
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
MINI REVIEW article
Front. Plant Sci. , 31 March 2016
Sec. Plant Physiology
Volume 7 - 2016 | https://doi.org/10.3389/fpls.2016.00354
This article is part of the Research Topic Ion Transport in Chloroplast and Mitochondria Physiology in Green Organisms View all 11 articles
 Luca Carraretto1Vanessa Checchetto1,2
Luca Carraretto1Vanessa Checchetto1,2 Sara De Bortoli1
Sara De Bortoli1 Elide Formentin1,3
Elide Formentin1,3 Alex Costa4,5
Alex Costa4,5 Ildikó Szabó1,6*Enrico Teardo1,6*
Ildikó Szabó1,6*Enrico Teardo1,6*Plants, being sessile organisms, have evolved the ability to integrate external stimuli into metabolic and developmental signals. A wide variety of signals, including abiotic, biotic, and developmental stimuli, were observed to evoke specific spatio-temporal Ca2+ transients which are further transduced by Ca2+ sensor proteins into a transcriptional and metabolic response. Most of the research on Ca2+ signaling in plants has been focused on the transport mechanisms for Ca2+ across the plasma- and the vacuolar membranes as well as on the components involved in decoding of cytoplasmic Ca2+ signals, but how intracellular organelles such as mitochondria are involved in the process of Ca2+ signaling is just emerging. The combination of the molecular players and the elicitors of Ca2+ signaling in mitochondria together with newly generated detection systems for measuring organellar Ca2+ concentrations in plants has started to provide fruitful grounds for further discoveries. In the present review we give an updated overview of the currently identified/hypothesized pathways, such as voltage-dependent anion channels, homologs of the mammalian mitochondrial uniporter (MCU), LETM1, a plant glutamate receptor family member, adenine nucleotide/phosphate carriers and the permeability transition pore (PTP), that may contribute to the transport of Ca2+ across the outer and inner mitochondrial membranes in plants. We briefly discuss the relevance of the mitochondrial Ca2+ homeostasis for ensuring optimal bioenergetic performance of this organelle.
Molecular identification and pharmacological characterization of mitochondria-located ion channels allowed a deep understanding of the crucial importance of these proteins for organelle function and even for determining cell fate in animals (Leanza et al., 2014; Szabo and Zoratti, 2014). In plant mitochondria the current knowledge is unfortunately more limited than in the animal system and only few electrophysiological studies deal with plant mitochondrial ion channels. These include the voltage-dependent anion channel (VDAC), e.g., (Abrecht et al., 2000; Berrier et al., 2015) of the outer mitochondrial membrane (OMM) and those of the inner membrane (IMM), i.e., a large conductance Ca2+-insensitive potassium channel (Matkovic et al., 2011), a mitochondrial chloride channel (Matkovic et al., 2011) presumably corresponding to PIMAC (plant inner membrane anion channel) studied by classical bioenergetics (for review see Laus et al., 2008), a large-conductance Ca2+-activated BK-type potassium channel (Koszela-Piotrowska et al., 2009) and an ATP-dependent potassium channel K (ATP; De Marchi et al., 2010; Jarmuszkiewicz et al., 2010; Matkovic et al., 2011). In almost all studies either purified inner mitochondrial vesicles or purified proteins have been exploited upon incorporation into artificial membrane via the black lipid bilayer technique, because application of the patch clamp electrophysiological technique is experimentally very demanding (De Marchi et al., 2010). In addition to electrophysiological investigation, in-depth bioenergetic studies proved the existence and the relevance of potassium-permeable pathways (e.g., Pastore et al., 1999; Trono et al., 2014, 2015), of anion transport (Laus et al., 2008), of the proton-gradient dissipating uncoupling proteins (Vercesi et al., 2006) and of the permeability transition pore (Vianello et al., 2012; Zancani et al., 2015) in the context of plant mitochondrial physiology. Plant mitochondria have been shown to contain an uptake system for Ca2+ as well (Hanson et al., 1965; Dieter and Marme, 1980; Martins and Vercesi, 1985; Carnieri et al., 1987; Silva et al., 1992; Zottini and Zannoni, 1993): however, the molecular identification of the pathways mediating Ca2+ flux is still far from complete. Figure 1 summarizes the currently known/hypothesized pathways in plant mitochondria.
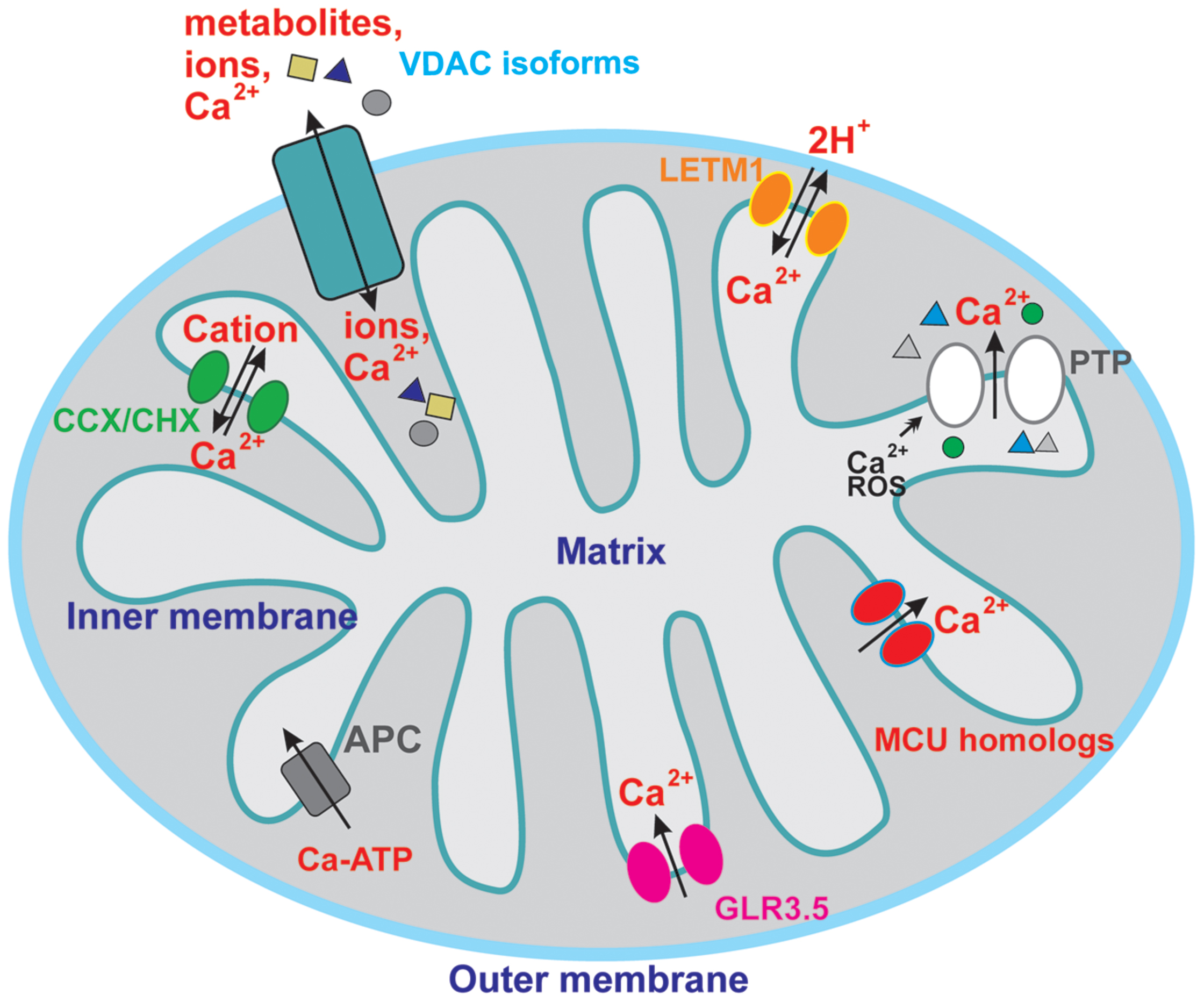
FIGURE 1. Overview of the putative calcium-permeable ion channels and transporters in plant mitochondria. For sake of clarity, only channels and transporters putatively involved in calcium transport are shown, so the channels mediating flux of other ions are not depicted here. See text for further details.
Understanding the routes and regulation of mitochondrial Ca2+ entry and exit would bring to an important advancement in the field and would help to determine the impact of Ca2+ homeostasis on bioenergetic efficiency and on the function of this organelle in plants. It has to be underlined that amongst the different signal transduction mechanisms Ca2+ plays a prominent role as a secondary messenger. A highly negative membrane potential in mitochondria (-150 to -180 mV) represents a huge driving force for the uptake of Ca2+ and other cations (Szabo and Zoratti, 2014) therefore calcium influx and efflux have to be under tight control in order to avoid calcium overload. In animal cells, transient accumulation of Ca2+ in intracellular organelles shapes cytosolic Ca2+ signals (Rizzuto et al., 2012) and a similar concept has been suggested for plants (Stael et al., 2012; Nomura and Shiina, 2014). The emerging view is that the influx of Ca2+ into the mitochondrial matrix can be induced by a variety of abiotic stresses such as heat, oxidative and salt stress, anoxia or extracellular ATP and extracellular Ca2+ (Logan and Knight, 2003; Xiong et al., 2006; Loro et al., 2012; Schwarzlander et al., 2012; Zhao et al., 2013; Rikhvanov et al., 2014; He et al., 2015; Pu et al., 2015), but the extent of uptake is variable among different tissues and species (Martins and Vercesi, 1985) as is the case also for the animals, (e.g., Fieni et al., 2012). The so far available data suggest that Ca2+ fluxes into and out of mitochondria might also shape the cytosolic “Ca2+ signature” (Stael et al., 2012; Nomura and Shiina, 2014), however, direct in vivo experimental proof has to be obtained in favor of this idea. In fact, in a recent study, where the calcium uptake into mitochondria has been impaired by deletion of the MCU regulator MICU (see below), cytoplasmic calcium transients appeared unaltered (Wagner et al., 2015).
Clear-cut demonstration of stimuli-induced Ca2+ uptake into mitochondria in intact plant tissues has been obtained thanks to the mitochondria-targeted genetically encoded Ca2+ sensors, aequorin (Logan and Knight, 2003) and Cameleon (Loro et al., 2012). Mitochondrial Ca2+ accumulation correlated to the intensity of Ca2+ increase in the cytoplasm. However, the molecular players of Ca2+ uptake and release remain to be largely elucidated. In addition, mostly indirect evidence indicate that in plant mitochondria, similarly to animal ones (Rizzuto et al., 2012), matrix Ca2+ levels regulate tricarboxylic acid (TCA) cycle enzymes (He et al., 2015) and oxidative phosphorylation (Wagner et al., 2015).
Several players participating in calcium homeostasis have been identified over the last decades. We briefly mention these pathways, although it has to be kept in mind, that while knowledge from the animal field might be a source of inspiration, plant mitochondria do not necessarily take advantage of the same systems.
Voltage-dependent anion channel (VDAC) isoforms are universally recognized as the main pathways for flux of metabolites and ions across the OMM (see, e.g., Shoshan-Barmatz et al., 2010; Checchetto et al., 2014; Mertins et al., 2014; Madamba et al., 2015). VDACs, although defined as anion channels, can conduct a substantial flow of Ca2+, as demonstrated both in vitro and in vivo for the mammalian protein (Gincel et al., 2001; Rapizzi et al., 2002; Bathori et al., 2006; Israelson et al., 2007; Rizzuto et al., 2009; Shoshan-Barmatz et al., 2010). As to Ca2+ uptake across the IMM, the 40 kDa protein (MCU) has been proposed to be the channel-forming component of the Ca2+ uniporter (Baughman, 2011; De Stefani et al., 2011). The uptake of Ca2+ via the uniporter has been linked to energy production (De Stefani et al., 2015). Lack of MCU in knock-out mice causes only modest defects in skeletal muscle strength and minor metabolic changes (Pan et al., 2013; Pendin et al., 2014) but MCU was shown to be important for bioenergetic performance in other models (Huang et al., 2013; Wu et al., 2015). In addition to MCU, several additional proteins such as MICU1 (Perocchi et al., 2010), EMRE (Sancak et al., 2013), MCUR1 (Mallilankaraman et al., 2012), and MCUb (a dominant negative MCU isoform; Raffaello et al., 2013) as well as MICU2 (Patron et al., 2014) were reported to be essential components and/or regulators of the mammalian MCU complex (MCUC; De Stefani et al., 2015; Foskett and Philipson, 2015). MICU1/2 proteins are EF-hand proteins with the ability to regulate mitochondrial Ca2+ uptake and MCU channel activity upon Ca2+ binding (Patron et al., 2014). EMRE was proposed to mediate the physical interaction between MCU and MICU1/MICU2 dimer and has recently been shown to fine-tune calcium-regulation of the channel activity on the matrix side (Vais et al., 2016). MCUR1 also affects mitochondrial Ca2+ uptake, however the underlying mechanism is still highly debated (Paupe et al., 2015; Vais et al., 2015).
Mitochondrial Ca2+ uptake might take place via additional or alternative components as well. In animals, LETM1 has been proposed to possess a Ca2+/2H+ electroneutral antiporter activity and to take up Ca2+ (Jiang et al., 2013; Doonan et al., 2014; Tsai et al., 2014). The yeast homolog Mdm38 however affects potassium homeostasis (Nowikovsky et al., 2004) and impacts mitochondrial translation, independently of its ion transporter function (Bauerschmitt et al., 2010). The nature of the ions transported by LETM1 is still highly debated (Nowikovsky et al., 2012; Nowikovsky and Bernardi, 2014). In addition, the solute carrier 25A23 (SLC25A23) has been shown to interact with MCU and was proposed to play an important role in mitochondrial Ca2+ influx (Hoffman et al., 2014).
It also has to be mentioned that calcium uptake is linked to cell death pathways via induction of the PTP (Bernardi et al., 2015). The PTP, first characterized in mammalian cells, is a channel responsible for the permeability increase of the inner mitochondrial membrane under specific conditions (Zoratti and Szabo, 1995; Bernardi et al., 2015). PTP can be activated by different stimuli such as high matrix Ca2+ concentration (in 100s μM range) and oxidative stress, leading to swelling of mitochondria and dissipation of energy. In animals, when PTP opens only transiently, it possibly mediates Ca2+ release from mitochondria (Bernardi and von Stockum, 2012) together with the recently identified 3Na+/Ca2+ antiporter (NCX; Palty et al., 2010). In addition, a still unidentified Na+-insensitive Ca2+ release system, possibly a H+/Ca2+ antiporter plays a role (Nowikovsky et al., 2012). Over the last decades VDAC, the adenine nucleotide carrier, the benzodiazepine receptor and cyclophilin D (CypD) were proposed in different combinations as the main components of the mammalian PTP (Zoratti and Szabo, 1995; Bernardi et al., 2015). The discovery that oxidative stress and application of elevated [Ca2+] results in channel formation by the dimeric form of the F-ATP synthase, opened a new perspective to the field (Giorgio et al., 2013). A recent work proposed instead, that mitochondrial spastic paraplegia 7 (SPG7), a nuclear-encoded mitochondrial metalloprotease (m-AAA) which interacts with CypD and VDAC1 and with a paraplegin-like protein AFG3L2, is essential for the PTP complex formation (Shanmughapriya et al., 2015). However, PTP could still be opened in the absence of SPG7, although at higher matrix calcium concentrations, suggesting that SPG7, similarly to CypD, acts a regulator rather than a crucial pore-forming moiety of the PTP (Bernardi and Forte, 2015). Interestingly, apart from their proteolytic roles, the m-AAA proteases mediate ATP-dependent membrane dislocation of the heme-binding reactive oxygen scavenger protein Ccp1 (Tatsuta et al., 2007), possibly linking PTP activation to oxidative stress.
In higher plants, similarly to animals, functionally distinct isoforms of VDAC exist. In particular, in Arabidopsis various isoforms displaying distinct subcellular localization (in the plasma membrane, mitochondria, chloroplasts, and plastids) and function have been identified (Smack and Colombini, 1985; Pottosin, 1993; Clausen et al., 2004; Tateda et al., 2011; Homble et al., 2012; Robert et al., 2012; Takahashi and Tateda, 2013; Michaud et al., 2014). Their roles, as assessed mostly by using T-DNA insertion knockout mutants of Arabidopsis, include regulation of development (Tateda et al., 2011; Robert et al., 2012; Pan et al., 2014), regulation of the hypersensitive response/programmed cell death (Lacomme and Roby, 1999; Tateda et al., 2009, 2011), of the response to abiotic stress (Li et al., 2013; Zhang et al., 2015) and import of tRNA into mitochondria (Salinas et al., 2006). Whether any of these functions requires Ca2+ flux across the OMM (or across other membranes) mediated by VDACs is unclear, however AtVDAC1 has been shown to interact in two-hybrid yeast system with CBL1, a Ca2+-sensor (Li et al., 2013). Despite detailed electrophysiological characterization of several isoforms (Blumenthal et al., 1993; Pottosin, 1993; Mlayeh et al., 2010; Godbole et al., 2011; Berrier et al., 2015), experimental evidence is still missing to understand whether and how VDAC proteins influence Ca2+ flux across plant endomembranes.
In the Arabidopsis thaliana genome six genes are present which can be identified as putative MCU channel proteins with predicted mitochondrial targeting, since they display sequence similarity with the mammalian MCU counterparts and contain the conserved DVME (Asp-Val-Met-Glu) selectivity filter sequence (Stael et al., 2012). Varying number of homologs can be identified in the genome of other higher plants as well (see Aramemnon http://aramemnon.uni-koeln.de/). Whether all these isoforms are indeed targeted to mitochondria, whether they form ion channels able to provide a permeation pathway for Ca2+ and whether the various isoforms operate in different tissues and/or at different developmental stages still awaits clarification. Recently obtained experimental evidence indicates that at least one of the isoforms is indeed targeted to mitochondria in Arabidopsis (Carraretto et al., 2016). The discovery showing that lack of the only existing isoform of the regulator, AtMICU, in Arabidopsis alters mitochondrial Ca2+ uptake points to a functional conservation of the core-components of the MCU complex in plants (Wagner et al., 2015). Interestingly, even though Ca2+ uptake into mitochondria and basal Ca2+ are significantly higher in AtMICU-less plants than in WT plants, respiration and mitochondrial morphology are only slightly affected and plant development is normal (Wagner et al., 2015). As to EMRE, its close homologs do not seem to be present in higher plants, while one of the two homologs of MCUR1 in Arabidopsis has been described as a plant specific subunit of complex IV (Millar et al., 2004; Klodmann et al., 2011). In summary, the plant homologs of MCU and MICU1 are certainly excellent candidates to be key players in mitochondrial Ca2+ homeostasis, but experimental proof for the ability of MCU proteins to form Ca2+-permeable channels is still lacking.
Similarly to the animal mitochondria, alternative calcium flux-mediating pathways seem to exist also in higher plants. Our knowledge in this respect is restricted mostly to Arabidopsis, since T-DNA insertion mutants of this model plant are available and widely used. In Arabidopsis deletion of both isoforms of LETM is lethal, probably due to the requirement of LETM proteins for mitochondrial protein translation and accumulation (Zhang et al., 2012). A recent study provided evidence that members of the APC family of Arabidopsis mediate a time dependent uptake of [45Ca] (in the form of Ca-ATP) in vitro (Lorenz et al., 2015). This system is homolog of the above mentioned SLC25A23. The transport rate of AtAPC2 was however low and was completely blocked by 25-fold excess of Mg2+, suggesting that in vivo Ca2+ flux might take place via this transporter only under specific conditions in plant mitochondria.
Another study located an alternative spliced isoform of a member of the glutamate receptor family, AtGLR3.5 to mitochondria. Although there is no direct evidence thus far demonstrating that the subfamily 3 member AtGLR3.5 functions as Ca2+-permeable ion channel, the close homolog AtGLR3.4 and the AtGLR1.4 and AtGLR1.1 pores behave as Ca2+-permeable non-selective cation channels when expressed in heterologous systems (Tapken and Hollmann, 2008; Vincill et al., 2012; Tapken et al., 2013). In addition, inner membrane vesicles isolated from spinach chloroplasts and containing members of the GLR subfamily 3 harbor a glutamate/glycine-induced Ca2+-permeable activity which is sensitive to known animal ionotropic glutamate receptor antagonists (Teardo et al., 2010). Studies using Atglr3.3 mutant plants showed that Ca2+ uptake induced by glutamate in Arabidopsis into hypocotyls and root cells is correlated with the presence of AtGLR3.3 (Qi et al., 2006). Therefore, similarly to the other members of the subfamily 3, AtGLR3.5 is expected to work as Ca2+-permeable channel. In accordance, Ca2+ dynamics measurements performed using the Cameleon probe targeted to mitochondria in WT and mutant plants lacking AtGLR3.5 revealed that AtGLR3.5 might mediate Ca2+ uptake into mitochondria, at least in response to specific stimuli (e.g., wounding (Teardo et al., 2015).
In agreement with the mitochondrial localization and predicted activity of AtGLR3.5, plants lacking this protein harbor mitochondria with profoundly altered ultrastructure: a dramatic loss of cristae and swelling with the matrix becoming translucent can be observed. The exact mechanism leading to these changes is not known, even though AtGLR3.5 topology indicates that the glutamate (agonist)-binding domains are located in the cytosol, possibly sensing the cytosolic glutamate/aminoacid concentration (Teardo et al., 2015). Whether, a reduced Ca2+ uptake in the Atglr3.5 KO plants leads to morphological changes via reduction of the oxidative phosphorylation remains to be established. It is interesting to note that complex I inhibition in mammalian mitochondria leads to similar morphological changes to that observed for Atglr3.5 KO plants (Ramonet et al., 2013).
Plant mitochondria can also undergo Ca2+-induced permeability transition, (Arpagaus et al., 2002; Petrussa et al., 2004; Vianello et al., 2012), an event linked to nitric oxide-induced cell death (Saviani et al., 2002) and to programmed cell death (for updated review see Zancani et al., 2015). Whether the “life-sustaining” F-ATP synthase forms the PTP in plants as well, still has to be clarified, but the thylakoid membrane, which also contains this ATP-producing machinery, harbors a high-conductance channel resembling PTP (Hinnah and Wagner, 1998). Ccp1 (see above) shares high sequence homology with ascorbate peroxidase (APX), with one of the isoforms dually targeted to mitochondria and chloroplasts (Chew et al., 2003). This further supports previous findings in the literature that ROS regulates PTP also in plants. A BLAST search in the Arabidopsis database reveals that it contains several ATP-dependent metalloproteases (FtsH proteases) that show high aminoacid sequence similarity to SPG7 (see above; e-values ranging from 2e-169 to 8e-58, with aminoacid identities in the range of 36–49%) and to AFG3L2 (e-value of 0 and 50% identity is found for AtFtsH3 and AtFtsH10). Among those with the highest score, AtFtsH3 and AtFtsH10 are present in mitochondria (Piechota et al., 2010).
Several plant putative Ca2+/cation exchangers, for example CCX1, CCX3, and CCX4 (Schwacke et al., 2003), score for mitochondrial localization according to Aramemnon database although in most species rather poorly. Experimental evidence in favor of their localization in mitochondria versus secretory pathways or of a dual localization is missing. Furthermore, for AtCCX3 it has been established that it functions as an endomembrane-localized H+-dependent K+ transporter (Morris et al., 2008). A putative cation/proton exchanger, AtCHX25 has also a predicted mitochondrial localization, but whether it mediates K+/H+ exchange like some members of the AtCHX family or Ca2+/H+ exchange has still to be established (Sze et al., 2004; Evans et al., 2012; Chanroj et al., 2013).
Under biologically relevant conditions (e.g., during oxidative stress), the above-described uptake and efflux pathways for calcium might cooperate. For example, Ca2+ influx-triggered Ca2+ release has been linked to pulsing of the mitochondrial membrane potential, a phenomena proposed to yield a transient uncoupling leading to reduced ROS production (Schwarzlander et al., 2012).
In summary, a combination of genetics, fluorescent probe imaging, electrophysiology, bioenergetics and physiology will hopefully provide answers to the numerous open questions related to mitochondrial calcium homeostasis in plants.
All authors contributed to the works published by our groups which are described in the minireview. Furthermore, all authors actively participated in writing the manuscript.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The authors are grateful to the Italian Ministry (PRIN 2010CSJX4F to IS and AC), and to Human Frontiers Science Program (HFSP0052 to IS) and the FIRB program (RBFR10S1LJ_001 to AC) for funding. We also thank all co-authors who participated in the works mentioned in the text that were published by our groups.
Abrecht, H., Wattiez, R., Ruysschaert, J. M., and Homble, F. (2000). Purification and characterization of two voltage-dependent anion channel isoforms from plant seeds. Plant Physiol. 124, 1181–1190. doi: 10.1104/pp.124.3.1181
Arpagaus, S., Rawyler, A., and Braendle, R. (2002). Occurrence and characteristics of the mitochondrial permeability transition in plants. J. Biol. Chem. 277, 1780–1787. doi: 10.1074/jbc.M109416200
Bathori, G., Csordas, G., Garcia-Perez, C., Davies, E., and Hajnoczky, G. (2006). Ca2+-dependent control of the permeability properties of the mitochondrial outer membrane and voltage-dependent anion-selective channel (VDAC). J. Biol. Chem. 281, 17347–17358. doi: 10.1074/jbc.M600906200
Bauerschmitt, H., Mick, D. U., Deckers, M., Vollmer, C., Funes, S., Kehrein, K., et al. (2010). Ribosome-binding proteins Mdm38 and Mba1 display overlapping functions for regulation of mitochondrial translation. Mol. Biol. Cell 21, 1937–1944. doi: 10.1091/mbc.E10-02-0101
Baughman, J. M. (2011). Integrative genomics identifies MCU as an essential component of the mitochondrial calcium uniporter. Nature 476, 341–345. doi: 10.1038/nature10234
Bernardi, P., and Forte, M. (2015). Commentary: SPG7 is an essential and conserved component of the mitochondrial permeability transition pore. Front. Physiol. 6:320. doi: 10.3389/fphys.2015.00320
Bernardi, P., Rasola, A., Forte, M., and Lippe, G. (2015). The mitochondrial permeability transition pore: channel formation by F-ATP synthase, integration in signal transduction, and role in pathophysiology. Physiol. Rev. 95, 1111–1155. doi: 10.1152/physrev.00001.2015
Bernardi, P., and von Stockum, S. (2012). The permeability transition pore as a Ca2+ release channel: new answers to an old question. Cell Calcium 52, 22–27. doi: 10.1016/j.ceca.2012.03.004
Berrier, C., Peyronnet, R., Betton, J. M., Ephritikhine, G., Barbier-Brygoo, H., Frachisse, J. M., et al. (2015). Channel characteristics of VDAC-3 from Arabidopsis thaliana. Biochem. Biophys. Res. Commun. 459, 24–28. doi: 10.1016/j.bbrc.2015.02.034
Blumenthal, A., Kahn, K., Beja, O., Galun, E., Colombini, M., and Breiman, A. (1993). Purification and characterization of the voltage-dependent anion-selective channel protein from wheat mitochondrial membranes. Plant Physiol. 101, 579–587.
Carnieri, E. G., Martins, I. S., and Vercesi, A. E. (1987). The mechanism and biological role of calcium transport by plant mitochondria. Brazil. J. Med. Biol. Res. 20, 635–638.
Carraretto, L., Teardo, E., Checchetto, V., Finazzi, G., Uozumi, N., and Szabo, I. (2016). Ion channels in plant bioenergetic organelles chloroplast and mitochondria: from molecular identification to function. Mol. Plant 9, 371–395. doi: 10.1016/j.molp.2015.12.004
Chanroj, S., Padmanaban, S., Czerny, D. D., Jauh, G. Y., and Sze, H. (2013). K+ transporter AtCHX17 with its hydrophilic C tail localizes to membranes of the secretory/endocytic system: role in reproduction and seed set. Mol. Plant 6, 1226–1246. doi: 10.1093/mp/sst032
Checchetto, V., Reina, S., Magri, A., Szabo, I., and De Pinto, V. (2014). Recombinant human voltage dependent anion selective channel isoform 3 (hVDAC3) forms pores with a very small conductance. Cell. Physiol. Biochem. 34, 842–853. doi: 10.1159/000363047
Chew, O., Whelan, J., and Millar, A. H. (2003). Molecular definition of the ascorbate-glutathione cycle in Arabidopsis mitochondria reveals dual targeting of antioxidant defenses in plants. J. Biol. Chem. 278, 46869–46877. doi: 10.1074/jbc.M307525200
Clausen, C., Ilkavets, I., Thomson, R., Philippar, K., Vojta, A., Mohlmann, T., et al. (2004). Intracellular localization of VDAC proteins in plants. Planta 220, 30–37. doi: 10.1007/s00425-004-1325-3
De Marchi, U., Checchetto, V., Zanetti, M., Teardo, E., Soccio, M., Formentin, E., et al. (2010). ATP-sensitive cation-channel in wheat (Triticum durum Desf.): identification and characterization of a plant mitochondrial channel by patch-clamp. Cell. Physiol. Biochem. 26, 975–982. doi: 10.1159/000324010
De Stefani, D., Patron, M., and Rizzuto, R. (2015). Structure and function of the mitochondrial calcium uniporter complex. Biochim. Biophys. Acta 1853, 2006–2011. doi: 10.1016/j.bbamcr.2015.04.008
De Stefani, D., Raffaello, A., Teardo, E., Szabo, I., and Rizzuto, R. (2011). A forty-kilodalton protein of the inner membrane is the mitochondrial calcium uniporter. Nature 476, 336–340. doi: 10.1038/nature10230
Dieter, P., and Marme, D. (1980). Ca2+ transport in mitochondrial and microsomal fractions from higher plants. Planta 150, 1–8. doi: 10.1007/BF00385606
Doonan, P. J., Chandramoorthy, H. C., Hoffman, N. E., Zhang, X., Cardenas, C., Shanmughapriya, S., et al. (2014). LETM1-dependent mitochondrial Ca2+ flux modulates cellular bioenergetics and proliferation. FASEB J. 28, 4936–4949. doi: 10.1096/fj.14-256453
Evans, A. R., Hall, D., Pritchard, J., and Newbury, H. J. (2012). The roles of the cation transporters CHX21 and CHX23 in the development of Arabidopsis thaliana. J. Exp. Bot. 63, 59–67. doi: 10.1093/jxb/err271
Fieni, F., Lee, S. B., Jan, Y. N., and Kirichok, Y. (2012). Activity of the mitochondrial calcium uniporter varies greatly between tissues. Nat. Commun. 3:1317. doi: 10.1038/ncomms2325
Foskett, J. K., and Philipson, B. (2015). The mitochondrial Ca2+ uniporter complex. J. Mol. Cell Cardiol. 78, 3–8. doi: 10.1016/j.yjmcc.2014.11.015
Gincel, D., Zaid, H., and Shoshan-Barmatz, V. (2001). Calcium binding and translocation by the voltage-dependent anion channel: a possible regulatory mechanism in mitochondrial function. Biochem. J. 358, 147–155. doi: 10.1042/bj3580147
Giorgio, V., von Stockum, S., Antoniel, M., Fabbro, A., Fogolari, F., Forte, M., et al. (2013). Dimers of mitochondrial ATP synthase form the permeability transition pore. Proc. Natl. Acad. Sci. U.S.A. 110, 5887–5892. doi: 10.1073/pnas.1217823110
Godbole, A., Mitra, R., Dubey, A. K., Reddy, P. S., and Mathew, M. K. (2011). Bacterial expression, purification and characterization of a rice voltage-dependent, anion-selective channel isoform, OsVDAC4. J. Membr. Biol. 244, 67–80. doi: 10.1007/s00232-011-9399-x
Hanson, J. B., Malhotra, S. S., and Stoner, C. D. (1965). Action of calcium on corn mitochondria. Plant Physiol. 40, 1033–1040. doi: 10.1104/pp.40.6.1033
He, L., Li, B., Lu, X., Yuan, L., Yang, Y., Yuan, Y., et al. (2015). The effect of exogenous calcium on mitochondria, respiratory metabolism enzymes and ion transport in cucumber roots under hypoxia. Sci. Rep. 5:11391. doi: 10.1038/srep11391
Hinnah, S. C., and Wagner, R. (1998). Thylakoid membranes contain a high-conductance channel. Eur. J. Biochem. 253, 606–613. doi: 10.1046/j.1432-1327.1998.2530606.x
Hoffman, N. E., Chandramoorthy, H. C., Shanmughapriya, S., Zhang, X. Q., Vallem, S., Doonan, P. J., et al. (2014). SLC25A23 augments mitochondrial Ca2+ uptake, interacts with MCU, and induces oxidative stress-mediated cell death. Mol. Biol. Cell 25, 936–947. doi: 10.1091/mbc.E13-08-0502
Homble, F., Krammer, E. M., and Prevost, M. (2012). Plant VDAC: facts and speculations. Biochim. Biophys. Acta 1818, 1486–1501. doi: 10.1016/j.bbamem.2011.11.028
Huang, G., Vercesi, A. E., and Docampo, R. (2013). Essential regulation of cell bioenergetics in Trypanosoma brucei by the mitochondrial calcium uniporter. Nat. Commun. 4:2865. doi: 10.1038/ncomms3865
Israelson, A., Abu-Hamad, S., Zaid, H., Nahon, E., and Shoshan-Barmatz, V. (2007). Localization of the voltage-dependent anion channel-1 Ca2+-binding sites. Cell Calcium 41, 235–244. doi: 10.1016/j.ceca.2006.06.005
Jarmuszkiewicz, W., Matkovic, K., and Koszela-Piotrowska, I. (2010). Potassium channels in the mitochondria of unicellular eukaryotes and plants. FEBS Lett. 584, 2057–2062. doi: 10.1016/j.febslet.2010.01.023
Jiang, D., Zhao, L., Clish, C. B., and Clapham, D. E. (2013). Letm1, the mitochondrial Ca2+/H+ antiporter, is essential for normal glucose metabolism and alters brain function in Wolf-Hirschhorn syndrome. Proc. Natl. Acad. Sci. U.S.A. 110, E2249–E2254. doi: 10.1073/pnas.1308558110
Klodmann, J., Senkler, M., Rode, C., and Braun, H. P. (2011). Defining the protein complex proteome of plant mitochondria. Plant Physiol. 157, 587–598. doi: 10.1104/pp.111.182352
Koszela-Piotrowska, I., Matkovic, K., Szewczyk, A., and Jarmuszkiewicz, W. (2009). A large-conductance calcium-activated potassium channel in potato (Solanum tuberosum) tuber mitochondria. Biochem. J. 424, 307–316. doi: 10.1042/BJ20090991
Lacomme, C., and Roby, D. (1999). Identification of new early markers of the hypersensitive response in Arabidopsis thaliana (1). FEBS Lett. 459, 149–153. doi: 10.1016/S0014-5793(99)01233-8
Laus, M. N., Soccio, M., Trono, D., Cattivelli, L., and Pastore, D. (2008). Plant inner membrane anion channel (PIMAC) function in plant mitochondria. Plant Cell Physiol. 49, 1039–1055. doi: 10.1093/pcp/pcn082
Leanza, L., Zoratti, M., Gulbins, E., and Szabo, I. (2014). Mitochondrial ion channels as oncological targets. Oncogene 33, 5569–5581. doi: 10.1038/onc.2013.578
Li, Z. Y., Xu, Z. S., He, G. Y., Yang, G. X., Chen, M., Li, L. C., et al. (2013). The voltage-dependent anion channel 1 (AtVDAC1) negatively regulates plant cold responses during germination and seedling development in Arabidopsis and interacts with calcium sensor CBL1. Int. J. Mol. Sci. 14, 701–713. doi: 10.3390/ijms14010701
Logan, D. C., and Knight, M. R. (2003). Mitochondrial and cytosolic calcium dynamics are differentially regulated in plants. Plant Physiol. 133, 21–24. doi: 10.1104/pp.103.026047
Lorenz, A., Lorenz, M., Vothknecht, U. C., Niopek-Witz, S., Neuhaus, H. E., and Haferkamp, I. (2015). In vitro analyses of mitochondrial ATP/phosphate carriers from Arabidopsis thaliana revealed unexpected Ca2+-effects. BMC Plant Biol. 15:238. doi: 10.1186/s12870-015-0616-0
Loro, G., Drago, I., Pozzan, T., Schiavo, F. L., Zottini, M., and Costa, A. (2012). Targeting of Cameleons to various subcellular compartments reveals a strict cytoplasmic/mitochondrial Ca2+ handling relationship in plant cells. Plant J. 71, 1–13. doi: 10.1111/j.1365-313X.2012.04968.x
Madamba, S. M., Damri, K. N., Dejean, L. M., and Peixoto, P. M. (2015). Mitochondrial ion channels in cancer transformation. Front. Oncol. 5:120. doi: 10.3389/fonc.2015.00120
Mallilankaraman, K., Cardenas, C., Doonan, P. J., Chandramoorthy, H. C., Irrinki, K. M., Golenar, T., et al. (2012). MCUR1 is an essential component of mitochondrial Ca2+ uptake that regulates cellular metabolism. Nat. Cell Biol. 14, 1336–1343. doi: 10.1038/ncb2622
Martins, I. S., and Vercesi, A. E. (1985). Some characteristics of Ca2+ transport in plant mitochondria. Biochem. Biophys. Res. Commun. 129, 943–948. doi: 10.1016/0006-291X(85)91982-5
Matkovic, K., Koszela-Piotrowska, I., Jarmuszkiewicz, W., and Szewczyk, A. (2011). Ion conductance pathways in potato tuber (Solanum tuberosum) inner mitochondrial membrane. Biochim. Biophys. Acta 1807, 275–285. doi: 10.1016/j.bbabio.2010.12.001
Mertins, B., Psakis, G., and Essen, L. O. (2014). Voltage-dependent anion channels: the wizard of the mitochondrial outer membrane. Biol. Chem. 395, 1435–1442. doi: 10.1515/hsz-2014-0203
Michaud, M., Ubrig, E., Filleur, S., Erhardt, M., Ephritikhine, G., Marechal-Drouard, L., et al. (2014). Differential targeting of VDAC3 mRNA isoforms influences mitochondria morphology. Proc. Natl. Acad. Sci. U.S.A. 111, 8991–8996. doi: 10.1073/pnas.1402588111
Millar, A. H., Eubel, H., Jansch, L., Kruft, V., Heazlewood, J. L., and Braun, H. P. (2004). Mitochondrial cytochrome c oxidase and succinate dehydrogenase complexes contain plant specific subunits. Plant Mol. Biol. 56, 77–90. doi: 10.1007/s11103-004-2316-2
Mlayeh, L., Chatkaew, S., Leonetti, M., and Homble, F. (2010). Modulation of plant mitochondrial VDAC by phytosterols. Biophys. J. 99, 2097–2106. doi: 10.1016/j.bpj.2010.07.067
Morris, J., Tian, H., Park, S., Sreevidya, C. S., Ward, J. M., and Hirschi, K. D. (2008). AtCCX3 is an Arabidopsis endomembrane H+ -dependent K+ transporter. Plant Physiol. 148, 1474–1486. doi: 10.1104/pp.108.118810
Nomura, H., and Shiina, T. (2014). Calcium signaling in plant endosymbiotic organelles: mechanism and role in physiology. Mol. Plant 7, 1094–1104. doi: 10.1093/mp/ssu020
Nowikovsky, K., and Bernardi, P. (2014). LETM1 in mitochondrial cation transport. Front. Physiol. 5:83. doi: 10.3389/fphys.2014.00083
Nowikovsky, K., Froschauer, E. M., Zsurka, G., Samaj, J., Reipert, S., Kolisek, M., et al. (2004). The LETM1/YOL027 gene family encodes a factor of the mitochondrial K+ homeostasis with a potential role in the Wolf-Hirschhorn syndrome. J. Biol. Chem. 279, 30307–30315. doi: 10.1074/jbc.M403607200
Nowikovsky, K., Pozzan, T., Rizzuto, R., Scorrano, L., and Bernardi, P. (2012). Perspectives on: SGP symposium on mitochondrial physiology and medicine: the pathophysiology of LETM1. J. Gen. Physiol. 139, 445–454. doi: 10.1085/jgp.201110757
Palty, R., Silverman, W. F., Hershfinkel, M., Caporale, T., Sensi, S. L., Parnis, J., et al. (2010). NCLX is an essential component of mitochondrial Na+/Ca2+ exchange. Proc. Natl. Acad. Sci. U.S.A. 107, 436–441. doi: 10.1073/pnas.0908099107
Pan, X., Chen, Z., Yang, X., and Liu, G. (2014). Arabidopsis voltage-dependent anion channel 1 (AtVDAC1) is required for female development and maintenance of mitochondrial functions related to energy-transaction. PLoS ONE 9:e106941. doi: 10.1371/journal.pone.0106941
Pan, X., Liu, J., Nguyen, T., Liu, C., Sun, J., Teng, Y., et al. (2013). The physiological role of mitochondrial calcium revealed by mice lacking the mitochondrial calcium uniporter. Nat. Cell Biol. 15, 1464–1472. doi: 10.1038/ncb2868
Pastore, D., Stoppelli, M. C., Di Fonzo, N., and Passarella, S. (1999). The existence of the K(+) channel in plant mitochondria. J. Biol. Chem. 274, 26683–26690. doi: 10.1074/jbc.274.38.26683
Patron, M., Checchetto, V., Raffaello, A., Teardo, E., Vecellio Reane, D., Mantoan, M., et al. (2014). MICU1 and MICU2 finely tune the mitochondrial Ca2+ uniporter by exerting opposite effects on MCU activity. Mol. Cell. 53, 726–737. doi: 10.1016/j.molcel.2014.01.013
Paupe, V., Prudent, J., Dassa, E. P., Rendon, O. Z., and Shoubridge, E. A. (2015). CCDC90A (MCUR1) is a cytochrome c oxidase assembly factor and not a regulator of the mitochondrial calcium uniporter. Cell Metabol. 21, 109–116. doi: 10.1016/j.cmet.2014.12.004
Pendin, D., Greotti, E., and Pozzan, T. (2014). The elusive importance of being a mitochondrial Ca2+ uniporter. Cell Calcium 55, 139–145. doi: 10.1016/j.ceca.2014.02.008
Perocchi, F., Gohil, V. M., Girgis, H. S., Bao, X. R., McCombs, J. E., Palmer, A. E., et al. (2010). MICU1 encodes a mitochondrial EF hand protein required for Ca2+ uptake. Nature 467, 291–296. doi: 10.1038/nature09358
Petrussa, E., Casolo, V., Peresson, C., Braidot, E., Vianello, A., and Macri, F. (2004). The K(ATP)+ channel is involved in a low-amplitude permeability transition in plant mitochondria. Mitochondrion 3, 297–307. doi: 10.1016/j.mito.2004.01.002
Piechota, J., Kolodziejczak, M., Juszczak, I., Sakamoto, W., and Janska, H. (2010). Identification and characterization of high molecular weight complexes formed by matrix AAA proteases and prohibitins in mitochondria of Arabidopsis thaliana. J. Biol. Chem. 285, 12512–12521. doi: 10.1074/jbc.M109.063644
Pottosin, I. I. (1993). One of the chloroplast envelope ion channels is probably related to the mitochondrial VDAC. FEBS Lett. 330, 211–214. doi: 10.1016/0014-5793(93)80275-Y
Pu, X., Lv, X., Tan, T., Fu, F., Qin, G., and Lin, H. (2015). Roles of mitochondrial energy dissipation systems in plant development and acclimation to stress. Ann. Bot. 116, 583–600. doi: 10.1093/aob/mcv063
Qi, Z., Stephens, N. R., and Spalding, E. P. (2006). Calcium entry mediated by GLR3.3, an Arabidopsis glutamate receptor with a broad agonist profile. Plant Physiol. 142, 963–971. doi: 10.1104/pp.106.088989
Raffaello, A., De Stefani, D., Sabbadin, D., Teardo, E., Merli, G., Picard, A., et al. (2013). The mitochondrial calcium uniporter is a multimer that can include a dominant-negative pore-forming subunit. EMBO J. 32, 2362–2376. doi: 10.1038/emboj.2013.157
Ramonet, D., Perier, C., Recasens, A., Dehay, B., Bove, J., Costa, V., et al. (2013). Optic atrophy 1 mediates mitochondria remodeling and dopaminergic neurodegeneration linked to complex I deficiency. Cell Death. Differ. 20, 77–85. doi: 10.1038/cdd.2012.95
Rapizzi, E., Pinton, P., Szabadkai, G., Wieckowski, M. R., Vandecasteele, G., Baird, G., et al. (2002). Recombinant expression of the voltage-dependent anion channel enhances the transfer of Ca2+ microdomains to mitochondria. J. Cell Biol. 159, 613–624. doi: 10.1083/jcb.200205091
Rikhvanov, E. G., Fedoseeva, I. V., Pyatrikas, D. V., Borovskii, G. B., and Voinikov, V. K. (2014). Role of mitochondria in the operation of calcium signaling system in heatstressed plants. Russ. J. Plant Physiol. 61, 141–153. doi: 10.1134/S1021443714020125
Rizzuto, R., De Stefani, D., Raffaello, A., and Mammucari, C. (2012). Mitochondria as sensors and regulators of calcium signalling. Nat. Rev. Mol. Cell Biol. 13, 566–578. doi: 10.1038/nrm3412
Rizzuto, R., Marchi, S., Bonora, M., Aguiari, P., Bononi, A., De Stefani, D., et al. (2009). Ca2+ transfer from the ER to mitochondria: when, how and why. Biochim. Biophys. Acta 1787, 1342–1351. doi: 10.1016/j.bbabio.2009.03.015
Robert, N., d’Erfurth, I., Marmagne, A., Erhardt, M., Allot, M., Boivin, K., et al. (2012). Voltage-dependent-anion-channels (VDACs) in Arabidopsis have a dual localization in the cell but show a distinct role in mitochondria. Plant Mol. Biol. 78, 431–446. doi: 10.1007/s11103-012-9874-5
Salinas, T., Duchene, A. M., Delage, L., Nilsson, S., Glaser, E., Zaepfel, M., et al. (2006). The voltage-dependent anion channel, a major component of the tRNA import machinery in plant mitochondria. Proc. Natl. Acad. Sci. U.S.A. 103, 18362–18367. doi: 10.1073/pnas.0606449103
Sancak, Y., Markhard, A. L., Kitami, T., Kovacs-Bogdan, E., Kamer, K. J., Udeshi, N. D., et al. (2013). EMRE is an essential component of the mitochondrial calcium uniporter complex. Science 342, 1379–1382. doi: 10.1126/science.1242993
Saviani, E. E., Orsi, C. H., Oliveira, J. F., Pinto-Maglio, C. A., and Salgado, I. (2002). Participation of the mitochondrial permeability transition pore in nitric oxide-induced plant cell death. FEBS Lett. 510, 136–140. doi: 10.1016/S0014-5793(01)03230-6
Schwacke, R., Schneider, A., van der Graaff, E., Fischer, K., Catoni, E., Desimone, M., et al. (2003). ARAMEMNON, a novel database for Arabidopsis integral membrane proteins. Plant Physiol. 131, 16–26. doi: 10.1104/pp.011577
Schwarzlander, M., Logan, D. C., Johnston, I. G., Jones, N. S., Meyer, A. J., Fricker, M. D., et al. (2012). Pulsing of membrane potential in individual mitochondria: a stress-induced mechanism to regulate respiratory bioenergetics in Arabidopsis. Plant Cell 24, 1188–1201. doi: 10.1105/tpc.112.096438
Shanmughapriya, S., Rajan, S., Hoffman, N. E., Higgins, A. M., Tomar, D., Nemani, N., et al. (2015). SPG7 is an essential and conserved component of the mitochondrial permeability transition pore. Mol. Cell. 60, 47–62. doi: 10.1016/j.molcel.2015.08.009
Shoshan-Barmatz, V., De Pinto, V., Zweckstetter, M., Raviv, Z., Keinan, N., and Arbel, N. (2010). VDAC, a multi-functional mitochondrial protein regulating cell life and death. Mol. Aspects Med. 31, 227–285. doi: 10.1016/j.mam.2010.03.002
Silva, M. A., Carnieri, E. G., and Vercesi, A. E. (1992). Calcium transport by corn mitochondria : evaluation of the role of phosphate. Plant Physiol. 98, 452–457. doi: 10.1104/pp.98.2.452
Smack, D. P., and Colombini, M. (1985). Voltage-dependent channels found in the membrane fraction of corn mitochondria. Plant Physiol. 79, 1094–1097. doi: 10.1104/pp.79.4.1094
Stael, S., Wurzinger, B., Mair, A., Mehlmer, N., Vothknecht, U. C., and Teige, M. (2012). Plant organellar calcium signalling: an emerging field. J. Exp. Bot. 63, 1525–1542. doi: 10.1093/jxb/err394
Szabo, I., and Zoratti, M. (2014). Mitochondrial channels: ion fluxes and more. Physiol. Rev. 94, 519–608. doi: 10.1152/physrev.00021.2013
Sze, H., Padmanaban, S., Cellier, F., Honys, D., Cheng, N. H., Bock, K. W., et al. (2004). Expression patterns of a novel AtCHX gene family highlight potential roles in osmotic adjustment and K+ homeostasis in pollen development. Plant Physiol. 136, 2532–2547. doi: 10.1104/pp.104.046003
Takahashi, Y., and Tateda, C. (2013). The functions of voltage-dependent anion channels in plants. Apoptosis 18, 917–924. doi: 10.1007/s10495-013-0845-3
Tapken, D., Anschutz, U., Liu, L. H., Huelsken, T., Seebohm, G., Becker, D., et al. (2013). A plant homolog of animal glutamate receptors is an ion channel gated by multiple hydrophobic amino acids. Sci. Signal. 6:ra47. doi: 10.1126/scisignal.2003762
Tapken, D., and Hollmann, M. (2008). Arabidopsis thaliana glutamate receptor ion channel function demonstrated by ion pore transplantation. J. Mol. Biol. 383, 36–48. doi: 10.1016/j.jmb.2008.06.076
Tateda, C., Watanabe, K., Kusano, T., and Takahashi, Y. (2011). Molecular and genetic characterization of the gene family encoding the voltage-dependent anion channel in Arabidopsis. J. Exp. Bot. 62, 4773–4785. doi: 10.1093/jxb/err113
Tateda, C., Yamashita, K., Takahashi, F., Kusano, T., and Takahashi, Y. (2009). Plant voltage-dependent anion channels are involved in host defense against Pseudomonas cichorii and in Bax-induced cell death. Plant Cell Rep. 28, 41–51. doi: 10.1007/s00299-008-0630-x
Tatsuta, T., Augustin, S., Nolden, M., Friedrichs, B., and Langer, T. (2007). m-AAA protease-driven membrane dislocation allows intramembrane cleavage by rhomboid in mitochondria. EMBO J. 26, 325–335. doi: 10.1038/sj.emboj.7601514
Teardo, E., Carraretto, L., De Bortoli, S., Costa, A., Behera, S., Wagner, R., et al. (2015). Alternative splicing-mediated targeting of the Arabidopsis GLUTAMATE RECEPTOR3.5 to mitochondria affects organelle morphology. Plant Physiol. 167, 216–227. doi: 10.1104/pp.114.242602
Teardo, E., Segalla, A., Formentin, E., Zanetti, M., Marin, O., Giacometti, G. M., et al. (2010). Characterization of a plant glutamate receptor activity. Cell. Physiol. Biochem. 26, 253–262. doi: 10.1159/000320525
Trono, D., Laus, M. N., Soccio, M., and Pastore, D. (2014). Transport pathways–proton motive force interrelationship in durum wheat mitochondria. Int. J. Mol. Sci. 15, 8186–8215. doi: 10.3390/ijms15058186
Trono, D., Laus, M. N., Soccio, M., Alfarano, M., and Pastore, D. (2015). Modulation of potassium channel activity in the balance of ROS and ATP production by durum wheat mitochondria—an amazing defense tool against hyperosmotic stress. Front. Plant Sci. 6:1072. doi: 10.3389/fpls.2015.01072
Tsai, M. F., Jiang, D., Zhao, L., Clapham, D., and Miller, C. (2014). Functional reconstitution of the mitochondrial Ca2+/H+ antiporter Letm1. J. Gen. Physiol. 143, 67–73. doi: 10.1085/jgp.201311096
Vais, H., Mallilankaraman, K., Mak, D. O., Hoff, H., Payne, R., Tanis, J. E., et al. (2016). EMRE Is a Matrix Ca2+ sensor that governs gatekeeping of the mitochondrial Ca2+ Uniporter. Cell Rep. 14, 403–410. doi: 10.1016/j.celrep.2015.12.054
Vais, H., Tanis, J. E., Muller, M., Payne, R., Mallilankaraman, K., and Foskett, J. K. (2015). MCUR1, CCDC90A, is a regulator of the mitochondrial calcium uniporter. Cell Metabol. 22, 533–535. doi: 10.1016/j.cmet.2015.09.015
Vercesi, A. E., Borecky, J., Maia Ide, G., Arruda, P., Cuccovia, I. M., and Chaimovich, H. (2006). Plant uncoupling mitochondrial proteins. Annu. Rev. Plant Biol. 57, 383–404. doi: 10.1146/annurev.arplant.57.032905.105335
Vianello, A., Casolo, V., Petrussa, E., Peresson, C., Patui, S., Bertolini, A., et al. (2012). The mitochondrial permeability transition pore (PTP) - an example of multiple molecular exaptation? Biochim. Biophys. Acta 1817, 2072–2086. doi: 10.1016/j.bbabio.2012.06.620
Vincill, E. D., Bieck, A. M., and Spalding, E. P. (2012). Ca2+ conduction by an amino acid-gated ion channel related to glutamate receptors. Plant Physiol. 159, 40–46. doi: 10.1104/pp.112.197509
Wagner, S., Behera, S., De Bortoli, S., Logan, D. C., Fuchs, P., Carraretto, L., et al. (2015). The EF-Hand Ca2+ binding protein MICU choreographs mitochondrial Ca2+ dynamics in Arabidopsis. Plant Cell doi: 10.1105/tpc.15.00509
Wu, Y., Rasmussen, T. P., Koval, O. M., Joiner, M. L., Hall, D. D., Chen, B., et al. (2015). The mitochondrial uniporter controls fight or flight heart rate increases. Nat. Commun. 6:6081. doi: 10.1038/ncomms7081
Xiong, T. C., Bourque, S., Lecourieux, D., Amelot, N., Grat, S., Briere, C., et al. (2006). Calcium signaling in plant cell organelles delimited by a double membrane. Biochim. Biophys. Acta 1763, 1209–1215. doi: 10.1016/j.bbamcr.2006.09.024
Zancani, M., Casolo, V., Petrussa, E., Peresson, C., Patui, S., Bertolini, A., et al. (2015). The permeability transition in plant mitochondria: the missing link. Front. Plant Sci. 6:1120. doi: 10.3389/fpls.2015.01120
Zhang, B., Carrie, C., Ivanova, A., Narsai, R., Murcha, M. W., Duncan, O., et al. (2012). LETM proteins play a role in the accumulation of mitochondrially encoded proteins in Arabidopsis thaliana and AtLETM2 displays parent of origin effects. J. Biol. Chem. 287, 41757–41773. doi: 10.1074/jbc.M112.383836
Zhang, M., Takano, T., Liu, S., and Zhang, X. (2015). Arabidopsis mitochondrial voltage-dependent anion channel 3 (AtVDAC3) protein interacts with thioredoxin m2. FEBS Lett. 589, 1207–1213. doi: 10.1016/j.febslet.2015.03.034
Zhao, Y., Pan, Z., Zhang, Y., Qu, X., Zhang, Y., Yang, Y., et al. (2013). The actin-related Protein2/3 complex regulates mitochondrial-associated calcium signaling during salt stress in Arabidopsis. Plant Cell 25, 4544–4559. doi: 10.1105/tpc.113.117887
Zoratti, M., and Szabo, I. (1995). The mitochondrial permeability transition. Biochim. Biophys. Acta 1241, 139–176. doi: 10.1016/0304-4157(95)00003-A
Keywords: higher plants, mitochondria, calcium channels and transporters, calcium homeostasis, physiological processes
Citation: Carraretto L, Checchetto V, De Bortoli S, Formentin E, Costa A, Szabo I and Teardo E (2016) Calcium Flux across Plant Mitochondrial Membranes: Possible Molecular Players. Front. Plant Sci. 7:354. doi: 10.3389/fpls.2016.00354
Received: 08 November 2015; Accepted: 07 March 2016;
Published: 31 March 2016.
Edited by:
Richard Sayre, New Mexico Consortium at Los Alamos National Labs, USAReviewed by:
Michael G. Palmgren, University of Copenhagen, DenmarkCopyright © 2016 Carraretto, Checchetto, De Bortoli, Formentin, Costa, Szabo and Teardo. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Enrico Teardo, ZW5yaWNvLnRlYXJkb0B1bmlwZC5pdA==; Ildiko Szabo, aWxkaUBjaXYuYmlvLnVuaXBkLml0
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.