 Xiahezi Kuai
Xiahezi Kuai Brandon J. MacLeod
Brandon J. MacLeod Charles Després
Charles Després
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
PERSPECTIVE article
Front. Plant Sci. , 10 April 2015
Sec. Plant Pathogen Interactions
Volume 6 - 2015 | https://doi.org/10.3389/fpls.2015.00235
This article is part of the Research Topic Salicylic acid signaling networks View all 20 articles
Salicylic acid (SA) is a mandatory plant metabolite in the deployment of systemic acquired resistance (SAR), a broad-spectrum systemic immune response induced by local inoculation with avirulent pathogens. The NPR1 transcription co-activator is the central node positively regulating SAR. SA was the last of the major hormones to be without a known receptor. Recently, NPR1 was shown to be the direct link between SA and gene activation. This discovery seems to be controversial. NPR1 being an SA-receptor is reminiscent of the mammalian steroid receptors, which are transcription factors whose binding to DNA is dependent on the interaction with a ligand. Unlike steroid receptors, NPR1 does not bind directly to DNA, but is recruited to promoters by the TGA family of transcription factors to form an enhanceosome. In Arabidopsis, NPR1 is part of a multigene family in which two other members, NPR3 and NPR4, have also been shown to interact with SA. NPR3/NPR4 are negative regulators of immunity and act as substrate adaptors for the recruitment of NPR1 to an E3-ubiquitin ligase, leading to its subsequent degradation by the proteasome. In this perspective, we will stress-test in a friendly way the current NPR1/NPR3/NPR4 model.
Salicylic acid (SA) is an endogenous plant hormone essential to the deployment of a long-lasting, broad-based immunity termed systemic acquired resistance (SAR). SA protects plants from a wide range of phytopathogens by mediating immune response at both local and systemic level (Vlot et al., 2009). SA has also been found to participate in abiotic stress responses. For instance, exogenous SA applications induce tolerance to copper toxicity (Mostofa and Fujita, 2013). In addition to its role in biotic and abiotic stress resistances, SA can influence plant flowering and thermogenesis (Vlot et al., 2009). Due to its biological significance, the synthesis and signal transduction of SA has been intensely studied. Still, not much is known about the molecular details of the SA signaling pathway and the SA receptor remained unidentified for decades.
In 2012, two independent groups contributed new insights into the SA-perception and signaling-cascade. Interestingly, these advances are all centered on the NPR1 protein. One study showed that NPR1 can directly bind SA and acts as an SA-receptor (Wu et al., 2012). The other group proposed that two NPR1 paralogs, NPR3, and NPR4, bind SA and control the proteasome-mediated degradation of NPR1 through their interaction with NPR1 (Fu et al., 2012). Both groups however, demonstrated the indispensable role of NPR1 in SA signaling. The focus of this perspective centers on NPR1 as the mediator of SA-perception, while comparing the SA-binding properties and molecular mechanisms of NPR1, NPR3, and NPR4. These data are compiled in Table 1. Furthermore, we will address some shortcomings in our current understanding of the SA-signaling pathway in the context of plant immunity.
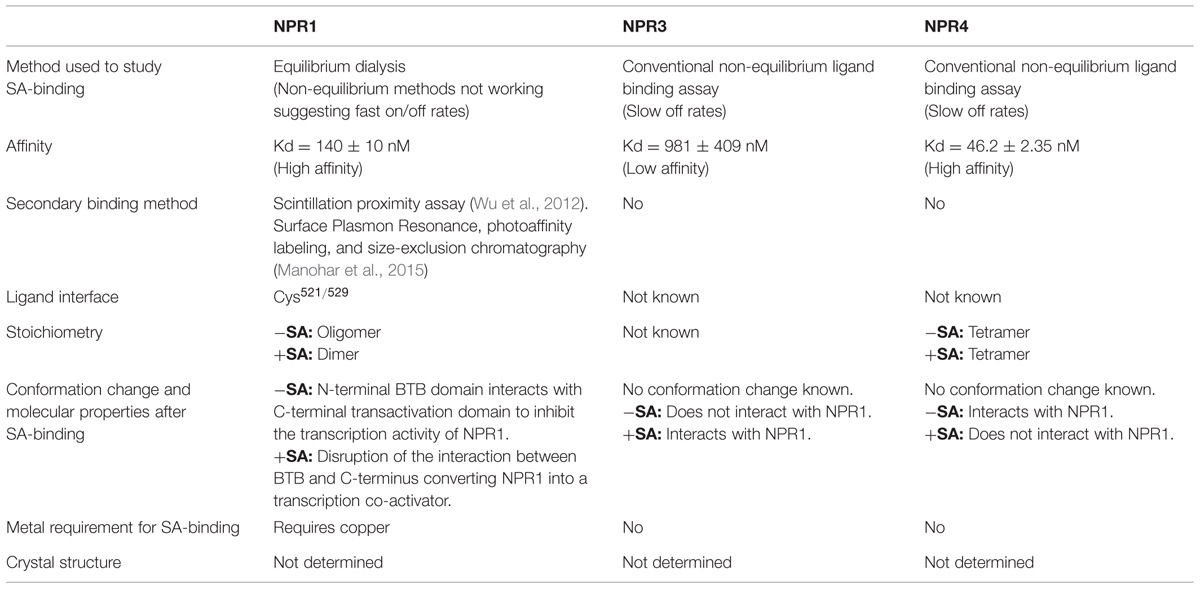
TABLE 1. Comparison of salicylic acid (SA)-binding properties between NPR1, NPR3, and NPR4.
NPR1 is a central regulator of plant immunity, which controls both local resistance and SAR. Plants lacking a functional NPR1 protein are unable to undergo SAR or express the SAR-marker gene PR1, and as a result succumb to biotrophic pathogenic challenges (Cao et al., 1994; Delaney et al., 1995). Later, it was shown that NPR1 is a transcription coactivator (Rochon et al., 2006). The molecular mechanisms of NPR1 function are best understood in the case of PR1. Transcription of PR1 is repressed by TGA2 transcription factor under SA concentration existing in naïve cells (Zhang et al., 2003; Rochon et al., 2006). Upon build-up of SA, NPR1 activates PR1 transcription by forming an enhansome with TGA2 on the promoter and negating the repressor activity of TGA2 (Boyle et al., 2009). The formation of the enhanceosome is well understood. However, the exact role played by SA leading to its formation remains unclear.
Structurally, NPR1 contains an N-terminal BTB/POZ domain, an ankyrin repeat domain, a C-terminal transactivation domain and a nuclear localization sequence. The ankyrin repeats of NPR1 are responsible for its interaction with TGA2 (Zhang et al., 1999). The BTB/POZ also contacts TGA2 masking its repressor domain (Boyle et al., 2009). Besides its role in converting TGA2 from a repressor to an activator, the BTB/POZ also acts as an autoinhibitory domain. In the absence of SA, it interacts with the NPR1 C-terminal transactivation domain, and inhibits the transcription co-activator function of NPR1 (Wu et al., 2012). Two cysteines (Cys521 and Cys529), located in the C-terminus of NPR1, are crucial for the SA-induced transactivation activity of NPR1 (Rochon et al., 2006). These same Cys are required for the direct binding of SA to Arabidopsis NPR1 (Wu et al., 2012). Mechanistically, the binding of SA leads to the disruption of the interaction between the BTB/POZ and the C-terminus, thus releasing the C-terminal transactivation domain from autoinhibition by the BTB/POZ domain and converting NPR1 into an activated transcription co-activator.
A novel and interesting feature of NPR1, aside from being a newly discovered and important phytohormone-receptor, is the requirement of the transition metal copper for SA-binding. Mutation of Cys521 and Cys529 of the C-terminal transactivation domain not only disrupts the SA-binding capacity of NPR1, but also eliminates the recruitment of copper by NPR1 (Wu et al., 2012). This is the first plant example of a copper-binding protein acting as a transcription regulator. The fact that NPR1 is a metalloprotein explains why it took so long to identify it as an SA receptor. Many researchers, by default, include EDTA as a chelator when preparing buffers. However, recruitment of SA by NPR1 is EDTA-sensitive and its presence in buffers precludes SA from binding to NPR1 (Wu et al., 2012). Despite the fact that NPR1 is the first copper-binding transcription-regulator discovered in plant, it is not the first time that copper is found to play a critical function in hormone signal-transduction pathway. The high-affinity binding-activity of the gaseous plant hormone, ethylene, to the ethylene receptor, ETR1, also requires copper as a cofactor (Rodriguez et al., 1999). As is the case of SA in NPR1, ethylene is coordinated to copper in the ETR1 hormone-binding pocket.
NPR1 is a positive regulator of SAR. Recently, additional members of the NPR family, NPR3, and NPR4, were shown to negatively regulate SAR (Liu et al., 2005; Zhang et al., 2006; Fu et al., 2012). Analysis of conceptual gene products revealed that NPR3 and NPR4, respectively, share 34.5 and 36.0% amino acid-conservation with NPR1, specifically in the BTB/POZ and ankyrin repeat domains (Liu et al., 2005). Protein alignments indicate that all three NPR share four (4) conserved Cys in their BTB/POZ domain, and a stretch of five (5) variable basic-amino acids at the C-terminal, that may be involved in nuclear localization (Shi et al., 2013). The structural similarities among these three protein appears to extend to their functional roles including SA-perception and interaction with members of the TGA family of transcription factors (Després et al., 2000; Kinkema et al., 2000; Subramaniam et al., 2001; Fan and Dong, 2002; Mou et al., 2003; Rochon et al., 2006).
At the organ level, expression of NPR1/NPR3/NPR4 appears to occur in different locations. Promoter-driven GFP expression observed with fluorescence stereomicroscopy, demonstrated that NPR1 was detectable only in leaves, NPR4 only in mature siliques and roots, while NPR3 was expressed in relatively high quantities in the young flower (Shi et al., 2013). At the subcellular level, NPR3/NPR4-TGA2 interactions have been observed primarily in the nucleus, when studied in onion epidermal cells and Arabidopsis mesophyll protoplasts (Zhang et al., 2006). While nuclear localization of NPR1 has been shown definitively, differing reports have suggested that NPR1 can also be observed in the cytoplast as well (Després et al., 2000).
The pathology surrounding npr1/npr3/npr4 mutants has displayed different phenotypes under the exact and differential conditions. Early experiments infecting npr4-1 plants with the fungi Erysiphe cichoracearum (powdery mildew) and bacterium Pseudomonas syringae pv. Tomato DC3000 (Pst DC3000) indicated that these plants were compromised in disease resistance (Liu et al., 2005). However, an independent study from Zhang et al. (2006), partially disagreed, rather observing that the npr4-3 and npr4-2 plants were not more susceptible to Pst DC3000 or P. syringae pv. maculicola ES4326 (Psm ES4326). When combined with the npr3-1 mutant (npr3-1npr4-3) plants were found to be more resistant (Zhang et al., 2006). Corroborating the results of Zhang et al. (2006), single npr3 or npr4 mutants showed little difference in SAR response when compared to Col-0. Furthermore, the double mutant (npr3npr4) was highly resistant in basal and induced SAR states (Fu et al., 2012). At the basal level, NPR3 deficient backgrounds have compromised fitness when measured by primary root length, average growth rates, and seed production. Most recently an npr3-3 mutant was generated and found to not differ from Col-0 plants in terms of quantity of bacterial growth when leaves were infiltrated with Pst DC3000, consistent with previous data. Conversely, transgenic plants overexpressing NPR3 were more susceptible to inoculation (Shi et al., 2013). Interestingly, the quantity of NPR3 transcripts was approximately threefold lower in flower petals when taken from the npr3-3 background in comparison to the npr3-2 mutant (Shi et al., 2013). Although, both backgrounds were created from homozygous T-DNA insertions in the third exon, the npr3-2 plant may nonetheless be a “weak allele” in flowers, at least (Shi et al., 2013). The discrepancies observed between laboratories when testing the same pathogens reflect the complexity of the disease resistance phenotype compared to the analysis of the SAR-marker gene PR1. Differences may result from the use of different mutant alleles. However, the theme emerging from these data is the functional redundancy, at least in leaves, of NPR3 and NPR4 with respect to immunity. This somewhat contrasts with the proposed role of NPR4 and NPR3 functioning as independent SA-receptors under low and high SA concentrations, respectively.
Contemporary analysis suggests that NPR4 is a CUL3 E3-ligase substrate-adapter in naïve cells, which can interact with NPR1, allowing for the continuous ubiquitylation and turnover of NPR1 by the proteasome. During SAR, the cellular accumulation of SA allows NPR4 to bind the hormone, disrupting the NPR4–NPR1 interaction and abolishing the adaptor-substrate complex. Conversely, NPR3 responds to the abundance of SA by presumably binding to the hormone allowing NPR3 to interact with NPR1, resuming ubiquitylation of NPR1, targeting it for degradation (Fu et al., 2012). Hence, NPR3 and NPR4 would function as both substrate adaptors and SA- receptors that mediate the degradation of NPR1 in the SAR induced and naïve cells, respectively.
Despite the attractiveness of this model, it has yet to be demonstrated how NPR3 or NPR4 actually interact and bind SA. Furthermore, no structural changes in these proteins were directly observed upon binding SA. Such conformational changes are the usual hallmark of receptor-ligand interactions. In what appears to be a controversial finding, the study by Fu et al. (2012) suggested that NPR1 was unable to bind SA. Interestingly, using the same non-equilibrium method (see Table 1), Wu et al. (2012) came to the same conclusion, as they also found that NPR1 could not bind SA under these conditions. However, NPR1 clearly binds SA under equilibrium conditions when appropriate methodologies are used and chelating agents are omitted from experimental buffers (Wu et al., 2012; Table 1). Furthermore, while this manuscript was under review, Manohar et al. (2015), demonstrated, using three alternative methods, that NPR1 binds SA, bringing to five the total number of methods tested to demonstrate that NPR1 is an SA-receptor (Table 1). While these data clearly confirm that NPR1 binds SA and should put an end to the controversy, they also clearly show the need to confirm that NPR3 and NPR4 can indeed bind SA, especially given the fact that they do not undergo conformational changes upon binding SA. Therefore, considering that NPR1 is also an SA-receptor that binds the hormone with a relatively high affinity in the presence of copper, it is also unclear in vivo whether the interaction between NPR1–NPR3/NPR4 is a result of SA bound to NPR1 or to NPR3/NPR4. Since yeast-two-hybrid assays were used to study the SA-dependent interactions between NPR1 and NPR3/NPR4, it is possible that cellular copper was present at quantities sufficient to allow NPR1 to bind SA. Transient BiFC assays in naïve onion epidermal cells have also indicated an interaction between NPR1 and NPR3. However, it is unclear whether basal levels of SA were present at sufficient concentrations in the naïve onion epidermal cells to allow SA perception by NPR1 or NPR3, making it unclear whether or not the interaction requires NPR1 bound SA, NPR3 bound SA, or whether the interaction requires the presence of SA at all in vivo (Shi et al., 2013). However, because NPR1 has a higher affinity for SA than NPR3, as observed by the respective dissociation constants, it would follow that NPR1 would outcompete NPR3 for the interaction with SA (Fu et al., 2012; Wu et al., 2012). Given that NPR1 is the only NPR (among NPR1/NPR3/NPR4) shown to display a conformational change upon binding to its ligand, NPR1 may in fact be the decisional entity responsible for dictating whether interaction with NPR3/NPR4 occurs, regardless of the SA-status of the system.
Although NPR3 and NPR4 appear to degrade NPR1 in an SA-dependent and independent model, respectively, the biochemical and phenotypic data observed from the npr3, npr4, and npr3npr4 mutant plants are not always in agreement with this hypothesis. For example, in the in vivo NPR1 degradation experiment (Figure 1A in Fu et al., 2012), in the npr4 mutant, in which the NPR3-mediated NPR1-degradation is not affected, NPR1 accumulates to the highest levels after 8 h SA application. This indicates that NPR4 and not NPR3 is responsible for degrading NPR1 under SA conditions, which is not consistent with the model. Furthermore, although NPR1 accumulates to some extent in npr3npr4 mutant before SA application, NPR1 accumulates to even greater extent in the npr3npr4 double mutant in response to SA treatment, which indicates that the npr3npr4 mutant is not completely insensitive to SA, suggesting that there is(are) other SA receptor(s) which mediate or trigger the accumulation of NPR1. Another indication, illustrating the presence of central receptor(s) of SA other than NPR3/NPR4, is the data showing that Psm ES4326 growth is significantly decreased in the npr3npr4 double mutant plant even without SAR induction (Figure 4A in Fu et al., 2012). This does not suggest that SAR is defective as proposed by the authors, but rather that SAR is already established in the npr3npr4 double mutant. Further inconsistencies with the model are revealed by the SAR sets of experiments. Although SA accumulation was not quantified in these experiments, treatment with the Psm avrRpt2 strain would presumably induce SAR and thus promote SA accumulation. Therefore the model would predict that, if NPR4 is a CUL3 substrate adaptor only in the absence of SA, the npr3npr4 mutant should not be more resistant than the single npr3 mutant.
On the PR1 front, the relative expression of the gene in naïve cells shows a slightly higher than wild-type induction in the npr3 plants and about the same induction as wild-type in npr4 plants. By contrast PR1 induction was several folds greater in the npr3npr4 plants when compared with wild-type or the single npr3 or npr4 mutants. The current NPR3/NPR4–NPR1 degradation model would predict rather that the npr4 plants should display similar PR1 induction as the npr3npr4 plants and that the npr3 plants should be no different from the wild-type. This is expected because of the lack of NPR3-targeted degradation of NPR1 in naïve cells. As proposed by Zhang et al. (2006), NPR3 and NPR4 appear to have redundant functions with respect to immunity, as opposed to the model proposed by Fu et al. (2012), where they have distinct non-overlapping functions.
The NPR3/NPR4-mediated NPR1 degradation is reminiscent of the emerging trend of ubiquitylation in plant hormone signaling (Santner and Estelle, 2009). Auxins act by stimulating the degradation of Aux/IAA transcriptional repressor through the ubiquitin-ligase complex SCFTIR1 (Gray et al., 2001). Jasmonates activate downstream gene transcription by promoting degradation of the JAZ family of repressors through SCFCOL1 E3 ubiquitin-ligase (Chini et al., 2007). The gibberellin receptor GID1 mediates ubiquitylation and degradation of DELLA repressor, thus activating gibberellin-responsive gene transcription (Griffiths et al., 2006). It seems that in many signaling pathways, plants use ubiquitin and the proteasome pathway to regulate the abundance of negative regulators of the corresponding system. However, in contrast to the aforementioned pathways, in the case of SA signaling, the proteasome targets the positive regulator NPR1. Although the biological importance and molecular mechanism of SA-regulated NPR1-degradation needs further investigation, ubiquitylation also plays a role in mediating SA signaling (Fu et al., 2012).
Cys521 and Cys529 responsible for the binding of SA to the Arabidopsis NPR1 are not universally conserved in NPR1 orthologs, such as those found in crops. However, metal interaction with proteins is not limited to Cys, since any amino acid harboring electronegative elements in its side chain can potentially participate in metal interaction (Wu et al., 2012; Figure S2 therein). Objectively, this leaves us with three possible scenarios: (1) NPR1 from crops could bind SA through metal-coordination, as does the Arabidopsis NPR1, using amino-acids other than Cys; (2) NPR1 from crops could bind SA without coordination through a metal; (3) NPR1 from crops are not receptors for SA. Further research on crop NPR1 should prove invaluable in assessing whether, in the case of NPR1, Arabidopsis can serve as a model system or whether it is the exception to the rule.
Since NPR1 is a transcriptional coactivator (Rochon et al., 2006), the discovery that two NPR1 family members are Cul3 substrate-adaptors (Fu et al., 2012) came as a surprise. Given that NPR3 and NPR4, just like NPR1, interact with the TGA family of transcription factors (Liu et al., 2005; Zhang et al., 2006), a role for these proteins in transcription regulation would have been anticipated. Nevertheless, as proposed (Zhang et al., 2006), a regulatory function for NPR3 and NPR4 involving transcriptional control may still be revealed in the future.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
CD is supported by the National Science and Engineering Research Council (NSERC) Discovery Grant and Accelerator Grant Programs. BM was supported by an NSERC Undergraduate Student Research Award.
Boyle, P., Le, S. E., Rochon, A., Shearer, H. L., Murmu, J., Chu, J. Y., et al. (2009). The BTB/POZ domain of the Arabidopsis disease resistance protein NPR1 interacts with the repression domain of TGA2 to negate its function. Plant Cell 21, 3700–3713. doi: 10.1105/tpc.109.069971
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Cao, H., Bowling, S. A., Gordon, A. S., and Dong, X. (1994). Characterization of an Arabidopsis mutant that is nonresponsive to inducers of systemic acquired resistance. Plant Cell 6, 1583–1592. doi: 10.1105/tpc.6.11.1583
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Chini, A., Fonseca, S., Fernandez, G., Adie, B., Chico, J. M., Lorenzo, O., et al. (2007). The JAZ family of repressors is the missing link in jasmonate signalling. Nature 448, 666–671. doi: 10.1038/nature06006
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Delaney, T. P., Friedrich, L., and Ryals, J. A. (1995). Arabidopsis signal transduction mutant defective in chemically and biologically induced disease resistance. Proc. Natl. Acad. Sci. U.S.A. 92, 6602–6606. doi: 10.1073/pnas.92.14.6602
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Després, C., DeLong, C., Glaze, S., Liu, E., and Fobert, P. R. (2000). The Arabidopsis NPR1/NIM1 protein enhances the DNA binding activity of a subgroup of the TGA family of bZIP transcription factors. Plant Cell 12, 279–290. doi: 10.1105/tpc.12.2.279
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Fan, W., and Dong, X. (2002). In vivo interaction between NPR1 and transcription factor TGA2 leads to salicylic acid-mediated gene activation in Arabidopsis. Plant Cell 14, 1377–1389. doi: 10.1105/tpc.001628
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Fu, Z. Q., Yan, S., Saleh, A., Wang, W., Ruble, J., Oka, N., et al. (2012). NPR3 and NPR4 are receptors for the immune signal salicylic acid in plants. Nature 486, 228–232. doi: 10.1038/nature11162
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Gray, W. M., Kepinski, S., Rouse, D., Leyser, O., and Estelle, M. (2001). Auxin regulates SCF(TIR1)-dependent degradation of AUX/IAA proteins. Nature 414, 271–276. doi: 10.1038/35104500
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Griffiths, J., Murase, K., Rieu, I., Zentella, R., Zhang, Z. L., Powers, S. J., et al. (2006). Genetic characterization and functional analysis of the GID1 gibberellin receptors in Arabidopsis. Plant Cell 18, 3399–3414. doi: 10.1105/tpc.106.047415
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Kinkema, M., Fan, W., and Dong, X. (2000). Nuclear localization of NPR1 is required for activation of PR gene expression. Plant Cell 12, 2339–2350. doi: 10.1105/tpc.12.12.2339
Liu, G., Holub, E. B., Alonso, J. M., Ecker, J. R., and Fobert, P. R. (2005). An Arabidopsis NPR1-like gene, NPR4, is required for disease resistance. Plant J. 41, 304–318. doi: 10.1111/j.1365-313X.2004.02296.x
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Manohar, M., Tian, M., Moreau, M., Park, S. W., Choi, H. W., Fei, Z., et al. (2015). Identification of multiple salicylic acid-binding proteins using two high throughput screens. Front. Plant Sci. 5:777. doi: 10.3389/fpls.2014.00777
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Mostofa, M. G., and Fujita, M. (2013). Salicylic acid alleviates copper toxicity in rice (Oryza sativa L.) seedlings by up-regulating antioxidative and glyoxalase systems. Ecotoxicology 22, 959–973. doi: 10.1007/s10646-013-1073-x
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Mou, Z., Fan, W., and Dong, X. (2003). Inducers of plant systemic acquired resistance regulate NPR1 function through redox changes. Cell 113, 935–944. doi: 10.1016/S0092-8674(03)00429-X
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Rochon, A., Boyle, P., Wignes, T., Fobert, P. R., and Despres, C. (2006). The coactivator function of Arabidopsis NPR1 requires the core of its BTB/POZ domain and the oxidation of C-terminal cysteines. Plant Cell 18, 3670–3685. doi: 10.1105/tpc.106.046953
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Rodriguez, F. I., Esch, J. J., Hall, A. E., Binder, B. M., Schaller, G. E., and Bleecker, A. B. (1999). A copper cofactor for the ethylene receptor ETR1 from Arabidopsis. Science 283, 996–998. doi: 10.1126/science.283.5404.996
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Santner, A., and Estelle, M. (2009). Recent advances and emerging trends in plant hormone signalling. Nature 459, 1071–1078. doi: 10.1038/nature08122
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Shi, Z., Maximova, S., Liu, Y., Verica, J., and Guiltinan, M. J. (2013). The salicylic acid receptor NPR3 is a negative regulator of the transcriptional defense response during early flower development in Arabidopsis. Mol. Plant 6, 802–816. doi: 10.1093/mp/sss091
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Subramaniam, R., Desveaux, D., Spickler, C., Michnick, S. W., and Brisson, N. (2001). Direct visualization of protein interactions in plant cells. Nat. Biotechnol. 19, 769–772. doi: 10.1038/90831
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Vlot, A. C., Dempsey, D. A., and Klessig, D. F. (2009). Salicylic Acid, a multifaceted hormone to combat disease. Annu. Rev. Phytopathol. 47, 177–206. doi: 10.1146/annurev.phyto.050908.135202
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Wu, Y., Zhang, D., Chu, J. Y., Boyle, P., Wang, Y., Brindle, I. D., et al. (2012). The Arabidopsis NPR1 protein is a receptor for the plant defense hormone salicylic acid. Cell Rep. 1, 639–647. doi: 10.1016/j.celrep.2012.05.008
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Zhang, Y., Cheng, Y. T., Qu, N., Zhao, Q., Bi, D., and Li, X. (2006). Negative regulation of defense responses in Arabidopsis by two NPR1 paralogs. Plant J. 48, 647–656. doi: 10.1111/j.1365-313X.2006.02903.x
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Zhang, Y., Fan, W., Kinkema, M., Li, X., and Dong, X. (1999). Interaction of NPR1 with basic leucine zipper protein transcription factors that bind sequences required for salicylic acid induction of the PR1 gene. Proc. Natl. Acad. Sci. U.S.A. 96, 6523–6528. doi: 10.1073/pnas.96.11.6523
Zhang, Y., Tessaro, M. J., Lassner, M., and Li, X. (2003). Knockout analysis of Arabidopsis transcription factors TGA2, TGA5, and TGA6 reveals their redundant and essential roles in systemic acquired resistance. Plant Cell 15, 2647–2653. doi: 10.1105/tpc.014894
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Keywords: NPR1, Arabidopsis, salicylic acid, receptor, plant immunity
Citation: Kuai X, MacLeod BJ, and Després C (2015) Integrating data on the Arabidopsis NPR1/NPR3/NPR4 salicylic acid receptors; a differentiating argument. Front. Plant Sci. 6:235. doi: 10.3389/fpls.2015.00235
Received: 15 January 2015; Accepted: 24 March 2015;
Published online: 10 April 2015
Edited by:
Loreto Holuigue, Pontificia Universidad Católica de Chile, ChileReviewed by:
Kemal Kazan, Commonwealth Scientific and Industrial Research Organization, AustraliaCopyright © 2015 Kuai, MacLeod, and Després. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Charles Després, Department of Biological Sciences, Brock University, St. Catharines, ON L2S 3A1, CanadaY2Rlc3ByZXNAYnJvY2t1LmNh
†These authors have contributed equally for this work.
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.