 Giao N. Nguyen
Giao N. Nguyen Steven J. Rothstein
Steven J. Rothstein German Spangenberg
German Spangenberg Surya Kant
Surya Kant
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
REVIEW article
Front. Plant Sci. , 13 August 2015
Sec. Plant Nutrition
Volume 6 - 2015 | https://doi.org/10.3389/fpls.2015.00629
This article is part of the Research Topic MicroRNAs and their role in plant nutrient stress response. View all 6 articles
Plant microRNAs (miRNAs) are a class of small non-coding RNAs which target and regulate the expression of genes involved in several growth, development, and metabolism processes. Recent researches have shown involvement of miRNAs in the regulation of uptake and utilization of nitrogen (N) and phosphorus (P) and more importantly for plant adaptation to N and P limitation conditions by modifications in plant growth, phenology, and architecture and production of secondary metabolites. Developing strategies that allow for the higher efficiency of using both N and P fertilizers in crop production is important for economic and environmental benefits. Improved crop varieties with better adaptation to N and P limiting conditions could be a key approach to achieve this effectively. Furthermore, understanding on the interactions between N and P uptake and use and their regulation is important for the maintenance of nutrient homeostasis in plants. This review describes the possible functions of different miRNAs and their cross-talk relevant to the plant adaptive responses to N and P limiting conditions. In addition, a comprehensive understanding of these processes at molecular level and importance of biological adaptation for improved N and P use efficiency is discussed.
In the past half century, there has been a marked increase in food production allowing a significant decline in food shortages worldwide although the world population has doubled during this time (Godfray et al., 2010). However, in the next half century to achieve a similar expansion of food production to meet the needs of the increased human population is quite challenging given that with the confounding factors of narrowing arable land due to urbanization, a shortage of water for irrigation, global climate change, changing human diet, and significant increase in the proportion of food used for feeding animals and livestock or for producing biofuels (Rothstein, 2007). Cost effective approaches to increase crop production include but are not limited to the usage of modern high yielding crop varieties including genetically modified crops to increase productivity per unit of cultivated land, and the application of advanced agricultural practices such as minimal soil tillage and improvements in water and fertilizer use efficiency (Good et al., 2004). Improved fertilizer use efficiency requires balanced fertilization with adequate macro-nutrients and micro-nutrients (Baligar and Fageria, 2015).
Among macro-nutrients, both nitrogen (N) and phosphorus (P) are key elements for crop production and are major constraints for plant growth, development, and yield since intensive crop production relies heavily on a large input of these fertilizers (Vance et al., 2003; Sinha et al., 2015). Annually it requires approximately 85–90 million tons of N and 50 million tons of P fertilizer for crop production, worldwide (Good et al., 2004; López-Arredondo et al., 2014). However, crop plants are only able to exploit up to 40% of the applied N and P fertilizers, while the residual is lost to the environment through leaching, surface runoff, hypertrophication, denitrification, volatilization, and microbial consumption (Poirier and Bucher, 2002; Good and Beatty, 2011). Under the current trend, there will be about 2.5 fold increased level of eutrophication caused by excessive use of N and P fertilizer by 2050 (Poirier and Bucher, 2002). All these factors lead to a greater production cost and very significant levels of environmental pollution (Giles, 2005). However, it is pertinent to note that about 70% of arable land worldwide are deficient of Pi (Hinsinger, 2001; Kirkby and Johnston, 2008). Unlike N fertilizer which can easily be produced from the unlimited ambient N2, natural P resources such as phosphate rock, apatite used to manufacture P fertilizers are non-renewable and increasingly limited making it a major challenge for sustainable crop production in the future (López-Arredondo et al., 2014). It is estimated that an increase in nitrogen use efficiency by 1% worldwide, would save approximately $1.1 billion annually (Kant et al., 2011a). Therefore, it is of urgent importance to develop crop varieties with higher fertilizer use efficiency.
Plant small RNAs are short non-coding RNAs, which can be classified into two major groups based on their origin and biogenesis (Axtell, 2013). Small RNAs that are generated from perfect double-stranded RNA precursors are referred to as small interfering RNAs (siRNAs), which can be further divided into several subclasses such as heterochromatic siRNAs (hc-siRNA) and trans-acting siRNAs (ta-siRNAs) (Fei et al., 2013). Small RNAs that are processed from a partially double-stranded region of single-stranded RNA precursors are known as microRNAs (miRNAs) (Jones-Rhoades et al., 2006; Voinnet, 2009). Interestingly, miRNAs can trigger the production of secondary siRNAs such as ta-siRNAs (Voinnet, 2009; Fei et al., 2013). In this review, we focused on the involvement of miRNAs in the regulation of plant adaptation responses to nutrient deficiency. Readers who have interests on other classes of plant regulatory small RNAs, their biogenesis and modes of action are referred to other excellent reviews (Axtell, 2013; Fei et al., 2013; Patil et al., 2014; Weiberg et al., 2014; Kamthan et al., 2015) and the references cited therein.
MiRNAs have been identified as potent regulators of plant growth, development (Jung et al., 2009; Sun, 2012; Wu, 2013) and stress-responses including biotic and abiotic stresses (Phillips et al., 2007; Khraiwesh et al., 2012; Sunkar et al., 2012; Ferdous et al., 2015). In the last decade, miRNAs have also been implicated in nutrient uptake, transport and assimilation. Moreover, miRNAs were also identified as signaling molecules between cells, tissues, and organs (Chitwood and Timmermans, 2010; Meng et al., 2010). MiRNAs are short (19–24 nucleotides), single-stranded, non-coding RNAs and serve as post-transcriptional regulators of gene expression in plants (Jones-Rhoades et al., 2006). They are initially transcribed from MIR genes by RNA polymerase II to form primary-microRNAs (pri-miRNAs) with stem loop structures. A DICER-LIKE 1 protein (DCL1) processes these long pri-miRNAs at stem loop regions to form pre-miRNAs. RNA duplexes excised from pre-miRNAs are exported from the nucleus into cytoplasm (Rogers and Chen, 2013). Subsequently, one of the small RNA strands referred to as miRNA is stably incorporated into AGO1, the effector nuclease of the RNA-induced silencing complexes (RISCs). The other strand, known as miRNA* is rapidly degraded. MiRNAs can regulate gene expression by guiding AGO1 to cleave target mRNAs with complementary target sites or to interfere with protein translation (Jones-Rhoades et al., 2006; Voinnet, 2009; Kamthan et al., 2015).
There are a number of publications describing the expression profiles of individual miRNAs in response to nutrient deficiency, only a few attempts have been made to comprehensively cover the molecular mechanisms where miRNAs are important for the adaptive responses. This review shed light on (i) recent progress in understanding the mechanism of N and P acquisition, assimilation and mobilization in plant; (ii) elaborate on how plants respond to N and P deficiency and in what ways miRNAs contribute to this physiological adaptation; and (iii) discuss the involvement of plant miRNAs in the crosstalk between N and P under limiting conditions.
N is an essential element for plant development as it is a key component of other cellular constituents such as nucleic acids, proteins, chlorophyll, and phytohormones (Hawkesford et al., 2012). Plants take up N mainly in the forms of nitrate (NO−3), ammonium (NH+4) or urea from the soil, however, NO−3 is preferred form for most arable plants (Crawford and Forde, 2002). Synergistic association of legumes, actinorhizal plants and several C4 grasses with symbiotic microorganisms can produce NH+4 that fix atmospheric N2 via bacterial enzyme nitrogenase (Andrews et al., 2013). Two main phases of N uptake and usage during life cycle of plants have been well described in the literature. The first phase is during the vegetative stage where N is taken up, stored and assimilated into amino acids or other nitrogenous compounds. The second phase is the remobilization of assimilated N at senescence, where these compounds will be released and remobilized to reproductive organs to support the developing seeds (Kant et al., 2011a).
The N uptake process from the soil can be conducted either directly by roots or indirectly via mycorrhizal fungi (Andrews et al., 2013). Two types of N uptake systems are well defined in plants i.e., high affinity transport system (HATS) and low affinity transport system (LATS), which functions at low external N concentrations (1 μm–1 mM) and high external N concentrations (>1 mM), respectively (Kraiser et al., 2011). Studies on the molecular mechanism of N uptake and translocation revealed the involvement of a number of genes for these processes (Masclaux-Daubresse et al., 2010; Kant et al., 2011a; Xu et al., 2012; Krapp et al., 2014). Four members of nitrate transporter families involved in the NO−3 uptake process are described in Arabidopsis i.e., nitrate transporter 1/peptide (NPD), nitrate transporter 2 (NRT2), the chloride channel (CLC), and slow anion channel associated homologs (SLAC/SLAH) (Krapp et al., 2014). NPD is a recent nomenclature of nitrate transporter 1 (NRT1) family recently proposed by Léran et al. (2014) since NRT1 transporters has been reported to transport NO−3 and other substrates such as auxin, ABA, and glucosinolates. However, here the original gene names as these were initially named are referred. While NO−3 entering plant cells by facilitation of nitrate transporters, NH+4 from external source is taken up by ammonium transporters (AMT) (Crawford and Forde, 2002). Upon entering plant cells, NO−3 is converted to nitrite then to NH+4 and finally to amino acids through the action of nitrate reductase, nitrite reductase, glutamine synthetase, and glutamate synthase (Crawford and Forde, 2002). During reproductive growth, there are three pathways to release the nitrogenous compounds: the chloroplast degradation pathway; the vacuolar and autophagic pathway; and the ubiquintin-26S proteasome pathway (Liu et al., 2008). After degradation, the released amino acids will be loaded into phloem and remobilized to the developing seeds by facilitation of amino acid transporters completing the plant N cycle (Kant et al., 2011a).
Similar to N, P is also an essential macro-nutrient for plant growth and development. P plays a vital role as a key constituent of nucleic acids, phospholipids and the high energetic phosphate compounds ATP, ADP, or AMP (Marschner, 2012). Despite its central role in cellular processes, P availability in soil for plant uptake is very limited compared to other mineral elements (Ramaekers et al., 2010; Shen et al., 2011). Plants take up phosphate (Pi) in the form of PO−4 and PO−24. Unfortunately, a majority of P existing in soil is insoluble due to its adsorption, precipitation with other cations or conversion into organic forms by microbes. Therefore, to maintain P homeostasis, plants have evolved a number of adaptive responses including activation of Pi transporters, modifications of root architecture, secretion of phosphatases and organic acids and symbiosis with mycorrhizal fungi (Raghothama, 1999; Poirier and Bucher, 2002; López-Arredondo et al., 2014).
The initial transport of Pi from soil into roots requires a mechanism allowing Pi movement against an extremely high endogenous Pi concentration gradient in root cells, which is usually 1000–10,000 times higher than external Pi concentration in the soil (Bieleski, 1973). Plants have several Pi transporters for acquisition of Pi when external Pi availability is low (Rausch and Bucher, 2002; López-Arredondo et al., 2014). Pi transporter genes have been identified and cloned in many crop species and each of them plays a specific role to maintain P homeostasis (Lin et al., 2009). Four Pi transporter families have been identified in Arabidopsis; PHT1, PHT2, PHT3, and PHT4 (Poirier and Bucher, 2002; Guo et al., 2008). Pi is taken up from soil to root cells via mediation of members of the high affinity PHT1 family, which employs an H+-gradient at the plasma membrane to modulate H+/Pi symport activity (Shen et al., 2011). PHT2 family transporters are located in chroloplast and affects whole plant Pi allocation (Versaw and Harrison, 2002). PHT3 family members are located in mitochondrial inner membrane (Poirier and Bucher, 2002) and PHT4 family transporters are located in chloroplasts, non-photosynthetic plastids, and the Golgi apparatus (Guo et al., 2008). In addition, genetic screens also identified several key genes involved in Pi acquisition and translocation in Arabidopsis. Among these are PHOSPHATE1 (PHO1) involved in Pi loading in xylem vessels in roots (Hamburger et al., 2002) and PHOSPHATE2 (PHO2) a negative regulator of Pi uptake (Aung et al., 2006; Bari et al., 2006). These Pi transporters, PHO1 and PHO2 work in coordination for the acquisition and translocation of Pi in plants.
Under N limiting conditions, miRNAs can be up- or down-regulated. Expression profiles of different miRNA families have been observed in various crop species such as maize (Xu et al., 2011; Trevisan et al., 2012; Zhao et al., 2013), rice (Cai et al., 2012; Yan et al., 2014), soybean (Wang et al., 2013), and Arabidopsis (Pant et al., 2009; Liang et al., 2012). Alteration in the expression pattern of these miRNAs results in plant adaptive responses to N limitation in the soil via mediation of the expression of their target genes (Zeng et al., 2014). Involvement of different miRNAs under a N limitation response is summarized in Table 1. Changes of the expression patterns of miRNAs have been shown to play crucial roles in modulating adaptive responses. These adaptations include enhanced N uptake and transport, changes in plant architecture, production of metabolites and radical scavengers, reduced growth and early flowering and modulation of metabolism (Fischer et al., 2013; Zeng et al., 2014; Sinha et al., 2015), as discussed in the following sub-sections.
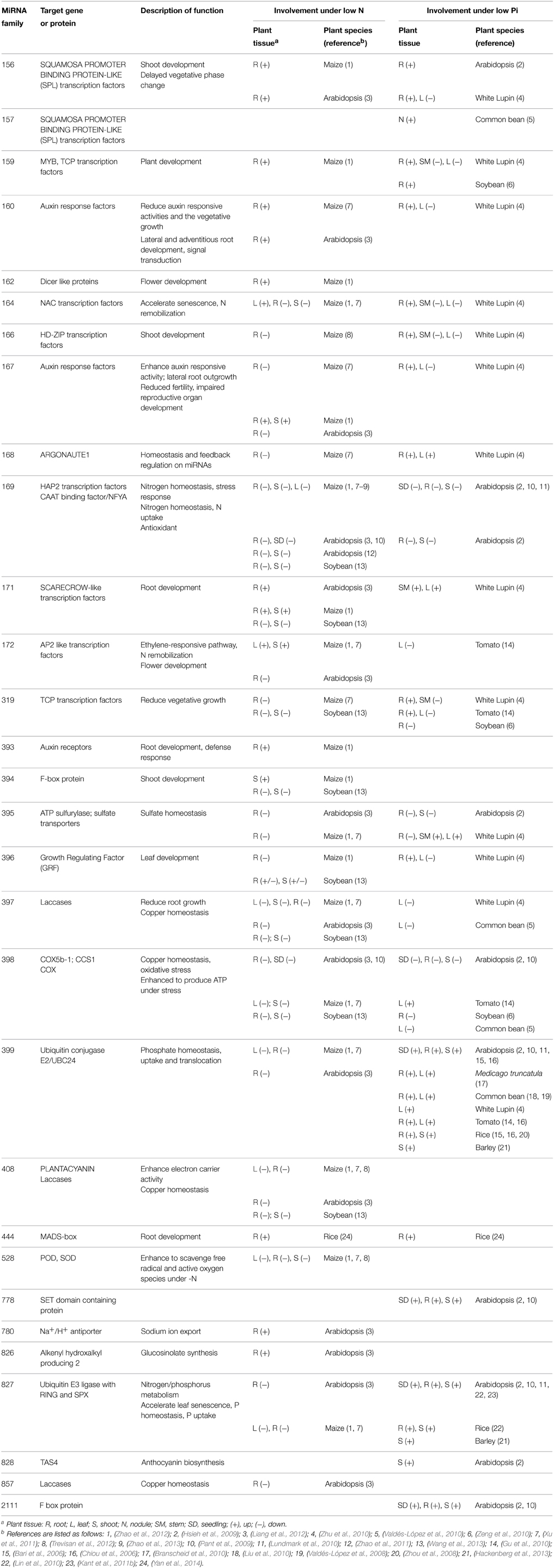
Table 1. Different miRNAs involved in N and P limitation with their target genes, functions, and tissue specific effects.
Several miRNAs have been reported to play vital roles in root growth and development under N deficiency (Xu et al., 2011; Liang et al., 2012). N deficiency modifies root architecture and morphology to improve the plant's ability to acquire nutrients from the soil more efficiently (Hermans et al., 2006). Recent studies have demonstrated the regulatory roles of miRNAs on root architecture and growth under N deficient conditions (Khan et al., 2011; Liang et al., 2012; Zhao et al., 2012; Wang et al., 2013). In Arabidopsis, two Auxin Response Factors (ARF transcription factors), ARF6 and ARF8, regulate development of reproductive organs and are targets of miR167 (Wu et al., 2006). Further it was shown that ARF8 is the regulator of lateral roots, where its expression was induced in pericycle and lateral root cap cells under N limiting conditions (Gifford et al., 2008). ARF proteins bind to auxin responsive cis-acting promoter elements and can induce or suppress gene expression in response to the plant phytohormone auxin (Hagen and Guilfoyle, 2002; Liscum and Reed, 2002). MiR167 also reportedly targets IAA-Ala Resistant3 (IAR3), whose protein hydrolyses the inactive auxin derivative indole-3-acetic acid alanine and releases bioactive auxin, for root development during high osmotic stress (Kinoshita et al., 2012). Over-expression of miR167 resulted in plant morphologies similar to arf6 and arf8 mutant phenotypes (Wu et al., 2006). Therefore, lower expression of miR167 under N deficiency might lift its inhibition on auxin transcription factors which could in turn induce lateral root growth (Liang et al., 2012). In contrary, miR160 was induced while ARF16 and ARF17 were down-regulated under N limitation (Liang et al., 2012). In Arabidopsis, ARF16 modulates root cap cell formation while ARF17 serves as a regulator of GH3-like early auxin response genes (Mallory et al., 2005; Wang et al., 2005). Studies showed that miR160 over-expressed transgenic plants had more developed lateral roots implying that induced expression of miR160 might promote lateral root growth via mediation of ARF16 and ARF17 under N deficiency (Liang et al., 2012). Further study demonstrated that modulation of the root system under N starvation was actually coordinated by a spatial regulatory complex of three miRNAs: miR160, miR167, and miR171 (Liang et al., 2012). Under N deficiency, root growth was increased by enhanced expression of miR160 and miR171 and reduced expression of miR167.
The NAC gene family encode transcription factors that play multiple roles in developmental processes in plants. NACs consist of three gene families; NAM (No Apical Meristem), ATAF (Arabidopsis Transcription Activation Factor), CUC (CUp shaped Cotyledon) (Olsen et al., 2005). MiR164 was reported to target five NAC domain containing genes, including NAC1 which is involved in auxin signal transduction for the growth and development of lateral roots (Guo et al., 2005) and CUC1 which is required for normal embryonic, vegetative, and floral development (Mallory et al., 2004). Down-regulation of miR164 concomitant with the up-regulation of NAC1 produced more lateral roots (Guo et al., 2005). A NAC locus has been reported to accelerate senescence and increase nutrient remobilization from leaves to the developing grains in wheat (Uauy et al., 2006). MiR164 is up-regulated in maize leaf under N limiting conditions (Xu et al., 2011). This might imply the role of miR164 in modulating both root and shoot development under N limitation adaptation.
MiRNA, miR169a is the only candidate reported so far, regulating the expression of key target N transporters under N limiting conditions. The MiR169 family in Arabidopsis consists of 14 members, among these miR169a is the main contributor to the total miR169 level (Zhao et al., 2011). In Arabidopsis, miR169 targets NFY (Nuclear Factor Y) a ubiquitous transcription factor consisting of 3 subunits A, B, and C, some of which bind to promoter regions and regulate expression of the nitrate transporters AtNRT2.1 and AtNRT1.1 (Zhao et al., 2011). Up-regulation of NFYA5, a target of miR169, reportedly enhanced drought tolerance by stimulating expression of several antioxidant genes (Li et al., 2008). Under N deficiency, miR169 was strongly down-regulated and its target NFYA family members were strongly induced in root and shoot tissues (Zhao et al., 2011). Furthermore, over-expression of MIR169a repressed expression of NFYA transcripts. These over-expresser transgenic plants were especially hypersensitive to N starvation, accumulating less N which resulted in leaf yellowing compared to the wild type (WT) plants (Zhao et al., 2011). The hypersensitivity of MIR169a over-expresser plants was associated with down-regulation of nitrate transporter genes AtNRT2.1 and AtNRT1.1 (CHL1) suggesting the regulatory role of miR169 in N uptake and remobilization. The chl1 mutant plants showed a reduced expression of AtNRT1.1 and its phenotype mimicked over-expressed MIR169a transgenic plants. These indicate that lower expression of miR169 is an adaptive response of plants under N limiting conditions.
The role of miRNAs involved in the production of antioxidants and anthocyanins has been reported (Kandlbinder et al., 2004; Shin et al., 2005; Liang et al., 2012). These are secondary metabolites protecting plants from photo-inhibition damage under abiotic stresses including N limitation. In Arabidopsis, the AOP2 gene which encodes 2-oxoglutarate-dependent dioxygenase and associates with glucosinolate biosynthesis is the target of miR826 (Liang et al., 2012). A recent study reported that under N limiting conditions miR826 was strongly induced while AOP2 transcripts were significantly repressed (He et al., 2014). In addition, expression of the nitrate transporter (NRT2.1) and ammonium transporter (AMT1.5) genes were also induced (He et al., 2014). MiR826 over-expresser Arabidopsis transgenic plants showed enhanced tolerance under N limiting conditions, and had higher biomass, more primary and lateral roots, increased chlorophyll and less glucosinolate and anthocyanin contents (He et al., 2014). It can be hypothesized that these transgenic lines were able to withstand N limiting conditions better than WT plants and had less need to accumulate stress induced secondary metabolites such as glucosinolates and anthocyanin.
N deficiency was reported to repress expression of miR398 in plants (Pant et al., 2009; Liang et al., 2012). MiR398 is a conserved miRNA in Arabidopsis, rice, Lotus, and Medicago (Sunkar and Zhu, 2004). This miRNA targets transcripts of multiple genes: cytosolic CSD1, chloroplastic CSD2, COX5b-1, and CCS1. CSD1 and CSD2 encode a Cu/Zn superoxide dismutase (SOD) an important radical scavenger that protects plants from oxidative stress damage (Sunkar et al., 2006; Jagadeeswaran et al., 2009). COX5b-1 encodes a subunit of the mitochondrial cytochrome c oxidase and CCS1 encodes the copper chaperone for SOD (Beauclair et al., 2010; Zhu et al., 2011). Over-expression of CSD2 was reported to confer tolerance to oxidative stress induced by high light (Sunkar et al., 2006). This suggests that down-regulation of miR398 might reduce its control on these target antioxidant genes and thus indirectly provided protection to the photosynthetic machinery from reactive oxygen species (ROS) generated from N deficiency (Kandlbinder et al., 2004; Shin et al., 2005).
In plants, miR156 targets transcripts of the Squamosa Promoter Binding Protein Like (SPL) family of transcription factors whose expressions were synergistically associated with anthocyanin biosynthesis (Gou et al., 2011). Over-expression of miR156 repressed the expression of SLP, concomitantly with an increased production of anthocyanin in Arabidopsis. The accumulation of anthocyanin and reduction of photosynthesis are adaptive responses of plants to N limiting condition, which protect them from photo-inhibition damage (Diaz et al., 2006; Peng et al., 2008). Thus, increased expression of miR156 might have resulted in higher levels of anthocyanin production conferring better protection of plants during N starvation (Liang et al., 2012).
MicroRNAs have been long known for controlling flowering time in plants (Yamaguchi and Abe, 2012; Spanudakis and Jackson, 2014). MiR156 was shown to regulate flowering, vegetative phase changes, fertility, and leaf formation via mediation of the SPL genes (Wu and Poethig, 2006; Wang et al., 2008, 2009; Wu et al., 2009; Xing et al., 2010). Transgenic plants over-expressing miR156 resulted in a prolonged juvenile phase, stunted growth, altered biomass production, and delayed flowering (Wu and Poethig, 2006; Xie et al., 2006; Chuck et al., 2007a; Zhang et al., 2011b; Fu et al., 2012; Shikata et al., 2012). MiR172 targets the AP2-like family of transcription factors including TOE1 and TOE2 and controls flowering time and floral organ identity in maize and Arabidopsis (Aukerman and Sakai, 2003; Chen, 2004; Chuck et al., 2007b; Zhao et al., 2007). In contrast to miR156, miR172 over-expressers were shown to promote flowering in Arabidopsis (Aukerman and Sakai, 2003; Chen, 2004; Jung et al., 2011). Further research revealed that miR156 regulates expression of miR172 via mediation of the transcription factors SPL9 and SPL10 (Wu et al., 2009). Over-expressed 35S::miR156a transgenic plants had only half of the normal transcript level of miR172, whereas 35S::MIM156 transgenic plants had more than double the miR172 level (Wu et al., 2009). Since N starvation is known to induce early flowering in plants (Vidal et al., 2014), the changes in the expression pattern of miR156 and miR172 under N deficiency (Liang et al., 2012) will likely result in modification of flowering time.
Differential expression patterns of miRNAs under Pi limitation have been observed in several plant species for example Arabidopsis (Hsieh et al., 2009; Lundmark et al., 2010), common bean (Valdés-López et al., 2010), soybean (Zeng et al., 2010), rice (Zhou et al., 2008; Lin et al., 2010; Yan et al., 2014), barley (Hackenberg et al., 2013), white Lupin (Zhu et al., 2010), tomato (Gu et al., 2010), and Medicago truncatula (Branscheid et al., 2010). Similar to their behavior in N limiting condition, changes in the level of miRNAs mediated the expression of target genes resulting in physiological and morphological changes of plants under low Pi conditions. These changes include modifications of root architecture, production of metabolites and biosynthesis of anthocyanin and oxidative radical scavengers. Involvement of different miRNAs under Pi deficiency is shown in Table 1.
MiRNAs are known to regulate root architecture (Guo et al., 2005; Mallory et al., 2005). Their differential expression patterns under Pi limiting conditions (Zhu et al., 2010) suggest their regulatory role in modulating root growth. Pi starvation induces changes in plant root system architecture as adaptive responses such as minimizing development of primary roots, increasing root branching, and stimulating elongation of lateral roots (Péret et al., 2014). Expression of miR160, miR164, and miR167 were up-regulated specifically in roots and were down-regulated in stem and leaves under Pi starvation in white Lupin (Zhu et al., 2010). Studies on Pi deficient white lupin reported that plants produced more lateral roots (Johnson et al., 1996), which enable the plant to secrete more organic acids and thus to facilitate liberation of precipitated Pi from the soil (Massonneau et al., 2001). Since miR160, miR164, and miR167 modulate root growth via mediation of NAC and auxin transcription factors (Guo et al., 2005; Mallory et al., 2005) it is arguable that their induced expression locally in the roots might be associated with lateral root development in response to Pi deficiency.
A number of miRNAs, their target genes and involvement in Pi uptake, relocation, and remobilization has been well characterized. As discussed in the earlier section, reduced expression of miR169 facilitated N uptake, and increased expression of the nitrate transporter genes AtNRT2.1 and AtNRT1.1 via mediation of NFYA transcription factors. A recent study showed that increased supply of NO−3 also stimulated root formation thus enhancing Pi uptake in the shrub legume (Maistry et al., 2014). It seems likely that the down-regulation of miR169 under a low Pi condition (Hsieh et al., 2009; Pant et al., 2009) was an adaptive response to facilitate N uptake and remobilization, which in turn indirectly stimulates Pi uptake.
The role of miR399 in regulation of P homeostasis via Pi acquisition and translocation is well characterized (Chiou, 2007; Liu et al., 2014). MiR399 targets PHO2 (or UBC24) gene which encodes a ubiquitin conjugating E2 enzyme (Aung et al., 2006; Bari et al., 2006). PHO2 plays a role to negatively regulate the level of Pi uptake, translocation, and remobilization when Pi supply is sufficient. pho2 mutant plants accumulate excessive amounts of Pi in shoots and thus exhibit Pi induced toxic symptoms in Arabidopsis (Delhaize and Randall, 1995; Aung et al., 2006). In agreement, miR399 over-expresser plants enhance Pi uptake and translocation in shoots and under Pi sufficient conditions causing Pi toxicity in plants (Fujii et al., 2005; Aung et al., 2006; Bari et al., 2006; Chiou et al., 2006). MiR399 was up-regulated and PHO2 was down-regulated under low Pi conditions (Chiou et al., 2006) suggesting the role of miR399 in maintaining Pi homeostasis in plants. Recent findings showed that PHO2 is located in the endomembrane system and mediates the degradation of PHT1 family members and PHO1 (Liu et al., 2012; Huang et al., 2013). As remobilization of Pi from old to young leaves was inhibited under Pi starvation (Chiou et al., 2006), induced miR399 expression helped maintain Pi homeostasis via enhancing Pi uptake, transport, and remobilization from root to shoot by down-regulating its target gene PHO2 (Kuo and Chiou, 2011). Other studies on plants such as common bean (Valdés-López et al., 2008; Liu et al., 2010) and rice (Hu et al., 2011) also reported similar interactions between miR399 and PHO2 homologs in modulating Pi homeostasis, suggesting the regulatory roles of miR399 are conserved in angiosperms.
Similar to miR399, miR827 also plays a crucial role in maintaining Pi homeostasis in plants (Kant et al., 2011b; Lin et al., 2013; Liu et al., 2014; Park et al., 2014). MiR827 target the 5′-untranslated region of the NITROGEN LIMITATION ADAPTATION (NLA) transcripts (Kant et al., 2011b), which encode a RING type ubiquitin E3 ligase (Peng et al., 2007). The expression of miR827 is induced under a low Pi condition, where the transcript level of NLA was repressed (Kant et al., 2011b). Initially, NLA was reported to be involved in the adaptive response to N deficiency, where nla mutant plants showed earlier onset of senescence compared to WT plants under N limiting conditions and an inability to accumulate anthocyanins (Peng et al., 2007). However, later studies showed that a mutation in either PHOSPHATE TRANSPORTER TRAFFIC FACILITATOR1 (PHF1) or PHOSPHATE TRANSPORTER1:1 (PHT1:1) genes rescued the early senescence phenotype in nla mutant in response to N starvation. The early senescence phenotype of the nla mutant indeed was caused by excessive Pi accumulation similar to pho2 mutant plants (Kant et al., 2011b). Pi over-accumulation was much more pronounced under low NO−3 conditions in both nla and pho2 mutant plants suggesting the important role of NLA and PHO2 in maintaining Pi homeostasis in a NO−3 dependent manner (Kant et al., 2011b). Further research showed that NLA is predominantly expressed in the plasma membrane. NLA; an E3 ubiquitin ligase interacts with PHO2; an E2 ubiquitin conjugase to degrade Pi transporter PHT1:4 by ubiquitination (Park et al., 2014). Up-regulation of miR399 and miR827 which are negative regulators of PHO2 and NLA, respectively, would therefore enhance Pi uptake in plants under Pi limiting conditions (Kant et al., 2011b).
Pi deficiency induce accumulation of anthocyanins in plants and several miRNAs are known to target genes involved in the anthocyanin biosynthesis pathway. MiR828 is up-regulated under Pi deficiency and its target is Trans-Acting SiRNA gene 4 (TAS4) non-coding RNA transcript, which results in the production of tasi-RNAs (Hsieh et al., 2009). TAS4-siR81 (−) one of the dominant TAS4 siRNAs, targets a set of MYB transcription factors PAP1/MYB75, PAP2/MYB90, and MYB113, which regulate genes in the anthocyanin biosynthesis pathway and leaf senescence (Rajagopalan et al., 2006). MiR828 also target MYB113, suggesting an inter-relationship between these MYB genes, miR828 and TAS4. Since anthocyanin biosynthesis is also induced under Pi starvation (Misson et al., 2005), it is assumed that there is a mechanistic cross-talk between miR828, TAS4-siR81 (−) and its target genes, which modulates anthocyanin accumulation. Pi starvation might activate expression of these MYB transcription factors leading to enhanced biosynthesis of anthocyanins. In addition, increased levels of PAP1/MYB75 trigger the production of TAS4-siR81 (−) via the activation of miR828 or TAS4, which in turn suppressed the expression of MYB transcription factors (Hsieh et al., 2009).
Oxidative stress conditions are a common phenomenon in plants under nutrient deficiency in general and under Pi limitation in particular (Kandlbinder et al., 2004; Shin et al., 2005). To cope with deleterious oxidative stress conditions, plants usually develop a protective mechanism by activating production of oxidant scavengers in order to maintain redox homeostasis. Under Pi starvation, two miRNAs, miR169 and miR398, were reportedly down-regulated (Hsieh et al., 2009; Pant et al., 2009) suggesting that their reduced expression might involve plant tolerance to oxidative stress conditions. Up-regulation of NFYA5, a target of miR169, reportedly enhanced drought tolerance where NFYA5 over-expresser transgenic Arabidopsis had increased expression of several antioxidant enzymes such as glutathione S-transferase and peroxidases (Li et al., 2008). Most likely, down-regulation of miR169 allows for coping with the oxidative condition generated from Pi limiting conditions. In Arabidopsis miR398 targets two genes CSD1 and CSD2, which encode Cu/Zn SOD antioxidant scavengers (Sunkar et al., 2006). Down-regulation of miR398 by oxidative stresses was reported to promote post-transcriptional mRNA accumulation of CSD1 and CSD2 and enhanced oxidative stress tolerance (Sunkar et al., 2006). Thus, down-regulation of miR398 could potentially increase expression of the antioxidant genes CSD1 and CSD2 enhancing plant tolerance to oxidative stress conditions caused by Pi starvation.
Amongst miRNAs identified so far, perhaps miR399 and miR827 are best characterized with their crucial roles in maintaining Pi homeostasis in plants (Liu et al., 2014). However, differential expression patterns of these miRNAs under N (Xu et al., 2011; Liang et al., 2012; Zhao et al., 2012) and P (Hsieh et al., 2009; Pant et al., 2009; Lundmark et al., 2010) limiting conditions suggest that their regulatory roles also depend upon the interaction between N and P. An adequate and balanced supply of nutrients is important to meet plant nutritional requirement, and efforts have been made to identify interactions amongst the response to different nutrients in general and between N and P in particular. It is clear that N and P uptake and assimilation in plants are not independent processes, but they interact with each other, in which the supply of one affects the other (Fageria, 2001). Increasing Pi supply was reported to increase nodulation and N fixation of subterranean clover by mediation of host plant growth (Robson et al., 1981). A recent study also shows that increasing N supply stimulates root formation thereby enhancing Pi acquisition in shrub legume (Maistry et al., 2014).
The form of N present can also have an effect on the type of interaction between N and Pi. For example, a competing interaction for uptake has been reported between NO−3 and Pi, both forms being anionic, while no antagonism for uptake was found between NH+4 and Pi (Kant et al., 2011b). Further, the suppression by high levels of NO−3 on Pi uptake was higher compared to the suppression by high levels of Pi on NO−3 (Kant et al., 2011b). The differential suppression by these ions could be due to their altered mobility in soil, since NO−3 diffusion is 3–4 times faster than that seen for Pi (Tinker and Nye, 2000). Higher NH+4 supply was reported to increase Pi uptake in some crop species (Riley and Barber, 1971; Gahoonia et al., 1992), while higher application of NO−3 was found to suppress Pi uptake (Kant et al., 2011b). The synergistic effects of NH+4 toward Pi uptake could be attributed to its ability to create changes in rhizosphere pH and alter root development, where supply of NH+4 reduces pH in rhizosphere therefore enabling the availability of externally soluble Pi; whereas supply of NO−3 increases pH in the surrounding areas of the roots affecting Pi uptake (Hinsinger, 2001; Jin et al., 2014). Rice plants supplemented with NH+4 were shown to have higher Pi content in both roots and shoots than those supplied with NO−3 (Zeng et al., 2012). Furthermore, Pi uptake in plants requires involvement of H+-ATPase proton pump and co-transporters (Raghothama, 1999; Poirier and Bucher, 2002). Involvement of root plasma membrane H+-ATPase in the adaptation of plants in response to Pi deficiency has been reported (Yan et al., 2002; Shen et al., 2006). Transgenic Arabidopsis plants overexpressing plasma membrane H+-ATPase absorbed more Pi under low Pi conditions compared to the WT plants (Shen et al., 2006). It was hypothesized that Pi deficiency contributed to the enhanced activity of plasma membrane H+-ATPase and H+ pump leading to acidification the rhizosphere (Zhang et al., 2011a) which in turn makes external Pi available for plants. Recent studies on effects of N on Pi acquisition reported that there is an association of involvement of plasma membrane H+-ATPase in stimulating Pi uptake by addition of NH+4 fertilizer.
Plants share common adaptation responses under N or Pi limiting conditions such as activation of high affinity transporters, development of lateral roots to facilitate uptake process, remobilization of nutrients from older leaves to young leaves and reproductive parts, retardation of growth and photosynthesis and production of antioxidant scavengers (Fang et al., 2009; Kant et al., 2011a). Under Pi limitation, NO−3 uptake and translocation from roots to shoots was reduced in different plant species such as tobacco (Nicotinana tabacum L.) (Rufty et al., 1990), barley (Hordeum vulgare L.) (Rufty et al., 1991), and soybean (Glycine max L.) (Rufty et al., 1993). Pi starvation also reduced the uptake of NH+4 (Taylor et al., 2010). It was proposed that the decrease in NO−3 uptake was resulted from the decrease of ATP pool (Rufty et al., 1993) and the feedback control mechanisms where uptake of a respective nutrient element not only depends on its presence but also on the availability of all other nutrients in rhizosphere (Amtmann and Blatt, 2009). Furthermore, Pi deficiency resulted in reduced activity of nitrate reductase in the roots of bean (Phaseolus vulgaris L.) (Gniazdowska and Rychter, 2000), an important enzyme of N assimilation pathway, which in turn inhibited N uptake from external source. Moreover, de Groot et al. (2003) suggested that reduced N uptake in tomato (Lycopersicon esculentum Mill.) under Pi limitation could probably be mediated by leaf cytokinin concentrations since low cytokinin levels might cause decrease in nitrate reductase activity. Transcriptional profiling of Arabiopsis and maize shed more light on this aspect. Low Pi supply was shown to reduce expression of nitrate reductase genes in Arabidopsis (Wu et al., 2003) and in maize (Schlüter et al., 2013). Therefore, it is reasonable to mention that genes in N and P metabolisms have orchestrated to maintain the nutrient balance under Pi limiting condition. As discussed in previous section miR399 and miR827 play important roles in maintaining Pi homeostasis in plants. Under Pi limitation, these miRNAs are up-regulated to release their inhibition on PHO2 and NLA. As a result, induced expression of PHT1 transporters will increase the uptake and translocation of Pi from roots to shoots (Liu et al., 2014). In contrast, under low NO−3 supply Pi accumulation was higher in Arabidopsis plants (Kant et al., 2011b; Krapp et al., 2011) and maize leaves (Schlüter et al., 2012), which might be resulted from feedback control responses (Amtmann and Blatt, 2009). It is well documented that under Pi deprivation remobilization of soluble carbohydrates from leaves to roots increased to facilitate transporter activity and Pi uptake (Hermans et al., 2006; Karthikeyan et al., 2007). It is likely that increased root growth and high level of soluble carbohydrate accumulation in the roots under N limitation stimulate Pi accumulation in plants (Paul and Stitt, 1993). Furthermore, low NO−3 supply down-regulates many Pi starvation responsive genes (Schlüter et al., 2012, 2013). Probably down-regulation of Pi starvation responsive genes in these conditions is to alleviate unnecessary Pi uptake and accumulation in plants. Nevertheless, this mechanism requires further research. Under N limitation, expression of miR399 and miR827 are significantly down-regulated to stimulate expression of PHO2 and NLA, which are negatively correlated with PHT1 activity and Pi uptake. Since low NO−3 supply causes Pi accumulation in different plants (Kant et al., 2011b; Krapp et al., 2011; Schlüter et al., 2012), down-regulation of miR399 and miR827 under such conditions is possibly a counter measure to control the over-accumulation of Pi to the toxic level.
A hypothetical model for the crosstalk between N and P with emphasis on the role of miR399 and miR827 and their target genes PHO2 and NLA is presented in Figure 1, which is based on the report of Kant et al. (2011b). They studied a range of combinations of N and P applications and has described under which combinations pho2 and nla mutants were showing Pi toxicity. There was a strong effect of interaction between N and P supply on both pho2 and nla phenotypes. When supply of N and Pi is sufficient, both WT and pho2 and nla mutant plants show a normal phenotype (Figure 1A_Sufficient N and Pi). Pi toxicity effect was not observed in pho2 and nla mutant plants although Pi concentration was two-fold higher in these plants. Probably Pi level might not reached the critical toxic point (Figure 1B_Sufficient N and Pi) (Delhaize and Randall, 1995; Kant et al., 2011b). Under low Pi supply, miR399 and miR827 expressions are induced with concomitant repression of PHO2 and NLA genes though these genes are still functional in WT plants imposing some repression on Pi uptake. Lower Pi supply and uptake result in anthocyanin accumulation in WT plants, a known phenomenon (Figure 1A_Low Pi) (Jiang et al., 2007; Kant et al., 2011b). Interestingly, in pho2 or nla mutant plants PHO2 and NLA genes are non-functional thereby letting more Pi uptake compared to WT plants resulting in normal growth of these plants (Figure 1B_Low Pi). Under low N supply, WT plants accumulate anthocyanin; a known phenomenon (Figure 1A_Low N) (Peng et al., 2007, 2008; Kant et al., 2011b). Expressions of miR399 and miR827 was reduced with enhanced expression of PHO2 and NLA genes regulating the controlled Pi uptake (Figure 1A_Low N). Since these genes are non-functional in pho2 or nla mutant plants, Pi uptake is not regulated resulting in excessive accumulation of Pi causing Pi toxicity in nla or pho2 mutant plants (Figure 1B_Low N) (Delhaize and Randall, 1995; Kant et al., 2011b).
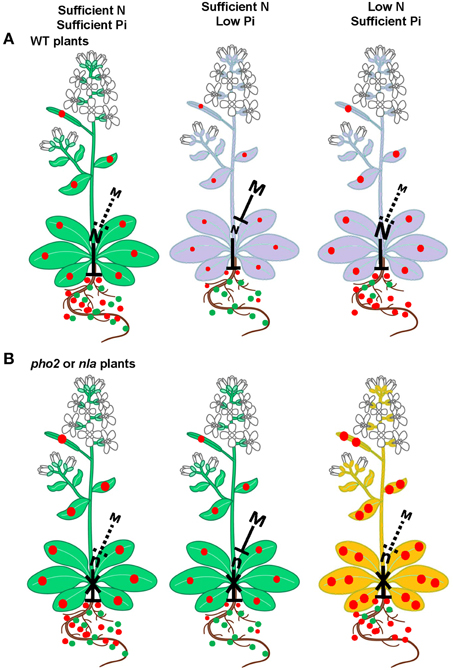
Figure 1. Hypothetical model for the role of miR399 and miR827 to maintain Pi homeostasis in plants. (A) In WT plants, PHO2 and NLA act as repressors for Pi uptake and these genes are targets of miR399 and miR827, respectively. Under sufficient N and Pi supply plants grow normal and under low N or Pi supply plants accumulate anthocyanin. (B) In pho2 or nla mutants, the PHO2 or NLA gene are not functional and these plants have higher Pi uptake in general compared to WT plants. Under sufficient N and Pi supply, mutant plants grow normal like WT. Under sufficient N and low Pi supply, since PHO2 or NLA gene are non-functional thereby letting more Pi uptake resulting in normal plant growth of these plants. Low N and sufficient Pi conditions accelerate excessive Pi accumulation causing Pi toxicity in these mutants. Purple leaves show N or Pi deficiency leading to anthocyanin accumulation, orange leaves indicate Pi toxicity. Dotted lines indicate less suppression. The size of the letter M or N correlates with expression level. The size and number of the solid green and red circles correlates with N and Pi concentration, respectively. M, miR399 or miR827; N, PHO2 or NLA (adapted from Kant et al., 2011b).
It is important for both economic and environmental reasons to improve N and P use efficiency in plants. Improving N and P uptake under their limiting supply would be a viable approach to utilize these nutrient elements more efficiently. The strategies for such improvement could include optimizing agricultural practices, molecular marker assisted breeding, and genetic engineering of genes involved in N and Pi uptake and metabolism (Ramaekers et al., 2010; McAllister et al., 2012; Guevara et al., 2014; López-Arredondo et al., 2014; Dass et al., 2015). A genetic engineering approach by manipulation of miRNA expression to improve N use efficiency was proposed (Fischer et al., 2013; Sinha et al., 2015), a similar approach is quite reasonable to propose for the improvement of P use efficiency.
Since the cloning of first plant miRNA in 2002 (Reinhart et al., 2002), a number of miRNAs have been identified. In the past five years, the number of newly cloned plant miRNAs has increased from 10,898 to 15,041 and target transcripts have been extended to 178,138 in 46 species (Yi et al., 2015). Several genetically engineered plants using different miRNAs have shown improved resistance against biotic and abiotic stresses (Kamthan et al., 2015). Still the understanding of the functionality of several miRNAs is unclear or their target genes have unknown function. Only a few of the miRNAs and their target genes have been completely characterized and experimentally validated especially those involved in adaptive response to N and Pi limitation conditions. Further research and studies are required and would be much helpful to decipher the regulatory roles of known miRNAs and their target gene functions involved in plant growth and development in general and improving efficient utilization of key macro-nutrient elements in particular.
To understand the role of miRNAs and involvement in crosstalk between N and P in plant is quite important. However, this interactive mechanism remains elusive and requires further investigations. The uptake and utilization processes of N and P in plants are complex and cannot be considered in isolation. Instead these processes are synergistic and depend on the nutritional requirements of the plants in a particular environment. Furthermore, levels of N and P cause affect not only the root and associated transport systems in plants but also the ion balance of other macro- (K, Ca, Mg, and S) and micro-nutrients (Mn, Fe, Zn, and Cu, etc.) (Schlüter et al., 2013).
Current knowledge of the involvement of various miRNAs for the regulation of N and P uptake, assimilation and utilization and plant adaptation to both N and P limitation conditions has been reviewed here along with how these processes affects the modifications in shoot and root growth, development, and architecture, effects on vegetative and reproductive phase and production of secondary metabolites. Nevertheless, a holistic approach to study the interaction of N and P along with other macro- and micro-nutrient elements and the involvement of miRNAs would be of much benefit and would thus require further research.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
We thank the reviewers for their constructive comments and suggestions to improve the manuscript and Dr Lancelot Maphosa for critical reading of the draft manuscript.
Amtmann, A., and Blatt, M. R. (2009). Regulation of macronutrient transport. New Phytol. 181, 35–52. doi: 10.1111/j.1469-8137.2008.02666.x
Andrews, M., Raven, J. A., and Lea, P. J. (2013). Do plants need nitrate? The mechanisms by which nitrogen form affects plants. Ann. Appl. Biol. 163, 174–199. doi: 10.1111/aab.12045
Aukerman, M. J., and Sakai, H. (2003). Regulation of flowering time and floral organ identity by a microRNA and its APETALA2-Like target genes. Plant Cell 15, 2730–2741. doi: 10.1105/tpc.016238
Aung, K., Lin, S. I., Wu, C. C., Huang, Y. T., Su, C. L., and Chiou, T. J. (2006). pho2, a phosphate overaccumulator, is caused by a nonsense mutation in a microRNA399 target gene. Plant Physiol. 141, 1000–1011. doi: 10.1104/pp.106.078063
Axtell, M. J. (2013). Classification and comparison of small RNAs from plants. Ann. Rev. Plant Biol. 64, 137–159. doi: 10.1146/annurev-arplant-050312-120043
Baligar, V. C., and Fageria, N. K. (2015). “Nutrient use ufficiency in plants: an overview,” in Nutrient Use Efficiency: from Basics to Advances, eds A. Rakshit, H. B. Singh, and A. Sen (New Delhi: Springer), 1–14.
Bari, R., Datt Pant, B., Stitt, M., and Scheible, W. R. (2006). PHO2, microRNA399, and PHR1 define a phosphate-signaling pathway in plants. Plant Physiol. 141, 988–999. doi: 10.1104/pp.106.079707
Beauclair, L., Yu, A., and Bouché, N. (2010). microRNA-directed cleavage and translational repression of the copper chaperone for superoxide dismutase mRNA in Arabidopsis. Plant J. 62, 454–462. doi: 10.1111/j.1365-313X.2010.04162.x
Bieleski, R. L. (1973). Phosphate pools, phosphate transport, and phosphate availability. Annu. Rev. Plant Physiol. 24, 225–252. doi: 10.1146/annurev.pp.24.060173.001301
Branscheid, A., Sieh, D., Pant, B. D., May, P., Devers, E. A., Elkrog, A., et al. (2010). Expression pattern suggests a role of miR399 in the regulation of the cellular response to local Pi increase during arbuscular mycorrhizal symbiosis. Mol. Plant Microbe Interact. 23, 915–926. doi: 10.1094/MPMI-23-7-0915
Cai, H., Lu, Y., Xie, W., Zhu, T., and Lian, X. (2012). Transcriptome response to nitrogen starvation in rice. J. Biosci. 37, 731–747. doi: 10.1007/s12038-012-9242-2
Chen, X. (2004). A microRNA as a translational repressor of APETALA2 in Arabidopsis flower development. Science 303, 2022–2025. doi: 10.1126/science.1088060
Chiou, T. J. (2007). The role of microRNAs in sensing nutrient stress. Plant Cell Environ. 30, 323–332. doi: 10.1111/j.1365-3040.2007.01643.x
Chiou, T. J., Aung, K., Lin, S. I., Wu, C. C., Chiang, S. F., and Su, C. L. (2006). Regulation of phosphate homeostasis by microRNA in Arabidopsis. Plant Cell 18, 412–421. doi: 10.1105/tpc.105.038943
Chitwood, D. H., and Timmermans, M. C. P. (2010). Small RNAs are on the move. Nature 467, 415–419. doi: 10.1038/nature09351
Chuck, G., Cigan, A. M., Saeteurn, K., and Hake, S. (2007a). The heterochronic maize mutant Corngrass1 results from overexpression of a tandem microRNA. Nat. Genet. 39, 544–549. doi: 10.1038/ng2001
Chuck, G., Meeley, R., Irish, E., Sakai, H., and Hake, S. (2007b). The maize tasselseed4 microRNA controls sex determination and meristem cell fate by targeting Tasselseed6/indeterminate spikelet1. Nat. Genet. 39, 1517–1521. doi: 10.1038/ng.2007.20
Crawford, N. M., and Forde, B. G. (2002). Molecular and developmental biology of inorganic nitrogen nutrition. Arabidopsis Book 1:e0011. doi: 10.1199/tab.0011
Dass, A., Jat, S., and Rana, K. S. (2015). “Resource conserving techniques for improving nitrogen-use efficiency,” in Nutrient Use Efficiency: from Basics to Advances, eds A. Rakshit, H. B. Singh, and A. Sen (New Delhi: Springer), 45–58. doi: 10.1007/978-81-322-2169-2_4
de Groot, C., Marcelis, L. M., van den Boogaard, R., Kaiser, W., and Lambers, H. (2003). Interaction of nitrogen and phosphorus nutrition in determining growth. Plant Soil 248, 257–268. doi: 10.1023/A:1022323215010
Delhaize, E., and Randall, P. J. (1995). Characterization of a phosphate-accumulator mutant of Arabidopsis thaliana. Plant Physiol. 107, 207–213.
Diaz, C., Saliba-Colombani, V., Loudet, O., Belluomo, P., Moreau, L., Daniel-Vedele, F., et al. (2006). Leaf yellowing and anthocyanin accumulation are two genetically independent strategies in response to nitrogen limitation in Arabidopsis thaliana. Plant Cell Physiol. 47, 74–83. doi: 10.1093/pcp/pci225
Fageria, V. D. (2001). Nutrient interactions in crop plants. J. Plant Nutr. 24, 1269–1290. doi: 10.1081/PLN-100106981
Fang, Z., Shao, C., Meng, Y., Wu, P., and Chen, M. (2009). Phosphate signaling in Arabidopsis and Oryza sativa. Plant Sci. 176, 170–180. doi: 10.1016/j.plantsci.2008.09.007
Fei, Q., Xia, R., and Meyers, B. C. (2013). Phased, secondary, small interfering RNAs in posttranscriptional regulatory networks. Plant Cell 25, 2400–2415. doi: 10.1105/tpc.113.114652
Ferdous, J., Hussain, S. S., and Shi, B. J. (2015). Role of microRNAs in plant drought tolerance. Plant Biotech. J. 13, 293–305. doi: 10.1111/pbi.12318
Fischer, J. J., Beatty, P. H., Good, A. G., and Muench, D. G. (2013). Manipulation of microRNA expression to improve nitrogen use efficiency. Plant Sci. 210, 70–81. doi: 10.1016/j.plantsci.2013.05.009
Fu, C., Sunkar, R., Zhou, C., Shen, H., Zhang, J. Y., Matts, J., et al. (2012). Overexpression of miR156 in switchgrass (Panicum virgatum L.) results in various morphological alterations and leads to improved biomass production. Plant Biotech. J. 10, 443–452. doi: 10.1111/j.1467-7652.2011.00677.x
Fujii, H., Chiou, T. J., Lin, S. I., Aung, K., and Zhu, J. K. (2005). A miRNA involved in phosphate-starvation response in Arabidopsis. Curr. Biol. 15, 2038–2043. doi: 10.1016/j.cub.2005.10.016
Gahoonia, T., Claassen, N., and Jungk, A. (1992). Mobilization of phosphate in different soils by ryegrass supplied with ammonium or nitrate. Plant Soil 140, 241–248. doi: 10.1007/BF00010600
Gifford, M. L., Dean, A., Gutierrez, R. A., Coruzzi, G. M., and Birnbaum, K. D. (2008). Cell-specific nitrogen responses mediate developmental plasticity. Proc. Natl. Acad. Sci. U.S.A. 105, 803–808. doi: 10.1073/pnas.0709559105
Giles, J. (2005). Nitrogen study fertilizes fears of pollution. Nature 433, 791–791. doi: 10.1038/433791a
Gniazdowska, A., and Rychter, A. M. (2000). Nitrate uptake by bean (Phaseolus vulgaris L.) roots under phosphate deficiency. Plant Soil 226, 79–85. doi: 10.1023/A:1026463307043
Godfray, H. C. J., Beddington, J. R., Crute, I. R., Haddad, L., Lawrence, D., Muir, J. F., et al. (2010). Food security: the challenge of feeding 9 billion people. Science 327, 812–818. doi: 10.1126/science.1185383
Good, A. G., and Beatty, P. H. (2011). Fertilizing nature: a tragedy of excess in the commons. PLoS Biol. 9:e1001124. doi: 10.1371/journal.pbio.1001124
Good, A. G., Shrawat, A. K., and Muench, D. G. (2004). Can less yield more? Is reducing nutrient input into the environment compatible with maintaining crop production? Trends Plant Sci. 9, 597–605. doi: 10.1016/j.tplants.2004.10.008
Gou, J. Y., Felippes, F. F., Liu, C. J., Weigel, D., and Wang, J. W. (2011). Negative regulation of anthocyanin biosynthesis in Arabidopsis by a miR156-Targeted SPL transcription factor. Plant Cell 23, 1512–1522. doi: 10.1105/tpc.111.084525
Gu, M., Xu, K., Chen, A., Zhu, Y., Tang, G., and Xu, G. (2010). Expression analysis suggests potential roles of microRNAs for phosphate and arbuscular mycorrhizal signaling in Solanum lycopersicum. Physiol. Plant. 138, 226–237. doi: 10.1111/j.1399-3054.2009.01320.x
Guevara, D. R., Bi, Y. M., and Rothstein, S. J. (2014). Identification of regulatory genes to improve nitrogen use efficiency. Can. J. Plant Sci. 94, 1009–1012. doi: 10.4141/cjps2013-154
Guo, B., Jin, Y., Wussler, C., Blancaflor, E. B., Motes, C. M., and Versaw, W. K. (2008). Functional analysis of the Arabidopsis PHT4 family of intracellular phosphate transporters. New Phytol. 177, 889–898. doi: 10.1111/j.1469-8137.2007.02331.x
Guo, H. S., Xie, Q., Fei, J. F., and Chua, N. H. (2005). MicroRNA directs mRNA cleavage of the transcription factor NAC1 to downregulate auxin signals for Arabidopsis lateral root development. Plant Cell 17, 1376–1386. doi: 10.1105/tpc.105.030841
Hackenberg, M., Shi, B. J., Gustafson, P., and Langridge, P. (2013). Characterization of phosphorus-regulated miR399 and miR827 and their isomirs in barley under phosphorus-sufficient and phosphorus-deficient conditions. BMC Plant Biol. 13:214. doi: 10.1186/1471-2229-13-214
Hagen, G., and Guilfoyle, T. (2002). Auxin-responsive gene expression: genes, promoters and regulatory factors. Plant Mol. Biol. 49, 373–385. doi: 10.1023/A:1015207114117
Hamburger, D., Rezzonico, E., MacDonald-Comber, P. J., Somerville, C., and Poirier, Y. (2002). Identification and characterization of the Arabidopsis PHO1 gene involved in phosphate loading to the xylem. Plant Cell 14, 889–902. doi: 10.1105/tpc.000745
Hawkesford, M., Horst, W., Kichey, T., Lambers, H., Schjoerring, J., Møller, I. S., et al. (2012). “Chapter 6 - Functions of macronutrients,” in Marschner's Mineral Nutrition of Higher Plants, 3rd Edn., ed P. Marschner (San Diego, CA: Academic Press), 135–189. doi: 10.1016/B978-0-12-384905-2.00006-6
He, H., Liang, G., Li, Y., Wang, F., and Yu, D. (2014). Two young microRNAs originating from target duplication mediate nitrogen starvation adaptation via regulation of glucosinolate synthesis in Arabidopsis thaliana. Plant Physiol. 164, 853–865. doi: 10.1104/pp.113.228635
Hermans, C., Hammond, J. P., White, P. J., and Verbruggen, N. (2006). How do plants respond to nutrient shortage by biomass allocation? Trends Plant Sci. 11, 610–617. doi: 10.1016/j.tplants.2006.10.007
Hinsinger, P. (2001). Bioavailability of soil inorganic P in the rhizosphere as affected by root-induced chemical changes: a review. Plant Soil 237, 173–195. doi: 10.1023/A:1013351617532
Hsieh, L. C., Lin, S. I., Shih, A. C. C., Chen, J. W., Lin, W. Y., Tseng, C. Y., et al. (2009). Uncovering small RNA-mediated responses to phosphate deficiency in Arabidopsis by deep dequencing. Plant Physiol. 151, 2120–2132. doi: 10.1104/pp.109.147280
Hu, B., Zhu, C., Li, F., Tang, J., Wang, Y., Lin, A., et al. (2011). LEAF TIP NECROSIS1 plays a pivotal role in the regulation of multiple phosphate starvation responses in rice. Plant Physiol. 156, 1101–1115. doi: 10.1104/pp.110.170209
Huang, T. K., Han, C. L., Lin, S. I., Chen, Y. J., Tsai, Y. C., Chen, Y. R., et al. (2013). Identification of downstream components of ubiquitin-conjugating enzyme PHOSPHATE2 by quantitative membrane proteomics in Arabidopsis roots. Plant Cell 25, 4044–4060. doi: 10.1105/tpc.113.115998
Jagadeeswaran, G., Saini, A., and Sunkar, R. (2009). Biotic and abiotic stress down-regulate miR398 expression in Arabidopsis. Planta 229, 1009–1014. doi: 10.1007/s00425-009-0889-3
Jiang, C., Gao, X., Liao, L., Harberd, N. P., and Fu, X. (2007). Phosphate starvation root architecture and anthocyanin accumulation responses are modulated by the gibberellin-DELLA signaling pathway in Arabidopsis. Plant Physiol. 145, 1460–1470. doi: 10.1104/pp.107.103788
Jin, J., Tang, C., Hogarth, T., Armstrong, R., and Sale, P. (2014). Nitrogen form but not elevated CO2 alters plant phosphorus acquisition from sparingly soluble phosphorus sources. Plant Soil 374, 109–119. doi: 10.1007/s11104-013-1870-2
Johnson, J. F., Vance, C. P., and Allan, D. L. (1996). Phosphorus deficiency in Lupinus albus (altered lateral root development and enhanced expression of phosphoenolpyruvate carboxylase). Plant Physiol. 112, 31–41. doi: 10.1104/pp.112.1.31
Jones-Rhoades, M. W., Bartel, D. P., and Bartel, B. (2006). MicroRNAs and their regulatory roles in plants. Annu. Rev. Plant Biol. 57, 19–53. doi: 10.1146/annurev.arplant.57.032905.105218
Jung, J. H., Seo, P., and Park, C. M. (2009). MicroRNA biogenesis and function in higher plants. Plant Biotechnol. Rep. 3, 111–126. doi: 10.1007/s11816-009-0085-8
Jung, J. H., Seo, P., Kang, S., and Park, C. M. (2011). miR172 signals are incorporated into the miR156 signaling pathway at the SPL3/4/5 genes in Arabidopsis developmental transitions. Plant Mol. Biol. 76, 35–45. doi: 10.1007/s11103-011-9759-z
Kamthan, A., Chaudhuri, A., Kamthan, M., and Datta, A. (2015). Small RNAs in plants: recent development and application for crop improvement. Front. Plant Sci. 6:208. doi: 10.3389/fpls.2015.00208
Kandlbinder, A., Finkemeier, I., Wormuth, D., Hanitzsch, M., and Dietz, K. J. (2004). The antioxidant status of photosynthesizing leaves under nutrient deficiency: redox regulation, gene expression and antioxidant activity in Arabidopsis thaliana. Physiol. Plant. 120, 63–73. doi: 10.1111/j.0031-9317.2004.0272.x
Kant, S., Bi, Y. M., and Rothstein, S. J. (2011a). Understanding plant response to nitrogen limitation for the improvement of crop nitrogen use efficiency. J. Exp. Bot. 62, 1499–1509. doi: 10.1093/jxb/erq297
Kant, S., Peng, M., and Rothstein, S. J. (2011b). Genetic regulation by NLA and MicroRNA827 for maintaining nitrate-dependent phosphate homeostasis in Arabidopsis. PLoS Genet. 7:e1002021. doi: 10.1371/journal.pgen.1002021
Karthikeyan, A. S., Varadarajan, D. K., Jain, A., Held, M. A., Carpita, N. C., and Raghothama, K. G. (2007). Phosphate starvation responses are mediated by sugar signaling in Arabidopsis. Planta 225, 907–918. doi: 10.1007/s00425-006-0408-8
Khan, G., Declerck, M., Sorin, C., Hartmann, C., Crespi, M., and Lelandais-Brière, C. (2011). MicroRNAs as regulators of root development and architecture. Plant Mol. Biol. 77, 47–58. doi: 10.1007/s11103-011-9793-x
Khraiwesh, B., Zhu, J. K., and Zhu, J. (2012). Role of miRNAs and siRNAs in biotic and abiotic stress responses of plants. Biochim. Biophys. Acta 1819, 137–148. doi: 10.1016/j.bbagrm.2011.05.001
Kinoshita, N., Wang, H., Kasahara, H., Liu, J., MacPherson, C., Machida, Y., et al. (2012). IAA-Ala Resistant3, an evolutionarily conserved target of miR167, mediates Arabidopsis root architecture changes during high ssmotic stress. Plant Cell 24, 3590–3602. doi: 10.1105/tpc.112.097006
Kirkby, E., and Johnston, A. E. (2008). “Soil and fertilizer phosphorus in relation to crop nutrition,” in The Ecophysiology of Plant-Phosphorus Interactions, eds P. White and J. Hammond (Netherlands: Springer), 177–223. doi: 10.1007/978-1-4020-8435-5_9
Kraiser, T., Gras, D. E., Gutiérrez, A. G., González, B., and Gutiérrez, R. A. (2011). A holistic view of nitrogen acquisition in plants. J. Exp. Bot. 62, 1455–1466. doi: 10.1093/jxb/erq425
Krapp, A., Berthomé, R., Orsel, M., Mercey-Boutet, S., Yu, A., Castaings, L., et al. (2011). Arabidopsis roots and shoots show distinct temporal adaptation patterns toward nitrogen starvation. Plant Physiol. 157, 1255–1282. doi: 10.1104/pp.111.179838
Krapp, A., David, L. C., Chardin, C., Girin, T., Marmagne, A., and Leprince, A. S. (2014). Nitrate transport and signalling in Arabidopsis. J. Exp. Bot. 65, 789–798. doi: 10.1093/jxb/eru001
Kuo, H. F., and Chiou, T. J. (2011). The role of microRNAs in phosphorus deficiency signaling. Plant Physiol. 156, 1016–1024. doi: 10.1104/pp.111.175265
Léran, S., Varala, K., Boyer, J. C., Chiurazzi, M., Crawford, N., Daniel-Vedele, F., et al. (2014). A unified nomenclature of NITRATE TRANSPORTER 1/PEPTIDE TRANSPORTER family members in plants. Trends Plant Sci. 19, 5–9. doi: 10.1016/j.tplants.2013.08.008
Li, W. X., Oono, Y., Zhu, J., He, X. J., Wu, J. M., Iida, K., et al. (2008). The Arabidopsis NFYA5 transcription factor is regulated transcriptionally and post-transcriptionally to promote drought resistance. Plant Cell 20, 2238–2251. doi: 10.1105/tpc.108.059444
Liang, G., He, H., and Yu, D. (2012). Identification of nitrogen starvation-responsive microRNAs in Arabidopsis thaliana. PLoS ONE 7:e48951. doi: 10.1371/journal.pone.0048951
Lin, S. I., Santi, C., Jobet, E., Lacut, E., El-Kholti, N., Karlowski, W. M., et al. (2010). Complex regulation of two target genes encoding SPX-MFS proteins by rice miR827 in response to phosphate starvation. Plant Cell Physiol. 51, 2119–2131. doi: 10.1093/pcp/pcq170
Lin, W. Y., Huang, T. K., and Chiou, T. J. (2013). NITROGEN LIMITATION ADAPTATION, a target of microRNA827, mediates degradation of plasma membrane–localized phosphate transporters to maintain phosphate homeostasis in Arabidopsis. Plant Cell 25, 4061–4074. doi: 10.1105/tpc.113.116012
Lin, W. Y., Lin, S. I., and Chiou, T. J. (2009). Molecular regulators of phosphate homeostasis in plants. J. Exp. Bot. 60, 1427–1438. doi: 10.1093/jxb/ern303
Liscum, E., and Reed, J. W. (2002). Genetics of Aux/IAA and ARF action in plant growth and development. Plant Mol. Biol. 49, 387–400. doi: 10.1023/A:1015255030047
Liu, J. Q., Allan, D. L., and Vance, C. P. (2010). Systemic signaling and local sensing of phosphate in common bean: cross-talk between photosynthate and microRNA399. Mol. Plant 3, 428–437. doi: 10.1093/mp/ssq008
Liu, J., Wu, Y., Yang, J., Liu, Y., and Shen, F. (2008). Protein degradation and nitrogen remobilization during leaf senescence. J. Plant Biol. 51, 11–19. doi: 10.1007/BF03030735
Liu, T. Y., Huang, T. K., Tseng, C. Y., Lai, Y. S., Lin, S. I., Lin, W. Y., et al. (2012). PHO2-dependent degradation of PHO1 modulates phosphate homeostasis in Arabidopsis. Plant Cell 24, 2168–2183. doi: 10.1105/tpc.112.096636
Liu, T. Y., Lin, W. Y., Huang, T. K., and Chiou, T. J. (2014). MicroRNA-mediated surveillance of phosphate transporters on the move. Trends Plant Sci. 19, 647–655. doi: 10.1016/j.tplants.2014.06.004
López-Arredondo, D. L., Leyva-González, M. A., González-Morales, S. I., López-Bucio, J., and Herrera-Estrella, L. (2014). Phosphate nutrition: improving low-phosphate tolerance in crops. Annu. Rev. Plant Biol. 65, 95–123. doi: 10.1146/annurev-arplant-050213-035949
Lundmark, M., Kørner, C. J., and Nielsen, T. H. (2010). Global analysis of microRNA in Arabidopsis in response to phosphate starvation as studied by locked nucleic acid-based microarrays. Physiol. Plant. 140, 57–68. doi: 10.1111/j.1399-3054.2010.01384.x
Maistry, P. M., Muasya, A. M., Valentine, A. J., and Chimphango, S. B. M. (2014). Increasing nitrogen supply stimulates phosphorus acquisition mechanisms in the fynbos species Aspalathus linearis. Funct. Plant Biol. 42, 52–62. doi: 10.1071/FP14100
Mallory, A. C., Bartel, D. P., and Bartel, B. (2005). MicroRNA-directed regulation of Arabidopsis AUXIN RESPONSE FACTOR17 is essential for proper development and modulates expression of early auxin response genes. Plant Cell 17, 1360–1375. doi: 10.1105/tpc.105.031716
Mallory, A. C., Dugas, D. V., Bartel, D. P., and Bartel, B. (2004). MicroRNA regulation of NAC-domain targets is required for proper formation and separation of adjacent embryonic, vegetative, and floral organs. Curr. Biol. 14, 1035–1046. doi: 10.1016/j.cub.2004.06.022
Masclaux-Daubresse, C., Daniel-Vedele, F., Dechorgnat, J., Chardon, F., Gaufichon, L., and Suzuki, A. (2010). Nitrogen uptake, assimilation and remobilization in plants: challenges for sustainable and productive agriculture. Ann. Bot. 105, 1141–1157. doi: 10.1093/aob/mcq028
Massonneau, A., Langlade, N., Léon, S., Smutny, J., Vogt, E., Neumann, G., et al. (2001). Metabolic changes associated with cluster root development in white lupin (Lupinus albus L.): relationship between organic acid excretion, sucrose metabolism and energy status. Planta 213, 534–542. doi: 10.1007/s004250100529
McAllister, C. H., Beatty, P. H., and Good, A. G. (2012). Engineering nitrogen use efficient crop plants: the current status. Plant Biotech. J. 10, 1011–1025. doi: 10.1111/j.1467-7652.2012.00700.x
Meng, Y., Ma, X., Chen, D., Wu, P., and Chen, M. (2010). MicroRNA-mediated signaling involved in plant root development. Biochem. Biophys. Res. Commun. 393, 345–349. doi: 10.1016/j.bbrc.2010.01.129
Misson, J., Raghothama, K. G., Jain, A., Jouhet, J., Block, M. A., Bligny, R., et al. (2005). A genome-wide transcriptional analysis using Arabidopsis thaliana Affymetrix gene chips determined plant responses to phosphate deprivation. Proc. Natl. Acad. Sci. U.S.A. 102, 11934–11939. doi: 10.1073/pnas.0505266102
Olsen, A. N., Ernst, H. A., Leggio, L. L., and Skriver, K. (2005). NAC transcription factors: structurally distinct, functionally diverse. Trends Plant Sci. 10, 79–87. doi: 10.1016/j.tplants.2004.12.010
Pant, B. D., Musialak-Lange, M., Nuc, P., May, P., Buhtz, A., Kehr, J., et al. (2009). Identification of nutrient-responsive Arabidopsis and rapeseed microRNAs by comprehensive real-time polymerase chain reaction profiling and small RNA sequencing. Plant Physiol. 150, 1541–1555. doi: 10.1104/pp.109.139139
Park, B. S., Seo, J. S., and Chua, N. H. (2014). NITROGEN LIMITATION ADAPTATION recruits PHOSPHATE2 to target the phosphate transporter PT2 for degradation during the regulation of Arabidopsis phosphate homeostasis. Plant Cell 26, 454–464. doi: 10.1105/tpc.113.120311
Patil, V. S., Zhou, R., and Rana, T. M. (2014). Gene regulation by non-coding RNAs. Crit. Rev. Biochem. Mol. 49, 16–32. doi: 10.3109/10409238.2013.844092
Paul, M. J., and Stitt, M. (1993). Effects of nitrogen and phosphorus deficiencies on levels of carbohydrates, respiratory enzymes and metabolites in seedlings of tobacco and their response to exogenous sucrose. Plant Cell Environ. 16, 1047–1057. doi: 10.1111/j.1365-3040.1996.tb02062.x
Peng, M., Hannam, C., Gu, H., Bi, Y. M., and Rothstein, S. J. (2007). A mutation in NLA, which encodes a RING-type ubiquitin ligase, disrupts the adaptability of Arabidopsis to nitrogen limitation. Plant J. 50, 320–337. doi: 10.1111/j.1365-313X.2007.03050.x
Peng, M., Hudson, D., Schofield, A., Tsao, R., Yang, R., Gu, H., et al. (2008). Adaptation of Arabidopsis to nitrogen limitation involves induction of anthocyanin synthesis which is controlled by the NLA gene. J. Exp. Bot. 59, 2933–2944. doi: 10.1093/jxb/ern148
Péret, B., Desnos, T., Jost, R., Kanno, S., Berkowitz, O., and Nussaume, L. (2014). Root architecture responses: in search of phosphate. Plant Physiol. 166, 1713–1723. doi: 10.1104/pp.114.244541
Phillips, J. R., Dalmay, T., and Bartels, D. (2007). The role of small RNAs in abiotic stress. FEBS Lett. 581, 3592–3597. doi: 10.1016/j.febslet.2007.04.007
Poirier, Y., and Bucher, M. (2002). Phosphate transport and homeostasis in Arabidopsis. Arabidopsis Book 1:e0024. doi: 10.1199/tab.0024
Raghothama, K. G. (1999). Phosphate acquisition. Annu. Rev. Plant Physiol. Plant Mol. Biol. 50, 665–693. doi: 10.1146/annurev.arplant.50.1.665
Rajagopalan, R., Vaucheret, H., Trejo, J., and Bartel, D. P. (2006). A diverse and evolutionarily fluid set of microRNAs in Arabidopsis thaliana. Genes Dev. 20, 3407–3425. doi: 10.1101/gad.1476406
Ramaekers, L., Remans, R., Rao, I. M., Blair, M. W., and Vanderleyden, J. (2010). Strategies for improving phosphorus acquisition efficiency of crop plants. Field Crops Res. 117, 169–176. doi: 10.1016/j.fcr.2010.03.001
Rausch, C., and Bucher, M. (2002). Molecular mechanisms of phosphate transport in plants. Planta 216, 23–37. doi: 10.1007/s00425-002-0921-3
Reinhart, B. J., Weinstein, E. G., Rhoades, M. W., Bartel, B., and Bartel, D. P. (2002). MicroRNAs in plants. Genes Dev. 16, 1616–1626. doi: 10.1101/gad.1004402
Riley, D., and Barber, S. A. (1971). Effect of ammonium and nitrate fertilization on phosphorus uptake as related to root-induced pH changes at the root-soil interface. Soil Sci. Soc. Am. J. 35, 301–306. doi: 10.2136/sssaj1971.03615995003500020035x
Robson, A., O'Hara, G., and Abbott, L. (1981). Involvement of phosphorus in nitrogen fixation by subterranean clover (Trifolium subterraneum L.). Funct. Plant Biol. 8, 427–436.
Rogers, K., and Chen, S. (2013). Biogenesis, turnover, and mode of action of plant microRNAs. Plant Cell 25, 2383–2399. doi: 10.1105/tpc.113.113159
Rothstein, S. J. (2007). Returning to our roots: making plant biology research relevant to future challenges in agriculture. Plant Cell 19, 2695–2699. doi: 10.1105/tpc.107.053074
Rufty, T. W., Israel, D. W., Volk, R. J., Qiu, J., and Sa, T. (1993). Phosphate regulation of nitrate assimilation in soybean. J. Exp. Bot. 44, 879–891. doi: 10.1093/jxb/44.5.879
Rufty, T. W., MacKown, C. T., and Israel, D. W. (1990). Phosphorus stress effects on assimilation of nitrate. Plant Physiol. 94, 328–333. doi: 10.1104/pp.94.1.328
Rufty, T. W., Siddiqi, M. Y., Glass, A. D. M., and Ruth, T. J. (1991). Altered 13NO−3 influx in phosphorus limited plants. Plant Sci. 76, 43–48. doi: 10.1016/0168-9452(91)90216-U
Schlüter, U., Colmsee, C., Scholz, U., Brautigam, A., Weber, A., Zellerhoff, N., et al. (2013). Adaptation of maize source leaf metabolism to stress related disturbances in carbon, nitrogen and phosphorus balance. BMC Genomics 14:442. doi: 10.1186/1471-2164-14-442
Schlüter, U., Mascher, M., Colmsee, C., Scholz, U., Bräutigam, A., Fahnenstich, H., et al. (2012). Maize source leaf adaptation to nitrogen deficiency affects not only nitrogen and carbon metabolism but also control of phosphate homeostasis. Plant Physiol. 160, 1384–1406. doi: 10.1104/pp.112.204420
Shen, H., Chen, J., Wang, Z., Yang, C., Sasaki, T., Yamamoto, Y., et al. (2006). Root plasma membrane H+-ATPase is involved in the adaptation of soybean to phosphorus starvation. J. Exp. Bot. 57, 1353–1362. doi: 10.1093/jxb/erj111
Shen, J., Yuan, L., Zhang, J., Li, H., Bai, Z., Chen, X., et al. (2011). Phosphorus dynamics: from soil to plant. Plant Physiol. 156, 997–1005. doi: 10.1104/pp.111.175232
Shikata, M., Yamaguchi, H., Sasaki, K., and Ohtsubo, N. (2012). Overexpression of Arabidopsis miR157b induces bushy architecture and delayed phase transition in Torenia fournieri. Planta 236, 1027–1035. doi: 10.1007/s00425-012-1649-3
Shin, R., Berg, R. H., and Schachtman, D. P. (2005). Reactive oxygen species and root hairs in Arabidopsis root response to nitrogen, phosphorus and potassium deficiency. Plant Cell Physiol. 46, 1350–1357. doi: 10.1093/pcp/pci145
Sinha, S., Srinivasan, R., and Mandal, P. K. (2015). “MicroRNA-based approach to improve nitrogen use efficiency in crop plants,” in Nutrient Use Efficiency: from Basics to Advances, eds A. Rakshit, H. B. Singh, and A. Sen (New Delhi: Springer), 221–235. doi: 10.1007/978-81-322-2169-2_15
Spanudakis, E., and Jackson, S. (2014). The role of microRNAs in the control of flowering time. J. Exp. Bot. 65, 365–380. doi: 10.1093/jxb/ert453
Sun, G. (2012). MicroRNAs and their diverse functions in plants. Plant Mol. Biol. 80, 17–36. doi: 10.1007/s11103-011-9817-6
Sunkar, R., and Zhu, J. K. (2004). Novel and stress-regulated microRNAs and other small RNAs from Arabidopsis. Plant Cell 16, 2001–2019. doi: 10.1105/tpc.104.022830
Sunkar, R., Kapoor, A., and Zhu, J. K. (2006). Post-transcriptional induction of two Cu/Zn superoxide dismutase genes in Arabidopsis is mediated by down-regulation of miR398 and important for oxidative stress tolerance. Plant Cell 18, 2051–2065. doi: 10.1105/tpc.106.041673
Sunkar, R., Li, Y. F., and Jagadeeswaran, G. (2012). Functions of microRNAs in plant stress responses. Trends Plant Sci. 17, 196–203. doi: 10.1016/j.tplants.2012.01.010
Taylor, M. D., Nelson, P. V., Frantz, J. M., and Rufty, T. W. (2010). Phosphorus deficiency in Pelargonium: effects on nitrate and ammonium uptake and acidity generation. J. Plant Nutr. 33, 701–712. doi: 10.1080/01904160903575923
Tinker, P., and Nye, P. (2000). Solute Movement in the Rhizosphere. Topics in sustainable agronomy. New York, NY: Oxford University Press.
Trevisan, S., Nonis, A., Begheldo, M., Manoli, A., Palme, K., Caporale, G., et al. (2012). Expression and tissue-specific localization of nitrate-responsive miRNAs in roots of maize seedlings. Plant Cell Environ. 35, 1137–1155. doi: 10.1111/j.1365-3040.2011.02478.x
Uauy, C., Distelfeld, A., Fahima, T., Blechl, A., and Dubcovsky, J. (2006). A NAC gene regulating senescence improves grain protein, zinc, and iron content in wheat. Science 314, 1298–1301. doi: 10.1126/science.1133649
Valdés-López, O., Arenas-Huertero, C., Ramirez, M., Girard, L., Sanchez, F., Vance, C. P., et al. (2008). Essential role of MYB transcription factor: PvPHR1 and microRNA: PvmiR399 in phosphorus-deficiency signalling in common bean roots. Plant Cell Environ. 31, 1834–1843. doi: 10.1111/j.1365-3040.2008.01883.x
Valdés-López, O., Yang, S. S., Aparicio-Fabre, R., Graham, P. H., Reyes, J. L., Vance, C. P., et al. (2010). MicroRNA expression profile in common bean (Phaseolus vulgaris) under nutrient deficiency stresses and manganese toxicity. New Phytol. 187, 805–818. doi: 10.1111/j.1469-8137.2010.03320.x
Vance, C. P., Uhde-Stone, C., and Allan, D. L. (2003). Phosphorus acquisition and use: critical adaptations by plants for securing a nonrenewable resource. New Phytol. 157, 423–447. doi: 10.1046/j.1469-8137.2003.00695.x
Versaw, W. K., and Harrison, M. J. (2002). A chloroplast phosphate transporter, PHT2;1, Influences allocation of phosphate within the plant and phosphate-starvation responses. Plant Cell 14, 1751–1766. doi: 10.1105/tpc.002220
Vidal, E. A., Moyano, T. C., Canales, J., and Gutiérrez, R. A. (2014). Nitrogen control of developmental phase transitions in Arabidopsis thaliana. J. Exp. Bot. 65, 5611–5618. doi: 10.1093/jxb/eru326
Voinnet, O. (2009). Origin, biogenesis, and activity of plant microRNAs. Cell 136, 669–687. doi: 10.1016/j.cell.2009.01.046
Wang, J. W., Czech, B., and Weigel, D. (2009). miR156-regulated SPL transcription factors define an endogenous flowering pathway in Arabidopsis thaliana. Cell 138, 738–749. doi: 10.1016/j.cell.2009.06.014
Wang, J. W., Schwab, R., Czech, B., Mica, E., and Weigel, D. (2008). Dual effects of miR156-targeted SPL genes and CYP78A5/KLUH on plastochron length and organ size in Arabidopsis thaliana. Plant Cell 20, 1231–1243. doi: 10.1105/tpc.108.058180
Wang, J. W., Wang, L. J., Mao, Y. B., Cai, W. J., Xue, H. W., and Chen, X. Y. (2005). Control of root cap formation by microRNA-targeted auxin response factors in Arabidopsis. Plant Cell 17, 2204–2216. doi: 10.1105/tpc.105.033076
Wang, Y., Zhang, C., Hao, Q., Sha, A., Zhou, R., Zhou, X., et al. (2013). Elucidation of miRNAs-mediated responses to low nitrogen stress by deep sequencing of two soybean genotypes. PLoS ONE 8:e67423. doi: 10.1371/journal.pone.0067423
Weiberg, A., Wang, M., Bellinger, M., and Jin, H. (2014). Small RNAs: A new paradigm in plant-microbe interactions. Annu. Rev. Phytopathol. 52, 495–516. doi: 10.1146/annurev-phyto-102313-045933
Wu, G. (2013). Plant microRNAs and development. J. Genet. Genomics 40, 217–230. doi: 10.1016/j.jgg.2013.04.002
Wu, G., and Poethig, R. S. (2006). Temporal regulation of shoot development in Arabidopsis thaliana by miR156 and its target SPL3. Development 133, 3539–3547. doi: 10.1242/dev.02521
Wu, G., Park, M. Y., Conway, S. R., Wang, J. W., Weigel, D., and Poethig, R. S. (2009). The sequential action of miR156 and miR172 regulates developmental timing in Arabidopsis. Cell 138, 750–759. doi: 10.1016/j.cell.2009.06.031
Wu, M. F., Tian, Q., and Reed, J. W. (2006). Arabidopsis microRNA167 controls patterns of ARF6 and ARF8 expression, and regulates both female and male reproduction. Development 133, 4211–4218. doi: 10.1242/dev.02602
Wu, P., Ma, L., Hou, X., Wang, M., Wu, Y., Liu, F., et al. (2003). Phosphate starvation triggers distinct alterations of genome expression in Arabidopsis roots and leaves. Plant Physiol. 132, 1260–1271. doi: 10.1104/pp.103.021022
Xie, K., Wu, C., and Xiong, L. (2006). Genomic organization, differential expression, and interaction of SQUAMOSA Promoter-Binding-Like transcription factors and microRNA156 in rice. Plant Physiol. 142, 280–293. doi: 10.1104/pp.106.084475
Xing, S., Salinas, M., Höhmann, S., Berndtgen, R., and Huijser, P. (2010). miR156-targeted and nontargeted SBP-Box transcription factors act in concert to secure male fertility in Arabidopsis. Plant Cell 22, 3935–3950. doi: 10.1105/tpc.110.079343
Xu, G., Fan, X., and Miller, A. J. (2012). Plant nitrogen assimilation and use efficiency. Annu. Rev. Plant Biol. 63, 153–182. doi: 10.1146/annurev-arplant-042811-105532
Xu, Z., Zhong, S., Li, X., Li, W., Rothstein, S. J., Zhang, S., et al. (2011). Genome-wide identification of microRNAs in response to low nitrate availability in maize leaves and roots. PLoS ONE 6:e28009. doi: 10.1371/journal.pone.0028009
Yamaguchi, A., and Abe, M. (2012). Regulation of reproductive development by non-coding RNA in Arabidopsis: to flower or not to flower. J. Plant Res. 125, 693–704. doi: 10.1007/s10265-012-0513-7
Yan, F., Zhu, Y., Müller, C., Zörb, C., and Schubert, S. (2002). Adaptation of H+-pumping and plasma membrane H+ ATPase activity in proteoid roots of white lupin under phosphate deficiency. Plant Physiol. 129, 50–63. doi: 10.1104/pp.010869
Yan, Y., Wang, H., Hamera, S., Chen, X., and Fang, R. (2014). miR444a has multiple functions in the rice nitrate-signaling pathway. Plant J. 78, 44–55. doi: 10.1111/tpj.12446
Yi, X., Zhang, Z., Ling, Y., Xu, W., and Su, Z. (2015). PNRD: a plant non-coding RNA database. Nucleic Acids Res. 43, D982–D989. doi: 10.1093/nar/gku1162
Zeng, H. Q., Zhu, Y. Y., Huang, S. Q., and Yang, Z. M. (2010). Analysis of phosphorus-deficient responsive miRNAs and cis-elements from soybean (Glycine max L.). J. Plant Physiol. 167, 1289–1297. doi: 10.1016/j.jplph.2010.04.017
Zeng, H., Liu, G., Kinoshita, T., Zhang, R., Zhu, Y., Shen, Q., et al. (2012). Stimulation of phosphorus uptake by ammonium nutrition involves plasma membrane H+ ATPase in rice roots. Plant Soil 357, 205–214. doi: 10.1007/s11104-012-1136-4
Zeng, H., Wang, G., Hu, X., Wang, H., Du, L., and Zhu, Y. (2014). Role of microRNAs in plant responses to nutrient stress. Plant Soil 374, 1005–1021. doi: 10.1007/s11104-013-1907-6
Zhang, R., Liu, G., Wu, N., Gu, M., Zeng, H., Zhu, Y., et al. (2011a). Adaptation of plasma membrane H+ ATPase and H+ pump to P deficiency in rice roots. Plant Soil 349, 3–11. doi: 10.1007/s11104-011-0774-2
Zhang, X., Zou, Z., Zhang, J., Zhang, Y., Han, Q., Hu, T., et al. (2011b). Over-expression of sly-miR156a in tomato results in multiple vegetative and reproductive trait alterations and partial phenocopy of the sft mutant. FEBS Lett. 585, 435–439. doi: 10.1016/j.febslet.2010.12.036
Zhao, L., Kim, Y., Dinh, T. T., and Chen, X. (2007). miR172 regulates stem cell fate and defines the inner boundary of APETALA3 and PISTILLATA expression domain in Arabidopsis floral meristems. Plant J. 51, 840–849. doi: 10.1111/j.1365-313X.2007.03181.x
Zhao, M., Ding, H., Zhu, J. K., Zhang, F., and Li, W. X. (2011). Involvement of miR169 in the nitrogen-starvation responses in Arabidopsis. New Phytol. 190, 906–915. doi: 10.1111/j.1469-8137.2011.03647.x
Zhao, M., Tai, H., Sun, S., Zhang, F., Xu, Y., and Li, W. X. (2012). Cloning and characterization of maize miRNAs involved in responses to nitrogen deficiency. PLoS ONE 7:e29669. doi: 10.1371/journal.pone.0029669
Zhao, Y., Xu, Z., Mo, Q., Zou, C., Li, W., Xu, Y., et al. (2013). Combined small RNA and degradome sequencing reveals novel miRNAs and their targets in response to low nitrate availability in maize. Ann. Bot. 112, 633–642. doi: 10.1093/aob/mct133
Zhou, J., Jiao, F., Wu, Z., Li, Y., Wang, X., He, X., et al. (2008). OsPHR2 is involved in phosphate-starvation signaling and excessive phosphate accumulation in shoots of plants. Plant Physiol. 146, 1673–1686. doi: 10.1104/pp.107.111443
Zhu, C., Ding, Y., and Liu, H. (2011). MiR398 and plant stress responses. Physiol. Plant. 143, 1–9. doi: 10.1111/j.1399-3054.2011.01477.x
Keywords: microRNA, fertilizer, nitrogen deficiency, phosphorous deficiency, nitrate and phosphate interaction
Citation: Nguyen GN, Rothstein SJ, Spangenberg G and Kant S (2015) Role of microRNAs involved in plant response to nitrogen and phosphorous limiting conditions. Front. Plant Sci. 6:629. doi: 10.3389/fpls.2015.00629
Received: 04 March 2015; Accepted: 30 July 2015;
Published: 13 August 2015.
Edited by:
Andrzej Miroslaw Pacak, Adam Mickiewicz University, PolandCopyright © 2015 Nguyen, Rothstein, Spangenberg and Kant. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Surya Kant, Biosciences Research, Department of Economic Development, Grains Innovation Park, 110 Natimuk Road, Horsham, VIC 3400, Australia,c3VyeWEua2FudEBlY29kZXYudmljLmdvdi5hdQ==
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.