
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
MINI REVIEW article
Front. Plant Sci. , 07 April 2015
Sec. Plant Physiology
Volume 6 - 2015 | https://doi.org/10.3389/fpls.2015.00210
This article is part of the Research Topic Frontiers of Sulfur Metabolism in Plant Growth, Development, and Stress Response View all 35 articles
 Naser A. Anjum1
Naser A. Anjum1 Ritu Gill2Manjeri Kaushik2
Ritu Gill2Manjeri Kaushik2 Mirza Hasanuzzaman3Eduarda Pereira1
Mirza Hasanuzzaman3Eduarda Pereira1 Iqbal Ahmad1,4
Iqbal Ahmad1,4 Narendra Tuteja5
Narendra Tuteja5 Sarvajeet S. Gill2*
Sarvajeet S. Gill2*
Sulfur (S) stands fourth in the list of major plant nutrients after N, P, and K. Sulfate (SO42-), a form of soil-S taken up by plant roots is metabolically inert. As the first committed step of S-assimilation, ATP-sulfurylase (ATP-S) catalyzes SO42--activation and yields activated high-energy compound adenosine-5′-phosphosulfate that is reduced to sulfide (S2-) and incorporated into cysteine (Cys). In turn, Cys acts as a precursor or donor of reduced S for a range of S-compounds such as methionine (Met), glutathione (GSH), homo-GSH (h-GSH), and phytochelatins (PCs). Among S-compounds, GSH, h-GSH, and PCs are known for their involvement in plant tolerance to varied abiotic stresses, Cys is a major component of GSH, h-GSH, and PCs; whereas, several key stress-metabolites such as ethylene, are controlled by Met through its first metabolite S-adenosylmethionine. With the major aim of briefly highlighting S-compound-mediated role of ATP-S in plant stress tolerance, this paper: (a) overviews ATP-S structure/chemistry and occurrence, (b) appraises recent literature available on ATP-S roles and regulations, and underlying mechanisms in plant abiotic and biotic stress tolerance, (c) summarizes ATP-S-intrinsic regulation by major S-compounds, and (d) highlights major open-questions in the present context. Future research in the current direction can be devised based on the discussion outcomes.
Abiotic and biotic stresses (in isolation and/or combination) are known to cause severe decline in crop productivity globally as a result of their impact on plant growth, development, and metabolism (Suzuki et al., 2014). Maintenance of plant-mineral nutrients status has been extensively evidenced to significantly improve the crop-productivity and -resistance to various stresses (Anjum and Lopez-Lauri, 2011; Gill and Tuteja, 2011). Sulfur (S) stands fourth in the list of major plant-nutrients after N, P, and K, and its importance is being increasingly emphasized in agriculture (Yi et al., 2010) and plant stress tolerance (Gill and Tuteja, 2011; Nazar et al., 2011). Nevertheless, S-deficiency in agricultural-soils is becoming widespread globally (Anjum et al., 2012a). Thus far, adopted approaches such as increased S-fertilization, -remobilization, and -uptake/accumulation may not be sufficient for S-deficiency-alleviation. Nevertheless, plant harbored-S is metabolically inert and is of no significance if it is not efficiently assimilated into physiologically/biochemically exploitable organic forms that is performed by the process of S-assimilation.
As the first committed step of primary S-assimilation in plants, ATP-sulfurylase (ATP-S; Adenylsulfurylase/ATP:sulfate adenylyltransferase; E.C. 2.7.7.4) catalyzes the activation of sulfate (SO42-) and yields adenosine-5′-phosphosulfate (APS) that is reduced to sulfide (S2-) and incorporated into cysteine (Cys). Having thiol (S2-)-residue and due to its strong nucleophilic-characteristics, Cys performs important metabolic-functions and actively mediates redox-reactions (Hell and Wirtz, 2011). Notably, as a major component of predominant thiol-peptide found in plants and as a direct/indirect precursor, Cys is involved in the synthesis of S-containing compounds including glutathione (GSH, γ-glutamyl-cysteinyl-glycine) and its analog homo-GSH (h-GSH, γ-glutamyl-cysteinyl-β-Ala), reported in several genera within Fabaceae; phytochelatins (PCs; γ-glutamyl-cysteinyl)nx; n = 2-11; x represents (Gly, Ser, β-Ala, Glu, Gln, or no residue), and metallothioneins (MTs), Cys-rich gene-encoded low-molecular-weight peptides. Previous S-compounds are known for their involvement in plant-tolerance to varied abiotic–biotic stresses, and metal/metalloid-homeostasis as well (Rausch and Wachter, 2005; Verbruggen et al., 2009; Anjum et al., 2010, 2012b, 2014a,b; Na and Salt, 2011; Seth et al., 2012; Gill et al., 2013). Additionally, in secondary SO42--assimilation, where instead of entering the reductive S-assimilation pathway after ATP-S-mediated activation, APS is phosphorylated in a APS kinase-catalyzed reaction to produce 3′-phosphoadenosine 5′-phosphosulfate (PAPS). PAPS is involved in the production of other S-containing methionine-derived (aliphatic) or tryptophan-derived (indolic) secondary metabolites such as glucosinolates (GSs). GSs (particularly indolic type) are reported to protect plants mainly against several biotic stress-factors such as herbivory and pathogenesis, and are required for plant-immunity (Frerigmann and Gigolashvili, 2014). Therefore, S-assimilation pathway-enzymes including ATP-S are the major target of current plant-nutrition research to achieve maximum benefits including improved productivity of crops and their resistance to multiple stresses with less S-input (Herrmann et al., 2014).
Thus, to briefly highlight S-compound-mediated role of ATP-S in plant stress tolerance, ATP-S structure/chemistry and occurrence are overviewed, recent literature available on ATP-S roles, regulations and underlying major mechanisms in plant abiotic and biotic stress tolerance is appraised, ATP-S intrinsic regulation by major S-compounds is summarized, and important open-questions in the topic considered are highlighted herein.
Described as monomers or homo-oligomeric complexes (which do not require GTPase for activation), plant-ATP-S has been reported to be a homotetramer of 52–54 kDa polypeptides, or a mono-functional, non-allosteric homodimer (100 kDa, formed by two ∼48 kDa monomers; Phartiyal et al., 2006; Ravilious et al., 2013; Bohrer et al., 2014; Koprivova and Kopriva, 2014; Prioretti et al., 2014). Photosynthetic organisms can exhibit a variable number of ATP-S isoforms (Koprivova and Kopriva, 2014; Prioretti et al., 2014). X-ray crystal structure of Glycine max ATP-S isoform 1 in complex with APS revealed the exhibition of several highly conserved substrate-binding motifs in the active site and a distinct dimerization interface compared with other ATP-S (Herrmann et al., 2014). Enzymes involved in S-assimilation are not equally expressed in all plant cell-types/ organelles. In particular, ATP-S, APS kinase, serine acetyltransferase, and O-acetylserine-(thiol)-lyase are present in both plastids and cytosol but APS reductase and sulfite reductase are localized only in plastids for catalyzing the reduction steps (Lopez-Martin et al., 2008; Bohrer et al., 2014; Koprivova and Kopriva, 2014). Occurrence of SO42--activation in cytosol and plastids also supports the presence of ATP-S in these locations (Koprivova and Kopriva, 2014). Seed-plants possess multiple ATP-S-isoforms. Four ATP-S genes (ATPS1, -2, -3, and -4) reported in Arabidopsis thaliana have N′-terminal extensions typical of plastid-transit-peptides, and are located on different chromosomes; however, one of them can also be cytosolic (Rotte and Leustek, 2000; Prioretti et al., 2014). Genetic-identity of cytosolic-ATP-S has been verified recently (Bohrer et al., 2015). A. thaliana ATPS2 was evidenced to be dually encode plastidic and cytosolic forms, where translational-initiation at AUGMet1 and AUGMet52 or AUGMet58 produced ATPS2 in plastid and cytosol, respectively (Bohrer et al., 2015). Oryza sativa has two ATP-S genes (ATPS1-2; Kopriva et al., 2007). Plastidic and/or mitochondrial localization of ATP-S genes (Glyma10g38760, Glyma20g28980, Glyma13g06940; Glyma19g05020) was reported in G. max (Yi et al., 2010).
ATP-sulfurylase can be involved in plant-tolerance to several abiotic stresses via different S-compounds. GSH, a non-protein S-containing tripeptide acts as a storage and transport form of reduced-S. Significant induction of GSH-based defense-system, its role in reactive oxygen species (ROS)-scavenging, and in the maintenance of reduced cellular-redox environment have been extensively evidenced in plants under various abiotic stresses including metal/metalloids (Anjum et al., 2010, 2012b, 2014a,b; Gill and Tuteja, 2010; Noctor et al., 2012; Talukdar, 2012; Gill et al., 2013; Talukdar and Talukdar, 2014) and salinity (Ruiz and Blumwald, 2002; Kocsy et al., 2004; Gill and Tuteja, 2010; Table 1). Cys-rich metal-chelating proteins – MTs and PCs maintain homeostasis of varied metals/metalloids and mitigate major detrimental effects of their elevated concentrations (Na and Salt, 2011; Anjum et al., 2014a). h-GSH is an effective antioxidant in Fabaceae plants, where it is argued to scavenge ROS, act as PCs-precursor, and found to be involved in xenobiotic defenses via GSH-sulfotransferases (Frendo et al., 2013). GSs provide plant-tolerance to varied abiotic stresses including drought/salinity, metals/metalloids, and nutritional-deficiencies (Martínez-Ballesta et al., 2013).

TABLE 1. Summary of representative studies on ATP-S activity or expression modulation/regulation in abiotic and biotic stressed plants.
Varied abiotic stresses differentially regulate ATP-S activity/expression in plants (Table 1). Among metals/metalloids, literature is full on Cd-accrued enhanced ATP-S activity and increased pools of Cys and GSH (Guo et al., 2009; Khan et al., 2009a; Masood et al., 2012; Bashir et al., 2013; Asgher et al., 2014). Up-regulation of ATP-S transcripts was reported in Cd-exposed Brassica juncea (Heiss et al., 1999) and A. thaliana (Harada et al., 2002). Enhanced ATP-S activity was evidenced in several Cd/Zn-hyperaccumulators including Sedum alfredii (Guo et al., 2009), A. halleri (Weber et al., 2006), and Thlaspi caerulescens (van de Mortel et al., 2008). Lower ATP-S activity-exhibiting Brassica juncea cv. (SS2) was reported to be salt-sensitive (Khan et al., 2009b). Chilling-stress can also mediate modulation of levels and also intercellular-distribution of ATP-S mRNAs (Kopriva et al., 2001). Reports also indicate the ATP-S activity/expression-regulation by light-regimes. Forty four hours of dark was reported to down-regulate ATPS1–ATPS3; whereas, ATPS4 was not affected (Huseby et al., 2013). However, after 3-h of re-illumination, ATPS1, ATPS3, and ATPS4 were induced by light but only ATPS2 reached the levels in control plants (Huseby et al., 2013).
Unknown for its essential-function in higher plants, Se, taken-up as selenate (SeO2-4)/or selenite (SeO2-3) was reported to enhance plant growth and antioxidant activity (Pilon-Smits and Quinn, 2010). ATP-S is also involved in Se-reductive-assimilation pathway and activates SeO2-4 to organic-metabolite, seleno-Cys (El Kassis et al., 2007; Pilon-Smits and Quinn, 2010). Recently, ability to hyperaccumulate and hypertolerate Se in Stanleya pinnata (Se-hyperaccumulator) was considered due to its potential to exhibit higher transcript levels of APS1, APS2, and APS4 (vs. Brassica juncea, a non-Se-hyperaccumulator; Schiavon et al., 2015). Additionally, under Se-exposure and S-deficiency, S. pinnata hyperaccumulates and tolerates Se due to its ability to convert SeO2-4 to non-toxic organic-seleno-compounds by down-regulating APS1, APS2, and APS4. However, under S-sufficient and Se-exposure, adoption of different types of regulatory mechanisms and subcellular-localization were revealed in S. pinnata and Brassica juncea, where Se up-regulated APS1 and APS4 but was not able to affect APS2 in S. pinnata (Schiavon et al., 2015). Earlier, compared to Camellia sinensis grown on Se un-enriched soil, young (or mature) leaves and roots were reported to exhibit a lower and higher APS1 and APS2 expression levels in Se-enriched soil-grown C. sinensis (Tao et al., 2012).
Extensive reports are available on S-depletion-mediated regulation of ATP-S activity/expression. ATP-S isoforms can be differentially expressed by S-depletion. AtAPS3 increased in S-deprived A. thaliana (Liang et al., 2010; Kawashima et al., 2011). However, response of AtAPS2 (a putative cytosolic-isoform) to S-depletion is inconsistent between different studies (Logan et al., 1996; Takahashi et al., 1997; Kawashima et al., 2011). Plant-ontogeny/developmental-stages can also modulate ATP-S-activity/expression under S-depleted conditions (Rotte and Leustek, 2000; Honsel et al., 2012). Confirmed by ATP-S protein-immunoblotting, ATP-S-activity exhibited a linear, threefold decline between 14 and 61 days after germination in S-depleted A. thaliana (Rotte and Leustek, 2000). Compared to young leaves, higher transcript-levels of PtaATPS3/4 were reported in Populus tremula× Populus alba after 21 days of S-depletion (Honsel et al., 2012). Contrarily, S-depletion did not lead any change in PtaATPS1/2-expression in young leaves; whereas, this ATP-S isoform increasingly expressed after 9 days in mature leaves (Honsel et al., 2012). In A. thaliana, both S-deficiency (-S/-Cd) and Cd (+S/+Cd) regulated APT-S activity (Bashir et al., 2013).
ATP-S gene-regulation has been discussed in different SO42--starved plants. APS1, APS3, and APS4 genes can be targeted to regulate root-shoot-SO42--accumulation by miR395 (small conserved non-coding RNAs with 20–24 nucleotides, specific sizes, and dedicated functions; Liang and Yu, 2010; Liang et al., 2010). In APS4-RNAi transgenic A. thaliana, loss-of-function of APS1 or/and APS4-genes can lead to 5-times higher SO42--accumulation in shoot (vs. wild-type plants). Additionally, enhanced miR395-expression in the absence of APS4 was considered as an indicator of a negative-feedback-loop between miR395 and APS4 (Liang et al., 2010). Moreover, unlike APS1 and APS4-mRNA, both miR395 and APS3 can exhibit a similar response to SO42- starvation; however, APS1 and APS3-expression can be regulated via miR395 (Liang and Yu, 2010). MiRNA395 can also cleave mRNAs encoding ATPS1 and ATPS4-isoforms (Jones-Rhoades and Bartel, 2004; Kawashima et al., 2009). Nevertheless, ATPS1 and ATPS4 were evidenced as the major targets of miRNA (miR395), in both leaves and roots (Kawashima et al., 2009). In a similar report, SO42--limitation decreased ATPS4-transcript-levels; whereas, ATPS1 levels were unaffected (Kawashima et al., 2011). It was argued in previous and other studies that for the SO42--limitation-mediated decreased ATPS4-transcripts that ATPS4 can undergo a canonical regulation by miR395 because its mRNA levels can decrease following miR395-induction (Kawashima et al., 2009, 2011; Liang et al., 2010). SO42--deficiency cannot affect (Kawashima et al., 2011) or can bring a slight decrease in the levels of ATPS1 mRNA (Liang et al., 2010). ATP-S activity/expression can also be controlled/modulated by S-Limitation 1 (SLIM1), a TF identical to Ethylene-Insensitive3-Like (EIL3) TF in Arabidopsis and the regulator of many S-deficiency responsive genes (Wawrzynska and Sirko, 2014). ATP-S-relation with ethylene is supported by the role of EIN3 and EIL1, two members of EI3/EIL TF family as central regulators of ethylene signaling (Maruyama-Nakashita et al., 2006). Relation among ATP-S-activity, GSH-content, ethylene-level, and decreased Cd-impacts was reported in Se-supplemented Cd-exposed Triticum aestivum (Khan et al., 2015). Nevertheless, a joint action of miR395 and SLIM 1 TF can maintain optimal-levels of ATP-S-transcripts in S-starved plants (Kawashima et al., 2011).
Through different S-compounds such as Cys, GSH, and GSs, ATP-S is also involved in plant-tolerance to several biotic stresses. Free-Cys and cytosolic Cys-homeostasis can orchestrate plant-pathogen responses (Gullner and Kömives, 2001; Álvarez et al., 2012). Pathogen-infection can trigger accumulation of GSH and also the modulation of transient changes in its redox-state (Noctor et al., 2012). Elevated GSH and Cys were reported to suppress and delay virus-symptoms, and decrease virus-content in zucchini yellow mosaic virus (ZYMV)-infected Cucurbita pepo (Zechmann et al., 2005, 2007; Zechmann and Müller, 2008; Király et al., 2012). Decreased GSH-pool and its redox-state in Lycopersicon esculentum signify their role against Botrytis cinerea infection (Kuźniak and Skłodowska, 2005). Elevated GSH metabolism can also counteract infection in plants with tobacco mosaic virus (Höller et al., 2010; Király et al., 2012), Pseudomonas syringae (Großkinsky et al., 2012) and B. cinerea (Simon et al., 2013). Zechmann (2014) recently reviewed the compartment-specific importance of GSH in biotic stressed plants.
Evidences confirm the requirement of a certain level of GSH for disease-resistance via synthesis of pathogen defense-related molecules such as camalexin, an indole-phytoalexin containing one S-atom per molecule with partly Cys-derived thiazole-ring (Noctor et al., 2012). A link between GSH-deficiency and plant-susceptibility to pathogens such as Pieris brassicae was reported in A. thaliana phytoalexin deficient 2-1 (pad2-1) mutant (Dubreuil-Maurizi and Poinssot, 2012). Earlier, a higher susceptibility of previous GSH-deficient-mutant to insect-herbivore Spodoptera littoralis was related with a lower GSs-accumulation therein (Schlaeppi et al., 2008). GSH (and also numerous GSH-sulfotransferases) is required for wound-induced resistance to B. cinerea (Chassot et al., 2008; Consonni et al., 2010). Expression of defense-related genes including PATHOGENESIS-RELATED1 (PR1) can be activated by exogenous-GSH-mediated mimicking of fungal-elicitors (reviewed by Noctor et al., 2012). Inner chloroplast-envelope-transporters export GSH across the chloroplast envelope. In Arabidopsis, CLT1, CLT2, and CLT3 genes encode these transporters (Maughan et al., 2010). Decreased PR1-expression and also lower resistance to the oomycete Pieris brassicae were reported in CLTs-defective Arabidopsis-mutants (Maughan et al., 2010). Numerous reports support an increased S-requirement in plants infected with Phytopthora infestans and/or B. cinerea and was advocated to be met, at least in part, by increased transcription of ATPS1, ATPS3, and ATPS4 genes (Matthewman, 2010). To this end, in A. thaliana, ATP-S genes namely ATPS1 and ATPS3 were reported to be linked with the regulation of biosynthetic networks of aliphatic and indolic GSs, respectively (Yatusevich et al., 2010). P. infestans and B. cinerea-infection in A. thaliana resulted in a similar increase in the transcript levels of ATPS1, ATPS3 and ATPS4 (Matthewman, 2010). Earlier, B. cinerea, P. Infestans, and aphid Myzus persicae were reported to induce a GSs-response in Arabidopsis (Kim and Jander, 2007; Rowe et al., 2010).
Literature is scarce on insights into S-compounds-mediated regulation of ATP-S activity/expression in plants. Among the thiol-compounds, GSH, rather than Cys can be used as a signal for regulating ATP-S (Lappartient et al., 1999; Vauclare et al., 2002). Externally supplied GSH-mediated increase in Cys and GSH accumulation can control both ATP-S activity and SO42--uptake (Vauclare et al., 2002). Compared to its lower level (up to 1.0 mM), Cys can significantly decrease ATP-S-activity at its higher level (2.0 mM). However, further increase in Cys-concentration can cause an additional accumulation of GSH that in turn can cause a decrease in ATP-S-mRNA, -protein, and -activity (Lappartient et al., 1999; Vauclare et al., 2002). ATP-S enzymatic activity might be sensitive to redox regulation in plants, where it can be a target for thioredoxins (reviewed by Prioretti et al., 2014). As a major redox regulator, GSH feeds into glutaredoxin system and subsequently into the thiol-redox-network (Dietz, 2008). Referring to the studies of Lappartient and Touraine (1996, 1997), cellular-redox-conditions and also that of GSH were advocated to modulate ATP-S-activity (reviewed by Yi et al., 2010). However, the authors suggested further biochemical- and structural-analysis of ATP-S to determine how, and to what extent, ATP-S responds to redox-changes. MiR395 is related with ATP-S-genes such as APS1, APS3, and APS4 (Liang et al., 2010). Recently, GSH-supplementation was reported to block accumulation of S-deprivation-inducible miR395 in S-deprived A. thaliana (Jagadeeswaran et al., 2014). Declined GSH-pools and induced miR395-levels in S-deprived A. thaliana were cross-talked (Kawashima et al., 2011; Matthewman et al., 2012). Nevertheless, biosynthesis of indolic-GSs in A. thaliana is regulated by MYB34, MYB51, and MYB122 TFs (Frerigmann and Gigolashvili, 2014). In A. thaliana, expression of both ATPS1 and ATPS3 isoforms was reported to be controlled by all six GSs-related MYB TFs namely MYB28, MYB29, and MYB76; MYB51, MYB34, and MYB122 (Yatusevich et al., 2010). ATPS1 and ATPS3 were expected to be strongly associated with the control of synthesis of aliphatic and indolic GSs, respectively. A. thaliana overexpressing or disruption in MYB51-gene showed alterations in ATP-S-transcript-levels and -activity (Matthewman, 2010; Figure 1).
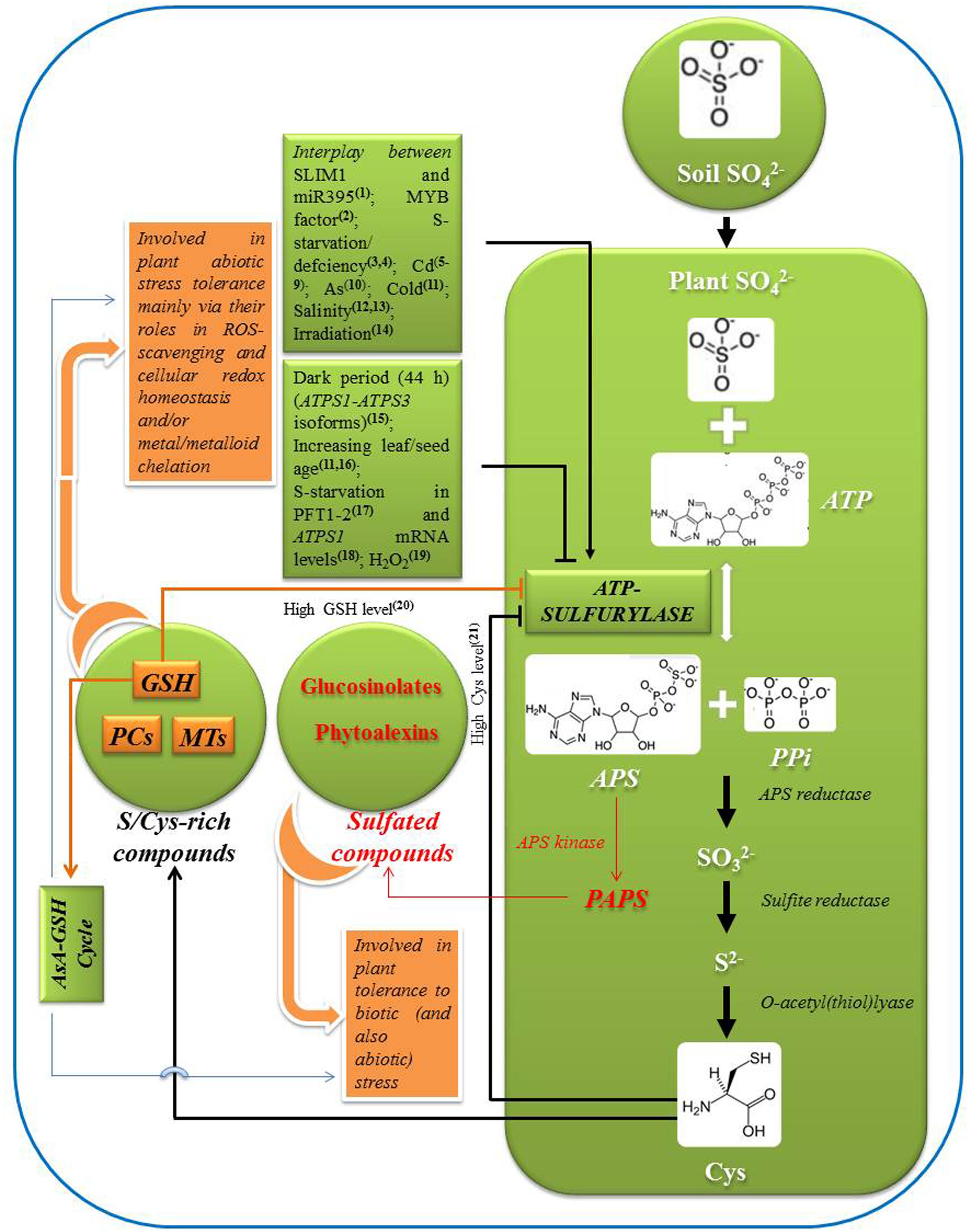
FIGURE 1. Schematic representation of pathway of sulfate assimilation, reaction catalyzed by ATP-sulfurylase (ATP-S), and its regulation by major factors. Role of ATP-S in plant stress tolerance through sulfur/cysteine rich and sulfated compounds is outlined. Positive and negative regulation of ATP-S is indicated by arrows and blunt ends, respectively, [1Kawashima et al. (2011); 2Yatusevich et al. (2010); 3Hopkins et al. (2004); 4Schiavon et al. (2007); 5van de Mortel et al. (2008); 6Guo et al. (2009); 7Gill et al. (2012); 8Bashir et al. (2013); 9Asgher et al. (2014); 10Leao et al. (2014); 11Phartiyal et al. (2006); 12Ruiz and Blumwald (2002); 13Nazar et al. (2011); 14Passera et al. (1989); 15Huseby et al. (2013); 16Rotte and Leustek (2000); 17Takahashi et al. (1997); 18Liang et al. (2010); 19Lappartient and Touraine (1997); 20Lappartient and Touraine (1996); 21Vauclare et al. (2002)]. (APS, adenosine 5′-phosphosulfate; Cys, cysteine; AsA, ascorbate; GSH, reduced glutathione; PCs, phytochelatins; MTs, metallothioneins; ROS, reactive oxygen species).
S-containing compounds such as Met, GSH, h-GSH, PCs, and GSs, directly or indirectly modulated/regulated by ATP-S are involved in plant tolerance to both biotic and abiotic stresses. Much has been achieved on the subject considered herein; there remain numerous aspects to be enlightened and open-questions to be answered. Ample scope exists for getting more molecular-genetic insights into the energetically unfavorable-reaction that yields APS from SO42- and ATP with ATP-S-catalytic-function. Notably, compared to APR enzyme and its encoding genes, much less amplitude and significance has been given to ATP-S in mutant-experiments. Hence, molecular-genetic dissection of so far neglected significance of ATP-S as a major control in the initial step of S-assimilation pathway is required. ATPS has been evidenced as an integral part of GS-biosynthesis-regulatory network (Matthewman, 2010); however, unveiling insights into interrelationship of ATP-S transcripts with other secondary S-assimilation products will be rewarding. Though picture is clear regarding the relationship of ATPS1 and ATPS3-expression with MYB TFs (Yatusevich et al., 2010) effort is required to unveil potential relationships of MYB TFs with ATPS2 and ATPS4-expression (Prioretti et al., 2014). If done, these studies may shed light on the complexity of regulatory interactions between primary and secondary S-metabolism. Efforts are also required to dissect the molecular biology/genetics of interaction of ATP-S with ratios of oxidized and reduced GSH (GSSG/GSH) and that of oxidized (dehydroascorbate, DHA) and reduced ascorbate (AsA; DHA/AsA) in stressed plants since DHA can be recycled back to AsA at the expense of GSH (or NADPH) by the AsA–GSH cycle-enzymes (Anjum et al., 2010). Role of miR395 family of micro-RNAs in the regulation of ATPS1-4 is known (Maruyama-Nakashita et al., 2006; Kawashima et al., 2011); however, picture is unclear in context with functions and effects of miR395 on ATPS3 and ATPS4-expression. A cross-talk among GSH-pools, miR395-levels and ATP-S-transcripts/activity particularly under deprived condition of interdependent nutrients S and N can also be significant for the maintenance of the status of S-compounds, and S-N homeostasis.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
NA is grateful to the Portuguese Foundation for Science and Technology (FCT) financial support in the form of post-doctoral research grants (SFRH/BPD/64690/2009; SFRH/BPD/84671/2012). SG and RG acknowledge the funds from DST-SERB, CSIR, and UGC, Government of India, New Delhi. Authors apologize if some references related to the main theme of the current article could not be cited due to space constraint.
Álvarez, C., Bermúdez, M. A., Romero, L. C., Gotor, C., and García, I. (2012). Cysteine homeostasis plays an essential role in plant immunity. New Phytol. 193, 165–177. doi: 10.1111/j.1469-8137.2011.03889.x
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Anjum, N. A., Gill, S. S., and Gill, R. (2014a). “Cysteine – jack of all glutathione-based plant stress defense trades,” in Plant Adaptation to Environmental Change: Significance of Amino Acids and their Derivatives, eds N. A. Anjum, S. S. Gill, and R. Gill (Wallingford: CAB International), 35–52. doi: 10.1079/9781780642734.0035
Anjum, N. A., Aref, I. M., Duarte, A. C., Pereira, E., Ahmad, I., and Iqbal, M. (2014b). Glutathione and proline can coordinately make plants withstand the joint attack of metal(loid) and salinity stresses. Front. Plant Sci. 5:662. doi: 10.3389/fpls.2014.00662
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Anjum, N. A., Gill, S. S., Umar, S., Ahmad, I., Duarte, A. C., and Pereira, E. (2012a). Improving growth and productivity of oleiferous Brassicas under changing environment: significance of nitrogen and sulphur nutrition, and underlying mechanisms. Sci. World J. 2012:657808. doi: 10.1100/2012/657808
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Anjum, N. A., Ahmad, I., Mohmood, I., Pacheco, M., Duarte, A. C., Pereira, E., et al. (2012b). Modulation of glutathione and its related enzymes in plants’ responses to toxic metals and metalloids -a review. Environ. Exp. Bot. 75, 307–324. doi: 10.1016/j.envexpbot.2011.07.002
Anjum, N. A., and Lopez-Lauri, F. (2011). Plant Nutrition and Abiotic Stress Tolerance III. Ikenobe: Global Science Books.
Anjum, N. A., Umar, S., and Chan, M. T. (2010). Ascorbate-Glutathione Pathway and Stress Tolerance in Plants. Dordrecht: Springer. doi: 10.1007/978-90-481-9404-9
Asgher, M., Khan, N. A., Khan, M. I. R., Fatma, M., and Masood, A. (2014). Ethylene production is associated with alleviation of cadmium-induced oxidative stress by sulfur in mustard types differing in ethylene sensitivity. Ecotoxicol. Environ. Saf. 106, 54–61. doi: 10.1016/j.ecoenv.2014.04.017
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Bashir, H., Ahmad, J., Bagheri, R., Nauman, M., and Qureshi, M. I. (2013). Limited sulfur resource forces Arabidopsis thaliana to shift towards non-sulfur tolerance under cadmium stress. Environ. Exp. Bot. 94, 19–32. doi: 10.1016/j.envexpbot.2012.05.004
Bohrer, A., Kopriva, S., and Takahashi, H. (2014). Plastid-cytosol partitioning and integration of metabolic pathways for APS/PAPS biosynthesis in Arabidopsis thaliana. Front. Plant Sci. 5:751. doi: 10.3389/fpls.2014.00751
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Bohrer, A. S., Yoshimoto, N., Sekiguchi, A., Rykulski, N., Saito, K., and Takahashi, H. (2015). Alternative translational initiation of ATP sulfurylase underlying dual localization of sulfate assimilation pathways in plastids and cytosol in Arabidopsis thaliana. Front. Plant Sci. 5:750. doi: 10.3389/fpls.2014.00750
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Brunner, M., Kocsy, G., Rüegsegger, A., Schmutz, D., and Brunold, C. (1995). Effect of chilling on assimilatory sulfate reduction and glutathione synthesis in maize. J. Plant Physiol. 146, 743–747. doi: 10.1016/S0176-1617(11)81942-5
Chassot, C., Buchala, A., Schoonbeek, H. J., Metraux, J. P., and Lamotte, O. (2008). Wounding of Arabidopsis leaves causes a powerful but transient protection against Botrytis infection. Plant J. 55, 555–567. doi: 10.1111/j.1365-313X.2008.03540.x
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Consonni, C., Bednarek, P., Humphry, M., Francocci, F., Ferrari, S., Harzen, A., et al. (2010). Tryptophan-derived metabolites are required for antifungal defense in the Arabidopsis mlo2 mutant. Plant Physiol. 152, 1544–1561. doi: 10.1104/pp.109.147660
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Dietz, K. J. (2008). Redox signal integration: from stimulus to networks and genes. Physiol. Plant 133, 459–468. doi: 10.1111/j.1399-3054.2008.01120.x
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Dubreuil-Maurizi, C., and Poinssot, B. (2012). Role of glutathione in plant signaling under biotic stress. Plant Signal. Behav. 7, 210–212. doi: 10.4161/psb.18831
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
El Kassis, E., Cathala, N., Rouached, H., Fourcroy, P., Berthomieu, P., Terry, N., et al. (2007). Characterization of a selenate-resistant Arabidopsis mutant. Root growth as a potential target for selenate toxicity. Plant Physiol. 143, 1231–1241. doi: 10.1104/pp.106.091462
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Frendo, P., Matamoros, M. A., Alloing, G., and Becana, M. (2013). Thiol-based redox signaling in the nitrogen-fixing symbiosis. Front. Plant Sci. 4:376. doi: 10.3389/fpls.2013.00376
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Frerigmann, H., and Gigolashvili, T. (2014). MYB34, MYB51, and MYB122 distinctly regulate indolic glucosinolate biosynthesis in Arabidopsis thaliana. Mol. Plant 7, 814–828. doi: 10.1093/mp/ssu004
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Gill, S. S., Anjum, N. A., Hasanuzzaman, M., Gill, R., Trivedi, D. K., Ahmad, I., et al. (2013). Glutathione and glutathione reductase: a boon in disguise for plant abiotic stress defense operations. Plant Physiol. Biochem. 70, 204–212. doi: 10.1016/j.plaphy.2013.05.032
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Gill, S. S., Khan, N. A., and Tuteja, N. (2012). Cadmium at high dose perturbs growth, photosynthesis and nitrogen metabolism while at low dose it up regulates sulfur assimilation and antioxidant machinery in garden cress (Lepidium sativum L.). Plant Sci. 182, 112–120. doi: 10.1016/j.plantsci.2011.04.018
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Gill, S. S., and Tuteja, N. (2010). Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiol. Biochem. 48, 909–930. doi: 10.1016/j.plaphy.2010.08.016
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Gill, S. S., and Tuteja, N. (2011). Cadmium stress tolerance in crop plants probing the role of sulphur. Plant Signal. Behav. 6, 215–222. doi: 10.4161/psb.6.2.14880
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Großkinsky, D. K., Koffler, B. E., Roitsch, T., Maier, R., and Zechmann, B. (2012). Compartment-specific antioxidative defense in Arabidopsis against virulent and avirulent Pseudomonas syringae. Phytopathology 102, 662–673. doi: 10.1094/PHYTO-02-12-0022-R
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Gullner, G., and Kömives, T. (2001). “The role of glutathione and glutathione-related enzymes in plant-pathogen interactions,” in Significance of Glutathione to Plant Adaptation to the Environment, eds D. Grill, M. Tausz, and L. De Kok (Houten: Springer), 207–239. doi: 10.1007/0-306-47644-4_9
Guo, W. D., Liang, J., Yang, X. E., Chao, Y. E., and Feng, Y. (2009). Response of ATP sulfurylase and serine acetyltransferase towards cadmium in hyperaccumulator Sedum alfredii Hance. J. Zhejiang Univ. Sci. B 10, 251–257. doi: 10.1631/jzus.B0820169
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Harada, E., Yamaguchi, Y., Koizumi, N., and Hiroshi, S. (2002). Cadmium stress induces production of thiol compounds and transcripts for enzymes involved in sulfur assimilation pathways in Arabidopsis. J. Plant Physiol. 159, 445–448. doi: 10.1078/0176-1617-00733
Heiss, S., Schäfer, H. J., Haag-Kerwer, A., and Rausch, T. (1999). Cloning sulfur assimilation genes of Brassica juncea L.: cadmium differentially affects the expression of a putative low-affinity sulfate transporter and isoforms of ATP sulfurylase and APS reductase. Plant Mol. Boil. 39, 847–857. doi: 10.1023/A:1006169717355
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Hell, R., and Wirtz, M. (2011). Molecular biology, biochemistry and cellular physiology of cysteine metabolism in Arabidopsis thaliana. Arabidopsis Book 9:e0154. doi: 10.1199/tab.0154
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Herrmann, J., Ravilious, G. E., McKinney, S. E., Westfall, C. S., Lee, S. G., Baraniecka, P., et al. (2014). Structure and mechanism of soybean ATP sulfurylase and the committed step in plant sulfur assimilation. J. Biol. Chem. 289, 10919–10929. doi: 10.1074/jbc.M113.540401
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Höller, K., Király, L., Künstler, A., Müller, M., Gullner, G., Fattinger, M., et al. (2010). Enhanced glutathione metabolism is correlated with sulfur-induced resistance in tobacco mosaic virus-infected genetically susceptible Nicotiana tabacum plants. Mol. Plant Microbe Interact. 23, 1448–1459. doi: 10.1094/MPMI-05-10-0117
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Honsel, A., Kojima, M., Haas, R., Frank, W., Sakakibara, H., Herschbach, C., et al. (2012). Sulphur limitation and early sulphur deficiency responses in poplar: significance of gene expression, metabolites, and plant hormones. J. Exp. Bot. 63, 1873–1893. doi: 10.1093/jxb/err365
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Hopkins, L., Parmar, S., Bouranis, D. L., Howarth, J. R., and Hawkesford, M. J. (2004). Coordinated expression of sulfate uptake and components of the sulfate assimilatory pathway in maize. Plant Biol. 6, 408–414. doi: 10.1055/s-2004-820872
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Huseby, S., Koprivova, A., Lee, B. R., Saha, S., Mithen, R., Wold, A. B., et al. (2013). Diurnal and light regulation of sulphur assimilation and glucosinolate biosynthesis in Arabidopsis. J. Exp. Bot. 64, 1039–1048. doi: 10.1093/jxb/ers378
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Jagadeeswaran, G., Li, Y. F., and Sunkar, R. (2014). Redox signaling mediates the expression of a sulfate-deprivation-inducible microRNA395 in Arabidopsis. Plant J. 77, 85–96. doi: 10.1111/tpj.12364
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Jones-Rhoades, M. W., and Bartel, D. P. (2004). Computational identification of plant micro-RNAs and their targets, including a stress-induced miRNA. Mol. Cell 14, 787–799. doi: 10.1016/j.molcel.2004.05.027
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Kawashima, C. G., Matthewman, C. A., Huang, S., Lee, B. R., Yoshimoto, N., Koprivova, A., et al. (2011). Interplay of SLIM1 and miR395 in the regulation of sulfate assimilation in Arabidopsis. Plant J. 66, 863–876. doi: 10.1111/j.1365-313X.2011.04547.x
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Kawashima, C. G., Yoshimoto, N., Maruyama-Nakashita, A., Tsuchiya, Y. N., Saito, K., Takahashi, H., et al. (2009). Sulfur starvation induces the expression of microRNA-395 and one of its target genes but in different cell types. Plant J. 57, 313–321. doi: 10.1111/j.1365-313X.2008.03690.x
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Khan, M. I. R., Nazir, F., Asgher, M., Per, T. S., and Khan, N. A. (2015). Selenium and sulfur influence ethylene formation and alleviate cadmium-induced oxidative stress by improving proline and glutathione production in wheat. J. Plant Physiol. 173, 9–18. doi: 10.1016/j.jplph.2014.09.011
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Khan, N. A., Anjum, N. A., Nazar, R., and Iqbal, N. (2009a). Increased activity of ATP-sulfurylase and increased contents of cysteine and glutathione reduce high cadmium-induced oxidative stress in mustard cultivar with high photosynthetic potential. Russ. J. Plant Physiol. 56, 670–677. doi: 10.1134/S1021443709050136
Khan, N. A., Nazar, R., and Anjum, N. A. (2009b). Growth, photosynthesis and antioxidant metabolism in mustard (Brassica juncea L.) cultivars differing in ATP-sulfurylase activity under salinity stress. Sci. Hortic. 122, 455–460. doi: 10.1016/j.scienta.2009.05.020
Khan, N. A., Samiullah, Singh, S., and Nazar, R. (2007). Activities of antioxidative enzymes, sulphur assimilation, photosynthetic activity and growth of wheat (Triticum aestivum) cultivars differing in yield potential under cadmium stress. J. Agron. Crop Sci. 193, 435–444. doi: 10.1111/j.1439-037X.2007.00272.x
Kim, J. H., and Jander, G. (2007). Myzus persicae (green peach aphid) feeding on Arabidopsis induces the formation of a deterrent indole glucosinolate. Plant J. 49, 1008–1019. doi: 10.1111/j.1365-313X.2006.03019.x
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Király, L., Künstler, A., Höller, K., Fattinger, M., Juhász, C., Müller, M., et al. (2012). Sulfate supply influences compartment specific glutathione metabolism and confers enhanced resistance to tobacco mosaic virus during a hypersensitive response. Plant Physiol. Biochem. 59, 44–54. doi: 10.1016/j.plaphy.2011.10.020
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Kocsy, G., Szalai, G., and Galiba, G. (2004). Effect of osmotic stress on glutathione and hydroxymethylglutathione accumulation in wheat. J. Plant Physiol. 161, 785–794. doi: 10.1016/j.jplph.2003.12.006
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Kopriva, S., Jones, S., Koprivova, A., Suter, M., von Ballmoos, P., Brander, K., et al. (2001). Influence of chilling stress on the intercellular distribution of assimilatory sulfate reduction and thiols in Zea mays. Plant Biol. 3, 24–31. doi: 10.1055/s-2001-11745
Kopriva, S., Wiedemann, G., and Reski, R. (2007). Sulfate assimilation in basal land plants-what does genomic sequencing tell us? Plant Biol. 9, 556–564. doi: 10.1055/s-2007-965430
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Koprivova, A., and Kopriva, S. (2014). Molecular mechanisms of regulation of sulfate assimilation: first steps on a long road. Front. Plant Sci. 5:589. doi: 10.3389/fpls.2014.00589
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Kuźniak, E., and Skłodowska, M. (2005). Compartment-specific role of the ascorbate-glutathione cycle in the response of tomato leaf cells to Botrytis cinerea infection. J. Exp. Bot. 56, 921–933. doi: 10.1093/jxb/eri086
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Lappartient, A. G., and Touraine, B. (1996). Demand-driven control of root ATP sulfurylase activity and SO42–uptake in intact canola. The role of phloem-translocated glutathione. Plant Physiol. 111, 147–157.
Lappartient, A. G., and Touraine, B. (1997). Glutathione-mediated regulation of ATP sulfurylase activity, SO42–uptake, and oxidative stress response in intact canola roots. Plant Physiol. 114, 177–183.
Lappartient, A. G., Vidmar, J. J., Leustek, T., Glass, A. D. M., and Touraine, B. (1999). Inter-organ signaling in plants: regulation of ATP sulfurylase and sulfate transporter genes expression in roots mediated by phloem-translocated compound. Plant J. 18, 89–95. doi: 10.1046/j.1365-313X.1999.00416.x
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Leao, G. A., Oliveira, J. A., Farnese, F. S., Gusman, G. S., and Felipe, R. T. A. (2014). Sulfurmetabolism: different tolerances of two aquatic macrophytes exposed to arsenic. Ecotoxicol. Environ. Saf. 105, 36–42. doi: 10.1016/j.ecoenv.2014.03.011
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Lee, M. S., and Leustek, T. (1999). Identification of the gene encoding homoserine kinase from Arabidopsis thaliana and characterization of the recombinant enzyme derived from the gene. Arch. Biochem. Biophys. 372, 135–142. doi: 10.1006/abbi.1999.1481
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Liang, G., Yang, F., and Yu, D. (2010). MicroRNA395 mediates regulation of sulfate accumulation and allocation in Arabidopsis thaliana. Plant J. 62, 1046–1057. doi: 10.1111/j.1365-313X.2010.04216.x
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Liang, G., and Yu, D. (2010). Reciprocal regulation among miR395, APS and SULTR2;1 in Arabidopsis thaliana. Plant Signal. Behav. 5, 1257–1259. doi: 10.4161/psb.5.10.12608
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Logan, H. M., Cathala, N., Grignon, C., and Davidian, J. C. (1996). Cloning of a cDNA encoded by a member of the Arabidopsis thaliana ATP sulfurylase multigene family. Expression studies in yeast and in relation to plant sulfur nutrition. J. Biol. Chem. 271, 12227–12233. doi: 10.1074/jbc.271.21.12227
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Lopez-Martin, M. C., Romero, L. C., and Gotor, C. (2008). Cytosolic cysteine in redox signaling. Plant Signal. Behav. 3, 880–881. doi: 10.1104/pp.108.117408
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Martínez-Ballesta, M. C., Moreno, D. A., and Carvajal, M. (2013). The physiological importance of glu-cosinolates on plant response to abiotic stress in Brassica. Int. J. Mol. Sci. 14, 11607–11625. doi: 10.3390/ijms140611607
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Maruyama-Nakashita, A., Nakamura, Y., Tohge, T., Saito, K., and Takahashi, H. (2006). Arabidopsis SLIM1 is a central transcriptional regulator of plant sulfur response and metabolism. Plant Cell 18, 3235–3251. doi: 10.1105/tpc.106.046458
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Masood, A., Iqbal, N., and Khan, N. A. (2012). Role of ethylene in alleviation of cadmium-induced photosynthetic capacity inhibition by sulphur in mustard. Plant Cell Environ. 35, 524–533. doi: 10.1111/j.1365-3040.2011.02432.x
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Matthewman, C. A. (2010). Arabidopsis ATP Sulfurylase: Roles and Regulation of Individual Isoforms. Ph.D. thesis, submitted to the University of East Anglia, Norwich.
Matthewman, C. A., Kawashima, C. G., Húska, D., Csorba, T., Dalmay, T., and Kopriva, S. (2012). miR395 is a general component of the sulfate assimilation regulatory network in Arabidopsis. FEBS Lett. 586, 3242–3248. doi: 10.1016/j.febslet.2012.06.044
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Maughan, S. C., Pasternak, M., Cairns, N., Kiddle, G., Brach, T., Jarvis, R., et al. (2010). Plant homologs of the Plasmodium falciparum chloroquinoneresistance transporter, PfCRT, are required for glutathione homeostasis and stress responses. Proc. Natl. Acad. Sci. U.S.A. 107, 2331–2336. doi: 10.1073/pnas.0913689107
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Na, G., and Salt, D. E. (2011). The role of sulfur assimilation and sulfur-containing compounds in trace element homeostasis in plants. Environ. Exp. Bot. 72, 18–25. doi: 10.1016/j.envexpbot.2010.04.004
Nazar, R., Iqbal, N., Masood, A., Syeed, S., and Khan, N. A. (2011). Understanding the significance of sulfur in improving salinity tolerance in plants. Environ. Exp. Bot. 70, 80–87. doi: 10.1016/j.envexpbot.2010.09.011
Noctor, G., Mhamdi, A., Chaouch, S., Han, Y. I., Neukermans, J., Marquez-Garcia, B., et al. (2012). Glutathione in plants: an integrated overview. Plant Cell Environ. 35, 454–484. doi: 10.1111/j.1365-3040.2011.02400.x
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Nussbaum, S., Schmutz, D., and Brunold, C. (1988). Regulation of assimilatory sulfate reduction by cadmium in Zea mays L. Plant Physiol. 88, 1407–1410. doi: 10.1104/pp.88.4.1407
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Passera, C., Ghisi, R., and Ferretti, M. (1989). Light-activation of ATP-sulfurylase in leaves and chloroplasts of Zea mays. Photosynthetica 23, 166–172.
Phartiyal, P., Kim, W. S., Cahoon, R. E., Jez, J. M., and Krishnan, H. B. (2006). Soybean ATP sulfurylase, a homodimeric enzyme involved in sulfur assimilation, is abundantly expressed in roots and induced by cold treatment. Arch. Biochem. Biophys. 450, 20–29. doi: 10.1016/j.abb.2006.03.033
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Pilon-Smits, E. A. H., and Quinn, C. F. (2010). “Selenium metabolism in plants,” in Cell Biology of Metal and Nutrients, eds R. Hell and R. Mendel (Berlin: Springer), 225–241. doi: 10.1007/978-3-642-10613-2_10
Prioretti, L., Gontero, B., Hell, R., and Giordano, M. (2014). Diversity and regulation of ATP sulfurylase in photosynthetic organisms. Front. Plant Sci. 5:597. doi: 10.3389/fpls.2014.00597
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Rausch, T., and Wachter, A. (2005). Sulfur metabolism: a versatile platform for launching defence operations. Trends Plant Sci. 10, 503–509. doi: 10.1016/j.tplants.2005.08.006
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Ravilious, G. E., Herrmann, J., Goo Lee, S., Westfall, C. S., and Jez, J. M. (2013). Kinetic mechanism of the dimeric ATP sulfurylase from plants. Biosci. Rep. 33:e00053. doi: 10.1042/BSR20130073
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Reuveny, Z., Dougall, D. K., and Trinity, P. M. (1980). Regulatory coupling of nitrate and sulfate assimilation pathways in cultured tobacco cells. Proc. Natl. Acad. Sci. U.S.A. 77, 6670–6672. doi: 10.1073/pnas.77.11.6670
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Rotte, C., and Leustek, T. (2000). Differential subcellular localization and expression of ATP sulfurylase and APS reductase during ontogenesis of Arabidopsis thaliana leaves indicates that cytosolic and plastid forms of ATP sulfurylase may have specialized functions. Plant Physiol. 124, 715–724. doi: 10.1104/pp.124.2.715
Rowe, H. C., Walley, J. W., Corwin, J., Chan, E. K. F., Dehesh, K., and Kliebenstein, D. J. (2010). Deficiencies in jasmonate-mediated plant defense reveal quantitative variation in Botrytis cinerea pathogenesis. PLoS Pathog. 6:e1000861. doi: 10.1371/journal.ppat.1000861
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Ruiz, J., and Blumwald, E. (2002). Salinity-induced glutathione synthesis in Brassica napus. Planta 214, 965–969. doi: 10.1007/s00425-002-0748-y
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Schiavon, M., Pilon, M., Malagoli, M., and Pilon-Smits, E. A. H. (2015). Exploring the importance of sulfate transporters and ATP sulphurylases for selenium hyperaccumulation -a comparison of Stanleya pinnata and Brassica juncea (Brassicaceae). Front. Plant Sci. 6:2. doi: 10.3389/fpls.2015.00002
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Schiavon, M., Wirtz, M., Borsa, P., Quaggiotti, S., Hell, R., and Malagoli, M. (2007). Chromate differentially affects the expression of a high-affinity sulfate transporter and isoforms of components of the sulfate assimilatory pathway in Zea mays (L.). Plant Biol. 9, 662–671. doi: 10.1055/s-2007-965440
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Schlaeppi, K., Bodenhausen, N., Buchala, A., Mauch, F., and Reymond, P. (2008). The glutathione-deficient mutant pad2-1 accumulates lower amounts of glucosinolates and is more susceptible to the insect herbivore Spodoptera littoralis. Plant J. 55, 774–786. doi: 10.1111/j.1365-313X.2008.03545.x
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Seth, C. S., Remans, T., Keunen, E., Jozefczak, M., Gielen, H., Opdenakker, K., et al. (2012). Phytoextraction of toxic metals: a central role for glutathione. Plant Cell Environ. 35, 334–346. doi: 10.1111/j.1365-3040.2011.02338.x
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Simon, K. U., Polanschütz, L. M., Koffler, B. E., and Zechmann, B. (2013). High resolution imaging of temporal and spatial changes of subcellular ascorbate, glutathione and H2O2 distribution during Botrytis cinerea infection in Arabidopsis. PLoS ONE 8:e65811. doi: 10.1371/journal.pone.0065811
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Suzuki, N., Rivero, R. M., Shulaev, V., Blumwald, E., and Mittler, R. (2014). Abiotic and biotic stress combinations. New Phytol. 203, 32–43. doi: 10.1111/nph.12797
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Takahashi, H., Yamazaki, M., Sasakura, N., Watanabe, A., Leustek, T., de Almeida Engler, J., et al. (1997). Regulation of sulfur assimilation in higher plants: a sulfate transporter induced in sulfate-starved roots plays a central role in Arabidopsis thaliana. Proc. Natl. Acad. Sci. U.S.A. 94, 11102–11107. doi: 10.1073/pnas.94.20.11102
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Talukdar, D. (2012). An induced glutathione-deficient mutant in grass pea (Lathyrus sativus L.): modifications in plant morphology, alteration in antioxidant activities and increased sensitivity to cadmium. Biorem. Biodiv. Bioavail. 6, 75–86.
Talukdar, D., and Talukdar, T. (2014). Coordinated response of sulfate transport, cysteine biosynthesis, and glutathione-mediated antioxidant defense in lentil (Lens culinaris Medik.) genotypes exposed to arsenic. Protoplasma 251, 839–855. doi: 10.1007/s00709-013-0586-8
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Tao, S., Li, J., Gu, X., Wang, Y., Xia, Q., Qin, B., et al. (2012). Quantitative analysis of ATP sulfurylase and selenocysteine methyltransferase gene expression in different organs of tea plant (Camellia sinensis). Am. J. Plant Sci. 3, 51–59. doi: 10.4236/ajps.2012.31004
van de Mortel, J. E., Schat, H., Moerland, P. D., Ver Loren van Themaat, E., van der Ent, S., and Blankestijn, H.et al. (2008). Expression differences for genes involved in lignin, glutathione and sulphate metabolism in response to cadmium in Arabidopsis thaliana and the related Zn/Cd-hyperaccumulator Thlaspi caerulescens. Plant Cell Environ. 31, 301–324. doi: 10.1111/j.1365-3040.2007.01764.x
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Vauclare, P., Kopriva, S., Fell, D., Suter, M., Sticher, L., Von Ballmoos, P., et al. (2002). Flux control of sulphate assimilation in Arabidopsis thaliana: adenosine 5’-phosphosulphate reductase is more susceptible than ATP sulphurylase to negative control by thiols. Plant J. 31, 729–740. doi: 10.1046/j.1365-313X.2002.01391.x
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Verbruggen, N., Hermans, C., and Schat, H. (2009). Molecular mechanisms of metal hyperaccumulation in plants. New Phytol. 181, 759–776. doi: 10.1111/j.1469-8137.2008.02748.x
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Wawrzynska, A., and Sirko, A. (2014). To control and to be controlled - understanding the Arabidopsis SLIM1 function in sulfur deficiency through comprehensive investigation of the EIL protein family. Front. Plant Sci. 5:575. doi: 10.3389/fpls.2014.00575
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Weber, M., Trampczynska, A., and Clemens, S. (2006). Comparative transcriptome analysis of toxic metal responses in Arabidopsis thaliana and the Cd2+-hypertolerant facultative metallophyte Arabidopsis halleri. Plant Cell Environ. 29, 950–963. doi: 10.1111/j.1365-3040.2005.01479.x
Yatusevich, R., Mugford, S. G., Matthewman, C., Gigolashvili, T., Frerigmann, H., Delaney, S., et al. (2010). Genes of primary sulfate assimilation are part of the glucosinolate biosynthetic network in Arabidopsis thaliana. Plant J. 62, 1–11. doi: 10.1111/j.1365-313X.2009.04118.x
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Yi, H., Ravilious, G. E., Galant, A., Krishnan, H. B., and Jez, J. M. (2010). From sulfur to homoglutathione: thiol metabolism in soybean. Amino Acids 39, 963–978. doi: 10.1007/s00726-010-0572-9
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Zechmann, B. (2014). Compartment-specific importance of glutathione during abiotic and biotic stress. Front. Plant Sci. 5:566. doi: 10.3389/fpls.2014.00566
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Zechmann, B., and Müller, M. (2008). Effects of zucchini yellow mosaic virus infection on the subcellular distribution of glutathione and its precursors in a highly tolerant Cucurbita pepo cultivar. Botany 86, 1092–1100. doi: 10.1139/B08-048
Zechmann, B., Zellnig, G., and Müller, M. (2005). Changes in the subcellular distribution of glutathione during virus infection in Cucurbita pepo (L.). Plant Biol. 7, 49–57. doi: 10.1055/s-2004-830477
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Zechmann, B., Zellnig, G., Urbanek-Krajnc, A., and Müller, M. (2007). Artificial elevation of glutathione affects symptom development in ZYMV-infected Cucurbita pepo L. plants. Arch. Virol. 152, 747–762. doi: 10.1007/s00705-006-0880-2
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Keywords: ATP-sulfurylase, sulfur assimilation, organic S-compounds, stress tolerance
Citation: Anjum NA, Gill R, Kaushik M, Hasanuzzaman M, Pereira E, Ahmad I, Tuteja N and Gill SS (2015) ATP-sulfurylase, sulfur-compounds, and plant stress tolerance. Front. Plant Sci. 6:210. doi: 10.3389/fpls.2015.00210
Received: 03 November 2014; Accepted: 16 March 2015;
Published online: 07 April 2015.
Edited by:
Dibyendu Talukdar, University of Calcutta, IndiaReviewed by:
Ruediger Hell, University of Heidelberg, GermanyCopyright © 2015 Anjum, Gill, Kaushik, Hasanuzzaman, Pereira, Ahmad, Tuteja and Gill. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Sarvajeet S. Gill, Stress Physiology and Molecular Biology Lab, Centre for Biotechnology, Maharshi Dayanand University, Rohtak 124 001, Indiac3NnaWxsMTRAeWFob28uY28uaW4=
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.