 Anton Hartmann
Anton Hartmann Michael Rothballer
Michael Rothballer Burkhard A. Hense
Burkhard A. Hense Peter Schröder
Peter Schröder
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
OPINION article
Front. Plant Sci., 08 April 2014
Sec. Plant Pathogen Interactions
Volume 5 - 2014 | https://doi.org/10.3389/fpls.2014.00131
This article is part of the Research TopicThe plant microbiome and its importance for plant and human healthView all 19 articles
Higher organisms evolved in the omnipresence of microbes, which could be of pathogenic or symbiotic nature. A framework of response patterns evolved which is known as innate immunity. A major part of this response is the recognition of microbial-associated molecular patterns (MAMP) such as chitin or lipochitooligosaccharides, peptidoglycan, lipopolysaccharides or flagellum structures, and the initiation of efficient plant defence reactions (Janeway and Medzhitov, 2002; Jones and Dangl, 2006). However, there are many plant-associated endophytic bacteria known, which are living within plants without triggering persistent and apparent defence responses or visibly do not harm the plant. In some cases, even a stimulation of plant growth due to the presence of specific players within the plant microbiome was reported (Turner et al., 2013). It is now generally accepted, that plant performance and activities can only be characterized and understood completely, if the “holobiont,” the plant plus the intimately associated microbiota, is considered (Zilber-Rosenberg and Rosenberg, 2008). The evolutionary advantage of an integrated holobiontic system is characterized by a much better adaptability and flexibility towards rapidly changing adverse environmental conditions. It is still mostly unknown, which particular plant genetic loci are controlling the interactions with the plant microbiome and which signals are steering this cooperativity. Mutualistic microbes are able to overcome or short-circuit plant defence responses to enable successful colonization of the host (Zamioudis and Pieterse, 2012; Alqueres et al., 2013). Beneficial associations with microbes other than mycorhiza or Rhizobia are also controlled by systemically regulated or autoregulated processes on top of the basic innate immunity response. The induction of systemic immunity responses like ISR (induced systemic resistance) by some beneficial rhizosphere bacteria or the SAR (systemic acquired resistance) response provoked by pathogens are results of multiple response cascades employed by the plant host to respond to microbial and other environmental interactions. However, the entire response network is by far not yet revealed. For example, bacteria-induced plant responses resulting in improved resistance towards pathogens can also be due to the perception of secondary metabolites, like the surfactin lipopeptide, produced by certain biocontrol Bacilli (Garcia-Gutiérrez et al., 2013) or volatile compounds of plant-associated microbes (Yi et al., 2010). The biocontrol activity of microbial inoculants is probably due to multiple effects of their secondary metabolites to achieve direct inhibition of the pathogenic counterpart as well as an increase of systemic resistance of the plant host.
It is hypothesized, that eukaryotic organisms developed ways to sense microbes in addition to the recognition of their MAMPs by their diffusible small molecules. A very ancient and basic feature of unicellular bacteria is their way of environmental sensing and social communication. In many Gram-negative bacteria the synthesis of autoinducers of the N-acyl-homoserine lactone (AHL) type is tightly regulated in response to cell density or the cell “quorum” (Eberl, 1999). These metabolites are released into the cellular environment to sense the quality of the ecological niche in terms of diffusion space and the density and distribution of their own population. This environmental sensing mechanism helps to adapt the regulation of their gene expression to the given conditions in their habitat and thus optimizes the fitness of the population. Therefore, the generally known term “quorum sensing” (QS) was supplemented by the more broadly defined concept of “efficiency sensing” (Hense et al., 2007). Since this optimization of in situ gene expression is of very basic importance, autoinducer QS-molecules are widespread among bacteria and have quite different molecular structures. AHL are common in Gram-negative bacteria, while cyclic peptides as QS-signals are only to be found in Gram-positive bacteria. The detailed structure of the AHL-molecules can vary; the acyl side chain consists of 4–14 carbon atoms and may also contain double bonds. The C3-atom can be hydroxylated or oxidized to a carbonyl-carbon; thus, considerable information and quite different physicochemical properties can be present within these different AHL-structures. As is outlined below, also plants have obviously learned during their evolution to respond to these QS compounds in different specific ways. We speculate, that QS-compounds are early signals indicating that pathogens are in the surroundings to gather themselves for attack or that mutualists are about to interact with roots.
The first demonstration of specific responses of a plant to bacterial AHLs was demonstrated for the legumes Phaseolus vulgaris (Joseph and Phillips, 2003) and Medicago truncatula (Mathesius et al., 2003) (Table 1). AHLs from symbiotic (Sinorhizobium meliloti) or pathogenic (Pseudomonas aeruginosa) bacteria provoked at concentrations as low as nano- to micromolar significant changes in the accumulation of over 150 proteins. Auxin-responsive and flavonoid synthesis proteins were induced and also a secretion of plant metabolites that mimic QS compounds were found, which may have the potential to disrupt QS signaling by associated bacteria. In tomato plants, a specific induction of systemic resistance proteins after inoculation of the roots with C4- and C6-side chain AHL-producing Serratia liquefaciens MG1 was discovered independently (Hartmann et al., 2004; Schuhegger et al., 2006). The fungal leaf pathogen Alternaria alternata was much less effective, when S. liquefaciens MG 1 wild type had been inoculated to roots of tomato plants as compared to the AHL-negative mutant. It could be shown, that salicylic acid was increased as well as SA- and ethylene-dependent defence genes (i.e., PR1a) in MG1-inoculated plants. Furthermore, Serratia plymuthica HRO-C48, producing C4-/C6- and OHC4-/OHC6-homoserine lactones, is able to induce ISR-like systemic protection of bean and tomato plants against the fungal leaf pathogen Botrytis cinnera; this response was greatly reduced with mutants impaired in AHL-production (Liu et al., 2007; Pang et al., 2009). In contrast, Arabidopsis thaliana responds to short (C4- and C6-) N-acyl AHL-compounds in a different manner: C4- and C6- homoserine lactones alter the expression of selected hormonal regulated genes which results in changes of the plant's hormone content, in particular an increased auxin/cytokinin ratio (von Rad et al., 2008). However, no systemic resistance response was found to be induced in A. thaliana when roots were stimulated with short side-chain AHLs. Ortíz-Castro et al. (2008) found that C10-homoserine lactone elicited developmental changes in the root system in Arabidopsis plants by altering the expression of cell division and differentiation-related genes. Furthermore, Liu et al. (2012) and Jin et al. (2012) demonstrated that the root stimulatory effect of C6- and C8- homoserine lactones in Arabidopsis plants is mediated through the G-protein coupled receptor encoded by AtGPA1. In mung bean, oxoC10-homoserine lactone activates auxin-induced adventitious root formation via H2O2- and NO-dependent cyclic GMP signaling (Bai et al., 2012). On the other hand, N-acyl-AHLs with C12- and C14- side chains induce systemic resistance to the obligate biotrophic fungus Golovinomyces orontii in A. thaliana and to Blumeria graminis f. sp. hordei in barley (Hordeum vulgare) (Schikora et al., 2011). This response is mediated through altered activation of AtMPK6. The mitogen-activated protein kinases AtMPK3 and AtMPK6 were stronger activated by the model elicitor flg22 in the presence of C12- or C14-AHL compounds which resulted in a higher expression of the defence-related transcription factors WRKY26 and WRKY29 as well as the PR1 gene (Schikora et al., 2011). Thus, AHLs with short and medium side chain lengths induce developmental effects on root architecture, while long side chain AHLs induce systemic resistance in A. thaliana (Schenk et al., 2012). Furthermore, it was shown, that better water soluble short side chain AHL-compounds are actively taken up into plant roots and transported along the roots into the shoot; in contrast, the lipophilic long acyl side chain AHLs are not transported in barley and A. thaliana. (Götz et al., 2007; von Rad et al., 2008; Sieper et al., 2014). However, no uptake was detected in the legume yam bean (Pachyrhizus erosus (L.) Urban) (Götz et al., 2007). The latter finding corroborates the report of Delalande et al. (2005) that legumes like Lotus corniculatus produce lactonases which degrade AHLs and prevent their uptake and transport. In barley, it could further be demonstrated that C8- and C10-AHLs are taken up in a cell energy dependent manner by ABC-transporters into the root and transported via the central cylinder into the shoot (Sieper et al., 2014).
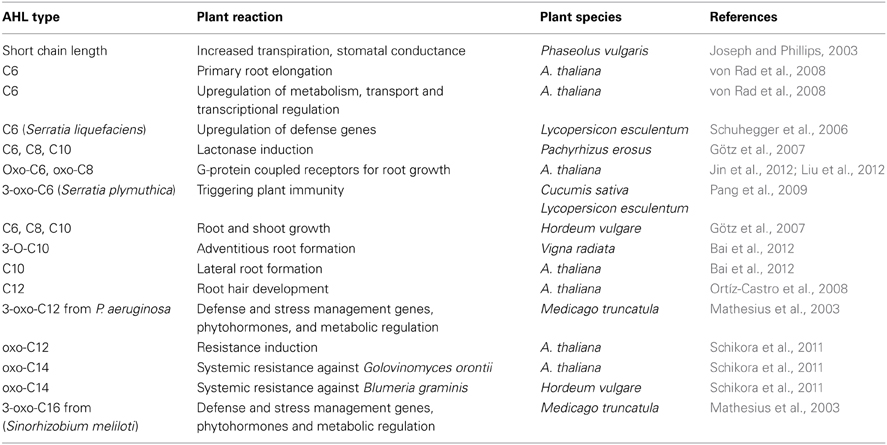
Table 1. Recent findings of direct AHL impact on different plants.
Interestingly, several plants have been demonstrated to produce AHL-mimic substances or to develop other activities influencing QS of plant associated bacteria (Gao et al., 2003; Bauer and Mathesius, 2004). Flavonoids released by legumes increase the expression of AHL synthesis genes in Rhizobia (Pérez-Montano et al., 2011). Indole acetic acid and cytokinin biosynthesis of Gypsophila was shown to influence QS, type III secretion system and gall formation activity by Pantoea plantarum (Chalupowicz et al., 2009). On the other hand, tobacco plants have been engineered to produce short- and long-side chain AHL-compounds which could be detected in substantial amounts at leaf and root surfaces as well as in soil (Scott et al., 2006). Constitutive expression of QS genes in transgenic tobacco plants leads to alteration in induced systemic resistance elicited by the rhizobacterium Serratia marcescens 90–166 (Ryu et al., 2013). Furthermore, transgenic tomato plants engineered to produce different AHL-compounds were demonstrated to alter the activity of plant growth promoting rhizobacteria and resulted, e.g., in increased salt tolerance (Barriuso et al., 2008). We hypothesize, that QS in a plant-microbe holobiont system should be regarded in a bidirectional way with influences from the plant and the microbial partners.
Uptake of AHL-compounds and specific perception of AHLs in animal cells were also studied intensively in recent years (Teplitski et al., 2011; Hartmann and Schikora, 2012). 3-oxo-C12-homoserine lactone (C12-AHL), the major AHL-compound of Pseudomonas aeruginosa, was shown to selectively impair the regulation of the nuclear transcription factor NF-κ B which controls innate immune responses in mammalian cells (Kravchenko et al., 2008). C12-AHL also impaired human dendritic cell functions required for priming of T-cells (Bernatowicz et al., submitted). Since the response to AHL-compounds in mammalian systems is complicated due to the interferences with the adaptive immune system, plants provide an ideal model for the detailed interaction studies of basic innate immune responses and developmental processes with N-acylhomoserine lactones as modifying bacterial effector molecules.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Alqueres, S., Menses, C., Rouws, L., Rothballer, M., Baldani, I., Schmid, M., et al. (2013). The bacterial superoxide dismutase and glutathione reductase are crucial for endophytic colonization of rice roots by Gluconacetobacter diazotrophicus strain PAL5. Mol. Plant-Microbe Interact. 26, 937–945. doi: 10.1094/MPMI-12-12-0286-R
Bai, X., Todd, C. D., Desikan, R., Yang, Y., and Hu, X. (2012). N-3-oxo-decanoyl-L-homoserine-lactone activates auxin-induced adventitious root formation via hydrogen peroxide- and nitric oxide-dependent cyclic GMP signaling in mung bean. Plant Physiol. 158, 725–736. doi: 10.1104/pp.111.185769
Barriuso, J., Solano, B. R., Fray, R. G., Cámara, M., Hartmann, A., and Gutierrez Manero, F. J. (2008). Transgenic tomato plants alter quorum sensing in plant growth-promoting rhizobacteria. Plant Biotech. J. 6, 442–452. doi: 10.1111/j.1467-7652.2008.00331.x
Bauer, W. D., and Mathesius, U. (2004). Plant responses to bacterial quorum sensing signals. Curr. Opin. Plant Biol. 7, 429–433. doi: 10.1016/j.pbi.2004.05.008
Chalupowicz, L., Barash, I., Panijel, M., Sessa, G., and Manulis-Sasson, S. (2009). Regulatory interaction between quorum-sensing, auxin, cytokinin, and the hrp regulon in relation to gall formation and epiphytic fitness of Pantoea agglomerans pv. Gypsophilae. Mol. Plant Microbe Interact. 22, 849–856. doi: 10.1094/MPMI-22-7-0849
Delalande, L., Faure, D., Raffoux, A., Uroz, S., D'Angelo-Picard, C., Elasri, M., et al. (2005). N-hexanoyl-L-homoserine lactone, a mediator of bacterial quorum sensing regulation, exhibits plant-dependent. FEMS Microbiol. Ecol. 52, 13–50.
Eberl, L. (1999). N-acyl-homoserine lactone-mediated gene regulation in Gram-negative bacteria. Syst. Appl. Microbiol. 22, 493–506. doi: 10.1016/S0723-2020(99)80001-0
Gao, M., Teplitski, M., Robinson, J. B., and Bauer, W. D. (2003). Production of substances by Medicago truncatula that affect bacterial quorum sensing. Mol. Plant Microbe Interact. 16, 827–834. doi: 10.1094/MPMI.2003.16.9.827
Garcia-Gutiérrez, L., Zeriouh, H., Romero, D., Cubero, J., de Vicente, A., and Pérez-Garcia A. (2013). The antagonistic strain Bacillus subtilis UMAF6639 also confers protection to melon plants against cucurbit powdery mildew by activation of jasmonate- and salicylic acid-dependent defence response. Microb. Biotechnol. 6, 264–274. doi: 10.1111/1751-7915.12028
Götz, C., Fekete, A., Gebefügi, I., Forczek, S., Fuksová, K., Li, X., et al. (2007). Uptake, degradation and chiral discrimination of N-acyl-D/L-homoserine lactones by barley (Hordeum vulgare) and yam bean (Pachyrhizus erosus) plants. Anal. Bioanal. Chem. 389, 1447–1457. doi: 10.1007/s00216-007-1579-2
Hartmann, A., Gantner, S., Schuhegger, R., Steidle, A., Dürr, C., Schmid, M., et al. (2004). “N-acyl-homoserine lactones of rhizosphere bacteria trigger systemic resistance in tomato plants” in Biology of Molecular Plant-Microbe Interactions, Vol. 4, eds B. Lugtenberg, I. Tikhonovich, and N. Provorov (St. Paul, Minnesota: MPMI-Press), 554–556.
Hartmann, A., and Schikora, A. (2012). Quorum sensing of bacteria and trans-kingdom interactions of N-acyl homoserine lactones with eukaryotes. J. Chem. Ecol. 38, 704–713. doi: 10.1007/s10886-012-0141-7
Hense, B. A., Kuttler, C., Müller, J., Rothballer, M., Hartmann, A., and Kreft, J.-U. (2007). Does efficiency sensing unify diffusion and quorum sensing? Nat. Rev. Microbiol. 5, 230–239. doi: 10.1038/nrmicro1600
Janeway, C. A. Jr., and Medzhitov, R. (2002). Innate immune recognition. Annu. Rev. Immunol. 20, 197–216. doi: 10.1146/annurev.immunol.20.083001.084359
Jin, G., Liu, F., Ma, H., Hao, S., Zhao, Q., Bian, Z., et al. (2012). Two G-protein-coupled-receptor candidates, Cand2 and Cand7, are involved in Arabidopsis root growth mediated by the bacterial quorum-sensing signals N-acyl- homoserine lactones. Biochem. Biophys. Res. Commun. 417, 991–995. doi: 10.1016/j.bbrc.2011.12.066
Jones, J. D. G., and Dangl, J. L. (2006). The plant immune system. Nature 444, 323–329. doi: 10.1038/nature05286
Joseph, C. M., and Phillips, D. A. (2003). Metabolites from soil bacteria affect plant water relations. Plant Physiol. Biochem. 41, 189–192. doi: 10.1016/S0981-9428(02)00021-9
Kravchenko, V. V., Kaufmann, G. F., Mathison, J. C., Scott, D. A., Katz, A. Z., Wood, M. R., et al. (2008). Modulation of gene expression via disruption of NF-κB signalling by a bacterial small molecule. Science 321, 259–263. doi: 10.1126/science.1156499
Liu, F., Bian, Z., Jia, Z., Zhao, Q., and Song, S. (2012). The GCR1 and GPA1 participate in promotion of Arabidopsis primary root elongation induced by N-acyl-homoserine lactones, the bacterial quorum-sensing system. Mol. Plant Microbe Interact. 25, 677–683. doi: 10.1094/MPMI-10-11-0274
Liu, X., Bimerew, M., Ma, Y., Müller, H., Ovadis, M., Eberl, L., et al. (2007). Quorum-sensing signaling is required for production of the antobiotic pyrrolnitrin in a rhizospheric biocontrol strain of Serratia plymuthica. FEMS Microbiol. Lett. 270, 299–305. doi: 10.1111/j.1574-6968.2007.00681.x
Mathesius, U., Mulders, S., Gao, M., Teplitski, M., Caetano-Anoliés, G., Rolfe, B. G., et al. (2003). Extensive and specific responses of a eukaryote to bacterial quorum sensing signals. Proc. Natl. Acad. Sci. U.S.A 100, 1444–1449. doi: 10.1073/pnas.262672599
Ortíz-Castro, R., Martinez-Trujillo, M., and López-Bucio, J. (2008). N-acyl-homoserine lactones: a class of bacterial quorum-sensing signals alter post-embryonic root development in Arabidopsis thaliana. Plant Cell Environ. 31, 1497–1509. doi: 10.1111/j.1365-3040.2008.01863.x
Pang, Y., Liu, X., Ma, Y., Chernin, L., Berg, G., and Gao, K. (2009). Induction of systemic resistance, root colonisation and biocontrol activities of the rhizospheric strain of Serratia plymuthica are dependent on N-acyl homoserine lactones. Eur. J. Plant Pathol. 124, 261–268. doi: 10.1007/s10658-008-9411-1
Pérez-Montano, F., Guasch-Vidal, B., González-Barroso, S., López-Baena, F. J., Cubo, T., Ollero, F. J., et al. (2011). Nodulation-gene-inducing flavonoids increase overall production of autoinducers and expression of N-acyl-homoserine lactone synthesis genes in Rhizobia. Res. Microbiol. 162, 715–723. doi: 10.1016/j.resmic.2011.05.002
Ryu, C.-M., Choi, H. K., Lee, C.-H., Murphy, J. F., Lee, J.-K., and Kloepper, J. W. (2013). Modulation of quorum sensing in acyl-homoserine lactone-producing or—degrading tobacco plants leads to alteration of induced systemic resistance elicited by the rhizobacterium Serratia marcescens 90–166. Plant Pathol. J. 29, 182–192. doi: 10.5423/PPJ.SI.11.2012.0173
Schenk, S. T., Stein, E., Kogel, K.-H., and Schikora, A. (2012). Arabidopsis growth and defense are modulated by bacterial quorum sensing molecules. Plant Signal. Behav. 7, 178–181. doi: 10.4161/psb.18789
Schikora, A., Schenk, S. T., Stein, E., Molitor, A., Zuccaro, A., and Kogel, K.-H. (2011). N-acyl-homoserine lactone confers resistance toward biotrophic and hemibiotrophic pathogens via altered activation of AtMPK6. Plant Physiol. 157, 1407–1418. doi: 10.1104/pp.111.180604
Schuhegger, R., Ihring, A., Gantner, S., Bahnweg, G., Knappe, C., Vogg, G., et al. (2006) Induction of systemic resistance in tomato plants by N-acyl-homoserine lactone–producing rhizosphere bacteria. Plant Cell Environ. 29, 909–918. doi: 10.1111/j.1365-3040.2005.01471.x
Scott, R. A., Weil, J., Le, P. T., Williams, P., Fray, R. G., von Bodman, S. B., et al. (2006). Long- and short chain plant-produced bacterial N-acyl-homoserine lactones become components of phyllosphere, rhizosphere, and soil. Mol. Plant-Microbe Interact. 19, 227–239. doi: 10.1094/MPMI-19-0227
Sieper, T., Forczek, S., Matucha, M., Krämer, P., Hartmann, A., and Schröder, P. (2014). N-acyl-homoserine lactone uptake and systemic transport in barley rest upon active parts of the plant. New Phytol. 201, 545–555. doi: 10.1111/nph.12519
Teplitski, M., Mathesius, U., and Rumbaugh, K. P. (2011). Perception and degradation of N-acyl-homoserine lactone quorum sensing signals by mammalian and plant cells. Chem. Rev. 111, 100–116. doi: 10.1021/cr100045m
Turner, T. R., James, E. K., and Poole, P. S. (2013). The plant microbiome. Genome Biol. 14, 209–219. doi: 10.1186/gb-2013-14-6-209
von Rad, U., Klein, I., Dobrev, P. I., Kottova, J., Zazimalova, E., Fekete, A., et al. (2008). The response of Arabidopsis thaliana to N-hexanoyl-DL-homoserine-lactone, a bacterial quorum sensing molecule produced in the rhizosphere. Planta 229, 73–85. doi: 10.1007/s00425-008-0811-4
Yi, H.-S., Ryu, C.-M., and Heil, M. (2010). Sweet smells prepare plants for future stress—airborne induction of plant disease immunity. Plant Signal. Behav. 5, 528–531. doi: 10.4161/psb.10984
Zamioudis, C., and Pieterse, C. M. J. (2012). Modulation of host immunity by beneficial microbes. Mol. Plant Microbe Interact. 25, 139–150. doi: 10.1094/MPMI-06-11-0179
Keywords: Quorum sensing, N-acyl-homoserine lactones, systemic responses, innate immunity, mutualists, endophytic bacteria
Citation: Hartmann A, Rothballer M, Hense BA and Schröder P (2014) Bacterial quorum sensing compounds are important modulators of microbe-plant interactions. Front. Plant Sci. 5:131. doi: 10.3389/fpls.2014.00131
Received: 30 September 2013; Accepted: 19 March 2014;
Published online: 08 April 2014.
Edited by:
Gabriele Berg, Graz University of Technology, AustriaReviewed by:
Ingyu Hwang, Seoul National University, South KoreaCopyright © 2014 Hartmann, Rothballer, Hense and Schröder. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence:YW50b24uaGFydG1hbm5AaGVsbWhvbHR6LW11ZW5jaGVuLmRl
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.