 Zhi-Zhen Fang
Zhi-Zhen Fang Dan-Rong Zhou
Dan-Rong Zhou
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Plant Sci. , 31 August 2016
Sec. Plant Breeding
Volume 7 - 2016 | https://doi.org/10.3389/fpls.2016.01338
Anthocyanins are important pigments and are responsible for red coloration in plums. However, little is known about the molecular mechanisms underlying anthocyanin accumulation in plum fruits. In this study, the RNA-seq technique was used to analyze the transcriptomic changes during fruit ripening in the red-fleshed plum (Prunus salicina Lindl.) cultivar ‘Furongli’. Over 161 million high-quality reads were assembled into 52,093 unigenes and 49.4% of these were annotated using public databases. Of these, 25,681 unigenes had significant hits to the sequences in the NCBI Nr database, 17,203 unigenes showed significant similarity to known proteins in the Swiss-Prot database and 5816 and 8585 unigenes had significant similarity to existing sequences in the Kyoto Encyclopedia of Genes and Genomes and the Cluster of Orthologous Groups databases, respectively. A total of 3548 unigenes were differentially expressed during fruit ripening and 119 of these were annotated as involved in “biosynthesis of other secondary metabolites.” Biological pathway analysis and gene ontology term enrichment analysis revealed that 13 differentially expressed genes are involved in anthocyanin biosynthesis. Furthermore, transcription factors such as MYB and bHLH, which may control anthocyanin biosynthesis, were identified through coexpression analysis of transcription factors, and structural genes. Real-time qPCR analysis of candidate genes showed good correlation with the transcriptome data. These results contribute to our understanding of the molecular mechanisms underlying anthocyanin biosynthesis in plum flesh. The transcriptomic data generated in this study provide a basis for further studies of fruit ripening in plum.
The plum is one of the traditional fruit trees in China and is widely distributed in the world (Carrasco et al., 2012). Its fruit is highly appreciated by consumers. Plums are rich in bioactive substances such as vitamin C, carotenoids, polyphenols, and anthocyanins (Valero et al., 2013) and have great health benefits (Hooshmand and Arjmandi, 2009; Lee et al., 2009; Shukitt-Hale et al., 2009; Johnson et al., 2011). Color is one of the most important determinants of fruit quality. As a result of the influence of culture, Chinese people prefer red color. Anthocyanin accumulation is responsible for red coloration in plums (Cheng et al., 2015). Anthocyanins are widespread secondary metabolites that play an important role in the pigmentation of fruits. They are powerful antioxidants and naturally-occurring dietary anthocyanins are beneficial to human health (Nemie-Feyissa et al., 2015). Santhakumar et al. (2015) demonstrated that consumption of anthocyanin-rich plum juice attenuates thrombogenesis by reducing platelet activation/hyper-coagulability and oxidative stress. It is therefore, desirable to increase the anthocyanin content in plums, especially in the flesh, through improved cultivation methods, postharvest handling, or breeding.
The anthocyanin biosynthetic pathway has been well studied in many plants. Structural genes in the anthocyanin pathway, including those encoding phenylalanine ammonialyase (PAL), cinnamate-4-hydroxylase (C4H), 4-coumaroyl:CoA-ligase (4CL), chalcone synthase (CHS), chalcone isomerase (CHI), flavanone 3-hydroxylase (F3H), flavonoid 3′-hydroxylase (F3′H), flavonoid 3′,5′-hydroxylase, dihydroflavonol 4-reductase (DFR), anthocyanidin synthase/leucoanthocyanidin dioxygenase (ANS/LDOX), and UDP-glucose: flavonoid 3-O-glucosyltransferase (UFGT), have been isolated and characterized in plants (Tanaka et al., 2008). After the synthesizing in the cytosol, anthocyanins must be transported to the vacuole to exhibit their brilliant colors (Winkel-Shirley, 2001). This process is mediated by proteins including glutathione S-transferase (GST), multidrug and toxic compound extrusion, and ATP-binding cassette transporters (Hu et al., 2016).
Anthocyanin biosynthesis is cooperatively regulated by transcriptional regulators including MYB proteins, basic helix-loop-helix (bHLH) proteins, and WD40 proteins (Lepiniec et al., 2006; Feller et al., 2011). These regulators form an MBW complex that binds to promoters and activates transcription of structural genes of the anthocyanin biosynthetic pathway (Rahim et al., 2014; Li et al., 2016). The role of MBW in anthocyanin biosynthesis has been elucidated in fruit trees such as grape (Kobayashi et al., 2002, 2004; Walker et al., 2007), Chinese bayberry (Niu et al., 2010; Liu et al., 2013a,b), mangosteen (Palapol et al., 2009), blood orange (Crifò et al., 2011; Butelli et al., 2012), kiwifruit (Fraser et al., 2013), litchi (Lai et al., 2014, 2016), sweet cherry (Shen et al., 2014), apple (Takos et al., 2006; Espley et al., 2007; Xie et al., 2012; Chagné et al., 2013; An et al., 2015), and peach (Rahim et al., 2014; Zhou et al., 2014; Tuan et al., 2015).
In addition, other regulatory factors also affect anthocyanin biosynthesis via interaction with MBW complexex or by modulating the transcription of structural genes directly. MYBs that act as repressors of the anthocyanin pathway have been identified in several plants (Matsui et al., 2008; Salvatierra et al., 2013; Xu et al., 2013; Huang et al., 2014; Jun et al., 2015; Yoshida et al., 2015). Shin et al. (2007) found that PIF3 and HY5 regulated anthocyanin synthesis by binding directly to promoters of anthocyanin biosynthetic genes in Arabidopsis thaliana. The Arabidopsis COP1/SPA complex represses anthocyanin accumulation by degrading PAP1 and PAP2 proteins (Maier et al., 2013; Maier and Hoecker, 2014). Li et al. (2012b) also demonstrated that MdCOP1 negatively regulates accumulation of anthocyanin in apple peel by modulating the degradation of the MdMYB1 protein. ANAC078 has been shown to promote accumulation of anthocyanins by inducing the expression of flavonoid biosynthesis genes under high-light (Morishita et al., 2009). Recently, Zhou et al. (2015) found that NAC transcription factor BL, which controls the red-fleshed trait, interacts with PpNAC1 to form a heterodimer, and activate the transcription of PpMYB10.1 to induce anthocyanin accumulation. This process is repressed by a SQUAMOSA promoter-binding protein-like (SPL) transcription factor PpSPL1. SPLs have been shown to inhibit the expression of anthocyanin biosynthetic genes and negatively regulate anthocyanin accumulation through destabilization of MBW (Gou et al., 2011). MADS-box genes are reported to be involved in regulation of anthocyanin accumulation (Jaakola et al., 2010; Wu et al., 2013). Jasmonates induce anthocyanin biosynthesis through degradation of jasmonate-ZIM-domain (JAZ) proteins and the subsequent release of MBW (Qi et al., 2011). DELLA proteins promote anthocyanin biosynthesis by sequestering MYBL2 and JAZ proteins, which repress the activity of MBW, to release bHLH/MYB subunits in Arabidopsis (Xie et al., 2016). Furthermore, epigenetic mechanisms also play important roles in anthocyanin biosynthesis (Wang et al., 2013; Zabala and Vodkin, 2014).
The ‘Furongli’ plum (Prunus salicina Lindl.), a red-skinned and red-fleshed cultivar, is native to Fujian, where it has been cultivated for more than 700 years. Fruit of ‘Furongli’ is popular for its attractive color, delicious taste, and health-promoting nutrients (Fang et al., 2014). The fruit can be eaten fresh and is used for the production of candied fruits. In recent years, RNA-seq-based transcriptome analysis has been extensively used for identification of functional genes in fruit trees. Rodamilans et al. (2014) analyzed the transcriptome changes in response to Plum pox virus infection in P. cerasifera. Jo et al. (2015) reported the leaf transcriptome assembly of two different P. salicina cultivars. More recently, González et al. (2016a) reported the fruit skin transcriptomes of two Japanese plum cultivars with different skin color and developed candidate EST–SSR markers for marker-assisted selection of fruit skin color in the Japanese plum (González et al., 2016b). In the present study, we analyzed the transcriptomic changes during the ripening of ‘Furongli’ plum fruits to identify candidate genes involved in the biosynthesis of anthocyanins. Based on the RNA-seq datasets generated, we identify several potential structural genes, and transcription factor genes related to anthocyanin biosynthesis. The transcriptomic data generated in this study provide a basis for further studies of fruit ripening in plum, and identify candidate genes involved in sugar accumulation, organic acid degradation, fruit softening, and pigmentation.
All samples were collected from 5-year-old field grown ‘Furongli’ plum (Prunus salicina Lindl.) trees in an orchard in Yongtai County, Fujian Province, China. Fruit samples were harvested at 105, 115, 125, and 135 days after flowering (DAF) from three trees in 2014 (Figure 1A). Twenty representative fruits were sampled from each tree at each developmental stage and sliced. The sliced samples were combined and immediately frozen in liquid nitrogen and kept at −80°C until use.

Figure 1. Images of ‘Furongli’ plum and anthocyanin content in the fruits. (A) ‘Furongli’ plums at different time-points selected for sequencing. Representative photographs of plums at 105, 115, 125, and 135 days after flowering (DAF). (B) Anthocyanin content in plums at different ripening stages. The vertical bars represent the standard error of triplicate experiments.
Anthocyanin content was quantified as described by Niu et al. (2010). Briefly, approximately 3 g of sample was ground to a fine powder in liquid nitrogen and extracted with 20 mL extraction solution (0.05% HCl in methanol) at 4°C for 24 h. After centrifugation at 8000 × g for 20 min, the supernatant was transferred into a clean tube. One milliliter of supernatant and 4 mL of either bufferA (0.4MKCl, adjusted to pH 1.0 with HCl) or buffer B (1.2 N citric acid, adjusted to pH 4.5 with Na2HPO4) were mixed and the absorbance at 510 and 700 nm (A510 and A700) measured for A and B buffers, respectively. The anthocyanin content was calculated according to Romero et al. (2008) using the following formula: TA = A × MW × 5 × 100 × V/e, where TA stands for total anthocyanin content (mg/100 g, as cyanidin-3-O-glucose equivalent), V for final volume (mL), and A = [A510 (pH 1.0) − A700 (pH 1.0)] − [A510 (pH 4.5) − A700 (pH 4.5)]. A molar absorptivity (e) of 26,900 and molecular weight (MW) of 449.2 were used according to Wrolstad et al. (1982). Three measurements were taken for each biological replicate.
Total RNA was extracted from each sample using a EZNA Plant RNA Kit (Omega Bio-tek). The concentration of RNA was quantified with a Qubit 2.0 Fluorometer (Invitrogen, Life Technologies, CA, USA), and RNA integrity was evaluated using an Agilent 2100 Bioanalyzer (Agilent Technologies, Santa Clara, CA, USA). Equal amounts of RNA from three samples at the same stage were mixed together. The RNA isolation, library construction, and RNA-seq were performed by staff at Beijing BioMarker Technologies (Beijing, China). cDNA libraries were constructed as described by Han et al. (2013). Poly-A mRNA was enriched using poly-T oligoattached magnetic beads, then broken into small pieces, and used as template for cDNA synthesis. First strand cDNA was synthesized using reverse transcriptase and random primers, followed by second strand cDNA synthesis using DNA Polymerase I and RNase H. The cDNA libraries were sequenced on an Illumina HiSeq™ 2500.
Before assembly, the raw reads in FASTQ format were filtered using in-house Perl scripts to discarding the reads containing the sequencing adapters and low-quality reads with ambiguous “N” bases or in which more than 10% of bases had a Q ≤ 20. The left files (read1 files) from all libraries/samples were pooled into one large left.fq file, and right files (read2 files) into one large right.fq file. De novo assembly of the clean reads was performed using Trinity (Grabherr et al., 2011) with min_kmer_cov set to 2 by default and all other parameters set to their default values.
To annotate unigene sequences of ‘Furongli’ plums, blastx search (E < 10−5) was used to search against the NCBI nonredundant protein (nr), UniProt/Swiss-Prot, Gene Ontology (GO), Cluster of Orthologous Groups of proteins (COG), and Kyoto Encyclopedia of Genes and Genomes (KEGG) and Swiss-Prot databases and retrieve protein functional annotations based on sequence similarity. GO terms were assigned to the unigenes using Blast2GO (Conesa et al., 2005) with E ≤ 10−5. The distribution of the GO functional classifications of the unigenes was plotted using WEGO software (Ye et al., 2006).
Reads from the four cDNA libraries were mapped to the assembled unigenes using Bowtie (Langmead et al., 2009). Unigene expression levels were quantified using fragments per kilobase of transcript per million mapped reads (FPKM). FPKM values were calculated using RSEM (Li and Dewey, 2011). For each sequenced library, the read counts were adjusted using edgeR (Robinson et al., 2010) with one normalization factor. Differential expression analysis of two samples was performed using the DEGseq R package (Wang et al., 2010). Unigenes differentially expressed between two samples were screened using false discovery rate (FDR) < 0.01 and absolute log2 fold changes ≥1 as the threshold.
To identify transcription factors expressed during ripening of ‘Furongli’ plum fruit, predicted peptide sequences of all unigenes were searched against transcription factors in PlantTFDB 3.0 using the Transcription Factor Prediction module (http://planttfdb.cbi.pku.edu.cn/prediction.php) with default parameters.
Correlation analysis of anthocyanin structural genes and transcription factors was performed as described by Ye et al. (2015). To exclude false positives, we only selected unigenes with a FPKM value ≥10 in at least one of the four stages during fruit ripening. Transcription factors with correlation coefficient values of ≥0.95 by t-test were considered to have an expression that was significantly correlated with the expression of genes in anthocyanin biosynthetic pathways. The formula used was t = (r√(n–2))/√(1–r2), at P < 0.05 and n = 4. A value of |t|>t0.05, 2 = 4.303, implying r > 0.95, means significant correlation. Coexpression analysis was carried out using the “CORREL” function in Microsoft Excel 2003 and confirmed using in-house Perl scripts and IBM SPSS statistics software.
Nineteen candidate differentially expressed genes involved in anthocyanin biosynthesis were selected for validation by real-time quantitative RT-PCR (qRT-PCR). Total RNA from fruit samples was extracted using a modified CTAB method as described by Xuan et al. (2015). The primer sequences used for qRT-PCR are listed in Table S1. The cDNA was transcribed from 500 ng of total RNA using the PrimeScript RT reagent kit with gDNA Eraser (Takara, Dalian, China). Quantitative RT-PCR was performed using the Eppendorf RealPlex4 real-time PCR system (Hamburg, Germany) in a total volume of 20 μL in each well containing 10 μL of 2 × SYBR Premix Ex Taq™ II (Tli RNaseH Plus, TaKaRa), 1 μL of cDNA (in 1:10 dilution), and 0.4 μL 10 μM primers. Quantitative PCR conditions were 5 min at 95°C, followed by 40 cycles of 5 s at 95°C, 15 s at annealing temperatures listed in Table S1, and 30 s at 72°C, followed by 60–95°C melting curve detection. The actin gene was used as the reference. The expression levels were calculated as described by Fang et al. (2013). Three biological and three technical replicates were performed.
As indicated in Figure 1A, the color of ‘Furongli’ plums changed from green to red during ripening and the flesh became pigmented before the skin. Anthocyanin accumulation is responsible for the red color of plums and the major anthocyanins in ‘Furongli’ plums are cyanidin 3-rutinoside and cyanidin 3-glucoside (Usenik et al., 2009; Zhang, 2009). The anthocyanin content of ‘Furongli’ plums increased from 0.13 mg/100 g FW to 7.0 mg/100 g FW as ripening proceeded (Figure 1B).
Four cDNA libraries were constructed from the total RNA of ‘Furongli’ plums at 105, 115, 125, and 135 DAF. These libraries were subjected to RNA-seq using an Illumina HiSeq2500, generating 41,555,484, 43,021,234, 40,991,934, and 37,023,409 raw 100-bp paired-end raw reads, respectively (Table 1). All of the raw reads are available in the NCBI SRA database (accession number SRP076001). After removing low-quality reads and trimming adapter sequences, 41,067,827, 42,872,666, 40,886,898 and 36,927,707 clean reads were obtained for the 105, 115, 125, and 135 DAF libraries, respectively.
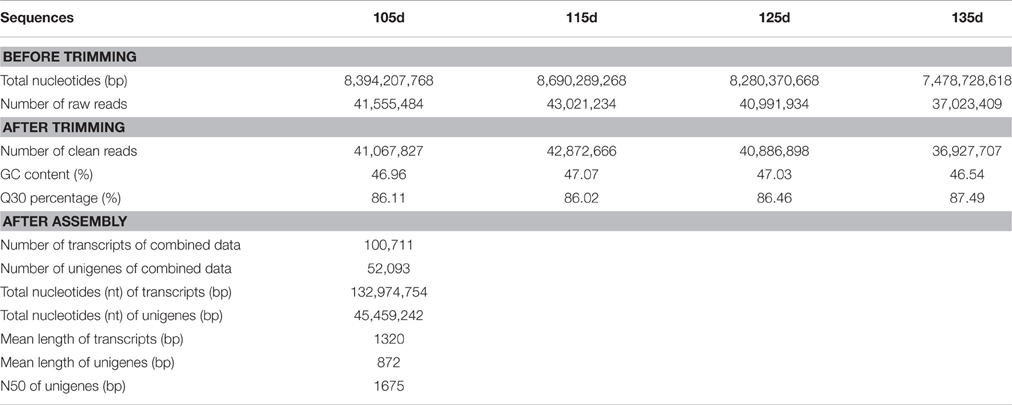
Table 1. Summary of sequencing and de novo assembly.
Using Trinity, the clean reads from the four libraries were assembled into 100,711 transcripts with an average length of 1320 bp, and 52,093 unigenes with an average length of 872 bp were obtained (Table 1). Unigenes shorter than 500 bp accounted for 55.43% of the total unigenes and 26.77% of the unigenes (13,944) were longer than 1 kb (Figure 2). These results suggest that the quality of the unigene data was high enough for the following analyses.
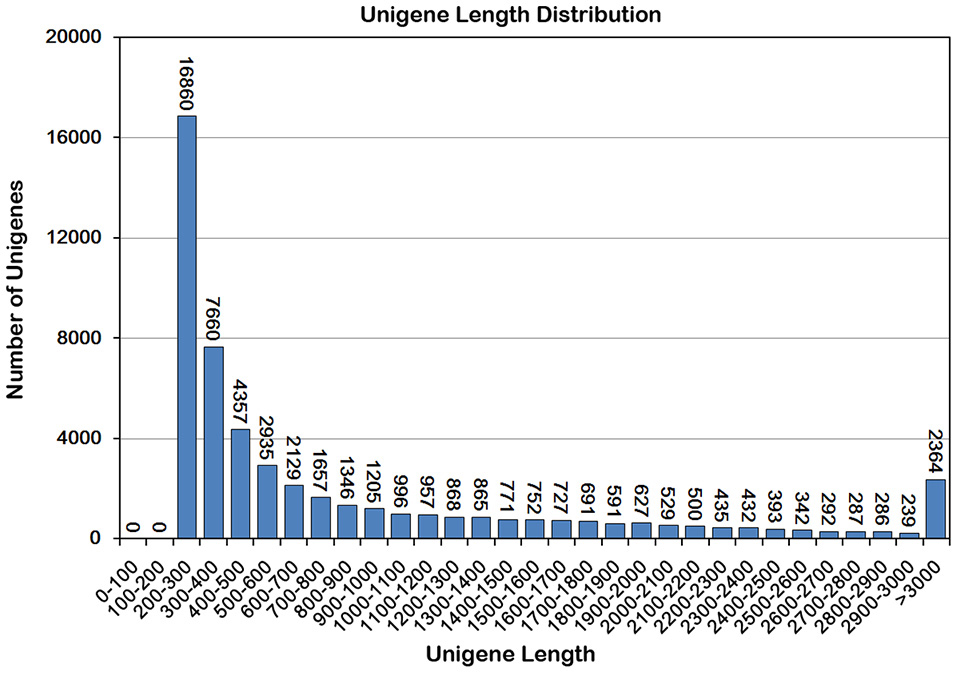
Figure 2. Sequence length distribution of the unigenes in ‘Furongli’ plum fruit transcriptomes. The x-axis indicates unigene length interval from 200 bp to > 3000 bp. The y-axis indicates the number of unigenes of each given sequence length.
To annotate the transcriptome of ‘Furongli’ plums, 52,093 unigenes were searched against five public databases (nr, UniProt/Swiss-Prot, GO, COG, and KEGG) with a cutoff E-value of 10−5. The functional annotation results are listed in Table 2. Only 49.4% of the unigenes (25,730) were identified. The remaining unigenes (50.6%) could not be annotated with known genes (Table 2), most likely because oflimitations in the genomic information and the presence of short sequences. Only 947 (3.6%) of the unannotated unigenes were longer than 1000 bp, while 47.1% were shorter than 300 bp. The species distribution of the best match results in nr is indicated in Figure 3. The ‘Furongli’ plum unigenes showed the closest matches with Prunus persica (77.80%), followed by Fragaria vesca (5.32), Vitis vinifera (1.51), Zea mays (1.06), Theobroma cacao (0.71), Populus trichocarpa (0.56), Hordeum vulgare (0.48), Ricinus communis (0.46), Medicago truncatula (0.38), and Glycine max (0.37%).
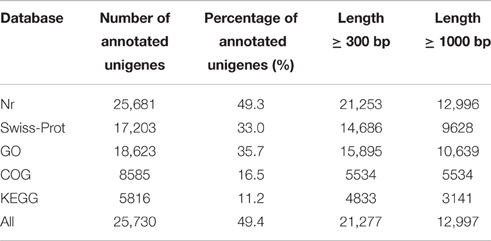
Table 2. Summary of functional annotations for the assembled ‘Furongli’ plum unigenes.
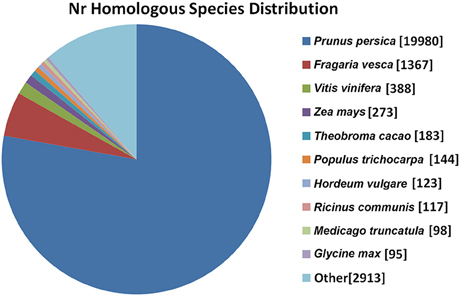
Figure 3. Species distribution of the BLAST search results in nr database. This figure shows the species distribution of unigene BLAST results against the nr database with a cutoff E-value of 10−5. Different species are indicated by different colors.
The ‘Furongli’ plum unigenes were searched against the GO database to classify standardized gene functions. At least one GO term was assigned to 18,623 of the unigenes (Table 2). Unigenes were assigned to three main GO categories, including biological process category, cellular component category, and molecular function category, and 58 subcategories shown in Figure 4. The terms “cell,” “cell part,” and “organelle” were dominant in the cellular component category, the term “binding” and “catalytic activity” was dominant in the molecular function category, and the terms “metabolic process” and “cellular process” were dominant in the biological process category.
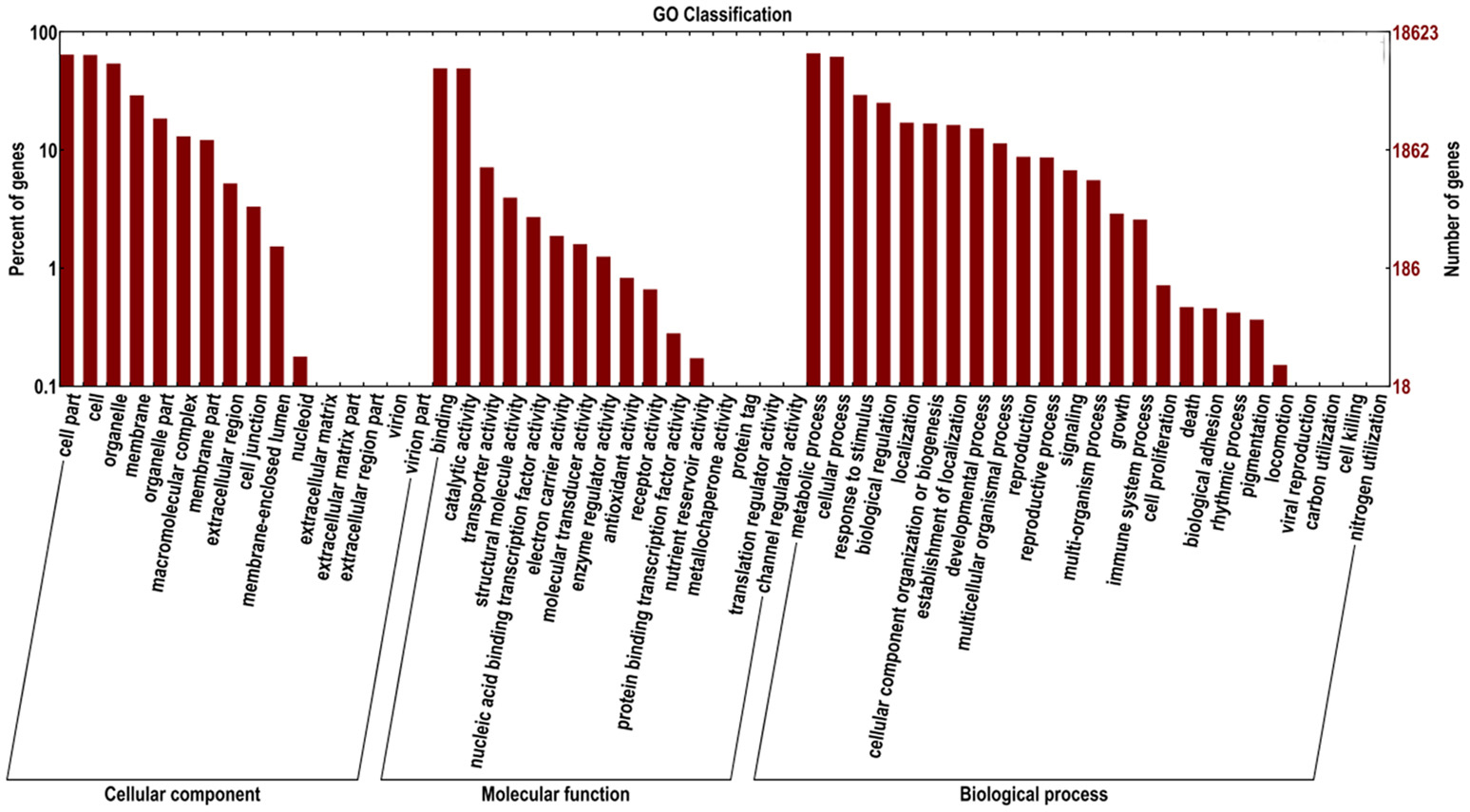
Figure 4. GO classifications of ‘Furongli’ plum unigenes.
The completeness of the plum transcriptome library and the validity of the annotations were further evaluated by COG annotation. Out of the 25,730 annotated unigenes, 8585 (33.37%) were clustered into 24 COG categories (Figure 5). The cluster for “general functional prediction only” (2260, 26.32%) represented the largest group, followed by “translation, ribosomal structure and biogenesis” (1095, 12.75%), “replication, recombination, and repair” (1074, 12.51%), “transcription” (1038, 12.09%), “Signal transduction mechanisms” (887, 10.33%), “posttranslational modification, protein turnover, chaperones” (840, 9.78%), “carbohydrate transport and metabolism” (652, 7.59%), “amino acid transport and metabolism” (553, 6.44%), and “energy production and metabolism” (516, 6.01%). “cell motility” (9, 0.1%) and “nuclear structure” (1, 0.01%) represented the smallest groups. No unigene was assigned to “extracellular structures.”
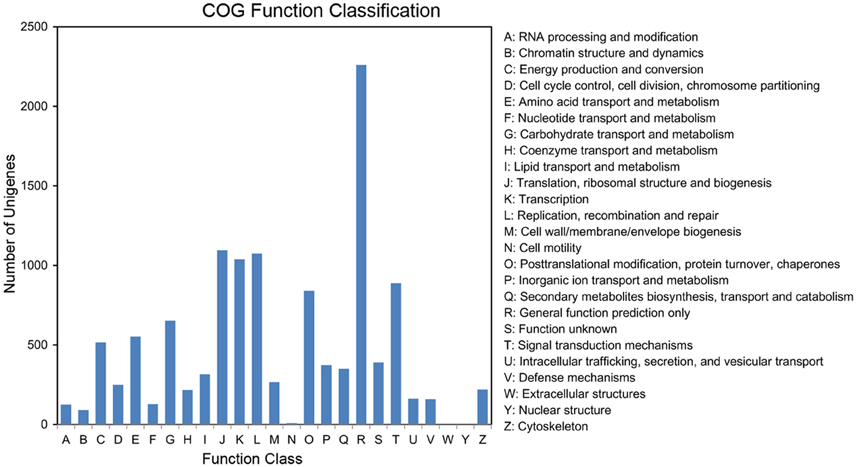
Figure 5. Histogram of the COG classifications. 8585 unigenes were grouped into 24 COG categories.
To better understand biological pathways involved in ripening of ‘Furongli’ plum fruits, all unigenes were searched against the reference canonical pathways in the KEGG database. Only 5816 (11.2%) unigenes had significant matches in the KEGG database and these were assigned to 115 KEGG pathways (Table S2). Among them, 2009 unigenes were assigned to metabolic pathways. As demonstrated in Figure 6A, 552 unigenes were assigned to carbohydrate metabolism, followed by Energy metabolism (548 unigenes), amino acid metabolism (426 unigenes), and lipid metabolism (195 unigenes). We concentrated on the “biosynthesis of other secondary metabolites” category in relation to fruit pigmentation. In this category, 119 unigenes were classified into seven subcategories (Figure 6B). Among these, the cluster for “phenylpropanoid biosynthesis” was the most represented, followed by “flavonoid biosynthesis.” “Flavone and flavonol biosynthesis” and “caffeine metabolism” appeared to be the smallest groups.
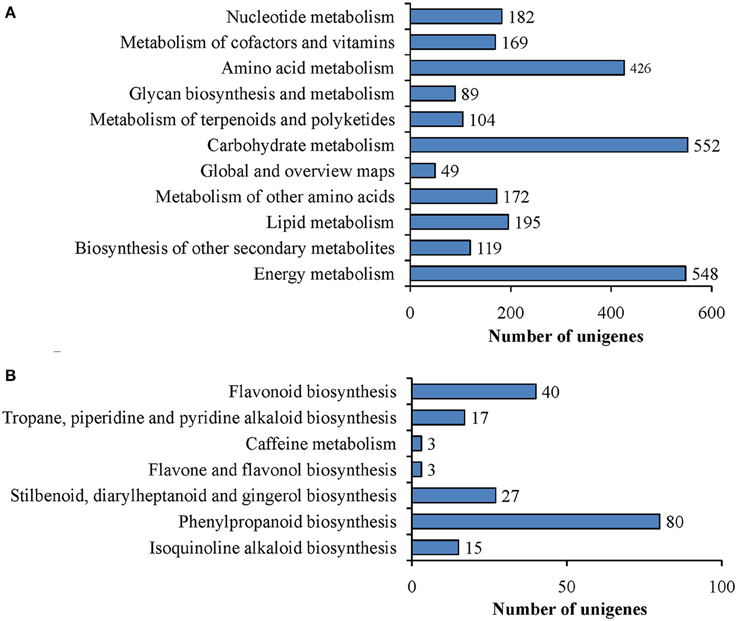
Figure 6. Pathway assignment based on KEGG. (A) Classification based on metabolism categories; (B) Classification based on “biosynthesis of other secondary metabolites” categories.
To study unigene expression during different developmental stages, the reads from the four libraries were mapped to the assembled transcriptome. As indicated in Figure 7, most of the unigenes (32,926, 63.21%) were expressed in all four stages of development. A total of 3891 unigenes (7.47%) were only detected at a single stage of development. Of these stage-specific expressed unigenes, 682 (1.31), 862 (1.65), 1280 (2.46), and 1.067 (2.05%) unigenes were expressed exclusively at 105, 115, 125, and 135 DAF, respectively.
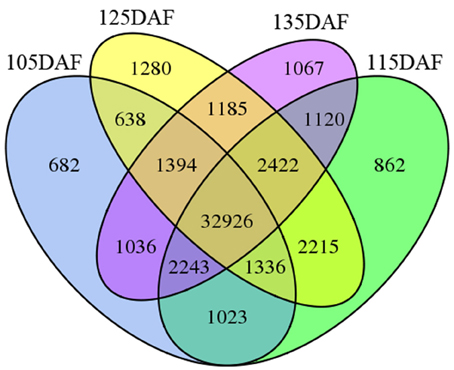
Figure 7. Venn diagram illustrating the number of unigenes expressed at different developmental stages.
By comparing the libraries, 507 differentially expressed unigenes (304 upregulated and 203 downregulated) were identified between 105 and 115 DAF, with 1674 differentially expressed unigenes (778 upregulated and 886 downregulated) between 115 and 125 DAF, as well as 1004 differentially expressed unigenes (523 up-regulated and 481 downregulated) between 125 and 135 DAF (Figure 8). Between 105 and 125 DAF, there are 2609 differentially expressed unigenes (1289 up-regulated and 1320 downregulated) showed differential expression. Between 105 and 135 DAF, there were 1743 differentially expressed unigenes (876 upregulated and 867 downregulated) show different expression. Between 115 and 135 DAF, there are 940 differentially expressed unigenes (467 upregulated and 473 downregulated). When considered together, 3548 unigenes were differentially expressed during fruit ripening. This result suggests that the developmental period with the most dynamic changes in the transcriptome was between 115 and 125 DAF, with almost half of the differentially expressed genes (47.18%) showing significant changes during this period.
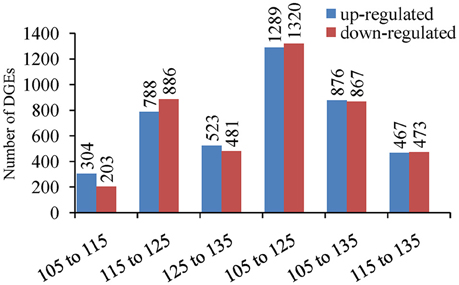
Figure 8. Number of differentially expressed genes (DEGs) between 105, 115, 125, and 135 DAF in ‘Furongli’ plums.
To identify unigenes involved in anthocyanin biosynthesis, KEGG functional enrichment was analyzed to characterize the functions of differentially expressed unigenes (Table S3). A total of 11 unigenes encoding enzymes, including PAL (c38398.graph_c0), C4H (c23939.graph_c0), 4CL (c30378.graph_c0), CHS (c37054.graph_c0), CHI (c28749.graph_c0), F3H (c23888.graph_c0), F3′H (c38186.graph_c0), DFR (c24831.graph_c0), ANS/LDOX (c29583.graph_c0), and UFGT (c19095.graph_c0), were assigned to the anthocyanin biosynthetic pathway based on KEGG (Table 3; Table S3). GO annotation was carried out to further identify anthocyanin-associated unigenes without annotation information in KEGG database (Table S4). The unigene c8193.graph_c0 (CHI) was assigned to “anthocyanin-containing compound biosynthetic process.” The unigene c25083.graph_c0 (flavonoid 3′-monooxygenase) was predicted to have flavonoid 3′, 5′-hydroxylase activity (GO:0033772). Furthermore, a GST (c29416.graph_c0), with best match to AtGSTF12 (AT5G17220), also showed differential expression. As shown in Table 3, the expression of these 13 unigenes was significantly upregulated during fruit maturation.
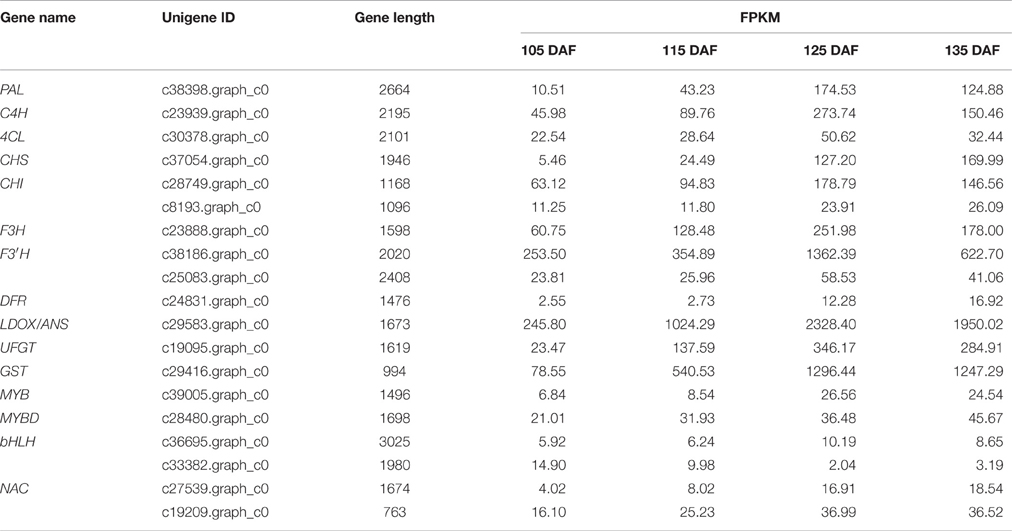
Table 3. Expression profiles of anthocyanin biosynthesis genes in ‘Furongli’ plums.
Transcription factors play important roles in the regulation of anthocyanin biosynthesis. In total, 791 unigenes (Table S5) were predicted to encode transcription factors from 55 different families (Table S6) and 147 of them were differentially expressed (Table S7). To identify transcription factors that were coexpressed with the candidate enzymatic genes involved in anthocyanin biosynthesis, a transcription abundance correlation analysis was carried out between the differentially expressed transcription factors and structural genes from the anthocyanin biosynthetic pathway. This identified 37 transcription factors whose expression levels were highly correlated with those of the candidate structural genes (Table 4; Table S8). Of these, 22 showed a significant correlation with five or more structural genes from the anthocyanin biosynthetic pathway. The identified transcription factors included homologs of Arabidopsis transcription factors that are implicated in regulating anthocyanin biosynthesis, such as MYB, bHLH, and NAC (Table 3).
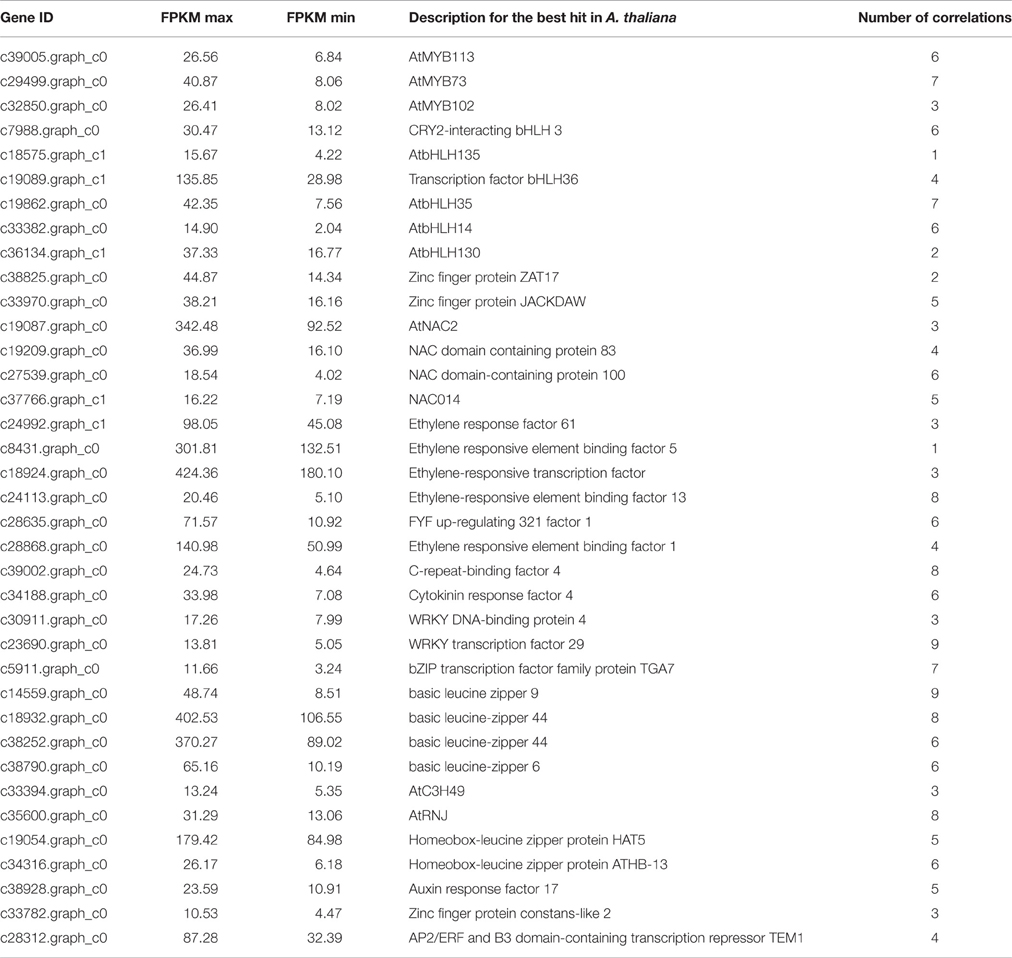
Table 4. Correlation analysis of structural genes involved in anthocyanin metabolism and transcription factors.
A total of 37 MYBs were differentially expressed during ripening of ‘Furongli’ plums and three of them (c39005.graph_c0, c29499.graph_c0 and c32850.graph_c0) were associated with the anthocyanin biosynthetic pathway (Table 4; Table S8). The unigene c39005.graph_c0 (homologous to AtMYB113) was upregulated, while c29499.graph_c0 (homologous to AtMYB73) and c32850.graph_c0 (homologous to AtMYB102) were downregulated. The expression level of the homolog of AtMYBD (c28480.graph_c0) also increased. However, it was correlated with none of the structural genes. Seven of the differentially expressed transcription factors annotated as bHLH showed a significant correlation with anthocyanin biosynthetic genes (Table 4; Table S8). Only two of the bHLH genes (c7988.graph_c0 and c18575.graph_c1) showed a positive correlation with the expression of structural genes, while most of them were negatively correlated with that of structural genes involved in the anthocyanin biosynthetic pathway. In addition, a plum bHLH (c36695.graph_c0, log2 fold change < 1.0), which is the best BLAST match to Arabidopsis AtbHLH42, was significantly correlated with seven anthocyanin biosynthetic structural genes (Table S8). WD40 encoding unigenes (c28377.graph_c0 and c10590.graph_c1), which are the homolog of AtTTG1, did not show differential expression during the ripening process. Apart from the MBW components, other differentially expressed transcription factors, such as NAC, were also found to be potentially related to the anthocyanin pathway. Four plum NAC genes (c19087.graph_c0, c19209.graph_c0, c27539.graph_c0, and c37766.graph_c1) were significantly correlated with structural genes (Table 4; Table S8). The unigenes c27539.graph_c0 (a homolog of AtNAC100) and c19209.graph_c0 were upregulated during plum fruit ripening. Peach homolog of AtNAC100 have been reported to be involved in the regulation of anthocyanin accumulation in fruit flesh. GO annotation results indicated that c19209.graph_c0 is involved in “biological process: positive regulation of flavonoid biosynthetic process” (GO:0009963).
We further analyzed the expression profiles of 19 candidate unigenes (13 structural genes and six transcription factors) involved in anthocyanin biosynthesis using qRT-PCR. The results indicated that there is a good correlation between RNA-seq data and qPCR data for most of the genes (Figure 9).
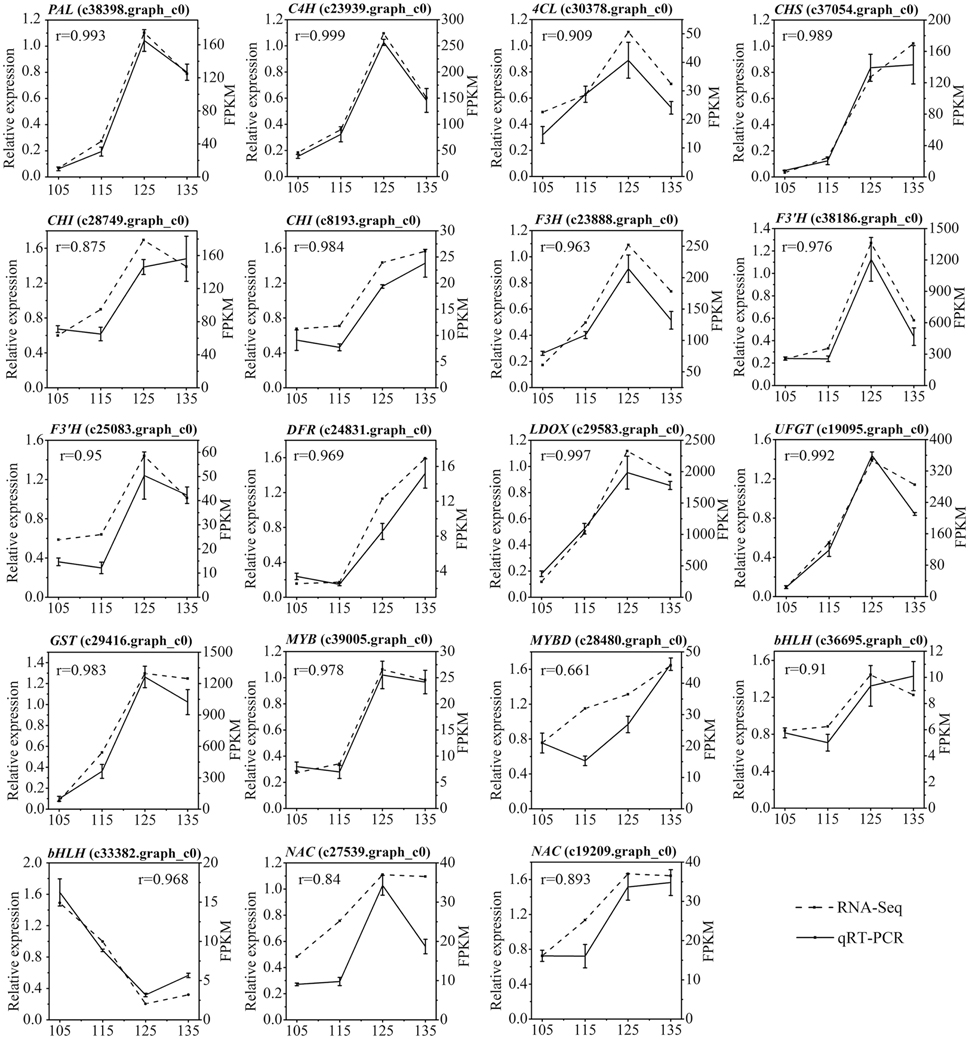
Figure 9. Expression analysis of 19 differentially expressed genes related to anthocyanin biosynthesis in ‘Furongli’ plums during fruit ripening. Actin was used as the internal control. The error bars represent the standard error of three biological replicates. The numbers above the graphics correspond to values obtained with the Pearson correlation. Pearson correlation between the RNA-seq data and qRT-PCR data was calculated using the log2 value of FPKM and the relative expression level.
In this study, we constructed a transcriptome of ‘Furongli’ plums during fruit maturation. In total, 52,093 unigenes were assembled with a mean length of 872 bp, which is comparable to 944 bp for sweet cherry (P. avium L.) (Wei et al., 2015). A large quantity of genomic data is available for many rosaceous plants, but only 49.4% of the plum unigenes were annotated to public databases (nr, Swiss-Prot, GO, COG, and KEGG). This means that more than half of the unigenes have no significant known homologs. The low rate of annotated unigenes could be a result of limitations in the genomic information available for P. salicina Lindl. as is the case in other non-model plant species (Yates et al., 2014; Wei et al., 2015; Wu et al., 2015). As expected, 77.80% of the unigenes annotated using nr show significant similarity to P. persica transcripts. The unannotated unigenes could be plum specific genes with novel functions. The transcriptome of ‘Furongli’ plums will serve as an important datasets for studying plum ripening processes such as sugar accumulation, organic acid degradation, fruit softening, and pigmentation.
Anthocyanin-rich plums are of great interest for their implications in human health (Santhakumar et al., 2015). The main objective of this study was to identify genes involved in anthocyanin biosynthesis in plums. RNA-seq-based comparative transcriptome analysis has been shown to be an efficient strategy for the investigation of genes involved in anthocyanin biosynthesis in several plants, such as kiwifruit (Li et al., 2015a), sweet cherry (Wei et al., 2015), zoysiagrass (Ahn et al., 2015), anthurium (Li et al., 2015b), and potato (Liu et al., 2015). In the later ripening stages, ‘Furongli’ plums accumulate anthocyanins rapidly (Figure 1). The expression of anthocyanin biosynthetic genes has been shown to be correlated with fruit anthocyanin content in Rosaceae such as apple (Feng et al., 2013, 2014; Vimolmangkang et al., 2014), pear (Li et al., 2012a; Yang et al., 2015), sweet cherry (Wei et al., 2015), strawberry (Xu et al., 2014), and plum (Cheng et al., 2015). In the present study, changes in gene expression between different stages of ripening were analyzed to identify differentially expressed genes implicated in anthocyanin biosynthesis, including PAL, C4H, 4CL, CHS, CHI, F3H, F3′H, DFR, ANS/LDOX, UFGT, and GST, were significantly upregulated in the late stages of fruit maturation (Table 3; Figure 9).
Anthocyanin biosynthesis is regulated by several well-studied transcription factors such as MYB, bHLH, and WD40 (Gonzalez et al., 2008). MYB transcription factors have been reported to play a pivotal role in anthocyanin biosynthesis regulation in several fruit trees (Chagné et al., 2013; Ravaglia et al., 2013; Umemura et al., 2013; Lai et al., 2014; Shen et al., 2014; Tuan et al., 2015; Zhai et al., 2015; Jin et al., 2016). Lin-Wang et al. (2010) demonstrated that R2R3 MYBs are highly conserved in rosaceous plants and MYBs from European plum and cherry plum are able to induce the anthocyanin accumulation in tobacco. Gu et al. (2015) proposed that constitutive activation of PcMYB10.6 is responsible for red pigmentation in purple-leaf plum. Cheng et al. (2015) indicated that PsMYB10 was involved in ethylene-regulated anthocyanin biosynthesis in plums. These studies suggested that MYBs play a role in anthocyanin biosynthesis in the plum. In this study, a plum MYB (c39005.graph_c0) was positively correlated with structural genes. Its Arabidopsis homolog (AtMYB113) is an activator of the anthocyanin pathway. Recently, AtMYBD was shown to enhance anthocyanin biosynthesis by repressing the negative regulator MYBL2 (Nguyen et al., 2015). The expression of an AtMYBD homolog (c28480.graph_c0) was upregulated in late ripening stages, but it does not show significant correlation with any structural genes. In addition, the anthocyanin biosynthetic pathway is also negatively controlled by MYB repressors in many plants, including Arabidopsis (Dubos et al., 2008; Matsui et al., 2008; Zhu et al., 2009), poplar (Yoshida et al., 2015), Medicago truncatula (Jun et al., 2015), grapevine (Cavallini et al., 2015; Pérez-Díaz et al., 2015), strawberry (Salvatierra et al., 2013), and apple (Lin-Wang et al., 2011). We found that two MYBs (c29499.graph_c0 and c32850.graph_c0) were negatively correlated with anthocyanin biosynthetic genes. Another important component of the MBW complex, bHLH proteins have been shown to be essential for or to enhance MYB-induced anthocyanin accumulation in transient expression assays (Liu et al., 2013b; Rahim et al., 2014; Feng et al., 2015; Starkevič et al., 2015; Wei et al., 2015; Lai et al., 2016). Our results indicated that c36695.graph_c0, which is highly homologous to AtTT8 (AT4G09820), accumulates to higher levels in late stages of ripening and shows significant correlation with anthocyanin biosynthetic genes. Conversely, the expression of c33382.graph_c0 was repressed as ripening proceeded. The unigene c33382.graph_c0 is a homolog of AtbHLH14 (AT4G00870), which belongs to bHLH subgroup IIId. Song et al. (2013) demonstrated that Arabidopsis lines overexpressing bHLH17 showed jasmonate-induced anthocyanin accumulation. NAC proteins have also been reported to be involved in anthocyanin synthesis in Arabidopsis (Morishita et al., 2009). Recently, a peach homolog of AtNAC100 was shown to be responsible for regulation of anthocyanin accumulation in flesh (Zhou et al., 2015). Our results indicated that a plum homolog of AtNAC100 (c27539.graph_c0) is upregulated and positively correlated with anthocyanin biosynthetic genes. It should be noted that coexpression analysis usually requires a large sample size and the small sample size in our study will reduce the reliability of our results. However, relatively small numbers of samples have recently been used to analyze the correlation of structural genes and regulators involved in biological processes, such as flavonoid biosynthesis (Zhai et al., 2015) and fruit ripening (Wu et al., 2016). The exact roles of these candidate transcription factor should be investigated in further studies.
In the current study, we used RNA-seq to analyze changes in the transcriptome during ripening of ‘Furongli’ plums. We generated 52,093 unigenes and over 50% of them were not annotated to public databases. Unigenes differentially expressed during fruit ripening were identified. Candidate genes encoding anthocyanin biosynthetic enzymes and transcription factors involved in anthocyanin biosynthesis were identified using functional annotation and coexpression analysis of differentially expressed genes. The expression patterns of some candidate genes encode anthocyanin biosynthetic enzymes and transcription factors were further validated by qRT-PCR. This provides an important datasets for studying fruit ripening processes, especially anthocyanin biosynthesis, in plums. Further studies are needed to determine whether the identified candidate genes are related to anthocyanin biosynthesis in plum.
This study was conceived by ZF and XY. The plant material preparation were carried out by ZF and SP. ZF, XY, CJ, and DZ analyzed the RNA-seq data. ZF, CJ, DZ, and SP performed the laboratory experiments and analyses. ZF drafted the manuscript. XY revised the manuscript. All authors read and approved the final manuscript.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
This work was funded by the Basic Scientific Research Funds of Public Welfare Scientific Research Institutes of Fujian Province (2014R1014-13), the Major Projects of Science and Technology Project of Fujian Province (2013NZ0002-1), the Youth Talent Plan of Fujian Academy of Agricultural Sciences (YC2015-9) and the Natural Science Foundation of Fujian Province (2016J01123 and 2015J05058).
The Supplementary Material for this article can be found online at: http://journal.frontiersin.org/article/10.3389/fpls.2016.01338
4CL, 4-coumaroyl:CoA-ligase; ANS/LDOX, anthocyanidin synthase/leucoanthocyanidin dioxygenase; bHLH, basic Helix-Loop-Helix; C4H, cinnamate-4-hydroxylase; CHI, chalcone isomerase; CHS, chalcone synthase; COG, Cluster of Orthologous Groups of proteins DFR, dihydroflavonol 4-reductase; DAF, days after flowering; F3H, flavanone 3-hydroxylase; F3′H, flavonoid 3′-hydroxylase; FPKM, Fragments Per Kilobase of transcript per Million mapped reads; GO, gene ontology; GST, glutathione S-transferase; KEGG, Kyoto Encyclopedia of Genes and Genomes; nr, NCBI nonredundant protein database; PAL, phenylalanine ammonialyase; qRT-PCR, real-time quantitative RT-PCR; UFGT, UDP-glucose: flavonoid 3-O-glucosyltransferase.
Ahn, J. H., Kim, J. S., Kim, S., Soh, H. Y., Shin, H., Jang, H., et al. (2015). De novo transcriptome analysis to identify anthocyanin biosynthesis genes responsible for tissue-specific pigmentation in Zoysiagrass (Zoysia japonica Steud.). PLoS ONE 10:e0124497. doi: 10.1371/journal.pone.0124497
An, X., Tian, Y., Chen, K., Liu, X., Liu, D., Xie, X., et al. (2015). MdMYB9 and MdMYB11 are involved in the regulation of the JA-induced biosynthesis of anthocyanin and proanthocyanidin in apples. Plant Cell Physiol. 56, 650–662. doi: 10.1093/pcp/pcu205
Butelli, E., Licciardello, C., Zhang, Y., Liu, J., Mackay, S., Bailey, P., et al. (2012). Retrotransposons control fruit-specific, cold-dependent accumulation of anthocyanins in blood oranges. Plant Cell 24, 1242–1255. doi: 10.1105/tpc.111. 095232
Carrasco, B., Díaz, C., Moya, M., Gebauer, M., and García-González, R. (2012). Genetic characterization of Japanese plum cultivars (Prunus salicina) using SSR and ISSR molecular markers. Cienc. Invest. Agra. 39, 533–543. doi: 10.4067/S0718-16202012000300012
Cavallini, E., Matus, J. T., Finezzo, L., Zenoni, S., Loyola, R., Guzzo, F., et al. (2015). The phenylpropanoid pathway is controlled at different branches by a set of R2R3-MYB C2 repressors in grapevine. Plant Physiol. 167, 1448–1470. doi: 10.1104/pp.114.256172
Chagné, D., Lin-Wang, K., Espley, R. V., Volz, R. K., How, N. M., Rouse, S., et al. (2013). An ancient duplication of apple MYB transcription factors is responsible for novel red fruit-flesh phenotypes. Plant Physiol. 161, 225–239. doi: 10.1104/pp.112.206771
Cheng, Y., Liu, L., Yuan, C., and Guan, J. (2015). Molecular characterization of ethylene-regulated anthocyanin biosynthesis in plums during fruit ripening. Plant Mol. Biol. Rep. 34, 777–785. doi: 10.1007/s11105-015-0963-x
Conesa, A., Götz, S., García-Gómez, J. M., Terol, J., Talón, M., and Robles, M. (2005). Blast2GO: a universal tool for annotation, visualization and analysis in functional genomics research. Bioinformatics 21, 3674–3676. doi: 10.1093/bioinformatics/bti610
Crifò, T., Petrone, G., Lo Cicero, L., and Lo Piero, A. R. (2011). Short cold storage enhances the anthocyanin contents and level of transcripts related to their biosynthesis in blood oranges. J. Agr. Food. Chem. 60, 476–481. doi: 10.1021/jf203891e
Dubos, C., Le Gourrierec, J., Baudry, A., Huep, G., Lanet, E., Debeaujon, I., et al. (2008). MYBL2 is a new regulator of flavonoid biosynthesis in Arabidopsis thaliana. Plant J. 55, 940–953. doi: 10.1111/j.1365-313X.2008.03564.x
Espley, R. V., Hellens, R. P., Putterill, J., Stevenson, D. E., Kutty-Amma, S., and Allan, A. C. (2007). Red colouration in apple fruit is due to the activity of the MYB transcription factor, MdMYB10. Plant J. 49, 414–427. doi: 10.1111/j.1365-313X.2006.02964.x
Fang, Z., Zhou, D., Liao, R., Jiang, C., Pan, S., and Ye, X. (2014). ISSR analysis of Furong plums from Ninggang and Fujian. South China Fruits 43, 12–14.
Fang, Z. Z., Lai, C. C., Zhang, Y. L., Lin, Y. L., and Lai, Z. X. (2013). Identification of a PTC-containing DlRan transcript and its differential expression during somatic embryogenesis in Dimocarpus longan. Gene 529, 37–44. doi: 10.1016/j.gene.2013.07.091
Feller, A., Machemer, K., Braun, E. L., and Grotewold, E. (2011). Evolutionary and comparative analysis of MYB and bHLH plant transcription factors. Plant J. 66, 94–116. doi: 10.1111/j.1365-313X.2010.04459.x
Feng, F., Li, M., Ma, F., and Cheng, L. (2013). Phenylpropanoid metabolites and expression of key genes involved in anthocyanin biosynthesis in the shaded peel of apple fruit in response to sun exposure. Plant Physiol. Biochem. 69, 54–61. doi: 10.1016/j.plaphy.2013.04.020
Feng, F., Li, M., Ma, F., and Cheng, L. (2014). The effects of bagging and debagging on external fruit quality, metabolites, and the expression of anthocyanin biosynthetic genes in ‘Jonagold’ apple (Malus domestica Borkh.). Sci. Hortic. 165, 123–131. doi: 10.1016/j.scienta.2013.11.008
Feng, S., Sun, S., Chen, X., Wu, S., Wang, D., and Chen, X. (2015). PyMYB10 and PyMYB10.1 interact with bHLH to enhance anthocyanin accumulation in pears. PLoS ONE 10:e0142112. doi: 10.1371/journal.pone.0142112
Fraser, L., Seal, A., Montefiori, M., McGhie, T., Tsang, G., Datson, P., et al. (2013). An R2R3 MYB transcription factor determines red petal colour in an Actinidia (kiwifruit) hybrid population. BMC Genomics 14:28. doi: 10.1186/1471-2164-14-28
Gonzalez, A., Zhao, M., Leavitt, J. M., and Lloyd, A. M. (2008). Regulation of the anthocyanin biosynthetic pathway by the TTG1/bHLH/Myb transcriptional complex in Arabidopsis seedlings. Plant J. 53, 814–827. doi: 10.1111/j.1365-313X.2007.03373.x
González, M., Maldonado, J., Salazar, E., Silva, H., and Carrasco, B. (2016a). De novo transcriptome assembly of ‘Angeleno’ and ‘Lamoon’ Japanese plum cultivars (Prunus salicina). Genomics Data 9, 35–36. doi: 10.1016/j.gdata.2016.06.010
González, M., Salazar, E., Castillo, J., Morales, P., Mura-Jornet, I., Maldonado, J., et al. (2016b). Genetic structure based on EST–SSR: a putative tool for fruit color selection in Japanese plum (Prunus salicina L.) breeding programs. Mol. Breed. 36, 1–15. doi: 10.1007/s11032-016-0491-x
Gou, J., Felippes, F. F., Liu, C., Weigel, D., and Wang, J. (2011). Negative regulation of anthocyanin biosynthesis in arabidopsis by a miR156-targeted SPL transcription factor. Plant Cell 23, 1512–1522. doi: 10.1105/tpc.111.084525
Grabherr, M. G., Haas, B. J., Yassour, M., Levin, J. Z., Thompson, D. A., Amit, I., et al. (2011). Full-length transcriptome assembly from RNA-Seq data without a reference genome. Nat. Biotechol. 29, 644–652. doi: 10.1038/nbt.1883
Gu, C., Liao, L., Zhou, H., Wang, L., Deng, X., and Han, Y. (2015). Constitutive activation of an anthocyanin regulatory gene PcMYB10.6 is related to red coloration in purple-foliage plum. PLoS ONE 10:e0135159. doi: 10.1371/journal.pone.0135159
Han, X.-J., Wang, Y.-D., Chen, Y.-C., Lin, L.-Y., and Wu, Q.-K. (2013). Transcriptome sequencing and expression analysis of terpenoid biosynthesis genes in Litsea cubeba. PLoS ONE 8:e76890. doi: 10.1371/journal.pone.0076890
Hooshmand, S., and Arjmandi, B. H. (2009). Viewpoint: Dried plum, an emerging functional food that may effectively improve bone health. Ageing Res. Rev. 8, 122–127. doi: 10.1016/j.arr.2009.01.002
Hu, B., Zhao, J., Lai, B., Qin, Y., Wang, H., and Hu, G. (2016). LcGST4 is an anthocyanin-related glutathione S-transferase gene in Litchi chinensis Sonn. Plant Cell Rep. 35, 831–841. doi: 10.1007/s00299-015-1924-4
Huang, Y. F., Vialet, S., Guiraud, J. L., Torregrosa, L., Bertrand, Y., Cheynier, V., et al. (2014). A negative MYB regulator of proanthocyanidin accumulation, identified through expression quantitative locus mapping in the grape berry. New Phytol. 201, 795–809. doi: 10.1111/nph.12557
Jaakola, L., Poole, M., Jones, M. O., Kämäräinen-Karppinen, T., Koskimäki, J. J., Hohtola, A., et al. (2010). A SQUAMOSA MADS box gene involved in the regulation of anthocyanin accumulation in bilberry fruits. Plant Physiol. 153, 1619–1629. doi: 10.1104/pp.110.158279
Jin, W., Wang, H., Li, M., Wang, J., Yang, Y., Zhang, X., et al. (2016). The R2R3 MYB transcription factor PavMYB10.1 involves in anthocyanin biosynthesis and determines fruit skin colour in sweet cherry (Prunus avium L.). Plant Biotechnol. J. doi: 10.1111/pbi.12568. [Epub ahead of print].
Jo, Y., Lian, S., Cho, J. K., Choi, H., Chu, H., and Cho, W. K. (2015). De novo transcriptome assembly of two different Prunus salicina cultivars. Genomics Data 6, 262–263. doi: 10.1016/j.gdata.2015.10.015
Johnson, C. D., Lucas, E. A., Hooshmand, S., Campbell, S., Akhter, M. P., and Arjmandi, B. H. (2011). Addition of fructooligosaccharides and dried plum to soy-based diets reverses bone loss in the ovariectomized rat. Evid. Based Complment. Alternat. Med. 2011:836267. doi: 10.1093/ecam/nen050
Jun, J. H., Liu, C., Xiao, X., and Dixon, R. A. (2015). The transcriptional repressor MYB2 regulates both spatial and temporal patterns of proanthocyandin and anthocyanin pigmentation in Medicago truncatula. Plant Cell 27, 2860–2879. doi: 10.1105/tpc.15.00476
Kobayashi, S., Goto-Yamamoto, N., and Hirochika, H. (2004). Retrotransposon-induced mutations in grape skin color. Science 304, 982. doi: 10.1126/science.1095011
Kobayashi, S., Ishimaru, M., Hiraoka, K., and Honda, C. (2002). Myb-related genes of the Kyoho grape (Vitis labruscana) regulate anthocyanin biosynthesis. Planta 215, 924–933. doi: 10.1007/s00425-002-0830-5
Lai, B., Du, L., Liu, R., Hu, B., Su, W., Qin, Y., et al. (2016). Two LcbHLH transcription factors interacting with LcMYB1 in regulating late structural genes of anthocyanin biosynthesis in Nicotiana and Litchi chinensis during anthocyanin accumulation. Front. Plant Sci. 7:166. doi: 10.3389/fpls.2016.00166
Lai, B., Li, X., Hu, B., Qin, Y., Huang, X., Wang, H., et al. (2014). LcMYB1 is a key determinant of differential anthocyanin accumulation among genotypes, tissues, developmental phases and ABA and light stimuli in Litchi chinensis. PLoS ONE 9:e86293. doi: 10.1371/journal.pone.0086293
Langmead, B., Trapnell, C., Pop, M., and Salzberg, S. L. (2009). Ultrafast and memory-efficient alignment of short DNA sequences to the human genome. Genome Biol. 10, 1–10. doi: 10.1186/gb-2009-10-3-r25
Lee, S. H., Lillehoj, H. S., Cho, S. M., Chun, H. K., Park, H. J., Lim, C. I., et al. (2009). Immunostimulatory effects of oriental plum (Prunus salicina Lindl.). Comp. Immunol. Microbiol. Infect. Dis. 32, 407–417. doi: 10.1016/j.cimid.2007.12.001
Lepiniec, L., Debeaujon, I., Routaboul, J.-M., Baudry, A., Pourcel, L., Nesi, N., et al. (2006). Genetics and biochemistry of seed flavonoids. Annu. Rev. Plant Biol. 57, 405–430. doi: 10.1146/annurev.arplant.57.032905.105252
Li, B., and Dewey, C. (2011). RSEM: accurate transcript quantification from RNA-Seq data with or without a reference genome. BMC Bioinformatics 12:323. doi: 10.1186/1471-2105-12-323
Li, L., Ban, Z.-J., Li, X.-H., Wu, M.-Y., Wang, A.-L., Jiang, Y.-Q., et al. (2012a). Differential expression of anthocyanin biosynthetic genes and transcription factor PcMYB10 in pears (Pyrus communis L.). PLoS ONE 7:e46070. doi: 10.1371/journal. pone.0046070
Li, P., Chen, B., Zhang, G., Chen, L., Dong, Q., Wen, J., et al. (2016). Regulation of anthocyanin and proanthocyanidin biosynthesis by Medicago truncatula bHLH transcription factor MtTT8. New Phytol. 210, 905–921. doi: 10.1111/nph.13816
Li, W., Liu, Y., Zeng, S., Xiao, G., Wang, G., Wang, Y., et al. (2015a). Gene expression profiling of development and anthocyanin accumulation in kiwifruit (Actinidia chinensis) based on transcriptome sequencing. PLoS ONE 10:e0136439. doi: 10.1371/journal.pone.0136439
Li, Y., Mao, K., Zhao, C., Zhao, X., Zhang, H., Shu, H., et al. (2012b). MdCOP1 ubiquitin E3 ligases interact with MdMYB1 to regulate light-induced anthocyanin biosynthesis and red fruit coloration in apple. Plant Physiol. 160, 1011–1022. doi: 10.1104/pp.112.199703
Li, Z., Wang, J., Zhang, X., and Xu, L. (2015b). Comparative transcriptome analysis of Anthurium “Albama” and its anthocyanin-loss mutant. PLoS ONE 10:0119027. doi: 10.1371/journal.pone.0119027
Lin-Wang, K., Bolitho, K., Grafton, K., Kortstee, A., Karunairetnam, S., McGhie, T., et al. (2010). An R2R3 MYB transcription factor associated with regulation of the anthocyanin biosynthetic pathway in Rosaceae. BMC Plant Biol. 10:50. doi: 10.1186/1471-2229-10-50
Lin-Wang, K., Micheletti, D., Palmer, J., Volz, R., Lozano, L., Espley, R., et al. (2011). High temperature reduces apple fruit colour via modulation of the anthocyanin regulatory complex. Plant Cell Environ. 34, 1176–1190. doi: 10.1111/j.1365-3040.2011.02316.x
Liu, X., Feng, C., Zhang, M., Yin, X., Xu, C., and Chen, K. (2013a). The MrWD40-1 gene of chinese bayberry (Myrica rubra) interacts with MYB and bHLH to enhance anthocyanin accumulation. Plant Mol. Biol. Rep. 31, 1474–1484. doi: 10.1007/s11105-013-0621-0
Liu, X., Yin, X., Allan, A., Lin-Wang, K., Shi, Y., Huang, Y., et al. (2013b). The role of MrbHLH1 and MrMYB1 in regulating anthocyanin biosynthetic genes in tobacco and Chinese bayberry (Myrica rubra) during anthocyanin biosynthesis. Plant Cell Tissue Organ Cult. 115, 285–298. doi: 10.1007/s11240-013-0361-8
Liu, Y., Lin-Wang, K., Deng, C., Warran, B., Wang, L., Yu, B., et al. (2015). Comparative transcriptome analysis of white and purple potato to identify genes involved in anthocyanin biosynthesis. PLoS ONE 10:e0129148. doi: 10.1371/journal.pone.0129148
Maier, A., and Hoecker, U. (2014). COP1/SPA ubiquitin ligase complexes repress anthocyanin accumulation under low light and high light conditions. Plant Signal. Behav. 10:e970440. doi: 10.4161/15592316.2014.970440
Maier, A., Schrader, A., Kokkelink, L., Falke, C., Welter, B., Iniesto, E., et al. (2013). Light and the E3 ubiquitin ligase COP1/SPA control the protein stability of the MYB transcription factors PAP1 and PAP2 involved in anthocyanin accumulation in Arabidopsis. Plant J. 74, 638–651. doi: 10.1111/tpj.12153
Matsui, K., Umemura, Y., and Ohme-Takagi, M. (2008). AtMYBL2, a protein with a single MYB domain, acts as a negative regulator of anthocyanin biosynthesis in Arabidopsis. Plant J. 55, 954–967. doi: 10.1111/j.1365-313X.2008.03565.x
Morishita, T., Kojima, Y., Maruta, T., Nishizawa-Yokoi, A., Yabuta, Y., and Shigeoka, S. (2009). Arabidopsis NAC transcription factor, ANAC078, regulates flavonoid biosynthesis under high-light. Plant Cell Physiol. 50, 2210–2222. doi: 10.1093/pcp/pcp159
Nemie-Feyissa, D., Heidari, B., Blaise, M., and Lillo, C. (2015). Analysis of interactions between heterologously produced bHLH and MYB proteins that regulate anthocyanin biosynthesis: quantitative interaction kinetics by Microscale Thermophoresis. Phytochemistry 111, 21–26. doi: 10.1016/j.phytochem.2015.01.004
Nguyen, N. H., Jeong, C. Y., Kang, G.-H., Yoo, S.-D., Hong, S.-W., and Lee, H. (2015). MYBD employed by HY5 increases anthocyanin accumulation via repression of MYBL2 in Arabidopsis. Plant J. 84, 1192–1205. doi: 10.1111/tpj.13077
Niu, S., Xu, C., Zhang, W., Zhang, B., Li, X., Lin-Wang, K., et al. (2010). Coordinated regulation of anthocyanin biosynthesis in Chinese bayberry (Myrica rubra) fruit by a R2R3 MYB transcription factor. Planta 231, 887–899. doi: 10.1007/s00425-009-1095-z
Palapol, Y., Ketsa, S., Lin-Wang, K., Ferguson, I., and Allan, A. (2009). A MYB transcription factor regulates anthocyanin biosynthesis in mangosteen (Garcinia mangostana L.) fruit during ripening. Planta 229, 1323–1334. doi: 10.1007/s00425-009-0917-3
Pérez-Díaz, J. R., Pérez-Díaz, J., Madrid-Espinoza, J., González-Villanueva, E., Moreno, Y., and Ruiz-Lara, S. (2015). New member of the R2R3-MYB transcription factors family in grapevine suppresses the anthocyanin accumulation in the flowers of transgenic tobacco. Plant Mol. Biol. 90, 63–76. doi: 10.1007/s11103-015-0394-y
Qi, T., Song, S., Ren, Q., Wu, D., Huang, H., Chen, Y., et al. (2011). The jasmonate-ZIM-domain proteins interact with the WD-repeat/bHLH/MYB complexes to regulate jasmonate-mediated anthocyanin accumulation and trichome initiation in Arabidopsis thaliana. Plant Cell 23, 1795–1814. doi: 10.1105/tpc.111.083261
Rahim, M. A., Busatto, N., and Trainotti, L. (2014). Regulation of anthocyanin biosynthesis in peach fruits. Planta 240, 913–929. doi: 10.1007/s00425-014-2078-2
Ravaglia, D., Espley, R., Henry-Kirk, R., Andreotti, C., Ziosi, V., Hellens, R., et al. (2013). Transcriptional regulation of flavonoid biosynthesis in nectarine (Prunus persica) by a set of R2R3 MYB transcription factors. BMC Plant Biol. 13:68. doi: 10.1186/1471-2229-13-68
Robinson, M. D., McCarthy, D. J., and Smyth, G. K. (2010). edgeR: a Bioconductor package for differential expression analysis of digital gene expression data. Bioinformatics 26, 139–140. doi: 10.1093/bioinformatics/btp616
Rodamilans, B., San León, D., Mühlberger, L., Candresse, T., Neumüller, M., Oliveros, J. C., et al. (2014). Transcriptomic analysis of Prunus domestica undergoing hypersensitive response to Plum pox virus infection. PLoS ONE 9:e100477. doi: 10.1371/journal.pone.0100477
Romero, I., Teresa Sanchez-Ballesta, M., Maldonado, R., Isabel Escribano, M., and Merodio, C. (2008). Anthocyanin, antioxidant activity and stress-induced gene expression in high CO2-treated table grapes stored at low temperature. J. Plant Physiol. 165, 522–530. doi: 10.1016/j.jplph.2006.12.011
Salvatierra, A., Pimentel, P., Moya-Leon, M. A., and Herrera, R. (2013). Increased accumulation of anthocyanins in Fragaria chiloensis fruits by transient suppression of FcMYB1 gene. Phytochemistry 90, 25–36. doi: 10.1016/j.phytochem.2013.02.016
Santhakumar, A. B., Kundur, A. R., Fanning, K., Netzel, M., Stanley, R., and Singh, I. (2015). Consumption of anthocyanin-rich Queen Garnet plum juice reduces platelet activation related thrombogenesis in healthy volunteers. J. Funct. Foods 12, 11–22. doi: 10.1016/j.jff.2014.10.026
Shen, X., Zhao, K., Liu, L., Zhang, K., Yuan, H., Liao, X., et al. (2014). A role for PacMYBA in ABA-regulated anthocyanin biosynthesis in red-colored sweet cherry cv. Hong Deng (Prunus avium L.). Plant Cell Physiol. 55, 862–880. doi: 10.1093/pcp/pcu013
Shin, J., Park, E., and Choi, G. (2007). PIF3 regulates anthocyanin biosynthesis in an HY5-dependent manner with both factors directly binding anthocyanin biosynthetic gene promoters in Arabidopsis. Plant J. 49, 981–994. doi: 10.1111/j.1365-313X.2006.03021.x
Shukitt-Hale, B., Kalt, W., Carey, A. N., Vinqvist-Tymchuk, M., McDonald, J., and Joseph, J. A. (2009). Plum juice, but not dried plum powder, is effective in mitigating cognitive deficits in aged rats. Nutrition 25, 567–573. doi: 10.1016/j.nut.2008.10.018
Song, S., Qi, T., Fan, M., Zhang, X., Gao, H., Huang, H., et al. (2013). The bHLH subgroup IIId factors negatively regulate jasmonate-mediated plant defense and development. PLoS Genet 9:e1003653. doi: 10.1371/journal.pgen.1003653
Starkevič, P., Paukštytė, J., Kazanavičiūtė, V., Denkovskienė, E., Stanys, V., Bendokas, V., et al. (2015). Expression and Anthocyanin Biosynthesis-Modulating Potential of Sweet Cherry (Prunus avium L.) MYB10 and bHLH Genes. PLoS ONE 10:e0126991. doi: 10.1371/journal.pone.0126991
Takos, A., Jaffe, F., Jacob, S. J. B., Robinson, S., and Walker, A. (2006). Light-induced expression of a MYB gene regulates anthocyanin biosynthesis in red apples. Plant Physiol. 142, 1216–1232. doi: 10.1104/pp.106
Tanaka, Y., Sasaki, N., and Ohmiya, A. (2008). Biosynthesis of plant pigments: anthocyanins, betalains, and carotenoids. Plant J. 54, 733–749. doi: 10.1111/j.1365-313X.2008.03447.x
Tuan, P., Bai, S., Yaegaki, H., Tamura, T., Hihara, S., Moriguchi, T., et al. (2015). The crucial role of PpMYB10.1 in anthocyanin accumulation in peach and relationships between its allelic type and skin color phenotype. BMC Plant Biol. 15:280. doi: 10.1186/s12870-015-0664-5
Umemura, H., Otagaki, S., Wada, M., Kondo, S., and Matsumoto, S. (2013). Expression and functional analysis of a novel MYB gene, MdMYB110a_JP, responsible for red flesh, not skin color in apple fruit. Planta 238, 65–76. doi: 10.1007/s00425-013-1875-3
Usenik, V., Štampar, F., and Veberič, R. (2009). Anthocyanins and fruit colour in plums (Prunus domestica L.) during ripening. Food Chem. 114, 529–534. doi: 10.1016/j.foodchem.2008.09.083
Valero, D., Díaz-Mula, H. M., Zapata, P. J., Guillén, F., Martínez-Romero, D., Castillo, S., et al. (2013). Effects of alginate edible coating on preserving fruit quality in four plum cultivars during postharvest storage. Postharvest Biol. Technol. 77, 1–6. doi: 10.1016/j.postharvbio.2012.10.011
Vimolmangkang, S., Zheng, D., Han, Y., Khan, M. A., Soria-Guerra, R. E., and Korban, S. S. (2014). Transcriptome analysis of the exocarp of apple fruit identifies light-induced genes involved in red color pigmentation. Gene 534, 78–87. doi: 10.1016/j.gene.2013.10.007
Walker, A. R., Lee, E., Bogs, J., McDavid, D. A. J., Thomas, M. R., and Robinson, S. P. (2007). White grapes arose through the mutation of two similar and adjacent regulatory genes. Plant J. 49, 772–785. doi: 10.1111/j.1365-313X.2006.02997.x
Wang, L., Feng, Z., Wang, X., Wang, X., and Zhang, X. (2010). DEGseq: an R package for identifying differentially expressed genes from RNA-seq data. Bioinformatics 26, 136–138. doi: 10.1093/bioinformatics/btp612
Wang, Z., Meng, D., Wang, A., Li, T., Jiang, S., Cong, P., et al. (2013). The methylation of the PcMYB10 promoter is associated with green-skinned sport in Max Red Bartlett pear. Plant Physiol. 162, 885–896. doi: 10.1104/pp.113.214700
Wei, H., Chen, X., Zong, X., Shu, H., Gao, D., and Liu, Q. (2015). Comparative transcriptome analysis of genes involved in anthocyanin biosynthesis in the red and yellow fruits of sweet cherry (Prunus avium L.). PLoS ONE 10:e0121164. doi: 10.1371/journal.pone.0121164
Winkel-Shirley, B. (2001). Flavonoid biosynthesis. A colorful model for genetics, biochemistry, cell biology, and biotechnology. Plant Physiol. 126, 485–493. doi: 10.1104/pp.126.2.485
Wrolstad, R. E., Culbertson, J. D., Cornwell, C. J., and Mattick, L. R. (1982). Detection of adulteration in blackberry juice concentrates and wines. J. Assoc. Off. Anal. Chem. 65, 1417–1423.
Wu, J., Fu, L., and Yi, H. (2016). Genome-wide identification of the transcription factors involved in citrus fruit ripening from the transcriptomes of a late-ripening sweet orange mutant and its wild type. PLoS ONE 11:e0154330. doi: 10.1371/journal.pone.0154330
Wu, J., Zhao, G., Yang, Y.-N., Le, W.-Q., Khan, M., Zhang, S.-L., et al. (2013). Identification of differentially expressed genes related to coloration in red/green mutant pear (Pyrus communis L.). Tree Genet. Genomes 9, 75–83. doi: 10.1007/s11295-012-0534-3
Wu, Z.-G., Jiang, W., Mantri, N., Bao, X.-Q., Chen, S.-L., and Tao, Z.-M. (2015). Transciptome analysis reveals flavonoid biosynthesis regulation and simple sequence repeats in yam (Dioscorea alata L.) tubers. BMC Genomics 16:346. doi: 10.1186/s12864-015-1547-8
Xie, X., Li, S., Zhang, R., Zhao, J., Chen, Y., Zhao, Q., et al. (2012). The bHLH transcription factor MdbHLH3 promotes anthocyanin accumulation and fruit colouration in response to low temperature in apples. Plant Cell Environ. 35, 1884–1897. doi: 10.1111/j.1365-3040.2012.02523.x
Xie, Y., Tan, H., Ma, Z., and Huang, J. (2016). DELLA proteins promote anthocyanin biosynthesis via sequestering MYBL2 and JAZ suppressors of the MYB/bHLH/WD40 complex in Arabidopsis thaliana. Mol. Plant. 9, 711–721. doi: 10.1016/j.molp.2016.01.014
Xu, F., Ning, Y., Zhang, W., Liao, Y., Li, L., Cheng, H., et al. (2013). An R2R3-MYB transcription factor as a negative regulator of the flavonoid biosynthesis pathway in Ginkgo biloba. Funct. Integr. Genomics 14, 177–189. doi: 10.1007/s10142-013-0352-1
Xu, W., Peng, H., Yang, T., Whitaker, B., Huang, L., Sun, J., et al. (2014). Effect of calcium on strawberry fruit flavonoid pathway gene expression and anthocyanin accumulation. Plant Physiol;. Biochem. 82, 289–298. doi: 10.1016/j.plaphy.2014.06.015
Xuan, J., Jia, Z., Qain, M., Zhang, J., Wang, G., and Guo, Z. (2015). Comparative of methods for RNA extraction from plum (Prunus salicina Lindl.) fruit flesh. North. Hort. 110–113. doi: 10.1093/jxb/erm213
Yang, Y.-N., Yao, G.-F., Zheng, D., Zhang, S.-L., Wang, C., Zhang, M.-Y., et al. (2015). Expression differences of anthocyanin biosynthesis genes reveal regulation patterns for red pear coloration. Plant Cell Rep. 34, 189–198. doi: 10.1007/s00299-014-1698-0
Yates, S. A., Swain, M. T., Hegarty, M. J., Chernukin, I., Matthew, L., and Allison, G. G. (2014). De novo assembly of red clover transcriptome based on RNA-Seq data provides insight into drought response, gene discovery and marker identification. BMC Genomics 15:453. doi: 10.1186/1471-2164-15-453
Ye, J., Fang, L., Zheng, H., Zhang, Y., Chen, J., Zhang, Z., et al. (2006). WEGO: a web tool for plotting GO annotations. Nucleic Acids Res 34, W293–W297. doi: 10.1093/nar/gkl031
Ye, J., Hu, T., Yang, C., Li, H., Yang, M., Ijaz, R., et al. (2015). Transcriptome profiling of tomato fruit development reveals transcription factors associated with ascorbic acid, carotenoid and flavonoid biosynthesis. PLoS ONE 10:e0130885. doi: 10.1371/journal.pone.0130885
Yoshida, K., Ma, D., and Constabel, C. P. (2015). The MYB182 protein downregulates proanthocyanidin and anthocyanin biosynthesis in poplar by repressing both structural and regulatory flavonoid genes. Plant Physiol. 167, 693–710. doi: 10.1104/pp.114.253674
Zabala, G., and Vodkin, L. O. (2014). Methylation affects transposition and splicing of a large CACTA transposon from a MYB transcription factor regulating anthocyanin synthase genes in soybean seed coats. PLoS ONE 9:e111959. doi: 10.1371/journal.pone.0111959
Zhai, R., Wang, Z., Zhang, S., Meng, G., Song, L., Wang, Z., et al. (2015). Two MYB transcription factors regulate flavonoid biosynthesis in pear fruit (Pyrus bretschneideri Rehd.). J. Exp. Bot. 67, 1275–1284. doi: 10.1093/jxb/erv524
Zhang, X. (2009). Research on Pigmentation and Mechanism of Anthocyanin Synthesis in Plum (Prunus spp.) Fruits. Ph.D. thesis, Zhejiang University, Hangzhou.
Zhou, H., Lin-Wang, K., Wang, H., Gu, C., Dare, A. P., Espley, R. V., et al. (2015). Molecular genetics of blood-fleshed peach reveals activation of anthocyanin biosynthesis by NAC transcription factors. Plant J. 82, 105–121. doi: 10.1111/tpj.12792
Zhou, Y., Zhou, H., Lin-Wang, K., Vimolmangkang, S., Espley, R. V., Wang, L., et al. (2014). Transcriptome analysis and transient transformation suggest an ancient duplicated MYB transcription factor as a candidate gene for leaf red coloration in peach. BMC Plant Biol. 14:388. doi: 10.1186/s12870-014-0388-y
Keywords: transcriptome, Prunus salicina Lindl., anthocyanin biosynthesis, fruit ripening, biosynthetic enzyme, transcription factor
Citation: Fang Z-Z, Zhou D-R, Ye X-F, Jiang C-C and Pan S-L (2016) Identification of Candidate Anthocyanin-Related Genes by Transcriptomic Analysis of ‘Furongli’ Plum (Prunus salicina Lindl.) during Fruit Ripening Using RNA-Seq. Front. Plant Sci. 7:1338. doi: 10.3389/fpls.2016.01338
Received: 31 May 2016; Accepted: 19 August 2016;
Published: 31 August 2016.
Edited by:
Claudio Bonghi, University of Padua, ItalyReviewed by:
Chuang Ma, Northwest Agricultural and Forestry University, ChinaCopyright © 2016 Fang, Zhou, Ye, Jiang and Pan. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Xin-Fu Ye, c3BlY2lhbGZydWl0QDEyNi5jb20=; eWV4aW5mdUAxMjYuY29t
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.