 Frontiers in Plant Science Editorial Office*
Frontiers in Plant Science Editorial Office*A retraction of the Original Research Article:
Mechanosensitive channel candidate MCA2 is involved in touch-induced root responses in Arabidopsis
by Nakano, M., Samejima, R., and Iida, H. (2014). Front. Plant Sci. 5:421. doi: 10.3389/fpls.2014.00421
The authors and the journal wish to retract the 21 Aug 2014 article cited above in light of new experimental evidence.
Following the publication of the study, we performed a DNA microarray analysis to detect genes with expression levels that were specifically lower in the mca2-null mutant than in the Col-0 wild type, and found that the expression level of the AXR4 gene (At1G54990), which encodes a protein required for the subcellular localization of the auxin influx carrier AUX1 (Dharmasiri et al., 2006), was significantly lower in the mca2-null mutant. To confirm this finding, we then performed a semi-quantitative reverse transcription-PCR analysis using the primers axr4-f1 and axr4-Cr1 (Figure 1A), and found that the RT-PCR product was detectable in some mca2-null seedlings at wild-type levels but not in other mca2-null seedlings at all. This result suggested that some mca2-null seedlings have a certain lesion in the AXR4 locus, and a PCR-based genomic deletion analysis (Figures 1B,C) followed by DNA sequencing confirmed this speculation. Our conclusion is that most of the mca2-null seedlings used in the study presented in the above paper had a homozygous 2592-bp deletion that started from the intron between exons 1 and 2 of the AXR4 gene and reached the intron between exons 1 and 2 of the adjacent gene AT1G55000 (Figure 1A). Therefore, the phenotypes presented in the above paper may be ascribable to the axr4 mutation, the at1g55000 mutation, or both or even triple mutations, but not to the mca2-null mutation. The AT1G55000 gene encodes the peptidoglycan-binding LysM domain-containing protein involved in a macromolecule catabolic process in the cell wall1.
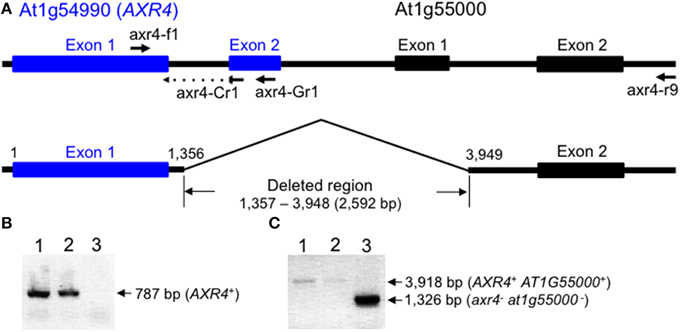
Figure 1. Schematic representation of the AXR4 locus and PCR data to check its integrity. (A) The AXR4 locus and its adjacent locus At1g55000 in Arabidopsis thaliana (ecotype Col-0). The upper line drawing shows intact loci, while the lower one shows loci with the 2592-bp deletion found in some mca2-null mutants. The position and orientation of the primers (axr4-f1, axr4-Cr1, axr4-Gr1, and axr4-r9) are presented by arrows. Nucleotide numbering starts from the 5′-end of exon 1 in the AXR4 gene. (B) An example of PCR data with the primers axr4-f1 and axr4-Gr1. (C) An example of PCR data with the primers arx4-f1 and axr4-r9. Genomic DNA was isolated from an mca2-null mutant with the mca2−/mca2− AXR4+/AXR4+ genotype (lane 1), the wild type with the MCA2+/MCA2+ AXR4+/AXR4+ genotype (lane 2), and another mca2-null mutant with the mca2−/mca2− axr4−/axr4− genotype (lane 3), and subjected to PCR followed by agarose gel electrophoresis. The nucleotide sequences of the primers axr4-f1, axr4-Cr1, axr4-Gr1, and axr4-r9 were 5′-GCAACGTGTAGCTAAGGCTCTTCC-3′, 5′-TCGCCGGATTTGCTTTCCTGAG-3′, 5′-GTAGTAGTCCATTCCTCACCAAG-3′, and 5′-AGCTCCATCTTCGTGTCTAG-3′, respectively, where the underlined nucleotide sequence is complementary to the coding strand of the 3′-terminal region of the exon 1 of the AXR4 gene. The PCR conditions used were as follows: initial activation of Taq DNA polymerase (Quick Taq™ HS DyeMix, TOYOBO, Osaka, Japan) at 94°C for 2 min, followed by 30 cycles of denaturation at 94°C for 30 s, annealing at 55°C for 30 s, and extension at 68°C for 2 min.
Our phenotypic study revealed that none of the mca2-null AXR4+ AT1G55000+ seedlings showed all the abnormal phenotypes reported in the above paper, regarding the skewing, waving, and bending responses of the root. In contrast, the seedlings of the axr4-1 (Ws-2 background) and axr4-2 (Col background) single mutants obtained from the Arabidopsis Biological Resource Center (ABRC germplasm names CS8018 and CS8019, respectively) showed the same abnormal phenotypes as those described in the above paper. We also confirmed that the abnormal phenotypes for the skewing, waving, and bending responses of the mca2-null arx4 at1g55000 triple mutant were identical to those of the axr4-1 and axr4-2 single mutants. These findings clearly demonstrated that the abnormal phenotypes described in the above paper were ascribed solely to the mutation in the AXR4 gene.
An important question is why did some of our mca2-null germplasms have the axr4−/axr4− at1g55000−/at1g55000− allele? We had never used axr4 mutants in our laboratory before the above paper was published. We speculated that some of the seeds of the mca2-null mutant (germplasm name: SALK_129208) obtained from the ABRC 12 years ago were heterozygous for the AXR4 and AT1G55000 loci (i.e., AXR4+/axr4− AT1G55000+/at1g55000−), and multiple self-pollinations performed by us to maintain seed viability produced seed stocks with axr4−/axr4− at1g55000−/at1g55000− as well as AXR4+/AXR4+ AT1G55000+/AT1G55000+ and AXR4+/axr4− AT1G55000+/at1g55000−, although all of the 20 seeds of the mca2-null mutant (germplasm name: SALK_129208C), which were newly obtained from the ABRC and tested, had the genotype of AXR4+/AXR4+ AT1G55000+/AT1G55000+. As for the MCA2 locus, 19 out of the 20 seeds had the mca2−/mca2− genotype and one had the MCA2+/mca2− genotype. The mca1-null mca2-null double mutant and mca2/MCA2 complementation lines used in the above paper were AXR4+/AXR4+ AT1G55000+/AT1G55000+. Furthermore, the mca2-null single and mca1-null mca2-null lines used in our previous study (Yamanaka et al., 2010) were also AXR4+/AXR4+ AT1G55000+/AT1G55000+.
We deeply regret any scientific misconceptions that have been caused by the above paper and apologize to the scientific community for any adverse consequences.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Footnotes
1. ^Database of The Arabidopsis Information Resource (TAIR) at http://www.arabidopsis.org/servlets/TairObject?type=locus&name=AT1G55000
References
Dharmasiri, S., Swarup, R., Mockaitis, K., Dharmasiri, N., Singh, S. K., Kowalchyk, M., et al. (2006). AXR4 is required for localization of the auxin influx facilitator AUX1. Science 312, 1218–1220. doi: 10.1126/science.1122847
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Yamanaka, T., Nakagawa, Y., Mori, K., Nakano, M., Imamura, T., Kataoka, H., et al. (2010). MCA1 and MCA2 that mediate Ca2+ uptake have distinct and overlapping roles in Arabidopsis. Plant Physiol. 152, 1284–1296. doi: 10.1104/pp.109.147371
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Keywords: mechanosensitive channel, Arabidopsis, root, skewing, waving, calcium, touch response, mechanical stress
Citation: Frontiers in Plant Science Editorial Office (2015) Retraction: Mechanosensitive channel candidate MCA2 is involved in touch-induced root responses in Arabidopsis. Front. Plant Sci. 6:153. doi: 10.3389/fpls.2015.00153
Received: 26 February 2015; Accepted: 26 February 2015;
Published online: 05 March 2015.
Approved by:
Steven Carl Huber, USDA-ARS, Urbana, USAAndreas P. M. Weber, Heinrich-Heine-Universität, Germany
Copyright © 2015 Frontiers in Plant Science Editorial Office. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Frontiers in Plant Science Editorial Office,cGxhbnRzY2llbmNlLmVkaXRvcmlhbC5vZmZpY2VAZnJvbnRpZXJzaW4ub3Jn