developed in the 1980s and has become the third key technology. For molecular breeding, accumulation of molecular biological information and resources on crops is of great value. DNA sequencing technology and bioinformatics continue to evolve and whole-genome sequencing has been accomplished in maize and wheat, following that of rice (Schnable et al., 2009; Brenchley et al., 2012; International Wheat Genome Sequencing Consortium (IWGSC), 2014; Michael and VanBuren, 2015). Exploiting this information for molecular breeding will depend on the identification and characterization of functions of cereal genes. For this purpose, various collections of plant resources and their databases (DBs) have recently been developed for rice and other cereals (Mochida and Shinozaki, 2010; Tsuchida-Mayama et al., 2010; Kersey et al., 2016; Tello-Ruiz et al., 2016).
Plant genetic resources artificially induced by various mutagens can be classified into two groups based on the types of mutation: loss- and gain-of-function resources. Production of loss-of-function resources for cereals became active at the end of the 20th century after the effectiveness of ionizing radiation, alkylating chemicals and various genetic elements as potent mutagens was established (Kurata et al., 2005; Krishnan et al., 2009; Morita et al., 2009; Satoh et al., 2010). Gain-of-function resources for cereals have become popular in the 21st century in coordination with the development of transformation technology (Hiei et al., 2014) and genomics, including genome sequences, and the information and clones for full-length cDNAs (fl-cDNAs) (Tsuchida-Mayama et al., 2010; Abdeeva et al., 2012). In this review, the current status of these resources developed for identifying and deciphering novel functions present in numerous cereal genes will be overviewed with emphasis on overexpression resources. Also, the potential of these research tools for identifying useful genes and phenotypes for application in crop breeding will be evaluated and discussed.
Loss-of-function resources can be classified into two groups. The first group of resources is induced by irradiation with physical mutagens including X-rays, γ-rays, fast neutrons (FNs), and heavy ions (ion beams), and treatment with chemical mutagens such as ethyl methanesulfonate (EMS) and N-methyl-N-nitrosourea (MNU). These mutagens are applicable to any species regardless of the availability of efficient transformation systems. Therefore, using these mutagens, numerous mutant populations of cereals have been produced and characterized with the aim of isolating genes responsible for respective mutations (Kurata et al., 2005; Wu et al., 2005; Morita et al., 2009; Yamaguchi et al., 2009; Jiang and Ramachandran, 2010; Satoh et al., 2010; Kurowska et al., 2012). Mutants with easily observable phenotypes have been screened from the mutagenized populations; the causative genes have then been identified by map-based cloning strategies. Causative genes have been genetically confirmed by complementation tests. Using this workflow, various cereal genes have been identified and functionally characterized using rice mutants (Huang et al., 2009; Gao et al., 2010; Jiang and Ramachandran, 2010; Yang et al., 2011).
New reverse genetic methods have recently been developed to identify and isolate mutants for individual target genes from chemically or physically mutagenized populations (Jiang and Ramachandran, 2010; Wang et al., 2012). Chemical reagents, such as EMS and MNU, efficiently induce chemical modification of nucleotides, resulting in various point mutations. Targeting Induced Local Lesions In Genomes (TILLING), a reverse genetic method for high-throughput identification of single-nucleotide polymorphisms (SNPs) in a target gene, is applicable to the mutagenized populations with point mutations and short insertions and deletions (INDELs) (Till et al., 2003; Wang et al., 2012). Suzuki et al. (2008) modified the TILLING system using unlabeled primers and fast capillary gel electrophoresis and efficiently detected SNPs in a mutant library of rice using individual zygotes treated with MNU to produce a high mutation rate: one mutation per 135 kbp of genome sequence (1/135). A frequency of 7.4 SNPs per 1 kbp of genome sequence was estimated for 1,000 M2 mutant lines of rice. A diploid einkorn wheat (Triticum monococcum), the first domesticated crop, is an attractive genetic model for studying the functions of genes conferring wheat-specific traits. Rawat et al. (2012) prepared 1,532 M2 families of the diploid wheat induced by EMS for TILLING analysis. The estimated mutation density was 1/92 kbp. Compared with this rate, the rates for hexaploid bread wheat (T. aestivum) were much higher [1 per 24–47 kbp; Chen et al., 2012; Rawat et al., 2012] because of the higher tolerance of polyploids to mutations. FNs with relatively high linear energy transfer (LET) can induce kilobase-scale deletions in plant genomes ranging from a single base to 60 kbp (Rogers et al., 2009; Shu et al., 2012). Compared with mutations produced by chemical mutagenesis, such deletions tend to produce null mutations with few background mutations. Reverse-genetic procedures, consisting of FN mutagenesis of a population followed by high-throughput PCR screening of the population, detected deletions in Arabidopsis and rice (Li et al., 2001) and Medicago (Rogers et al., 2009).
The procedures are convenient and powerful if the following conditions are met: large populations to be prepared because of low FN mutation frequencies and deletions must be of appropriate size to be flanked by designed PCR primers that generate mutant amplicons separable from those of the wild-type (Till et al., 2007). Recent findings of Morita et al. (2009) and Belfield et al. (2012) indicated that γ-rays and FNs induce base substitutions and deletions of a few bases more frequently than larger-scale deletions in rice and Arabidopsis, respectively. Bruce et al. (2009) applied oligonucleotide microarray-based comparative genomic hybridization (array-CGH) to identify both small and large deletions in rice lines irradiated with FNs and γ-rays. Belfield et al. (2014) identified, using array-CGH, small deletions (4–104 bases) in FN-irradiated mutants of Arabidopsis. Nishizawa-Yokoi et al. (2016) screened, with TILLING, 1,968 lines of gamma-irradiated rice for a defect in the DNA Ligase4 (OsLig4) locus and obtained an Oslig4 mutant bearing a 35-base deletion in exon 14. To retrieve mutants among soybean plants heavily mutagenized with EMS, Tsuda et al. (2015) designed and compared two novel screening strategies: high resolution melting (HRM) assay and indexed amplicon sequencing [DNA sequencing by next-generation sequencing (NGS) technologies of PCR fragments for target gene regions amplified from pooled samples], and reported that the latter detected mutated targets more efficiently.
The second group of resources is derived by insertion of endogenous or exogenous DNA insertion elements to generate loss-of-function mutations. In rice, large loss-of-function mutant populations have been generated by activation and random genome insertion of the Tos17 retrotransposon (Hirochika, 2001; Miyao et al., 2003, 2007), Ds and dSpm transposable elements (Upadhyaya et al., 2002; Kolesnik et al., 2004; Kumar et al., 2005; van Enckevort et al., 2005; Park et al., 2007), or T-DNAs (An et al., 2003; Chen et al., 2003; Ryu et al., 2004; Sallaud et al., 2004). These loss-of-function mutant resources are convenient for identification and functional analyses of genes disrupted and tagged by insertion of the DNA elements and can be widely used for both forward and reverse genetic approaches.
Ogawa et al. (2011) isolated a dwarf and short-root mutant (rice salt sensitive 1, rss1) under high salt stress among Tos17-mutagenized rice plants (M2 generation from ∼2,500 lines). They obtained two more alleles of rss1 mutation by PCR-based screening of DNA pools of genomic DNA mixtures from Tos17-insertion lines. Molecular analysis indicated that RSS1 encodes a novel protein that acts as a key factor in the maintenance of meristematic activity by ensuring cell division under saline conditions (Ogawa et al., 2011).
Zou et al. (2011) screened more than 100,000 T-DNA insertion lines of rice derived from several populations and isolated 312 mutant lines with rolled leaves. Among them, an outcurved leaf1 mutant showing abaxial (outside) leaf rolling was identified. This phenotype was caused by knockout of Rice outermost cell-specific gene5 (Roc5). Overexpression of Roc5 induced an opposite phenotype, adaxially rolled leaves, to the Roc5-knockout rice. Roc5 encodes a homeodomain leucine-zipper class IV TF, an ortholog of Arabidopsis GLABRA2. Further analyses indicated that Roc5 functions as a negative regulator controlling bulliform cell number and size in rice leaves (Zou et al., 2011). Ning et al. (2011) found a transgenic rice plant bearing a T-DNA insertion in the fourth exon of Increased Leaf Angle1 (ILA1) gene among a large T-DNA-insertion population of more than 130,000 lines developed for enhancer tagging (Rice Mutant Database1). The ila1 mutant displayed smaller vascular bundles and reduced cellulose content in leaf lamina joints, and the phenotypes were unrelated to response to brassinosteroids (BRs). Molecular analyses revealed that ILA1 encodes a Raf-like MAPKKK and is mainly expressed in vascular bundles of lamina joints. Ning et al. (2011) identified a novel mechanism of leaf angle regulation by ILA1, a key factor conferring mechanical strength to lamina joints. Ramamoorthy et al. (2011) screened visible mutant lines among approximately 20,000 lines of rice carrying Ds insertions and found a mutant with semi-dwarfism caused by defects in cell elongation. The mutant line included a single-copy Ds insertion in OsCYP96B4, a member of the cytochrome P450 multigene family.
As described in the previous section, many mutants have been isolated from loss-of-function resources and their causative genes have been identified and characterized. However, it would be difficult to observe loss-of-function phenotypes for gene families with functional redundancy, such as those for various TFs and genes essential for growth and development. To address this disadvantage, various overexpression systems for transgenes have been developed: gain-of-function resources and tools based on activation tagging and Full Length-cDNA OvereXpressor gene (FOX) hunting, and loss-of-function resources and tools on gene-silencing technologies with RNA interference (RNAi) and artificial microRNAs (amiRNAs), and on a dominant-negative strategy, Chimeric REpressor gene Silencing Technology (CRES-T) (Walden et al., 1994; Horiguchi, 2004; Eamens et al., 2008; Tsuchida-Mayama et al., 2010; Kondou et al., 2011; Mitsuda et al., 2011a; Abdeeva et al., 2012; Sun, 2012; Zheng and Qu, 2015). Researchers may select these resources depending on their research purposes (Figure 1). In the following paragraphs, transgenic resources based on both gain- and loss-of-function approaches (Table 1) and also some examples of characterization and application of gene functions in cereals (Table 2) are introduced.
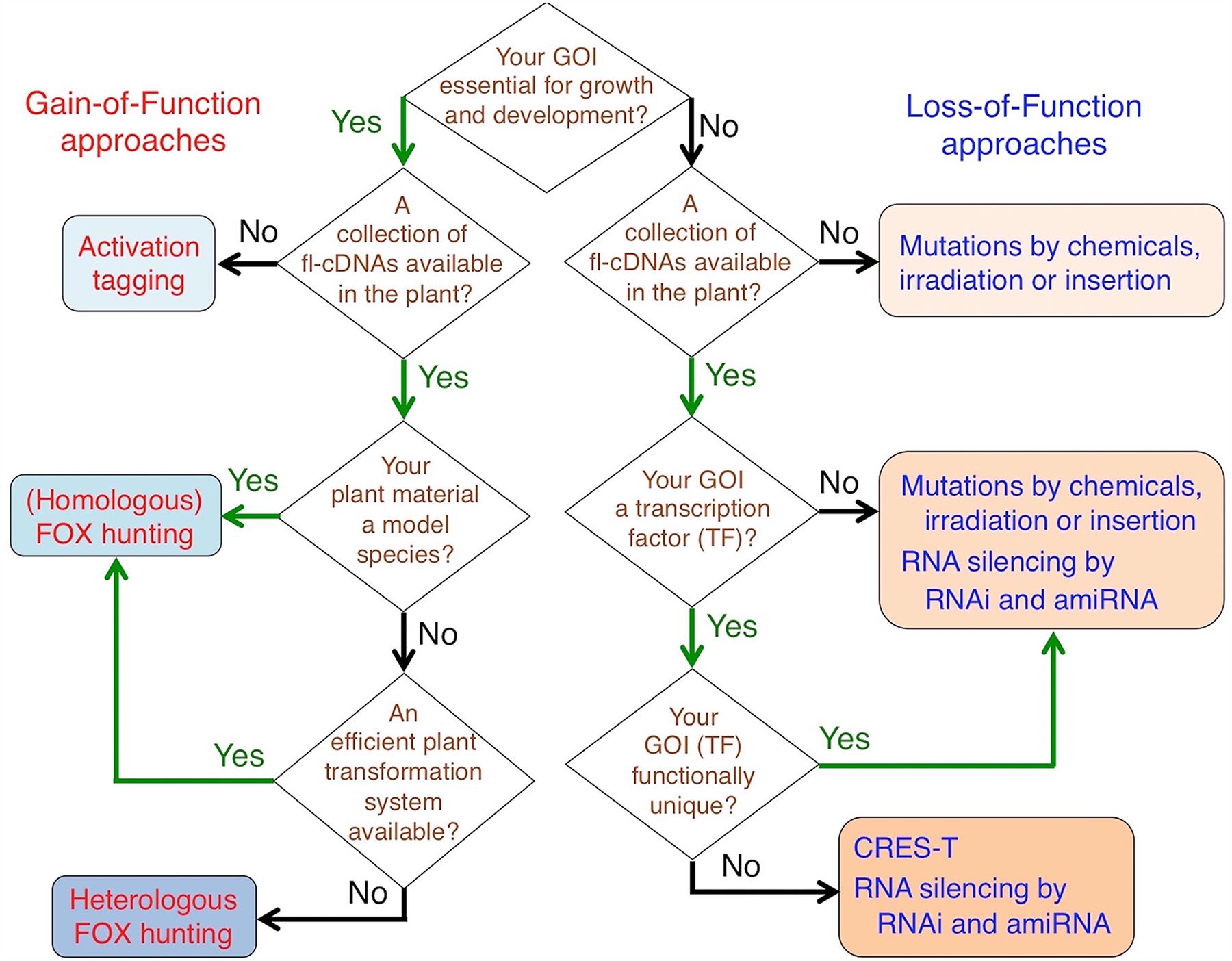
FIGURE 1. Choosing appropriate resources to isolate your gene(s) of interest by forward and/or reverse genetic approaches. GOI, Gene(s) of interest.
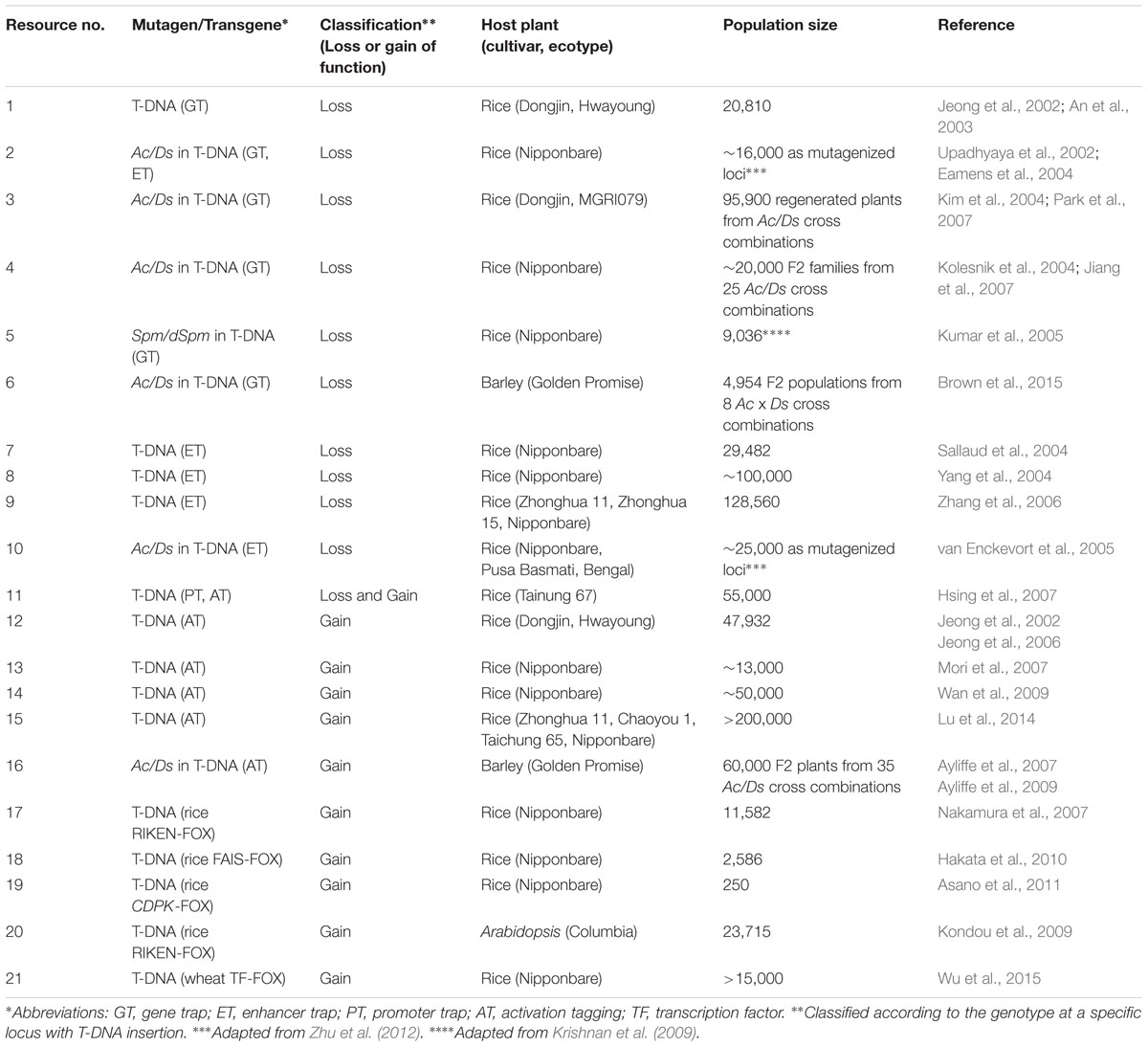
TABLE 1. Loss- and gain-of-function transgenic resources in cereals.
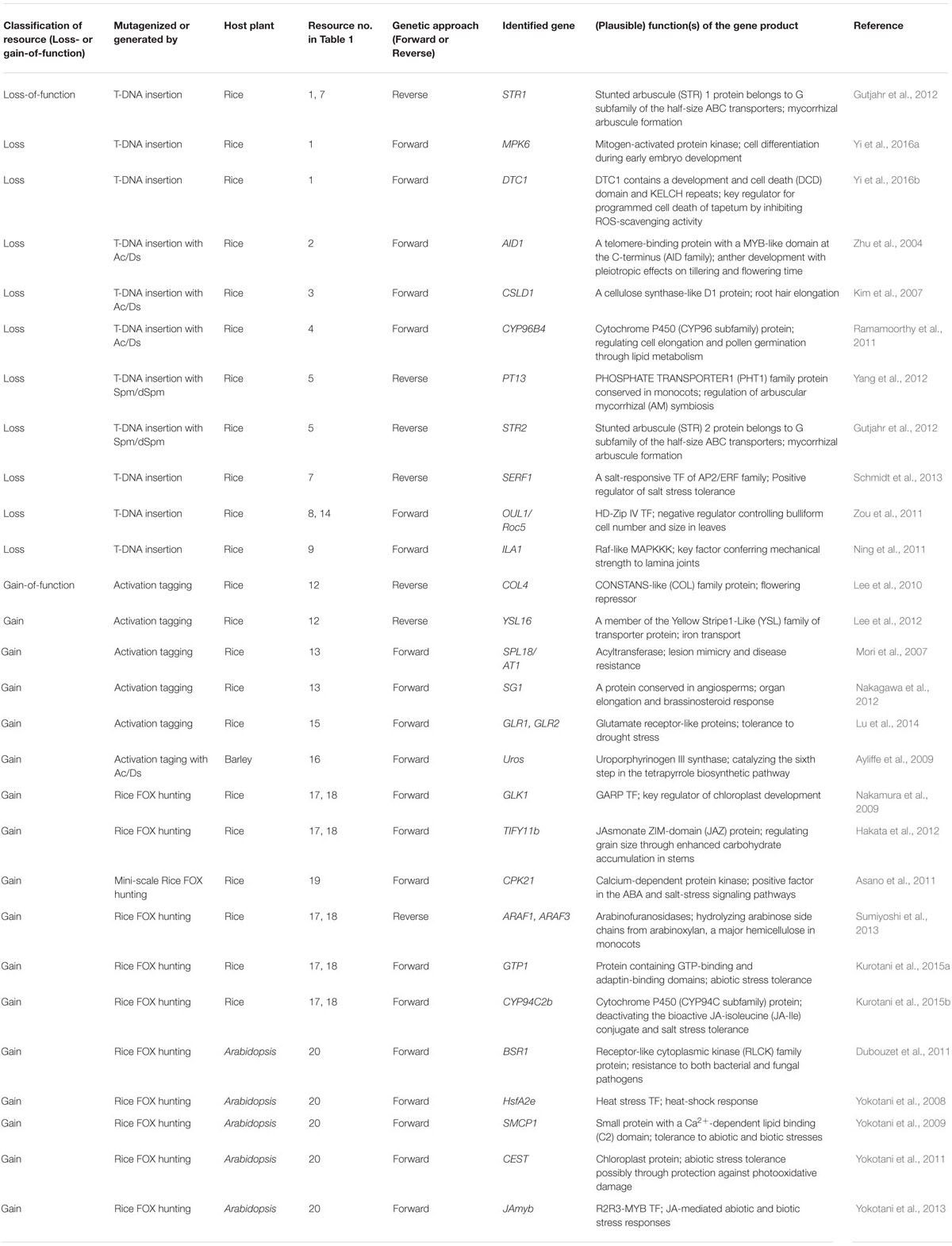
TABLE 2. Examples on the identification and characterization of genes by using transgenic resources in cereals.
The concept of activation tagging by T-DNA was first described by Walden et al. (1994). In the system, multiple transcriptional enhancer elements are randomly introduced into the plant genome. Gene(s) adjacent to an insertion site of the enhancer elements display enhanced expression and show dominant gain-of-function phenotypes. The first successful example of activation-tagged gene was Arabidopsis ICK1 encoding a histidine kinase homolog involved in cytokinin signaling (Kakimoto, 1996).
Activation tagging is applicable to both forward and reverse genetic screens, as is the case with loss-of-function resources generated by Tos17, Ds, and T-DNA insertions. Though activation tagging was first applied to generate gain-of-function resources in Arabidopsis (Weigel et al., 2000), numerous activation-tagged rice resources have been generated by the insertion of T-DNA or transposable elements (Jeong et al., 2002, 2006; Chern et al., 2007; Hsing et al., 2007; Mori et al., 2007; Qu et al., 2008; Wan et al., 2009). We first introduce two typical examples for elucidating gene functions using reverse genetic approach. Jeong et al. (2006) generated 47,932 activation-tagged lines by T-DNA and obtained 27,621 flanking sequence tags (FSTs) by inverse PCR. Using this resource and FST information, Lee et al. (2010) isolated an activation tagging line of the OsCOL4 gene (OsCOL4-D, D for Dominant), a member of the CONSTANS-like (COL) family. Given that Cauliflower mosaic virus 35S enhancer elements were inserted approximately 2 kbp upstream of OsCOL4, the transcript level was significantly upregulated in the line. Oscol4 null mutants flowered early under short and long days. In contrast, OsCOL4-D plants flowered late in either environment, indicating that OsCOL4 functions as a flowering repressor in rice (Lee et al., 2010). Lee et al. (2012) analyzed the function(s) of OsYSL16, which is encoded by a member of 18 Yellow Stripe1-like (YSL) genes and may be involved in iron transport in rice. Knockout mutation and antisense suppression of OsYSL16 did not alter Fe distribution and utilization. Two lines (OsYSL16-D1 and -D2) ectopically overexpressing OsYSL16 in the activation tagging population (Jeong et al., 2006) were identified. OsYSL16-D plants were more tolerant to iron deficiency than the wild type (WT), and OsYSL16 played a role in the mobilization of iron from roots to shoots (Lee et al., 2012). This is a typical example that function(s) of a gene of interest cannot be characterized by a loss-of-function mutation without altered phenotype(s) because of the possible functional redundancy, but can be characterized by gain-of-function mutation resulting in the altered phenotypes.
Next, three examples of the identification and characterization of genes by forward genetics using activation-tagged resources are shown. Mori et al. (2007) generated approximately 13,000 activation-tagged lines of rice using a binary vector carrying tetramerized 35S enhancer elements along with 35S minimum promoter and the first intron from the phaseolin gene in T-DNA with aim of activating flanking genes more strongly than by the enhancer alone. Among the population, a dominant lesion-mimic line, Spotted leaf 18 (Spl18), was obtained by visual screening. Spl18 showed enhanced resistance to both blast fungus and bacterial blight and harbored a T-DNA insertion 481 bp upstream of OsAT1 encoding an acyltransferase. Increased expression of OsAT1 was correlated with lesion mimicry and disease resistance (Mori et al., 2007). Nakagawa et al. (2012) identified a dominant mutation with short grain and semi-dwarfism (Sg1-D) by visual screening of the above-mentioned activation tagging population in rice (Mori et al., 2007). A gene with unknown function Os09g0459200 located 1.4-kbp downstream of T-DNA insertion was overexpressed and responsible for the Sg1-D mutation. This mutant also showed erect, short, wide, and dark-green leaves and also decreased cell division and response to BR. The results indicated that SG1 down-regulates BR signaling or response to BR and causes suppression of cell proliferation and finally organ elongation (Nakagawa et al., 2012). Lu et al. (2014) developed a population of more than 200,000 activation-tagged rice lines based on T-DNA bearing tetramerized 35S enhancers and identified a tagged line displaying enhanced drought tolerance by a forward genetic approach. In the mutant, T-DNA was inserted in the second intron of the glutamate receptor-like 1 gene (OsGLR1). Another GLR gene OsGLR2 was located adjacent to OsGLR1. Overexpressing OsGLR1 or OsGLR2 significantly enhanced drought tolerance in both rice and Arabidopsis, indicating that GLR genes play important roles in drought tolerance and could be used to improve agronomic performance of monocot and dicot crops (Lu et al., 2014).
Though the FOX-hunting system and CRES-T require fl-cDNA collection and cDNA clones of TFs, respectively, activation tagging can be applied to non-model plant species whose genome and/or cDNA sequences are not yet available. It is not realistic, however, to apply activation tagging in cereals without high-throughput transformation systems (Hiei et al., 2014). Accordingly, Ayliffe et al. (2007) applied activation tagging to barley, a non-model plant species, in combination with the Ac/Ds transposable element of maize. Approximately 20 primary transgenic barley lines carrying a modified Ds element (UbiDs) consisting of the terminal 5′ and 3′ ends of Ac and two copies of the maize Polyubiqiuitin (Ubi) promoter placed oppositely for ectopic overexpression of genes near T-DNA were each crossed to two Ac lines bearing the homozygous Ubi promoter::Ac transposase. In the hybrid progenies, transposition of the UbiDs element was observed at frequencies sufficient for mutagenesis. In a screen of 60,000 F2 seedlings, a mutant was identified that displayed necrotic phenotype with reduced seedling vigor. An UbiDs element was inserted downstream of the uroporphyrinogen III synthase (Uros) gene, whose product catalyzes the sixth step in the chlorophyll and heme biosynthetic pathway. In the mutant, Uros transcript was significantly reduced by the accumulation of antisense Uros transcript initiated by the UbiDs element (Ayliffe et al., 2009).
Although activation tagging is very useful for the identification and characterization of genes, as described above, caution is warranted. Activation-tagged lines sometimes show complex phenotypes, when the enhancer element affects the expression of several genes (Ichikawa et al., 2003; Tsuchida-Mayama et al., 2010). Besides acting as a gene-activating agent, the activation-tagging cassette can behave as an agent for gene disruption and antisense expression. Thus, activation tagging gives rise to both gain- and loss-of-function phenotypes, depending on the location and direction of insertions.
Because of remarkable progress in DNA sequencing technology and bioinformatics (Mochida and Shinozaki, 2010), whole-genome information of several grasses has recently been available, including rice (International Rice Genome Sequencing Project, 2005), maize (Schnable et al., 2009), sorghum (Paterson et al., 2009), Brachypodium (International Brachypodium Initiative, 2010), barley (International Barley Genome Sequencing Consortium, 2012) and wheat (International Wheat Genome Sequencing Consortium (IWGSC), 2014). Additionally, large-scale collections of fl-cDNA clones have been generated for rice (Rice Full-Length cDNA Consortium, 2003), barley (Mochida et al., 2009; Sato et al., 2009; Matsumoto et al., 2011), maize (Soderlund et al., 2009), wheat (Kawaura et al., 2009; Mochida et al., 2009), and Brachypodium (Mochida et al., 2013). Fl-cDNAs provide sequence information for mature mRNAs and proteins for the corresponding genes (Kondou et al., 2011). Thus, collections of characterized fl-cDNA clones are useful for analyzing functions of genes and proteins (Abdeeva et al., 2012). By exploiting an fl-cDNA collection of Arabidopsis, a novel gain-of-function technology was developed for the identification of functions of the genes and designated as FOX-hunting system (Ichikawa et al., 2006).
In the FOX-hunting system, individual fl-cDNAs are mixed at nearly equal molar ratios and then cloned downstream of a strong promoter in an expression vector. The resulting fl-cDNA expression library is transformed into plants via Agrobacterium, and a large population of transgenic plants can be obtained, many of which overexpress a single fl-cDNA. Among them, we can screen and isolate FOX lines displaying altered phenotypes of interest. Fl-cDNA(s) introduced in the FOX lines can be easily identified with T-DNA specific primers. In this manner, fl-cDNA that caused the altered phenotype(s) can be directly linked to a function. The FOX hunting system is applicable to various plants, including non-model species, provided that fl-cDNA clones and highly efficient transformation system are available (Ichikawa et al., 2006; Kondou et al., 2011).
In rice, excellent transformation systems using Agrobacterium are available (Hiei et al., 1994; Toki et al., 2006). Nakamura et al. (2007) and Hakata et al. (2010) have developed the FOX-hunting system in rice for the functional analysis of rice genes. Rice fl-cDNA clones were originally prepared and sequenced by two groups, the Institute of Physical and Chemical Research (RIKEN) and the Foundation for Advancement of International Science (FAIS) in Japan (Rice Full-Length cDNA Consortium, 2003). Nakamura et al. (2007) used a mixture of 13,980 rice fl-cDNA clones from RIKEN to construct a cDNA overexpression library for rice. Each fl-cDNA was cloned downstream of the maize Ubi promoter in a binary vector for overexpressing cDNAs. Approximately 12,000 rice FOX-rice lines were generated by transforming rice with the rice FOX-Agrobacterium library (Nakamura et al., 2007). Hakata et al. (2010) used 13,823 fl-cDNA clones from FAIS to generate a cDNA overexpression library. Each fl-cDNA was cloned downstream of the rice Actin-1 promoter in a Gateway-based expression vector. Approximately 2,600 rice FOX-rice lines were produced (Hakata et al., 2010). Introduced fl-cDNA(s) in each line in these two FOX populations were identified by amplification of a fragment bearing fl-cDNA by genomic PCR using the promoter- and terminator-specific primers, followed by direct sequencing of the PCR fragment. A total of 7,050 fl-cDNAs were contained in the rice FOX-rice lines, of which 332 were found in both FOX populations.
Functions of several genes have been characterized using these two populations of FOX-rice lines. Nakamura et al. (2009) found FOX-rice lines producing green calli even in the presence of auxin, which suppresses plastid differentiation and greening. The mutation was caused by ectopic overexpression of a cDNA encoding a GARP TF, Golden2-like 1 (OsGLK1). A large number of genes involved in chloroplast functions were highly expressed in the OsGLK1-FOX green calli. Well-developed thylakoid membranes and grana structures were observed in the plastids of OsGLK1-FOX calli. These results showed that OsGLK1 is a key regulator of chloroplast development (Nakamura et al., 2009). Hakata et al. (2012) identified FOX-rice lines overexpressing an fl-cDNA for OsTIFY11b/OsJAZ10 as a negative regulator of jasmonate (JA) signaling. The OsTIFY11b-FOX lines showed enhanced growth, including increased grain size, weight, leaf length, and plant height. The increases in grain size and weight were suggested to be caused by enhanced accumulation and translocation of carbohydrates in leaf sheaths and culms in the FOX lines (Hakata et al., 2012). As shown in these two examples, phenotype(s) altered by overexpression of each fl-cDNA for TF are useful clues to the function(s) of the gene product. Since duplicated GLK genes have functional redundancy, no altered phenotypes were observed in a single loss-of-function mutation in the two GLK genes (Wang P. et al., 2013).
A T1 population of 2,229 FOX-rice lines (Nakamura et al., 2007; Hakata et al., 2010) was treated with 300 mM NaCl for 5 weeks, and two lines showed salinity tolerance (Kurotani et al., 2015b). One of the two lines overexpressed an fl-cDNA for a protein, GTP1 containing putative GTP- and adaptin-binding domains, whose function remains to be elucidated. The other line carried an fl-cDNA encoding CYP94C2b, a cytochrome P450 family protein. CYP94C2b was involved in deactivation of the bioactive conjugate of JA, jasmonoyl isoleucine (JA-Ile) which activates JA signaling. CYP94C2b-FOX rice plants showed reduced responses to JA and enhanced shoot viability under salt stress, suggesting that the repression of JA action is responsible for enhanced salt tolerance (Kurotani et al., 2015a).
Asano et al. (2011) reported FOX-rice lines overexpressing calcium-dependent protein kinases (CDPKs), Ca2+-binding sensory kinases that regulate downstream components of calcium signaling. Because CDPKs constitute a large multigene family of 29 members in rice, biological functions of these CDPK genes would show functional redundancy and might be difficult to characterize by loss-of-function phenotypes. By using three Agrobacterium pools divided among 25 CDPK genes and the OsCCaMK1 gene for a calmodulin-dependent protein kinase, Asano et al. (2011) applied a mini-scale FOX hunting system (Fujita et al., 2007). They produced a total of 250 CDPK FOX-rice lines accounting for 24 of the 25 CDPK and OsCCaMK1 genes. Among FOX lines screened for salt stress tolerance, OsCPK21-FOX plants showed tolerance to salt stress by increasing sensitivity to ABA, suggesting the role of OsCPK21 as a positive factor in the ABA and salt-stress signaling pathways (Asano et al., 2011). This study is regarded as a test case for the mini-scale FOX hunting for a specific gene family. Considering the task of identification of fl-cDNA in each FOX line by genomic PCR and DNA sequencing, it might be convenient to transform host plants by fl-cDNA expression constructs one by one, when a multigene family consists of approximately 100 or fewer members. However, transforming rice with 1,000 fl-cDNA overexpression constructs one by one would be laborious. Provided that we generate an Agrobacterium library carrying 1,000 T-DNA vectors and transform rice with this single library, approximately 2,300 and 4,600 transgenic lines would require to cover 90 and 99% of fl-cDNAs, respectively, according to the equation formulated by Clarke and Carbon (1976). Although genomic PCR with a pair of specified primers followed by Sanger sequencing to identify individual fl-cDNA(s) in each transgenic line would be time-consuming, the FOX-hunting system would be a reasonable alternative to the one-by-one approach.
Sumiyoshi et al. (2013) reported FOX-rice lines overexpressing OsARAF1 or OsARAF3 for arabinofuranosidase (ARAF). ARAF dissolves arabinose side chains that bind to a xylose backbone and constitute a hemicellulose, arabinoxylan. Arabinose contents in the cell wall of OsARAF1- and OsARAF3-FOX lines decreased to 75.4 and 81.6%, and amounts of glucose increased to 128.2 and 134.2%, respectively, of that in WT. These FOX lines could be advantageous for producing bioethanol (Sumiyoshi et al., 2013). This is an example that FOX lines show valuable phenotypes of potential industrial use.
Kondou et al. (2009) developed a heterologous FOX-hunting system. An fl-cDNA expression library was constructed by individually placing approximately 13,000 rice fl-cDNAs under the control of the 35S promoter. By introduction of fl-cDNA overexpression constructs into Arabidopsis via in planta transformation, more than 23,000 independent rice FOX-Arabidopsis lines, theoretically encompassing 11,000 of the 13,000 fl-cDNAs, were generated. The advantage of this heterologous FOX-hunting system is in characterizing gene functions using another host plant with a short life cycle and compact size that is equipped with a quick and efficient transformation system. Given that Arabidopsis is one of the ideal host plants that fulfill all the features, it was exploited for the heterologous FOX-hunting system to characterize gene functions (Kondou et al., 2009, 2011; Higuchi-Takeuchi et al., 2013).
Using approximately 20,000 rice FOX-Arabidopsis lines, Dubouzet et al. (2011) screened mutant lines resistant against Pseudomonas syringae and obtained 74 resistant lines. One of the selected genes, BROAD-SPECTRUM RESISTANCE 1 (BSR1) encoding a receptor-like cytoplasmic kinase (RLCK) family protein, conferred strong resistance to both bacterial and fungal pathogens when overexpressed in both Arabidopsis and rice. Thus, the heterologous system could identify potentially useful gene(s) that confer multiple or broad-spectrum disease resistance on both dicot and monocot plants. Yokotani et al. (2008) screened the same population of rice FOX-Arabidopsis lines and isolated a mutant tolerant to heat stress. The rice fl-cDNA inserted in this line encoded OsHsfA2e, a member of heat stress TFs (HSFs) playing a central role in heat-shock response in many species (Yokotani et al., 2008). Yokotani et al. (2009) also screened 208 candidates for salt-tolerant lines among the FOX population. One of the mutants carried a rice cDNA encoding OsSMCP1, a small protein with a single C2 domain for Ca2+-dependent membrane binding. The OsSMCP1-FOX Arabidopsis plants showed improved tolerance to various abiotic and biotic stresses other than salinity (Yokotani et al., 2009). The other salt-tolerant lines carried an fl-cDNA for CHLOROPLAST PROTEIN-ENHANCING STRESS TOLERANCE (CEST), a novel chloroplast protein found in photosynthetic organisms (Yokotani et al., 2011), and JAmyb as a JA-responsive TF (Yokotani et al., 2013). These attractive examples show that the heterologous FOX-Arabidopsis resource is advantageous for high-throughput screening to yield plants with agronomically useful traits, although a homologous expression system would often be needed to confirm the function(s) of the gene of interest.
The heterologous FOX-hunting system could be a powerful tool for researchers working on non-model cereals, such as maize, barley and wheat, without appropriate facilities and skills of efficient transformation for those cereals but with large-scale collections of fl-cDNAs (Kawaura et al., 2009; Mochida et al., 2009; Sato et al., 2009; Soderlund et al., 2009; Matsumoto et al., 2011). Arabidopsis and rice with convenient transformation protocols would be suitable as host plants. Transgenic Arabidopsis plants can be efficiently generated with in planta transformation (Kondou et al., 2009). Accordingly, we need not consider background mutations caused by somaclonal variation, indicating the excellent linkage between mutant phenotypes and introduced cDNAs in the FOX-Arabidopsis lines. In contrast, rice transformation needs tissue culture. There is a tendency that the frequency of somaclonal variation is related to the duration of tissue culture. Minimizing the duration could suppress somaclonal variation in rice (Toki et al., 2006), indicating that the employment of high-speed transformation is important.
Wu et al. (2015) generated wheat FOX-rice plants using 1,455 fl-cDNAs of TFs from bread wheat and its relatives. TF cDNAs were divided into three size fractions: <1 kbp, 1–2 kbp, >2 kbp. Each fl-cDNA fraction was transformed into a Gateway-based binary vector to construct three wheat TF-FOX Agrobacterium libraries. By transforming rice with the libraries, more than 15,000 transgenic rice plants (T0 generation) were generated. Among 10,496 T0 plants that set seeds, 1,562 lines (14.9%) showed altered phenotypes at the T0 stage. Wu et al. (2015) then used 5,232 T1 lines to screen for salt and osmotic stress tolerance and identified seven TFs that functioned in stress tolerance.
Some graminaceous plants have recently emerged as new attractive model species. Brachypodium distachyon, a wild grass species, is one such species. Brachypodium, as well as wheat and barley, belongs to Pooideae, one of the three subfamilies of agronomical importance in the grass family Poaceae (Kellogg, 2001). This plant is small (∼20 cm), self-fertile and long-day with a short life cycle of a few months, and grows in the same facilities as Arabidopsis (Draper et al., 2001; Opanowicz et al., 2008). The compact genome of Brachypodium, approximately 272 Mbp on five chromosomes, has been sequenced by the International Brachypodium Initiative (2010). Moreover, molecular and genetic tools have been developed, including efficient transformation system preferentially using immature embryos (Garvin et al., 2008; Alves et al., 2009; Thole and Vain, 2012). Approximately 24,000 T-DNA insertion lines have been generated as of October, 2015 at the USDA-ARS2, and approximately 5,000 lines at the John Innes Centre (Thole et al., 2012). Application of the homologous FOX-hunting system to Brachypodium and/or that of the heterologous system to wheat and barley would be convenient and advantageous for high-throughput and accurate elucidation of functions of genes in Pooideae.
Setaria viridis (green foxtail), a wild ancestor of S. italica (foxtail millet), has also become popular as an attractive model species for the study of C4 grasses. S. viridis combines such convenient traits as diploidy (n = 9), short life cycle (6–9 weeks), small size (∼20 cm) and self-fertility with prolific seed production (∼13,000 seeds per plant) (Brutnell et al., 2010; Li and Brutnell, 2011; Brutnell et al., 2015). S. viridis has a relatively small genome (∼510 Mbp), and the whole genome sequence is available (Bennetzen et al., 2012; Zhang et al., 2012). The C4 photosynthetic pathway in association with a specialized leaf structure (Kranz anatomy) confers high productivity on several major food crops and bioenergy grasses, including maize, sugarcane, sorghum, switchgrass, and Miscanthus that belong to the Panicoideae subfamily (Brutnell et al., 2010). Recently, efficient procedures for Agrobacterium-mediated transformation of S. viridis have been developed using embryogenic calli derived from mature seeds (Brutnell et al., 2010; Martins et al., 2015b; Van Eck and Swartwood, 2015). Floral-dip transformation of S. viridis has been successful with a transformation efficiency of 0.6% (Martins et al., 2015a), and this could accelerate functional genomics in C4 grasses. For example, a population of S. viridis plants transformed with an expression library for maize fl-cDNAs (maize FOX-Setaria) could facilitate identification of functions of genes for C4 photosynthesis.
Antisense RNA and RNAi technologies are based on RNA-induced gene silencing [in other words, posttranscriptional gene silencing (PTGS)]. In the process, double-stranded small RNAs are generated and mediate sequence-specific RNA degradation for a target gene. RNA-induced gene silencing has been adopted extensively to plants since 1980’s for elucidating functions of particular gene(s) (Horiguchi, 2004; Eamens et al., 2008). In spite of the technological versatility, populations of transgenic plants overexpressing a variety of antisense RNA or RNAi constructs have not been generated until recently, probably because of the availability of various loss-of-function resources as described in the previous section. Wang L. et al. (2013) produced three hairpin RNA (hpRNA) libraries, OsHP2, OsHP4, and OsHP6, using size-fractionated cDNAs (200–400 bp, 400–600 bp, and 600–1,000 bp, respectively) from various rice tissues. Each T-DNA construct carried an RNAi cassette placed under the control of Ubi promoter for overexpression. The hpRNA library OsHP4 was introduced into Agrobacterium, and transformed into rice to generate more than 6,000 transgenic hpRNA plants. Among them, approximately 3,000 lines were deduced to possess the entire T-DNA regions, and more than 30% of them displayed poor growth and/or sterility. About 48% of the T1 lines derived from fertile T0 plants showed diverse mutations, and the ratio was much greater than that (3.5%) of a population of T-DNA insertion lines. The results suggested effective silencing of the target genes by the hpRNA library, and a high potential of the rice RNAi population for genomewide gene identification (Wang L. et al., 2013).
microRNAs (miRNAs), small RNAs of 20–24 nucleotides in length, have been found in almost all eukaryotes and play important roles through post-transcriptional control of genes in regulating almost all biological and metabolic processes including cell differentiation, organ development, response to biotic and abiotic stresses (Sun, 2012; Zheng and Qu, 2015). The combined genome and transcriptome sequencing studies have identified 427, 713, and 321 miRNAs in Arabidopsis, rice and maize, respectively (release 21 of miRBase) (Kozomara and Griffiths-Jones, 2014). A miRNA family, MIR528, is distributed only in monocot species (Cuperus et al., 2011). Warthmann et al. (2008) chose the 254-bp precursor of rice MIR528 (OsMIR528), modified its miRNA/miRNA∗ region to silence endogenous and exogenous target genes, and evaluated the silencing of three different rice genes. Selective silencing of the target genes in transgenic rice plants overexpressing each amiRNA construct was induced and stably inherited in the progenies. MIR390 family is highly conserved in plants (Cuperus et al., 2011). For high-throughput production of the amiRNAs and their overexpression in monocots, a series of expression vectors carrying the OsMIR390 precursor was constructed and validated in transgenic Brachypodium plants (Carbonell et al., 2015). Among the chimeric OsMIR390-based precursors tested, amiRNAs accumulated to high levels, was accurately processed when expressed from the OsMIR390-based precursor (OsMIR390-AtL) of which the distal stem–loop sequence (16 bp) was replaced with that (31 bp) of Arabidopsis MIR390a (AtMIR390a), and induced effective silencing of individual target genes. In combination with a web tool, Plant Small RNA Maker Site (P-SAMS), for automated design of amiRNAs (Fahlgren et al., 2016), binary vectors carrying the OsMIR390-AtL precursor enable direct cloning of various amiRNAs and could be used for generating large-scale amiRNA construct libraries for silencing genes in monocots (Carbonell et al., 2015).
Transcription factor is a protein that binds to specific DNA sequences typically located in the 5′-upstream regions of protein-coding sequences of the target genes and plays key role(s) in the control of gene expression in growth, development, and response to environmental stimuli in plants (Riechmann et al., 2000; Zhang, 2003). Most of plant TFs constitute multigene families and functions of TFs that belong to a subfamily often show redundancy. Therefore, it would be difficult to elucidate TF functions by knock-out or knock-down of the TF gene(s) of interest (Mitsuda et al., 2011a). To overcome such functional redundancy in TFs, Hiratsu et al. (2003) developed a novel gene silencing system designated as CRES-T. In this system, a TF is converted to a strong repressor (chimeric repressor) by fusion of the EAR-motif repression domain (SRDX). Overexpression of the chimeric repressor of the TF in host plants suppresses expression of target genes of both the TF and those with functional redundancy, and the dominant-negative phenotype(s) give clues to the function(s) of TF members (Hiratsu et al., 2003; Kubo et al., 2005; Mitsuda et al., 2011a).
To date, several TF genes have been characterized using CRES-T in rice (Mitsuda et al., 2006; Ogo et al., 2008; Mito et al., 2011; Tanaka et al., 2012; Todaka et al., 2012; Yoshida et al., 2013). For instance, Ogo et al. (2008) identified a novel TF of the NAC family IDEF2 that specifically binds to the iron deficiency-responsive cis element 2 (IDE2) in rice and barley. Transgenic rice plants in which the function of IDEF2 was disrupted by RNA interference (RNAi) or CRES-T (IDEF2::SRDX under the control of rice Actin-1 constitutive promoter) revealed aberrant iron homeostasis and repression of expression of genes induced under iron deficiency, indicating that IDEF2 functions as a key TF regulating iron-deficiency response (Ogo et al., 2008).
Yoshida et al. (2013) found two NAC TFs in rice, SECONDARY WALL NAC DOMAIN PROTEIN 1 and 2 (OsSWN1 and OsSWN2), that are orthologs of NAC SECONDARY WALL THICKNINGS FACTORs (NSTs) from Arabidopsis as master regulators of secondary wall formation. OsSWN1 transcription was highly active in sclerenchyma cells of leaf blades and less active in xylem cells. In contrast, OsSWN2 transcription was particularly active in xylem cells and less in sclerenchyma cells. OsSWN2 produces two alternatively spliced variants OsSWN2L and OsSWN2S of which the truncated ORF lacks a potential transcriptional activation domain. Transgenic rice plants expressing a chimeric repressor of OsSWN2S (OsSWN2S-SRDX) driven by the OsSWN2 promoter caused stunted growth and needle-like shape and browning in leaves. Transgenic lines carrying OsSWN2S-SRDX driven by the OsWN1 promoter showed drooping leaves, reduced thickness of secondary walls in sclerenchyma cells, and reduced lignin and xylose contents, resulting in enhanced cell-wall digestibility. These results show that OsSWNs regulate secondary wall formation in rice and would be useful for improving cereals and grasses for forage and industrial use of products, such as sugars and biofuels (Yoshida et al., 2013).
As described above, CRES-T is a powerful tool for characterization of various TFs even with functional redundancy. In ornamental and model plants, including torenia, chrysanthemum, cyclamen and Arabidopsis, a database of floral phenotypes induced by CRES-T has been developed (FioreDB3) (Mitsuda et al., 2011b). For comprehensive study and discovery of unknown TF functions, a large population of CRES-T (TF cDNA::SRDX driven by the 35S promoter) transgenic lines for 1,600 independent TFs from Arabidopsis was generated. Evaluation of CRES-T lines revealed that repressors for six TFs conferred high tolerance to salt and osmotic stresses on Arabidopsis plants (Kazama et al., 2013). CRES-T resources for TFs of monocots may be developed in some species, including rice and Brachypodium, with available collections of fl-cDNA clones.
In the Sections “Loss-of-Function Resources by Direct Modification of Genes” and “Gain- and Loss-of-Function Resources by Overexpression,” various loss- and gain-of-function resources are described with special emphasis on the overexpression resources (see Gain- and Loss-of-Function Resources by Overexpression). For comparison, advantages and disadvantages for respective mutant resources are summarized in Table 3.
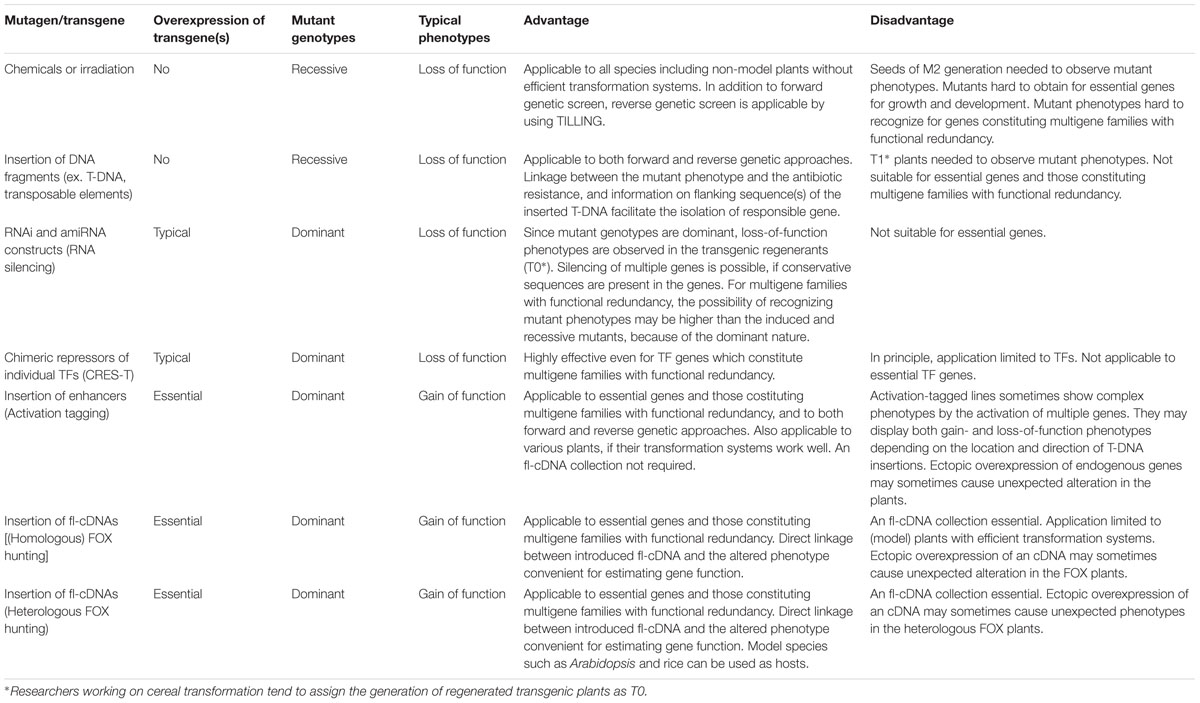
TABLE 3. Comarison of artificial mutant resources in plants induced by various mutagens.
Use of the gene-overexpression resources generated by activation tagging, FOX hunting, and CRES-T has resulted in the discovery and characterization of many cereal genes. When these resources are used for the identification, screening, and/or evaluation of genes and their functions of interest, the choice of appropriate resource(s), by considering their (plausible) expression patterns and networks and phenotypes, would be important. As DNA sequencing technology continues to progress, the cost and time required for whole-genome sequencing in many plants may be reduced. Then, collections of sequence-based resources, such as fl-cDNAs, can be developed for many plants. In cooperation with the development and improvement of transformation technology (Hiei et al., 2014), novel combinations of both homologous and heterologous FOX hunting systems may be developed. Numerous combinations of loss- or gain-of-function phenotypes with agronomically important traits may lead to “super cereals.” Most recently, genome editing technologies, including transcription activator-like effector nucleases (TALENs) and clustered regularly interspaced short palindromic repeats (CRISPR)/CRISPR-associated (Cas) system, have been developed in cereals (Zhang et al., 2014; Osakabe and Osakabe, 2015). By adequately combining homology-dependent recombination (i.e., gene targeting), the genome-editing technologies provide on-demand insertion, deletion, or replacement at the target locus in a genome. Therefore, novel plant materials may be generated, for example, a pinpoint targeted base substitution conferring a single amino-acid change, deletion, or substitution of a functional domain to alter the function and/or activity of a target protein (Endo et al., 2016; Schiml and Puchta, 2016; Sun et al., 2016). Appropriate combinations of classical and novel technologies and resources will thus provide novel materials for crop improvement and broaden the potential of cereal breeding.
All authors listed, have made substantial, direct and intellectual contribution to the work, and approved it for publication.
Our studies described in this review were partly supported by the funding for biological resources from National Institute of Agrobiological Sciences and grants from the Ministry of Agriculture, Forestry and Fisheries (MAFF) of Japan: Green Technology Project EF-1004 and Genomics for Agricultural Innovation AMR-0001.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
We thank all of our past and present colleagues who have contributed to the production of transgenic rice resources. We also thank Dr. Seiichi Toki for helpful suggestions and Enago (www.enago.jp) for the English language review.
Abdeeva, I., Abdeev, R., Bruskin, S., and Piruzian, E. (2012). “Transgenic plants as a tool for plant functional genomics,” in Transgenic Plants – Advances and Limitations, ed. Y. O. Çiftçi (Rijeka: InTech), 259–284. doi: 10.5772/33553
Alves, A. C., Worland, B., Thole, V., Snape, J. W., Bevan, M. W., and Vain, P. (2009). A protocol for Agrobacterium-mediated transformation of Brachypodium distachyon community standard line Bd21. Nat. Protoc. 4, 638–649. doi: 10.1038/nprot.2009.30
An, S., Park, S., Jeong, D.-H., Lee, D.-Y., Kang, H.-G., Yu, J.-H., et al. (2003). Generation and analysis of end sequence database for T-DNA tagging lines in rice. Plant Physiol. 133, 2040–2047. doi: 10.1104/pp.103.030478
Asano, T., Hakata, M., Nakamura, H., Aoki, N., Komatsu, S., Ichikawa, H., et al. (2011). Functional characterisation of OsCPK21, a calcium-dependent protein kinase that confers salt tolerance in rice. Plant Mol. Biol. 75, 179–191. doi: 10.1007/s11103-010-9717-1
Ayliffe, M. A., Agostino, A., Clarke, B. C., Furbank, R., von Caemmerer, S., and Pryor, A. J. (2009). Suppression of the barley uroporphyrinogen III synthase gene by a Ds activation tagging element generates developmental photosensitivity. Plant Cell 21, 814–831. doi: 10.1105/tpc.108.063685
Ayliffe, M. A., Pallotta, M., Langridge, P., and Pryor, A. J. (2007). A barley activation tagging system. Plant Mol. Biol. 64, 329–347. doi: 10.1007/s11103-007-9157-8
Basu, S. K., Dutta, M., Goyal, A., Bhowmik, P. K., Kumar, J., Nandy, S., et al. (2010). Is genetically modified crop the answer for the next green revolution? GM Crops 1, 68–79. doi: 10.4161/gmcr.1.2.11877
Belfield, E. J., Brown, C., Gan, X., Jiang, C., Baban, D., Mithani, A., et al. (2014). Microarray-based ultra-high resolution discovery of genomic deletion mutations. BMC Genomics 15:224. doi: 10.1186/1471-2164-15-224
Belfield, E. J., Gan, X., Mithani, A., Brown, C., Jiang, C., Franklin, K., et al. (2012). Genome-wide analysis of mutations in mutant lineages selected following fast-neutron irradiation mutagenesis of Arabidopsis thaliana. Genome Res. 22, 1306–1315. doi: 10.1101/gr.131474.111
Bennetzen, J. L., Schmutz, J., Wang, H., Percifield, R., Hawkins, J., Pontaroli, A. C., et al. (2012). Reference genome sequence of the model plant Setaria. Nat. Biotechnol. 30, 555–561. doi: 10.1038/nbt.2196
Brenchley, R., Spannagl, M., Pfeifer, M., Barker, G. L., D’Amore, R., Allen, A. M., et al. (2012). Analysis of the bread wheat genome using whole-genome shotgun sequencing. Nature 491, 705–710. doi: 10.1038/nature11650
Brown, R. H., Singh, J., Singh, S., Dahleen, L. S., Lemaux, P. G., Stein, N., et al. (2015). Behavior of a modified Dissociation element in barley: a tool for genetic studies and for breeding transgenic barley. Mol. Breed. 35, 85. doi: 10.1007/s11032-015-0193-9
Bruce, M., Hess, A., Bai, J., Mauleon, R., Diaz, M. G., Sugiyama, N., et al. (2009). Detection of genomic deletions in rice using oligonucleotide microarrays. BMC Genomics 10:129. doi: 10.1186/1471-2164-10-129
Brutnell, T. P., Bennetzen, J. L., and Vogel, J. P. (2015). Brachypodium distachyon and Setaria viridis: model genetic systems for the grasses. Annu. Rev. Plant Biol. 66, 465–485. doi: 10.1146/annurev-arplant-042811-105528
Brutnell, T. P., Wang, L., Swartwood, K., Goldschmidt, A., Jackson, D., Zhu, X.-G., et al. (2010). Setaria viridis: a model for C4 photosynthesis. Plant Cell 22, 2537–2544. doi: 10.1105/tpc.110.075309
Carbonell, A., Fahlgren, N., Mitchell, S., Cox, K. L. Jr., Reilly, K. C., Mockler, T. C., et al. (2015). Highly specific gene silencing in a monocot species by artificial microRNAs derived from chimeric miRNA precursors. Plant J. 82, 1061–1075. doi: 10.1111/tpj.12835
Chen, L., Huang, L., Min, D., Phillips, A., Wang, S., Madgwick, P. J., et al. (2012). Development and characterization of a new TILLING population of common bread wheat (Triticum aestivum L.). PLoS ONE 7:e41570. doi: 10.1371/journal.pone.0041570
Chen, S., Jin, W., Wang, M., Zhang, F., Zhou, J., Jia, Q., et al. (2003). Distribution and characterization of over 1000 T-DNA tags in rice genome. Plant J. 36, 105–113. doi: 10.1046/j.1365-313X.2003.01860.x
Chern, C. G., Fan, M. J., Yu, S. M., Hour, A. L., Lu, P. C., Lin, Y. C., et al. (2007). A rice phenomics study – Phenotype scoring and seed propagation of a T-DNA insertion-induced rice mutant population. Plant Mol. Biol. 65, 427–438. doi: 10.1007/s11103-007-9218-z
Clarke, L., and Carbon, J. (1976). A colony bank containing synthetic Col El hybrid plasmids representative of the entire E. coli genome. Cell 9, 91–99. doi: 10.1016/0092-8674(76)90055-6
Cuperous, J. T., Fahlgren, N., and Carrington, J. C. (2011). Evolution and functional diversification of MIRNA genes. Plant Cell 23, 431–442. doi: 10.1105/tpc.110.082784
Draper, J., Mur, L. A. J., Jenkins, G., Ghosh-Biswas, G. C., Bablak, P., Hasterok, R., et al. (2001). Brachypodium distachyon. A new model system for functional genomics in grasses. Plant Physiol. 127, 1539–1555. doi: 10.1104/pp.010196
Dubouzet, J. G., Maeda, S., Sugano, S., Ohtake, M., Hayashi, N., Ichikawa, T., et al. (2011). Screening for resistance against Pseudomonas syringae in rice-FOX Arabidopsis lines identified a putative receptor-like cytoplasmic kinase gene that confers resistance to major bacterial and fungal pathogens in Arabidopsis and rice. Plant Biotechnol. J. 9, 466–485. doi: 10.1111/j.1467-7652.2010.00568.x
Eamens, A., Wang, M. B., Smith, N. A., and Waterhouse, P. M. (2008). RNA silencing in plants: yesterday, today, and tomorrow. Plant Physiol. 147, 456–468. doi: 10.1104/pp.108.117275
Eamens, A. L., Blanchard, C. L., Dennis, E. S., and Upadhyaya, N. M. (2004). A bidirectional gene trap construct suitable for T-DNA and Ds-mediated insertional mutagenesis in rice (Oryza sativa L.). Plant Biotechnol. J. 2, 367–380. doi: 10.1111/j.1467-7652.2004.00081.x
Endo, M., Mikami, M., and Toki, S. (2016). Biallelic gene targeting in rice. Plant Physiol. 170, 667–677. doi: 10.1104/pp.15.01663
Fahlgren, N., Hill, S. T., Carrington, J. C., and Carbonell, A. (2016). P-SAMS: a web site for plant artificial microRNA and synthetic trans-acting small interfering RNA design. Bioinformatics 32, 157–158. doi: 10.1093/bioinformatics/btv534
Fujita, M., Mizukado, S., Fujita, Y., Ichikawa, T., Nakazawa, M., Seki, M., et al. (2007). Identification of stress-tolerance-related transcription-factor genes via mini-scale full-length cDNA Over-eXpressor (FOX) gene hunting system. Biochem. Biophys. Res. Commun. 364, 250–257. doi: 10.1016/j.bbrc.2007.09.124
Gao, X., Liang, W., Yin, C., Ji, S., Wang, H., Su, X., et al. (2010). The SEPALLATA-like gene OsMADS34 is required for rice inflorescence and spikelet development. Plant Physiol. 153, 728–740. doi: 10.1104/pp.110.156711
Garvin, D. F., Gu, Y. Q., Hasterok, R., Hazen, S. P., Jenkins, G., Mockler, T. C., et al. (2008). Development of genetic and genomic research resources for Brachypodium distachyon, a new model system for grass crop research. Crop Sci. 48, S69–S84. doi: 10.2135/cropsci2007.06.0332tpg
Gutjahr, C., Radovanovic, D., Geoffroy, J., Zhang, Q., Siegler, H., Chiapello, M., et al. (2012). The half-size ABC transporters STR1 and STR2 are indispensable for mycorrhizal arbuscule formation in rice. Plant J. 69, 906–920. doi: 10.1111/j.1365-313X.2011.04842.x
Hakata, M., Kuroda, M., Ohsumi, A., Hirose, T., Nakamura, H., Muramatsu, M., et al. (2012). Overexpression of a rice TIFY gene increases grain size through enhanced accumulation of carbohydrates in the stem. Biosci. Biotechnol. Biochem. 76, 2129–2134. doi: 10.1271/bbb.120545
Hakata, M., Nakamura, H., Iida-Okada, K., Miyao, A., Kajikawa, M., Imai-Toki, N., et al. (2010). Production and characterization of a large population of cDNA-overexpressing transgenic rice plants using Gateway-based full-length cDNA expression libraries. Breed. Sci. 60, 575–585. doi: 10.1270/jsbbs.60.575
Hiei, Y., Ishida, Y., and Komari, T. (2014). Progress of cereal transformation technology mediated by Agrobacterium tumefaciens. Front. Plant Sci. 5:628. doi: 10.3389/fpls.2014.00628
Hiei, Y., Ohta, S., Komari, T., and Kumashiro, T. (1994). Efficient transformation of rice (Oryza sativa L.) mediated by Agrobacterium and sequence analysis of the boundaries of the T-DNA. Plant J. 6, 271–282. doi: 10.1046/j.1365-313X.1994.6020271.x
Higuchi-Takeuchi, M., Mori, M., and Matsui, M. (2013). High-throughput analysis of rice genes by means of the heterologous full-length cDNA overexpressor (FOX)-hunting system. Int. J. Dev. Biol. 57, 517–523. doi: 10.1387/ijdb.130176mm
Hiratsu, K., Matsui, K., Koyama, T., and Ohme-Takagi, M. (2003). Dominant repression of target genes by chimeric repressors that include the EAR motif, a repression domain, in Arabidopsis. Plant J. 34, 733–739. doi: 10.1046/j.1365-313X.2003.01759.x
Hirochika, H. (2001). Contribution of the Tos17 retrotransposon to rice functional genomics. Cur. Opin. Plant Biol. 4, 118–122. doi: 10.1016/S1369-5266(00)00146-1
Horiguchi, G. (2004). RNA silencing in plants: a shortcut to functional analysis. Differentiation 72, 65–73. doi: 10.1111/j.1432-0436.2004.07202005.x
Hsing, Y. I., Chern, C. G., Fan, M. J., Lu, P. C., Chen, K. T., Lo, S. F., et al. (2007). A rice gene activation/knockout mutant resource for high throughput functional genomics. Plant Mol. Biol. 63, 351–364. doi: 10.1007/s11103-006-9093-z
Huang, C. F., Yamaji, N., Mitani, N., Yano, M., Nagamura, Y., and Ma, J. F. (2009). A bacterial-type ABC transporter is involved in aluminum tolerance in rice. Plant Cell 21, 655–667. doi: 10.1105/tpc.108.064543
Ichikawa, T., Nakazawa, M., Kawashima, M., Iizumi, H., Kuroda, H., Kondou, Y., et al. (2006). The FOX hunting system: an alternative gain-of-function gene hunting technique. Plant J. 45, 974–985. doi: 10.1111/j.1365-313X.2006.02924.x
Ichikawa, T., Nakazawa, M., Kawashima, M., Muto, S., Gohda, K., Suzuki, K., et al. (2003). Sequence database of 1172 T-DNA insertion sites in Arabidopsis activation-tagging lines that showed phenotypes in T1 generation. Plant J. 36, 421–429. doi: 10.1046/j.1365-313X.2003.01876.x
International Barley Genome Sequencing Consortium (2012). A physical, genetic and functional sequence assembly of the barley genome. Nature 491, 711–716. doi: 10.1038/nature11543
International Brachypodium Initiative (2010). Genome sequencing and analysis of the model grass Brachypodium distachyon. Nature 463, 763–768. doi: 10.1038/nature08747
International Rice Genome Sequencing Project (2005). The map-based sequence of the rice genome. Nature 436, 793–800. doi: 10.1038/nature03895
International Wheat Genome Sequencing Consortium (IWGSC) (2014). A chromosome-based draft sequence of the hexaploid bread wheat (Triticum aestivum) genome. Science 345:1251788. doi: 10.1126/science.1251788
Jeong, D. H., An, S., Kang, H. G., Moon, S., Han, J. J., Park, S., et al. (2002). T-DNA insertional mutagenesis for activation tagging in rice. Plant Physiol. 130, 1636–1644. doi: 10.1104/pp.014357
Jeong, D. H., An, S., Park, S., Kang, H. G., Park, G. G., Kim, S. R., et al. (2006). Generation of a flanking sequence-tag database for activation-tagging lines in japonica rice. Plant J. 45, 123–132. doi: 10.1111/j.1365-313X.2005.02610.x
Jiang, S. Y., Bachmann, D., La, H., Ma, Z., Venkatesh, P. N., Ramamoorthy, R., et al. (2007). Ds insertion mutagenesis as an efficient tool to produce diverse variations for rice breeding. Plant Mol. Biol. 65, 385–402. doi: 10.1007/s11103-007-9233-0
Jiang, S. Y., and Ramachandran, S. (2010). Natural and artificial mutants as valuable resources for functional genomics and molecular breeding. Int. J. Biol. Sci. 6, 228–251. doi: 10.7150/ijbs.6.228
Kakimoto, T. (1996). CKI1, a histidine kinase homolog implicated in cytokinin signal transduction. Science 274, 982–985. doi: 10.1126/science.274.5289.982
Kawaura, K., Mochida, K., Enju, A., Totoki, Y., Toyoda, A., Sakaki, Y., et al. (2009). Assessment of adaptive evolution between wheat and rice as deduced from full-length common wheat cDNA sequence data and expression patterns. BMC Genomics 10:271. doi: 10.1186/1471-2164-10-271
Kazama, D., Itakura, M., Kurusu, T., Mitsuda, N., Ohme-Takagi, M., and Tada, Y. (2013). Identification of chimeric repressors that confer salt and osmotic stress tolerance in Arabidopsis. Plants 2, 769–785. doi: 10.3390/plants2040769
Kellogg, E. A. (2001). Evolutionary history of the grasses. Plant Physiol. 125, 1198–1205. doi: 10.1104/pp.125.3.1198
Kersey, P. J., Allen, J. E., Armean, I., Boddu, S., Bolt, B. J., Carvalho-Silva, D., et al. (2016). Ensembl Genomes 2016: more genomes, more complexity. Nucleic Acids Res. 44, D574–D580. doi: 10.1093/nar/gkv1209
Khush, G. S. (1999). Green revolution: preparing for the 21st century. Genome 42, 646–655. doi: 10.1139/gen-42-4-646
Kim, C. M., Park, S. H., Je, B. I., Park, S. H., Park, S. J., Piao, H. L., et al. (2007). OsCSLD1, a cellulose synthase-like D1 gene, is required for root hair morphogenesis in rice. Plant Physiol. 143, 1220–1230. doi: 10.1104/pp.106.091546
Kim, C. M., Piao, H. L., Park, S. J., Chon, N. S., Je, B. I., Sun, B., et al. (2004). Rapid, large-scale generation of Ds transposant lines and analysis of the Ds insertion sites in rice. Plant J. 39, 252–263. doi: 10.1111/j.1365-313X.2004.02116.x
Kolesnik, T., Szeverenyi, I., Bachmann, D., Kumar, C. S., Jiang, S., Ramamoorthy, R., et al. (2004). Establishing an efficient Ac/Ds tagging system in rice: large-scale analysis of Ds flanking sequences. Plant J. 37, 301–314. doi: 10.1046/j.1365-313X.2003.01948.x
Kondou, Y., Higuchi, M., Ichikawa, T., and Matsui, M. (2011). Application of full-length cDNA resources to gain-of-function technology for characterization of plant gene function. Methods Mol. Biol. 729, 183–197. doi: 10.1007/978-1-61779-065-2_12
Kondou, Y., Higuchi, M., Takahashi, S., Sakurai, T., Ichikawa, T., Kuroda, H., et al. (2009). Systematic approaches to using the FOX hunting system to identify useful rice genes. Plant J. 57, 883–894. doi: 10.1111/j.1365-313X.2008.03733.x
Kozomara, A., and Griffiths-Jones, S. (2014). miRBase: annotating high confidence microRNAs using deep sequencing data. Nucleic Acids Res. 42, D68–D73. doi: 10.1093/nar/gkt1181
Krishnan, A., Guiderdoni, E., An, G., Hsing, Y. I., Han, C. D., Lee, M. C., et al. (2009). Mutant resources in rice for functional genomics of the grasses. Plant Physiol. 149, 165–170. doi: 10.1104/pp.108.128918
Kubo, M., Udagawa, M., Nishikubo, N., Horiguchi, G., Yamaguchi, M., Ito, J., et al. (2005). Transcription switches for protoxylem and metaxylem vessel formation. Genes Dev. 19, 1855–1860. doi: 10.1101/gad.1331305
Kumar, C. S., Wing, R. A., and Sundaresan, V. (2005). Efficient insertional mutagenesis in rice using the maize En/Spm elements. Plant J. 44, 879–892. doi: 10.1111/j.1365-313X.2005.02570.x
Kurata, N., Miyoshi, K., Nonomura, K. I., Yamazaki, Y., and Ito, Y. (2005). Rice mutants and genes related to organ development, morphogenesis and physiological traits. Plant Cell Physiol. 46, 48–62. doi: 10.1093/pcp/pci506
Kurotani, K., Hayashi, K., Hatanaka, S., Toda, Y., Ogawa, D., Ichikawa, H., et al. (2015a). Elevated levels of CYP94 family gene expression alleviate the jasmonate response and enhance salt tolerance in rice. Plant Cell Physiol. 56, 779–789. doi: 10.1093/pcp/pcv006
Kurotani, K., Yamanaka, K., Toda, Y., Ogawa, D., Tanaka, M., Kozawa, H., et al. (2015b). Stress tolerance profiling of a collection of extant salt-tolerant rice varieties and transgenic plants overexpressing abiotic stress tolerance genes. Plant Cell Physiol. 56, 1867–1876. doi: 10.1093/pcp/pcv106
Kurowska, M., Labocha-Pawłowska, A., Gnizda, D., Maluszynski, M., and Szarejko, I. (2012). Molecular analysis of point mutations in a barley genome exposed to MNU and gamma rays. Mutat. Res. 73, 52–70. doi: 10.1016/j.mrfmmm.2012.08.008
Lee, S., Ryoo, N., Jeon, J. S., Guerinot, M. L., and An, G. (2012). Activation of rice Yellow Stripe1-Like 16 (OsYSL16) enhances iron efficiency. Mol. Cells 33, 117–126. doi: 10.1007/s10059-012-2165-9
Lee, Y.-S., Jeong, D.-H., Lee, D.-Y., Yi, J., Ryu, C.-H., Kim, S. L., et al. (2010). OsCOL4 is a constitutive flowering repressor upstream of Ehd1 and downstream of OsphyB. Plant J. 63, 18–30. doi: 10.1111/j.1365-313X.2010.04226.x
Li, P., and Brutnell, T. P. (2011). Setaria viridis and Setaria italica, model genetic systems for the Panicoid grasses. J. Exp. Bot. 62, 3031–3037. doi: 10.1093/jxb/err096
Li, X., Song, Y., Century, K., Straight, S., Ronald, P., Dong, X., et al. (2001). A fast neutron deletion mutagenesis-based reverse genetics system for plants. Plant J. 27, 235–242. doi: 10.1046/j.1365-313x.2001.01084.x
Lu, G., Wang, X., Liu, J., Yu, K., Gao, Y., Liu, H., et al. (2014). Application of T-DNA activation tagging to identify glutamate receptor-like genes that enhance drought tolerance in plants. Plant Cell Rep. 33, 617–631. doi: 10.1007/s00299-014-1586-7
Martins, P. K., Nakayama, T. J., Ribeiro, A. P., Dias Brito da Cunha, B. A., Nepomuceno, A. L., Harmon, F. G., et al. (2015a). Setaria viridis floral-dip: a simple and rapid Agrobacterium-mediated transformation method. Biotechnol. Rep. 6, 61–63. doi: 10.1016/j.btre.2015.02.006
Martins, P. K., Ribeiro, A. P., Dias Brito da Cunha, B. A., Kobayashi, A. K., and Molinari, H. B. C. (2015b). A simple and highly efficient Agrobacterium-mediated transformation protocol for Setaria viridis. Biotechnol. Rep. 6, 41–44. doi: 10.1016/j.btre.2015.02.002
Matsumoto, T., Tanaka, T., Sakai, H., Amano, N., Kanamori, H., Kurita, K., et al. (2011). Comprehensive sequence analysis of 24,783 barley full-length cDNAs derived from 12 clone libraries. Plant Physiol. 156, 20–28. doi: 10.1104/pp.110.171579
Michael, T. P., and VanBuren, R. (2015). Progress, challenges and the future of crop genomes. Curr. Opin. Plant Biol. 24, 71–81. doi: 10.1016/j.pbi.2015.02.002
Mito, T., Seki, M., Shinozaki, K., Ohme-Takagi, M., and Matsui, K. (2011). Generation of chimeric repressors that confer salt tolerance in Arabidopsis and rice. Plant Biotechnol. J. 9, 736–746. doi: 10.1111/j.1467-7652.2010.00578.x
Mitsuda, N., Hiratsu, K., Todaka, D., Nakashima, K., Yamaguchi-Shinozaki, K., and Ohme-Takagi, M. (2006). Efficient production of male and female sterile plants by expression of a chimeric repressor in Arabidopsis and rice. Plant Biotechnol. J. 4, 325–332. doi: 10.1111/j.1467-7652.2006.00184.x
Mitsuda, N., Matsui, K., Ikeda, M., Nakata, M., Oshima, Y., Nagatoshi, Y., et al. (2011a). CRES-T, an effective gene silencing system utilizing chimeric repressors. Methods Mol. Biol. 754, 87–105. doi: 10.1007/978-1-61779-154-3_5
Mitsuda, N., Takiguchi, Y., Shikata, M., Sage-Ono, K., Ono, M., Sasaki, K., et al. (2011b). The new fioreDB database provides comprehensive information on plant transcription factors and phenotypes induced by CRES-T in ornamental and model plants. Plant Biotechnol. 28, 123–130. doi: 10.5511/plantbiotechnology.11.0106a
Miyao, A., Iwasaki, Y., Kitano, H., Itoh, J. I., Maekawa, M., Murata, K., et al. (2007). A large-scale collection of phenotypic data describing an insertional mutant population to facilitate functional analysis of rice genes. Plant Mol. Biol. 63, 625–635. doi: 10.1007/s11103-006-9118-7
Miyao, A., Tanaka, K., Murata, K., Sawaki, H., Takeda, S., Abe, K., et al. (2003). Target site specificity of the Tos17 retrotransposon shows a preference for insertion within genes and against insertion in retrotransposon-rich regions of the genome. Plant Cell 15, 1771–1780. doi: 10.1105/tpc.012559
Mochida, K., and Shinozaki, K. (2010). Genomics and bioinformatics resources for crop improvement. Plant Cell Physiol. 51, 497–523. doi: 10.1093/pcp/pcq027
Mochida, K., Uehara-Yamaguchi, Y., Takahashi, F., Yoshida, T., Sakurai, T., and Shinozaki, K. (2013). Large-scale collection and analysis of full-length cDNAs from Brachypodium distachyon and integration with Pooideae sequence resources. PLoS ONE 8:e75265. doi: 10.1371/journal.pone.0075265
Mochida, K., Yoshida, T., Sakurai, T., Ogihara, Y., and Shinozaki, K. (2009). TriFLDB: a database of clustered full-length coding sequences from Triticeae with applications to comparative grass genomics. Plant Physiol. 150, 1135–1146. doi: 10.1104/pp.109.138214
Mori, M., Tomita, C., Sugimoto, K., Hasegawa, M., Hayashi, N., Dubouzet, J. G., et al. (2007). Isolation and molecular characterization of a Spotted leaf 18 mutant by modified activation-tagging in rice. Plant Mol. Biol. 63, 847–860. doi: 10.1007/s11103-006-9130-y
Morita, R., Kusaba, M., Iida, S., Yamaguchi, H., Nishio, T., and Nishimura, M. (2009). Molecular characterization of mutations induced by gamma irradiation in rice. Genes Genet. Syst. 84, 361–370. doi: 10.1266/ggs.84.361
Nakagawa, H., Tanaka, A., Tanabata, T., Ohtake, M., Fujioka, S., Nakamura, H., et al. (2012). SHORT GRAIN1 decreases organ elongation and brassinosteroid response in rice. Plant Physiol. 158, 1208–1219. doi: 10.1104/pp.111.187567
Nakamura, H., Hakata, M., Amano, K., Miyao, A., Toki, N., Kajikawa, M., et al. (2007). A genome-wide gain-of-function analysis of rice genes using the FOX-hunting system. Plant Mol. Biol. 65, 357–371. doi: 10.1007/s11103-007-9243-y
Nakamura, H., Muramatsu, M., Hakata, M., Ueno, O., Nagamura, Y., Hirochika, H., et al. (2009). Ectopic overexpression of the transcription factor OsGLK1 induces chloroplast development in non-green rice cells. Plant Cell Physiol. 50, 1933–1949. doi: 10.1093/pcp/pcp138
Ning, J., Zhang, B., Wang, N., Zhou, Y., and Xiong, L. (2011). Increased leaf angle1, a Raf-like MAPKKK that interacts with a nuclear protein family, regulates mechanical tissue formation in the Lamina joint of rice. Plant Cell 23, 4334–4347. doi: 10.1105/tpc.111.093419
Nishizawa-Yokoi, A., Cermak, T., Hoshino, T., Sugimoto, K., Saika, H., Mori, A., et al. (2016). A defect in DNA Ligase4 enhances the frequency of TALEN-mediated targeted mutagenesis in rice. Plant Physiol. 170, 653–666. doi: 10.1104/pp.15.01542
Ogawa, D., Abe, K., Miyao, A., Kojima, M., Sakakibara, H., Mizutani, M., et al. (2011). RSS1 regulates the cell cycle and maintains meristematic activity under stress conditions in rice. Nat. Commun. 2:278. doi: 10.1038/ncomms1279
Ogo, Y., Kobayashi, T., Nakanishi Itai, R., Nakanishi, H., Kakei, Y., Takahashi, M., et al. (2008). A novel NAC transcription factor, IDEF2, that recognizes the iron deficiency-responsive element 2 regulates the genes involved in iron homeostasis in plants. J. Biol. Chem. 283, 13407–13417. doi: 10.1074/jbc.M708732200
Opanowicz, M., Vain, P., Draper, J., Parker, D., and Doonan, J. H. (2008). Brachypodium distachyon: making hay with a wild grass. Trends Plant Sci. 13, 172–177. doi: 10.1016/j.tplants.2008.01.007
Osakabe, Y., and Osakabe, K. (2015). Genome editing with engineered nucleases in plants. Plant Cell Physiol. 56, 389–400. doi: 10.1093/pcp/pcu170
Park, S. H., Jun, N. S., Kim, C. M., Oh, T. Y., Huang, J., Xuan, Y. H., et al. (2007). Analysis of gene-trap Ds rice populations in Korea. Plant Mol. Biol. 65, 373–384. doi: 10.1007/s11103-007-9192-5
Paterson, A. H., Bowers, J. E., Bruggmann, R., Dubchak, I., Grimwood, J., Gundlach, H., et al. (2009). The Sorghum bicolor genome and the diversification of grasses. Nature 457, 551–556. doi: 10.1038/nature07723
Qu, S., Desai, A., Wing, R., and Sundaresan, V. (2008). A versatile transposon-based activation tag vector system for functional genomics in cereals and other monocot plants. Plant Physiol. 146, 189–199. doi: 10.1104/pp.107.111427
Ramamoorthy, R., Jiang, S. Y., and Ramachandran, S. (2011). Oryza sativa cytochrome P450 family member OsCYP96B4 reduces plant height in a transcript dosage dependent manner. PLoS ONE 6:e28069. doi: 10.1371/journal.pone.0028069
Rawat, N., Sehgal, S. K., Joshi, A., Rothe, N., Wilson, D. L., McGraw, N., et al. (2012). A diploid wheat TILLING resource for wheat functional genomics. BMC Plant Biol. 12:205. doi: 10.1186/1471-2229-12-205
Rice Full-Length cDNA Consortium (2003). Collection, mapping, and annotation of over 28,000 cDNA clones from japonica rice. Science 301, 376–379. doi: 10.1126/science.1081288
Riechmann, J. L., Heard, J., Martin, G., Reuber, L., Jiang, C., Keddie, J., et al. (2000). Arabidopsis transcription factors: genome-wide comparative analysis among eukaryotes. Science 290, 2105–2110. doi: 10.1126/science.290.5499.2105
Rogers, C., Wen, J., Chen, R., and Oldroyd, G. (2009). Deletion-based reverse genetics in Medicago truncatula. Plant Physiol. 151, 1077–1086. doi: 10.1104/pp.109.142919
Ryu, C. H., You, J. H., Kang, H. G., Hur, J., Kim, Y. H., Han, M. J., et al. (2004). Generation of T-DNA tagging lines with a bidirectional gene trap vector and the establishment of an insertion-site database. Plant Mol. Biol. 54, 489–502. doi: 10.1023/B:PLAN.0000038257.93381.05
Sallaud, C., Gay, C., Larmande, P., Bès, M., Piffanelli, P., Piégu, B., et al. (2004). High throughput T-DNA insertion mutagenesis in rice: a first step towards in silico reverse genetics. Plant J. 39, 450–464. doi: 10.1111/j.1365-313X.2004.02145.x
Sato, K., Shin-I, T., Seki, M., Shinozaki, K., Yoshida, H., Takeda, K., et al. (2009). Development of 5006 full-length CDNAs in barley: a tool for accessing cereal genomics resources. DNA Res. 16, 81–89. doi: 10.1093/dnares/dsn034
Satoh, H., Matsusaka, H., and Kumamaru, T. (2010). Use of N-methyl-N-nitrosourea treatment of fertilized egg cells for saturation mutagenesis of rice. Breed. Sci. 60, 475–485. doi: 10.1270/jsbbs.60.475
Schiml, S., and Puchta, H. (2016). Revolutionizing plant biology: multiple ways of genome engineering by CRISPR/Cas. Plant Methods 12, 8. doi: 10.1186/s13007-016-0103-0
Schmidt, R., Mieulet, D., Hubberten, H. M., Obata, T., Hoefgen, R., Fernie, A. R., et al. (2013). SALT-RESPONSIVE ERF1 regulates reactive oxygen species-dependent signaling during the initial response to salt stress in rice. Plant Cell 25, 2115–2131. doi: 10.1105/tpc.113.113068
Schnable, P. S., Ware, D., Fulton, R. S., Stein, J. C., Wei, F., Pasternak, S., et al. (2009). The B73 maize genome: complexity, diversity, and dynamics. Science 326, 1112–1115. doi: 10.1126/science.1178534
Shu, Q. Y., Shirasawa, K., Hoffmann, M., Hurlebaus, J., and Nishio, T. (2012). “Molecular techniques and methods for mutation detection and screening in plants,” in Plant Mutation Breeding and Biotechnology, eds Q. Y. Shu, B. P. Forster, and H. Nakagawa (Vienna: Joint FAO/IAEA Programme), 241–256.
Soderlund, C., Descour, A., Kudrna, D., Bomhoff, M., Boyd, L., Currie, J., et al. (2009). Sequencing, mapping, and analysis of 27,455 maize full-length cDNAs. PLoS Genet. 5:e1000740. doi: 10.1371/journal.pgen.1000740
Sumiyoshi, M., Nakamura, A., Nakamura, H., Hakata, M., Ichikawa, H., Hirochika, H., et al. (2013). Increase in cellulose accumulation and improvement of saccharification by overexpression of arabinofuranosidase in rice. PLoS ONE 8:e78269. doi: 10.1371/journal.pone.0078269
Sun, G. (2012). MicroRNAs and their diverse functions in plants. Plant Mol. Biol. 80, 17–36. doi: 10.1007/s11103-011-9817-6
Sun, Y., Zhang, X., Wu, C., He, Y., Ma, Y., Hou, H., et al. (2016). Engineering herbicide-resistant rice plants through CRISPR/Cas9-mediated homologous recombination of acetolactate synthase. Mol. Plant 9, 628–631. doi: 10.1016/j.molp.2016.01.001
Suzuki, T., Eiguchi, M., Kumamaru, T., Satoh, H., Matsusaka, H., Moriguchi, K., et al. (2008). MNU-induced mutant pools and high performance TILLING enable finding of any gene mutation in rice. Mol. Genet. Genomics 279, 213–223. doi: 10.1007/s00438-007-0293-2
Tanaka, W., Toriba, T., Ohmori, Y., Yoshida, A., Kawai, A., Mayama-Tsuchida, T., et al. (2012). The YABBY gene TONGARI-BOUSHI1 is involved in lateral organ development and maintenance of meristem organization in the rice spikelet. Plant Cell 24, 80–95. doi: 10.1105/tpc.111.094797
Tello-Ruiz, M. K., Stein, J., Wei, S., Preece, J., Olson, A., Naithani, S., et al. (2016). Gramene 2016: comparative plant genomics and pathway resources. Nucleic Acids Res. 44, D1133–D1140. doi: 10.1093/nar/gkv1179
Thole, V., Peraldi, A., Worland, B., Nicholson, P., Doonan, J. H., and Vain, P. (2012). T-DNA mutagenesis in Brachypodium distachyon. J. Exp. Bot. 63, 567–576. doi: 10.1093/jxb/err333
Thole, V., and Vain, P. (2012). Agrobacterium-mediated transformation of Brachypodium distachyon. Methods Mol. Biol. 847, 137–149. doi: 10.1007/978-1-61779-558-9_12
Till, B. J., Cooper, J., Tai, T. H., Colowit, P., Greene, E. A., Henikoff, S., et al. (2007). Discovery of chemically induced mutations in rice by TILLING. BMC Plant Biol. 7:19. doi: 10.1186/1471-2229-7-19
Till, B. J., Reynolds, S. H., Greene, E. A., Codomo, C. A., Enns, L. C., Johnson, J. E., et al. (2003). Large-scale discovery of induced point mutations with high-throughput TILLING. Genome Res. 13, 524–530. doi: 10.1101/gr.977903
Todaka, D., Nakashima, K., Maruyama, K., Kidokoro, S., Osakabe, Y., Ito, Y., et al. (2012). Rice phytochrome-interacting factor-like protein OsPIL1 functions as a key regulator of internode elongation and induces a morphological response to drought stress. Proc. Natl. Acad. Sci. U.S.A. 109, 15947–15952. doi: 10.1073/pnas.1207324109
Toki, S., Hara, N., Ono, K., Onodera, H., Tagiri, A., Oka, S., et al. (2006). Early infection of scutellum tissue with Agrobacterium allows high-speed transformation of rice. Plant J. 47, 969–976. doi: 10.1111/j.1365-313X.2006.02836.x
Tsuchida-Mayama, T., Nakamura, H., Hakata, M., and Ichikawa, H. (2010). Rice transgenic resources with gain-of-function phenotypes. Breed. Sci. 60, 493–501. doi: 10.1270/jsbbs.60.493
Tsuda, M., Kaga, A., Anai, T., Shimizu, T., Sayama, T., Takagi, K., et al. (2015). Construction of a high-density mutant library in soybean and development of a mutant retrieval method using amplicon sequencing. BMC Genomics 16:1014. doi: 10.1186/s12864-015-2079-y
Upadhyaya, N. M., Zhou, X.-R., Zhu, Q.-H., Ramm, K., Wu, L., Eamens, A., et al. (2002). An iAc/Ds gene and enhancer trapping system for insertional mutagenesis in rice. Funct. Plant Biol. 29, 547–559. doi: 10.1071/PP01205
Van Eck, J., and Swartwood, K. (2015). “Setaria viridis,” in Methods in Molecular Biology, Vol. 1223: Agrobacterium Protocols, Vol. 1 , ed. K. Wang (New York, NY: Springer Science+Business Media), 57–67. doi: 10.1007/978-1-4939-1695-5_5
van Enckevort, L. J. G., Droc, G., Piffanelli, P., Greco, R., Gagneur, C., Weber, C., et al. (2005). EU-OSTID: a collection of transposon insertional mutants for functional genomics in rice. Plant Mol. Biol. 59, 99–110. doi: 10.1007/s11103-005-8532-6
Walden, R., Fritze, K., Hayashi, H., Miklashevichs, E., Harling, H., and Schell, J. (1994). Activation tagging: a means of isolating genes implicated as playing a role in plant growth and development. Plant Mol. Biol. 26, 1521–1528. doi: 10.1007/BF00016488
Wan, S., Wu, J., Zhang, Z., Sun, X., Lv, Y., Gao, C., et al. (2009). Activation tagging, an efficient tool for functional analysis of the rice genome. Plant Mol. Biol. 69, 69–80. doi: 10.1007/s11103-008-9406-5
Wang, L., Zheng, J., Luo, Y., Xu, T., Zhang, Q., Zhang, L., et al. (2013). Construction of a genomewide RNAi mutant library in rice. Plant Biotechnol. J. 11, 997–1005. doi: 10.1111/pbi.12093
Wang, P., Fouracre, J., Kelly, S., Karki, S., Gowik, U., Aubry, S., et al. (2013). Evolution of GOLDEN2-LIKE gene function in C3 and C4 plants. Planta 237, 481–495. doi: 10.1007/s00425-012-1754-3
Wang, T. L., Uauy, C., Robson, F., and Till, B. (2012). TILLING in extremis. Plant Biotechnol. J. 10, 761–772. doi: 10.1111/j.1467-7652.2012.00708.x
Warthmann, N., Chen, H., Ossowski, S., Weigel, D., and Hervé, P. (2008). Highly specific gene silencing by artificial miRNAs in rice. PLoS ONE 3:e1829. doi: 10.1371/journal.pone.0001829
Weigel, D., Ahn, J. H., Blázquez, M. A., Borevitz, J. O., Christensen, S. K., Fankhauser, C., et al. (2000). Activation tagging in Arabidopsis. Plant Physiol. 122, 1003–1013. doi: 10.1104/pp.122.4.1003
Wu, J., Zhang, Z., Zhang, Q., Liu, Y., Zhu, B., Cao, J., et al. (2015). Generation of wheat transcription factor FOX rice lines and systematic screening for salt and osmotic stress tolerance. PLoS ONE 10:e0132314. doi: 10.1371/journal.pone.0132314
Wu, J. L., Wu, C., Lei, C., Baraoidan, M., Bordeos, A., Madamba, M. R. S., et al. (2005). Chemical- and irradiation-induced mutants of indica rice IR64 for forward and reverse genetics. Plant Mol. Biol. 59, 85–97. doi: 10.1007/s11103-004-5112-0
Yamaguchi, H., Hase, Y., Tanaka, A., Shikazono, N., Degi, K., Shimizu, A., et al. (2009). Mutagenic effects of ion beam irradiation on rice. Breed. Sci. 59, 169–177. doi: 10.1270/jsbbs.59.169
Yang, S. Y., Grønlund, M., Jakobsen, I., Grotemeyer, M. S., Rentsch, D., Miyao, A., et al. (2012). Nonredundant regulation of rice arbuscular mycorrhizal symbiosis by two members of the PHOSPHATE TRANSPORTER1 gene family. Plant Cell 24, 4236–4251. doi: 10.1105/tpc.112.104901
Yang, W., Ren, S., Zhang, X., Gao, M., Ye, S., Qi, Y., et al. (2011). BENT UPPERMOST INTERNODE1 encodes the class II formin FH5 crucial for actin organization and rice development. Plant Cell 23, 661–680. doi: 10.1105/tpc.110.081802
Yang, Y., Peng, H., Huang, H., Wu, J., Jia, S., Huang, D., et al. (2004). Large-scale production of enhancer trapping lines for rice functional genomics. Plant Sci. 167, 281–288. doi: 10.1016/j.plantsci.2004.03.026
Yi, J., Lee, Y. S., Lee, D. Y., Cho, M. H., Jeon, J. S., and An, G. (2016a). OsMPK6 plays a critical role in cell differentiation during early embryogenesis in Oryza sativa. J. Exp. Bot. 67, 2425–2437. doi: 10.1093/jxb/erw052
Yi, J., Moon, S., Lee, Y. S., Zhu, L., Liang, W., Zhang, D., et al. (2016b). Defective Tapetum Cell Death 1 (DTC1) regulates ROS levels by binding to metallothionein during tapetum degeneration. Plant Physiol. 170, 1611–1623. doi: 10.1104/pp.15.01561
Yokotani, N., Higuchi, M., Kondou, Y., Ichikawa, T., Iwabuchi, M., Hirochika, H., et al. (2011). A novel chloroplast protein, CEST induces tolerance to multiple environmental stresses and reduces photooxidative damage in transgenic Arabidopsis. J. Exp. Bot. 62, 557–569. doi: 10.1093/jxb/erq290
Yokotani, N., Ichikawa, T., Kondou, Y., Iwabuchi, M., Matsui, M., Hirochika, H., et al. (2013). Role of the rice transcription factor JAmyb in abiotic stress response. J. Plant Res. 126, 131–139. doi: 10.1007/s10265-012-0501-y
Yokotani, N., Ichikawa, T., Kondou, Y., Maeda, S., Iwabuchi, M., Mori, M., et al. (2009). Overexpression of a rice gene encoding a small C2 domain protein OsSMCP1 increases tolerance to abiotic and biotic stresses in transgenic Arabidopsis. Plant Mol. Biol. 71, 391–402. doi: 10.1007/s11103-009-9530-x
Yokotani, N., Ichikawa, T., Kondou, Y., Matsui, M., Hirochika, H., Iwabuchi, M., et al. (2008). Expression of rice heat stress transcription factor OsHsfA2e enhances tolerance to environmental stresses in transgenic Arabidopsis. Planta 227, 957–967. doi: 10.1007/s00425-007-0670-4
Yoshida, K., Sakamoto, S., Kawai, T., Kobayashi, Y., Sato, K., Ichinose, Y., et al. (2013). Engineering the Oryza sativa cell wall with rice NAC transcription factors regulating secondary wall formation. Front. Plant Sci. 4:383. doi: 10.3389/fpls.2013.00383
Zhang, G., Liu, X., Quan, Z., Cheng, S., Xu, X., Pan, S., et al. (2012). Genome sequence of foxtail millet (Setaria italica) provides insights into grass evolution and biofuel potential. Nat. Biotechnol. 30, 549–554. doi: 10.1038/nbt.2195
Zhang, H., Zhang, J., Wei, P., Zhang, B., Gou, F., Feng, Z., et al. (2014). The CRISPR/Cas9 system produces specific and homozygous targeted gene editing in rice in one generation. Plant Biotechnol. J. 12, 797–807. doi: 10.1111/pbi.12200
Zhang, J., Li, C., Wu, C., Xiong, L., Chen, G., Zhang, Q., et al. (2006). RMD: a rice mutant database for functional analysis of the rice genome. Nucleic Acids Res. 34, D745–D748. doi: 10.1093/nar/glj016
Zhang, J. Z. (2003). Overexpression analysis of plant transcription factors. Curr. Opin. Plant Biol. 6, 430–440. doi: 10.1016/S1369-5266(03)00081-5
Zheng, L. L., and Qu, L. H. (2015). Application of microRNA gene resources in the improvement of agronomic traits in rice. Plant Biotechnol. J. 13, 329–336. doi: 10.1111/pbi.12321s
Zhu, Q. H., Ramm, K., Shivakkumar, R., Dennis, E. S., and Upadhyaya, N. M. (2004). The ANTHER INDEHISCENCE1 gene encoding a single MYB domain protein is involved in anther development in rice. Plant Physiol. 135, 1514–1525. doi: 10.1104/pp.104.041459
Zhu, Q. H., Upadhyaya, N., and Helliwell, C. (2012). “Transposon mutagenesis for functional genomics,” in Plant Mutation Breeding and Biotechnology, eds Q. Y. Shu, B. P. Forster, and H. Nakagawa (Vienna: Joint FAO/IAEA Programme), 507–522.
Keywords: cereals, rice, transgenic plants, resources, overexpression, FOX hunting, chimeric repressors, activation tagging
Citation: Abe K and Ichikawa H (2016) Gene Overexpression Resources in Cereals for Functional Genomics and Discovery of Useful Genes. Front. Plant Sci. 7:1359. doi: 10.3389/fpls.2016.01359
Received: 18 May 2016; Accepted: 26 August 2016;
Published: 21 September 2016.
Edited by:
Toshihiko Komari, Japan Tobacco Inc., JapanReviewed by:
Narayana Mithur Upadhyaya, Commonwealth Scientific and Industrial Research Organisation, AustraliaCopyright © 2016 Abe and Ichikawa. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Hiroaki Ichikawa, aGljaGt3QGFmZnJjLmdvLmpw
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.